
Effects of equine chorionic gonadotropin and type of ovulatory stimulus in a timed-AI protocol on...
12
UNCORRECTED PROOF + Models THE 10941 1–12 Please cite this article in press as: Souza AH, et al. Effects of equine chorionic gonadotropin and type of ovulatory stimulus in a timed-AI protocol on reproductive responses in dairy cows. Theriogenology (2009), doi:10.1016/j.theriogenology.2008.12.025 Effects of equine chorionic gonadotropin and type of ovulatory stimulus in a timed-AI protocol on reproductive responses in dairy cows A.H. Souza a , S. Viechnieski b , F.A. Lima c , F.F. Silva d , R. Arau ´jo a , G.A. Bo ´ e , M.C. Wiltbank f , P.S. Baruselli a, * a Department of Animal Reproduction, FMVZ-USP, Brazil b Iguac ¸u Farm-StarMilk, Cascavel, PR, Brazil c Veterinary Policlinic-Pioneiros, Carambeı ´, PR, Brazil d Department of Informatics, UFV, Brazil e Institute of Animal Reproduction-Co ´rdoba, IRAC, Co ´rdoba, Argentina f Department of Dairy Science, UW-Madison, Madison, WI, USA Received 1 July 2008; received in revised form 1 December 2008; accepted 20 December 2008 Abstract The objectives were to evaluate the effects of equine chorionic gonadotropin (eCG) supplementation (with or without eCG) and type of ovulatory stimulus (GnRH or ECP) on ovarian follicular dynamics, luteal function, and pregnancies per AI (P/AI) in Holstein cows receiving timed artificial insemination (TAI). On Day 0, 742 cows in a total of 782 breedings, received 2 mg of estradiol benzoate (EB) and one intravaginal progesterone (P4) insert (CIDR). On Day 8, the CIDR was removed, and all cows were given PGF2a and assigned to one of four treatments in a 2 2 factorial arrangement: (1) CG: GnRH 48 h later; (2) CE: ECP; (3) EG: eCG + GnRH 48 h later; (4) EE: eCG + ECP. There were significant interactions for eCG ovulatory stimulus and eCG BCS. Cows in the CG group were less likely (28.9% vs. 33.8%; P < 0.05) to become pregnant compared with those in the EG group (odds ratio [OR] = 0.28). There were no differences in P/AI between CE and EE cows (30.9% vs. 29.1%; OR = 0.85; P = 0.56), respectively. Thinner cows not receiving eCG had lower P/AI than thinner cows receiving eCG (15.2% vs. 38.0%; OR = 0.20; P < 0.01). Treatment with eCG tended to increase serum progestesterone concentrations during the diestrus following synchronized ovulation (P < 0.10). However, the treatment used to induce ovulation did not affect CL volume or serum progesterone concentrations. In conclusion, both ECP and GnRH yielded comparable P/AI. However, eCG treatment at CIDR removal increased pregnancy rate in cows induced to ovulate with GnRH and in cows with lower BCS. # 2009 Elsevier Inc. All rights reserved. Keywords: eCG; Estrogen; Progesterone; Timed AI; Dairy cow 1. Introduction Timed artificial insemination (TAI) protocols, to inseminate without the need for detection of estrus, have been developed to increase service rates in cows [1–3]. Generally, these protocols use GnRH and PGF2a (Ovsynch protocol [1]) to precisely synchronize the www.theriojournal.com Available online at www.sciencedirect.com Theriogenology xxx (2009) xxx–xxx 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 35 36 37 38 39 40 41 * Corresponding author at: Departamento de Reproduc ¸a ˜o Animal – VRA, Av: Prof. Dr. Orlando Marques de Paiva, no. 87 – Cidade Universita ´ ria, CEP: 05508-000, Sa ˜o Paulo – SP, Brazil. Tel.: +55 11 3091 7674; fax: +55 11 3091 7412. E-mail address: [email protected] (P.S. Baruselli). 0093-691X/$ – see front matter # 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.theriogenology.2008.12.025
Transcript of Effects of equine chorionic gonadotropin and type of ovulatory stimulus in a timed-AI protocol on...

+ Models
THE 10941 1–12
1
2
3
4
5
6
7
8
9
10
11
1213
141516
17
18
19
20
21
22
23
24
25
26
27
28
29
30
3132
33
34
35
36
CO
RR
EC
TED
PR
OO
F
Effects of equine chorionic gonadotropin and type of ovulatory
stimulus in a timed-AI protocol on reproductive responses
in dairy cows
A.H. Souza a, S. Viechnieski b, F.A. Lima c, F.F. Silva d, R. Araujo a,G.A. Bo e, M.C. Wiltbank f, P.S. Baruselli a,*
a Department of Animal Reproduction, FMVZ-USP, Brazilb Iguacu Farm-StarMilk, Cascavel, PR, Brazil
c Veterinary Policlinic-Pioneiros, Carambeı, PR, Brazild Department of Informatics, UFV, Brazil
e Institute of Animal Reproduction-Cordoba, IRAC, Cordoba, Argentinaf Department of Dairy Science, UW-Madison, Madison, WI, USA
Received 1 July 2008; received in revised form 1 December 2008; accepted 20 December 2008
Abstract
The objectives were to evaluate the effects of equine chorionic gonadotropin (eCG) supplementation (with or without eCG) and
type of ovulatory stimulus (GnRH or ECP) on ovarian follicular dynamics, luteal function, and pregnancies per AI (P/AI) in
Holstein cows receiving timed artificial insemination (TAI). On Day 0, 742 cows in a total of 782 breedings, received 2 mg of
estradiol benzoate (EB) and one intravaginal progesterone (P4) insert (CIDR). On Day 8, the CIDR was removed, and all cows were
given PGF2a and assigned to one of four treatments in a 2 � 2 factorial arrangement: (1) CG: GnRH 48 h later; (2) CE: ECP; (3)
EG: eCG + GnRH 48 h later; (4) EE: eCG + ECP. There were significant interactions for eCG � ovulatory stimulus and
eCG � BCS. Cows in the CG group were less likely (28.9% vs. 33.8%; P < 0.05) to become pregnant compared with those
in the EG group (odds ratio [OR] = 0.28). There were no differences in P/AI between CE and EE cows (30.9% vs. 29.1%;
OR = 0.85; P = 0.56), respectively. Thinner cows not receiving eCG had lower P/AI than thinner cows receiving eCG (15.2% vs.
38.0%; OR = 0.20; P < 0.01). Treatment with eCG tended to increase serum progestesterone concentrations during the diestrus
following synchronized ovulation (P < 0.10). However, the treatment used to induce ovulation did not affect CL volume or serum
progesterone concentrations. In conclusion, both ECP and GnRH yielded comparable P/AI. However, eCG treatment at CIDR
removal increased pregnancy rate in cows induced to ovulate with GnRH and in cows with lower BCS.
# 2009 Elsevier Inc. All rights reserved.
Keywords: eCG; Estrogen; Progesterone; Timed AI; Dairy cow
www.theriojournal.com
Available online at www.sciencedirect.com
Theriogenology xxx (2009) xxx–xxx
UN
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
35
36
37
38
39
40
41
* Corresponding author at: Departamento de Reproducao Animal –
VRA, Av: Prof. Dr. Orlando Marques de Paiva, no. 87 – Cidade
Universitaria, CEP: 05508-000, Sao Paulo – SP, Brazil.
Tel.: +55 11 3091 7674; fax: +55 11 3091 7412.
E-mail address: [email protected] (P.S. Baruselli).
0093-691X/$ – see front matter # 2009 Elsevier Inc. All rights reserved.
doi:10.1016/j.theriogenology.2008.12.025
1. Introduction
Timed artificial insemination (TAI) protocols, to
inseminate without the need for detection of estrus, have
been developed to increase service rates in cows [1–3].
Generally, these protocols use GnRH and PGF2a
(Ovsynch protocol [1]) to precisely synchronize the
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
Cascavel-PR,
Original text:
Inserted Text
-PR,
Original text:
Inserted Text
IRAC-C
Original text:
Inserted Text
1)
Original text:
Inserted Text
2) CE: ECP; 3)
Original text:
Inserted Text
4)
Original text:
Inserted Text
(28.9 vs.
Original text:
Inserted Text
(30.9 vs.
Original text:
Inserted Text
(15.2 vs.
Original text:
Inserted Text
VRA;
Original text:
Inserted Text
°
Original text:
Inserted Text
ria; CEP: 05508-000; São Paulo
Original text:
Inserted Text
+11

R
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx2
+ Models
THE 10941 1–12
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
UN
CO
R
time of AI. In addition, short-acting estrogens in
conjunction with progestagen treatments have been
used to synchronize the emergence of a new follicular
wave during TAI protocols [3] and to avoid the
production of persistent follicles during progesterone-
based protocols [4]. Although service rates are
generally improved during TAI procedures, fertility
following synchronized ovulation is sub-optimal [5].
One potential problem with TAI protocols in
lactating dairy cows is the decrease in circulating
estradiol concentrations prior to TAI [6]. This reduction
in circulating estradiol is partly caused by the high rate
of steroid metabolism in dairy cows, due to increased
hepatic blood flow, in association with the elevated feed
intake of high-producing cows [7]. In addition, TAI
protocols induce premature ovulation of the follicle in
order to synchronize ovulation in all cows; this can
further reduce peak circulating estradiol concentrations
[6,8]. Previous studies have reported some benefits of
supplementing estrogen during the final stages of TAI
protocols [6,9–11]. Thus, supplementation with estro-
gens near TAI may be a rational approach to improve
fertility in TAI protocols for lactating cows.
A further consequence of prematurely inducing
ovulation during TAI protocols is that the diameter of
the ovulatory follicle is somewhat reduced [8,12,13].
Vasconcelos et al. [12] reported a close relationship
between diameter of the ovulatory follicle and CL
volume as well as circulating progesterone (P4)
concentrations. Thus, premature ovulation following
TAI procedures might induce ovulation of smaller
follicles, formation of a smaller CL, and with
concurrent high liver metabolism of ovarian steroids,
decrease circulating P4 during the luteal phase of high-
producing cows. Several studies have indicated that low
circulating P4 may suppress early embryonic develop-
ment [14,15]. Treatment with eCG prior to ovulation
has increased circulating P4 during the subsequent
luteal phase [16]. Studies in beef cattle in which P4-
based TAI protocols were supplemented with eCG
concurrent with removal of the P4 insert provided
evidence that eCG increased the percentage of cows that
ovulated to the TAI protocol, increased circulating P4
following TAI, and increased fertility to TAI [17].
Accordingly, strategies to increase circulating P4 after
ovulation could improve embryo survival for TAI
protocols.
We chose estradiol cypionate (ECP), a long-acting
conjugated estradiol, as a source of estrogen to be given
concurrent with P4 insert removal, to increase
circulating estrogen concentrations during the peri-
ovulatory period and minimize animal handling. In
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
EC
TED
PR
OO
F
addition, cows were treated with or without eCG at
CIDR removal, to increase subsequent luteal function.
The main hypotheses of this study were: (1) eCG
treatment will increase CL volume, circulating P4 and
P/AI; (2) ECP will increase P/AI; and (3) combining
eCG with ECP will produce the greatest increase in P/
AI following TAI. Therefore, this experiment was
designed to test whether eCG treatment at CIDR
removal (with or without eCG) and type of stimulus
(ECP or GnRH) used to induce ovulation, can increase
fertility in lactating cows following a TAI protocol.
2. Materials and methods
2.1. Cows and management
Lactating Holstein cows were used in this experiment
(ovarian follicular dynamics, n = 96 cows; field fertility
test, n = 742 cows in a total of 782 breedings) having
average (mean � S.E.M.) body condition score [18] of
2.88 � 0.01 (CG = 2.89 � 0.02; CE = 2.89 � 0.02;
EG = 2.87 � 0.02; and EE = 2.86 � 0.02), milk produc-
tion (daily average from Day 0 to Day 8) of
36.2 � 0.4 kg/d (CG = 35.9 � 0.6 kg/d; CE = 36.8 �0.6 kg/d; EG = 35.4 � 0.6 kg/d; and EE = 36.6 �0.6 kg/d), and average days in milk of 151.6 � 3.5
(CG = 151.1 � 6.7; CE = 153.7 � 7.0; EG = 145.5 �6.8; and EE = 156.2 � 7.8). The study was replicated
in 22 free-stall dairy herds in southeastern Brazil. Healthy
postpartum cows with >50 d in milk and cows not
pregnant from the last AI before enrollment in the study
were assigned to hormonal treatments. The experimental
period began in December 2005 and ended in September
2006. On all farms, cows were milked twice daily and fed
a TMR that consisted of corn silage and alfalfa silage as
forage, with a corn-soybean meal-based concentrate. The
TMR was balanced to meet or exceed minimum
nutritional requirements for lactating dairy cows [19].
All animal procedures were approved by the Animal Care
Committee of the Faculty of Veterinary Medicine and
Husbandry, University of Sao Paulo, Sao Paulo, SP,
Brazil.
2.2. Hormones and experimental design
All cows received 2 mg of estradiol benzoate (EB;
Estrogin; Farmavet Produtos Veterinarios, SP, Brazil)
and one intravaginal progesterone insert (CIDR; 1.9 g
of P4; Pfizer Animal Health, SP, Brazil) on Day 0. At
CIDR removal (Day 8), all cows received 25 mg of
PGF2a (Lutalyse Sterile Solution; Pfizer Animal
Health). Simultaneously (Day 8), cows were randomly
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
1)
Original text:
Inserted Text
2)
Original text:
Inserted Text
3)
Original text:
Inserted Text
SEM)

RE
CTE
D P
RO
OF
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx 3
+ Models
THE 10941 1–12
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
Fig. 1. Experimental design. US = ovarian ultrasound examination and BS = blood sample. *Ultrasound evaluation performed every 12 h for 96 h
after CIDR removal or until ovulation was detected. **Blood samples were collected every 4 h from 30 to 58 h after CIDR removal. ***Blood
Q2 samples and ovarian ultrasound examinations were performed every 2–3 d for 21 d after synchronized ovulation. Pregnancy diagnosis was
performed by ultrasound 33 d after timed AI.
UN
CO
R
assigned within each farm to four treatments (Fig. 1), in
a 2 � 2 factorial design, as follows: (1) Group CG:
GnRH (100 mg Fertagyl; Intervet Schering-Plough, SP,
Brazil) 48 h after CIDR removal (Day 10); (2) Group
CE: ECP (1 mg ECP; Pfizer Animal Health) at CIDR
removal (Day 8); (3) Group EG: eCG (400 IU; Folligon;
Intervet Schering-Plough) on Day 8 + GnRH (100 mg)
on Day 10; (4) Group EE: eCG (400 IU) + ECP (1 mg)
on Day 8.
2.3. Follicular dynamics
In a subset of the cows (Subset A; n = 45), transrectal
ovarian ultrasound examinations were performed every
24 h from Day 0 (CIDR insertion) to Day 8 (CIDR
removal). Wave emergence was defined retrospectively
when the dominant follicle was 4–5 mm in diameter, as
described previously [20]. After CIDR removal, ovarian
ultrasound examinations were performed every 12 h in
two subsets of cows (Subset A, n = 45 and Subset
B = 51; total = 96 cows) from CIDR removal to
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
disappearance of the ovulatory follicle, or 96 h after
CIDR withdrawal, whichever occurred first. In another
subset of cows (Subset B; LH analysis, n = 40) blood
samples were collected every 4 h from 30 to 58 h after
CIDR removal to evaluate the time of the LH peak.
Cows in Subsets A and B were not inseminated.
Subsequently, ovulating cows in Subset A were
evaluated every 2 or 3 d after synchronized ovulation
for 21 d. The total volume of the CL formed after TAI in
ovulating cows was calculated with the following
formula: V = (4/3)pR3; R was defined as R = (Da/
2 + Db/2)/2, where Da and Db are the perpendicular
diameters of the CL. If the CL had a cavity, it was
calculated with the same formula and deleted from the
total volume of the CL. An ultrasound scanner with
linear-array transducer (6.0/8.0 MHz) was used to
perform all measurements of ovarian structures (Falco
100, Pie Medical Equipment B.V., Maastricht, Hol-
land). On the day of CIDR removal (Day 8), all cows in
Subsets A and B (n = 96) were fitted with a Kamar (heat
mount patch) and visually checked for signs of estrus
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
1)
Original text:
Inserted Text
2)
Original text:
Inserted Text
3)
Original text:
Inserted Text
4)
Original text:
Inserted Text
(subset
Original text:
Inserted Text
4 to 5
Original text:
Inserted Text
subset
Original text:
Inserted Text
4/3×π×R
Original text:
Inserted Text
perpendicular
Original text:
Inserted Text
examination.
Original text:
Inserted Text
2 to 3 d for 21 d

R
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx4
+ Models
THE 10941 1–12
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
UN
CO
R
(defined as standing estrus, with an activated Kamar)
every 4 h for 48 h after CIDR removal. Day 0 of the
estrous cycle subsequent to hormonal treatments was
defined as the day of the synchronized ovulation.
2.4. Field fertility trial
In the fertility study, 742 cows (in a total of 782
breedings) randomly received the same hormonal
treatments as a 2 � 2 factorial design (Fig. 1). The
term ‘‘cow’’ and ‘‘breeding’’ is used interchangeably
throughout the manuscript to define our experimental
unit ‘‘breeding’’. Semen from 30 sires was used and
homogenously distributed among treatments and farms.
Timed AI was performed 58 h after CIDR removal and
pregnancy diagnosis was performed by ultrasonography
33 d later. Pregnancy was confirmed based on the
echodensity of the fluid in the uterine horns and the
presence of a heartbeat in the embryo.
2.5. Hormone assays
Blood samples were collected by puncture of
coccygeal vessels, immediately refrigerated, centrifuged
(3000 � g for 20 min), and serum samples were stored at
�20 8C until assayed for P4 concentrations. Circulating
P4 was evaluated from unextracted sera using an
antibody-coated-tube RIA kit (Diagnostic Products
Corporation, Los Angeles, CA, USA). The intra- and
inter-assay CVs were 4.3% and 4.6%, respectively.
Circulating concentrations of LH were determined by
RIA, as previously described [21]. The intra- and inter-
assay CVs for LH were2.3% and 3.6%, respectively. The
LH peak was defined as an increase in LH< 2SD above
the overall within-cow mean of LH concentrations [22].
2.6. Statistical analyses
The binomial distribution was assumed for the
categorical response variables, such as expression of
estrus, ovulation rate, double ovulation rate, and P/AI.
The variable P/AI was analyzed by using procedure
GLIMMIX of SAS [23], with cow treated as a random
effect. Variables initially considered for inclusion in the
models were eCG (with or without eCG), type of
ovulatory stimulus (GnRH or ECP), farm within season
(warmer or cooler months), parity (categorized as 1 or
�2), days in milk (continuous variable), BCS (categor-
ized as <2.75 or �2.75), milk production (continuous
variable), sire, CL presence at CIDR insertion, and
interactions. The variable farm was nested within
season in the model, because not all farms had cows
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
EC
TED
PR
OO
F
treated in both seasons. The final logistic regression
model removed variables by backward elimination,
based on the Wald statistics criterion when P > 0.20.
Variables that were included in the final model for
analysis of P/AI were: eCG (with or without eCG), type
of ovulatory stimulus (GnRH or ECP), BCS, interaction
eCG � type of ovulatory stimulus, interaction
eCG � BCS, interaction among eCG � type of ovula-
tory stimulus � BCS, and farm within season. Some
variables such as expression of estrus, number of LH
peaks detected, ovulation rate, and double ovulation
rate, were analyzed with the GENMOD procedure of
SAS. The final model for these dependent variables
accounted for effects of eCG, ovulatory stimulus, and
interaction between eCG and ovulatory stimulus.
Comparisons of CL volume and circulating P4 during
the cycle following synchronized ovulation were
performed using the MIXED procedure of SAS [24].
Square root transformation was used for CL volume and
log transformation was used for circulating progester-
one to attain normality. The model included the effects
of eCG, ovulatory stimulus, BCS, time, interaction
between eCG � time, interaction between ovulatory
stimulus � time, interaction between BCS � time,
interaction among eCG � ovulatory stimulus � time,
milk production, number of ovulations, and cow, which
was treated as a random effect and was the subject for
the repeated measures. Variables such as time to
ovulation, follicular wave emergence, dominant follicle
size 48 h after CIDR removal, time to LH peak, LH
peak amplitude, and area under the curve of LH release
(calculated by the trapezoid method), were analyzed
using the GLM procedure of SAS, and the final model
accounted for the effects of eCG, ovulatory stimulus,
and interaction between eCG � ovulatory stimulus.
Correlations between milk production and day of
follicular wave emergence after CIDR insertion were
evaluated with the CANCORR procedure of SAS. The
correlation model considered effects of days in milk,
parity and BCS. Levene’s test was used to compare
variances in the time of LH peak and ovulation after
CIDR removal. Probabilities of P < 0.05 were con-
sidered significant, whereas probabilities between 0.05
and 0.10 were considered tendencies.
3. Results
3.1. Follicular dynamics and hormonal responses
following CIDR + EB
Combining a CIDR with 2 mg of EB was effective in
synchronizing follicular wave emergence, with 84.4%
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
4.3 and
Original text:
Inserted Text
2.3 and
Original text:
Inserted Text
LH >2 SD
Original text:
Inserted Text
within cow mean of LH concentrations.
Original text:
Inserted Text
ECP).Effects of equine chorionic
Original text:
Inserted Text
ECP).Table 2Proportion of cows
R
PR
OO
F
Q1
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx 5
+ Models
THE 10941 1–12
275
276
277
278
279
280
281
282
283
284
285
286286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
323
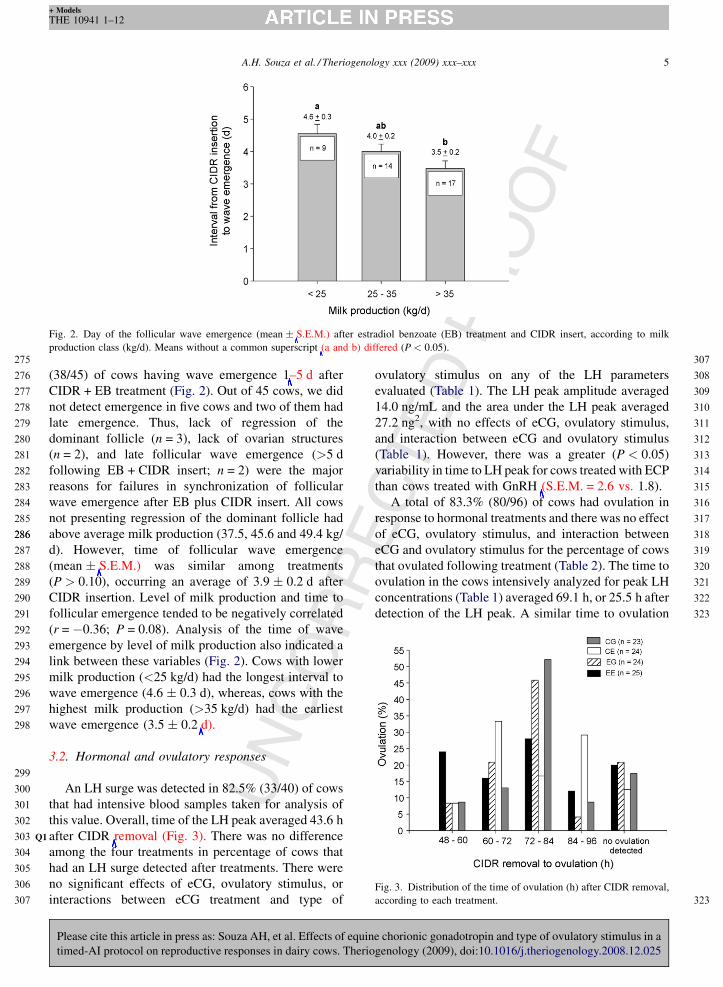
Fig. 2. Day of the follicular wave emergence (mean � S.E.M.) after estradiol benzoate (EB) treatment and CIDR insert, according to milk
production class (kg/d). Means without a common superscript (a and b) differed (P < 0.05).
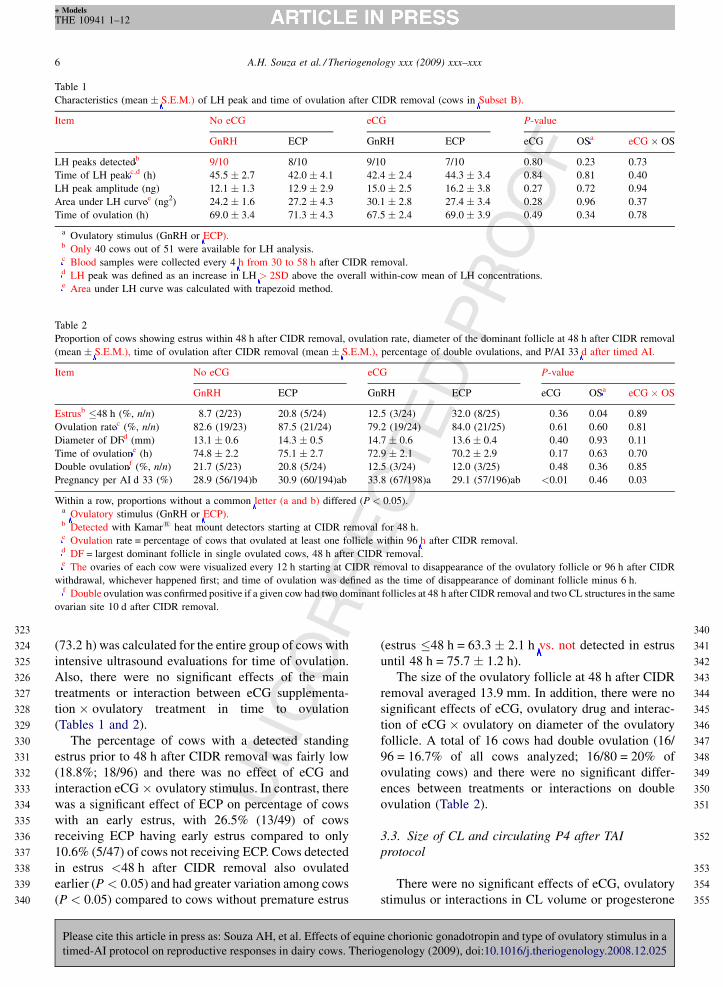
Fig. 3. Distribution of the time of ovulation (h) after CIDR removal,
according to each treatment.
UN
CO
R(38/45) of cows having wave emergence 1–5 d after
CIDR + EB treatment (Fig. 2). Out of 45 cows, we did
not detect emergence in five cows and two of them had
late emergence. Thus, lack of regression of the
dominant follicle (n = 3), lack of ovarian structures
(n = 2), and late follicular wave emergence (>5 d
following EB + CIDR insert; n = 2) were the major
reasons for failures in synchronization of follicular
wave emergence after EB plus CIDR insert. All cows
not presenting regression of the dominant follicle had
above average milk production (37.5, 45.6 and 49.4 kg/
d). However, time of follicular wave emergence
(mean � S.E.M.) was similar among treatments
(P > 0.10), occurring an average of 3.9 � 0.2 d after
CIDR insertion. Level of milk production and time to
follicular emergence tended to be negatively correlated
(r = �0.36; P = 0.08). Analysis of the time of wave
emergence by level of milk production also indicated a
link between these variables (Fig. 2). Cows with lower
milk production (<25 kg/d) had the longest interval to
wave emergence (4.6 � 0.3 d), whereas, cows with the
highest milk production (>35 kg/d) had the earliest
wave emergence (3.5 � 0.2 d).
3.2. Hormonal and ovulatory responses
An LH surge was detected in 82.5% (33/40) of cows
that had intensive blood samples taken for analysis of
this value. Overall, time of the LH peak averaged 43.6 h
after CIDR removal (Fig. 3). There was no difference
among the four treatments in percentage of cows that
had an LH surge detected after treatments. There were
no significant effects of eCG, ovulatory stimulus, or
interactions between eCG treatment and type of
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
EC
TED
ovulatory stimulus on any of the LH parameters
evaluated (Table 1). The LH peak amplitude averaged
14.0 ng/mL and the area under the LH peak averaged
27.2 ng2, with no effects of eCG, ovulatory stimulus,
and interaction between eCG and ovulatory stimulus
(Table 1). However, there was a greater (P < 0.05)
variability in time to LH peak for cows treated with ECP
than cows treated with GnRH (S.E.M. = 2.6 vs. 1.8).
A total of 83.3% (80/96) of cows had ovulation in
response to hormonal treatments and there was no effect
of eCG, ovulatory stimulus, and interaction between
eCG and ovulatory stimulus for the percentage of cows
that ovulated following treatment (Table 2). The time to
ovulation in the cows intensively analyzed for peak LH
concentrations (Table 1) averaged 69.1 h, or 25.5 h after
detection of the LH peak. A similar time to ovulation
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
1 to 5 d
Original text:
Inserted Text
SEM)
Original text:
Inserted Text
0.2 d).Fig. 33.2
Original text:
Inserted Text
removal.
Original text:
Inserted Text
(SEM=2.6 vs
Original text:
Inserted Text
SEM)
Original text:
Inserted Text
differed
RE
CTE
D P
RO
OF
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx6
+ Models
THE 10941 1–12
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
Table 1
Characteristics (mean � S.E.M.) of LH peak and time of ovulation after CIDR removal (cows in Subset B).
Item No eCG eCG P-value
GnRH ECP GnRH ECP eCG OSa eCG � OS
LH peaks detectedb 9/10 8/10 9/10 7/10 0.80 0.23 0.73
Time of LH peakc,d (h) 45.5 � 2.7 42.0 � 4.1 42.4 � 2.4 44.3 � 3.4 0.84 0.81 0.40
LH peak amplitude (ng) 12.1 � 1.3 12.9 � 2.9 15.0 � 2.5 16.2 � 3.8 0.27 0.72 0.94
Area under LH curvee (ng2) 24.2 � 1.6 27.2 � 4.3 30.1 � 2.8 27.4 � 3.4 0.28 0.96 0.37
Time of ovulation (h) 69.0 � 3.4 71.3 � 4.3 67.5 � 2.4 69.0 � 3.9 0.49 0.34 0.78
a Ovulatory stimulus (GnRH or ECP).b Only 40 cows out of 51 were available for LH analysis.c Blood samples were collected every 4 h from 30 to 58 h after CIDR removal.d LH peak was defined as an increase in LH > 2SD above the overall within-cow mean of LH concentrations.e Area under LH curve was calculated with trapezoid method.
Table 2
Proportion of cows showing estrus within 48 h after CIDR removal, ovulation rate, diameter of the dominant follicle at 48 h after CIDR removal
(mean � S.E.M.), time of ovulation after CIDR removal (mean � S.E.M.), percentage of double ovulations, and P/AI 33 d after timed AI.
Item No eCG eCG P-value
GnRH ECP GnRH ECP eCG OSa eCG � OS
Estrusb �48 h (%, n/n) 8.7 (2/23) 20.8 (5/24) 12.5 (3/24) 32.0 (8/25) 0.36 0.04 0.89
Ovulation ratec (%, n/n) 82.6 (19/23) 87.5 (21/24) 79.2 (19/24) 84.0 (21/25) 0.61 0.60 0.81
Diameter of DFd (mm) 13.1 � 0.6 14.3 � 0.5 14.7 � 0.6 13.6 � 0.4 0.40 0.93 0.11
Time of ovulatione (h) 74.8 � 2.2 75.1 � 2.7 72.9 � 2.1 70.2 � 2.9 0.17 0.63 0.70
Double ovulationf (%, n/n) 21.7 (5/23) 20.8 (5/24) 12.5 (3/24) 12.0 (3/25) 0.48 0.36 0.85
Pregnancy per AI d 33 (%) 28.9 (56/194)b 30.9 (60/194)ab 33.8 (67/198)a 29.1 (57/196)ab <0.01 0.46 0.03
Within a row, proportions without a common letter (a and b) differed (P < 0.05).a Ovulatory stimulus (GnRH or ECP).b Detected with Kamar1 heat mount detectors starting at CIDR removal for 48 h.c Ovulation rate = percentage of cows that ovulated at least one follicle within 96 h after CIDR removal.d DF = largest dominant follicle in single ovulated cows, 48 h after CIDR removal.e The ovaries of each cow were visualized every 12 h starting at CIDR removal to disappearance of the ovulatory follicle or 96 h after CIDR
withdrawal, whichever happened first; and time of ovulation was defined as the time of disappearance of dominant follicle minus 6 h.f Double ovulation was confirmed positive if a given cow had two dominant follicles at 48 h after CIDR removal and two CL structures in the same
ovarian site 10 d after CIDR removal.
UN
CO
R
(73.2 h) was calculated for the entire group of cows with
intensive ultrasound evaluations for time of ovulation.
Also, there were no significant effects of the main
treatments or interaction between eCG supplementa-
tion � ovulatory treatment in time to ovulation
(Tables 1 and 2).
The percentage of cows with a detected standing
estrus prior to 48 h after CIDR removal was fairly low
(18.8%; 18/96) and there was no effect of eCG and
interaction eCG � ovulatory stimulus. In contrast, there
was a significant effect of ECP on percentage of cows
with an early estrus, with 26.5% (13/49) of cows
receiving ECP having early estrus compared to only
10.6% (5/47) of cows not receiving ECP. Cows detected
in estrus <48 h after CIDR removal also ovulated
earlier (P < 0.05) and had greater variation among cows
(P < 0.05) compared to cows without premature estrus
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
(estrus �48 h = 63.3 � 2.1 h vs. not detected in estrus
until 48 h = 75.7 � 1.2 h).
The size of the ovulatory follicle at 48 h after CIDR
removal averaged 13.9 mm. In addition, there were no
significant effects of eCG, ovulatory drug and interac-
tion of eCG � ovulatory on diameter of the ovulatory
follicle. A total of 16 cows had double ovulation (16/
96 = 16.7% of all cows analyzed; 16/80 = 20% of
ovulating cows) and there were no significant differ-
ences between treatments or interactions on double
ovulation (Table 2).
3.3. Size of CL and circulating P4 after TAI
protocol
There were no significant effects of eCG, ovulatory
stimulus or interactions in CL volume or progesterone
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
vs not
Original text:
Inserted Text
SEM)
Original text:
Inserted Text
subset B).No eCGeCGP-value
Original text:
Inserted Text
f
Original text:
Inserted Text
a9/10
Original text:
Inserted Text
b,c
Original text:
Inserted Text
d
Original text:
Inserted Text
ECP),
Original text:
Inserted Text
bBlood
Original text:
Inserted Text
4 h from 30 to 58 h after CIDR
Original text:
Inserted Text
removal.
Original text:
Inserted Text
<2 SD
Original text:
Inserted Text
dArea
Original text:
Inserted Text
SEM),
Original text:
Inserted Text
SEM),
Original text:
Inserted Text
33
Original text:
Inserted Text
eeCG×OSLH peaks detecteda9/10
Original text:
Inserted Text
b
Original text:
Inserted Text
c
Original text:
Inserted Text
d
Original text:
Inserted Text
e
Original text:
Inserted Text
superscript
Original text:
Inserted Text
Detected with Kamar® heat
Original text:
Inserted Text
ECP),
Original text:
Inserted Text
bOvulation
Original text:
Inserted Text
96 h after CIDR removal.cDF=
Original text:
Inserted Text
removal.
Original text:
Inserted Text
dThe
Original text:
Inserted Text
eDouble
RE
CTE
D P
RO
OF
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx 7
+ Models
THE 10941 1–12
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
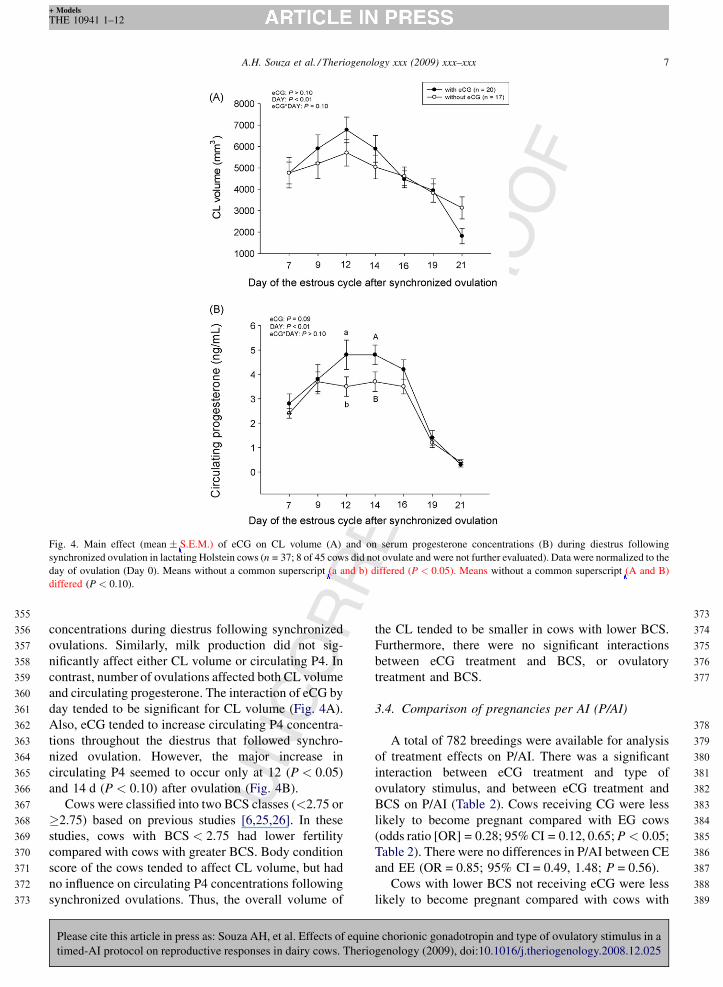
Fig. 4. Main effect (mean � S.E.M.) of eCG on CL volume (A) and on serum progesterone concentrations (B) during diestrus following
synchronized ovulation in lactating Holstein cows (n = 37; 8 of 45 cows did not ovulate and were not further evaluated). Data were normalized to the
day of ovulation (Day 0). Means without a common superscript (a and b) differed (P < 0.05). Means without a common superscript (A and B)
differed (P < 0.10).
UN
CO
Rconcentrations during diestrus following synchronized
ovulations. Similarly, milk production did not sig-
nificantly affect either CL volume or circulating P4. In
contrast, number of ovulations affected both CL volume
and circulating progesterone. The interaction of eCG by
day tended to be significant for CL volume (Fig. 4A).
Also, eCG tended to increase circulating P4 concentra-
tions throughout the diestrus that followed synchro-
nized ovulation. However, the major increase in
circulating P4 seemed to occur only at 12 (P < 0.05)
and 14 d (P < 0.10) after ovulation (Fig. 4B).
Cows were classified into two BCS classes (<2.75 or
�2.75) based on previous studies [6,25,26]. In these
studies, cows with BCS < 2.75 had lower fertility
compared with cows with greater BCS. Body condition
score of the cows tended to affect CL volume, but had
no influence on circulating P4 concentrations following
synchronized ovulations. Thus, the overall volume of
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
the CL tended to be smaller in cows with lower BCS.
Furthermore, there were no significant interactions
between eCG treatment and BCS, or ovulatory
treatment and BCS.
3.4. Comparison of pregnancies per AI (P/AI)
A total of 782 breedings were available for analysis
of treatment effects on P/AI. There was a significant
interaction between eCG treatment and type of
ovulatory stimulus, and between eCG treatment and
BCS on P/AI (Table 2). Cows receiving CG were less
likely to become pregnant compared with EG cows
(odds ratio [OR] = 0.28; 95% CI = 0.12, 0.65; P < 0.05;
Table 2). There were no differences in P/AI between CE
and EE (OR = 0.85; 95% CI = 0.49, 1.48; P = 0.56).
Cows with lower BCS not receiving eCG were less
likely to become pregnant compared with cows with
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
SEM)
Original text:
Inserted Text
differed
Original text:
Inserted Text
differed (P<0.05).Fig. 3Distribution
R
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx8
+ Models
THE 10941 1–12
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
419
420
421
422
423
424424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
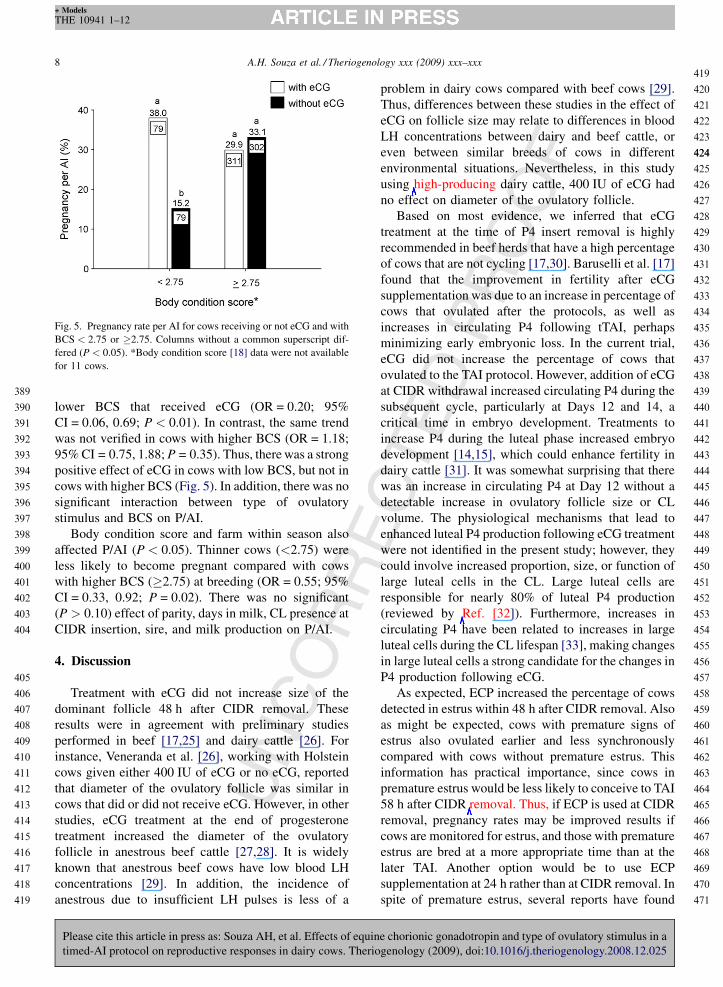
Fig. 5. Pregnancy rate per AI for cows receiving or not eCG and with
BCS < 2.75 or �2.75. Columns without a common superscript dif-
fered (P < 0.05). *Body condition score [18] data were not available
for 11 cows.
UN
CO
Rlower BCS that received eCG (OR = 0.20; 95%
CI = 0.06, 0.69; P < 0.01). In contrast, the same trend
was not verified in cows with higher BCS (OR = 1.18;
95% CI = 0.75, 1.88; P = 0.35). Thus, there was a strong
positive effect of eCG in cows with low BCS, but not in
cows with higher BCS (Fig. 5). In addition, there was no
significant interaction between type of ovulatory
stimulus and BCS on P/AI.
Body condition score and farm within season also
affected P/AI (P < 0.05). Thinner cows (<2.75) were
less likely to become pregnant compared with cows
with higher BCS (�2.75) at breeding (OR = 0.55; 95%
CI = 0.33, 0.92; P = 0.02). There was no significant
(P > 0.10) effect of parity, days in milk, CL presence at
CIDR insertion, sire, and milk production on P/AI.
4. Discussion
Treatment with eCG did not increase size of the
dominant follicle 48 h after CIDR removal. These
results were in agreement with preliminary studies
performed in beef [17,25] and dairy cattle [26]. For
instance, Veneranda et al. [26], working with Holstein
cows given either 400 IU of eCG or no eCG, reported
that diameter of the ovulatory follicle was similar in
cows that did or did not receive eCG. However, in other
studies, eCG treatment at the end of progesterone
treatment increased the diameter of the ovulatory
follicle in anestrous beef cattle [27,28]. It is widely
known that anestrous beef cows have low blood LH
concentrations [29]. In addition, the incidence of
anestrous due to insufficient LH pulses is less of a
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
EC
TED
PR
OO
F
problem in dairy cows compared with beef cows [29].
Thus, differences between these studies in the effect of
eCG on follicle size may relate to differences in blood
LH concentrations between dairy and beef cattle, or
even between similar breeds of cows in different
environmental situations. Nevertheless, in this study
using high-producing dairy cattle, 400 IU of eCG had
no effect on diameter of the ovulatory follicle.
Based on most evidence, we inferred that eCG
treatment at the time of P4 insert removal is highly
recommended in beef herds that have a high percentage
of cows that are not cycling [17,30]. Baruselli et al. [17]
found that the improvement in fertility after eCG
supplementation was due to an increase in percentage of
cows that ovulated after the protocols, as well as
increases in circulating P4 following tTAI, perhaps
minimizing early embryonic loss. In the current trial,
eCG did not increase the percentage of cows that
ovulated to the TAI protocol. However, addition of eCG
at CIDR withdrawal increased circulating P4 during the
subsequent cycle, particularly at Days 12 and 14, a
critical time in embryo development. Treatments to
increase P4 during the luteal phase increased embryo
development [14,15], which could enhance fertility in
dairy cattle [31]. It was somewhat surprising that there
was an increase in circulating P4 at Day 12 without a
detectable increase in ovulatory follicle size or CL
volume. The physiological mechanisms that lead to
enhanced luteal P4 production following eCG treatment
were not identified in the present study; however, they
could involve increased proportion, size, or function of
large luteal cells in the CL. Large luteal cells are
responsible for nearly 80% of luteal P4 production
(reviewed by Ref. [32]). Furthermore, increases in
circulating P4 have been related to increases in large
luteal cells during the CL lifespan [33], making changes
in large luteal cells a strong candidate for the changes in
P4 production following eCG.
As expected, ECP increased the percentage of cows
detected in estrus within 48 h after CIDR removal. Also
as might be expected, cows with premature signs of
estrus also ovulated earlier and less synchronously
compared with cows without premature estrus. This
information has practical importance, since cows in
premature estrus would be less likely to conceive to TAI
58 h after CIDR removal. Thus, if ECP is used at CIDR
removal, pregnancy rates may be improved results if
cows are monitored for estrus, and those with premature
estrus are bred at a more appropriate time than at the
later TAI. Another option would be to use ECP
supplementation at 24 h rather than at CIDR removal. In
spite of premature estrus, several reports have found
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
high producing
Original text:
Inserted Text
[32
Original text:
Inserted Text
cDF=largest dominant follicle

R
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx 9
+ Models
THE 10941 1–12
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
UN
CO
R
positive effects on fertility by supplementing estrogen
in the pro-estrus period [6,34]. However, other studies
[35–37], including the present results, did not detect an
overall positive effect of estrogen on conception rates.
The timing, dose, or type of estrogen, as well as breed
and BCS of cow and experimental environment, may all
have contributed to differences between studies in the
effects of estrogen supplementation.
Cows with lower BCS had lower P/AI, tended to have
reduced CL volume, but had no significant difference in
circulating P4 during diestrus after synchronized ovula-
tion. Santos et al. [31], reported that lactating cows with
lower BCS (�2.75) tended to have lower circulating P4 in
diestrus compared to cows with higher BCS. Data from
Santos et al. [31] might indicate that cows with lower
BCS have either greater steroid metabolism and/or lower
luteal P4 production. It has been shown that dairy cows
with lower BCS have greater feed intake than higher BCS
cows [38], potentially increasing rates of steroid
metabolism in the liver [7] and decreasing circulating
P4. In addition, Gansworthy and Topps [38] found that
cows with lower BCS produced more milk than fatter
cows. However, in the current trial, milk production was
similar between cows with lower and higher BCS (lower
BCS = 33.0 � 2.6 kg/d vs. higher BCS = 35.6 � 1.0 kg/
d; P > 0.10). Regardless, in the current trial, there was a
significant interaction between eCG and BCS on P/AI.
Perhaps eCG treatment either improved ovulation rate or
CL development, particularly in cows with lower BCS,
and this may have lead to increased fertility in these cows.
Due to a limited number of cows with lower BCS during
the follicular dynamics evaluation, we were unable to
detect a significant interaction between eCG treatment
and BCS on ovulation results. Thus, other eCG-induced
mechanisms underlying the enhancement of fertility in
low BCS cows, such as improved ovulation rate,
follicular or oocyte function or increased circulating
estradiol, cannot be excluded.
Lactating cows with BCS < 2.75 had lower fertility
compared with cows with greater BCS [6,39,40].
Interestingly, in the current trial, cows with lower
BCS not receiving eCG were less likely to become
pregnant than cows with lower BCS that received eCG.
Veneranda et al. [26] reported that lactating cows
supplemented with eCG had greater fertility than
control cows. However, in a second study done by this
author [29], no fertility differences were detected in
cows treated with eCG compared with controls.
Nevertheless, when all three experiments were taken
together [41], pregnancy rates were higher (P < 0.01) in
eCG-treated cows than in non-eCG-treated cows. Other
studies suggested that eCG increased conception
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
EC
TED
PR
OO
F
outcomes in cows not cycling before TAI treatments
[42]. In the current trial, any improvements in P/AI
following the use of eCG seemed to be limited to cows
with lower BCS, but not to cyclicity before TAI.
Regardless, future studies using eCG supplementation
in cattle should compare treatments with different eCG
doses, and also take into account the possibility of anti-
eCG antibody after repeated treatments with high doses
of eCG [43].
There was no significant effect of ovulatory stimulus
or interaction between type of ovulatory stimulus and
BCS on P/AI. These results were not in agreement with
recent reports that described beneficial effects of
supplementing with a short-acting estrogen [6] in
protocols for TAI in lactating cows. A more recent
report [37], also using lactating cows, using the same
long-acting estrogen used in the current trial (ECP) did
not report improvements in fertility in animals with low
BCS treated for TAI. Thus, based on this previous study
and our present results, we inferred that improvements
in fertility with ECP supplementation in TAI protocols
were somewhat inconsistent. Because more cows
showed premature estrus in ECP group, improvements
in fertility might be possible with additional estrus
detection and breeding of cows expressing premature
estrus, or by delaying ECP treatment to avoid premature
estruses. However, this topic warrants further research.
The time of LH peak after CIDR removal was not
influenced by eCG, ovulatory treatment or their
interaction, and occurred on average 43.6 � 1.5 h after
CIDR removal. Consequently, the mean time of
ovulation was also similar among treatments. Colazo
et al. [8], using beef heifers, reported that long-lasting
estrogens, e.g. ECP, can be effectively used concurrent
with removal of the P4 insert, to synchronize ovulation
in TAI protocols. In the present study using high-
producing cows, it seems likely that ECP, also given at
CIDR removal, decreased the synchrony of the LH peak
and ovulation, compared to cows treated with GnRH
48 h later. In addition, more frequent sampling for the
LH peak (every 4 h) allowed detection of the greater
variation in this variable; whereas, greater numbers of
cows were required to statistically detect differences in
variation for time to ovulation, due to less frequent
evaluations (every 12 h) to detect ovulation following
CIDR removal. Together, data presented in the current
report clearly detected differences in variation in the
time of ovulation in cows treated with ECP versus
GnRH; there were some indications that time to
ovulation was more variable in ECP-treated cows.
Milk production might also contribute to variation in
the timing of the LH peak and ovulation in ECP-treated
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
vs higher
Original text:
Inserted Text
non-eCG treated
Original text:
Inserted Text
cLH peak was defined as an
Original text:
Inserted Text
high producing

R
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx10
+ Models
THE 10941 1–12
575
576
577
578
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
618
619
620
621
622
623
624
625
626
627
627
628
629
630
631
632
633
634
635
636
637
638
639
640
641
642
643
644
645
646
647
648
649
650
651
652
653
654
655
656
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
UN
CO
R
cows, due to greater estradiol metabolism in cows with
greater milk production. Thus, greater variation in the
time of ovulation in ECP-treated cows might be
expected to compromise fertility due to inconsistent
and, in some cases, inadequate intervals from AI to
ovulation [44,45]. Nonetheless, in spite of the increased
variability in timing of the LH peak and ovulation in
ECP-treated cows, we did not detect any differences in
fertility between groups that had ovulation synchro-
nized with ECP or GnRH. Perhaps, the increased
variability in time of ovulation in cows treated with ECP
was not sufficient to compromise fertility. Alternatively,
an increase in circulating estradiol concentrations in
ECP-treated cows may have provided some enhance-
ment in fertility, as previously observed [6,10,11] that
overcame any negative effects of increased variability in
time of ovulation. Additionally, in GnRH-treated cows,
it is also likely that the ideal (�16 h) interval from
GnRH treatment to insemination used in the current trial
(�10 h) was not attained as suggested by previous
research [45], which could also have lowered overall
conception rate results in GnRH-treated cows. Thus,
more experiments comparing these two types of
ovulatory stimulus, given at different times in relation
to device removal and TAI, are needed.
The reasons for the significant interaction between
eCG and type of ovulatory stimulus on P/AI are unclear.
However, differences in time to ovulation following
CIDR removal between cows treated with ECP or
GnRH might provide some rationale for this interaction.
Accordingly, it is likely that EG cows underwent a
better combination of synchronization of ovulation
produced by the GnRH treatment associated with
enhancements in CL function produced by the eCG
treatment. Thus, in cows treated only with GnRH, CL
function could have been compromised. Alternatively,
ECP groups had greater variation in time to ovulation.
Thus, cows ovulating prematurely were bred somewhat
late and eCG was not able to improve fertility, due to
inadequate insemination-to-ovulation intervals. Con-
versely, in cows with delayed ovulations, perhaps the
overexposure of the oocyte to the LH-like actions of
eCG hastened oocyte maturation, causing the ovulation
of an aged oocyte. Another physiological explanation
for the significant eCG by type of ovulatory stimulus
interactions was potential increases to more optimal
blood estradiol concentrations near the time of AI in EG
but not CG cows, and/or excessive estrogen concentra-
tions near AI in EE but not CE cows.
An interesting finding in the current trial, not
prospectively hypothesized, was that level of milk
production seemed to affect the time of follicular
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
EC
TED
PR
OO
F
emergence after EB and CIDR insertion. Burke et al.
[46], studying the effects of four doses of EB (0, 1, 2, or
4 mg) with P4 in a P4-based TAI protocol, reported that
greater doses of estradiol delayed follicular emergence.
Because follicular emergence only occurred after
circulating estradiol concentrations returned to basal,
these researchers indicated that the time of follicular
emergence seemed to depend on the rate of estradiol
clearance from the blood. In this regard, Sangsritavong
et al. [7] found that rate of steroid metabolism by the liver
was much higher in lactating dairy cows compared to
non-lactating cows. It is possible that in the current study
higher producing cows had greater estradiol clearance
that could lead to an earlier return of circulated estradiol
to basal concentrations after an EB treatment, with
subsequent earlier emergence of the next follicular wave.
Thus, based on our results combined with this previous
information, we speculated that greater estrogen doses
might be required in cows with greater milk production in
order to consistently cause dominant follicle atresia and
eventually improve the synchrony of follicular emer-
gence in these TAI protocols.
In conclusion, improvements in fertility in response to
eCG were limited to cows with lower BCS. Estradiol
cypionate can be successfully used to induce ovulation in
P4-based TAI protocols for lactating cows. Nevertheless,
in this study, there was greater variation in time of
ovulation with ECP compared to GnRH. Although no
fertility differences were detected between these two
treatments to induce ovulation, perhaps better timing of
ECP treatment following CIDR removal might reduce
variation in time of ovulation and improve fertility. Thus,
additional estrus detection and breeding seem to be
required when ECP is used at CIDR removal. More
studies are necessary toexplain the underlyingphysiology
for eCG enhancement of fertility and its interactions with
BCS and type of ovulatory stimulus on the bovine CL.
Acknowledgments
The authors thank dairy producers from Parana, Rio
Grande do Sul, and Sao Paulo, Brazil and their staff, for
all the help and the use of their herds to conduct these on-
farm trials. We also thank the reviewers for their critical
inputs in data analysis. This research was supported by
FAPESP of Brazil (process number 05/59009-0).
References
[1] Pursley JR, Mee MO, Wiltbank MC. Synchronization of ovula-
tion in dairy cows using PGF2 and GnRH. Theriogenology
1995;44:915–23.
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025
Original text:
Inserted Text
witj

R
A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx 11
+ Models
THE 10941 1–12
675
676
677
678
679
680
681
682
683
684
685
686
687
688
689
690
691
692
693
694
695
696
697
698
699
700
701
702
703
704
705
706
707
708
709
710
711
712
713
714
715
716
717
718
719
720
721
722
723
724
725
726
727
728
729
730
731
732
733
734
735
736
737
737
738
739
740
741
742
743
744
745
746
747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
763
764
765
766
767
768
769
770
771
772
773
774
775
776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
793
794
795
796
797
798
799
UN
CO
R
[2] Martinez MF, Adams GP, Kastelic JP, Bergfelt DR, Mapletoft
RJ. Induction of follicular wave emergence for estrus synchro-
nization and artificial insemination in heifers. Theriogenology
2000;54: 757–69.
[3] Bo GA, Adams GP, Caccia M, Martinez M, Pierson RA,
Mapletoft RJ. Ovarian follicular wave emergence after treatment
with progestagen and estradiol in cattle. Anim Reprod Sci
1995;39: 193–204.
[4] Kinder JE, Kojima FN, Bergfeld EG, Wehrman ME, Fike KE.
Progestin and estrogen regulation of pulsatile LH release and
development of persistent ovarian follicles in cattle. J Anim Sci
1996;74:1424–40.
[5] Santos JEP, Juchem SO, Cerri RLA, Galvao KN, Chebel RC,
Thatcher WW, et al. Effect of bST and reproductive management
on reproductive and lactational performance of Holstein dairy
cows. J Dairy Sci 2004;87:868–81.
[6] Souza AH, Gumen A, Silva EPB, Cunha AP, Guenther JN, Peto
CM, et al. Supplementation with estradiol-17b before the last
GnRH of the Ovsynch protocol in lactating dairy cows. J Dairy
Sci 2007;90:4623–34.
[7] Sangsritavong S, Combs DK, Sartori R, Armentano LE, Wilt-
bank MC. High feed intake increases liver blood flow and
metabolism of progesterone and estradiol-17b in dairy cattle.
J Dairy Sci 2002;85:2831–42.
[8] Colazo MG, Kastelic JP, Mapletoft RJ. Effects of estradiol
cypionate (ECP) on ovarian follicular dynamics, synchrony of
ovulation, and fertility in CIDR-based, fixed-time AI programs
in beef heifers. Theriogenology 2003;60:855–65.
[9] Hanlon DW, Williamson NB, Wichtell JJ, Steffert IJ, Craigie
AL, Pfeiffer DU. The effect of estradiol benzoate administration
on estrus response and synchronized pregnancy rate in dairy
heifers after treatment with exogenous progesterone. Theriogen-
ology 1996;45:775–85.
[10] Cerri RL, Santos JE, Juchem SO, Galvao KN, Chebel RC. Timed
artificial insemination with estradiol cypionate or insemination
at estrus in high-producing dairy cows. J Dairy Sci 2004;87:
3704–15.
[11] Colazo MG, Kastelic JP, Whittaker PR, Gavaga QA, Wilde R,
Mapletoft RJ. Fertility in beef cattle given a new or previously
used CIDR insert and estradiol, with or without progesterone.
Anim Reprod Sci 2004;81:25–34.
[12] Vasconcelos JL, Silcox RW, Rosa GJ, Pursley JP, Wiltbank MC.
Synchronization rate, size of the ovulatory follicle, and preg-
nancy rate after synchronization of ovulation beginning on
different days of the estrous cycle in lactating dairy cows.
Theriogenology 1999;52:1067–78.
[13] Mussard ML, Burke CR, Behlke EJ, Gasser CL, Day ML.
Influence of premature induction of a luteinizing hormone surge
with gonadotropin-releasing hormone on ovulation, luteal func-
tion, and fertility in cattle. J Anim Sci 2007;85:937–43.
[14] Garrett GE, Geisert RD, Zavy MT, Morgan GL. Evidence for
maternal regulation of early conceptus growth and development
in beef cattle. J Reprod Fertil 1988;84:437–46.
[15] Mann GE, Fray MD, Lamming GE. Effects of time of proges-
terone supplementation on embryo development and interferon-t
production in the cow. Vet J 2006;171:500–3.
[16] Murphy BD, Martinuk SD. Equine chorionic gonadotropin.
Endocr Rev 1991;12:27–43.
[17] Baruselli PS, Reis EL, Marques MO, Nasser LF, Bo GA. The use
of hormonal treatments to improve reproductive performance of
anestrous beef cattle in tropical climates. Anim Reprod Sci
2004;82/83:479–86.
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
EC
TED
PR
OO
F
[18] Edmonson AJ, Lean IJ, Weaver LD, Farver T, Webster G. A body
condition scoring chart for Holstein dairy cows. J Dairy Sci
1989;72:68–78.
[19] NRC. Nutrient requirements of dairy cattle, 7th rev. ed.,
Washington, DC: National Academic Science; 2001.
[20] Ginther OJ, Knopf L, Kastelic JP. Temporal associations among
ovarian events in cattle during oestrous cycles with two and three
follicular waves. J Reprod Fertil 1989;87:223–30.
[21] Bolt DJ, Scott V, Kiracofe GH. Plasma LH and FSH after
estradiol, norgestomet and Gn-RH treatment in ovariectomized
beef heifers. Anim Reprod Sci 1990;23:263–71.
[22] Haughian JM, Ginther OJ, Kot K, Wiltbank MC. Relationships
between FSH patterns and follicular dynamics and the temporal
associations among hormones in natural and GnRH-induced
gonadotropin surges in heifers. Reproduction 2004;127:23–33.
[23] SAS Institute. SAS User’s Guide: Statistics, Version 9. 1 for
Windows. Cary, NC: SAS Ins.; 2002–2003.
[24] Littell RC, Milliken GA, Stroup W, Wolfinger RD. SAS system
of mixed models. Cary, NC: Statistical Analysis System Insti-
tute; 1996.
[25] Small GA, Colazo MG, Kastelic JP, Mapletoft RJ. Effects of
progesterone presynchronization and eCG on pregnancy rates to
GnRH-based, timed-AI in beef cattle. Theriogenology 2009;71:
698–706.
[26] Veneranda G, Filippi L, Racca D, Romero G, Balla E, Cutaia L,
et al. Pregnancy rates in dairy cows treated with intravaginal
progesterone devices and different fixed-time AI protocols.
Reprod Fertil Dev 2006;18:118.
[27] Sa Filho MF, Reis EL, Viel Jr JO, Nichi M, Madureira EH,
Baruselli PS. Follicular dynamics in anestrous lactating Nelore
treated with ear implant, eCG and GnRH. Act Sci Vet 2004;32:
235.
[28] Marana D, Cutaia L, Peres L, Pincinato D, Borges LFK, Bo GA.
Ovulation and pregnancy rates in postpartum Bos indicus cows
treated with progesterone vaginal inserts and estradiol benzoate,
with or without eCG and temporary weaning. Reprod Fertil Dev
2006;18:116–7.
[29] Wiltbank MC, Gumen A, Sartori R. Physiological classification
of anovulatory conditions in cattle. Theriogenology 2002;57:21–
52.
[30] Bo GA, Baruselli PS, Martinez MF. Pattern and manipulation of
follicular development in Bos indicus cattle. Anim Reprod Sci
2003;78:307–26.
[31] Santos JE, Thatcher WW, Pool L, Overton MW. Effect of human
chorionic gonadotropin on luteal function and reproductive
performance of high-producing lactating Holstein dairy cows.
J Anim Sci 2001;79:2881–94.
[32] Diaz FJ, Anderson LE, Wu YL, Rabot A, Tsai SJ, Wiltbank MC.
Regulation of progesterone and prostaglandin F2a production in
the CL. Mol Cell Endocrinol 2002;191:65–80.
[33] Alila HW, Hansel W. Origin of different cell types in the bovine
corpus luteum as characterized by specific monoclonal antibo-
dies. Biol Reprod 1984;31:1015.
[34] Lopes AS, Butler ST, Gilbert RO, Butler WR. Relationship of
pre-ovulatory follicle size, estradiol concentrations and season to
pregnancy outcome in dairy cows. Anim Reprod Sci 2007;99:
34–43.
[35] Stevenson JS, Tiffany SM, Lucy MC. Use of estradiol cypionate
as a substitute for GnRH in protocols for synchronizing ovula-
tion in dairy cattle. J Dairy Sci 2004;87:3298–305.
[36] Sellars CB, Dalton JC, Manzo R, Day J, Ahmadzadeh A. Time
and incidence of ovulation and conception rates after incorpor-
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025

A.H. Souza et al. / Theriogenology xxx (2009) xxx–xxx12
+ Models
THE 10941 1–12
799800
801
802
803
804
805
806
807
808
809
810
811
812
813
814
815
816
817
818
819
820
820
821
822
823
824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841841
ating estradiol cypionate into a timed artificial insemination
protocol. J Dairy Sci 2006;89:620–6.
[37] Hillegass J, Lima FS, Sa Filho MF, Santos JEP. Effect of time of
artificial insemination and supplemental estradiol on reproduc-
tion of lactating cows. J Dairy Sci 2008;91:4226–37.
[38] Gansworthy PC, Topps JH. The effect of body condition of dairy
cows at calving on their food intake and performance when given
complete diets. Anim Prod 1982;35:113–9.
[39] Moreira F, Risco C, Pires MF, Ambrose JD, Drost M, DeLorenzo
M, et al. Effect of body condition on reproductive efficiency of
lactating dairy cows receiving a timed insemination. Theriogen-
ology 2000;53:1305–19.
[40] Galvao KN, Santos JE, Juchem SO, Cerri RL, Coscioni AC,
Villasenor M. Effect of addition of a progesterone intravaginal
insert to a timed insemination protocol using estradiol cypionate
on ovulation rate, pregnancy rate, and late embryonic loss in
lactating dairy cows. J Anim Sci 2004;82:3508–17.
[41] Bo GA, Cutaia L, Souza AH, Baruselli PS. Systematic repro-
ductive management in dairy herds. In: New Zealand Veterinary
Association (NZVA) Conference. New Zealand, July 5–July 7:
Christchurch Convention Centre; 2007. p. 155–68.
UN
CO
RR
Please cite this article in press as: Souza AH, et al. Effects of equin
timed-AI protocol on reproductive responses in dairy cows. Therio
PR
OO
F
[42] Bryan MA, Emslie R, Heuer C. Comparative efficacy of an 8-day
Cue-Mate/estradiol benzoate program with or without inclusion
of equine chorionic gonadotropin in anestrus dairy cows. Reprod
Fertil Dev 2008;20:85.
[43] Drion PV, De Roover R, Houtain J, McNamara EM, Remy B,
Sulon J, et al. Increase of plasma eCG binding rate after
administration of repeated high dose of eCG to cows. Reprod
Nutr Dev 2001;41:207–15.
[44] Dransfield MBG, Nebel RL, Pearson RE, Warnick LD. Timing
of insemination for dairy cows identified in estrus by a radio-
telemetric estrus detection system. J Dairy Sci 1998;81:1874–
82.
[45] Pursley RJ, Silcox RW, Wiltbank MC. Effect of time of
artificial insemination on pregnancy rates, calving rates,
pregnancy loss, and gender ratio after synchronization of
ovulation in lactating dairy cows. J Dairy Sci 1998;81:
2139–44.
[46] Burke CR, Mussard ML, Gasser CL, Grum DE, Day ML.
Estradiol benzoate delays new follicular wave emergence in a
dose-dependent manner after ablation of the dominant ovarian
follicle in cattle. Theriogenology 2003;60:647–58.
EC
TED
e chorionic gonadotropin and type of ovulatory stimulus in a
genology (2009), doi:10.1016/j.theriogenology.2008.12.025




















