
Effects of calcium deficiency on Coffea arabica. Nutrient changes and
10
Plantand Soil 172: 87-96, 1995. © 1995KluwerAcademicPublishers. PrintedintheNetherlands. Effects of calcium deficiency on Coffea arabica. Nutrient changes and correlation of calcium levels with some photosynthetic parameters Jos6 C. Ramalho, M. C. Rebelo, M. EnuqiaSantos, M. Lufsa Antunes and M. Antonieta Nunes lnstituto de Investigaf~o Cientffica Tropical - Centro de Estudos de Produfao e Tecnologia Agrfcolas, Tapada da Ajuda, Apartado 3014, 1301 Lisboa Codex, Portugal Received8 July 1994.Accepted in revisedform31 October1994 Key words: calcium, Coffea arabica, fluorescence analysis, nutrient relations, photosynthesis Abstract Calcium deficiency was induced in hydroponically grown 1.5-years-old coffee plants with 12-14 pairs of leaves. Calcium was given in the form of Ca(NO3)2: 5, 2.5, 0.1, 0.01 and 0 mM. After 71 days of Ca-treatment root and shoot as well as total biomass were decreased by severe Ca-deficiency. However, a stronger decrease was observed for shoot growth as revealed by the increase in the root/shoot ratio. New leaves were affected showing decreases in the total leaf area and in Leaf Area Duration (LAD). After 91 days of deficiency, leaf protein concentration decreased (by about 45%) in the top leaves while nitrate reductase activity (NRA) and NO3 content showed no significant changes. Total nitrogen and mineral concentrations (P, K, Ca, Mg and Na) were also determined in leaves and roots. With the decrease in calcium concentration in Ca-deficiency conditions, we observed concomitant increases in the concentrations of K +, Mg 2+ and Na + in leaves (maximal changes of 32% for K +, 96% for Mg 2+ and 438% for Na +) and in roots (108% for K +, 86% for Mg 2+ and 38% for Na+). Accordingly, the ratio between elements changed, including the ratio N/P, showing a non-equilibrium in the balance of nutrients. Significant correlations were obtained between Ca 2+ concentration and some photosynthetic parameters. Ca-deficiency conditions would increase the loss of energy as expressed by the rise in qE and decrease the photochemical efficiency, which confirms the importance of this element in the stabilization of chlorophyll and in the maintenance of good photochemical efficiency at PS II level. Abbreviations: Chl - Chlorophyll, Fv/Fm - ratio of variable to maximal fluorescence, LAD - leaf area duration, LHC II - light harvesting complex of PS II, NRA - nitrate reductase activity, PC - photosynthetic capacity, PS II - photosystem II, P680 - reaction center of PS II, qN - non-photochemical quenching, qE -- high-energy dependent quenching, qp - photochemical quenching, SLA - specific leaf area. Introduction Calcium has an important role in plant metabolism, since it is required for numerous physiological pro- cesses. Adequate supply of Ca 2+ in the soil solution is an important factor for the control of the severity of specific ion toxicities, namely of aluminium and heavy metals ions (Grattan and Grieves, 1993; Hawkins and Lewis, 1993b; Rengel, 1992). Under saline condi- tions, high Ca 2+ levels have protective effects, since Na + influx and consequently its damaging effects are reduced (Jacoby, 1993). Its presence is essential for a good growth, in density and length, of root hairs which are very important for nutrient uptake (Jaunin and Hofer, 1988). Apart from the structural function of calcium namely in the stabilisation of the cell wall and plasma membrane, it has a recognised role on the processes of cell division and elongation, in the poly- merisation of proteins, and as an enzyme regulator. As reviewed by Moncrief et al. (1990) over 150 different calcium-modulated proteins have been characterized, e.g. NAD kinases, ATPases, dehydrogenases and poly- merases. Considerable evidence also shows that calci- um acts as signal transducer (Roberts and Harmon,
-
Upload
isa-ulisboa -
Category
Documents
-
view
1 -
download
0
Transcript of Effects of calcium deficiency on Coffea arabica. Nutrient changes and

Plant and Soil 172: 87-96, 1995. © 1995KluwerAcademicPublishers. Printedin theNetherlands.
Effects of calcium deficiency on Coffea arabica. Nutrient changes and correlation of calcium levels with some photosynthetic parameters
Jos6 C. Ramalho, M. C. Rebelo, M. Enuqia Santos, M. Lufsa Antunes and M. Antonieta Nunes lnstituto de Investigaf~o Cientffica Tropical - Centro de Estudos de Produfao e Tecnologia Agrfcolas, Tapada da Ajuda, Apartado 3014, 1301 Lisboa Codex, Portugal
Received 8 July 1994. Accepted in revised form 31 October 1994
Key words: calcium, Coffea arabica, fluorescence analysis, nutrient relations, photosynthesis
A b s t r a c t
Calcium deficiency was induced in hydroponically grown 1.5-years-old coffee plants with 12-14 pairs of leaves. Calcium was given in the form of Ca(NO3)2: 5, 2.5, 0.1, 0.01 and 0 mM. After 71 days of Ca-treatment root and shoot as well as total biomass were decreased by severe Ca-deficiency. However, a stronger decrease was observed for shoot growth as revealed by the increase in the root/shoot ratio. New leaves were affected showing decreases in the total leaf area and in Leaf Area Duration (LAD). After 91 days of deficiency, leaf protein concentration decreased (by about 45%) in the top leaves while nitrate reductase activity (NRA) and NO3 content showed no significant changes. Total nitrogen and mineral concentrations (P, K, Ca, Mg and Na) were also determined in leaves and roots. With the decrease in calcium concentration in Ca-deficiency conditions, we observed concomitant increases in the concentrations of K +, Mg 2+ and Na + in leaves (maximal changes of 32% for K +, 96% for Mg 2+ and 438% for Na +) and in roots (108% for K +, 86% for Mg 2+ and 38% for Na+). Accordingly, the ratio between elements changed, including the ratio N/P, showing a non-equilibrium in the balance of nutrients. Significant correlations were obtained between Ca 2+ concentration and some photosynthetic parameters. Ca-deficiency conditions would increase the loss of energy as expressed by the rise in qE and decrease the photochemical efficiency, which confirms the importance of this element in the stabilization of chlorophyll and in the maintenance of good photochemical efficiency at PS II level.
Abbreviations: Chl - Chlorophyll, Fv/Fm - ratio of variable to maximal fluorescence, LAD - leaf area duration, LHC II - light harvesting complex of PS II, NRA - nitrate reductase activity, PC - photosynthetic capacity, PS II - photosystem II, P680 - reaction center of PS II, qN - non-photochemical quenching, qE -- high-energy dependent quenching, qp - photochemical quenching, SLA - specific leaf area.
I n t r o d u c t i o n
Calcium has an important role in plant metabolism, since it is required for numerous physiological pro- cesses. Adequate supply of Ca 2+ in the soil solution is an important factor for the control of the severity of specific ion toxicities, namely of aluminium and heavy metals ions (Grattan and Grieves, 1993; Hawkins and Lewis, 1993b; Rengel, 1992). Under saline condi- tions, high Ca 2+ levels have protective effects, since Na + influx and consequently its damaging effects are reduced (Jacoby, 1993). Its presence is essential for
a good growth, in density and length, of root hairs which are very important for nutrient uptake (Jaunin and Hofer, 1988). Apart from the structural function of calcium namely in the stabilisation of the cell wall and plasma membrane, it has a recognised role on the processes of cell division and elongation, in the poly- merisation of proteins, and as an enzyme regulator. As reviewed by Moncrief et al. (1990) over 150 different calcium-modulated proteins have been characterized, e.g. NAD kinases, ATPases, dehydrogenases and poly- merases. Considerable evidence also shows that calci- um acts as signal transducer (Roberts and Harmon,

88
1992), namely, in the expression of specific genes and in new protein synthesis under water stress conditions, resulting in an increase in the tolerance of water deficits (Funkhouser et al., 1993).
It is also known that calcium deficiency inter- fers with the photosynthetic process causing decreas- es, namely, in carboxylation efficiency, photosynthet- ic capacity and quantum yield, which might account for the reduced biomass production of affected plants (Atkinson et al., 1989; Ramalho et al., 1993). This ele- ment appears to be fundamental for the normal func- tioning of PS II and for the attainment of high rates of water oxidation (Ghanotakis and Yocum, 1990).
Very little attention has been given to the effects of external calcium concentrations on coffee plants, although low calcium levels are common in highly weathered tropical soils (Bell et al., 1989). Among the most important factors limiting the production of cof- fee in Brazil are the acidity and the calcium content of the soil (Matiello, 1985). In 1985 it was estimated that at least 60% of plantations in Brazil were estab- lished in acid soils, poor in calcium and magnesium. In such conditions, application of lime improved pro- duction up to 200% (Matiello, 1985). That is due to the importance of calcium, since it is known that this element is quantitatively the third element absorbed by coffee plants, corresponding to up to 12% of total macronutrients in the plant.
In this work the influence of calcium on the biomass production of young coffee plants was studied, with emphasis on the mineral composition of the leaf and on correlations between calcium concentrations and some photosynthetic parameters.
Material and methods
Growth conditions
Plants of Coffea arabica var. Catuai were grown (between 1991 and 1993) in a complete nutrient solu- tion (Hoagland and Snyder, 1933) with iron given as sodium ethylenediaminetetraacetate (Na-EDTA), and pH around 6.0. Irradiance of 120-140 #mol m -2 s - I at the top of plant canopy was provided by mercury vapour pressure lamps in a growth room with 60-70% RH and 26/18°C (day/night). Different Ca treatments were imposed to 1.5 years old plants (12 to 14 pairs of leaves, before the reproductive stage) by modifying the concentration of Ca(NO3)2 in the nutrient solution:
5, 2.5, 0.1, 0.01 and 0 mM (severe deficiency). The treatments lasted for 91 days.
Growth analysis
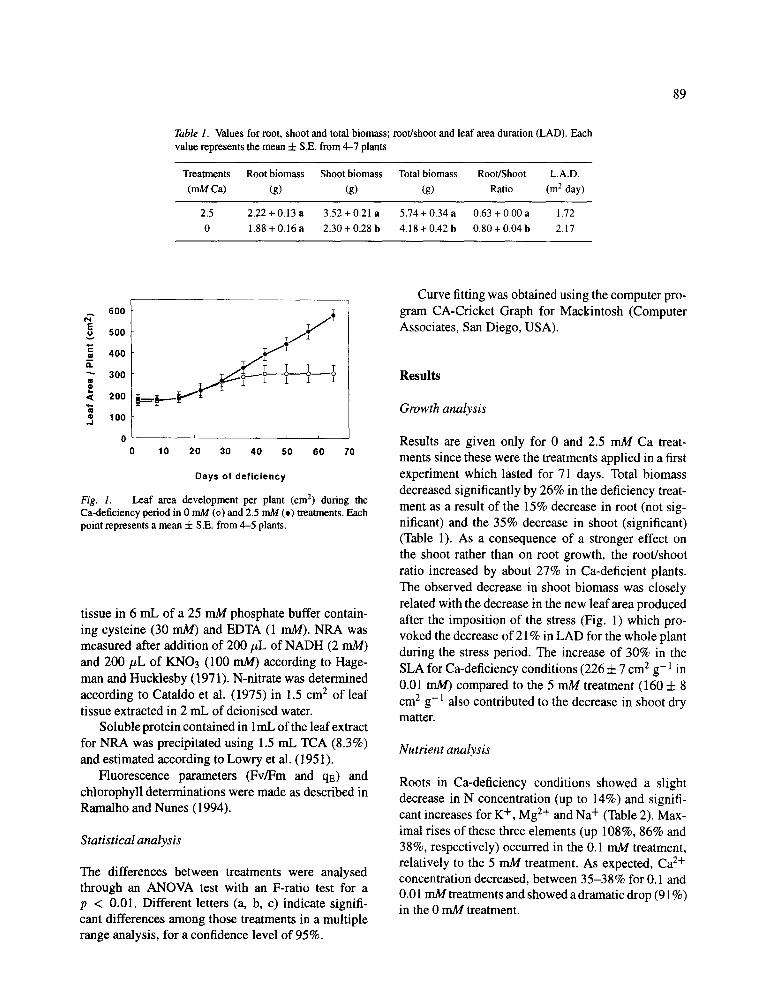
Leaf area was monitored during the deficiency period measuring the length and width of each leaf and using a linear regression to calculate the real leaf area (Y = 0.639X - 2.016, r 2 = 0.99, where Y is the effec- tive leaf area and X is the product of the measured length and width of each leaf). A destructive analysis was done after 71 days of Ca-treatment for shoot, root and total biomass. Leaf area duration (LAD) was cal- culated by the integration of the area below the line for each treatment shown in Figure 1. These measure- ments were performed only for the 0 mM and 2.5 mM Ca treatments.
Nutrient analysis
The concentration of several mineral nutrients was determined in the top pair of mature leaves and in roots.
Total nitrogen concentration was determined in 100 mg of dry material by the micro Kjeldahl method. Digestion was performed in 3 mL of H2SO4. A mixture of potassium sulphate and red mercuric oxide (1:10) was used as catalyst, and salicylic acid was included to reduce nitrate.
For the determination of P, K, Ca, Mg and Na, 3 mL of concentrated nitric acid were added to 200 mg of plant tissue and that mixture submitted to 30 min/60°C plus 1 h/150°C. Concentrated perchloric acid (1.5 mL) was then added and the sample submitted to 1 h/220°C. Phosphorus was evaluated by the method of vanadate- molybdate (Johnson and Ulrich, 1959; Chapman and Pratt, 1961), using a Unicam SP8 400 spectropho- tometer (Pye Unicam, Cambridge, U.K.) for colori- metric readings at 420 nm. K and Na concentrations were obtained by photometric emission, using a pho- tometer FLM3 (FLM, Copenhagen, Denmark) with a flame of propane and lithium as a standard. For Mg and Ca, atomic absorption spectrophotometry was used, in a spectrophotometer Philips PU 9100 (Pye Unicam, Cambridge, U.K.) and strontium chloride to release the elements.
Biochemical analysis
For determination of in vitro nitrate reductase activity (NRA) a leaf extract was prepared with 0.5 g fresh
89
Table 1. Values for root, shoot and total biomass; root/shoot and leaf area duration (LAD). Each value represents the mean + S,E. from 4-7 plants
Treatments Root biomass Shoot biomass Total biomass Root/Shoot L.A.D. (mM Ca) (g) (g) (g) Ratio (m 2 day)
2.5 2.22 +0.13 a 3.52 +0.21 a 5.74+ 0.34 a 0.63 + 0.00a 1.72
0 1.88 +0.16a 2.30 +0.28 b 4.18 + 0.42 b 0.80 + 0.04 b 2.17
600 P4 E o 500
¢= == 400
f t .
300
< 200
100 . J
o
0 10 20 30 40 50 60 70
Days of def ic iency
Fig. 1. Leaf area development per plant (cm 1) during the Ca-deficiency period in 0 mM (o) and 2.5 mM (e) treatments. Each point represents a mean + S.E. from 4-5 plants.
tissue in 6 mL of a 25 mM phosphate buffer contain- ing cysteine (30 mM) and EDTA (1 mM). NRA was measured after addition of 200 #L of NADH (2 mM) and 200/zL of KNO3 (100 mM) according to Hage- man and Hucklesby (1971). N-nitrate was determined according to Cataldo et al. (1975) in 1.5 cm 2 of leaf tissue extracted in 2 mL of deionised water.
Soluble protein contained in 1 mL of the leaf extract for NRA was precipitated using 1.5 mL TCA (8.3%) and estimated according to Lowry et al. (1951).
Fluorescence parameters (Fv/Fm and qE) and chlorophyll determinations were made as described in Ramalho and Nunes (1994).
Statistical analysis
The differences between treatments were analysed through an ANOVA test with an F-ratio test for a p < 0.01. Different letters (a, b, c) indicate signifi- cant differences among those treatments in a multiple range analysis, for a confidence level of 95%.
Curve fitting was obtained using the computer pro- gram CA-Cricket Graph for Mackintosh (Computer Associates, San Diego, USA).
Results
Growth analysis
Results are given only for 0 and 2.5 mM Ca treat- ments since these were the treatments applied in a first experiment which lasted for 71 days. Total biomass decreased significantly by 26% in the deficiency treat- ment as a result of the 15% decrease in root (not sig- nificant) and the 35% decrease in shoot (significant) (Table 1). As a consequence of a stronger effect on the shoot rather than on root growth, the root/shoot ratio increased by about 27% in Ca-deficient plants. The observed decrease in shoot biomass was closely related with the decrease in the new leaf area produced after the imposition of the stress (Fig. 1) which pro- voked the decrease of 21% in LAD for the whole plant during the stress period. The increase of 30% in the SLA for Ca-deficiency conditions (226 + 7 cm 2 g- 1 in 0.01 mM) compared to the 5 mM treatment (160 -4- 8 cm e g- 1 also contributed to the decrease in shoot dry matter.
Nutrient analysis
Roots in Ca-deficiency conditions showed a slight decrease in N concentration (up to 14%) and signifi- cant increases for K +, Mg 2+ and Na + (Table 2). Max- imal rises of these three elements (up 108%, 86% and 38%, respectively) occurred in the 0.1 mM treatment, relatively to the 5 mM treatment. As expected, Ca 2+ concentration decreased, between 35-38% for 0.1 and 0.01 mM treatments and showed a dramatic drop (91%) in the 0 mM treatment.
90
Table 2. Foliar concentrations (mg g - 1 dry weight) of several nutrients in the youngest mature leaves 91 days after the imposition of Ca-deficiency. Each value represents a mean 4- S.E. from 3-5 leaves or roots from different plants.
Part of Tremment N P K Ca Mg Na
~eplant (mMCa)
~ a f 0 - 2 . 0 + 0 . 0 ~ 31.6+1.6a 1.7+0.9c 6 .3+0 .3b
0.01 48.8+0.5b 1.5+0.3b - 1 .8+0.2c 6 .0+0 .4b 8 .6+1 .4b
0.1 45.7+1.2b 2 . 0 + 0 . 8 ~ 3 5 . 6 + 0 . 7 ~ 2 .0+0.3c 5 .7+0 .2b 9 .3+0 .5b
2.5 - 2 .2+0 .1a 32.2+2.2a 14.1+0.6b 3 .2+0 .0a -
5 37.6+0.4a 1 . 8 + 0 . 4 ~ 27.0+0.7a 11.2+0.4a - 1 .6+0.1a
Roots 0 - 1 .8+0.1a 22.8+1.3a 3 .9+0 .2b 9 .0+0 .6b -
0.01 31.9+1.1b - 20.9+2.9a 3 0 . 1 + 1 . 5 ~ 8 .9+0.5b 1.6+0.0c
0.1 3 3 . 5 + 1 . 3 ~ - 24.2+1.0a 28.6+2.1c 9 .7+0 .5b 1 .8+0.0b
2.5 - 1 .7+0.1a 23.1+1.6a 15.1+4.2b 8 .0+0.4b -
5 37.3+0.4a - 11.6+0.4b 46 .0+1.6a 5 .2+0 .3a 1 .3+0.1a
Table 3. Leaf concentration (mg g - l dry weight) of several elements, in conditions of Ca deficiency, adequate and excess to coffee plants (data from Amorim et al., 1968; Carvajal, 1984; Malavolta, 1963, 1993 and Matiello, 1985).
N P K Ca Mg
Deficiency < 20 < 0.5-0.9 < 0.5-10 < 5-8 < 1.0-1.5
Low 20-25 0.9-1.2 10-18 8-11 1.6-2.5
Adequate 25-30 1.2-2.0 18-20 11-15 2.5-4.0
High 30--40 2.0-3.5 20-25 15-17 4.0-5.0
Excess > 40 > 3.5 > 25 > 17 > 5.0
However, the major changes provoked by Ca- deficiency occurred in the leaf tissue. Nitrogen con- centrations increased by 21% (0.1 mM) and 30% (0.01 mM), while Mg 2+ showed a strong increase of 78% for the 0.1 mM treatment and 97% for 0 mM treat- ment (Table 2). K + concentration of the leaf tissue also changed with Ca-deficiency having a 32% max- imal significant increase for the 0.1 mM treatment at the end of the stress period. However, it was Na + that showed the stronger change, with an increase high- er than 4-fold for 0.1 and 0.01 mM treatments. Ca 2+ contents also denote significant changes in the deficien- cy treatments with decreases between 82-85%. Criti- cal values of the main mineral nutrients were already described for young coffee plants. Those results from other authors (Table 3) were compiled for a better and easier comparison with our own results.
Phosphorus was the only element that did not show any change either in root or leaf tissues.
Some relationships between the studied elements were considered, since they can contribute to a bet- ter diagnostic analysis of the nutrient status of coffee plants. From data presented on Table 2 were calculat- ed significant relations between the main cations (K +, Ca 2+ and Mg 2+) and also between two other important elements (N and P), which are expressed in Table 4. Apart from the obvious changes in the ratios where cal- cium is present (there is always an increase in the pro- portion of the other elements), we may conclude from the K:Mg ratio that Mg 2+ showed a higher increase than K +. Concerning the N:P ratio, we observed a signifcant increase of 36% in the Ca-deficient plants when 5 and 0.01 mM treatments are compared, but only a small increase of 8% when comparing 5 with 0.1 mM (Table 4). Those increases were a consequence

91
Table 4. Ratios between mineral elements in coffee leaves calculated from the mean values expressed in Table 2
Treatments Ca:Mg K : C a K:Mg N:P
(mM Ca)
0 0.27:1 18.6:1 5:1 -
0.01 0.30:1 - - 32.5:1
0.1 0.35:1 17.8:1 6.2:1 22.8:1
2.5 4.41:1 2.28:1 10.1:1 -
5 - 2.41:1 - 20.9:1
Table 5. Results of soluble protein, nitrate reductase activity and nitrate content in coffee leaves after 91 days of Ca-deficiency. Each value repre- sents the mean -4- S.E. from 3-6 leaves of different plants
Treatments Soluble Protein NRA NO 3
(mM Ca) (/~g cm -2) (nmol m -2 s - l ) (rag g - l d.w.)
5 817.2 + 73.9 a 66.9+21.1 a 3.89+ 1.08a
0.I 433.0+ 21.5 b 68.5+ 15.3a 4.76+ 0.12a
0.01 461.3 +31.1 b 65.5 + 8.9a 5.30+ 0.9 a
of the rise in leaf N-concentration in defciency condi- tions associated with quite stable P concentrations in all the treatments.
Biochemical analysis
The results of some biochemical determinations are presented in Table 5. Protein concentrations showed significant decreases of 48% (0.1 mM) and 44% (0.01 mM) after 91 days of Ca-deficiency.
NRA in leaves was not affected during the defi- ciency period (Table 5). Ca-deficiency caused a non- significant increase in NO 3 concentration in leaves (22 and 36%, respectively for 0.1 and 0.01 mM Ca treatments).
Correlation of Ca 2+ with photosynthetic parameters
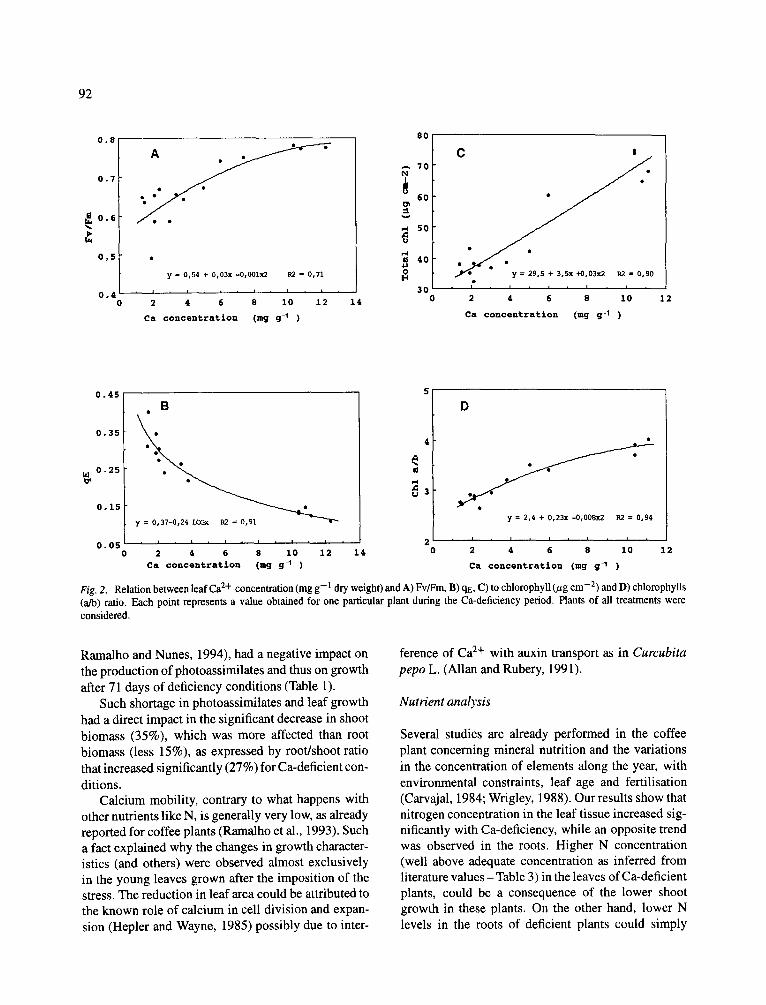
Several correlations between Ca 2+ leaf concentration and some photosynthetic parameters were established, to explore possible relations of this ion with compo- nents of the photosynthetic process (Fig. 2). The corre- lations obtained were quite good. Relating Ca 2+ levels with the photochemical efficiency, given here by the
Fv/Fm parameter, a positive correlation of r 2 = 0.71 was obtained (Fig. 2A). Concerning the effect of Ca- deficiency on the loss of energy in the form of heat (given by qE), we observed a very good correlation of energy loss with calcium depletion (r 2 = 0.91) (Fig. 2B). Very good positive correlations of calcium con- centration with the concentration of total chlorophyll (r 2 = 0.90) and with the ratio between chlorophylls a and b (r 2 --- 0.94) (Figs. 2C and D) were also obtained.
Discussion
Growth analysis
Calcium deficiency affected growth, predominantly that of new leaves. Reduction of leaf expansion and chlorosis (which became very severe with time and evolved to necrosis in extreme conditions of stress) were observed especially in the 2nd and 3rd pairs of leaves grown under deficiency conditions. The decreases in unit leaf area and LAD (which decreased the intercepted light during the stress period) and the impairment in photosynthetic performance (see
92
0.8
0.7
~ 0.6
0,5
y = 0,54 + 0,03x -0,001x2 R2 = 0,71
0 . 4 ' ~ ' ' ' 0 2 4 6 8 10 12 14
Ca concentration (mg g-1 )
8 0
7O ('4
6O
v
5 0
~ 40 x l
3 0
C $
I O I I I I
2 4 6 8 10
Ca concentration (mg g-1 )
12
L~
0 . 4 5
0.35
0.25
0.15
4
B
y = 0,37-0,24 LOGx ~ = 0,91
I I I I I I
2 4 6 8 i0 12
Ca concentration (mg g4 )
D
J .
y = 2,4 + 0,23x -0,008x2 R2 = 0,94
0 . 0 5 2 ' ~ ~ ' ' 0 1 4 0 2 4 6 8 1 0 1 2
Ca c o n c e n t r a t i o n ( m g g - 1 )
Fig. 2. Relation between leaf Ca 2+ concentration (mg g- 1 dry weight) and A) Fv/Fm, B) qE, C) to chlorophyll (/zg cm -2) and D) chlorophylls (a/b) ratio. Each point represents a value obtained for one particular plant during the Ca-deficiency period. Plants of all treatments were considered.
Ramalho and Nunes, 1994), had a negative impact on the production of photoassimilates and thus on growth after 71 days of deficiency conditions (Table 1).
Such shortage in photoassimilates and leaf growth had a direct impact in the significant decrease in shoot biomass (35%), which was more affected than root biomass (less 15%), as expressed by root/shoot ratio that increased significantly (27%) for Ca-deficient con- ditions.
Calcium mobility, contrary to what happens with other nutrients like N, is generally very low, as already reported for coffee plants (Ramalho et al., 1993). Such a fact explained why the changes in growth character- istics (and others) were observed almost exclusively in the young leaves grown after the imposition of the stress. The reduction in leaf area could be attributed to the known role of calcium in cell division and expan- sion (Hepler and Wayne, 1985) possibly due to inter-
ference of Ca 2+ with auxin transport as in Curcubita pepo L. (Allan and Rubery, 1991).
Nutrient analysis
Several studies are already performed in the coffee plant concerning mineral nutrition and the variations in the concentration of elements along the year, with environmental constraints, leaf age and fertilisation (Carvajal, 1984; Wrigley, 1988). Our results show that nitrogen concentration in the leaf tissue increased sig- nificantly with Ca-deficiency, while an opposite trend was observed in the roots. Higher N concentration (well above adequate concentration as inferred from literature values- Table 3) in the leaves of Ca-deficient plants, could be a consequence of the lower shoot growth in these plants. On the other hand, lower N levels in the roots of deficient plants could simply

result from a lower root efficiency for N-absorption. In the coffee leaves, Carvajal (1984) observed that the deficiency of Ca 2+ provoked a little decrease in the N concentration. Also Fenn and Taylor (1990) and Fenn et al. (1991) found (for radish and onion, respectively) that an increase in Ca 2+ concentration in the nutrient solution increased the N concentration in the leaves and decreased it in the roots, which is the opposite of our results.
Major ions can influence each other's absorption by competitive interactions or by affecting ion selectivity of membranes. Examples of these kinds of effect are the excess Na + that can induce Ca 2+ and/or K + defi- ciencies and the excess Ca 2+ that can induce Mg 2+ deficiency (Grattan and Grieves, 1993). Our results show that under Ca-deficiency conditions Mg 2+ and Na + increased significantly in the leaf tissue. Also K + shows a slight tendency to increase. These increases may represent a compensation to maintain the elec- trical and chemical balance of the cell, since Ca 2+, Mg 2+, K + (and also Na +) are cations that may substi- tute for each other in case of lack or excess of one of them (Iyengar and Reddy, 1993).
Na + showed the strongest increase of more than 4- fold for 0.1 and 0.01 mM Ca treatments. Other works (Hawkins and Lewis, 1993a; Jacoby, 1993) showed that Ca 2+ (and also K +) decreased Na + influx to the plant in adverse saline conditions, i.e. Ca 2+ and Na + presented a negative relationship, which seems to agree with our results, where the decrease in Ca 2+ was asso- ciated with a strong increase in Na + levels. In the above referred saline conditions additional Ca 2+ also improved biomass production and partially restored NO 3 uptake (Hawkins and Lewis, 1993a, b). Ca 2+ has an important role in the maintenance of membrane integrity (Alam, 1993) and its absence cause a loss of selectivity and other problems in the plasmatic mem- branes, which could allow the increase of Na + in the cell. The excessive increase in this element could then increase membrane porosity and contribute to mem- brane depolarisation (Hawkins and Lewis, 1993a).
Mg 2+ is usually considered as a partial substitute of Ca 2+ when this one is present in low levels. So, the observed significant increase to almost the double of the leaf concentration of Mg 2+ (the only studied biva- lent cation) in the Ca-deficient treatments seems under- standable. Mg 2+ showed a rise from adequate levels to concentrations that in normal conditions would be considered as in excess, a fact already observed in previous work (Ramalho et al., 1993).
93
K + also showed increases in leaf tissue, in compar- ison to the values for the 5 mM treatment. However, we must note that even the lowest leaf K + concentration was above the level considered as adequate (Table 3). The concentration in roots was lower than in shoots, without significant differences between the treatments, except for the 5 mM treatment where roots showed a significant lower level concomitantly to a significant higher Ca 2+ concentration. Roots from plants of the 5 mM treatment presented less K + and Mg 2+.
Phosphorus is the macronutrient required in lower amounts in coffee leaves (see Table 3) and its concen- tration was similar in both root and shoot. This ion did not show any significant change in Ca-deficiency con- ditions, an effect which only occurred with the other studied anion, NO 3 .
Despite the fact that no single ratio of nutrients can be validly used for diagnostic purposes of min- eral nutrition, some have been established to study the balance of nutrients in plants in general. Among these studied relations expressed as ratios are those of Table 4. According to Carvajal (1984) the relations K:Ca and K:Mg determined for well-nutritioned cof- fee plants were around, respectively, 3.1:1 and 14.3:1 which indicates that the leaves of plants from 0.1 and 0 mM treatments have imbalance problems. The changes in the ratio K:Ca confirms that K + increases relative- ly to Ca 2+, due both to the slight increase in K + and the strong decrease in Ca 2+. As already discussed, Mg 2+ and K + showed increases for the Ca-deficient treatments. The decrease in the ratio K:Mg shows, as we already suspected, that the increase in Mg 2+ was proportionally higher than that of K +.
For the deficient treatments the ratio Ca:Mg suf- fered a complete inversion, showing not only the Ca 2+ decrease but also the Mg 2+ increase as already observed elsewhere (Ramalho et al., 1993).
Another studied relation was the ratio N:P. Accord- ing to Wrigley (1988) this ratio could range from 24:1 (at low leaf P level of 0.06%) to 15:1 (at high leaf P level of 0.2% of the dry matter). In our experiment the obtained values of N:P were within optimal range both for 5 mM (20.9:1 ) and for 0.1 mM (22.8:1) treatments. That shows that even in mild Ca-deficiency conditions N and P seem to maintain their balance, despite the observed rise in N-levels. However, in a close obser- vation we can see that for the obtained P contents (around 0.2%) a ratio close to 15:1 could be expected. That did not happen, precisely due to the higher levels of N in the deficiency treatments. Only in the 0.01 mM treatment the strongest rise of N concentration cause a

94
rise in the N:P ratio, provoking a clear imbalance also between these two nutrients.
Biochemical determinations
Our data show that the soluble protein decreased signif- icantly due to Ca-deficiency. That decrease is frequent in several severe stress conditions as a consequence of altered N-metabolism and reduced rates of protein synthesis (Rabe, 1993). On the other hand, NRA which is related to the assimilation of nitrogen did not show significant differences between all the treatments. In fact, Ca-deficiency did not affect NRA, which does not support for our coffee plants the possible role of Ca 2+ as activator for NR as suggested for coffee (Carvajal, 1984) and also for Amaranthus (Sane et al., 1987). Since NRA did not change, the significant decrease observed in protein content in Ca-deficient plants could result from the lower rates of photosynthesis (Ramal- ho and Nunes, 1994) which induces shortage in energy molecules and C-chain molecules to accept the NH +. In agreement with this, a tendency for accumulation of N-nitrate occurred in the Ca-deficient leaves.
Correlation Ca 2+ vs. photosynthetic parameters
Ca 2+ is very important to the plant metabolism in gen- eral and for photosynthetic processes in particular. It has been suggested that the presence of Ca 2+ has a dual function. One Ca 2+ ion bound to PS II is involved in the oxygen evolution, while a second Ca 2+ ion has a structural role in the peripheral antenna assembly (Katoh and Han, 1993). The stabilisation of chloro- phyll and apoprotein from LHC II (Tanaka et al., 1992), the existence of high rates of oxygen evolution and a better yield production of energy molecules from the /kpH (Chiang and Diley, 1989) are some parts of a global effect of calcium in the photosynthetic process. As already observed in coffee plants, Ca-deficiency induces impairment of photosynthesis, both at stomatal and mesophyllic levels (Ramalho et al., 1993; Ramal- ho and Nunes, 1994). The present work shows evi- dence concerning the role of Ca 2+ at the mesophyll level. In fact, Ca 2+ concentration showed a reason- able (r 2 = 0.71) positive correlation with Fv/Fm, i.e. a gradual decrease in the photochemical efficiency of the PS II accompanied the decrease in Ca 2+ leaf concen- trations. We suggest that such a loss in the photochem- ical efficiency is linked to the lower 02 evolution rates observed in Ca-deficiency conditions (Ghanotakis and Yocum, 1990; Ramalho and Nunes, 1994).
A good correlation (r 2 = 0.91) established with qE shows that a probable rise in ApH occurs with lower levels of Ca 2+. The rise in the intrathylacoidal pro- ton gradient indicates both the existence of electron transport after the PS II (responsable for its build-up) and a relative decrease in the utilisation of that pro- ton gradient. Thus the impairement of photosynthesis does not occur only at PS II level. The decrease in the utilisation of the ApH could be due to a decrease in ATPase activity or in the use of ATP, e.g. for carboxyla- tion. However, previous results (Ramalho and Nunes, 1994) showed that Rubisco activity was not sensitive to Ca-deficiency. So we suggest that the build-up of the ApH is a result of a decrease in the activity of the ATPase.
It is known that when leaves receive more energy that they can use for photosynthesis a high proton gra- dient builds up exceeding the requirements for current ATP-synthesis, which provokes an increase in high energy quenching and, possibly, inactivation of PS II (Krause and Weis, 1991). In these circumstances, Krieger and Weis (1993) observed that the capacity of oxygen evolution decreases with the increase in ApH, especially when pH of thylacoid lumen goes below 5.5. Furthermore the inactivation of oxygen evolution was accompanied by release of Ca 2+ ions at PS II level. Those authors postulated that Ca 2+ could be related to high energy quenching (qE) and also with zeaxanthin formation as a way of controlling trapping and flux of energy in the antennae, and also of electron transport in PS II. The presence of Ca 2+ will then provoke a decrease in photoinactivation by the control of redox balance at PS II level (Krieger and Weis, 1993).
Those results are consistent with our own results concerning qE increase with Ca 2+ decrease, which is also accompanied by decreases in photochemical effi- ciency and photosynthetic capacity measured by oxy- gen evolution (Ramalho and Nunes, 1994).
Good correlations were also found both for the total chlorophyll content (r 2 -- 0.90) and for the ratio Chl (a/b) (r 2 = 0.94), reflecting the important role of Ca 2+ in the maintenance ofChl (Ramalho and Nunes, 1994) probably by acting in the stabilization and aggregation of Chl-apoprotein molecules in the antennae complex (Katoh and Han, 1993; Tanaka et al., 1992). Calcium levels seem to correlate better with the preferential loss of Chl a as expressed by the ratio Chl (a/b), suggesting the possibility of a selective photobleaching of Chl a from P S I (Miller and Carpentier, 1991; Williams et al., 1986). Such reduction in Chl concentration in Ca-deficiency conditions had surely contributed to the

observed loss in photochemical efficiency and low 02 evolution at PS II level.
Despite the gradual trend of these Ca-correlations, it seems that at least until Ca 2+ leaf concentrations of 1% there were no significant modifications of the stud- ied parameters. We think that the studied effects should be considered in a global view, since it is known that Ca-deficiency affects membrane stabilisation which would affect general cell metabolism, not only by the lack of Ca 2+ but also by the imbalance of other ions.
Conclusions
Due to its low capacity to move from old to young developing parts of the plant, Ca-deficient plants showed significant changes in the structures devel- oped after the imposition of the deficiency conditions, with reduced growth and modifications on the rela- tion root/shoot. Leaf expansion was depressed severe- ly, reducing the size of the photosynthetic active area. Moreover, mesophyllic components of photosynthesis were affected, namely at the levels of PS II and eventu- ally PS I and ATPase activity. The loss of chlorophylls in Ca-deficient leaves may have been a very important component in the decrease of photochemical efficiency by PS II.
Changes in mineral concentrations indicates that calcium deficiency had a strong overall effect on tissue concentration of other elements, changing the ionic balance of the cell. In this way, Ca-deficiency stress will induce also an imbalance stress, apparent in some nutrient ratios. These modifications will probably be related to changes in general metabolism and in photo- synthetic processes, both at stomata and mesophyllic levels.
Acknowledgements
The authors wish to thank Eng Maria Jos6 Silva (Cen- tro de Estudos de Produq[to e Tecnologia Agrfcolas - Instituto de Investiga~o Cientifica Tropical) for sta- tistical assistance, Prof Hans Lambers (Dept. Plant Ecology and Evolutionary Biology - Utrecht Univ.) for his critical comments on the manuscript and E A N - Dept. of Plant Physiology for the experimental facilities provided.
95
References
Alam S M 1993 Nutrient uptake by plants under stress conditions. In Handbook of Plant and Crop Stress. Ed. M Pessarakli. pp 227-246. Marcel Dekker Inc., New York.
Allan A C and Rubery P H 1991 Calcium deficiency and auxin transport in Curcubita pepo L. seedlings. Planta 183,604-612.
Amorim H V, Scoton L C, Haag H P and Malavolta E 1968 Estudos sobre a alimentaq~o mineral no cafeeiro. XX. Efeito da variaq~o do pH no desenvolvirnento e composiqAo quimica do cafeeiro (Coffea arabica L. var. Mundo Novo) cultivado em solu~o nutritiva. An. Esc. Super. Agr~ria Lufz Queiroz (Piracicaba) 25, 121-135.
Atkinson C J, Mansfield T A, Kean A M and Davies W J 1989 Control of stomatal aperture by calcium in isolated epidermal tissue and whole leaves of Commelina communis L. New Phytol. 111,9-17.
Bell R W, Edwards D G and Asher C J 1989 Effects of calcium supply on uptake of calcium and selected mineral nutrients by tropical food legumes in solution culture. Aust. J. Agric. Res. 40, 1003-1013.
Carvajal J 17 1984 Cafeto - Cultivo y Fertilization. Instituto Intema- cional de la Potassa, Berna.
Cataldo D A, Haroon M, Schrader L E and Youngs V L 1975 Rapid colorimetfic determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 6, 71-80.
Chapman H D and Pratt P F 1961 Methods of analysis for soils, plants and waters. Univ. of California, Division of Agricultural Sciences.
Chiang G G and Dilley R A 1989 Ca ++ regulation of proton fluxes in chloroplasts. The 8 KDa CF0 subunit is a Ca ++ -controlled H + flux gating struture. Physiol. Plant. 76, A160.
Fenn L B and Taylor R M 1990 Calcium stimulation of ammonium absorption in radish. Agron. J. 82, 81-84.
Fenn L B, Taylor R M, Binzel M L and Burks C M 1991 Calcium stimulation of ammonium absorption in onion. Agron. J. 83, 840-843.
Funkhouser E A, Cairney J, Chang S, Dilip M A, Dias L, Newton R J and Artlip T S 1993 Cellular and molecular responses to water deficit stress in woody plants. In Handbook of Plant and Crop Stress. Ed. M Pessarakli. pp 347-362. Marcel Dekker Inc., New York.
Ghanotakis D F and Yocum C F 1990 Photosystem II and the oxygen- evolving complex. Annu. Rev. Plant Physiol. Plant Mol. Biol. 41,255-276.
Grattan S R and Grieves C M 1993 Mineral nutrient acquisition and response by plants grown in saline environments. In Handbook of Plant and Crop Stress. Ed. M Pessarakli. pp 203-226. Marcel Dekker Inc., New York.
Hageman R H and Hucklesby D P 1971 Nitrate Reductase from Higher Plants. Methods Enzymol. XXIII, 491-503.
Hawkins H-J and Lewis O A 1993a Combination effect of NaC1 salinity, nitrogen form and calcium concentration on the growth, ionic content and gaseous exchange properties of Triticum aes- tivum L. cv. Gamtoos. New Phytol. 124, 161-170.
Hawkins H-J and Lewis O A 1993b Effect of NaC1 salinity, nitrogen form, calcium and potassium concentration on nitrogen uptake and kinetics in Triticum aestivum L. cv. Gamtoos. New Phytol. 124, 171-177.
Hepler P K and Wayne R O 1985 Calcium and plant development. Ann. Rev. Plant Physiol. 36, 397--439.
Hoagland D R and Snyder W C 1933 Nutrition of strawberry plant under controlled conditions. Proc. Am. Soc. Hort. Sci. 30, 288.

96
Iyengar E R and Reddy M P 1993 Crop response to salt stress: seawater application and prospects. In Handbook of Plant and Crop Stress. Ed. M Pessarakli. pp 183-201. Marcel Dekker Inc., New York.
Jacoby B 1993 Mechanisms involved in salt tolerance by plants. In Handbook of Plant and Crop Stress. Ed. M Pessarakli. pp 97-123. Marcel Dekker Inc., New York.
Jaunin F and Hofer R M 1988 Calcium and rhizodermal differentia- tion in primary maize roots. J. Exp. Bot. 39, 587-593.
Johnson C M and Ulrich A 1959 Analytical methods for use in plant sciences. Bull. Calif. Agric. Exp. St. no. 799.
Katoh S and Han K-C 1993 Different localization of two Ca 2+ in spinach oxygen-evolving photosystem II membranes. Evidence for involvement of only one Ca 2+ in oxygen evolution. Plant Cell Physiol. 34, 585-593.
Krause G H and Weis E 1991 Chlorophyll fluorescence and photo- synthesis: The basics. Ann. Rev. Plant Physiol. Plant Mol. Biol. 42, 313-349.
Krieger A and Weis E 1993 The role of calcium in the pH-dependent control of photosystem II. Photosynth. Res. 37, 117-130.
Lowry O H, Rosebrough N J, Farr A L and Randall R J 1951 Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193, 265-275.
Malavolta E 1963 Bothnica e melhoramento. In Cultura e Aduba~o do Cafeeiro. pp 143-190. Instituto Brasileiro da Potassa, S~o Paulo.
Maiavolta E 1993 Nutri~o mineral e aduba~o do cafeeiro. Col- heitas econ6micas m(tximas. Editora Agron6mica Ceres, S~o Paulo,
Matiello J 1985 Cultura de Caf6 no Brasil - Manual de Recomenda~es. Instituto Brasileiro do Caf6 - G.E.R.C.A., Rio de Janeiro.
Miller N and Carpentier R 1991 Energy dissipation and photopro- tection mechanisms during chlorophyll photobleaching in thy- lacoid membranes. Photochem. Photobiol. 54, 465-472.
Moncrief N, Kretsinger R and Goodman M 1990 Evolution of EF- hand calcium-modulated proteins. I. Relationships based on amino acid sequences. J. Mol. Evol. 30, 522-562.
Rabe E 1993 Altered nitrogen metabolism under environmental stress conditions. In Handbook of Plant and Crop Stress. Ed. M Pessarakli. pp 261-276. Marcel Dekker Inc., New York.
Ramalho J C and Nunes M A 1994 Effects of calcium deficiency in Coffea arabica photosynthesis. Gas exchanges and fluorescence analysis. (In preparation).
Ramalho J C, Rebelo M C and Nunes M A 1993 Efeitos da defi- ci~ncia de e~ilcio em algumas caracteristicas fotossint6ticas e de crescimento em Coffea arabica. Garcia de Orta 19. (In press).
Rengel Z 1992 Role of calcium in aluminium toxicity. New Phytol. 121,499-513.
Roberts D M and Harmon A C 1992 Calcium-modulated proteins: targets of intracellular calcium signals in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43,375-414.
Sane P V, Kumar N, Singh K K and Kochhar V K 1987 Activation of nitrate reductase by calcium and calmodulin. Phytochem. 26, 1289-1291.
Tanaka A, Tanaka Y and Tsuji H 1992 Calcium-induced accumu- lation of light-harvesting chlorophyll a/b-protein complex. In Research in Photosynthesis. Ed. N Murata. Vol. I, pp 299-302. Kluwer Academic Publishers, Dordrecht.
Williams W P, Sen A and Fork D C 1986 Selective photobleaching of PS I-related chlorophylls in the heat-stressed pea chloroplasts. Photosynth. Res. 10, 75-92.
Wrigley G 1988 Coffee. Tropical Agriculture Series. Longman Sci- entific and Technical, Singapore.
Section editor: H Lambers




















