
Inter Simple Sequence Repeat (ISSR) Analysis of Diploid Coffee Species and Cultivated Coffea arabica...
10
-1 Inter simple sequence repeat (ISSR) analysis of diploid coffee species and cultivated Coffea arabica L. from Tanzania Linus I. Masumbuko 1,2, * and Tomas Bryngelsson 2 1 Mikocheni Agricultural Research Institute, Box 6226 Dar es Salaam; 2 The Swedish University of Agricul- tural Sciences, Department of Crop Science, P.O. Box 44, SE 230 53 Alnarp. Sweden; *Author for correspondence (e-mail: [email protected]) Received 29 September 2003; accepted in revised form 9 April 2004 Key words: Coffea arabica, Cluster analysis, Diversity, Molecular markers Abstract Inter simple sequence repeat (ISSR) markers were used to evaluate levels of genetic similarity among Coffea arabica L. accessions from Tanzania and to estimate levels of genetic similarities in C. arabica and diploid coffee species. The six ISSR primers used generated a total of 82 fragments and the dissimilarity values ranged from 0.21 to 1. Mean dissimilarity values between provenances (0.56–0.85) were higher than within provenances (0.37–0.68). Cluster analysis based on Nei’s genetic distances showed C. arabica provenances grouping based on geographical origin. Two major clusters were formed that constituted of provenances from Kilimanjaro and Arusha in one sub-cluster; Tanga and Morogoro in the other; the second cluster had Mbeya provenances and diploid species, respectively. The implication is that Mbeya provenances are different from the rest of Tanzanian C. arabica. A principal coordinate analysis (PCA), whose first three coordinates explained 43% of the variation, showed similar groupings as in the cluster analysis. A separate cluster analysis of diploid species showed a distinct separation of the three species used. ISSR data gave results similar to previous findings from random amplified polymorphic DNA(RAPD) analysis. The results also confirm the limited diversity present in cultivated C. arabica in Tanzania. Introduction Coffea arabica L. and Coffea canephora Pierre are the two most important beverage species in the Rubiaceae family. While the former is self-fertile, the latter is self-incompatible. The primary centre of genetic diversity for C. arabica is the highlands of southwestern Ethiopia and the Boma Plateau of Sudan (Lashermes et al. 1999). There are also re- ports of C. arabica populations on the Imatong and Marsabit mountains in Sudan and Kenya, respectively (Berthaud and Charrier 1988). The first C. arabica introduction to Tanzania was in the late 19th century by Catholic missionaries. After abortive attempts to establish it in Baga- moyo and later on Mandera mission stations, success was finally achieved at Kilema on the slopes of Mount Kilimanjaro, because of the favourable climate for coffee growing (Kieran 1966). The history of the spread of coffee cultivation, gave rise to two major base populations, which were later described as distinct varieties. C. arabica var. arabica (syn. C. arabica var. typica Cramer) originated from progenies of a single plant from Indonesia, which were grown in Paris and Genetic Resources and Crop Evolution (2006) 53: 357–366 ȑ Springer 2006 DOI 10.1007/s10722-004-0577-x
Transcript of Inter Simple Sequence Repeat (ISSR) Analysis of Diploid Coffee Species and Cultivated Coffea arabica...

-1
Inter simple sequence repeat (ISSR) analysis of diploid coffee species and
cultivated Coffea arabica L. from Tanzania
Linus I. Masumbuko1,2,* and Tomas Bryngelsson21Mikocheni Agricultural Research Institute, Box 6226 Dar es Salaam; 2The Swedish University of Agricul-tural Sciences, Department of Crop Science, P.O. Box 44, SE 230 53 Alnarp. Sweden; *Author forcorrespondence (e-mail: [email protected])
Received 29 September 2003; accepted in revised form 9 April 2004
Key words: Coffea arabica, Cluster analysis, Diversity, Molecular markers
Abstract
Inter simple sequence repeat (ISSR) markers were used to evaluate levels of genetic similarity among Coffeaarabica L. accessions from Tanzania and to estimate levels of genetic similarities in C. arabica and diploidcoffee species. The six ISSR primers used generated a total of 82 fragments and the dissimilarity valuesranged from 0.21 to 1. Mean dissimilarity values between provenances (0.56–0.85) were higher than withinprovenances (0.37–0.68). Cluster analysis based on Nei’s genetic distances showed C. arabica provenancesgrouping based on geographical origin. Two major clusters were formed that constituted of provenancesfrom Kilimanjaro and Arusha in one sub-cluster; Tanga and Morogoro in the other; the second cluster hadMbeya provenances and diploid species, respectively. The implication is that Mbeya provenances aredifferent from the rest of Tanzanian C. arabica. A principal coordinate analysis (PCA), whose first threecoordinates explained 43% of the variation, showed similar groupings as in the cluster analysis. A separatecluster analysis of diploid species showed a distinct separation of the three species used. ISSR data gaveresults similar to previous findings from random amplified polymorphic DNA(RAPD) analysis. The resultsalso confirm the limited diversity present in cultivated C. arabica in Tanzania.
Introduction
Coffea arabica L. and Coffea canephora Pierre arethe two most important beverage species in theRubiaceae family. While the former is self-fertile,the latter is self-incompatible. The primary centreof genetic diversity for C. arabica is the highlandsof southwestern Ethiopia and the Boma Plateau ofSudan (Lashermes et al. 1999). There are also re-ports of C. arabica populations on the Imatongand Marsabit mountains in Sudan and Kenya,respectively (Berthaud and Charrier 1988). Thefirst C. arabica introduction to Tanzania was in
the late 19th century by Catholic missionaries.After abortive attempts to establish it in Baga-moyo and later on Mandera mission stations,success was finally achieved at Kilema on theslopes of Mount Kilimanjaro, because of thefavourable climate for coffee growing (Kieran1966).
The history of the spread of coffee cultivation,gave rise to two major base populations, whichwere later described as distinct varieties. C. arabicavar. arabica (syn. C. arabica var. typica Cramer)originated from progenies of a single plant fromIndonesia, which were grown in Paris and
Genetic Resources and Crop Evolution (2006) 53: 357–366 � Springer 2006
DOI 10.1007/s10722-004-0577-x

Amsterdam Botanic Gardens from whence theyspread to Latin America (Chevalier and Dagron1928; Carvalho 1946). C. arabica var. bourbon(B. Rodr.) Choussy, is the other variety, whichoriginated from Yemen and spread to the rest ofthe world via Bourbon Island, now Reunion(Haarer 1956). The two varieties are commonlycalled Typica and Bourbon, respectively (Kruget al. 1939; Carvalho et al. 1969).
Tanzania has had coffee introductions from thecentre of coffee genetic diversity, Ethiopia, andfrom both Typica and Bourbon base populations.These were all C. arabica, the only tetraploid(2n = 4x = 44) in the genus Coffea that alsohappens to be self-fertile. The country also hasseveral native diploid coffee species, e.g. C. zan-guebariae Lour., C. eugenioides Moore, C. mufin-diensis Bridson and C. canephora Pierre. Thedistribution of diploid coffee species in Tanzaniahas not been fully documented. C. zanguebariaehowever, grows at low altitude, it has been docu-mented in Zanzibar Island (Purseglove 1981).Samples have been gathered in the Coast region atUtete and Mkongo, the east side of Morogororegion (Kitulangalo) and in Uzigua which liesalong the Dar es Salaam-Tanga road. C. mufindi-ensis has been found on the highlands i.e. over1600 m., in Mamiwa and Mufindi, Iringa region(Berthaud et al. 1982). C. eugenoides grows on thewestern part of Tanzania and C. canephora in thenorthwest.
However, C. arabica is the species of economicimportance, which is cultivated at altitudes of1000–2000 m above sea level, in the northernhighlands (Arusha and Kilimanjaro regions) andsouthern highlands (Morogoro, Mbeya, Iringa,and Ruvuma regions). Emphasis in the coffee-breeding programme is now being directed todisease resistance, hence knowledge of geneticdiversity in coffee is critical for future coffeeimprovement. This has been necessitated by thesevere losses to coffee berry disease (CBD) andcoffee leaf rust (CLR), incurred by coffee farmers.
Molecular markers have been used to studydiversity in coffee. Orozco-Castillo et al. (1994)used random amplified polymorphic DNA(RAPD) markers to detect genetic diversity be-tween coffee species and between C. arabicagenotypes. The narrow genetic base of commercialcultivars of coffee was confirmed when cultivatedand sub-spontaneous accessions of C. arabica were
analysed by RAPD markers (Lashermes et al.1996). RAPD markers have also been used tostudy genetic diversity of Ethiopian C. arabica(Anthony et al. 2001; Aga et al. 2003). Masum-buko et al. (2003) used RAPD markers for analysisof Tanzanian cultivated C. arabica. The resultsshowed a uniform genetic distance, implyinglimited variability.
Using amplified fragment length polymorphism(AFLP) and simple sequence repeats (SSRs)markers, Anthony et al. (2002) were able to show asuccessive reduction of genetic diversity during thedissemination of coffee from its primary centre ofdiversity. AFLP and SSR markers were used toassess polymorphism between and within cultivarsfrom Typica and Bourbon, and relating these tofour Yemen cultivars and eleven sub-spontaneousaccessions from the primary centre of diversity.
This paper reports the use of inter-simple se-quence repeat (ISSR) markers, a PCR-basedmarker system that combines most of the advan-tages of AFLP (can detect several polymorphismsin a single assay and are as reproducible asrestriction fragment length polymorphism analy-sis) and microsatellite analysis (they are robust andcan be exchanged between laboratories and theirdata is highly informative) with the universality ofRAPD (Zietkiewics et al. 1994; Kantety et al.1995). Although ISSRs segregate mostly as domi-nant markers following Mendelian inheritance(Gupta et al. 1994; Tsumura et al. 1996; Ratna-parkhe et al. 1998; Wang et al. 1998), they havealso been shown to segregate as co-dominantmarkers in some cases thus enabling distinctionbetween homozygotes and heterozygotes (Wuet al. 1994; Akagi et al. 1996; Wang et al 1998;Sankar and Moore 2001).
Single microsatellite primers, usually 16–25 bplong are used in ISSR polymerase chain reaction(PCR). The microsatellite repeats in primers canbe di-nucleotides, tri-nucleotides, tetra-nucleotidesor penta-nucleotides. The technique amplifies aDNA segment present at an amplifiable distancebetween two identical microsatellite repeat regionsoriented in opposite direction. Primers can be ei-ther unanchored (Meyer et al. 1993; Wu et al.1994; Gupta et al. 1994) or more usually anchoredat the 3¢ or 5¢ end with 1–4 degenerate bases ex-tended into the flanking sequences (Zietkiewicset al. 1994). ISSR markers have been used suc-cessfully for cultivar identification in Olea euro-
358

paea L. and Cicer arietinum L. (Antonella et al.2001; Chowdhury et al. 2002). ISSR markers wereused to determine genetic relationships in thegenus Cicer, in Sesamum indicum L., diversity inOryza sativa L. and genetic mapping in Citrus(Parsons et al. 1997; Sankar and Moore 2001;Iruela et al. 2002; Kim et al. 2002).
Utilising the same accessions used in the RAPDstudy by Masumbuko et al. (2003), the objectivesof this research were (1) to evaluate geneticdiversity among Tanzanian C. arabica accessionsusing ISSR markers (2) to estimate levels of ge-netic similarity between C. arabica and diploidcoffee species; and among the diploid coffee spe-cies; C. eugenoides, C. zanguiberae and C. mufin-diensis. (3) to complement previous resultsobtained from RAPD analysis.
Materials and methods
Plant material
A total of 100 accessions of Coffea arabica L. fromKilimanjaro, Arusha, Tanga, Mbeya and Moro-goro regions (Figure 1) and 10 accessions of diploidcoffee were used in this study (Table 1). C. arabicaleaf samples were collected along a transect acrosseach farmat every 100–400 m, depending on the sizeof the farm. Samples of diploid coffee species com-prising two accessions from Coffea eugenoides, fivefrom Coffea zanguiberae and three from Coffeamufindiensis, were obtained from the field gene bankat the Tanzania Coffee Research Institute, Ly-amungu. Every ten accessions of Coffea arabicarepresent one collection site.
DNA extraction
DNA was isolated from young leaves by a modi-fied CTAB procedure (Aga et al. 2003) and theDNA concentration was determined by fluorescentspectrophotometry (Masumbuko et al. 2003). Thequality of the DNA was examined electrophoreti-cally on a 1% agarose gel.
PCR amplification and electrophoresis
Nine ISSR primers were tested, six from the Bio-technology Laboratory, University of BritishColumbia (UBC), USA, and three synthesized by
DNA Technology A/S, Denmark. The primerswere screened against three accessions from eachprovenance for polymorphism. Six primers thatshowed polymorphism and reproducibility werefinally selected for further analysis (Table 2).Amplification was carried out in 25 lL reactionscontaining 5 ng of genomic DNA, 800 lM dNTPs(Amersham Pharmacia Biotech, USA), 2 mMMgCl2, 2% formamide (v/v), 200 and 400 nM ofUBC and DNA Technology A/S primers, respec-tively, 1x PCR buffer (75 mM Tris-HCl pH 8.8,20 mM (NH4)2 SO4, 0.01% (v/v) Tween 20) and 1unit Taq polymerase (ABgene House, Surrey,UK). Amplification was done using a GENE AmpPCR System 9700 thermocycler (Hitachi Ltd.Tokyo, Japan). The programme consisted of thefollowing steps: step 1: 94 �C for 1 min; step 2:94 �C for 1 min; step 3: ramp rate of 0.5 �C/sec-ond to 55 �C; step 4: 55 �C for 2 min; step 5: 72 �Cfor 0.5 min; step 6: ramp rate of 1.3 �C/second to94 �C; step 7: final extension at 72 �C for 5 min;with 45 cycles between steps 2 and 5.
For separating amplified products, a poly-acrylamide gel (CleanGel 48S, Pharmacia Bio-tech, USA), was rehydrated for one hour in25 mL of rehydrating buffer (0.112 M Tris–ace-tate, pH 6.4). Sample loading buffer consisted of20% sucrose (w/v), 10% ficoll (w/v), 0.005%bromophenol blue (w/v), 5 M urea and 1 mMEDTA. Five microliter of the loading buffer wasadded to PCR products before 6 lL of theresulting mixture was loaded onto the gel. Elec-trode strips were soaked with 25 mL of electrodebuffer (0.2 M Tris, pH 8.0; 0.2 M Tricine; 0.55%w/v SDS) and then placed at the top and bottomof the gel. The gel was run on a horizontalMultiphor II Electrophoresis Unit at 450 V anda current of 20 mA for 20 min and then at50 mA for 1.5 h. Separated PCR products werevisualized by silver staining the gel using aHoefer Automated Gel Stainer (Pharmacia Bio-tech, USA).
Data analysis
Polymorphic bands were scored manually as pres-ent ‘1’ or absent ‘0’. Data were analysed usingEXCEL, NTSYS and the MAPRF6-DDAT pro-gram developed by E. Ritter, Neiker, Apartado 46,01080 Vitoria, Spain. Ten accessions from each of
359

Figure 1. The five regions of Tanzania where coffee leaf samples were collected.
Table 1. Regional locations from where Coffea arabica leaf samples were collected.
Region District Location Number of samples
Kilimanjaro (KIL) Moshi Rural Kilema RC Parish (Ki) 10
Moshi Rural Kibohehe (Kb) 10
Moshi Rural Chombo (Ch) 10
Arusha (AR) Arumeru Tengeru Farm (Te) 10
Arumeru Nkoanenkoli (Nk) 10
Tanga (TAN) Lushoto Gare (Ga) 10
Lushoto Bazo (Ba) 10
Mbeya (MBY) Tukuyu Bugoba Masebe (Bg) 10
Ileje Ileje (Ij) 10
Morogoro (MOR) Morogoro Rural Luale (Lu) 10
Kilimanjaro (DpSp) Lyamungu Gene Bank 10a
Total 110
aDiploid coffee species: C. zanguiberae (Zan), C. mufindiensis (Muf) and C. eugenoides (Eug), 5, 3 and 2 accessions, respectively.
360

the 11 provenances (total of 110 accessions) wereused to generate a dissimilarity matrix based onJaccard’s (1908) coefficient. The matrix was analy-sed by unweighted pair-group method arithmeticaverages (UPGMA) method (Sneath and Sokal1973), of the NTSYS-PC version 2.10t, �AppliedBiostatics, Inc. USA to generate a dendrogram. Acophenetic matrix was computed, in order to mea-sure the goodness of fit, by comparing the obtainedtree matrix with the dissimilarity matrix. Furtheranalysis involved performing a principal coordinateanalysis (PCA) and finally a cluster analysis basedon Nei (1978) genetic distance. A similar analysiswas repeated for the diploid coffee species only.
Results
The six ISSR primers generated a total of 82fragments ranging in size from 200 to 3500 bp. An
example of banding pattern is shown in Figure 2.Dissimilarity values between all accessions rangedfrom 0.21 to 1. The overall mean dissimilarityvalue between-provenances 0.73, was higher thanthat of within-provenance 0.55 (available fromauthors upon request). Among the ten C. arabicaprovenances used in the analysis, Bazo and Garewere the most similar while Kilema and Ileje werethe most dissimilar with values of 0.56 and 0.85,respectively.
Cluster analysis based on dissimilarity values ofall accessions generated a dendrogram with fourclusters (not shown due to size of figure). The firstand second clusters had most accessions fromKilimanjaro (Kibohehe, Chombo and Kilemaprovenances) and Arusha (Tengeru and Nkoa-nenkoli provenances) regions, except for threeaccessions from Nkoanenkoli and one from Teng-eru that fell into the third cluster. The third clusterhad accessions from Tanga (Bazo and Gare prov-enances) and Morogoro (Luale provenance) whilethe fourth cluster had Mbeya (Bugoba and Ilejeprovenances) and diploid coffee accessions. Thetwo C. eugenoides accessions in this cluster werecloser to the C. arabica accessions compared toaccessions from the other diploid species. Com-parison of the cophenetic matrix to the dissimilaritymatrix gave a cophenetic correlation of 78%.
A PCA of the 110 accessions was performed.The plot of the first three coordinates explained
Table 2. ISSR primers used in the analysis of Tanzanian
Arabica coffee.
Primer Sequence (5¢ to 3¢)
855 (C-01) ACA CAC ACA CAC ACA CYT
888 (C-02) BDB CAC ACA CAC ACA CA
889 (C-03) DBD ACA CAC ACA CAC AC
B2669 (B-12) ACA CAC ACA CAC ACA CG
811 GAG AGA GAG AGA GAG AC
825 ACA CAC ACA CAC ACA CT
Figure 2. ISSR patterns of four Coffea arabica accessions generated by primer B12; Bp, 100 base pair ladder (arrow shows 800 bp);
Ga, Gare; Lu, Luale; Ij, Ileje; Bg, Bugoba.
361
43% of the variation observed. The results showeda similar grouping of the accessions as in thecluster analysis. There were three distinct group-ings. The first had Kilimanjaro and Arusha prov-enances (Kilema, Kibohehe, Chombo, Tengeruand Nkoanenkoli); the second had Tanga andMorogoro provenances (Bazo, Gare and Luale)and the last group was comprised of Mbeya(Bugoba and Ileje) and diploid coffee species.
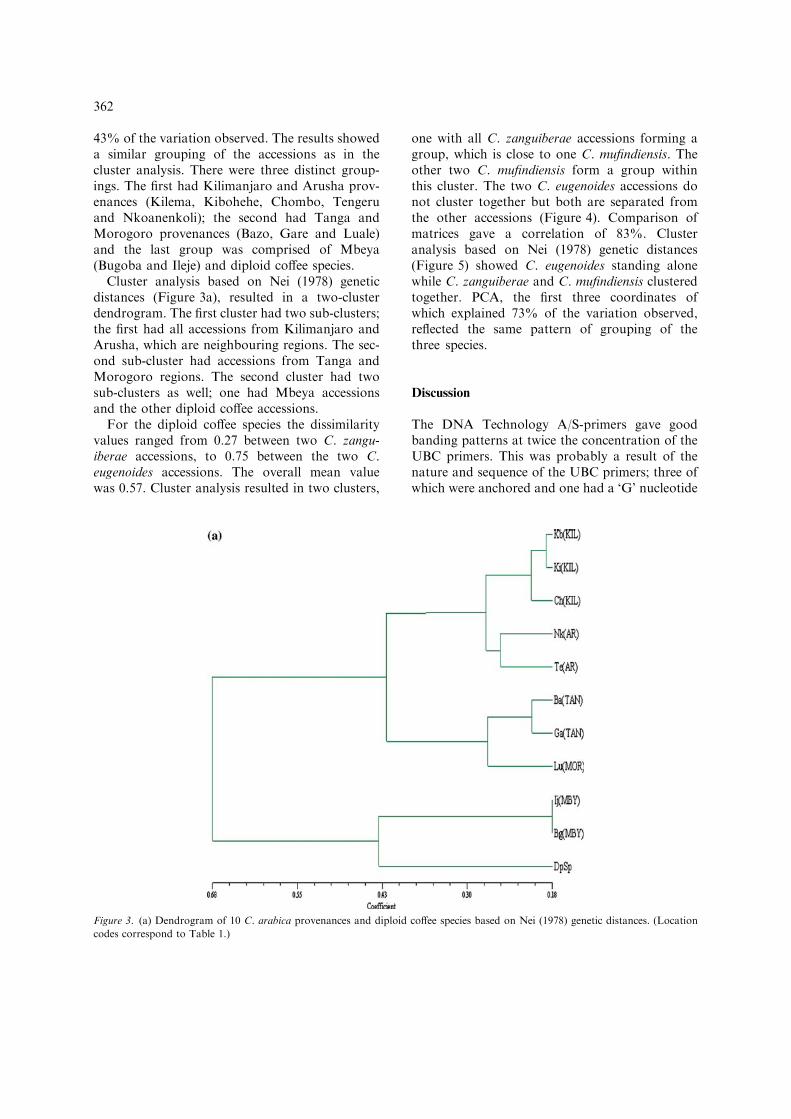
Cluster analysis based on Nei (1978) geneticdistances (Figure 3a), resulted in a two-clusterdendrogram. The first cluster had two sub-clusters;the first had all accessions from Kilimanjaro andArusha, which are neighbouring regions. The sec-ond sub-cluster had accessions from Tanga andMorogoro regions. The second cluster had twosub-clusters as well; one had Mbeya accessionsand the other diploid coffee accessions.
For the diploid coffee species the dissimilarityvalues ranged from 0.27 between two C. zangu-iberae accessions, to 0.75 between the two C.eugenoides accessions. The overall mean valuewas 0.57. Cluster analysis resulted in two clusters,
one with all C. zanguiberae accessions forming agroup, which is close to one C. mufindiensis. Theother two C. mufindiensis form a group withinthis cluster. The two C. eugenoides accessions donot cluster together but both are separated fromthe other accessions (Figure 4). Comparison ofmatrices gave a correlation of 83%. Clusteranalysis based on Nei (1978) genetic distances(Figure 5) showed C. eugenoides standing alonewhile C. zanguiberae and C. mufindiensis clusteredtogether. PCA, the first three coordinates ofwhich explained 73% of the variation observed,reflected the same pattern of grouping of thethree species.
Discussion
The DNA Technology A/S-primers gave goodbanding patterns at twice the concentration of theUBC primers. This was probably a result of thenature and sequence of the UBC primers; three ofwhich were anchored and one had a ‘G’ nucleotide
Figure 3. (a) Dendrogram of 10 C. arabica provenances and diploid coffee species based on Nei (1978) genetic distances. (Location
codes correspond to Table 1.)
362

at the 3¢ end. In unanchored primers where thesimple sequence repeats are used as primers, theprimer tends to slip within the repeat units duringamplification resulting in smears instead of clearbands.
The uniform within-provenance dissimilarityvalues show there is limited variability within thecultivated C. arabica provenances. The mean dis-similarity values between provenances 0.73 werehigher than within provenances 0.55 whichunderscores the low variability and the inbreedingnature of cultivated Arabica coffee. Lashermeset al. (1996), found similar results when comparingcultivated and wild accessions of coffee. A similarstudy by Masumbuko et al. (2002) using RAPDmarkers showed a similar trend.
Provenances from the same region clusteredtogether. Interviews with farmers during the sam-ple collection mission had shown that Bazo andBugoba accessions had originated from Gare andIleje, respectively; this was verified by the resultingdendrogram, where the provenances appear close
to each other. Mbeya accessions, which share thesame cluster with diploid coffee species, are iso-lated from other accessions from Kilimanjaro,Arusha, Tanga and Morogoro regions. Similarresults were obtained with RAPD analysis (Ma-sumbuko et al. 2002), although diploid specieswere not included in that study (Figure 3b). Thisshows Mbeya accessions to be different from therest of the Arabica coffee in Tanzania. Inciden-tally, Ileje provenance from Mbeya, had a within-provenance dissimilarity value of 0.68, which wasthe highest. Bazo and Ileje provenances had thehighest between-provenance means (0.78), becauseof the big differences with provenances fromKilimanjaro and Arusha (Kibohehe, Kilema,Chombo, Nkoanenkoli and Tengeru) whichranged from 0.82 to 0.85 (Table 2).
Cluster analysis of accessions from the diploidcoffee showed distinct separation of the threespecies. C. eugenoides that appeared closer toC. arabica accessions in the dendrogram consistingof all accessions is known to be one of its pro-
Figure 3. (b) RAPD analysis dendrogram of 16 Arabica coffee provenances from five regions of Tanzania based on Nei (1978)
distances. Kif, Kifumbu; Ke, Keiti; Zi, Ziwai; Ma, Maweni; Bw, Bwenda; Sh, Shiwanda.
363

genitors (Raina et al. 1998). The possibility of in-tercrossing C. arabica with diploid species (Bert-haud and Charrier 1988), offers an opportunity toobtain desirable genes from the diploid species.These results would have been more informative ifmore anchored ISSR primers were used and if eachof the diploid species were represented by several
accessions. Efforts should be made to start utilisingdiploid coffee species as a source of useful genes.
ISSR markers have shown their ability to iden-tify variation in coffee, in so doing it has comple-mented results obtained from a previous RAPDanalysis. It was also evident that Mbeya coffeeaccessions were different from the rest of Tanza-
Figure 4. Cluster analysis of diploid coffee species from dissimilarity matrix generated by Jaccard’s (1908) coefficient. Eug, eugenoides;
Zan, zanguibariae; Muf, mufindiensis.
Figure 5. UPGMA dendrogram of three diploid coffee species using Nei (1978) genetic distances. Eug, eugenoides; Zan, zanguibariae;
Muf, mufindiensis.
364

nian Arabica coffee; they could probably be ofinterest to the coffee improvement programme inTanzania.
Acknowledgements
The authors wish to thank Dr Enrique Ritter ofCentro de Investigacion y Mejora Agraria,Apartado 46 01080 Vitoria, Spain for availingthe use of statistical package DDAT, a compo-nent of MAPRF, in data analysis. We wouldalso like to thank Britt Green for introducing theISSR technique.
References
Aga E., Bryngelsson T., Bekele E. and Salomon B. 2003.
Genetic diversity of forest arabica coffee (Coffea arabica) in
Ethiopia as revealed by random amplified polymorphic DNA
(RAPD) analysis. Hereditas 138: 36–46.
Akagi H., Yokozeki Y., Inagaki A., Nakamura A. and Fu-
jimura T. 1996. A co-dominant DNA marker closely linked
to the rice nuclear restorer gene, Rf-1, identified with inter-
SSR fingerprinting. Genome 39: 1205–1209.
Anthony F., Bertrand B., Quiros O., Wilches A., Lashermes P.,
Berthaud J. and Charrier A. 2001. Genetic diversity of wild
coffee (Coffea arabica L.) using molecular markers. Euphy-
tica 118: 53–65.
Anthony F., Combes M.C., Astorga C., Bertrand B., Graziosi
G., Charrier A. and Lashermes P. 2002. The origin of culti-
vated Coffea arabica L. varieties revealed by AFLP and SSR
markers. Theor. Appl. Genet. 104: 894–900.
Antonella P., Caponio F. and Blanco A. 2001. Inter-simple
sequence repeat DNA markers for identification of drupes
from different Olea europaea L. cultivars. Eur. Food Res.
Technol. 213: 240–243.
Berthaud J. and Charrier A. 1988. Genetic resources of Coffea.
In: Lashermes P., Combes M-C., Robert J., Trouslot P.,
D’Hont A., Anthony F. and Charrier A. (eds), 1999.
Molecular Characterisation and Origin of the Coffea arabica
L. Genome. Mol. Gen. Genet. 261: 259–266.
Berthaud J., Anthony F. and Lourd M. 1982. Wild coffee trees
in Tanzania: Results of a prospecting mission. (March 6–
April 11, 1982).
Carvalho A. 1946. Distribuicao geografica e classificacao
botanica do genero Coffea com referencia especial a especie
Arabica. In: Anthony F., Bertrand B., Quiros O., Wilches A.,
Lashermes P., Berthaud J. and Charrier A. (eds), 2001.
Genetic Diversity of Wild Coffee (Coffea arabica L.) Using
Molecular Markers. Euphytica 118: 53–65.
Carvalho A., Ferwerda F.P., Frahm-Leliveld J.A., Medina
P.M., Mendes A.J.T. and Monaco L.C. 1969. Coffee. In:
Anthony F., Bertrand B., Quiros O., Wilches A., Lashermes
P., Berthaud J. and Charrier A. (eds), 2001. Genetic Diversity
of Wild Coffee (Coffea arabica L.) Using Molecular Markers.
Euphytica 118: 53–65.
Chevalier A. and Dagron M. 1928. Recherches historiques sur
les debuts de la culture du cafeier en Amerique. Communi-
cations et Actes de l’Academie des Sciences Coloniales, Paris.
In: Anthony F., Bertrand B., Quiros O., Wilches A., La-
shermes P., Berthaud J. and Charrier A. (eds), 2001. Genetic
Diversity of Wild Coffee (Coffea arabica L.) Using Molecular
Markers. Euphytica 118: 53–65.
Chowdhury M.A., Vandenberg B. and Warkentin T. 2002.
Cultivar identification and genetic relationship among se-
lected breeding lines in chickpea (Cicer arietinum L.).
Euphytica 127: 317–325.
Gupta M., Chyi Y-S., Romero-Severson J. and Owen J.L. 1994.
Amplification of DNA markers from evolutionary diverse
regions using single primers of simple-sequence repeats.
Theor. Appl. Genet. 89: 998–1006.
Haarer A.E. 1956. Modern coffee production. In: Anthony
F., Bertrand B., Quiros O., Wilches A., Lashermes P.,
Berthaud J. and Charrier A. (eds), 2001. Genetic Diversity
of Wild Coffee (Coffea arabica L.) Using Molecular
Markers. Leonard Hill (books) Limited, London. Euphy-
tica 118: 53–65.
Iruela M., Rubio J., Cubero J.I., Gil J. and Millan T. 2002.
Phylogenetic analysis in the genus Cicer and cultivated
chickpea using RAPD and ISSR markers. Theor. Appl.
Genet. 104: 643–651.
Jaccard P. 1908. Nouvelles recherches sur la distribution florale.
Bull. Soc. Vaud. Sci. Nat. 44: 223–270.
Kantety R.V., Zeng X.P., Bennetzen J.L. and Zehr B.E. 1995.
Assessment of genetic diversity in dent and pop corn (Zea
mays L.) inbred lines using inter-simple sequence repeat
(ISSR) amplification. Mol. Breed. 1: 365–373.
Kieran J.A. 1966. The Holy Ghost Fathers and Coffee in East
Africa-Unpublished Report- University College Nairobi. In:
The Improvement of Arabic coffee in East Africa: With
particular reference to Tanzania.
Kim D.H., Zur G., Danin-Poleg Y., Lee S.W., Shim K.B.,
Kang C.W. and Kashi Y. 2002. Genetic relationships of
sesame germplasm collection as revealed by inter-simple se-
quence repeats. Plant Breed. 121: 259–262.
Krug C.A., Mendes J.E.T. and Carvalho A. 1939. Taxonomia
de Coffea arabica L. Boletim Tecnico no 62. In: Anthony F.,
Bertrand B., Quiros O., Wilches A., Lashermes P., Berthaud
J. and Charrier A. (eds), 2001. Genetic diversity of wild coffee
(Coffea arabica L.) using molecular markers.. Euphytica 118:
53–65.
Lashermes P., Combes M.-C., Robert J., Trouslot P., D’Hont
A., Anthony F. and Charrier A. 1999. Molecular charac-
terisation and origin of the Coffea arabica L. genome. Mol.
Gen. Genet. 261: 259–266.
Lashermes P., Trouslot P., Anthony F., Combes M.C. and
Charrier A. 1996. Genetic diversity for RAPD markers be-
tween cultivated and wild accessions of Coffea arabica.
Euphytica 87: 59–64.
Masumbuko L.I., Bryngelsson T., Mneney E.E. and Salomon
B. 2003. Genetic diversity in Tanzanian Arabica coffee using
random amplified polymorphic DNA (RAPD) markers.
Hereditas 139: 56–63.
Meyer W., Mitchell T.G., Freedman E.Z. and Vilgays R. 1993.
Hybridization probes for conventional DNA fingerprinting
used as single primers in the polymerase chain reaction to
365

distinguish strains of Cryptococcus neoformans. J. Clin.
Microbiol. 31: 2274–2280.
Nei M. 1978. Estimation of average heterozygosity and genetic
distance from a small number of individuals. Genetics 89:
583–590.
Orozco-Castillo C., Chalmers K.J., Waugh R. and Powell W.
1994. Detection of genetic diversity and selective gene intro-
gression in coffee using RAPD markers. Theor. Appl. Genet.
87: 934–940.
Parsons B.J., Newbury H.J., Jackson M.T. and Ford-Lloyd B.
1997. Contrasting genetic diversity relationships are revealed
in rice (Oryza sativa L.) using different marker types. Mol.
Breed. 3: 115–125.
Purseglove J.W. 1981. Tropical Crops: Dicotyledon. Longman
Singapore Publishers (Pte) Ltd, pp. 458–492.
Raina S.N., Mukai Y. and Yamamoto M. 1998. In situ hy-
bridisation identifies the diploid progenitor species of
C. arabica (Rubiaceae). Theor. Appl. Genet. 97: 1204–1209.
Ratnaparke M.B., Tekeoglu M. and Muehlbauer F.J. 1998.
Inter-simple sequence repeat (ISSR) polymorphisms are
useful for finding markers associated with disease resistance
gene clusters. Theor. Appl. Genet. 97: 515–519.
Sankar A.A. and Moore G.A. 2001. Evaluation of inter-simple
sequence repeat analysis for mapping in Citrus and extension
of the genetic linkage map. Theor. Appl. Genet. 102: 206–
214.
Sneath P.H.A. and Sokal R.R. 1973. Numerical Taxonomy.
W.H. Freeman & Co, San Francisco.
Tsumura Y., Ohba K. and Strauss S.H. 1996. Diversity and
inheritance of inter-simple sequence repeat polymorphisms in
Douglas-fir (Pseudotsuga menziesii) and sugi (Cryptomeria
japonica). Theor. Appl. Genet. 92: 40–45.
Wang G., Mahalingan R. and Knap H.T. 1998. (C-A) and
(G-A) anchored simple sequence repeats (ASSRs) generated
polymorphism in soybean, Glycine max (L.). Merr. Theor.
Appl. Genet. 96: 1086–1096.
Wu K., Jones R., Dannaeberger L. and Scolnik P.A. 1994.
Detection of microsatellite polymorphism without cloning.
Nucleic Acids Res. 22: 3257–3258.
Zietkiewics E., Rafalski A. and Labuda D. 1994. Genome
fingerprinting by simple sequence repeat (SSR)-anchored
polymerase chain reaction amplification. Genomics 20:
176–183.
366




















