Effect of wastewater colloids on membrane removal of antibiotic resistance genes
11
Effect of wastewater colloids on membrane removal of antibiotic resistance genes Maria V. Riquelme Breazeal, John T. Novak, Peter J. Vikesland, Amy Pruden* Via Department of Civil and Environmental Engineering, Virginia Tech, Blacksburg, VA 24061, USA article info Article history: Received 21 June 2012 Received in revised form 21 September 2012 Accepted 22 September 2012 Available online 4 October 2012 Keywords: Antibiotic resistance genes Wastewater colloids Microfiltration Ultrafiltration abstract Recent studies have demonstrated that wastewater treatment plants (WWTPs) signifi- cantly alter the magnitude and distribution of antibiotic resistance genes (ARGs) in receiving environments, indicating that wastewater treatment represents an important node for limiting ARG dissemination. This study examined the potential for membrane treatment of microconstituent ARGs and the effect of native wastewater colloids on the extent of their removal. Plasmids containing vanA (vancomycin) and bla TEM (b-lactam) ARGs were spiked into three representative WWTP effluents versus a control buffer and tracked by quantitative polymerase chain reaction through a cascade of microfiltration and ultrafiltration steps ranging from 0.45 mm to 1 kDa. Significant removal of ARGs was ach- ieved by membranes of 100 kDa and smaller, and presence of wastewater colloids resulted in enhanced removal by 10 kDa and 1 kDa membranes. ARG removal was observed to correlate significantly with the corresponding protein, polysaccharide, and total organic carbon colloidal fractions. Alumina membranes removed ARGs to a greater extent than polyvinylidene fluoride membranes of the same pore size (0.1 mm), but only in the presence of wastewater material. Control studies confirmed that membrane treatment was the primary mechanism of ARG removal, versus other potential sources of loss. This study suggests that advanced membrane treatment technology is promising for managing public health risks of ARGs in wastewater effluents and that removal may even be enhanced by colloids in real-world wastewaters. ª 2012 Elsevier Ltd. All rights reserved. 1. Introduction Antibiotic resistance and, particularly, multidrug resistance, is an increasingly critical problem affecting human health (Davies and Davies, 2010). Globally, antibiotic resistance has been re- ported to be widespread (Allen et al., 2010) and on the rise (Zhang et al., 2006). Although most attention has been focused on nosocomial (i.e., hospital acquired) infections, environmental factors are also important contributors to the transport and spread of antibiotic resistance in the community (Larson, 2007). Recent studies have demonstrated that wastewater treatment plants can significantly alter the magnitude and distribution of antibiotic resistance genes (ARGs) in receiving environments (Storteboom et al., 2010). Given that antibiotic resistant bacteria (ARB) and ARGs of clinical concern have been documented to arise from environmental sources (Martinez, 2008), there is growing interest in limiting dissemination pathways between humans and the environment (Baquero et al., 2008). Thus, wastewater treatment plants may represent an important node for limiting the spread of antibiotic resistance. The presence of low levels of ARB and ARGs in wastewater is inevitable and several studies have documented their * Corresponding author. Tel.: þ1 540 231 3980. E-mail addresses: [email protected] (M.V. Riquelme Breazeal), [email protected] (J.T. Novak), [email protected] (P.J. Vikesland), apruden@engr. colostate.edu, [email protected] (A. Pruden). Available online at www.sciencedirect.com journal homepage: www.elsevier.com/locate/watres water research 47 (2013) 130 e140 0043-1354/$ e see front matter ª 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.watres.2012.09.044
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Effect of wastewater colloids on membrane removal of antibiotic resistance genes

ww.sciencedirect.com
wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0
Available online at w
journal homepage: www.elsevier .com/locate /watres
Effect of wastewater colloids on membrane removal ofantibiotic resistance genes
Maria V. Riquelme Breazeal, John T. Novak, Peter J. Vikesland, Amy Pruden*
Via Department of Civil and Environmental Engineering, Virginia Tech, Blacksburg, VA 24061, USA
a r t i c l e i n f o
Article history:
Received 21 June 2012
Received in revised form
21 September 2012
Accepted 22 September 2012
Available online 4 October 2012
Keywords:
Antibiotic resistance genes
Wastewater colloids
Microfiltration
Ultrafiltration
* Corresponding author. Tel.: þ1 540 231 398E-mail addresses: [email protected] (M.V. Riq
colostate.edu, [email protected] (A. Pruden).0043-1354/$ e see front matter ª 2012 Elsevhttp://dx.doi.org/10.1016/j.watres.2012.09.044
a b s t r a c t
Recent studies have demonstrated that wastewater treatment plants (WWTPs) signifi-
cantly alter the magnitude and distribution of antibiotic resistance genes (ARGs) in
receiving environments, indicating that wastewater treatment represents an important
node for limiting ARG dissemination. This study examined the potential for membrane
treatment of microconstituent ARGs and the effect of native wastewater colloids on the
extent of their removal. Plasmids containing vanA (vancomycin) and blaTEM (b-lactam)
ARGs were spiked into three representative WWTP effluents versus a control buffer and
tracked by quantitative polymerase chain reaction through a cascade of microfiltration and
ultrafiltration steps ranging from 0.45 mm to 1 kDa. Significant removal of ARGs was ach-
ieved by membranes of 100 kDa and smaller, and presence of wastewater colloids resulted
in enhanced removal by 10 kDa and 1 kDa membranes. ARG removal was observed to
correlate significantly with the corresponding protein, polysaccharide, and total organic
carbon colloidal fractions. Alumina membranes removed ARGs to a greater extent than
polyvinylidene fluoride membranes of the same pore size (0.1 mm), but only in the presence
of wastewater material. Control studies confirmed that membrane treatment was the
primary mechanism of ARG removal, versus other potential sources of loss. This study
suggests that advanced membrane treatment technology is promising for managing public
health risks of ARGs in wastewater effluents and that removal may even be enhanced by
colloids in real-world wastewaters.
ª 2012 Elsevier Ltd. All rights reserved.
1. Introduction plants can significantly alter the magnitude and distribution of
Antibiotic resistance and, particularly, multidrug resistance, is
an increasingly critical problemaffecting humanhealth (Davies
and Davies, 2010). Globally, antibiotic resistance has been re-
ported tobewidespread(Allenetal., 2010) andontherise (Zhang
et al., 2006). Although most attention has been focused on
nosocomial (i.e., hospital acquired) infections, environmental
factors are also important contributors to the transport and
spread of antibiotic resistance in the community (Larson, 2007).
Recent studies have demonstrated that wastewater treatment
0.uelme Breazeal), jtnov@v
ier Ltd. All rights reserve
antibiotic resistance genes (ARGs) in receiving environments
(Storteboom et al., 2010). Given that antibiotic resistant bacteria
(ARB) and ARGs of clinical concern have been documented to
arise from environmental sources (Martinez, 2008), there is
growing interest in limiting dissemination pathways between
humans and the environment (Baquero et al., 2008). Thus,
wastewater treatment plantsmay represent an important node
for limiting the spread of antibiotic resistance.
The presence of low levels of ARB and ARGs in wastewater
is inevitable and several studies have documented their
t.edu (J.T. Novak), [email protected] (P.J. Vikesland), apruden@engr.
d.

wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0 131
proliferation and persistence in wastewater treatment plants
(Auerbach et al., 2007; Borjesson et al., 2010; Kim et al., 2010;
Szczepanowski et al., 2009; Zhang et al., 2009). Traditionally
the objective of disinfection following water or wastewater
treatment is to kill or inactivate bacterial cells. However, to
address the challenge of antibiotic resistance, DNA removal or
destruction is ideal prior to discharge, reuse, or other appli-
cations. This is because ARGs are typically carried on highly
transmissible DNA elements, such as plasmids and integrons,
which can remain functional and be assimilated by down-
stream bacteria. Because of this ability of ARGs to transcend
their bacterial host, ARGs themselves are considered to be the
primary “contaminant” of concern (Pruden et al., 2006). Once
released, ARGs can persist in the environment (Biyela et al.,
2004; Storteboom et al., 2010), and have even been observed
to establish and proliferate in drinking water biofilms
(Schwartz et al., 2003; Xi et al., 2009). Although the effect of
some disinfection techniques on the fate of ARGs has been
investigated in a few studies (Munir et al., 2011; Oncu et al.,
2011), the fate of these microconstituents after wastewater
disinfection and release is still largely unknown.
The fate of ARGs through membrane removal processes is
of particular interest as membranes are growing in popularity
for enhancing sustainability of water resources and address-
ing concerns for other emerging contaminants (Dolar et al.,
2009, 2012; Munir et al., 2011; Wintgens et al., 2004). The
present study is the first to our knowledge to examine the
potential formembrane removal of ARGs.Membrane filtration
processes are commonly used for the removal of a wide
variety of contaminants, but most would not be expected to
impart significant removal of DNA based on its molecular
properties. However, wastewater effluents are complex in
their chemistries and could significantly affect membrane
removal of DNA. Thus, the focus of this study was on the
potential for the advanced membrane treatment of ARGs
within real-world wastewater effluent matrices.
Of specific interest to this study was the effect of DNA
interactions with wastewater effluent colloidal particles. DNA
is well known to interact with clay minerals and various soil
colloidal particles (Cai et al., 2007, 2006a, 2006b; Crecchio and
Stotzky, 1998; Nguyen et al., 2010; Nguyen and Elimelech,
2007a,b; Romanowski et al., 1991; Saeki et al., 2010). Consid-
ering that significant amounts of suspended colloidal particles
are present in wastewaters, these interactions represent
a promising and unexploited approach for enhanced removal
of ARGs from wastewater. However, most of the available
studies concerning DNAecolloid interactions have been
carried out in simplified or artificial systems (Mitra et al., 2001;
Nguyen and Elimelech, 2007a,b; Romanowski et al., 1991);
thus, there is a need for studies that emphasize the behavior
and fate of DNA in situ. In addition, the relationship between
the removal of ARGecolloid complexes and membrane pore
size has not been investigated in natural systems.
The overall objective of this project was to investigate the
effect of wastewater colloids on the removal of ARGs by
membrane filtration. It was hypothesized that colloidal
interactions with DNA would enhance membrane removal of
ARGs. To test this hypothesis, plasmids containing vanA and
blaTEM ARGs were spiked into three representative WWTP
effluents and a control buffer, and subjected to a cascade of
microfiltration (MF) and ultrafiltration (UF) steps ranging from
0.45 mm to 1 kDa. The vanA and blaTEM ARGs encode resistance
to vancomycin and b-lactam antibiotics (e.g., ampicillin),
respectively, with the former used typically as a last-resort
antibiotic when other treatments fail. Other potential sour-
ces of DNA loss, such as degradation, were also monitored to
verify that observed ARG removal was due to the membrane
treatment and also to examine the potential for wastewater
colloids to protect ARGs from deoxyribonuclease (DNase),
a naturally-occurring DNA-degrading enzyme. The effect of
membrane composition (alumina versus polyvinylidene
fluoride) was compared for the 0.1 mm pore size. Building
a knowledge base of the effect of membrane treatment on
ARGs is critical for enabling comprehensive management
strategies for emerging contaminants inwastewater effluents.
2. Materials and methods
A cascade of MF and UF steps was used to compare the
removal of ARGs from colloid-containing wastewater effluent
versus colloid-free aqueous buffer samples. Effluents from
three representative WWTPs were employed in order to test
the occurrence of DNAecolloid interactions in a complex
environmental matrix, as well as the reproducibility of the
results. Tris buffer (pH 8) was used as colloid-free material in
order tomaintain DNA stability in the controls throughout the
experiment, as low salt concentrations could cause DNA
instability, while high salt concentrations can promote the
sorption of the genetic material to the glass containers. The
key characteristics of the WWTPs selected for this study and
effluent quality are summarized in Table 1. An additional
experiment was carried out to further explore the potential for
DNAecolloid interactions to protect ARGs from degradation
by DNase.
2.1. Plasmid solution preparation
The pCR� 4-TOPO� plasmids (Invitrogen, Carlsbad, CA) con-
taining the ARGs of interestwere used to spike the appropriate
test solutions. This plasmid contains blaTEM and was modified
to contain vanA. A 732 bp portion of the vanA gene was PCR-
amplified from vancomycin-resistant Enterococcus genomic
DNA using previously described primers (Dutka-Malen et al.,
1995). The 25 mL PCR amplification reaction contained 1�Master Taq reaction buffer (5 Prime, Gaithersburg, MD), 1�Taq Master PCR enhancer (5 Prime), 1.5 mM Mg2þ solution (5
Prime), 0.05 mM of each deoxynucleoside triphosphate
(Promega, Madison, WI), 0.2 mM of each primer, 1.75 U Taq
DNA polymerase (5 Prime), 1 mL template DNA, and autoclaved
nanopure water to a final volume of 25 mL. The thermal cycle
consisted of an initial 3 min denaturing step at 95 �C; followed
by 50 cycles of a 30 s denaturing step at 95 �C, a 30 s annealing
step at 54 �C, and a 30 s extension step at 72 �C; and a 7 min
final extension step at 72 �C. The vanA PCR productwas ligated
to the pCR� 4-TOPO� plasmid and cloned using the TOPO TA
cloning kit (Invitrogen), following the manufacturer’s
instructions. Plasmids were extracted using the Qiagen
Plasmid Mega kit (Qiagen, Valencia, CA) and quantified by
qPCR targeting blaTEM and vanA ARGs.

Table 1 e General WWTP and effluent characteristics and effluent sampling dates.
WWTP Capacity(MGD)
Influent composition Key processesb TSSb
(mg/L)pHb Otherb
(mg/L)Collection
date
A 6 Residential, industrial Conventional,a UV disinfection 7.7 7.1 DO (8.1)
BOD (19.6)
4/21/11
B 9 Residential, hospital,
industrial
Conventional,a chlorine disinfection 2 7 DO (9.5) 3/1/11
C 6 Residential, hospital,
high Na2SO4 industrial
Conventional,a nitrification,
chlorine disinfection
7 6.7 BOD (<5) 4/7/11
An additional sampling event occurred on 1/30/12 from all WWTPs, for DNase-degradation test.
a Conventional treatment: primary clarification, aeration, secondary clarification, and disinfection.
b WWTP and effluent characteristics obtained by phone from WWTP operators.
wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0132
2.2. WWTP effluent collection and initial filtration
Grab samples of final effluent were collected from each of the
test wastewater treatment facilities (WWTPs A, B, or C) at
most 6 h before the experiment was carried out. Effluent
collection occurred during cool (2e15 �C) working week
mornings, avoiding rain or snow events taking place during
the previous 24 h. The effluent was immediately transported
to the lab where it was kept at 4 �C prior to use. As
a pretreatment to eliminate particles above the colloid size
range, the effluent was vacuum-filtered through a 1.2 mm
pore-sizemixed cellulose estermembrane (Millipore, Billerica,
MA) using a sterilized Nalgene� polysulfone filter holder
(Thermo Fisher Scientific, Rochester, NY).
2.3. Cascading filtration experiments
Each of the WWTP effluents was subjected to cascading
filtration experiments and compared with appropriate
controls: (1) Autoclaved 10 mM Tris buffer pH 8 (blank); (2)
Autoclaved 10mMTris pH 8 spikedwith vanA gene-containing
plasmids to a final concentration of 1 � 107 to 1 � 108 plasmid
copies/mL based on blaTEM gene concentrations (no-colloid
control); (3) filtered WWTP effluent (background); and (4)
filtered WWTP effluent spiked with vanA gene-containing
plasmids to a final concentration of 1 � 107 to
1� 108 plasmid copies/mL based on blaTEM gene concentrations
(wastewater colloid treatment). Four 1000-mL flasks were fil-
led with 600 mL of corresponding solution and lightly agitated
for 4e5 h using a Burrell wrist action shaker (Burrell Scientific,
Pittsburg, PA) to enable DNA interactions with wastewater
colloids prior to filtration. 10-mL subsamples were collected
from each of the four flasks during this stage to test for
alternative sources of DNA loss.
Sequential cascading filtration steps were carried out for
each of the four flask contents. First, 500 mL were filtered
through the membrane with the largest pore size (0.45 mm-
pore size membrane). Next, 10-mL sub-samples of the filtrate
were pipetted into previously baked 30-mL Qorpak (Bridge-
ville, PA) clear graduated glass bottles and immediately frozen
at �80 �C for subsequent lyophilization and DNA extraction.
Filtrate sub-samples originating from the unspiked WWTP
effluent were also collected for colloid characterization and
stored at 4 �C. The remaining water/buffer filtrate was then
filtered through the subsequent membrane (0.1 mm-pore size).
The processwas repeated for each of the filtrates, applying the
membranes in order of decreasing pore size:
0.45 mm/ 0.1 mm/ 100 kDa/ 10 kDa/ 1 kDa. Tominimize
carryover, the flask contents were processed in the following
order: (1) buffer blank, (2) unspiked background wastewater,
(3) spiked no-colloid control buffer, (4) spiked wastewater
containing colloids. The latter was processed after the 4-h
period allotted for DNAecolloid interactions to occur.
MF was carried out using 0.45 mm and 0.1 mm pore size
Durapore hydrophilic polyvinylidene fluoride (PVDF)
membranes (Millipore, Billerica, MA), and a Nalgene� poly-
sulfone filter holder (Thermo Fisher Scientific, Rochester, NY).
To compare the effect of membrane type, Anodisc 47 alumina
0.1 mm pore size membranes (Whatman GmbH, Germany)
were compared to the Durapore membrane of the same pore
size. UF was carried out using 100 kDa, 10 kDa, and 1 kDa
MWCO Ultracel regenerated cellulose membranes (Millipore,
Billerica, MA) and an 8200 Ultrafiltration Stirred Cell (Milli-
pore). UF filtrates flowed into baked glass flasks shielded by
baked aluminum foil. Because the UF membranes were reus-
able, they were cleansed between usages using DNA Away
(Molecular Bio Products, San Diego, CA) followed by a thor-
ough rinsing step that included filtering water through the
membrane for at least 10 min. A list of the membrane types
and pore sizes is provided in Table S1.
2.4. DNA stability in pretreated wastewater and buffer
To determine the extent to which membrane filtration
accounted for removal of ARGs in the above experiments,
changes in gene concentrations in pretreated wastewater and
buffer were monitored over the time allotted for DNAecolloid
interactions prior to filtration. A 10-mL sample was collected
immediately after plasmid spiking, and 2e3 additional
samples were collected during the following 4e5 h. Gene
fractions at times t > 0 were calculated with respect to the
initial concentration measured at t ¼ 0 h.
2.5. DNase-degradation experiment
The potential for colloideDNA interactions to protect ARGs
from degradation was investigated using DNase-degradation
experiments. All three WWTP effluents were sampled on the

wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0 133
same day and compared in parallel to the colloid-free Tris
buffer control. 500mL of either Tris buffer (pH 8) or the filtered
effluents were collected in fluorinated HDPE Nalgene bottles
(Thermo Fisher Scientific, Rochester, NY) and spiked to
a concentration of 1� 107 plasmids/mL, and placed in a shaking
incubator at 125 rpm and room temperature for 4 h to allow
DNAecolloid interactions. Sub-samples were collected at 0, 2,
and 4 h to test for background DNA losses. After 4 h, 200 mL of
the contents of each bottle (i.e., buffer orWWTP effluent) were
split into two new bottles (100 mL each), one of which was
spiked with 10.2 U of RQ1 RNase-free DNase (Promega) and
subject to 2.5 mM MgCl2 and 0.5 mM CaCl2 at 37 �C, and the
other only subject to the same salt and temperature conditions
used to activate theDNase. The bottleswere incubated at 37 �Cand shaken at 125 rpm. This approach allowed direct
comparison of the effect of the DNase versus the background
salt and temperature conditions required to activate the
DNase. 10 mL sub-samples were collected after light manual
vortexing, immediately after salt and/or DNase addition (i.e.,
shortly after hour 4), and at 5 and 7 h from the beginning of the
experiment. All collected samples were immediately stored at
�80 �C for further analysis.
2.6. DNA concentration and extraction
All samples were concentrated by lyophilization using
a FreeZone Plus 2.5 L Benchtop Freeze Dry System (Labconco,
Kansas City, Missouri). Isolation of DNA originating from
WWTP effluent samples was carried out using the FastDNA
Spin Kit for Soil (MP Biomedicals, Solon, OH) as recommended
in the kit manual. Isolation of DNA originating from buffer
samples was carried out using the PowerClean DNA Cleanup
Kit (MoBio Laboratories, Carlsbad, CA).
2.7. DNA quantification by qPCR
Quantification of vanA and blaTEM was carried out using real-
time quantitative polymerase chain reaction (qPCR) on
a CFX96 Touch� Real-Time PCR Detection System (Bio-Rad,
Hercules, CA). 1:10 DNA extract dilutions were analyzed in
triplicate reactions containing 40 nM of each primer, 1�SsoFast� EvaGreen� Supermix (Bio-Rad), 1 mL template DNA,
and sterile nanopure to a final volume of 10 mL. The primers
used for amplification of the vanA gene were vanAstF
(Bockelmann et al., 2009) and vana3RP (Volkmann et al., 2004).
The blaTEM gene was amplified using previously described
primers (Bibbal et al., 2007). The temperature programs con-
sisted of a 95 �C initial denaturing step for 30 s; 40 cycles of
denaturing at 95 �C for 5 s, annealing and extension at 60 �Cfor 5 s, and an additional extension step at 72 �C for 5 s for
vanA amplifications only. A melt curve was incorporated in
every reaction to verify specificity by ramping the tempera-
ture from 65 �C to 95 �C by 0.5 �C every 5 s. A calibration curve
was constructed for each set of reactions using at least five
standards. The qPCR standards were prepared by serially
diluting M13 PCR products obtained from cloned positive
controls (vanA or blaTEM). Concentrations were determined by
gel densitometry and converted to gene copies using the
relationship described by Pei et al. (2006). A limit of detection
(LOD) was calculated for each gene by averaging gene
concentrations in 10 reaction blanks and multiplying that
value by 3. The background gene concentrations detected in
each analytical blank was subtracted from the measured
sample concentration. Each individual measurement greater
than the detection limit was then multiplied by the dilution
factor (10) and the extraction volume, and divided by the
volume of each sample in microliters (1 � 104 mL) to obtain
the final concentration in gene copies/mL of each sample.
The three analytical measurements per sample were then
averaged to obtain the estimated concentration for that
sample.
2.8. Colloid characterization
Non-purgeable organic carbon concentrations weremeasured
using the Shimadzu TOC-VCSN analyzer (Shimadzu Scientific
Instruments, Columbia, MD) and potassium hydrogen
phthalate (KHP) as the standard. Polysaccharide concentra-
tions were measured by the phenolesulfuric acid method
(Dubois et al., 1956) using dextrose as standard. Protein
concentrations were measured by the Lowry method (Lowry
et al., 1977) using bovine serum albumin (BSA) as the stan-
dard. Briefly, five reagents were prepared as followse Reagent
1: 143 mM NaOH and 270 mM Na2CO3; Reagent 2: 57 mM
CuSO4; Reagent 3: 124 mM Na-tartrate; Reagent 4: mix of
reagents 1, 2, and 3 at a ratio of 100:1:1 (made daily); and
Reagent 5 contained folin ciocalteus reagent diluted 5:6 with
distilled water. 1-mL samples were added by sequential
addition and mixing of Reagents 4 and 5, respectively. The
samples containing the reagents were incubated for 45 min at
room temperature before measuring the absorbance at
750 nm.
2.9. Statistical analyses
Statistical analyseswere performed using R statistics software
(www.r-project.org). Statistical differences between the
membrane-removal of ARGs in the presence versus the
absence of colloids were calculated using a two-way analysis
of variance (ANOVA). The two-way ANOVAwas applied to the
log of the filtrate gene fraction from spiked WWTP effluent or
spiked buffer as the dependent variable. The filtrate gene
fraction was calculated as C/C0, where C was the gene
concentration in the filtrate and C0 was the average gene
concentration in the unfiltered samples used to test for
degradation. The presence/absence of colloids and the
membrane pore size or type were independent variables. The
analyses were repeated individually for each of the genes
monitored. To test the significance of the DNA degradation in
the water or buffer samples prior to treatment (i.e., prior to
filtration or application of DNase-degrading conditions),
sampling times after the addition of the plasmid spike were
binned as A (1e2 h), B (2.5e3 h), and C (4e5 h). A one-way
ANOVA test was applied to the log of the gene fractions
present in the water or buffer samples for each bin. Statistical
significance was set at a ¼ 0.05 ( p < 0.05). Correlation analysis
between the filtrate fraction of each of the colloidal compo-
nents and the logarithm of the filtrate gene fraction was
conducted using R, applying a one-sided Pearson correlation
test.
wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0134
3. Results
3.1. Membrane removal of ARGs and effect ofwastewater colloids
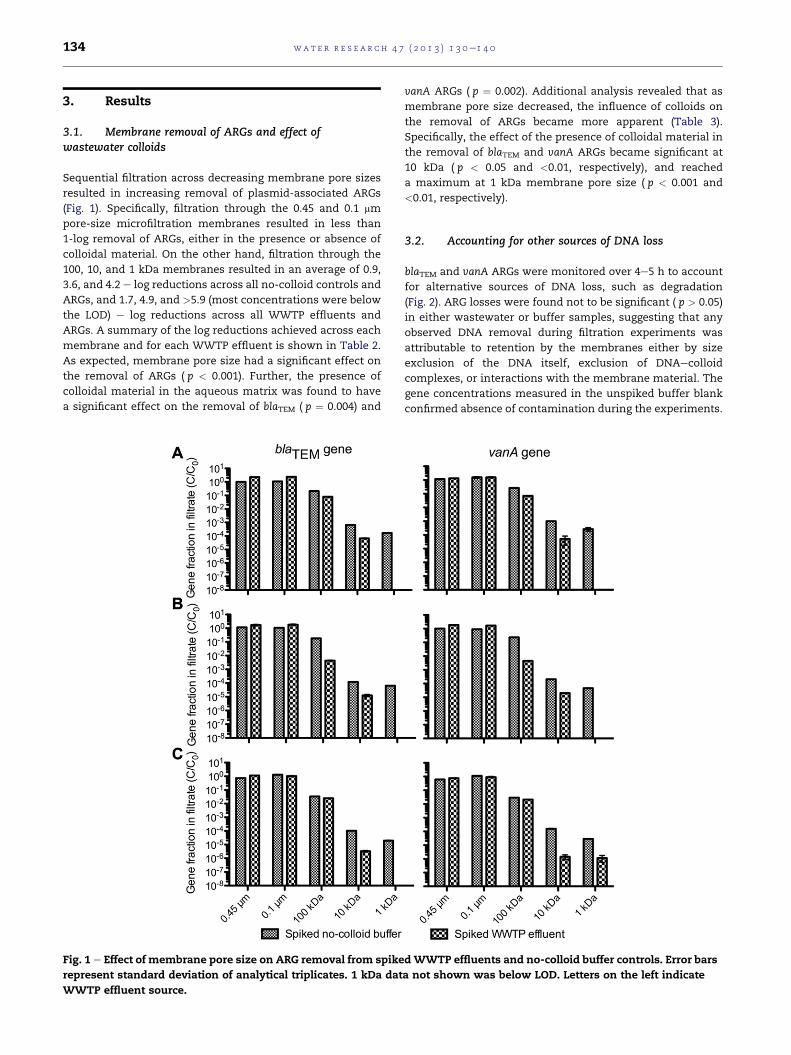
Sequential filtration across decreasing membrane pore sizes
resulted in increasing removal of plasmid-associated ARGs
(Fig. 1). Specifically, filtration through the 0.45 and 0.1 mm
pore-size microfiltration membranes resulted in less than
1-log removal of ARGs, either in the presence or absence of
colloidal material. On the other hand, filtration through the
100, 10, and 1 kDa membranes resulted in an average of 0.9,
3.6, and 4.2 e log reductions across all no-colloid controls and
ARGs, and 1.7, 4.9, and >5.9 (most concentrations were below
the LOD) e log reductions across all WWTP effluents and
ARGs. A summary of the log reductions achieved across each
membrane and for each WWTP effluent is shown in Table 2.
As expected, membrane pore size had a significant effect on
the removal of ARGs ( p < 0.001). Further, the presence of
colloidal material in the aqueous matrix was found to have
a significant effect on the removal of blaTEM ( p ¼ 0.004) and
Fig. 1 e Effect of membrane pore size on ARG removal from spike
represent standard deviation of analytical triplicates. 1 kDa dat
WWTP effluent source.
vanA ARGs ( p ¼ 0.002). Additional analysis revealed that as
membrane pore size decreased, the influence of colloids on
the removal of ARGs became more apparent (Table 3).
Specifically, the effect of the presence of colloidal material in
the removal of blaTEM and vanA ARGs became significant at
10 kDa ( p < 0.05 and <0.01, respectively), and reached
a maximum at 1 kDa membrane pore size ( p < 0.001 and
<0.01, respectively).
3.2. Accounting for other sources of DNA loss
blaTEM and vanA ARGs were monitored over 4e5 h to account
for alternative sources of DNA loss, such as degradation
(Fig. 2). ARG losses were found not to be significant ( p > 0.05)
in either wastewater or buffer samples, suggesting that any
observed DNA removal during filtration experiments was
attributable to retention by the membranes either by size
exclusion of the DNA itself, exclusion of DNAecolloid
complexes, or interactions with the membrane material. The
gene concentrations measured in the unspiked buffer blank
confirmed absence of contamination during the experiments.
dWWTP effluents and no-colloid buffer controls. Error bars
a not shown was below LOD. Letters on the left indicate
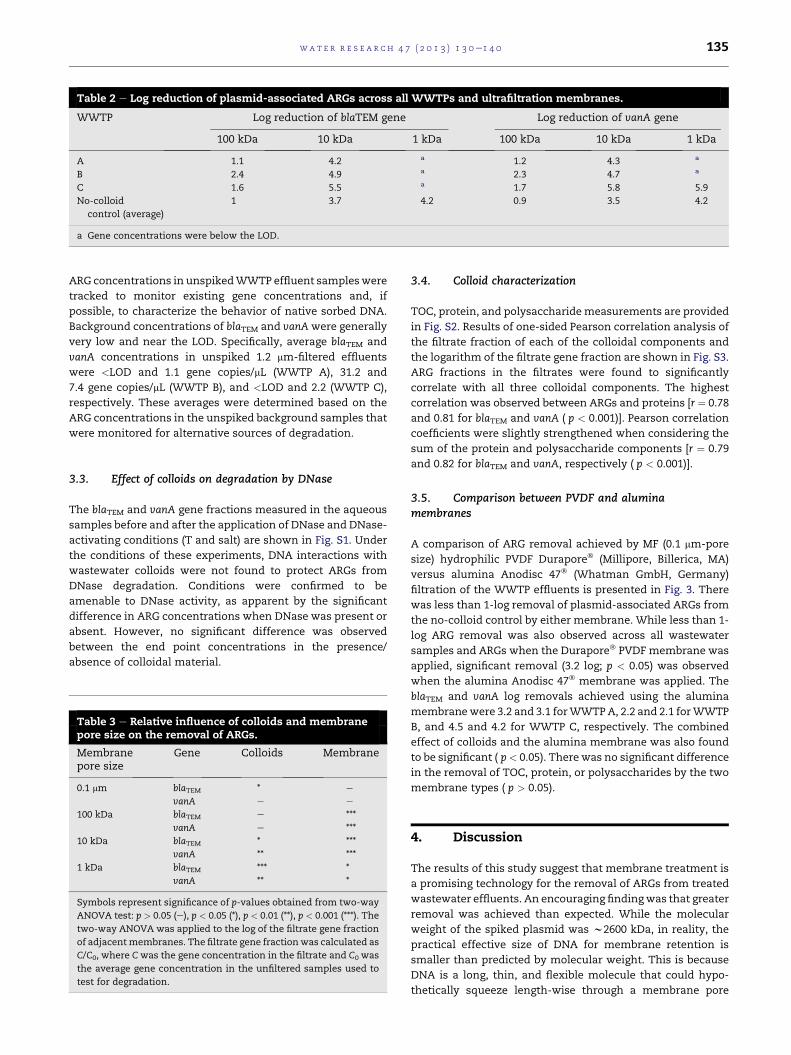
Table 2 e Log reduction of plasmid-associated ARGs across all WWTPs and ultrafiltration membranes.
WWTP Log reduction of blaTEM gene Log reduction of vanA gene
100 kDa 10 kDa 1 kDa 100 kDa 10 kDa 1 kDa
A 1.1 4.2 a 1.2 4.3 a
B 2.4 4.9 a 2.3 4.7 a
C 1.6 5.5 a 1.7 5.8 5.9
No-colloid
control (average)
1 3.7 4.2 0.9 3.5 4.2
a Gene concentrations were below the LOD.
wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0 135
ARG concentrations in unspikedWWTP effluent sampleswere
tracked to monitor existing gene concentrations and, if
possible, to characterize the behavior of native sorbed DNA.
Background concentrations of blaTEM and vanAwere generally
very low and near the LOD. Specifically, average blaTEM and
vanA concentrations in unspiked 1.2 mm-filtered effluents
were <LOD and 1.1 gene copies/mL (WWTP A), 31.2 and
7.4 gene copies/mL (WWTP B), and <LOD and 2.2 (WWTP C),
respectively. These averages were determined based on the
ARG concentrations in the unspiked background samples that
were monitored for alternative sources of degradation.
3.3. Effect of colloids on degradation by DNase
The blaTEM and vanA gene fractions measured in the aqueous
samples before and after the application of DNase and DNase-
activating conditions (T and salt) are shown in Fig. S1. Under
the conditions of these experiments, DNA interactions with
wastewater colloids were not found to protect ARGs from
DNase degradation. Conditions were confirmed to be
amenable to DNase activity, as apparent by the significant
difference in ARG concentrations when DNase was present or
absent. However, no significant difference was observed
between the end point concentrations in the presence/
absence of colloidal material.
Table 3 e Relative influence of colloids and membranepore size on the removal of ARGs.
Membranepore size
Gene Colloids Membrane
0.1 mm blaTEM * e
vanA e e
100 kDa blaTEM e ***
vanA e ***
10 kDa blaTEM * ***
vanA ** ***
1 kDa blaTEM *** *
vanA ** *
Symbols represent significance of p-values obtained from two-way
ANOVA test: p > 0.05 (e), p < 0.05 (*), p < 0.01 (**), p < 0.001 (***). The
two-way ANOVA was applied to the log of the filtrate gene fraction
of adjacentmembranes. The filtrate gene fractionwas calculated as
C/C0, where Cwas the gene concentration in the filtrate and C0 was
the average gene concentration in the unfiltered samples used to
test for degradation.
3.4. Colloid characterization
TOC, protein, and polysaccharidemeasurements are provided
in Fig. S2. Results of one-sided Pearson correlation analysis of
the filtrate fraction of each of the colloidal components and
the logarithm of the filtrate gene fraction are shown in Fig. S3.
ARG fractions in the filtrates were found to significantly
correlate with all three colloidal components. The highest
correlation was observed between ARGs and proteins [r ¼ 0.78
and 0.81 for blaTEM and vanA ( p < 0.001)]. Pearson correlation
coefficients were slightly strengthened when considering the
sum of the protein and polysaccharide components [r ¼ 0.79
and 0.82 for blaTEM and vanA, respectively ( p < 0.001)].
3.5. Comparison between PVDF and aluminamembranes
A comparison of ARG removal achieved by MF (0.1 mm-pore
size) hydrophilic PVDF Durapore� (Millipore, Billerica, MA)
versus alumina Anodisc 47� (Whatman GmbH, Germany)
filtration of the WWTP effluents is presented in Fig. 3. There
was less than 1-log removal of plasmid-associated ARGs from
the no-colloid control by either membrane. While less than 1-
log ARG removal was also observed across all wastewater
samples and ARGs when the Durapore� PVDF membrane was
applied, significant removal (3.2 log; p < 0.05) was observed
when the alumina Anodisc 47� membrane was applied. The
blaTEM and vanA log removals achieved using the alumina
membranewere 3.2 and 3.1 forWWTPA, 2.2 and 2.1 forWWTP
B, and 4.5 and 4.2 for WWTP C, respectively. The combined
effect of colloids and the alumina membrane was also found
to be significant ( p< 0.05). There was no significant difference
in the removal of TOC, protein, or polysaccharides by the two
membrane types ( p > 0.05).
4. Discussion
The results of this study suggest that membrane treatment is
a promising technology for the removal of ARGs from treated
wastewater effluents. An encouraging findingwas that greater
removal was achieved than expected. While the molecular
weight of the spiked plasmid was w2600 kDa, in reality, the
practical effective size of DNA for membrane retention is
smaller than predicted by molecular weight. This is because
DNA is a long, thin, and flexible molecule that could hypo-
thetically squeeze length-wise through a membrane pore

Fig. 2 e Stability of vanA and blaTEM ARGs in spiked WWTP effluents and no-colloid buffer controls. Error bars represent
standard deviation of analytical triplicates, but are smaller than the symbols in most cases. Values in parentheses indicate
minimum (in gene copies/mL), maximum (in gene copies/mL), and coefficient of variation. Letters on the left indicate WWTP
effluent source.
wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0136
(Marko et al., 2011; Travers, 2004). Correspondingly, the 1 kDa
membranes did not completely exclude passage of the
plasmid in the spiked buffer control (Fig. 1). The overall find-
ings highlight the importance of DNA conformation and
behavior, which can vary widely depending on water chem-
istry and composition.
To confirm that DNA removal was actually a result of
membrane filtration, ARG concentrations were monitored to
account for alternative sources of DNA loss, such as biodeg-
radation or sorption to the glass container. Interestingly, the
ARG concentrations did not change significantly, which
supports the conclusion that removal was driven by the
membranes. These control experiments also bring to light
the overall persistence of ARGs in treated wastewater
effluent. This result was somewhat unforeseen as the
extracellular DNA was expected to be more susceptible to
biodegradation given the numerous enzymes and other
cellular components present within a wastewater effluent
matrix. This observation further emphasizes the potential
importance of advanced treatment of wastewater effluents to
remove ARGs that can be transported and protected by larger
molecules.
Previous studies have noted the persistence of ARGs under
environmentally-relevant conditions, highlighting the poten-
tial for DNA protection through interactions with environ-
mental substrates. Specifically, the presence of ARB and ARGs,
including vanA, has been reported in drinking water sources,
treated drinking water, and treated wastewater (Auerbach
et al., 2007; Pruden et al., 2006; Schwartz et al., 2003; Xi et al.,
2009; Zhang et al., 2009; LaPara et al., 2011). The present
study found trace levels of vanA in all three WWTP effluents.
In addition, several studies have previously shown that water
disinfection processes such as chlorination can select for ARB
(Armstrong et al., 1982; Murray et al., 1984; Shrivastava et al.,
2004).
Previous studies have shown that DNA can be protected
against DNase degradationwhen it is bound to othermaterials
(Table 4). Contrary to these studies, the wastewater colloid-
associated ARGs remained susceptible to DNase degradation.
It is worth noting that the DNase degradation experiments in

Fig. 3 e Comparison of DNA removal between 0.1 mm PVDF
and alumina membranes. Error bars associated with
WWTPs represent standard deviations of analytical
measurements. Error bars associated with the control
represent standard deviations of experimental
measurements done independently in parallel with the
experiments involving each of the WWTPs.
wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0 137
this study were performed under conditions of excess
enzyme, and it is not certainwhether the DNAwas completely
sorbed to suspended material in the wastewater. Further
investigations of varying concentrations of DNA and DNase,
including environmentally-relevant concentrations, within
the wastewater matrix would be of interest.
Importantly, this study demonstrated that membrane
removal of ARGs is actually enhanced in the presence of
Table 4 e Summary of materials observed to interact with andtransformation ability of sorbed DNA.
Material Sorption Protection againstDNase degradation
T
Organic and
inorganic clays
þ þ
Soil colloids
and particles
þ þ
Silica/sand þ þ
Charcoal þ a
Humic acids þ þNOM þ a
a Data not available.
wastewater. Filtration experiment results revealed a signifi-
cant effect of 1.2 mm-filtered wastewater effluent matrix on
the removal of blaTEM and vanA genes present on the spiked
plasmids. This effect is presumably a result of interactions
with colloidal material present in the effluent, and is sup-
ported by the correlations observedwith the TOC, protein, and
polysaccharide fractions present in the filtrates. Previous
reports provide evidence of interactions between DNA and
organic and inorganic materials that are likely present in
wastewaters and also in natural waters (Table 4). However,
these studies were carried out only in simplified artificial
systems. Thus, the results of the present study provide insight
into the potential for membranes to remove ARGs from actual
wastewater effluents. Correlations with TOC, proteins, and
polysaccharides highlight the possibility that the interactions
taking place in the wastewater are complex and involve
agglomeration with various materials rather than the simple
interaction between DNA and discrete particles. The overall
results indicate that wastewater colloideARG interactions
could facilitate ARG removal by larger pore size membranes,
thus reducing the energy requirement for membrane filtra-
tion. The findings are congruent with those of a recent study
that demonstrated significantly higher ARB and ARG removals
from wastewater treated with a membrane bioreactor
compared to conventional treatment (Munir et al., 2011). This
finding further substantiates the hypothesis of DNAecolloid
interactions because, based on the molecular properties of
DNA, a conventional MBR system would not be expected to
significantly remove extracellular ARGs.
Effluents from three WWTPs were collected and used as
natural colloid-containingmedia in this study.According to the
literature, DNA has been found to bind to a variety of organic
and inorganic materials including different soil colloidal
particles, different types of clay, and NOM (Cai et al., 2007,
2006a, 2006b; Crecchio and Stotzky, 1998; Nguyen et al., 2010;
Nguyen and Elimelech, 2007a,b). However, DNA sorption
behavior not only varieswith type of sorbent, but alsowith pH,
ionic strength, type of cations present and DNA conformation
(Cai et al., 2006a; Mitra et al., 2001; Nguyen et al., 2010; Nguyen
and Elimelech, 2007b; Saeki et al., 2010). For this reason, it is
not possible to generate adsorption isotherms that would be
consistent across effluents. Even within one WWTP effluent,
/or protect DNA against enzymatic degradation, and
ransformationability
References
þ Cai et al. (2007), Cai et al. (2006a),
Cai et al. (2006b), Khanna and Stotzky (1992)
þ Cai et al. (2007), Cai et al. (2006a),
Cai et al. (2006b), Saeki et al. (2010)a Lorenz and Wackernagel (1987), Mitra et al.
(2001), Nguyen et al. (2010), Nguyen and
Elimelech (2007b), Romanowski et al. (1991)a Mitra et al. (2001)
þ Crecchio and Stotzky (1998)a Nguyen et al. (2010)

wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0138
the composition would likely change seasonally or even daily.
Based on adsorption times reported in the literature (Mitra
et al., 2001) and on typical WWTP retention times, a 4-h sorp-
tion timewaschosen toallow theDNAto interactwith colloidal
material in the wastewater effluent. Recent studies have indi-
cated trends of DNA transmission through UF membranes at
different ionic strengths, pH, andpressure (Arkhangelsky et al.,
2008; Latulippe and Zydney, 2008). Thus, the ionic strength of
the no-colloid control and the working pressure for all UF
membranes were kept constant throughout this study.
Comparisonof themembrane compositionprovided further
insight intomechanisms of ARG removal. The removal of ARGs
was most significantly enhanced by the combination of the
alumina membrane and the presence of wastewater material.
Interestingly, reductions observed in TOC, protein, and poly-
saccharidecolloidal components followingaluminamembrane
filtration (Fig. S2)were not significant ( p> 0.05).WhileDNAhas
been previously reported to interact with alumina and
aluminumspeciesat intermediatepH(Karliketal., 1980;Maand
Yeung, 2010), the removal of DNA in the buffer control was
negligible (Fig. 3). Although this finding suggests that DNA-
ealumina interactionsmay not be a significant factor affecting
the enhanced DNA removal by alumina membranes, it is also
possible that DNA retention was enhanced by DNAealumina
interactions within the wastewater matrix, but perhaps not
necessarily by the presence of colloidal material. One of the
reported attributes of the Anodisc aluminamembrane is that it
is made of a non-deformable material with no crossover
between individual pores. It is possible that although the
average reported pore size of the PVDF membrane was equiv-
alent (0.1 mm), either the actual range of pore sizes was higher
and reached pore sizes above 0.1 mm, or there were crossovers
among the pores. It is also possible that the PVDF membrane
was slightly deformable, thus allowing the transmission of
larger particles. Theseobservationshighlight the importanceof
investigating thedifferencesamongpotentialmembranesprior
to wider-scale use, as certain membranes could have the
potential to remove a wider range of contaminants under
similar or even lower energy demands.
Fouling is an important factor to consider for any
membrane treatment technology. Membranes subjected to
colloids andwastewater effluents are expected to foul as these
materials build up at the membrane surface (Ang et al., 2011).
The present study was purposefully not designed to examine
the implications of membrane fouling on ARG removal;
however, based upon the observed enhancement in ARG
removal in the presence of wastewater colloids it can be
hypothesized that ARGs will be more efficiently removed by
membranes that are partially fouled with organic material
than pristine membranes alone. This latter hypothesis,
however, requires additional experimental testing under
a variety of operating conditions.
5. Conclusions
Overall, this study demonstrates that membranes are
a promising treatment technology for the removal of ARGs
fromwastewater effluents. Fourmain findings are highlighted
from this study:
� ARGs could be effectively removed by membrane filtration,
with best performance observed for 10 kDamembranes (UF)
and smaller. Membrane removal of ARGs was better than
expected based on the molecular properties of DNA.
� ARGs spiked into the pre-filteredwastewater effluentmatrix
remained stable during the 4-h monitoring period, indi-
cating that ARGs can persist in wastewater effluents and
supporting the need to investigate treatment technologies.
� Associations of ARGswith natural colloidalmaterial present
in wastewater effluent actually resulted in enhanced
removal of ARGs. These results are encouraging, and
suggest that cost and energy requirements for ARG treat-
ment may be less than would be predicted by molecular
properties alone.
� Membrane material and other characteristics also have an
impact on ARG removal and should be optimized in future
studies.
Membrane treatments are likely to grow in their imple-
mentation for advancing water reuse and the results provide
important baseline information for the development of
a comprehensive water sustainability strategy. In particular,
membranes have been demonstrated to be effective for the
removal of other emerging contaminants, such as pharma-
ceuticals and personal care products. Benefits of membrane
treatment could be extended to ARGs as WWTPs consider
upgrades. Considering the growing problem of antibiotic
resistance and the demonstrated links between WWTPs and
ARGs in receiving environments, incorporating membrane
treatment of WWTP effluents may be advantageous for
minimizing potential risk to public health.
Acknowledgments
Financial support for this project was provided by the Water
Environment Research Foundation Project INFR8SG09, by the
NSF CBET CAREER award #0547342, and by the Institute for
Critical Technology and Applied Science award TSTS-11-26.
The findings do not reflect the opinions of the sponsors.
Special thanks to Dr. Dan Gallagher at Virginia Tech for his
insights regarding statistical analyses and Dr. Tony Fane of
the Singapore Membrane Technology Centre for consultation
in experimental design.
Appendix A. Supplementary data
Supplementary data related to this article can be found at
http://dx.doi.org/10.1016/j.watres.2012.09.044.
r e f e r e n c e s
Allen, H.K., Donato, J., Wang, H.H., Cloud-Hansen, K.A., Davies, J.,Handelsman, J., 2010. Call of the wild: antibiotic resistancegenes in natural environments. Nature Reviews Microbiology8 (4), 251e259.
Ang, W.S., Tiraferri, A., Chen, K.L., Elimelech, M., 2011. Foulingand cleaning of RO membranes fouled by mixtures of organic

wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0 139
foulants simulating wastewater effluent. Journal of MembraneScience 376 (1e2), 196e206.
Arkhangelsky, E., Steubing, B., Ben-Dov, E., Kushmaro, A.,Gitis, V., 2008. Influence of pH and ionic strength ontransmission of plasmid DNA through ultrafiltrationmembranes. Desalination 227 (1e3), 111e119.
Armstrong, J.L., Calomiris, J.J., Seidler, R.J., 1982. Selection ofantibiotic-resistant standard plate-count bacteria duringwater treatment. Applied and Environmental Microbiology 44(2), 308e316.
Auerbach, E.A., Seyfried, E.E., McMahon, K.D., 2007. Tetracyclineresistance genes in activated sludge wastewater treatmentplants. Water Research 41 (5), 1143e1151.
Baquero, F., Martinez, J.L., Canton, R., 2008. Antibiotics andantibiotic resistance in water environments. Current Opinionin Biotechnology 19 (3), 260e265.
Bibbal, D., Dupouy, V., Ferre, J.P., Toutain, P.L., Fayet, O.,Prere, M.F., Bousquet-Melou, A., 2007. Impact of threeampicillin dosage regimens on selection of ampicillinresistance in Enterobacteriaceae and excretion of blaTEM genesin swine feces. Applied and Environmental Microbiology 73(15), 4785e4790.
Biyela, P.T., Lin, J., Bezuidenhout, C.C., 2004. The role of aquaticecosystems as reservoirs of antibiotic resistant bacteria andantibiotic resistance genes. Water Science and Technology 50(1), 45e50.
Bockelmann, U., Dorries, H.H., Ayuso-Gabella, M.N., deMarcay, M.S., Tandoi, V., Levantesi, C., Masciopinto, C., VanHoutte, E., Szewzyk, U., Wintgens, T., Grohmann, E., 2009.Quantitative PCR monitoring of antibiotic resistance genesand bacterial pathogens in three European artificialgroundwater recharge systems. Applied and EnvironmentalMicrobiology 75 (1), 154e163.
Borjesson, S., Matussek, A., Melin, S., Lofgren, S., Lindgren, P.E.,2010. Methicillin-resistant Staphylococcus aureus (MRSA) inmunicipal wastewater: an uncharted threat? Journal ofApplied Microbiology 108 (4), 1244e1251.
Cai, P., Huang, Q., Chen, W., Zhang, D., Wang, K., Jiang, D.,Liang, W., 2007. Soil colloids-bound plasmid DNA: effect ontransformation of E. coli and resistance to DNase Idegradation. Soil Biology & Biochemistry 39 (5), 1007e1013.
Cai, P., Huang, Q., Zhang, X., Chen, H., 2006a. Adsorption of DNAon clay minerals and various colloidal particles from anAlfisol. Soil Biology & Biochemistry 38 (3), 471e476.
Cai, P., Huang, Q.Y., Zhang, X.W., 2006b. Interactions of DNA withclay minerals and soil colloidal particles and protectionagainst degradation by DNase. Environmental Science &Technology 40 (9), 2971e2976.
Crecchio, C., Stotzky, G., 1998. Binding of DNA on humic acids:effect on transformation of Bacillus subtilis and resistance toDNase. Soil Biology & Biochemistry 30 (8e9), 1061e1067.
Davies, J., Davies, D., 2010. Origins and evolution of antibioticresistance. Microbiology and Molecular Biology Reviews 74 (3),417e433.
Dolar, D., Kosutic, K., Pavlovic, D.M., Kunst, B., 2009. Removal ofemerging contaminants of industrial origin by NF/RO e a pilotscale study. Desalination and Water Treatment 6 (1e3),197e203.
Dolar, D., Zokic, T.I., Kosutic, K., Asperger, D., Pavlovic, D.M., 2012.RO/NF membrane treatment of veterinary pharmaceuticalwastewater: comparison of results obtained on a laboratoryand a pilot scale. Environmental Science and PollutionResearch 19 (4), 1033e1042.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith, F.,1956. Colorimetric method for determination of sugars andrelated substances. Analytical Chemistry 28 (3), 350e356.
Dutka-Malen, S., Evers, S., Courvalin, P., 1995. Detection ofglycopeptide resistance genotypes and identification to the
species level of clinically relevant Enterococci by PCR. Journal ofClinical Microbiology 33 (1), 24e27.
Karlik, S.J., Eichhorn, G.L., Lewis, P.N., Crapper, D.R., 1980.Interaction of aluminum species with deoxyribonucleic-acid.Biochemistry 19 (26), 5991e5998.
Khanna, M., Stotzky, G., 1992. Transformation of Bacillus subtilisby DNA bound on montmorillonite and effect of DNase on thetransforming ability of bound DNA. Applied andEnvironmental Microbiology 58 (6), 1930e1939.
Kim, S., Park, H., Chandran, K., 2010. Propensity of activatedsludge to amplify or attenuate tetracycline resistance genesand tetracycline resistant bacteria: a mathematical modelingapproach. Chemosphere 78 (9), 1071e1077.
LaPara, T.M., Burch, T.R., McNamara, P.J., Tan, D.T., Yan, M.,Eichmiller, J.J., 2011. Tertiary-treated municipal wastewater isa significant point source of antibiotic resistance genes intoDuluth-Superior harbor. Environmental Science & Technology45 (22), 9543e9549.
Larson, E., 2007. Community factors in the development ofantibiotic resistance. Annual Review of Public Health 28,435e447.
Latulippe, D.R., Zydney, A.L., 2008. Salt-induced changes inplasmid DNA transmission through ultrafiltrationmembranes. Biotechnology and Bioengineering 99 (2),390e398.
Lorenz, M.G., Wackernagel, W., 1987. Adsorption of DNA to sandand variable degradation rates of adsorbed DNA. Applied andEnvironmental Microbiology 53 (12), 2948e2952.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1977.Protein measurement with Folin phenol reagent. CurrentContents (1), 7.
Ma, C.B., Yeung, E.S., 2010. Entrapment of individual DNAmolecules and nanoparticles in porous alumina membranes.Analytical Chemistry 82 (2), 654e657.
Marko, A., Denysenkov, V., Margraft, D., Cekan, P., Schiemann, O.,Sigurdsson, S.T., Prisneet, T.F., 2011. Conformational flexibilityof DNA. Journal of the American Chemical Society 133 (34),13375e13379.
Martinez, J.L., 2008. Antibiotics and antibiotic resistance genes innatural environments. Science 321 (5887), 365e367.
Mitra, A., Chakraborty, P., Chattoraj, D.K., 2001. Kinetics ofadsorption of DNA at solideliquid interfaces. Journal of theIndian Chemical Society 78 (10e12), 689e696.
Munir, M., Wong, K., Xagoraraki, I., 2011. Release of antibioticresistant bacteria and genes in the effluent and biosolids offive wastewater utilities in Michigan. Water Research 45 (2),681e693.
Murray, G.E., Tobin, R.S., Junkins, B., Kushner, D.J., 1984. Effect ofchlorination on antibiotic-resistance profiles of sewage-related bacteria. Applied and Environmental Microbiology 48(1), 73e77.
Nguyen, T.H., Chen, K.L., Elimelech, M., 2010. Adsorption kineticsand reversibility of linear plasmid DNA on silica surfaces:influence of alkaline earth and transition metal ions.Biomacromolecules 11 (5), 1225e1230.
Nguyen, T.H., Elimelech, M., 2007a. Adsorption of plasmid DNA toa natural organic matter-coated silica surface: kinetics,conformation, and reversibility. Langmuir 23 (6), 3273e3279.
Nguyen, T.H., Elimelech, M., 2007b. Plasmid DNA adsorption onsilica: kinetics and conformational changes in monovalentand divalent salts. Biomacromolecules 8 (1), 24e32.
Oncu, N.B., Menceloglu, Y.Z., Balcioglu, I.A., 2011. Comparison ofthe effectiveness of chlorine, ozone, and photocatalyticdisinfection in reducing the risk of antibiotic resistancepollution. Journal of Advanced Oxidation Technologies 14 (2),196e203.
Pei, R.T., Kim, S.C., Carlson, K.H., Pruden, A., 2006. Effect of riverlandscape on the sediment concentrations of antibiotics and

wat e r r e s e a r c h 4 7 ( 2 0 1 3 ) 1 3 0e1 4 0140
corresponding antibiotic resistance genes (ARG). WaterResearch 40 (12), 2427e2435.
Pruden, A., Pei, R.T., Storteboom, H., Carlson, K.H., 2006.Antibiotic resistance genes as emerging contaminants: studiesin northern Colorado. Environmental Science & Technology 40(23), 7445e7450.
Romanowski,G., Lorenz,M.G.,Wackernagel,W., 1991.AdsorptionofplasmidDNA tomineral surfaces andprotection againstDNaseI.Applied and Environmental Microbiology 57 (4), 1057e1061.
Saeki, K., Kunito, T., Sakai, M., 2010. Effects of pH, ionic strength,and solutes on DNA adsorption by andosols. Biology andFertility of Soils 46 (5), 531e535.
Schwartz, T., Kohnen, W., Jansen, B., Obst, U., 2003. Detection ofantibiotic-resistant bacteria and their resistance genes inwastewater, surface water, and drinking water biofilms. FEMSMicrobiology Ecology 43 (3), 325e335.
Shrivastava, R., Upreti, R.K., Jain, S.R., Prasad, K.N., Seth, P.K.,Chaturvedi, U.C., 2004. Suboptimal chlorine treatment ofdrinking water leads to selection of multidrug-resistantPseudomonas aeruginosa. Ecotoxicology and EnvironmentalSafety 58 (2), 277e283.
Storteboom, H., Arabi, M., Davis, J.G., Crimi, B., Pruden, A., 2010.Tracking antibiotic resistance genes in the South Platte riverbasin using molecular signatures of urban, agricultural, andpristine sources. Environmental Science & Technology 44 (19),7397e7404.
Szczepanowski, R., Linke, B., Krahn, I., Gartemann, K.H.,Gutzkow, T., Eichler, W., Puhler, A., Schluter, A., 2009.
Detection of 140 clinically relevant antibiotic-resistance genesin the plasmid metagenome of wastewater treatment plantbacteria showing reduced susceptibility to selectedantibiotics. Microbiology-Sgm 155, 2306e2319.
Travers, A.A., 2004. The structural basis of DNA flexibility.Philosophical Transactions of the Royal Society of LondonSeries A-Mathematical Physical and Engineering Sciences 362(1820), 1423e1438.
Volkmann, H., Schwartz, T., Bischoff, P., Kirchen, S., Obst, U.,2004. Detection of clinically relevant antibiotic-resistancegenes in municipal wastewater using real-time PCR (TaqMan).Journal of Microbiological Methods 56 (2), 277e286.
Wintgens, T., Gallenkernper, M., Melin, T., 2004. Removal ofendocrine disrupting compounds with membrane processesin wastewater treatment and reuse. Water Science andTechnology 50 (5), 1e8.
Xi, C.W., Zhang, Y.L., Marrs, C.F., Ye, W., Simon, C., Foxman, B.,Nriagu, J., 2009. Prevalence of antibiotic resistance in drinkingwater treatment and distribution systems. Applied andEnvironmental Microbiology 75 (17), 5714e5718.
Zhang, R.F., Eggleston, K., Rotimi, V., Zeckhauser, R.J., 2006.Antibiotic resistance as a global threat: evidence fromChina, Kuwait and the United States. Globalization andHealth 2.
Zhang, Y.L., Marrs, C.F., Simon, C., Xi, C.W., 2009. Wastewatertreatment contributes to selective increase of antibioticresistance among Acinetobacter spp. Science of the TotalEnvironment 407 (12), 3702e3706.