Effect of retinoic acid on HL-60 cells infected with human immunodeficiency virus type 1
9
Effect of Retinoic Acid on HL-60 Cells Infected With Human Immunodeficiency Virus Type 1 By Marianne Semmel, Antonio Macho, Dominique Coulaud, Abdelkrim Alileche, Stephane Plaisance, Jose Aguilar, and Claude Jasmin HL-60 cells infected withhuman immunodeficiency virus type 1 (HIV 1) can be induced to differentiate along the gran- ulocyte pathway by retinoic acid. In these cells, HIV mRNA synthesis is stimulated, but synthesis of viral proteins and ELLS OF THE monocyte/macrophage lineage are con- sidered to be a reservoir of human immunodeficiency virus (HIV). The virus infects these cells without killing them and the cells produce moderate amounts of virus in- definitely.' During the maturation of the cells, HIV replica- tion can be modified.' In particular, HL-60 cells induced to differentiate before infection become permissive for mono- cytotropic strains of HIV type 1' and infection with HIV can modify the capacity of the cells to differentiate in response to inducer^.',^ U937, a monocyte/macrophage cell line, has been studied extensively. Differentiation of these cells upregulates HIV expression at the level of HIV mRNA5 and virus produ~tion.',~ Modification of HIV expression can be mediated by sev- eral cytokines: tumor necrosis factor a (TNFa) upregulates HIV replication in a T-cell line and in U937 cell^^.^; the interferons (IFNs) downregulate it.4,7,'0"2 However, IFNy can upregulate HIV replication if added to the culture before infection.13Transforming growth factor P (TGFP) also has dichotomous effects. If added to differentiating monocytes, it upregulates HIV replication but downregulates it in imma- ture monocytes and modifies HIV replication after induction of the cells with some, but not all, inducer^.'^.'^ Retinoic acid appears to act through TGFP." Turpin et all' report an increase of HIV mRNA transcription in monocytes, the monocyte-like U937, and the myeloid THP-1 cells treated with retinoic acid. Granulocyte-macrophage colony-stimu- lating factor (GM-CSF) has been reported to upregulate HIV expression in U1 cells (a clone derived from U937 cell~),'~*'' but todownregulate it in the parent line, U937.'" Interleukin- 1 (IL-I),lq IL-3," and IL-6" can upregulate HIV expression. Upregulation of HIV expression appears to be linked to the activation of protein kinase C,I8 but also to the induction of Differentiation induces the production of cyto- C NFKB,?1-2' From Unite' 268 INSERM, Villejug the Laboratoire de Chimie des Prote'ines, IRSC, Villejuif;and the Service de Microscopie Cellu- laire et Mole'culaire, IGR, VillejuiJ France. Submitted November I, 1993; accepted June 13, 1994. Supported by the Agence Nationale de Recherches sur le SIDA and by the Association Nouvelles Recherches Biomedicales. A.M. had a fellowship from the Ministerio de Educacion y Sciencia (Spain). Address reprint requests to Marianne Semmel, MD, U.268 1N- SERM, 14-16 AV. P.V. Couturier, Villejuif94800, France. The publication costs of this article weredefrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 1994 by The American Society of Hematology. 0006-497I/94/8408-0027$3.00/0 2480 virus replication are blocked and HIV-infected cells die after becoming apoptotic andlor vacuolized. 0 1994 by The American Society of Hematology. kines in cells of the monocyte/macrophage lineage; their amount will depend on the response of the cells to the in- ducer and the infecting virus. Because the relation between cytokines, differentiation, and HIV infection are very complex and because most of the studies have been performed with U937 cells that differ- entiate into macrophage-like cells, it seemed worthwhile to investigate the less intensively studied HL-60 cells, a line of myeloid parentage that can be induced to differentiate into granulocyte-like cells by retinoic acidz4 but is less differ- entiated than THPl cells.'' HL-60 cells are permissive for some but not all strains of HIV.z,25-27 We report here on the effect of retinoic acid on HL-60 cells infected with HIVl and on virus replication in these cells. MATERIALS AND METHODS Cells and virus. HL-60 cellsz8 were a gift from G. Milon (Institut Pasteur, Paris, France). CEM clone 11, a T-cell line, was a gift from F. Barrk-Sinoussi (Institut Pasteur). HT4-Lac21 cells were a gift from J.P. Nicolas (Institut Pasteur). CEM and HL-60 cells were grown in RPM1 1640 medium (GIBCO, BRL, Gaithersburg, MD) containing 10% heat-inactivated fetal calf serum (GIBCO), 2 mmoll L glutamine, 100 U penicillin, 10 bg streptomycin, and 0.25 pg amphothericidml. HT4-Lac21 cells were grown as described by Rocancourt et al." According to the supplier, endotoxin levels are very low in the media; the lot of serum used contained 43.8 ng endotoxidml. Cells were seeded at 2 X lo5 cells/mL and were incubated at 37°C in a humidified atmosphere containing 5% CO,. Medium was renewed and cell concentration readjusted to 2 X 10' cells twice weekly. HIV type 1 strain LA1 (formerly BRU)3'1 was a gift from F. Bank-Sinoussi. Stock virus was grown on CEM clone 11 cells and stored at -80°C. HL-60 cells were infected by adding cell-free supernatant (2 X IO" cpm RT HIV/106 cells) to the culture. Cell-free supematant was stored at -80°C. To obtain chronically infected cells, HL 60 cells were passaged for at least 70 days after infection. HIV infection was assayed weekly by coculture with HT4- Lac21 cells. Clones of chronically infected cells were obtained by limiting dilution and subcloned once. Continuing infection by HIV was monitored twice weekly by coculture with HT4-LacZ1 cells. To induce differentiation, cells were grown in the presence of 1 pmol/L all-trans retinoic acid (Sigma, St. Louis, MO). Cell viability was monitored by trypan blue exclusion. For fluorescence-activated cell sorter (FACS) analysis, cells were washed with phosphate-buffered saline (PBS), fixed with 1 % formal- dehyde and 0.1% glutaraldehyde, colored with iodine, and analyzed with an EPICS Profile I1 Analyzer (Coulter, Hialeah, FL). To observe cell morphology, 2 X lo4 cells were deposited on slides with a cytospin centrifuge (Shandon; BioBlock, Paris, France), fixed with May-Griinwald reagent (Biolyon, Lyon, France), colored with the Giemsa reagent (Prosciences, Paris, France), and visualized in a Leitz Dialux 20 microscope using oil immersion. For electron microscopy, cells were fixed in 4% glutaraldehyde in PBS and pelleted at low speed. The pellet was washed in 67 mmol/L phosphate buffer, pH 7.4, postfixed in 2% osmium tetroxide, Blood, Vol 84, No 8 (October 15). 1994: pp 2480-2488
Transcript of Effect of retinoic acid on HL-60 cells infected with human immunodeficiency virus type 1

Effect of Retinoic Acid on HL-60 Cells Infected With Human Immunodeficiency Virus Type 1
By Marianne Semmel, Antonio Macho, Dominique Coulaud, Abdelkrim Alileche, Stephane Plaisance, Jose Aguilar, and Claude Jasmin
HL-60 cells infected with human immunodeficiency virus type 1 (HIV 1) can be induced to differentiate along the gran- ulocyte pathway by retinoic acid. In these cells, HIV mRNA synthesis is stimulated, but synthesis of viral proteins and
ELLS OF THE monocyte/macrophage lineage are con- sidered to be a reservoir of human immunodeficiency
virus (HIV). The virus infects these cells without killing them and the cells produce moderate amounts of virus in- definitely.' During the maturation of the cells, HIV replica- tion can be modified.' In particular, HL-60 cells induced to differentiate before infection become permissive for mono- cytotropic strains of HIV type 1' and infection with HIV can modify the capacity of the cells to differentiate in response to inducer^.',^ U937, a monocyte/macrophage cell line, has been studied extensively. Differentiation of these cells upregulates HIV expression at the level of HIV mRNA5 and virus produ~tion.',~
Modification of HIV expression can be mediated by sev- eral cytokines: tumor necrosis factor a (TNFa) upregulates HIV replication in a T-cell line and in U937 cell^^.^; the interferons (IFNs) downregulate it.4,7,'0"2 However, IFNy can upregulate HIV replication if added to the culture before infection.13 Transforming growth factor P (TGFP) also has dichotomous effects. If added to differentiating monocytes, it upregulates HIV replication but downregulates it in imma- ture monocytes and modifies HIV replication after induction of the cells with some, but not all, inducer^.'^.'^ Retinoic acid appears to act through TGFP." Turpin et all' report an increase of HIV mRNA transcription in monocytes, the monocyte-like U937, and the myeloid THP-1 cells treated with retinoic acid. Granulocyte-macrophage colony-stimu- lating factor (GM-CSF) has been reported to upregulate HIV expression in U1 cells (a clone derived from U937 cell~), '~*' ' but to downregulate it in the parent line, U937.'" Interleukin- 1 (IL-I),lq IL-3," and IL-6" can upregulate HIV expression. Upregulation of HIV expression appears to be linked to the activation of protein kinase C,I8 but also to the induction of
Differentiation induces the production of cyto-
C
NFKB,?1-2'
From Unite' 268 INSERM, Villejug the Laboratoire de Chimie des Prote'ines, IRSC, Villejuif; and the Service de Microscopie Cellu- laire et Mole'culaire, IGR, VillejuiJ France.
Submitted November I , 1993; accepted June 13, 1994. Supported by the Agence Nationale de Recherches sur le SIDA
and by the Association Nouvelles Recherches Biomedicales. A.M. had a fellowship from the Ministerio de Educacion y Sciencia (Spain).
Address reprint requests to Marianne Semmel, MD, U.268 1N- SERM, 14-16 AV. P.V. Couturier, Villejuif94800, France.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 1994 by The American Society of Hematology. 0006-497I/94/8408-0027$3.00/0
2480
virus replication are blocked and HIV-infected cells die after becoming apoptotic andlor vacuolized. 0 1994 by The American Society of Hematology.
kines in cells of the monocyte/macrophage lineage; their amount will depend on the response of the cells to the in- ducer and the infecting virus.
Because the relation between cytokines, differentiation, and HIV infection are very complex and because most of the studies have been performed with U937 cells that differ- entiate into macrophage-like cells, it seemed worthwhile to investigate the less intensively studied HL-60 cells, a line of myeloid parentage that can be induced to differentiate into granulocyte-like cells by retinoic acidz4 but is less differ- entiated than THPl cells.'' HL-60 cells are permissive for some but not all strains of HIV.z,25-27 We report here on the effect of retinoic acid on HL-60 cells infected with HIVl and on virus replication in these cells.
MATERIALS AND METHODS
Cells and virus. HL-60 cellsz8 were a gift from G. Milon (Institut Pasteur, Paris, France). CEM clone 11, a T-cell line, was a gift from F. Barrk-Sinoussi (Institut Pasteur). HT4-Lac21 cells were a gift from J.P. Nicolas (Institut Pasteur). CEM and HL-60 cells were grown in RPM1 1640 medium (GIBCO, BRL, Gaithersburg, MD) containing 10% heat-inactivated fetal calf serum (GIBCO), 2 mmoll L glutamine, 100 U penicillin, 10 bg streptomycin, and 0.25 pg amphothericidml. HT4-Lac21 cells were grown as described by Rocancourt et al." According to the supplier, endotoxin levels are very low in the media; the lot of serum used contained 43.8 ng endotoxidml. Cells were seeded at 2 X lo5 cells/mL and were incubated at 37°C in a humidified atmosphere containing 5% CO,. Medium was renewed and cell concentration readjusted to 2 X 10' cells twice weekly. HIV type 1 strain LA1 (formerly BRU)3'1 was a gift from F. Bank-Sinoussi. Stock virus was grown on CEM clone 1 1 cells and stored at -80°C. HL-60 cells were infected by adding cell-free supernatant (2 X IO" cpm RT HIV/106 cells) to the culture. Cell-free supematant was stored at -80°C. To obtain chronically infected cells, HL 60 cells were passaged for at least 70 days after infection. HIV infection was assayed weekly by coculture with HT4- Lac21 cells. Clones of chronically infected cells were obtained by limiting dilution and subcloned once. Continuing infection by HIV was monitored twice weekly by coculture with HT4-LacZ1 cells. To induce differentiation, cells were grown in the presence of 1 pmol/L all-trans retinoic acid (Sigma, St. Louis, MO). Cell viability was monitored by trypan blue exclusion.
For fluorescence-activated cell sorter (FACS) analysis, cells were washed with phosphate-buffered saline (PBS), fixed with 1 % formal- dehyde and 0.1% glutaraldehyde, colored with iodine, and analyzed with an EPICS Profile I1 Analyzer (Coulter, Hialeah, FL).
To observe cell morphology, 2 X lo4 cells were deposited on slides with a cytospin centrifuge (Shandon; BioBlock, Paris, France), fixed with May-Griinwald reagent (Biolyon, Lyon, France), colored with the Giemsa reagent (Prosciences, Paris, France), and visualized in a Leitz Dialux 20 microscope using oil immersion.
For electron microscopy, cells were fixed in 4% glutaraldehyde in PBS and pelleted at low speed. The pellet was washed in 67 mmol/L phosphate buffer, pH 7.4, postfixed in 2% osmium tetroxide,
Blood, Vol 84, No 8 (October 15). 1994: pp 2480-2488
RETlNOlC ACID ON HL-60 CELLS WITH HIV
dehydrated with ethanol, and included in Epon resin by the usual techniques. Sections of cells were colored with uranyl acetate and lead citrate and observed with a Zeiss EM 902 microscope. Enhanced contrast was obtained by selecting elastic electrons using the slit of the spectrophotometer.
Determination of reverse transcriptase and tissue culture infec- tious units. HIV replication was assayed by measuring reverse tran- scriptase activity in the supernatant according to Oysten-J~nassen.~' Briefly, 50 pL of cell-free supernatant was incubated for 12 hours at 37°C in a mixture containing 0.05% Triton X100, 50 mmoVL KCl, 3 mmoYL dithiothreitol, 30 mmol/L Tris, pH 7.8, 0.3 mmol/ L EGTA, 0.13 U poly-rAdT as primer (Pharmacia, Brussels, Bel- gium), and 0.0378 mCi 3H-deoxythymidin triphosphate (Amersham, UK). The reaction was stopped with 10% trichloracetic acid con- taining 2 mmoYL Na pyrophosphate. The precipitate was washed 6 times with 5% trichloracetic acid containing 12 mmoVL Na pyro- phosphate, using an Automash 2000 Harvester (Dynatech; BioBlock) and Skatron filtermats. The filters were counted in scintillation liquid in a Beckman counter. Tissue culture infectious units (TCIU) were determined by titrating the virus in the supernatant or in the cells according to Rocancourt et al.29 Briefly, cell-free supernatants or cell suspensions were added to cultures of HT4-Lac Z1 cells
A
d 4 W 8 4
0 0 4 7 11 14
days E m
I 0 4 7
st inc
11
18
duction
14 18
days post induction
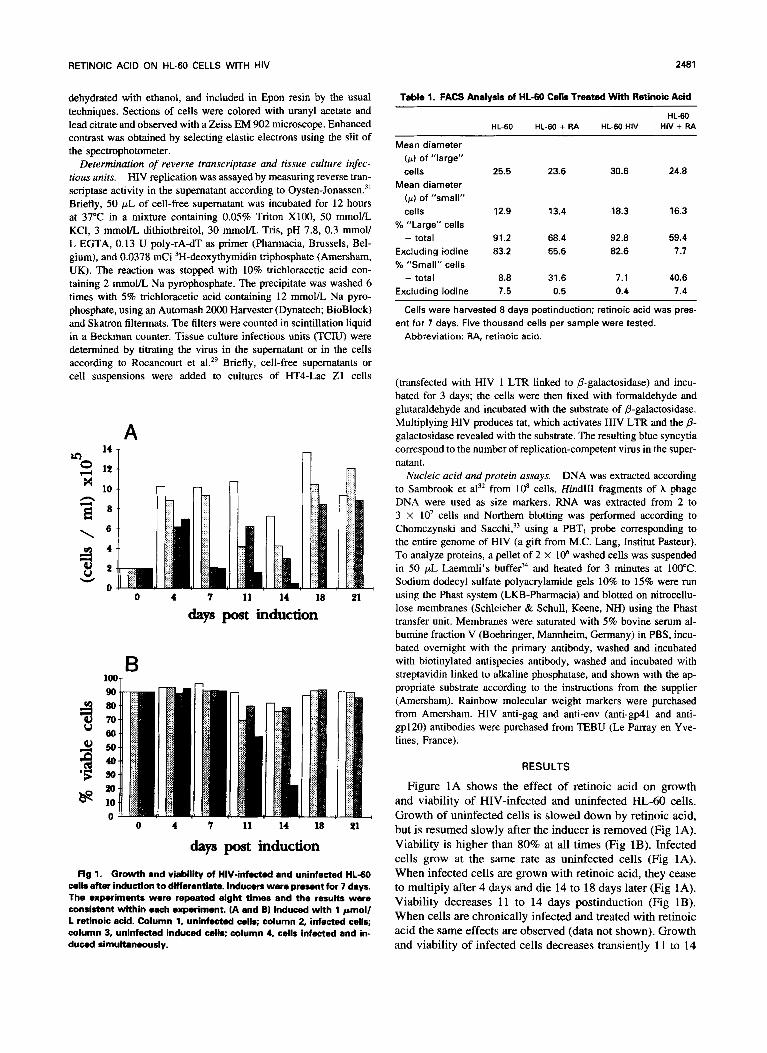
Fig l. Growth and viability of HW-infected and uninfected HL-60 cells after induction to diffarentiate. Inducers were present for 7 days. The expsrimonts were repeated eight times and the results were consistent within each experiment. (A and B) Induced with 1 pmoll L retinoic acid. Column 1, uninfected cells; column 2, infected cells; column 3, unlnfected induced cells; column 4, cells infected and in- duced simultaneously.
2481
Table 1. FACS Analysis of HL-60 Cells Traated With Retinoic Acid
HL-60 HL-60 HL-60 + RA HL-60 HIV HIV + RA
Mean diameter ( p ) of "large" cells 25.5 23.6 30.6 24.8
Mean diameter (1) of "small" cells 12.9 13.4 18.3 16.3
% "Large" cells - total 91.2 68.4 92.8 59.4
Excluding iodine 83.2 65.6 82.6 7 .l % "Small" cells - total 8.8 31.6 1.1 40.6
Excluding iodine 7.5 0.5 0.4 7.4
Cells were harvested 8 days postinduction; retinoic acid was pres-
Abbreviation: RA, retinoic acid. ent for 7 days. Five thousand cells per sample were tested.
(transfected with HIV 1 LTR linked to P-galactosidase) and incu- bated for 3 days; the cells were then fixed with formaldehyde and glutaraldehyde and incubated with the substrate of @-galactosidase. Multiplying HIV produces tat, which activates HIV LTR and the p- galactosidase revealed with the substrate. The resulting blue syncytia correspond to the number of replication-competent virus in the super- natant.
Nucleic acid and protein assays. DNA was extracted according to Sambrook et aI3* from lo8 cells. Hind111 fragments of X phage DNA were used as size markers. RNA was extracted from 2 to 3 x lo7 cells and Northern blotting was performed according to Chomczynski and S a ~ c h i , ~ ~ using a PBTI probe corresponding to the entire genome of HIV (a gift from M.C. Lang, Institut Pasteur). To analyze proteins, a pellet of 2 X IO6 washed cells was suspended in 50 pL Laemmli's buffef14 and heated for 3 minutes at 100°C. Sodium dodecyl sulfate polyacrylamide gels 10% to 15% were run using the Phast system (LKB-Pharmacia) and blotted on nitrocellu- lose membranes (Schleicher & Schull, Keene, NH) using the Phast transfer unit. Membranes were saturated with 5% bovine serum al- bumine fraction V (Boehringer, Mannheim, Germany) in PBS, incu- bated overnight with the primary antibody, washed and incubated with biotinylated antispecies antibody, washed and incubated with streptavidin linked to alkaline phosphatase, and shown with the ap- propriate substrate according to the instructions from the supplier (Amersham). Rainbow molecular weight markers were purchased from Amersham. HIV anti-gag and anti-env (anti-gp41 and anti- gp120) antibodies were purchased from TEBU (Le Parray en Yve- lines, France).
RESULTS
Figure 1A shows the effect of retinoic acid on growth and viability of HIV-infected and uninfected HL-60 cells. Growth of uninfected cells is slowed down by retinoic acid, but is resumed slowly after the inducer is removed (Fig 1A). Viability is higher than 80% at all times (Fig 1B). Infected cells grow at the same rate as uninfected cells (Fig 1A). When infected cells are grown with retinoic acid, they cease to multiply after 4 days and die 14 to 18 days later (Fig 1A). Viability decreases 1 1 to 14 days postinduction (Fig 1B). When cells are chronically infected and treated with retinoic acid the same effects are observed (data not shown). Growth and viability of infected cells decreases transiently 1 l to 14

2482 SEMMEL ET AL
1
Fig 2. Electron micrograph of HL-60 cells at low magnification. Cells were harvested 8 days postinduction. Scale bars = l 0 p. la and b l Untreated; IC and d) treated with retinoic acid. Uninfected (a) and infected lb) cells are undistinguishable; after treatment, most uninfected cells IC) have normal density, nuclei are multilobular, and two cells are apoptotic with dense cytoplasm, surface blebbing, and electron dense micronuclei. All infected cells (dl are apoptotic with dense vacuolized cytoplasm; apoptotic bodies are frequent.
RETlNOlC ACID ON HL-60 CELLS WITH HIV 2483
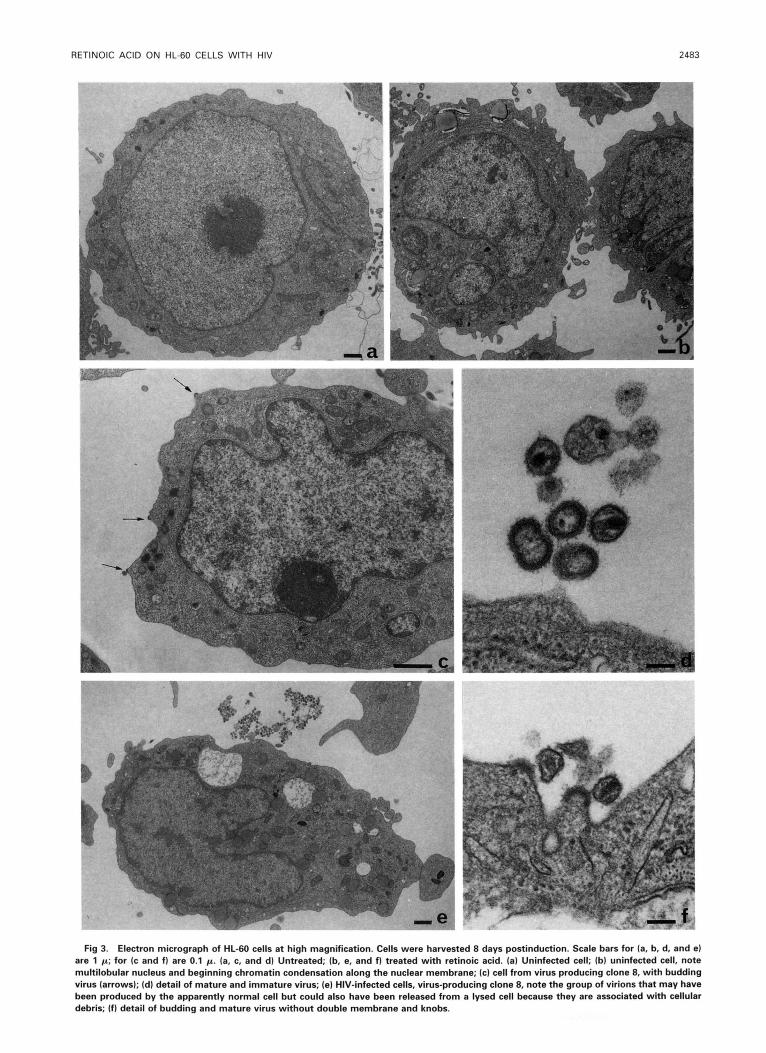
Fig 3. Electron micrograph of HL-60 cells at high magnification. Cells were harvested 8 days postinduction. Scale bars for la, b. d. and el are 1 p; for (c and f l are 0.1 p. (a, c, and d l Untreated; (b. e, and 1) treated with retinoic acid. (a) Uninfected cell; (bl uninfected cell, note multilobular nucleus and beginning chromatin condensation along the nuclear membrane; (c) cell from virus producing clone 8, with budding virus (arrows); (dl detail of mature and immature virus; (e) HIV-infected cells, virus-producing clone 8, note the group of virions that may have been produced by the apparently normal cell but could also have been released from a lysed cell because they are associated with cellular debris; (f) detail of budding and mature virus without double membrane and knobs.
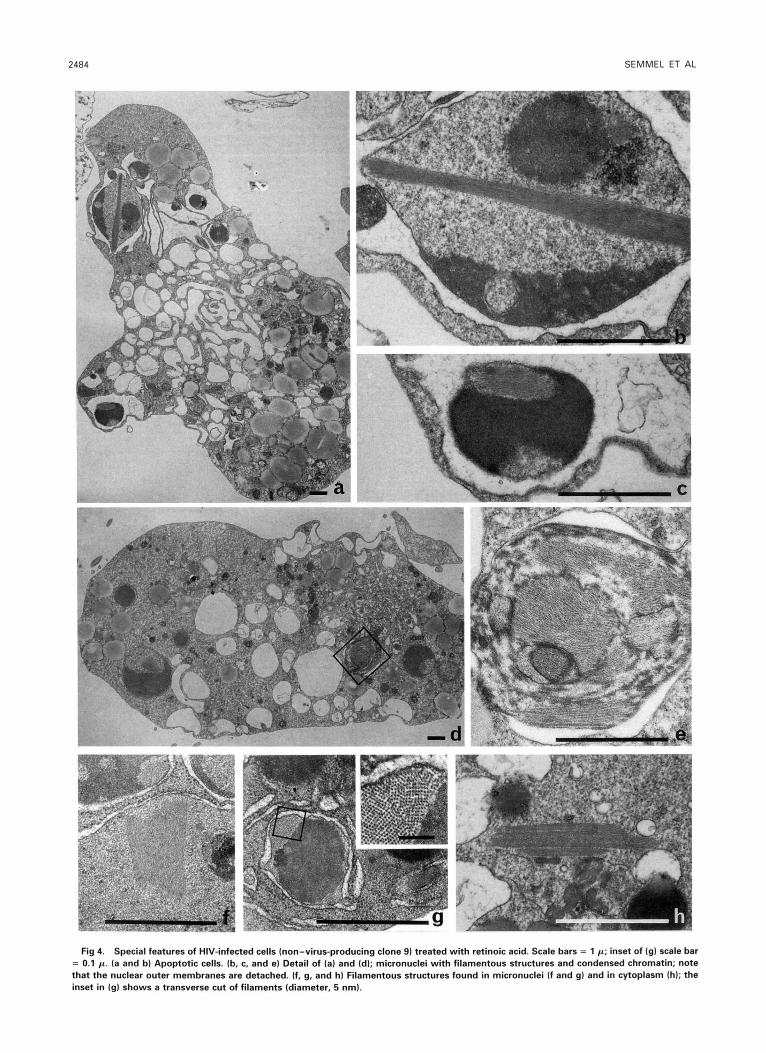
Fig 4. Special features of HIV-infected cells (non-virus-producing clone 91 treated with retinoic acid. Scale bars = 1 p; inset of (g1 scale bar = 0.1 p. (a and b) Apoptotic cells. (b. c, and e) Detail of (a) and (dl; micronuclei with filamentous structures and condensed chromatin; note that the nuclear outer membranes are detached. (f, g, and h) Filamentous structures found in micronuclei (f and g1 and in cytoplasm (h); the inset in (g) shows a transverse cut of filaments (diameter, 5 nm).

RETlNOlC ACID ON HL-60 CELLS WITH HIV 2485
Table 2. Effect of Retinoic Acid on HL-60 Cell Morphology
% of Abnormal Cells (Chromatin Segregated, Vacuolization,
Partial Lvsis)
Cells Induction 8 Days Postinduction Before
HL-60, uninfected 6 22
HL-60, HIV-infected clone 8, producing virus 31 72
producing few virions 5 93
One hundred cells were counted.
HL-60, HIV-infected, clone 9,
days postinfection (Fig 1A and B and Semmel et al”). HL- 60 cells treated with retinoic acid, whether infected or not, assume a granulocyte like morphology. Up to 80% of the cells appear to be differentiated between days 4 and 11 post- induction. Only 6% of the uninfected cells are differentiated 21 days postinduction; the percentage of differentiated in- fected cells also decreases, but these cells die 14 to 18 days postinfection. On day 7 postinduction, many of the infected cells that do not exclude trypan blue appear to be smaller than the corresponding uninfected cells. We therefore ana- lyzed chronically infected and uninfected cells 8 days postin- duction by FACS. HL-60 cells, whether infected or not, are heterogeneous in respect to size and constitute two subpopu- lations. HIV-infected cells are larger than uninfected cells; treatment with retinoic acid causes a decrease of the size, and the percentage of smaller cells increases. Iodine pene- trates cells more readily after treatment and when the cells are “large.” The results of these experiments are summa- rized in Table 1.
Chronically infected cells were cloned. All clones had subpopulations of “large” and “small” cells, but differed in respect to virus production as determined by coculture
5 1 A
with HT4-LacZ1 cells. Some clones consistently produce large amounts of virions, others produce, occasionally, a few virions. Electron microscopy (Figs 2 through 4) shows that treatment with retinoic acid decreases the size of uninfected HL-60 cells. The nuclei become horseshoe shaped and cren- allated, ressembling the nuclei of granulocytes. In some cells, chromatin is segregated, a few cells are vacuolized, and there are some apoptotic bodies (Fig 2c). At low magnification, infected cells are undistinguishable from uninfected cells before treatment with retinoic acid. At high magnifications, budding virions can be seen at the membrane of infected cells (Fig 3c) and most of the extracellular virions have an electron-dense double membrane and knobs (Fig 3d). After retinoic acid treatment, the cells and many cells are vac- uolized (Fig 2d), whether or not the cells produce virus. Budding virions are rare and there are no extracellular virions with double membranes (Fig 3e and f). The nuclear mem- brane is often detached (Fig 4e) and, in addition, both cyto- plasm and nuclei contain filaments (Fig 4). To our knowl- edge, such formations have not been described previously. Their origin is unknown, but their presence appears to be related to apoptosis, because they have also been observed in apoptotic cells of other lymphocytic cell lines (Ramos et al, personal communication, 1993). Table 2 summarizes quantitative results of electron microscopy.
Chromatin segregation and detachment of the nuclear membrane are considered to be a symptom of apoptosis, as is fragmentation of DNA in a typical L ‘DNA ladder” pattern.35 DNA extracted from HL-60 cells chronically infected with HIV does not differ from DNA of uninfected cells. After induction with retinoic acid, less DNA per cell is extracted, but the migration pattern is the same whether the cells were infected or not, showing only a band of high molecular weight material (data not shown).
HIV replication is blocked in HL-60 cells grown with retinoic acid regardless of whether the cells were chronically
- 6 iliil E
2 g 3
0 0 4 7 11 14 18 21
days post Induction days post induction
m 0 2 Fig 5. HIV replication in HL-60 cells. The experi- x
7
men- were repeated eight times and the results 2 - 120 were consistent within each experiment. (A and C) Reverse transcriptase activity. (B and Dl TCIU. (0)
noic acid. Inducer was present for 7 days. (A and B) Cells were simultaneously infected and induced. (C and D) Chronically infected cells were induced. days post induction days post induction
Uninduced controls; (A) induced with 1 pmol/L reti-
0 3 7 9 11 14 O O 3 7 9 11 14
2486 SEMMEL ET AL
A . .. ~
a
b
C"
d e
C 10
a 0.05
b 3.68
0 4.62
d 4.32+
e 9.27+
B
5 2.5 1.25pg
0.07. 0.06 . 0.11.
3.07 2.24 0.79.
5.63 4.98 5.17
5.63 7.11 5.77
11.63+ 13.39+ 18-26
+underestimated, HIV signal too strong for measurement *HIV signal too low for measurement
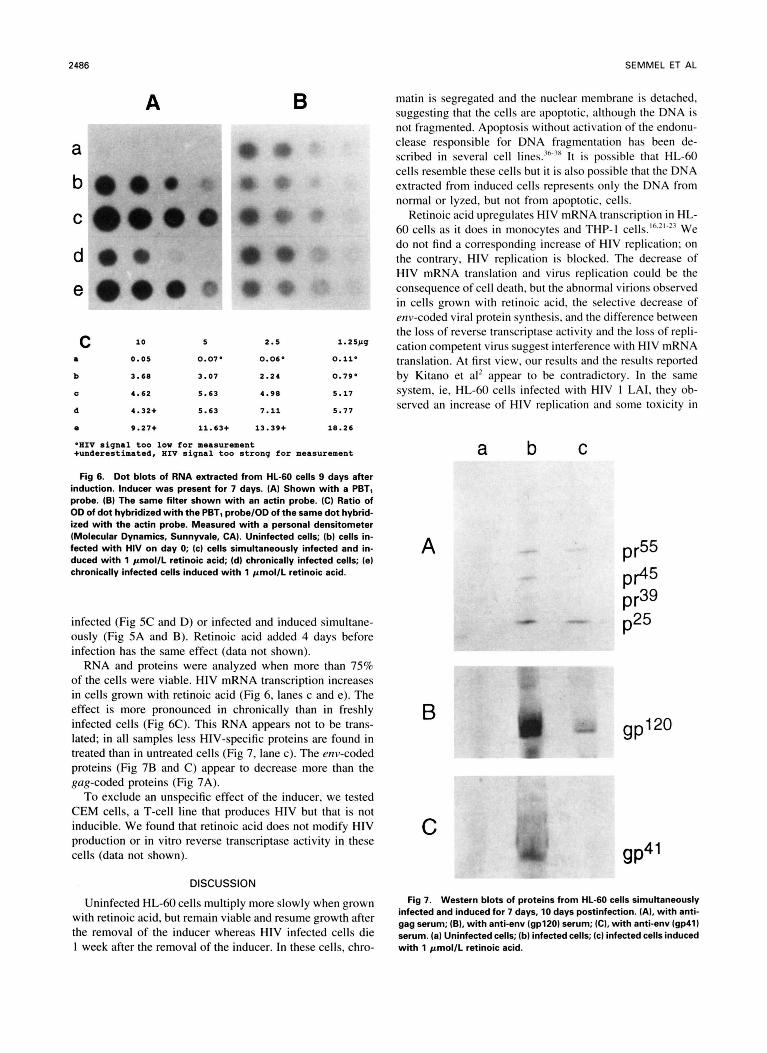
Fig 6. Dot blots of RNA extracted from HL-60 cells 9 days after induction. Inducer was present for 7 days. (A) Shown with a PBTl probe. (B) The same filter shown with an actin probe. (C) Ratio of OD of dot hybridized with the PBTl probe/OD of the same dot hybrid- ized with the actin probe. Measured with a personal densitometer (Molecular Dynamics, Sunnyvale, CA). Uninfected cells; (bl cells in- fected with HIV on day 0; (cl cells simultaneously infected and in- duced with 1 pmol/L retinoic acid; (dl chronically infected cells; (el chronically infected cells induced with 1 pmol/L retinoic acid.
infected (Fig 5C and D) or infected and induced simultane- ously (Fig 5A and B). Retinoic acid added 4 days before infection has the same effect (data not shown).
RNA and proteins were analyzed when more than 75% of the cells were viable. HIV mRNA transcription increases in cells grown with retinoic acid (Fig 6, lanes c and e). The effect is more pronounced in chronically than in freshly infected cells (Fig 6C). This RNA appears not to be trans- lated; in all samples less HIV-specific proteins are found in treated than in untreated cells (Fig 7, lane c). The env-coded proteins (Fig 7B and C) appear to decrease more than the gag-coded proteins (Fig 7A).
To exclude an unspecific effect of the inducer, we tested CEM cells, a T-cell line that produces HIV but that is not inducible. We found that retinoic acid does not modify HIV production or in vitro reverse transcriptase activity in these cells (data not shown).
DISCUSSION
Uninfected HL-60 cells multiply more slowly when grown with retinoic acid, but remain viable and resume growth after the removal of the inducer whereas HIV infected cells die 1 week after the removal of the inducer. In these cells, chro-
matin is segregated and the nuclear membrane is detached, suggesting that the cells are apoptotic, although the DNA is not fragmented. Apoptosis without activation of the endonu- clease responsible for DNA fragmentation has been de- scribed in several cell lines.".3x It is possible that HL-60 cells resemble these cells but it is also possible that the DNA extracted from induced cells represents only the DNA from normal or lyzed, but not from apoptotic, cells.
Retinoic acid upregulates HIV mRNA transcription in HL- 60 cells as it does in monocytes and THP-I cells.'h.'"23 We do not find a corresponding increase of HIV replication; on the contrary, HIV replication is blocked. The decrease of HIV mRNA translation and virus replication could be the consequence of cell death, but the abnormal virions observed in cells grown with retinoic acid, the selective decrease of env-coded viral protein synthesis, and the difference between the loss of reverse transcriptase activity and the loss of repli- cation competent virus suggest interference with HIV mRNA translation. At first view, our results and the results reported by Kitano et al' appear to be contradictory. In the same system, ie, HL-60 cells infected with HIV 1 LAI, they ob- served an increase of HIV replication and some toxicity in
a b C
A
B m
pr55
PP5 pr39
gP'*O
C 9P4'
Fig 7. Western blots of proteins from HL-60 cells simultaneously infected and induced for 7 days, 10 days postinfection. (AI, with anti- gag serum; (B), with anti-env (gpl20) serum; (C), with anti-env lgp41) serum. (a) Uninfected cells; (b) infected cells; (c) infected cells induced with 1 pmol/L retinoic acid.

RETlNOlC ACID ON HL-60 CELLS WITH HIV
cells treated with retinoic acid. However, their conditions differed from ours; they treated the cells before infection, we treated them during and after infection. Also, the HL-60 strains used appear to be different. Only a subpopulation of the HL-60 cells used by Kitano et a12 expressed the CD4 antigen and not all cells were infected, whereas 92% of our HL-60 cells expressed this antigen and more than 95% of the cells were infected”; therefore, one would expect more toxicity when more cells were infected.
The morphology of the cells shows that retinoic acid in- duces differentiation in both uninfected and HIV-infected cells and that undifferentiated cells overgrow the differenti- ated uninfected cells, whereas the HIV-infected cells die when the inducer is removed.
Many published results, including ours, indicate the exis- tence, in cells of the monocyte/macrophage lineage, of a stage of differentiation that restricts HIV replication at one or more levels of the replication cycle, whereas, at other stages of differentiation, HIV replication is upregulated. Each stage of differentiation corresponds to a balance of cytokines and it appears reasonable to expect that the out- come of HIV infection in a given cell (replication of the virus, silent infection, or cell death) will be determined by this balance. Retinoic acid, while stimulating HIV mRNA transcription, appears to block HIV replication and to cause the death of infected but not of uninfected HL-60 cells, possibly by modifying the balance of cytokines. Immature granulocytes are the normal counterpart of HL-60 cells. If the effect of retinoic acid on the normal cells in vivo were the same as that on HL-60 cells in vitro, retinoic acid would promote the death of HIV-infected cells, thereby reducing the reservoir of HIV-infected cells, and would cause the emergence of replication defective virus, thus reducing the virus load; it might then be considered useful for the treat- ment of acquired immunodeficiency syndrome. On the other hand, the absence of mature granulocytes might facilitate opportunistic infections because this cell is active in the host defense.
REFERENCES 1. Gendelman HE, Orenstein JM, Baca LM, Weiser B, Burger
H, Kalter DCh, Meltzer MS: The macrophage in the persistence and pathogenesis of HIV infection. AIDS 3:475, 1989
2. Kitano K, Baldwin GC, Raines MA, Golde DW: Differentiat- ing agents facilitate infection of myeloid leukemia cell lines by monocytotropic HIV strains. Blood 76:1980, 1990
3. Pautrat G, Suzan M, Salaun D, Corbeau P, Allasia C, Morel G, Filippi P: Human immunodeficiency virus type 1 infection of U937 cells promotes cell differentiation and a new pathway of viral assembly. Virology 179:749, 1990
4. Gazzolo L, Mack K: Regulation of HIV 1 replication in pro- monocytic U937 cells. Res Virol 141:259, 1990
5. Pauza CD, Galindo J, Richman DD: Human immunodeficiency virus infection of monoblastoid cells: Cellular differentiation deter- mines the pattern of virus replication. J Virol 62:3558, 1988
6. Folks TM, Justement J, Kinter A, Schnittman S, Orenstein J, Poli G, Fauci AS: Characterization of a promonocyte clone chroni- cally infected with HIV and inducible by 13-phorbol-l2-myristate acetate. J Immunol 140: I 1 17, 1988
7. Locardi Ch, Petrini C, Boccoli G, Testa U, Dieffenbach C, Butto S, Belardelli F: Increased human immunodeficiency virus
(HIV) expression in chronically infected U937 cells upon in vitro differentiation by hydroxyvitamin D3: Roles of interferon and tumor necrosis factor in regulation of HIV production. J Virol 64:5874, 1990
8. Duh ET, Maury WJ, Folks TM, Fauci AS, Rabson AB: Tumor necrosis factor a activates human immunodeficiency virus type 1 through induction of nuclear factor binding to the NF-K B sites in long terminal repeat. Proc Natl Acad Sci USA 86:5974, 1989
9. Poli G, Kinter A, Justement JS, Kehrl JH, Bressler P, Stanley S, Fauci AS: Tumor necrosis factor a functions in an autocrine manner in the induction of human immunodeficiency virus expres- sion. Proc Natl Acad Sci USA 87:782, 1990
IO. Hammer SM, Gillis JM, Groopman JE, Rose R M : In vitro modification of human immunodeficiency virus infection by granulo- cyte-macrophage colony-stimulating factor and gamma interferon. Proc Natl Acad Sci USA 83:8734, 1986
1 I . Dubreuil M, Sportza L, D’Addario M, Lacoste J, Rooke R, Wainberg MA, Hiscott J. Inhibition of HIV-I transmission by inter- feron and 3’-azido-3’ deoxythymidine during de novo infection of promonocytic cells. Virol 179:388, 1990
12. Mack K, Gazzolo L: Interferon regulated viral replication in chronically HIV-infected promonocytic U937 cells. Res Virol 142:213, 1991
13. Koyanagi Y, O’Brien WA, Zhao JQ, Golde DW, Gasson JC, Chen ISY: Cytokines alter production of HIV-I from primary mononuclear phagocytes. Science 241: 1673, 1988
14. Lazdins JK, Klimkait T, Alteri E, Walker M, Woods-Cook K, Cox D, Bilbe G, Shipman R, Cerletti N, McMasters G: TGF-P: Upregulator of HIV replication in macrophages. Res Virol 142239, 1991
15. Poli G, Kinter AL. Justement JS, Bressler P, Kehrl JH, Fauci AS: Retinoic acid mimics transforming growth factor P in the regula- tion of human immunodeficiency virus expression in monocytic cells. Proc Natl Acad Sci USA 89:2689, 1992
16. Turpin JA, Vargo M, Meltzer MS: Enhanced HIV-I replica- tion in retinoid-treated monocytes. Retinoid mediated effects through mechanisms related to cell differentiation and to a direct transcrip- tional action on viral gene expression. J Immunol 1482539, 1992
17. Folks TM, Justement J, Kinter A, Dinarello CA, Fauci AS: Cytokine-induced expression of HIV 1 in a chronically infected promonocyte line. Science 238:800, 1987
18. Kinter AL, Poli G, Maury W, Folks TM, Fauci AS: Direct and cytokine-mediated activation of protein kinase C induces human immunodeficiency virus expression in chronically infected promono- cytic cells. J Virol 64:4306, 1990
19. von Briesen H, von Mallinckrodt C, Esser R, Muller S, Becker K, Rubsamen-Waigmann H, Andreesen R: Effects of cytokines and lipopolysaccharides on HIV infection of human macrophages. Res Virol 142:197, 1991
20. Kalebic T, Kinter A, Poli G, Anderson ME, Meister A, Fauci AS: Suppression of human immunodeficiency virus expression in chronically infected monocytic cells by glutathione, glutathione es- ter, and N-acetylcysteine. Proc Natl Acad Sci USA 88:986, 1991
21. Bachelerie F, Alcami J, Arenzana-Seisdedos F, Virelizier JL: HIV enhancer activity perpetuated by NF-KB induction on infection of monocytes. Nature 350:709, 1991
22. Griffin GE, Leung K, Folks TM, Kunkel S, Nabel GJ: Induc- tion of NF-K B during monocyte differentiation is associated with activation of HIV-gene expression. Res Virol 142:233, 1991
23. Raziuddin, Mikovits JA, Calvert I, Ghosh S, Kung HF, Rus- cetti F W : Negative regulation of human immunodeficiency virus type 1 expression in monocytes: Role of the 65-kDa plus 50 kDa NF K B dimer. Proc Natl Acad Sci USA 88:9426, 1991
24. Breitman TR, Selonick S, Collins SJ: Induction of differentia-

2400 SEMMEL ET AL
tion of the human promyelocytic cell line HL-60 by retinoic acid. Proc Natl Acad Sci USA 77:2936, 1980
25. Levy JA, Shimabukuro J, McHugh T, Casavant C, Stites D, Oshiro L: AIDS-associated retroviruses (ARV) can productively in- fect other cells besides human T helper cells. Virology 147:441, 1985
26. Clapham PR, Weiss RA, Dalgleish AG, Exley M, Whitby D, Hogg N: Human immunodeficiency virus infection of monocytic and T-lymphocytic cells: Receptor modulation and differentiation induced by phorbol esters. Virology 158:44, 1987
27. Semmel M, Macho A, Morozov V, Coulaud D, Alileche A, Plaisance S, Jasmin C: Growth of human immunodeficiency virus type 1 (HIVI), in HL 60 cells. Res Virol 143:249, 1992
28. Collins SJ, Gallo RC, Gallagher RE: Continuous growth and differentiation of human myeloid leukaemic cells in suspension cul- ture. Nature 270:347, 1977
29. Rocancourt D, Bonnerot C, Jouin H, Emerman M, Nicolas JF: Activation of a betagalactosidase recombinant provirus: Applica- tion to titration of human immunodeficiency virus (HIV) and HIV infected cells. J Virol 64:2660, 1990
30. Wain-Hobson S, Vartanian S, Henry JP, Chencinas N, Chey- nier R, Delassus S, Martins LP, Samla M, Nugeyre MT, GuCtard D, Klatzmann D, Gluckmann JC, Rozenbaum W, BarrC-Sinoussi F. Montagnier L: LAV revisited: The origin of the early HIV-1 isolates from Institut Pasteur. Science 252:961. 1991
3 I . Oysten-Jonassen T: Direct measurement of reverse tran- scriptase activity in the medium from HTLV 11-LAV infected cells: An application of the Skatron Harvester. Technical Note. Skatron, I986
32. Sambrook J , Fritsch EF, Maniatis T: Molecular Cloning (ed 2). Cold Spring Harbor, NY, Cold Spring Harbor Laboratory, 1989
33. Chomczynski P, Sacchi N: Single step method of RNA isola- tion by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162:156, 1987
34. Laemmli UK: Cleavage of structural proteins during the as- sembly of the head of bacteriophage T4. Nature 227:680, 1970
35. Williams GT: Programmed cell death: Apoptosis and onco- genesis. Cell 65: 1097, 1991
36. Thompson EA: Insensitivity to the cytolytic effects of gluco- cortoids in vivo is associated with a novel “slow death” phenotype. Cancer Res 5 1:5544, 1991
37. Boe R, Gjertsen BT, Vintermyr OK, Houge G, Lanotte M, Doeskeland SO: The phosphatase inhibitor okadaic acid induces morphological changes typical of apoptosis in mammalian cells. Exp Cell Res 195:237, 1991
38. Oberhammer F, Fritsch G, Schmied M, Pavelka M, Printz D, Purchio T, Lassmann H, Schulte-Hermann R: Condensation of chromatin at the membrane of an apoptotic nucleus is not associ- ated with activation of endonuclease. J Cell Sci 104:317, 1993