
Subtropical Brazilian mangroves as a refuge of crab (Decapoda: Brachyura) diversity
Upload
independentCategory
view
0download
0
This article was downloaded by: [186.223.255.241]On: 10 April 2012, At: 12:18Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Invertebrate Reproduction & DevelopmentPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tinv20
Early juvenile development of Armases rubripes(Rathbun 1897) (Crustacea, Brachyura, Sesarmidae)and comments on the morphology of the megalopa andfirst crabMaria Lucia Negreiros-Fransozo a , Camile Sorbo Fernandes a , Stella Maris Januário Da Silvaa & Adilson Fransozo aa NEBECC (Crustacean Biology, Ecology and Culture Study Group), Departamento deZoologia, Instituto de Biociências, Universidade Estadual Paulista, 18618-000 Botucatu, SãoPaulo, Brasil
Available online: 24 Jan 2011
To cite this article: Maria Lucia Negreiros-Fransozo, Camile Sorbo Fernandes, Stella Maris Januário Da Silva & AdilsonFransozo (2011): Early juvenile development of Armases rubripes (Rathbun 1897) (Crustacea, Brachyura, Sesarmidae) andcomments on the morphology of the megalopa and first crab, Invertebrate Reproduction & Development, 55:1, 53-64
To link to this article: http://dx.doi.org/10.1080/07924259.2010.548645
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

Invertebrate Reproduction & DevelopmentVol. 55, No. 1, March 2011, 53–64
Early juvenile development of Armases rubripes (Rathbun 1897) (Crustacea, Brachyura,
Sesarmidae) and comments on the morphology of the megalopa and first crab
Maria Lucia Negreiros-Fransozo*, Camile Sorbo Fernandes, Stella Maris Januario Da Silva andAdilson Fransozo
NEBECC (Crustacean Biology, Ecology and Culture Study Group), Departamento de Zoologia, Instituto de Biociencias,Universidade Estadual Paulista, 18618-000 Botucatu, Sao Paulo, Brasil
(Received 14 April 2010; final version received 8 October 2010)
This article provides a description of the early juvenile stages of Armases rubripes and indicates some differencesand additional details found in the megalopa and first juvenile crab stage compared with previous descriptions.The main characters that permit identification of the subsequent juvenile stages were assessed, as well as themoult interval and increment during successive moults and the beginning of development of the secondary sexualfeatures. Megalopae of A. rubripes were collected from the Ubatuba region, Sao Paulo, Brazil using plankton andneuston nets. Morphological changes in body parts have been recorded for each stage and sex, as well as theduration of the intermoult period was noted. Possible effects of laboratory holding time on growth were checkedby means of analysis of covariance. The importance of standardization of the terminology of setae is emphasized.
Keywords: larval stage; morphological description; neuston; sexual differentiation
Introduction
The identification of the species of juvenile brachyur-
ans is often difficult (Martin et al. 1984) and has been
an impediment to ecological research on their func-
tional importance in estuarine habitats (O’Connor
1990). Papers that focus, therefore, on an accurate
description of the juveniles of distinct species would
permit ecological studies on these often-neglected
stages.According to Hebling et al. (1982), most authors
who have raised larvae did not go beyond than the
megalopa stage due to rearing difficulties and low
survival of juvenile crabs in the laboratory. The
available information on post-larval ontogeny of
crabs, therefore, is much reduced compared to that
on larval development (Guerao and Rotllant 2009).The most detailed literature available on several
post-larval stages of crabs are concerned with the
acquisition of adult characters, juvenile growth and
sexual differentiation (Shen 1935; Payen 1974; Ingle
1977; Hebling et al. 1982; Felder et al. 1985; Fransozo
1987; Fransozo and Negreiros-Fransozo 1987; Rieger
and Nakagawa 1995; Flores et al. 1998; Rieger and
Beltrao 2000; Flores et al. 2002; Guimaraes and
Negreiros-Fransozo 2005; Negreiros-Fransozo et al.
2007; Bolla Jr et al. 2008).Guerao and Rotllant (2009) emphasized that whilst
morphological characteristics of early juveniles have
not usually been applied in taxonomic studies of crabs,
a description of the early instars is necessary to assist in
identification of juvenile stages of crabs collected in
field studies (Ingle and Rice 1984; Guimaraes and
Negreiros-Fransozo 2005). Felder et al. (1985) have
demonstrated the importance of a better knowledge on
the first post-larval stages, as it can help in the
evaluation of body size, morphology, moulting fre-
quency, growth rate, feeding behaviour and even the
elucidation of the phylogeny of some groups of
Decapoda, when it is associated with information
related to larvae and adults.Many larval stages of Sesarmidae have been
described from American waters over the past
50 years (Costlow Jr and Bookhout 1960, 1962; Dıaz
and Ewald 1968; Fransozo and Hebling 1986; Schubart
and Cuesta 1998; Cuesta and Anger 2001; Cuesta et al.
2006), but papers that focus the morphological
description of first juvenile stages, as for example
Fransozo (1986, 1987), Rieger and Nakagawa (1995),
Luppi and Spivak (2007), and Guerao et al. (2007), are
still scarce.Hartnoll (1982) reported the growth of animals that
possess a hard exoskeleton, such as crustaceans, as an
essentially discontinuous process. There are successive
moults separated by intermoult periods and almost all
the growth occurs immediately after ecdysis before the
hardening of the new tegument. During the intermoult,
*Corresponding author. Email: [email protected]
ISSN 0792–4259 print/ISSN 2157–0272 online
� 2011 Taylor & Francis
DOI: 10.1080/07924259.2010.548645
http://www.informaworld.com
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

a little growth may occur due to the flexibility of themembranes that join the sclerites which link the hardexoskeleton.
The moulting frequency and the size increase permoult may be influenced independently by changes in
environmental conditions. Thus, to best determinegrowth rates, both the intermoult duration and thegrowth over that period need to be measured (Cadmanand Weinstein 1988).
The present lack of information makes larval
studies necessary and important, as they provide asounder basis for systematic classification and phylo-genetics (Guimaraes and Negreiros-Fransozo 2005), aswell as the identification of megalopae and juvenilesobtained directly from the field.
The aim of this study is to describe the earlyjuvenile development of Armases rubripes. The maincharacters that permit the identification of the earlyjuvenile stages are described, as well as the moultinterval and the increment during successive moultsand the beginning of the external secondary sexualdifferentiation. This study also provides some addi-tional details on the megalopa and first juveniledescription of A. rubripes, as a complement to theoriginal paper by Dıaz and Ewald (1968). Using theinformation obtained about the morphologic charac-ters, it is possible to compare the A. rubripes megalopaand juvenile with other species, from both planktonand benthos.
Materials and methods
Samples were obtained during October, November andDecember 2005 and February and April 2006.Megalopae were collected using plankton and neustonnets at Itamambuca mangrove (23�240S; 45�030W) andat Ubatuba Bay (23�250S; 45�030W), both at Ubatuba,Sao Paulo, Brazil. The collected material was placed ina bucket containing previously aerated seawater. Thesampled material was examined and megalopae wereremoved using a disposable pipette. Subsequently, eachmegalopa was transferred to a clean acrylic vialcontaining about 60mL of aerated seawater, and themegalopae were taken to the laboratory for culture.
Figure 1. Armases rubripes. Pereopods of megalopa stage:(a) cheliped (scale bar 0.2mm); (b) second pereopod; (c) thirdpereopod; (d) fourth pereopod and (e) fifth pereopod (scalebar 0.5mm).
Figure 2. Armases rubripes. Ventral view of thoracic sternitesof megalopa stage.
54 M.L. Negreiros-Fransozo et al.
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

They were held separately in numbered plastic vialscontaining seawater at a salinity of 15 at 25�C. Eachlarva received about 30 nauplii of Artemia sp, daily forfeeding. Individuals were daily checked for moult, andthe water of the vials was changed every 2 days.The intermoult period was recorded. The exuviaeobtained in the process were preserved in correctlyidentified bottles, containing 80% ethanol and pureglycerin, maintaining 1:1 proportion. Similarly, the
Figure 5. Armases rubripes. Pereopods of the first juvenilestage: (a) cheliped; (b) second pereopod; (c) third pereopod;(d) fourth pereopod; (e) fifth pereopod.
Figure 3. Armases rubripes. Pleopods of megalopa stage: (a) second pleopod; (b) third pleopod; (c) fourth pleopod;(d) fifth pleopod.
Figure 4. Armases rubripes. First juvenile stage: (a) dorsalview; (b) dorsal view of the carapace.
Invertebrate Reproduction & Development 55
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

dead specimens were identified and preserved in 80%alcohol. For reference, the biological material ofA. rubripes is stored in NEBECC larval collection,(numbers: 0141–0170), Zoology Department,Biosciences Institute, Universidade Estadual Paulista,Botucatu, State of Sao Paulo, Brazil.
After rearing, 130 individuals obtained in thejuvenile stages (about 10 stages), only 100 specimenswere identified as being A. rubripes according to Melo(1996), and also confirmed as such by G.S. Melo fromthe Museum of Zoology of Sao Paulo University(MZUSP). The most important diagnostic features forthis species are: carapace narrowed posteriorly; meso-gastric and cardiac regions well marked; lateral parts
obliquely striated; surface densely punctuated; irregu-lar anteriorly and with short lines of granules; frontapproximately two-thirds of the carapace width (CW);external angle of the orbit, obtuse and short lackingantero-lateral teeth; chelipeds with the superior faceirregular, except the teeth; flat pereopods, being thepropodus of the second pereopod wider and moresetose than the remainder (Melo 1996).
The morphology of megalopa and first juvenile ofA. rubripes was examined and changes in anatomywere recorded for each subsequent one by obtainedstage and sex. Mortality occurred progressively as thejuvenile development proceeded with only 10 crabsreaching the eleventh stage.
Table 1. Armases rubripes (Rathbun 1897): comparison of the original and the present description of megalopa stage.
Structure Diaz and Ewald (1968) This article
Antennule Basal peduncle St: 2; 2; 3 St: 2 pl and 8 ss; 1 ss; 1 ssExopod 3 segments. St: 0; 5 and 2; 5 3 segments. St: 0; 4 ae; 6 ae and
1 pl (long)Maxillule Coxal endite 11 St: 3 pl; 5 pl and 2 ss(10)
Maxilla Coxal endite Proximal lobe 4 ter, 7 sub 3 pd and 1 ss ter, 5 pl and 1 sssub
Distal lobe 2 4 plBasial endite Distal lobe 5 3 pd, 1 sparsely pl, 2 ssEndopod Without setae 1 small sp, 1 plScaphognathite 5 3 ss
Maxilliped I Coxal endite 6 5 ss stout (ter) and 4 ss (sub)Epipod 3 short and 4 long 6 ss (3 short and 3 long)Basial endite 9 7 ss (6 stout and 1 ss)Endopod 2 Unarmed
Maxilliped II Endopod St: 0, 1, 3, 6 St: 0; 1 ss; 2 ss (1 stout); 1 seand 4 cp
Exopod St: 1 prx; 5 dis St: 1 ss prx; 4 ss (long) dis
Maxilliped III Protopod 9 7 pl and 1 ssEpipod 20 (stout), 18 4 pl (long), 13 ss (12 long þ 1
short)Endopod St: 8, 8–9, 3, 4, 6–7 St: 6 ss (stout) and 1 se; 5 se
and 3 ss; 1 se and 2 ss; 3 seand 1 ss; 2 se and 1 ss (sub)and 4 se (ter)
Cheliped (Figure 1a) Ischium Nd 1 spCarpus Nd 2 ss (long and stout)
Second pereopod (Figure 1b) Basis Nd 4 plPropodus Nd 2 ss (stout) and 2 seDactylus Nd 1 ss (stout) and 4 se
Third pereopod (Figure 1c) Propodus Nd 1 ss (stout) and 2 seDactylus Nd 1 ss (stout) and 4 se
Fourth pereopod (Figure 1d) Basis Nd 1 spPropodus Nd 1 ss (stout) and 1 seDactylus Nd 1 ss (stout) and 1 se
Fifth pereopod (Figure 1e) Basis Nd 1 sp (usually)Dactylus 3 hooked setae 3 hooked setae and 1 ss (stout)
Sternum (Figure 2) Nd 4þ 2þ 2 ssTelson 1 pl; 2 lateral, small seta 4 ss each side, 1 terminal and
sp-like seta, 3 central ss
Note: Only the differences concerning setation pattern or additional structures were mentioned.Nd – no data; st – setation; ss – simple setae; pl – plumose setae; pd – plumodenticulate setae; se – serrate setae; ae – aesthetascs;sp – spine; cp – cuspidate; dis – distal; prx – proximal; sub – subterminal; ter – terminal.
56 M.L. Negreiros-Fransozo et al.
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12
About 30 exuviae of megalopae were dissectedusing sharpened needles under a Zeiss (SV6) stereomi-croscope. After this procedure, the appendages wereplaced on a slide covered with glycerin and the cover
was sealed with clear nail varnish. This type ofpreparation is semi-permanent, and lasts for anumber of years.
Using a Zeiss compound microscope (Axioskop 2plus) equipped with a drawing tube, appendages wereanalysed and drawn. Later, all the structures and setaewere observed under another Zeiss microscope
(Axioskop 2) equipped with Normaski differentialinterference contrast, using 1000X lens under immer-sion, for observing the details of setae. Illustrationswere then transferred to a vegetal paper by means of anink pen of 0.25mm.
The terminology adopted in this study uses that sug-gested by Clark et al. (1996) and Pohle et al.(1999). The setae description agrees with Pohle and
Telford (1981). In addition, appendages were describedsequentially by proximal article to the distal one, andsomites were described from anterior to posterior one.Only the setation pattern (number of setae) wascompared with the original paper (Diaz and Ewald1968).
To check for possible effects of laboratory holdingtime on growth, the procedure described by Tweedaleet al. (1993) was adopted, in which an analyses ofcovariance (Sokal and Rohlf 1995) were used tocompare the regression line generated using allmoults with the regression line generated using onlythe first two laboratory moults (the moults presumedto be least affected by laboratory holding time).To define the relationship between moults increment(post-moults CW minus pre-moults CW) and crab size,a regression line was generated, yielding the modelY¼ aþ bX, where Y¼moults increment andX¼pre-moults CW (Hiatt 1948). CW is defined as
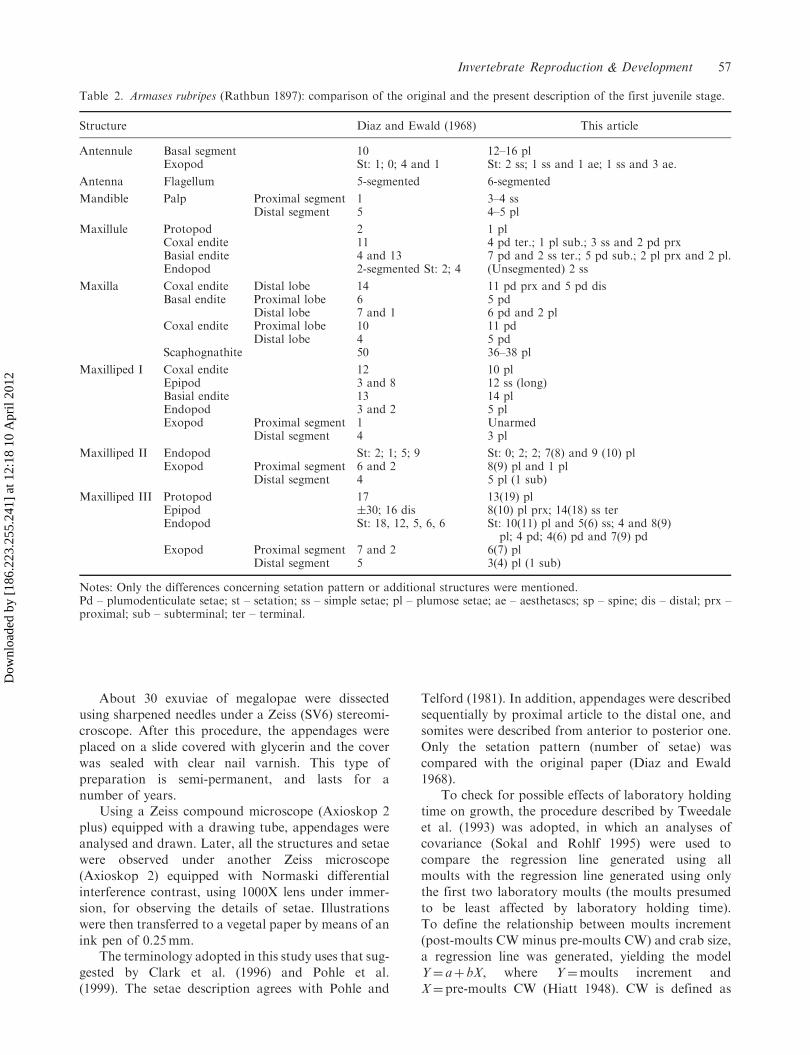
Table 2. Armases rubripes (Rathbun 1897): comparison of the original and the present description of the first juvenile stage.
Structure Diaz and Ewald (1968) This article
Antennule Basal segment 10 12–16 plExopod St: 1; 0; 4 and 1 St: 2 ss; 1 ss and 1 ae; 1 ss and 3 ae.
Antenna Flagellum 5-segmented 6-segmented
Mandible Palp Proximal segment 1 3–4 ssDistal segment 5 4–5 pl
Maxillule Protopod 2 1 plCoxal endite 11 4 pd ter.; 1 pl sub.; 3 ss and 2 pd prxBasial endite 4 and 13 7 pd and 2 ss ter.; 5 pd sub.; 2 pl prx and 2 pl.Endopod 2-segmented St: 2; 4 (Unsegmented) 2 ss
Maxilla Coxal endite Distal lobe 14 11 pd prx and 5 pd disBasal endite Proximal lobe 6 5 pd
Distal lobe 7 and 1 6 pd and 2 plCoxal endite Proximal lobe 10 11 pd
Distal lobe 4 5 pdScaphognathite 50 36–38 pl
Maxilliped I Coxal endite 12 10 plEpipod 3 and 8 12 ss (long)Basial endite 13 14 plEndopod 3 and 2 5 plExopod Proximal segment 1 Unarmed
Distal segment 4 3 pl
Maxilliped II Endopod St: 2; 1; 5; 9 St: 0; 2; 2; 7(8) and 9 (10) plExopod Proximal segment 6 and 2 8(9) pl and 1 pl
Distal segment 4 5 pl (1 sub)
Maxilliped III Protopod 17 13(19) plEpipod �30; 16 dis 8(10) pl prx; 14(18) ss terEndopod St: 18, 12, 5, 6, 6 St: 10(11) pl and 5(6) ss; 4 and 8(9)
pl; 4 pd; 4(6) pd and 7(9) pdExopod Proximal segment 7 and 2 6(7) pl
Distal segment 5 3(4) pl (1 sub)
Notes: Only the differences concerning setation pattern or additional structures were mentioned.Pd – plumodenticulate setae; st – setation; ss – simple setae; pl – plumose setae; ae – aesthetascs; sp – spine; dis – distal; prx –proximal; sub – subterminal; ter – terminal.
Invertebrate Reproduction & Development 57
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

Table
3.Arm
asesrubripes
(Rathbun1897).Measurements
ofjuvenilecrabs(m
ean�
standard
deviation;values
inmillimetres).
Bodyparts
Juvenilecrabstages
MFirst
Second
Third
Fourth
Fifth
Sixth
Seventh
Eighth
Ninth
Tenth
N10
10
10
10
10
10
10
10
98
6CL
1.04�0.06
1.26�0.04
1.51�0.21
1.63�0.36
2.18�0.11
2.54�0.18
3.00�0.20
3.54�0.32
4.07�0.38
4.53�0.39
4.91�0.46
CW
0.88�0.03
1.18�0.05
1.50�0.16
1.91�0.23
2.43�0.05
2.83�0.32
3.32�0.40
4.25�0.33
4.93�0.37
5.70�0.49
6.14�0.66
DO
0.43�0.03
0.62�0.06
0.76�0.07
0.88�0.31
1.36�0.08
1.56�0.16
1.78�0.47
2.44�0.15
3.02�0.48
3.44�0.33
3.74�0.48
AW
0.25�0.02
0.32�0.03
0.38�0.05
0.47�0.09
0.59�0.06
0.64�0.09
0.83�0.13
1.21�0.26
1.55�0.33
1.95�0.50
2.37�0.69
RPL
0.33�0.05
0.43�0.04
0.53�0.10
0.60�0.11
0.73�0.10
0.94�0.15
1.04�0.13
1.55�0.20
1.86�0.67
2.00�0.31
2.24�0.32
RPH
0.16�0.02
0.18�0.01
0.22�0.02
0.28�0.04
0.34�0.02
0.36�0.07
0.48�0.07
0.68�0.12
0.89�0.30
0.92�0.13
1.01�0.30
ML4p
0.43�0.04
0.58�0.09
0.75�0.04
1.03�0.29
1.27�0.22
1.56�0.21
1.85�0.22
2.56�0.21
2.96�0.34
3.38�0.33
3.72�0.67
MW
0.12�0.01
0.18�0.01
0.28�0.02
0.36�0.14
0.48�0.10
0.61�0.07
0.77�0.10
1.01�0.10
1.24�0.10
1.39�0.11
1.59�0.18
GD
––
––
––
–0.06
0.07
0.1
0.18
Notes:N
–number
ofspecim
ens;M
–megalopastage;first,second,third,fourth,fifth,sixth,seventh,eighth,ninth
andtenth
representsjuvenilestages,respectively;CL–carapace
length;CW
–carapace
width;DO
–distance
betweenorbits;AW
–abdominalwidth
(fifth
somite);RPL–rightpropodlength;RPH
–rightpropodheight;ML4p–meruslength
(fourthpereopod);MW
–meruswidth
(fourthpereopod);GD
–gonopore
diameter.
58 M.L. Negreiros-Fransozo et al.
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12
the distance measured between the antero-lateralspines, under a Zeiss microscope stereoscope equippedwith camera and distance measurement system.
Results
Some setal details and additions to the original descrip-tion of megalopa (Figures 1–3) and first juvenile(Figures 4 and 5) are summarized in Tables 1 and 2,respectively. Only the differences concerningsetation, level of details or additional structures aredescribed.
The morphological changes which occurred fromsecond to tenth juvenile stages are shown inFigures 7–9. The external morphology of subsequentjuvenile stages is slightly modified with the increasingsize (Table 3) and setation of certain structures. Thecarapace becomes wider than longer throughout thedevelopment of stages. The buttons on sternitedecrease in size as well as also do ventral depressionson the abdominal sixth somite.
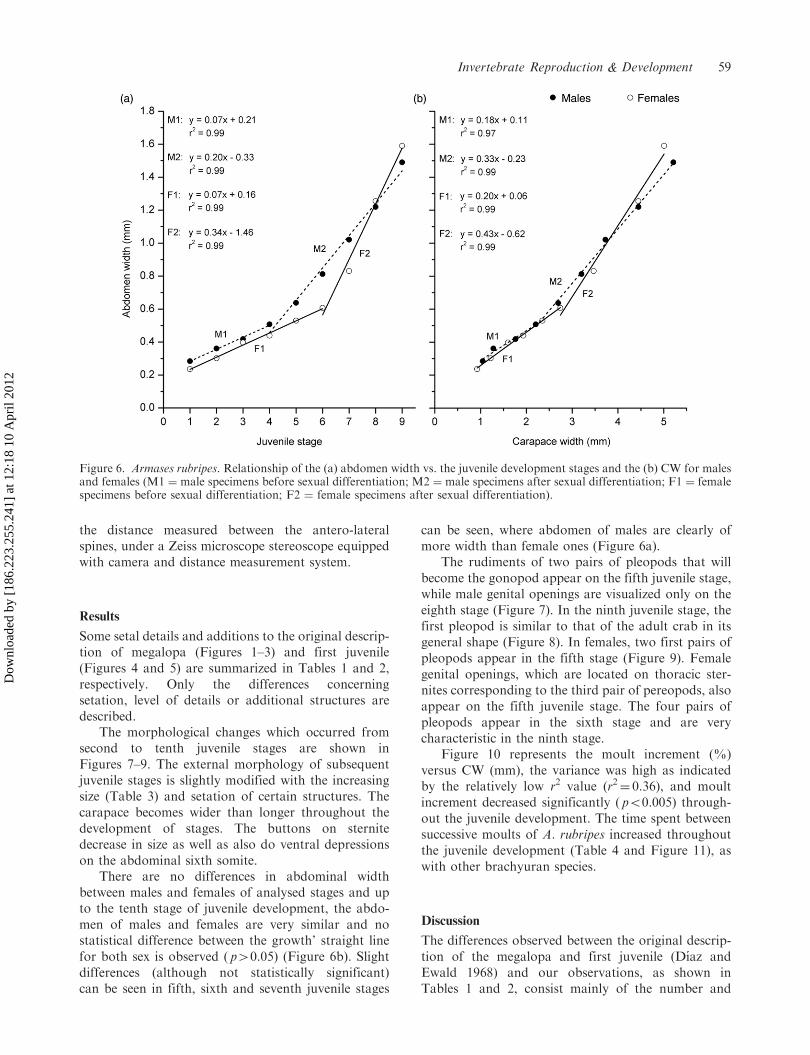
There are no differences in abdominal widthbetween males and females of analysed stages and upto the tenth stage of juvenile development, the abdo-men of males and females are very similar and nostatistical difference between the growth’ straight linefor both sex is observed ( p40.05) (Figure 6b). Slightdifferences (although not statistically significant)can be seen in fifth, sixth and seventh juvenile stages
can be seen, where abdomen of males are clearly ofmore width than female ones (Figure 6a).
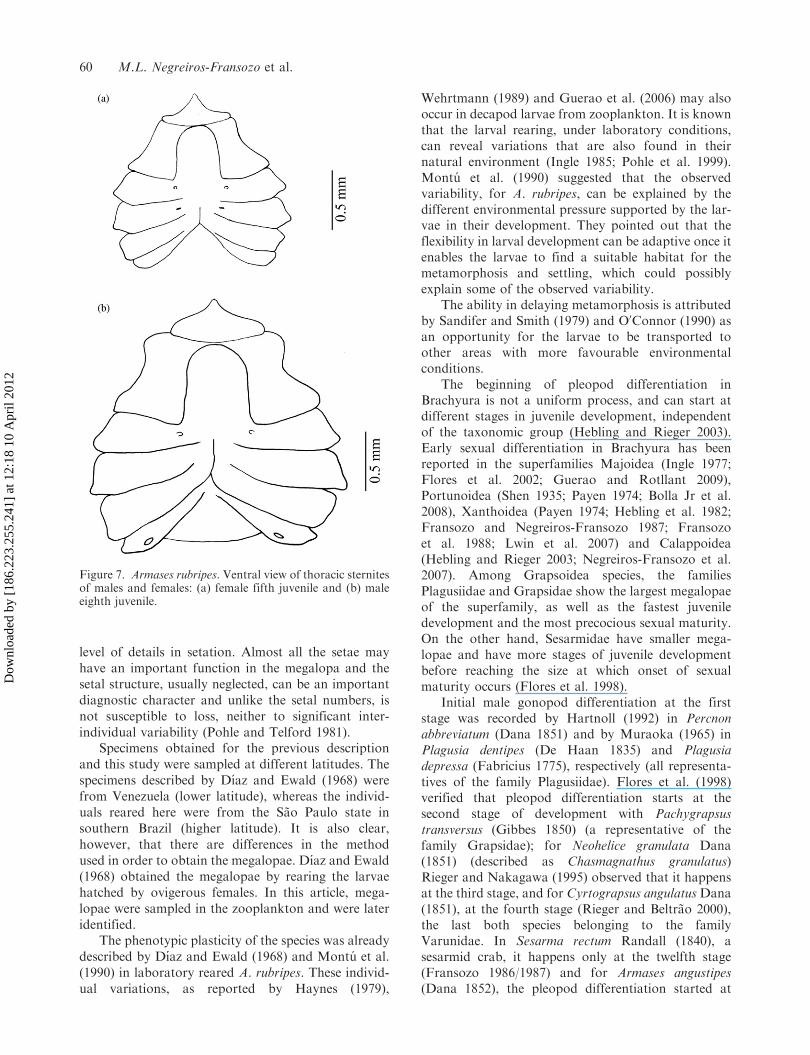
The rudiments of two pairs of pleopods that willbecome the gonopod appear on the fifth juvenile stage,while male genital openings are visualized only on theeighth stage (Figure 7). In the ninth juvenile stage, thefirst pleopod is similar to that of the adult crab in itsgeneral shape (Figure 8). In females, two first pairs ofpleopods appear in the fifth stage (Figure 9). Femalegenital openings, which are located on thoracic ster-nites corresponding to the third pair of pereopods, alsoappear on the fifth juvenile stage. The four pairs ofpleopods appear in the sixth stage and are verycharacteristic in the ninth stage.
Figure 10 represents the moult increment (%)versus CW (mm), the variance was high as indicatedby the relatively low r2 value (r2¼ 0.36), and moultincrement decreased significantly ( p50.005) through-out the juvenile development. The time spent betweensuccessive moults of A. rubripes increased throughoutthe juvenile development (Table 4 and Figure 11), aswith other brachyuran species.
Discussion
The differences observed between the original descrip-tion of the megalopa and first juvenile (Dıaz andEwald 1968) and our observations, as shown inTables 1 and 2, consist mainly of the number and
Figure 6. Armases rubripes. Relationship of the (a) abdomen width vs. the juvenile development stages and the (b) CW for malesand females (M1 ¼ male specimens before sexual differentiation; M2 ¼ male specimens after sexual differentiation; F1 ¼ femalespecimens before sexual differentiation; F2 ¼ female specimens after sexual differentiation).
Invertebrate Reproduction & Development 59
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12
level of details in setation. Almost all the setae mayhave an important function in the megalopa and thesetal structure, usually neglected, can be an importantdiagnostic character and unlike the setal numbers, isnot susceptible to loss, neither to significant inter-individual variability (Pohle and Telford 1981).
Specimens obtained for the previous descriptionand this study were sampled at different latitudes. Thespecimens described by Dıaz and Ewald (1968) werefrom Venezuela (lower latitude), whereas the individ-uals reared here were from the Sao Paulo state insouthern Brazil (higher latitude). It is also clear,however, that there are differences in the methodused in order to obtain the megalopae. Dıaz and Ewald(1968) obtained the megalopae by rearing the larvaehatched by ovigerous females. In this article, mega-lopae were sampled in the zooplankton and were lateridentified.
The phenotypic plasticity of the species was alreadydescribed by Dıaz and Ewald (1968) and Montu et al.(1990) in laboratory reared A. rubripes. These individ-ual variations, as reported by Haynes (1979),
Wehrtmann (1989) and Guerao et al. (2006) may alsooccur in decapod larvae from zooplankton. It is knownthat the larval rearing, under laboratory conditions,can reveal variations that are also found in theirnatural environment (Ingle 1985; Pohle et al. 1999).Montu et al. (1990) suggested that the observedvariability, for A. rubripes, can be explained by thedifferent environmental pressure supported by the lar-vae in their development. They pointed out that theflexibility in larval development can be adaptive once itenables the larvae to find a suitable habitat for themetamorphosis and settling, which could possiblyexplain some of the observed variability.
The ability in delaying metamorphosis is attributedby Sandifer and Smith (1979) and O0Connor (1990) asan opportunity for the larvae to be transported toother areas with more favourable environmentalconditions.
The beginning of pleopod differentiation inBrachyura is not a uniform process, and can start atdifferent stages in juvenile development, independentof the taxonomic group (Hebling and Rieger 2003).Early sexual differentiation in Brachyura has beenreported in the superfamilies Majoidea (Ingle 1977;Flores et al. 2002; Guerao and Rotllant 2009),Portunoidea (Shen 1935; Payen 1974; Bolla Jr et al.2008), Xanthoidea (Payen 1974; Hebling et al. 1982;Fransozo and Negreiros-Fransozo 1987; Fransozoet al. 1988; Lwin et al. 2007) and Calappoidea(Hebling and Rieger 2003; Negreiros-Fransozo et al.2007). Among Grapsoidea species, the familiesPlagusiidae and Grapsidae show the largest megalopaeof the superfamily, as well as the fastest juveniledevelopment and the most precocious sexual maturity.On the other hand, Sesarmidae have smaller mega-lopae and have more stages of juvenile developmentbefore reaching the size at which onset of sexualmaturity occurs (Flores et al. 1998).
Initial male gonopod differentiation at the firststage was recorded by Hartnoll (1992) in Percnonabbreviatum (Dana 1851) and by Muraoka (1965) inPlagusia dentipes (De Haan 1835) and Plagusiadepressa (Fabricius 1775), respectively (all representa-tives of the family Plagusiidae). Flores et al. (1998)verified that pleopod differentiation starts at thesecond stage of development with Pachygrapsustransversus (Gibbes 1850) (a representative of thefamily Grapsidae); for Neohelice granulata Dana(1851) (described as Chasmagnathus granulatus)Rieger and Nakagawa (1995) observed that it happensat the third stage, and for Cyrtograpsus angulatusDana(1851), at the fourth stage (Rieger and Beltrao 2000),the last both species belonging to the familyVarunidae. In Sesarma rectum Randall (1840), asesarmid crab, it happens only at the twelfth stage(Fransozo 1986/1987) and for Armases angustipes(Dana 1852), the pleopod differentiation started at
Figure 7. Armases rubripes. Ventral view of thoracic sternitesof males and females: (a) female fifth juvenile and (b) maleeighth juvenile.
60 M.L. Negreiros-Fransozo et al.
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

sixth stage (Kowalczuk 1994). Guerao et al. (2004)have described the complete larval development andthe first stage of juvenile development of Perisesarmafasciatum (Lanchester 1900), one of the most conspic-uous and species-rich genera of African, Asian and
Figure 8. Armases rubripes. Development of male’s pleopods. pl 1, first pleopod; pl 2, second pleopod; JV, fifth juvenile;JVI, sixth juvenile; JVII, seventh juvenile; JVIII, eighth juvenile; JIX, ninth juvenile.
Figure 9. Armases rubripes. Development of female’s pleo-pods. pl 2, second pleopod; pl 3, third pleopod; pl 4, fourthpleopod; pl 5, fifth pleopod; JV, fifth juvenile; JVI, sixthjuvenile; JVII, seventh juvenile; JVIII, eighth juvenile; JIX,ninth juvenile.
Figure 10. Armases rubripes. Relationship of growth param-eters to crab size (premoult CW for laboratory-rearedjuvenile). Dashed lines means 95% confidence intervals.
Table 4. Armases angustipes. Intermoult period (days)during the early juvenile stages.
Juvenile stages Intermoult period (days) Mean� SD
JI 10.5� 5.13JII 9.3� 3.78JIII 10.9� 3.60JIV 13.6� 4.39JV 14.6� 4.00JVI 18.3� 4.03JVII 20.1� 5.73JVIII 19.4� 5.21
Invertebrate Reproduction & Development 61
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

Australian mangroves. The latter authors verified thatat the first stage pleopods are reduced and until thefourth stage, juveniles grew in size but no changesrelated to the appearing of the secondary sexualcharacters were observed.
This article shows that the beginning of the pleopoddifferentiation in A. rubripes occurs when individualsreach the fifth stage of juvenile development, which isvery similar to that which has been observed forA. angustipes, another species of the same genus, whosesexual differentiation started at the sixth stage ofjuvenile development (Kowalczuk 1994) but differentto that observed by Fransozo (1986/1987) withS. rectum. The related species are phylogeneticallyclose (Schubart et al. 2002) and show megalopae withsizes that are very similar, so the difference at thebeginning of pleopod differentiation among the repre-sentatives of the genus Armases and Sesarma mayindicate an intrinsic strategy for each species, not apattern of the family, as suggested by Flores et al.(1998). This is one more character that demonstratesthe difficulty that taxonomists must face clarifying,more precisely, the position of each genus and familyin the superfamily Grapsoidea that was changedin 2001–2002 (Martin and Davis 2001; Schubartet al. 2002).
The growth parameters documented in laboratorystudies are frequently presumed to be representative ofthe growth parameters of animals in the wild(Tweedale et al. 1993); however, laboratory studies ofgrowth for which there are no comparisons with fieldobservations should be interpreted conservatively(Hartnoll 1982). O’Connor (1990) reported that shereared hundreds of field-caught fiddler crab instars upto the seventh juvenile developmental stage and did notobserve size differences between juvenile Uca pugnaxand Uca pugilator comparable to those seen in crabsreared from eggs in the laboratory. Hartnoll (1982),
however, has observed some differences when thegrowth of crabs reared in laboratory were comparedwith those caught in the field, namely a decrease in themoult increment and an increase in the intermoultperiod.
Although no comparative studies on the growth ofjuveniles reared in the laboratory and those obtainedfrom the nature has been made with A. rubripes, it ispossible that what was observed here (decrease in themoult increment and increase in the intermoult period)is present in individuals in the field. Negreiros-Fransozo and Fransozo (1991) have observed thesame pattern with other brachyuran species of differentfamilies (S. rectum, Eurypanopeus abbreviates, andEriphia gonagra) and also Hirose and Negreiros-Fransozo (2008) in Uca maracoani.
Acknowledgements
The authors are grateful to the Fundacao de Amparo aPesquisa do Estado de Sao Paulo – FAPESP – for thefinancial support (Nos 94/4878-4; 98/3136-4; 04/15194-6),and to FUNDUNESP (No. 00606/05 DFP) to the firstauthor and to FAPESP for a scholarship to the third author.The authors are grateful to anonymous reviewers for theiruseful suggestions and NEBECC members for their helpduring field and laboratory activities. Sampling was per-formed according to Brazilian state and federal lawsconcerning wild animals.
References
Bolla Jr EA, Negreiros-Fransozo ML, Fransozo A. 2008.
Juvenile development of Callinectes ornatus Ordway, 1863
(Crustacea: Decapoda: Portunidae), from megalopae
obtained in the neuston. Zootaxa. 1788:1–20.Cadman LR, Weinstein MP. 1988. Effects of temperature
and salinity on the growth of laboratory-reared juvenile
blue crabs Callinectes sapidus Rathbun. Journal of
Experimental Marine Biology and Ecology. 121:193–207.Clark PF, Calazans DK, Pohle GW. 1996. Accuracy and
standardization of Brachyuran larval descriptions.
Invertebrate Reproduction and Development.
33(2–3):127–144.Costlow Jr JD, Bookhout CG. 1960. The complete larval
development of Sesarma cinereum (Bosc) reared in the
laboratory. Biological Bulletin. 118(2):203–214.Costlow Jr JD, Bookhout CG. 1962. The larval development
of Sesarma reticulatum Say reared in the laboratory.
Crustaceana. 4:281–294.
Cuesta JA, Anger K. 2001. Larval morphology of the
Sesarmid crab Armases angustipes Dana, 1852 (Decapoda,
Brachyura, Grapsoidea). Journal of Crustacean Biology.
21(3):821–838.Cuesta JA, Garcıa-Guerrero MU, Rodrıguez A,
Hendrickx ME. 2006. Larval morphology of the sesarmid
crab Aratus pisonii (H. Milne Edwards, 1837) (Decapoda,
Brachyura, Grapsoidea), from laboratory reared material.
Crustaceana. 79(2):175–196.
Figure 11. Armases rubripes. Relationship between inter-moult period (days) and CW throughout the early juveniledevelopment.
62 M.L. Negreiros-Fransozo et al.
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

Dıaz H, Ewald JJ. 1968. A comparison of the larval
development of Metasesarma rubripes (Rathbun) and
Sesarma ricordi H. Milne Edwards (Brachyura,
Grapsidae) reared under laboratory conditions.
Crustaceana. 2:225–248.Felder DL, Martin JW, Goy JW. 1985. Patterns in early
postlarval development of decapods. In: Wenner AM,
editor. Crustacean issues II. Larval growth. Boston (MA):
Balkema. p. 163–225.Flores AAV, Marques FPL, Negreiros-Fransozo ML. 2002.
Postlarval stages and growth patterns of the spider crab
Pyromaia tuberculata (Brachyura, Majidae) from labora-
tory-reared material. Journal of Crustacean Biology.
22(2):314–327.
Flores AAV, Negreiros-Fransozo ML, Fransozo A. 1998.
The megalopa and juvenile development of Pachygrapsus
transversus (Gibbes 1850) (Decapoda, Brachyura) com-
pared with other grapsid crabs. Crustaceana.
71(2):197–222.
Fransozo A. 1986/1987. Desenvolvimento dos estagios
juvenis de Sesarma (Holometopus) rectum Randall, 1840
(Decapoda, Grapsidae) obtidos em laboratorio. Naturalia.
11/12:77–87.Fransozo A, Hebling NJ. 1986. Desenvolvimento larval de
Sesarma (Holometopus) rectum, Randall 1840 (Decapoda,
Grapsidae) em laboratorio. Revista Brasileira de Biologia.
46(2):353–364.Fransozo A, Negreiros-Fransozo ML. 1987. Morfologia dos
primeros estagios juvenis de Eriphia gonagra (Fabricius
1781) e Eurypanopeus abbreviatus (Stimpson 1860)
(Crustacea, Decapoda, Xanthidae), obtidos em labora-
torio. Papeis Avulsos de Zoologia. 36(22):257–277.
Fransozo A, Negreiros-Fransozo ML, Hiyodo CM. 1988.
Developpement juvenile de Menippe nodifrons Stimpson,
1859 (Crustacea, Decapoda, Xanthidae) au laboratoire.
Revue de Hydrobiologie Tropical. 21(4):297–308.Guerao G, Abello P, dos Santos A. 2006. Morphological
variability of the megalopa of Liocarcinus depurator
(Brachyura: Portunidae) in Mediterranean and Atlantic
populations. Journal of Natural History.
40(32–34):1851–1866.Guerao G, Anger K, Nettelmann UWE, Schubart CD. 2004.
Complete larval and early juvenile development of the
mangrove crab Perisesarma fasciatum (Crustacea:
Brachyura: Sesarmidae) from Singapore, with a larval
comparison of Parasesarma and Perisesarma. Journal of
Plankton Research. 26(12):1389–1408.Guerao G, Anger K, Schubart CD. 2007. Larvae and
first-stage juveniles of the American genus Armases
Abele 1992 (Brachyura: Sesarmidae): a morphological
description of two complete developments and one first
zoeal stage. Journal of Natural History. 41(29–32):
1811–1839.
Guerao G, Rotllant G. 2009. Post-larval development and
sexual dimorphism of the spider crab Maja brachydactyla
(Brachyura: Majidae). Scientia Marina. 73(4):797–808.Guimaraes FJ, Negreiros-Fransozo ML. 2005. Juvenile
development and growth patterns in the mud crab
Eurytium limosum (Say 1818) (Decapoda, Brachyura,
Xanthidae) under laboratory conditions. Journal of
Natural History. 39(23):2145–2161.
Hartnoll RG. 1982. Growth. In: Abele LG, editor. Vol. 2.
The biology of crustacea. New York: Academic Press.
p. 111–196.Hartnoll RG. 1992. Megalopae and early post-larval stages
of East African Percnon abbreviatum (Decapoda:
Brachyura: Grapsidae). Journal of Zoology. 228:51–67.
Haynes E. 1979. Description of larvae of the northern
shrimp, Pandalus borealis, reared in situ in Kachemak Bay,
Alaska. Fishery Bulletin. 77:157–173.Hebling NJ, Fransozo A, Negreiros-Fransozo ML. 1982.
Desenvolvimento dos primeiros estagios juvenis de
Panopeus herbstii H. Milne Edwards, 1834 (Crustacea,
Decapoda, Xanthidae), criados em laboratorio. Naturalia.
7:177–188.
Hebling NJ, Rieger PJ. 2003. Desenvolvimento juvenil de
Hepatus pudibundus (Herbst) (Crustacea, Decapoda,
Calappidae), em laboratorio. Revista Brasileira de
Zoologia. 20(3):531–539.Hiatt RW. 1948. The biology of the lined shore crab
Pachygrapsus crassipes Randall. Pacific Science.
11:135–212.Hirose GL, Negreiros-Fransozo ML. 2008. Growth and
juvenile development of Uca maracoani Latreille,
1802-1803 in laboratory conditions (Crustacea,
Decapoda, Brachyura, Ocypodidae). Senckenbergiana
biologica. 88(2):161–168.Ingle RW. 1977. The larval and post-larval development of
the scorpion spider crab, Inachus dorsettensis (Pennant)
(family: Majidae), reared in the laboratory. Bulletin of the
British Museum of Natural History (Zoology).
30:331–348.
Ingle RW, Rice AL. 1984. The juvenile stages of eight
swimming crab species (Crustacea: Brachyura:
Portunidae): a comparative study. Bulletin of the British
Museum of Natural History (Zoology). 46(4):345–354.Ingle RW. 1985. Larval development of the mud crab
Panopeus occidentalis de Saussure, from Bermuda
(Crustacea: Xanthoidea: Panopeidae). Bulletin of the
British Museum of Natural History (Zoology).
48(4):233–248.Kowalczuk VGL. 1994. Estrutura populacional de Armases
angustipes (Dana 1852) (Decapoda: Brachyura: Grapsidae)
da Ilha do Farol, Caioba, PR e seu desenvolvimento pos-
embrionario sob condicoes de laboratorio [unpublished
MSc dissertation]. [Curitiba, Brazil]: Universidade Federal
do Parana, p. 100.Luppi IA, Spivak ED. 2007. Morphology of megalopa and
first crab of Cyrtograpsus angulatus, with comments on the
presence of an anomalous first crab stage on Brachyuran
crabs. Journal of Crustacean Biology. 27(1):80–89.Lwin TT, Doi W, Yokota M, Strussmann CA, Watanabe S.
2007. Juvenile morphology of the xanthid crab Leptodius
exaratus (H. Milne-Edwards 1834) (Decapoda:
Brachyura), with notes on the appearance of sexual
dimorphism. Invertebrate Reproduction and
Development. 50(4):191–201.Martin JW, Davis GE. 2001. An updated classification of the
recent Crustacea. Natural Museum of Los Angeles
County, Science Series. 39:1–124.Martin JW, Felder DL, Truesdale FM. 1984. A comparative
study of morphology and ontogeny in juvenile stages
of four Western Atlantic xanthoid crabs
Invertebrate Reproduction & Development 63
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12

(Crustacea: Decapoda: Brachyura). PhilosophicalTransactions of the Royal Society of London.
303:537–604.Melo GAS. 1996. Manual de identificacao dos Brachyura(Caranguejos e Siris) do litoral brasileiro. Sao Paulo:Pleiade/FAPESP, 603p.
Montu M, Anger K, Bakker C. 1990. Variability in the larvaldevelopment of Metasesarma rubripes (Decapoda,Grapsidae) reared in the laboratory. Nerıtica.
5(1):113–128.Muraoka K. 1965. On the post-larval stage of Plagusiadepressa. Research on Crustaceans. 2:83–90.
Negreiros-Fransozo ML, Fransozo A. 1991. Growth and agedetermination of three juvenile crab species (Crustacea,Decapoda, Brachyura). Papeis Avulsos de Zoologia.37(18):277–283.
Negreiros-Fransozo ML, Wenner EL, Knott DM,Fransozo A. 2007. The megalopa and early juvenilestages of Calappa tortugae Rathbun, 1933 (Crustacea,
Brachyura) reared in the laboratory from South Carolinaneuston samples. Proceedings of the Biological Society ofWashington. 120(4):469–485.
O’Connor NJ. 1990. Morphological differentiation andmoulting of juvenile fiddler crabs (Uca pugilator andU. pugnax). Journal of Crustacean Biology. 10(4):608–612.
Payen G. 1974. Morphogenese sexuelle de quelquesBrachyoures (Cyclometopes) au cours du developpementembryonnaire, larvaire et postlarvaire. Bulletin duMuseum Nationale d’Histoire Naturelle, (3n serie), 209,
Zoology. 139:202–261.Pohle G, Mantelatto FLM, Negreiros-Fransozo ML,fransozo A. 1999. Larval Decapoda (Brachyura).
In: Boltovskoy D, editor. Vol. 2. South AtlanticZooplankton. Rotterdam, The Netherlands: Backhuys.p. 1280–1351.
Pohle G, Telford M. 1981. Morphology and classification ofdecapod crustacean larval setae: a scanning electron
microscope study of Dissodactylus crinitichelis Moreira,1901 (Brachyura, Pinnotheridae). Bulletin of Marine
Science. 31(3):736–752.Rieger PJ, Beltrao R. 2000. Desenvolvimento juvenil deCyrtograpsus angulatus Dana (Crustacea, Decapoda,Grapsidae), em laboratorio. Revista Brasileira de
Zoologia. 17(2):405–420.Rieger PJ, Nakagawa C. 1995. Desenvolvimento juvenil deChasmagnathus granulata Dana, 1851 (Crustacea,
Decapoda, Grapsidae), em laboratorio. Nauplius. 3:59–74.Sandifer PA, Smith IJ. 1979. Possible significance ofvariation in the larval development of Palaemonid
shrimp. Journal of Experimental Marine Biology andEcology. 39:55–64.
Schubart CD, Cuesta JA. 1998. First zoeal stages of fourspecies of Sesarma from Panama (Crustacea, Brachyura,
Grapsidae) described from the laboratory hatched materialwith a identification key and phylogenetic remarks.Journal of Plankton Research. 20(1):61–84.
Schubart CD, Cuesta JA, Diesel R. 2002. Glyptograpsidae, anew brachyuran family from Central America: larval andadult morphology, and a molecular phylogeny of the
Grapsoidea. Journal of Crustacean Biology. 22:28–44.Shen CJ. 1935. An investigation of the post-larval develop-ment of the shore-crab Carcinus maenas, with the special
reference to the external secondary sexual characters.Proceedings of the Zoological Society of London. 1:1–33.
Sokal RR, Rohlf FJ. 1995. Biometry: the principles andpractice of statistic in biological research. New York:
W. H. Freeman. p. 887.Tweedale WA, Bert TM, Brown SD. 1993. Growth ofpostsettlement juveniles of the Florida stone crab,Menippe
mercenaria (Say) (Decapoda: Xanthidae) in the laboratory.Bulletin of Marine Science. 52(3):873–885.
Wehrtmann I. 1989. Seasonal occurrence and abundance of
caridean larvae of Helgoland. German Bight. HelgolanderMeeresunters. 43:87–112.
64 M.L. Negreiros-Fransozo et al.
Dow
nloa
ded
by [
186.
223.
255.
241]
at 1
2:18
10
Apr
il 20
12
Copyright © 2022 FDOKUMEN




















