
Dynamics of a two-prey–one-predator system with predator switching regulated by a catabolic...
13
Ecological Modelling 186 (2005) 345–357 Dynamics of a two-prey–one-predator system with predator switching regulated by a catabolic repression control-like mode D.V. Vayenas a , G. Aggelis b , V. Tsagou b , S. Pavlou c,d,∗ a Department of Environmental and Natural Resources Management, School of Natural Resources and Enterprise Management, University of Ioannina, Agrinion, Greece b Laboratory of General and Agricultural Microbiology, Department of Agricultural Biotechnology, Agricultural University of Athens, Athens, Greece c Department of Chemical Engineering, University of Patras, GR-26500 Patras, Greece d Institute of Chemical Engineering and High Temperature Chemical Processes, FORTH, Patras, Greece Received 3 June 2004; received in revised form 10 January 2005; accepted 17 January 2005 Available online 3 March 2005 Abstract The strategy of alternative prey (switching), regulated by a catabolic repression control-like (CRCL) mode, in which the desirable prey acts as repressor on the predator’s attack on the alternative prey, was studied in a two-prey–one-predator chemostat system. In this system, alternative prey has to survive predation and competition for nutrients from the desirable prey. Therefore, when the alternative prey has no competitive advantage for the common substrate over the desirable prey, its survival depends on the protection offered by the desirable prey via CRCL. In this case, CRCL allows the coexistence of both desirable and alternative prey and predator populations. However, when the alternative prey has the competitive advantage over the desirable prey, CRCL negatively affects both the state of survival of the desirable prey and the coexistence state. © 2005 Elsevier B.V. All rights reserved. Keywords: Prey–predator dynamics; Switching; Alternative prey; Catabolic repression control mode; Bifurcations 1. Introduction Several strategies have been proposed so far, to il- lustrate the balanced attack of predators on their prey in a natural ecosystem (Alexander, 1981). Examples include competition among predators in high preda- ∗ Corresponding author. Tel.: +30 2610997640; fax: +30 2610997640. E-mail address: [email protected] (S. Pavlou). tors density, predators under biological control, genetic adaptation of the prey population, prey refuge in some places of the ecosystem, switching (facultative preda- tion, alternative prey strategy), etc. In a previous paper we studied the facultative predation strategy using an one-prey–one-facultative- predator dynamic model (Aggelis et al., 2005). This model assumed that both prey and predator popula- tions grow together in a chemostat, which is fed with medium containing the necessary nutrients for sapro- 0304-3800/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.ecolmodel.2005.01.032
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Dynamics of a two-prey–one-predator system with predator switching regulated by a catabolic...

Ecological Modelling 186 (2005) 345–357
Dynamics of a two-prey–one-predator system with predatorswitching regulated by a catabolic repression control-like mode
D.V. Vayenasa, G. Aggelisb, V. Tsagoub, S. Pavlouc,d,∗
a Department of Environmental and Natural Resources Management, School of Natural Resources and Enterprise Management,University of Ioannina, Agrinion, Greece
b Laboratory of General and Agricultural Microbiology, Department of Agricultural Biotechnology,Agricultural University of Athens, Athens, Greece
c Department of Chemical Engineering, University of Patras, GR-26500 Patras, Greeced Institute of Chemical Engineering and High Temperature Chemical Processes, FORTH, Patras, Greece
Received 3 June 2004; received in revised form 10 January 2005; accepted 17 January 2005Available online 3 March 2005
Abstract
The strategy of alternative prey (switching), regulated by a catabolic repression control-like (CRCL) mode, in which thedesirable prey acts as repressor on the predator’s attack on the alternative prey, was studied in a two-prey–one-predator chemostatsystem. In this system, alternative prey has to survive predation and competition for nutrients from the desirable prey. Therefore,when the alternative prey has no competitive advantage for the common substrate over the desirable prey, its survival dependso rable anda e desirablep©
K
1
lii
f
eticomeeda-
tivetive-
pula-ithpro-
0
n the protection offered by the desirable prey via CRCL. In this case, CRCL allows the coexistence of both desilternative prey and predator populations. However, when the alternative prey has the competitive advantage over threy, CRCL negatively affects both the state of survival of the desirable prey and the coexistence state.2005 Elsevier B.V. All rights reserved.
eywords:Prey–predator dynamics; Switching; Alternative prey; Catabolic repression control mode; Bifurcations
. Introduction
Several strategies have been proposed so far, to il-ustrate the balanced attack of predators on their preyn a natural ecosystem (Alexander, 1981). Examplesnclude competition among predators in high preda-
∗ Corresponding author. Tel.: +30 2610997640;ax: +30 2610997640.
E-mail address:[email protected] (S. Pavlou).
tors density, predators under biological control, genadaptation of the prey population, prey refuge in splaces of the ecosystem, switching (facultative prtion, alternative prey strategy), etc.
In a previous paper we studied the facultapredation strategy using an one-prey–one-facultapredator dynamic model (Aggelis et al., 2005). Thismodel assumed that both prey and predator potions grow together in a chemostat, which is fed wmedium containing the necessary nutrients for sa
304-3800/$ – see front matter © 2005 Elsevier B.V. All rights reserved.doi:10.1016/j.ecolmodel.2005.01.032

346 D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357
phytic growth of both populations. The attack of thefacultative predator on the prey population is regulatedby the abundance of the common resource for sapro-phytic growth, via a catabolic repression mechanism.The common substrate for saprophytic growth of bothpopulations acted as repressor on the attack of preda-tors on the prey population. It was demonstrated thatthe catabolic repression control favors domination ofthe prey over the predator and, under certain circum-stances (e.g. when the predator has the competitive ad-vantage at high substrate concentration) coexistence ofboth prey and predator populations.
Another interesting strategy, which allows the coex-istence of prey and their predators in the natural ecosys-tems, is that of the alternative predation (Verity, 1991;Morin and Lawler, 1995; Flynn et al., 1996; Fagan,1997; Holyoak and Sachdev, 1998; Morin, 1999). Sev-eral authors have proposed models in order to describethe balanced attack of the predators on their alterna-tive prey. Namely, the effects of adaptive switchingon the Lotka–Volterra population dynamics have beenstudied byKrivan (1996, 1997a, 1997b, 1998), Krivanand Sikder (1999), Krivan and Eisner (2003)andVanBaalen et al., (2001). It has been demonstrated thatthe optimal behavior of animals leads to persistenceof predator–prey systems and reduction of oscillationsin population densities.Pelletier (2000)suggested thatin multi-species Lotka–Volterra systems with predatorswitching, predators have a stabilizing effect on theecosystem, irrespective of diversity and/or connected-n dc sys-t dt enceo
bys ajorm msi -g gi, al-g ha-r ro-p iffer-e de-s thano siz-i iza-t 6;
Hraber and Milne, 1997). Control modes as inductionand repression, however, act at the level of genes andcause changes in the biosynthesis rate of these enzymes(Abrams, 1980; Bailey and Ollis, 1986). In a similarmannerBrandt et al. (2004)suggested that in the natu-ral environment microorganisms encounter changes insubstrate availability, involving either nutrient concen-tration or nutrient types. They have, therefore, to adaptto the new conditions in order to survive. The microbialgrowth on mixed substrates plays a crucial role in thebalance of nature and has been investigated by severalauthors (see, e.g.,Kompala et al., 1986; Egli, 1995;Brandt et al., 2004).
In the present paper a catabolic repression control-like (CRCL) mode is introduced in a two-prey–one-predator dynamic system. The two prey populations(desirable and alternative) grow in the same chemostattogether with their predator, while the desirable preypopulation represses the attack of the predator on thealternative prey through CRCL. The effect of switch-ing, as regulated by CRCL, on the dynamics and sur-vival of both prey and predator populations is studiedusing bifurcation analysis and is discussed, with somepractical conclusions of ecological importance beingdrawn.
2. Regulation of alternative prey strategy andmodeling
a witht et byt sentst at-t ep y ar s as-s es oft d forN
tionm monr s thep onsb andc s as-
ess.Kooi et al. (1999)have shown that short foohains are resistant to predators (when invade in aem), whileKooi and Kooijman (2000)demonstratehat the presence of a predator can allow coexistf two competing prey populations.
The enzymatic destruction of an organismpecies of another organism appears to be a mechanism for the killing of several microorganis
n natural ecosystems (Alexander, 1981). The microoranisms that possess resistant structures (e.g. funae) contain in their cell walls structural polysaccides, depolymerization of which results from appriate enzymes excreted by the predator. Since dnt enzymes are generally required for each preytruction, the predators that can consume morene prey must be sufficiently adaptive (by synthe
ng appropriate enzyme systems) for effective utilion of the prey mass (Saint-Cyr and Cloutier, 199
It is assumed that two preys (N1 and N2) grows saprophytes in the same chemostat together
heir predator (P). N1, having a cell wall structurhat is more accessible to the depolymerizationhe enzymes excreted by the predator, reprehe desirable prey. The predator will graduallyack the alternative preyN2 when the density of thopulationN1 decreases, as this is regulated bepression–derepression control mechanism. It iumed that the desirable prey represses the genhe predator responsible for the enzymes require2 destruction.This assumption resembles the intraguild preda
ode where both prey and predator feed on a comesource. In general, intraguild predation decreaserobability for stable coexistence of the populatiecause the prey has to survive both predationompetition for the common resource. However, a

D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357 347
sumed here, the desirable prey exerts two opposite ef-fects on the alternative prey population: a negative one,since both desirable and alternative prey populationscompete for the common resource, and a positive one,since, as the density of the desirable prey populationincreases, the predator’s attack on the alternative preyis gradually repressed via CRCL.
The equations of the model are as follows:Balance of the desirable prey population:
dN1
dt= −DN1 + rN1(S)N1 − 1
YP/N1
qP1(N1)P (1)
Balance of the alternative prey population:
dN2
dt= −DN2 + rN2(S)N2 − 1
YP/N2
qP2(N2, Q)P
(2)
Balance of the predator population:
dP
dt= −DP + qP1(N1)P + qP2(N2, Q)P (3)
Balance of the limiting factor for saprophytic growthof N1 andN2:
dS
dt=D(SF − S)− 1
YN1/S
rN1(S)N1 − 1
YN2/S
rN2(S)N2
(4)
whereD is the chemostat dilution rate;rN1(S) andrN2(S) the specific rates for saprophytic growth ofN1andN2, respectively;qP (N1) andqP (N2, Q) are thesayta ndo ons,t aren
thep yM
r
wt thr c-t
The specific growth rate of the predator on the de-sirable prey,qP1(N1), is given by
qP1(N1) = qP1,maxN1
KPN1+ N1
(6)
whereqP1,max is the maximum specific growth rate ofP grown onN1; KPN1
the saturation constant forN1.The specific growth rate of the predator on the al-
ternative prey,qP2(N2, Q), is given by
qP2(N2, Q) = qP2,maxN2
KPN2+ N2
Q (7)
whereqP2,max is the maximum specific growth rate ofPgrown onN2; KPN2
the saturation constant forN2; Qis a function of the desirable prey (N1).
The factorQ accounts for modification of the rateof predation on theN2 population by repressing en-zyme synthesis (indispensable forN2 destruction) viaCRCL. Originally the catabolic repression mechanismrefers to the repression of the transcription of genescoding for certain inducible enzyme systems by severalreadily utilizable carbon sources (for the mechanismof catabolic repression see,Imanaka and Aiba, 1977;Bailey and Ollis, 1986; Atlas, 1997). We assume herethat some metabolic products, excreted by the desirableprey in the growth environment, may act as repressorson the predator’s attack on the alternativeN1 prey popu-lation. Since the concentration of the repressors (extra-c eN sedf
Q
w nderc
mesw ificr r’sy
e-dwt re-q ina y.
1 2
pecific growth rates of the predator population onN1ndN2, respectively;YN1/S andYN2/S theN1 andN2ield coefficients onS, respectively;YP/N1 andYP/N2
heP yield coefficients onN1 andN2, respectively;SFndS the limiting factor concentration in the inlet autlet medium, respectively. In the above equati
erms of maintenance and death of all populationseglected.
The specific rates for saprophytic growth ofrey populations (rN1(S) andrN2(S)) are modeled bonod’s model (Monod, 1942):
Ni (S) = rNi,maxS
KNiS + S(5)
herei = 1 for the desirable prey andi = 2 for the al-ernative prey;rNi,max is the maximum specific growate;KNiS the saturation constant for the limiting faor.
ellular products ofN1) could be linearly related to th1 population density, the following equation was u
or Q (Aggelis et al., 2005):
(N1) = 1
1 + BN1(8)
hereB is a parameter related to operators found uatabolic repression.
Assuming that CRCL affects synthesis of enzyhich are related to the killing of the prey, the spec
ate of predation (qP2,max) rather than the predatoield should be affected byQ.
From Eqs.(7) and (8) it is apparent that the pration ability ofP on N2 is repressed whenB→ ∞,hereas this ability is maximized whenB→ 0. When
he density ofN1 decreases, the enzyme synthesisuired for killingN2 is gradually increased resultingn increase of the predation on the alternative pre
348 D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357
3. Methods
The long-term behavior of the system depends onthe values of its parameters. Of importance is the effectof the operating parameters and in particular of thechemostat dilution rateD, controlled by the flowratethrough the reactor. Another important parameter isB,which determines the extent of predator’s attack on theN2 prey or, in other words, the genetic adaptation ofthe predators to coexist in the same environment withtheir prey.
An illustrative way to present the effect of thoseparameters on the system is through bifurcation dia-grams. In such a diagram each region represents pa-rameter values for which the system exhibits differentdynamic behavior. The boundaries of those regions arelocations where bifurcations occur and the system un-dergoes a qualitative change in its dynamic behavior.This qualitative change consists in the change of thenumber or the character (stability) of steady and pe-riodic states. Construction of a bifurcation diagram isachieved by tracing the location of bifurcations in thetwo-parameter space. This is accomplished with thehelp of numerical algorithms that we have developedand are based on the continuation algorithm AUTO(Doedel et al., 1997).
For the above-mentioned system of alternative preystrategy there are six possible steady states:
(1) Extinction of all populations:N1 =N2 =P= 0
( .( .(
(
(
lity ofso inw varyp
emd n thes . Of
Table 1Sets of parameter values selected for the study of the effect ofcatabolic repression control-like mode on the population dynamics
Parameter values
A B C D
rNi,max (h−1) 0.4 0.4 0.4 0.4qPi,max (h−1) 0.3 0.3 0.3 0.3KN1S (mg/L) 0.4 0.8 0.4 0.8KN2S (mg/L) 0.6 0.6 0.6 0.6KPN1
(mg/L) 0.7 0.7 0.4 0.4KPN2
(mg/L) 0.4 0.4 0.7 0.7YNi/S 0.3 0.3 0.3 0.3YP/Ni
0.4 0.4 0.4 0.4
i = 1 for the desirable prey;i = 2 for the alternative prey.
particular importance are the parameter values in thespecific growth rate of the predator, since faster growthof the predator on the desirable prey or on the alter-native prey yields qualitative different cases. Also, theparameter values of the specific growth rates of thetwo prey populations play a very important role, sincethese values determine which of the two populationsgrows faster on the substrate and thus has the compet-itive advantage. The population dynamics was studiedfor all possible qualitatively different cases, the param-eter values of which are listed inTable 1. Selection ofthe prey for which the predator exhibits higher growthrate can be done by appropriate choice of the values ofthe saturation constantsKPN1
andKPN2in the specific
growth rate of the predator on each prey. Selection ofthe population that has the competitive advantage be-tween the two prey populations can be done by appro-priate choice of the values of the saturation constantsKN1S andKN2S in the specific growth rates of the preypopulations on the limiting nutrient.
4. Results
4.1. Predator grows faster on the alternative thanon the desirable prey (KPN1
> KPN2): cases A and
B
4.1.1. Case A: competitive advantage of thedesirable prey over the alternative prey population(
w e de-
(washout state).2) Survival ofN1 only:N1 > 0,N2 = 0,P= 0 (N1 state)3) Survival ofN2 only:N1 = 0,N2 > 0,P= 0 (N2 state)4) Survival ofN1 andP only: N1 > 0, N2 = 0, P> 0
(N1P state).5) Survival ofN2 andP only: N1 = 0, N2 > 0, P> 0
(N2P state).6) Coexistence of all three populations:N1 > 0,
N2 > 0,P> 0.
Besides these steady states, there is the possibitable periodic states of survival ofN1 andP, of survivalf N2 andP, or of coexistence of all populations,hich the population densities in the chemostateriodically with time.
The form of the bifurcation diagram of the systepends on the values of the kinetic parameters ipecific growth rate expressions of the populations
KN1S < KN2S)In case A ofTable 1theKS values forN1 andN2
ere selected so that the specific growth rate of th
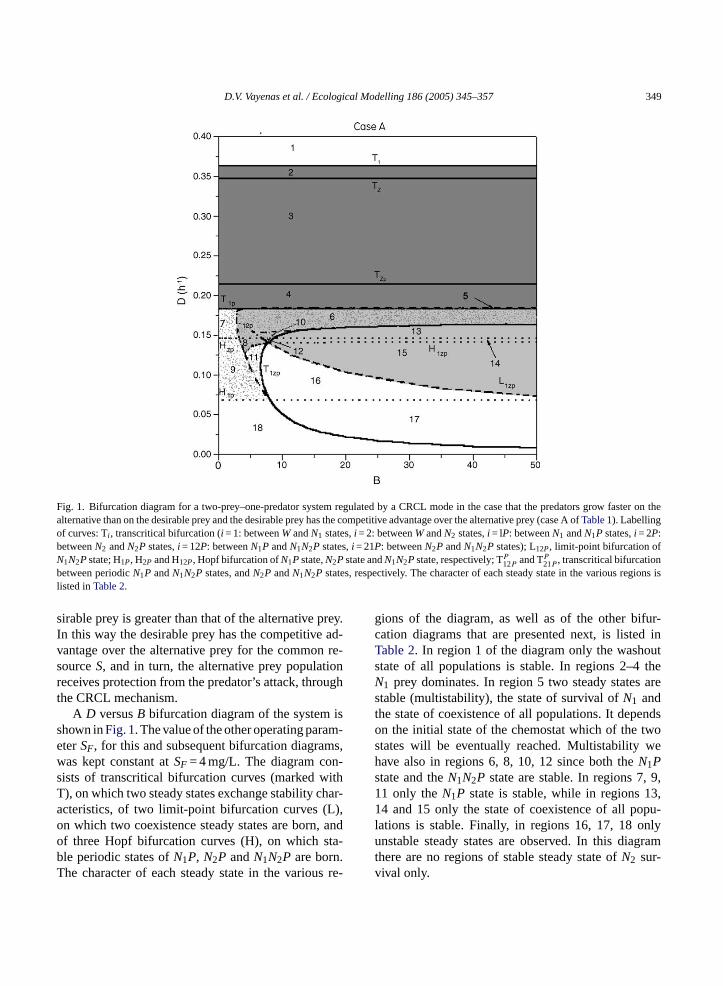
D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357 349
Fig. 1. Bifurcation diagram for a two-prey–one-predator system regulated by a CRCL mode in the case that the predators grow faster on thealternative than on the desirable prey and the desirable prey has the competitive advantage over the alternative prey (case A ofTable 1). Labellingof curves: Ti , transcritical bifurcation (i = 1: betweenWandN1 states,i = 2: betweenWandN2 states,i = lP: betweenN1 andN1P states,i = 2P:betweenN2 andN2P states,i = 12P: betweenN1P andN1N2P states,i = 21P: betweenN2P andN1N2P states); L12P, limit-point bifurcation ofN1N2Pstate; H1P, H2P and H12P, Hopf bifurcation ofN1Pstate,N2Pstate andN1N2Pstate, respectively; TP12P and TP
21P , transcritical bifurcationbetween periodicN1P andN1N2P states, andN2P andN1N2P states, respectively. The character of each steady state in the various regions islisted inTable 2.
sirable prey is greater than that of the alternative prey.In this way the desirable prey has the competitive ad-vantage over the alternative prey for the common re-sourceS, and in turn, the alternative prey populationreceives protection from the predator’s attack, throughthe CRCL mechanism.
A D versusB bifurcation diagram of the system isshown inFig. 1. The value of the other operating param-eterSF, for this and subsequent bifurcation diagrams,was kept constant atSF = 4 mg/L. The diagram con-sists of transcritical bifurcation curves (marked withT), on which two steady states exchange stability char-acteristics, of two limit-point bifurcation curves (L),on which two coexistence steady states are born, andof three Hopf bifurcation curves (H), on which sta-ble periodic states ofN1P, N2P andN1N2P are born.The character of each steady state in the various re-
gions of the diagram, as well as of the other bifur-cation diagrams that are presented next, is listed inTable 2. In region 1 of the diagram only the washoutstate of all populations is stable. In regions 2–4 theN1 prey dominates. In region 5 two steady states arestable (multistability), the state of survival ofN1 andthe state of coexistence of all populations. It dependson the initial state of the chemostat which of the twostates will be eventually reached. Multistability wehave also in regions 6, 8, 10, 12 since both theN1Pstate and theN1N2P state are stable. In regions 7, 9,11 only theN1P state is stable, while in regions 13,14 and 15 only the state of coexistence of all popu-lations is stable. Finally, in regions 16, 17, 18 onlyunstable steady states are observed. In this diagramthere are no regions of stable steady state ofN2 sur-vival only.
350 D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357
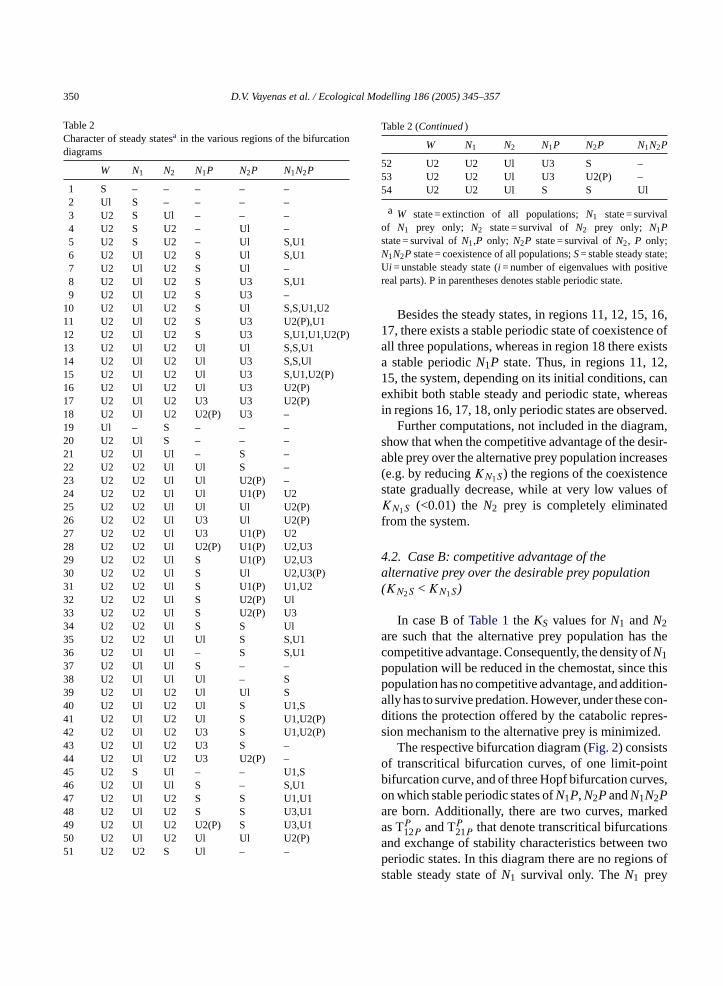
Table 2Character of steady statesa in the various regions of the bifurcationdiagrams
W N1 N2 N1P N2P N1N2P
1 S – – – – –2 Ul S – – – –3 U2 S Ul – – –4 U2 S U2 – Ul –5 U2 S U2 – Ul S,U16 U2 Ul U2 S Ul S,U17 U2 Ul U2 S Ul –8 U2 Ul U2 S U3 S,U19 U2 Ul U2 S U3 –
10 U2 Ul U2 S Ul S,S,U1,U211 U2 Ul U2 S U3 U2(P),U112 U2 Ul U2 S U3 S,U1,U1,U2(P)13 U2 Ul U2 Ul Ul S,S,U114 U2 Ul U2 Ul U3 S,S,Ul15 U2 Ul U2 Ul U3 S,U1,U2(P)16 U2 Ul U2 Ul U3 U2(P)17 U2 Ul U2 U3 U3 U2(P)18 U2 Ul U2 U2(P) U3 –19 Ul – S – – –20 U2 Ul S – – –21 U2 Ul Ul – S –22 U2 U2 Ul Ul S –23 U2 U2 Ul Ul U2(P) –24 U2 U2 Ul Ul U1(P) U225 U2 U2 Ul Ul Ul U2(P)26 U2 U2 Ul U3 Ul U2(P)27 U2 U2 Ul U3 U1(P) U228 U2 U2 Ul U2(P) U1(P) U2,U329 U2 U2 Ul S U1(P) U2,U330 U2 U2 Ul S Ul U2,U3(P)31 U2 U2 Ul S U1(P) U1,U232 U2 U2 Ul S U2(P) Ul33 U2 U2 Ul S U2(P) U334 U2 U2 Ul S S Ul35 U2 U2 Ul Ul S S,U136 U2 Ul Ul – S S,U137 U2 Ul Ul S – –38 U2 Ul Ul Ul – S39 U2 Ul U2 Ul Ul S40 U2 Ul U2 Ul S U1,S41 U2 Ul U2 Ul S U1,U2(P)42 U2 Ul U2 U3 S U1,U2(P)43 U2 Ul U2 U3 S –44 U2 Ul U2 U3 U2(P) –45 U2 S Ul – – U1,S46 U2 Ul Ul S – S,U147 U2 Ul U2 S S U1,U148 U2 Ul U2 S S U3,U149 U2 Ul U2 U2(P) S U3,U150 U2 Ul U2 Ul Ul U2(P)51 U2 U2 S Ul – –
Table 2 (Continued)
W N1 N2 N1P N2P N1N2P
52 U2 U2 Ul U3 S –53 U2 U2 Ul U3 U2(P) –54 U2 U2 Ul S S Ul
a W state = extinction of all populations;N1 state = survivalof N1 prey only; N2 state = survival ofN2 prey only; N1Pstate = survival ofN1,P only; N2P state = survival ofN2, P only;N1N2P state = coexistence of all populations;S= stable steady state;Ui = unstable steady state (i = number of eigenvalues with positivereal parts). P in parentheses denotes stable periodic state.
Besides the steady states, in regions 11, 12, 15, 16,17, there exists a stable periodic state of coexistence ofall three populations, whereas in region 18 there existsa stable periodicN1P state. Thus, in regions 11, 12,15, the system, depending on its initial conditions, canexhibit both stable steady and periodic state, whereasin regions 16, 17, 18, only periodic states are observed.
Further computations, not included in the diagram,show that when the competitive advantage of the desir-able prey over the alternative prey population increases(e.g. by reducingKN1S) the regions of the coexistencestate gradually decrease, while at very low values ofKN1S (<0.01) theN2 prey is completely eliminatedfrom the system.
4.2. Case B: competitive advantage of thealternative prey over the desirable prey population(KN2S < KN1S)
In case B ofTable 1theKS values forN1 andN2are such that the alternative prey population has thecompetitive advantage. Consequently, the density ofN1population will be reduced in the chemostat, since thispopulation has no competitive advantage, and addition-ally has to survive predation. However, under these con-ditions the protection offered by the catabolic repres-sion mechanism to the alternative prey is minimized.
The respective bifurcation diagram (Fig. 2) consistsof transcritical bifurcation curves, of one limit-pointb es,oa keda nsa twop s ofs
ifurcation curve, and of three Hopf bifurcation curvn which stable periodic states ofN1P,N2PandN1N2Pre born. Additionally, there are two curves, mars TP
12P and TP21P that denote transcritical bifurcatio
nd exchange of stability characteristics betweeneriodic states. In this diagram there are no regiontable steady state ofN1 survival only. TheN1 prey

D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357 351
Fig. 2. Bifurcation diagram for a two-prey–one-predator system regulated by a CRCL mode in the case that the predators grow faster on thealternative than on the desirable prey and the alternative prey has the competitive advantage over the desirable prey (case B ofTable 1). Labellingof curves as inFig. 1.The character of each steady state in the various regions is listed inTable 2.
can survive at stable steady states at low values ofB,in coexistence withP population (regions 29–34) or incoexistence with bothN2 andP (regions 35 and 36). AthighD values (regions 19 and 20) only the dominationof N2 is stable. The coexistence ofN2 andP is the onlystable steady state in regions 21 and 22, whereas inregion 34 two steady states are stable, theN1Pstate andtheN2Pstate. Multistability is also observed in regions35, 36 since both theN2Pstate and the coexistence stateare stable. There are no stable steady states in regions23–28.
With regard to periodic states, stable periodic coex-istence state is observed in regions 26, 27, 30, stableperiodicN1P state in region 28, and stable periodicN2P state in regions 23, 24, 27, 28, 29, 31, 32, 33.An interesting observation is that in region 28 we havemultistability of periodic states, since both the periodicN1P andN2P states are stable.
4.3. Predator grows faster on the desirable thanon the alternative prey (KPN1
< KPN2): cases C
and D
4.3.1. Case C: competitive advantage of thedesirable prey over the alternative prey population(KN1S < KN2S)
In case C ofTable 1the predators consume fasterthe desirable than the alternative prey population. How-ever, the desirable prey can survive since it has the com-petitive advantage for the common substrate over thealternative prey population.
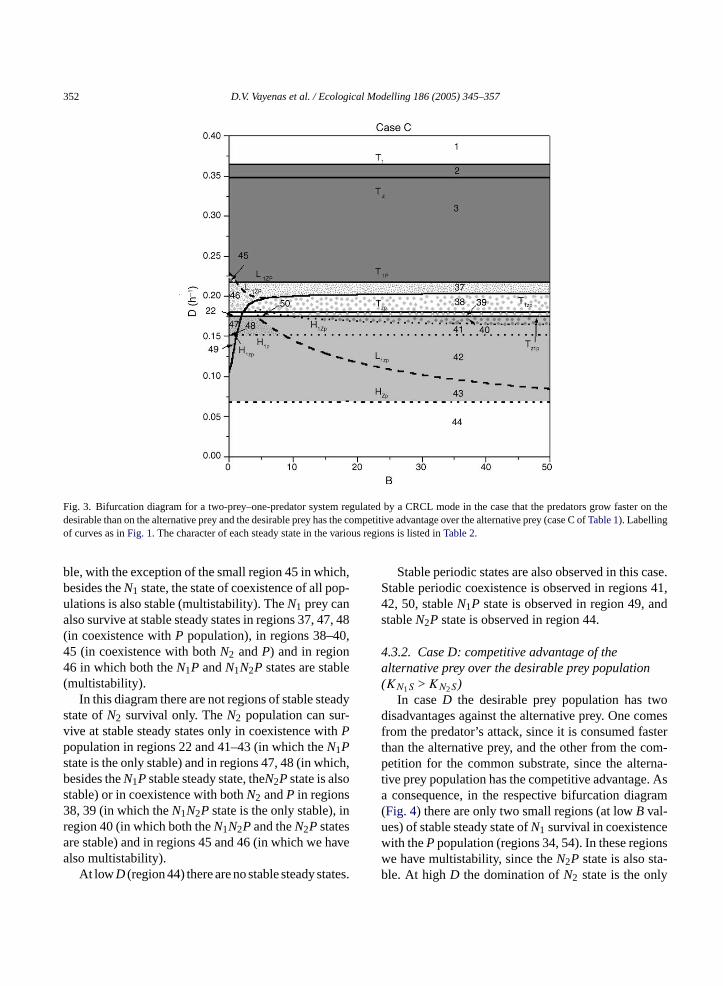
The bifurcation diagram of this case (Fig. 3) in-cludes several transcritical bifurcation curves, twolimit-point bifurcation curves, and three Hopf bifur-cation curves, on which stable periodic states ofN1P,N2PandN1N2Pare born. At high values ofD (regions2 and 3) the state of domination ofN1 is the only sta-
352 D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357
Fig. 3. Bifurcation diagram for a two-prey–one-predator system regulated by a CRCL mode in the case that the predators grow faster on thedesirable than on the alternative prey and the desirable prey has the competitive advantage over the alternative prey (case C ofTable 1). Labellingof curves as inFig. 1. The character of each steady state in the various regions is listed inTable 2.
ble, with the exception of the small region 45 in which,besides theN1 state, the state of coexistence of all pop-ulations is also stable (multistability). TheN1 prey canalso survive at stable steady states in regions 37, 47, 48(in coexistence withP population), in regions 38–40,45 (in coexistence with bothN2 andP) and in region46 in which both theN1P andN1N2P states are stable(multistability).
In this diagram there are not regions of stable steadystate ofN2 survival only. TheN2 population can sur-vive at stable steady states only in coexistence withPpopulation in regions 22 and 41–43 (in which theN1Pstate is the only stable) and in regions 47, 48 (in which,besides theN1Pstable steady state, theN2Pstate is alsostable) or in coexistence with bothN2 andP in regions38, 39 (in which theN1N2P state is the only stable), inregion 40 (in which both theN1N2Pand theN2P statesare stable) and in regions 45 and 46 (in which we havealso multistability).
At lowD (region 44) there are no stable steady states.
Stable periodic states are also observed in this case.Stable periodic coexistence is observed in regions 41,42, 50, stableN1P state is observed in region 49, andstableN2P state is observed in region 44.
4.3.2. Case D: competitive advantage of thealternative prey over the desirable prey population(KN1S > KN2S)
In caseD the desirable prey population has twodisadvantages against the alternative prey. One comesfrom the predator’s attack, since it is consumed fasterthan the alternative prey, and the other from the com-petition for the common substrate, since the alterna-tive prey population has the competitive advantage. Asa consequence, in the respective bifurcation diagram(Fig. 4) there are only two small regions (at lowB val-ues) of stable steady state ofN1 survival in coexistencewith thePpopulation (regions 34, 54). In these regionswe have multistability, since theN2P state is also sta-ble. At highD the domination ofN2 state is the only
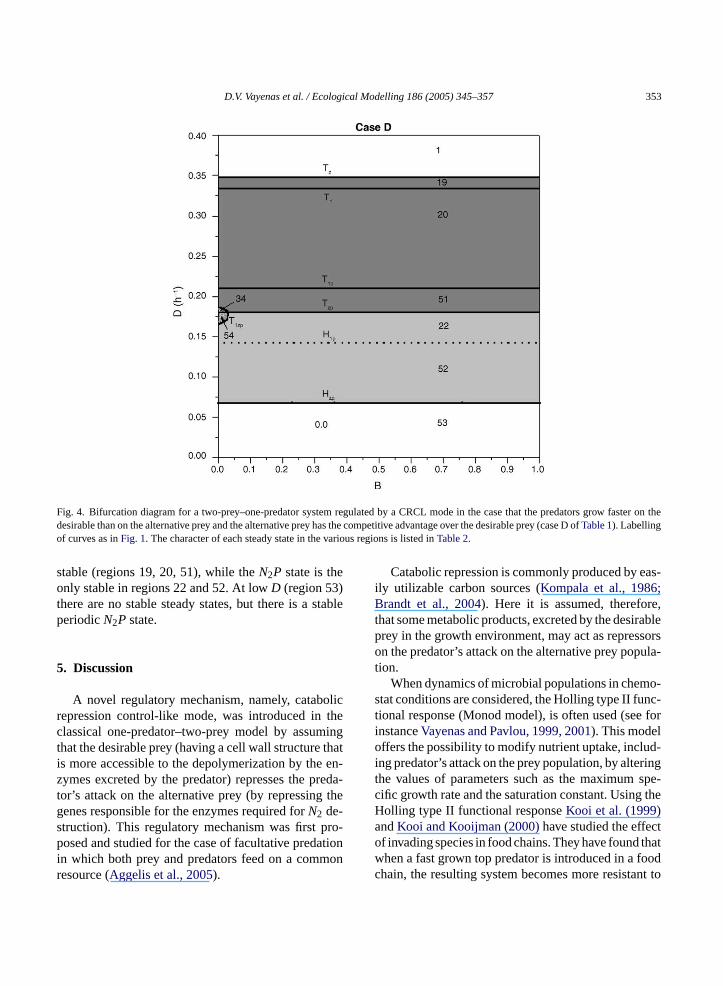
D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357 353
Fig. 4. Bifurcation diagram for a two-prey–one-predator system regulated by a CRCL mode in the case that the predators grow faster on thedesirable than on the alternative prey and the alternative prey has the competitive advantage over the desirable prey (case D ofTable 1). Labellingof curves as inFig. 1. The character of each steady state in the various regions is listed inTable 2.
stable (regions 19, 20, 51), while theN2P state is theonly stable in regions 22 and 52. At lowD (region 53)there are no stable steady states, but there is a stableperiodicN2P state.
5. Discussion
A novel regulatory mechanism, namely, catabolicrepression control-like mode, was introduced in theclassical one-predator–two-prey model by assumingthat the desirable prey (having a cell wall structure thatis more accessible to the depolymerization by the en-zymes excreted by the predator) represses the preda-tor’s attack on the alternative prey (by repressing thegenes responsible for the enzymes required forN2 de-struction). This regulatory mechanism was first pro-posed and studied for the case of facultative predationin which both prey and predators feed on a commonresource (Aggelis et al., 2005).
Catabolic repression is commonly produced by eas-ily utilizable carbon sources (Kompala et al., 1986;Brandt et al., 2004). Here it is assumed, therefore,that some metabolic products, excreted by the desirableprey in the growth environment, may act as repressorson the predator’s attack on the alternative prey popula-tion.
When dynamics of microbial populations in chemo-stat conditions are considered, the Holling type II func-tional response (Monod model), is often used (see forinstanceVayenas and Pavlou, 1999, 2001). This modeloffers the possibility to modify nutrient uptake, includ-ing predator’s attack on the prey population, by alteringthe values of parameters such as the maximum spe-cific growth rate and the saturation constant. Using theHolling type II functional responseKooi et al. (1999)andKooi and Kooijman (2000)have studied the effectof invading species in food chains. They have found thatwhen a fast grown top predator is introduced in a foodchain, the resulting system becomes more resistant to

354 D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357
further invasion (Kooi et al., 1999). They concluded,however, that, since the growth rate decreases with thetrophic level, this may enable short food chains to beresistant to invaders. On the other hand it is shown thatthe invasion of a competitor of the prey can stabilizean oscillatory nutrient–prey–predator bi-trophic foodchain (Kooi and Kooijman, 2000).
The preference of several predators for a specifictype of prey is well known in ecology. It may be ex-plained on grounds of size (even if size is consideredas a rather weak selection criterion), chemical com-position, or combination of both (Flynn et al., 1996).Although in several species predation may dependmainly on the probability of collision, some prey popu-lations appear to be selectively predated. For instance,the preyDunaliella primolectais selectively predatedby Oxyrrhis marina, which may be attracted towardsamines leaking fromDunaliella primolecta(Flynn andFielder, 1989), thus movement of the predator maynot be completely random (Verity, 1991). Tschirhart(2004)proposed that predator switching behavior de-pends on the relative energy expenditures for the preyspecies, and the predator may specialize in one prey orswitch gradually between prey.
According to the optimal foraging theory (Charnov,1976) the desirable prey is always included in thepredator’s diet, while the alternative prey is includedonly if the density of the desirable prey falls belowa critical threshold. The optimal foraging theory issupported by experimental data (Stephens and Krebs,1 tivep
-p l for-a nce,w r( per-s f os-ce hatt zoa( -na ym notb zy-m ismf
The CRCL mode, introduced in a two-prey–one-predator system in order to model the balanced attack ofthe predator on the alternative prey, has various effectson the population dynamics, which are dependent onthe values of the kinetic parameters of the populations.In general, the effect of parameterB (which is related tooperators found under catabolic repression and deter-mines the extent of the predator’s attack on theN2 prey)on the population dynamics is greater when predatorsgrow faster on the alternative than on the desirable prey.Especially, when the system a priori favors the domi-nation of the desirable prey (meaning that the desirableprey has the competitive advantage over the alternativeprey), the increase of parameterB, could positively af-fect the stable state of coexistence of all populations.Under these conditions there are no regions of stableN2 orN2Pstates in theD versusBbifurcation diagram,suggesting that the survival of the alternative prey de-pends on the presence of the desirable prey. With theincrease of parameterB the region of stable steady (orperiodic) state of coexistence extends over low valuesof D, while at high values ofD the only possible sta-ble state is theN1 state, since this population has thecompetitive advantage over the alternative prey.
When the alternative prey has the competitive ad-vantage over the desirable prey, the density of theN1population will be reduced in the chemostat and, conse-quently, the positive effect of the increase of parameterB on the survival of the alternative prey will decreaseor completely disappear. It is worth noting that, undert pro-m thec sir-a )a h thei eso ob-s lter-n duetc thee thec
thed f thep Es-p itivea f the
986), although the predator’s attack on the alternarey is more gradual than predicted by the theory.
Krivan and Sikder (1999), considering an oneredator–two-prey system, suggested that optimaging behavior of predators may promote coexistehile Krivan (1997a, 1998)and Krivan and Eisne
2003)showed that the optimal behavior leads toistence of prey–predator systems and reduction oillations in population densities. However,Malloryt al. (1983)have experimentally demonstrated t
he strategy of alternative prey is used by protoi.e., Tetrahymena thermothilus) to completely elimiate bacterial species such asSalmonella typhimuriumndKlebsiella pneumoniaein raw sewage. This studay suggest that optimal foraging theory coulde applied to microbial systems in which the enatic activity of the predators is a major mechan
or killing prey.
hese conditions, the catabolic repression controlotes the elimination of the desirable prey from
hemostat, since the regions of survival of the deble prey (in coexistence withP or N2P populationst stable steady states are drastically reduced wit
ncrease of parameterB, while stable periodic statf N1P or coexistence of all three populations areerved only when predators grow faster on the aative than on the desirable prey. This is probably
o the positive effect of the high values ofB on theompetitively favored alternative prey, resulting inxtinction of the desirable prey population fromhemostat.
In the case where the predator grows faster onesirable than on the alternative prey, the effect oarameterBon the population dynamics is smaller.ecially, when the desirable prey has the competdvantage over the alternative prey, the survival o

D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357 355
alternative prey at stable steady or periodic states incoexistence withP or N1P is possible at all values ofB. It is noted that, under these conditions, the presenceof the desirable prey is not indispensable for the sur-vival of the alternative prey. On the contrary, when thealternative prey has the competitive advantage over thedesirable prey population, the coexistence state is notpossible. The alternative prey dominates overN1 andPin a wide range ofD values, irrespective of theBvalues,whereas the desirable prey could survive only at lowB values (in a small range ofD values) in coexistencewith theP population.
Besides the ecological importance of the predatorswitching in the conservation and stability of naturalecosystems, some conclusions of agricultural or indus-trial interest could be drawn from the present work.The introduction and maintenance of particular speciesof microorganisms into the cultivated soil or bioreac-tors in particular is of great interest. For example, theinoculation of cultivated soils withAzospirillumspp.,or other free-living nitrogen-fixing bacteria, is consid-ered as an alternative method for supplying nitrogen,and probably other nutrients, and offering protectionagainst pathogens to the cultivated plants (Kefalogianniand Aggelis, 2002; Tsagou et al., 2003). Furthermore,the introduction of some bacteria, e.g.Pseudomonasspp., offers protection against many soil born plantpathogens (Thomashow and Weller, 1990), whereassome species are vastly used in the waste treatment orin the bioremediation of soils (Wiggins and Alexander,1 hon,1 iesa odelp pop-u cifics usedb ey),c
6
pa-r arry-i eda-t thea r thefi e of
prey (alternative prey) is additionally regulated by theabundance of an easily digested prey (desirable prey).Specifically, we have assumed that the desirable preycould act as a repressor on key-genes, which are respon-sible for the synthesis of essential (for the predator’s at-tack on the alternative prey population) enzyme(s). Theregulation of these genes, and therefore of the behav-ior of the predator population occurs via a catabolic re-pression mechanism. Furthermore, the proposed modelassumes that not only the abundance but also the typeof the desirable prey plays an important role in the reg-ulation of the attack of the predator on the alternativeprey population: the presence of a desirable prey shouldincrease the probability for alternative prey survival orcoexistence only when it can operate as an effective re-pressor in enzyme synthesis. The ability of the predatorto regulate its attack on the alternative prey in the pres-ence of a desirable prey is quantitatively expressed bythe parameterB, which is a parameter of biologicalsignificance. SimilarlyHraber and Milne (1997)haveproposed that, in adaptive systems, individuals processinformation and then alter their behavior according tosome objective.
The proposed assumption predicts, therefore, allpossible effects of the facultative predation strategyon the survival of the prey population. Namely, it canpredict the positive effects of intraguild predation onprey survival and coexistence (observed in experimen-tal food webs described, for example, byMorin andLawler, 1995; Fagan, 1997; Holyoak and Sachdev,1 onp
ofu o bea at thei da-tl er-r 01t sir-a tablep1 di ientu pop-ue presst
988; Gurijala and Alexander, 1990; Tso and Tag999; Rønn et al., 2001). However, these useful specre sometimes prey for the soil predators. The mroposed in this paper could be used for predictinglation dynamics in several microbial systems. Speystems (e.g., in which alternative prey strategy isy protozoa to completely eliminate desirable prould be explained by the present model.
. Conclusions
Switching behavior is regulated by ecologicalameters such as the specific growth rate, the cng capacity of the ecosystem, the saturation of prors, the competition among populations, etc. Inpproach presented in this paper we propose forst time that the predator’s attack on a specific typ
998; Morin, 1999), as well as the negative effectsrey survival (described byMallory et al., 1983).
It is important to know the chances of survivalseful microorganisms in a natural system, and tble to enhance these chances. In the case th
ntroduced microorganism is a prey for soil preors (e.g., nematodes, microarthropods, protozoa,Bde-ovibrio spp., etc) (Morin, 1999; Jurgens and Montsat Sala, 2000; Mamilov et al., 2001; Rønn et al., 20),he co-inoculation together with a fast growing deble prey may promote the establishment of the suiopulation by repressing the predator’s attack (Verity,991; Flynn et al., 1996). With the model propose
n this paper it is possible to modify, besides nutrptake, the predators attack on the alternative preylation by altering the value of the parameterB, whichxpresses the capability of the desirable prey to rehe predator’s attack on the alternative prey.

356 D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357
Acknowledgments
Financial support was provided by the bilateralproject between Greece and Albania: microbial con-version of residues from olive-oil industry and otheragricultural industries, into biotechnological products.
References
Abrams, P.A., 1980. Are competition coefficients constant? Inductiveversus deductive approaches. Am. Nat. 116, 730–735.
Aggelis, G., Vayenas, D.V., Tsagou, V., Pavlou, S., 2005.Prey–predator dynamics with predator switching regulated bya catabolic repression control mode. Ecol. Mod. 183, 453–464.
Alexander, M., 1981. Why microbial predators and parasites do noteliminate their prey and hosts. Ann. Rev. Microbiol. 35, 113–133.
Atlas, R.M., 1997. Principles of Microbiology, 2nd ed. McGraw-Hill,Boston, USA, pp. 283–285.
Bailey, J.E., Ollis, D.F., 1986. Biochemical Engineering Fundamen-tals, 2nd ed. McGraw-Hill, Singapore, pp. 307–372, 854–902.
Brandt, B.W., Kelpin, F.D.L., van Leeuwen, I.M.M., Kooijman,S.A.L.M., 2004. Modelling microbial adaptation to changingavailability of substrates. Water Res. 38, 1003–1013.
Charnov, E.L., 1976. Optimal foraging, the marginal value theorem.Theor. Popul. Biol. 9, 129–136.
Doedel, E.J., Champneys, A.R., Fairgrieve, T.F., Kuznetsov, Y.A.,Sandstede, B., Wang, X., 1997. AUTO 97: Continuation and Bi-furcation Software for Ordinary Differential Equations: User’sManual, Center for Research on Parallel Computing, CaliforniaInstitute of Technology, Pasadena, USA.
Egli, T., 1995. The ecological and physiological significance of thegrowth of heterotrophic microorganisms with mixtures of sub-strates. Adv. Microb. Ecol. 14, 305–386.
F com-
F tionand96,
F ex-oro-teo ascol.
G hy-nv-
H ple
H odel
I sionnol.
Jurgens, K., Montserrat Sala, M., 2000. Predation-mediated shifts insize distribution of microbial biomass and activity during detritusdecomposition. Oikos 91, 29–40.
Kefalogianni, I., Aggelis, G., 2002. Modelling growth and biochem-ical activities ofAzospirillumspp. Appl. Microbiol. Biotechnol.58, 352–357.
Kooi, B.W., Boer, M.P., Kooijman, S.A.L.M., 1999. Resistance ofa food chain to invasion by a top predator. Math. Biosci. 157,217–236.
Kooi, B.W., Kooijman, S.A.L.M., 2000. Invading species can stabi-lize simple trophic systems. Ecol. Model. 133, 57–72.
Kompala, D.S., Ramkrishna, D., Jansen, N.B., Tsao, G.T., 1986.Investigation of bacterial growth on mixed substrates: experi-mental evaluation of cybernetic models. Biotechnol. Bioeng. 28,1044–1055.
Krivan, V., 1996. Optimal foraging and predator–prey dynamics.Theor. Popul. Biol. 49, 265–290.
Krivan, V., 1997a. Dynamic ideal free distribution: effects of op-timal patch choice on predator–prey dynamics. Am. Nat. 149,164–178.
Krivan, V., 1997b. Dynamical consequences of optimal host-feedingon host–parasitoid population dynamics. Bull. Math. Biol. 59,809–831.
Krivan, V., 1998. Effects of optimal antipredator behavior of prey onpredator–prey dynamics: the role of refuges. Theor. Popul. Biol.53, 131–142.
Krivan, V., Sikder, A., 1999. Optimal foraging and predator–preydynamics II. Theor. Popul. Biol. 55, 111–126.
Krivan, V., Eisner, J., 2003. Optimal foraging and predator–prey dy-namics III. Theor. Popul. Biol. 63, 269–279.
Mallory, L.M., Yuk, C.-S., Liang, L.-N., Alexander, M., 1983. Al-ternative prey: a mechanism for elimination of bacterial speciesby protozoa. Appl. Environ. Microbiol. 46, 1073–1079.
Mamilov, A.S., Byzov, B.A., Zvyagintsev, D.G., Dilly, O.M., 2001.Predation on fungal and bacterial biomass in a soddy-podzolic
ppl.
M acteri-
M pula-col.
M ula-52–
P ble? Aiosci.
R addi-
3,
Siol.
S Uni-
agan, W.F., 1997. Omnivory as a stabilizing feature of naturalmunities. Am. Nat. 150, 554–567.
lynn, K.J., Davidson, K., Cunningham, A., 1996. Prey selecand rejection by a microflagellate; implications for the studyoperation of microbial food webs. J. Exp. Mar. Biol. Ecol. 1357–372.
lynn, K.J., Fielder, J., 1989. Changes in intracellular andtracellular amino acids during the predation of the chlphyteDunaliella primolectaby the heterotrophic dinoflagellaOxyrrhis marinaand the use of the glutamine/glutamate ratian indicator of nutrient status in mixed population. Mar. EProg. Ser. 53, 117–127.
urijala, K.R., Alexander, M., 1990. Effect of growth rate anddrophobicity on bacteria surviving protozoa grazing. Appl. Eiron. Microbiol. 56, 1631–1635.
olyoak, M., Sachdev, S., 1998. Omnivory and the stability of simfood webs. Oecologia 117, 413–419.
raber, R.T., Milne, B.T., 1997. Community assembly in a mecosystem. Ecol. Model. 103, 267–285.
manaka, T., Aiba, S., 1977. A kinetic model of catabolite represin the dual control mechanism in microorganisms. BiotechBioeng. 19, 757–764.
soil amended with starch, wheat straw and alfalfa meal. ASoil Ecol. 16, 131–139.
onod, J., 1942. Recherches sur la croissance des cultures bennes. Actualites Scientifiques et Industrielles 911, 1–215.
orin, P.J., Lawler, S.M., 1995. Food web architecture and potion dynamics: theory and empirical evidence. Ann. Rev. ESyst. 26, 505–529.
orin, P., 1999. Productivity, intraguild predation, and poption dynamics in experimental food webs. Ecology 80, 7760.
elletier, J.D., 2000. Are large complex ecosystems more unstatheoretical reassessment with predator switching. Math. B163, 91–96.
ønn, R., Grunert, J., Ekelund, F., 2001. Protozoan response totion of the bacteriaMycobacterium chlorophenolicumandPseu-domonas chlororaphisto soil microcosms. Biol. Fertil. Soils 3126–131.
aint-Cyr, J.-F., Cloutier, C., 1996. Prey preference by stinkbugPer-illus bioculatus, a predator of the colorado potato beetle. BContr. 7, 251–258.
tephens, D.W., Krebs, J.R., 1986. Foraging Theory. Princetonversity Press, Princeton, NJ.

D.V. Vayenas et al. / Ecological Modelling 186 (2005) 345–357 357
Thomashow, L.S., Weller, D.M., 1990. Application of fluorescentpseudomonads to control root diseases of wheat and some mech-anisms of disease suppression. In: Hornby, D. (Ed.), BiologicalControl of Soil-borne Plant Pathogens. CAB International, Oxon,pp. 109–122.
Tsagou, V., Kefalogianni, I., Sini, K., Aggelis, G., 2003. Metabolicactivities in Azospirilum lipoferumgrown in the presence ofNH+
4 . Appl. Microbiol. Biotechnol. 62, 574–578.Tschirhart, J., 2004. A new adaptive system approach to
predator–prey modeling. Ecol. Model. 176, 255–276.Tso, S.F., Taghon, G.L., 1999. Factors affecting predation byCy-
clidiumsp. andEuplotessp. on PAH-degrading bacteria. Microb.Ecol. 37, 3–12.
Van Baalen, M., Krivan, V., Van Rijn, P.C.J., Sabelis, M., 2001.Alternative food, switching predators, and the persistence ofpredator–prey systems. Am. Nat. 157, 512–524.
Verity, P.G., 1991. Feeding on planktonic protozoan – evidence fornonrandom acquisition. J. Protozool. 38, 69–76.
Vayenas, D.V., Pavlou, S., 1999. Chaotic dynamic of a food web ina chemostat. Math. Biosci. 162, 69–84.
Vayenas, D.V., Pavlou, S., 2001. Chaotic dynamic of a micro-bial system of coupled food chains. Ecol. Model. 136, 285–295.
Wiggins, B.A., Alexander, M., 1988. Role of protozoa in microbialacclimation for mineralization of organic chemical in sewage.Can. J. Microbiol. 34, 661–666.




















