
Proteolytic Cleavage of Protein Tyrosine Phosphatase Regulates Glioblastoma Cell Migration
Upload
independentCategory
view
6download
0
and Kathy K. GriendlingAlejandra San Martín, Moo Yeol Lee, Holly C. Williams, Kensaku Mizuno, Bernard Lassègue
Muscle CellsInduced Migration of Human Aortic Smooth−Controls Platelet-Derived Growth Factor
Dual Regulation of Cofilin Activity by LIM Kinase and Slingshot-1L Phosphatase
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2007 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.107.1589232008;102:432-438; originally published online December 20, 2007;Circ Res.
http://circres.ahajournals.org/content/102/4/432World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2007/12/20/CIRCRESAHA.107.158923.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from

Dual Regulation of Cofilin Activity by LIM Kinaseand Slingshot-1L Phosphatase Controls Platelet-Derived
Growth Factor–Induced Migration of HumanAortic Smooth Muscle Cells
Alejandra San Martın, Moo Yeol Lee, Holly C. Williams, Kensaku Mizuno,Bernard Lassegue, Kathy K. Griendling
Abstract—Platelet-derived growth factor (PDGF) plays a central role in vascular healing, atherosclerosis, and restenosis,partly by stimulating vascular smooth muscle cell (VSMC) migration. Migration requires rapid turnover of actinfilaments, which is partially controlled by cofilin. Although cofilin is negatively regulated by Ser3 phosphorylation, theupstream signaling pathways have not been defined, nor has its role in VSMC migration been studied. We hypothesizedthat PDGF-induced migration of VSMCs involves cofilin activation and that this is regulated by the serine kinase LIMkinase (LIMK) and the novel phosphatase Slingshot (SSH)1L. In human VSMCs, stimulation with PDGF increasedG-actin incorporation into the actin cytoskeleton. PDGF transiently activated the cofilin kinase, LIMK, with a peak at5 minutes. However, cofilin was dephosphorylated between 5 and 45 minutes, with a maximum of 43�5%dephosphorylation at 30 minutes, suggesting that PDGF also activates a cofilin phosphatase. We found that VSMCsexpress SSH1L, which is induced and activated (564�73 versus 1021�141 picomoles of PO4; P�0.015) by PDGF. Ofimportance, small interfering RNA directed against SSH1L blocked cofilin dephosphorylation and decreased migration(528�33 versus 318�25 cells/field; P�0.01). Taken together, our results suggest that PDGF participates in actindynamics by dual regulation of cofilin activity via LIMK and SSH1L. (Circ Res. 2008;102:432-438.)
Key Words: vascular smooth muscle � slingshot phosphatase � platelet-derived growth factor� migration � LIMK
Directed cell migration is an integrated, dynamic, andcyclical process that plays a key role in mounting
immune responses and repairing injured tissues, as well as inguiding the morphogenesis of the embryo during develop-ment. In addition, migration contributes to pathologies in-cluding chronic inflammatory diseases, postangioplasty reste-nosis, atherosclerosis, and cancer. Vascular smooth musclecell (VSMC) migration is influenced by many factors but invivo is largely a consequence of stimulation of the platelet-derived growth factor (PDGF)-� receptors by PDGF.1 How-ever, the signaling pathways by which PDGF controls VSMCmigration are still not completely understood.
Regardless of the cell type, a fast and efficient reorgani-zation of the actin cytoskeleton is absolutely essential formigration. Different proteins, including actin-related proteins2 and 3 (Arp2/3), Wiscott–Aldrich syndrome protein(WASP), profilin, mammalian diaphanous proteins, and co-filin, have been shown to regulate the assembly and disas-sembly of the actin cytoskeleton in response to multiple
extracellular stimuli.2 Of these, cofilin, a protein belonging tothe actin depolarizing factor (ADF) family of proteins, iscapable of stimulating disassembly and severance of actinfilaments at or near the pointed ends, thereby continuouslysupplying actin monomers for polymerization and rapidturnover of actin filaments.3 Thus, the activation of cofilinplays an essential role in the protrusion of lamellipodia at theleading edge of migrating cells. The activity of cofilin isnegatively regulated by the LIM kinase (LIMK) family ofserine/threonine kinases through phosphorylation at Ser3 ofcofilin.4–6 Suppression of cofilin activity by LIMK overex-pression abolishes lamellipodium formation and polarizedcell migration, which implicates cofilin in cell polarity andmigration.7,8
Previous studies have revealed that phospho-cofilin isdephosphorylated, and therefore activated, in response tovarious extracellular stimuli.9–11 Several proteins have beenshown to be cofilin phosphatases, including chronophin andthe Slingshot (SSH) family of protein phosphatases.12,13
Original received June 28, 2007; revision received November 20, 2007; accepted December 7, 2007.From the Division of Cardiology (A.S.M., M.Y.L., H.C.W., B.L., K.K.G.), Emory University, Atlanta, Ga; and Department of Biomolecular Sciences
(K.M.), Tohoku University, Aramaki-aza-aoba, Aoba-ku, Sendai, Japan.This manuscript was sent to Mark Taubman, Consulting Editor, for review by expert referees, editorial decision, and final disposition.Correspondence to Kathy K. Griendling, Division of Cardiology, Emory University, 1639 Pierce Dr, 319 WMB, Atlanta, GA 30322. E-mail
[email protected]© 2008 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.107.158923
432 by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from

Chronophin is highly expressed in brain, heart, and liver butis nearly absent in smooth muscle–containing tissues,13
whereas SSH family members are widely expressed in mousetissues.12 Of the SSH family of proteins, SSH1L, the longform of SSH1 that has an actin-binding site near the Cterminus, has been shown to be important in chemotaxis.14
SSH1L colocalizes with F-actin and its phosphatase activitycorrelates with the level of cofilin dephosphorylation.10
Based on these observations, we hypothesized that PDGF-induced changes in VSMC actin polymerization and migra-tion involve cofilin activation, and that this activation is theresult of regulation of LIMK and SSH1L phosphatase. Wefound that a net dephosphorylation of cofilin results fromsimultaneous activation of both enzymes, thus providing amechanism for fine control of VSMC migration.
Materials and MethodsRecombinant human PDGF-BB was purchased from R&D SystemsInc (Minneapolis, Minn) and was used in all the experiments.
Cell CultureHuman aortic smooth muscle cells were purchased from Clonetics(San Diego, Calif). The cells were cultured in Clonetics SmGM-2medium containing human epidermal growth factor 0.5 ng/mL,insulin 5 �g/mL, hFGF 2 ng/mL, 5% FBS, gentamicin 50 �g/mL,and amphotericin B 50 ng/mL. Cell cultures were incubated at 37°Cin a humidified atmosphere of 5% CO2. VSMCs between passages 6and 12 were used for the experiments. Cultures at 70% to 80%confluence were made quiescent by incubation in serum free mediafor 24 to 48 hours before the experiments.
Western BlottingAfter treatment, cells were lysed in 1% Triton–containing lysisbuffer and analyzed by Western blotting as described previously.15
Membranes were incubated with primary antibodies againstphospho-cofilin(Ser3), cofilin, phospho-LIMK1(Thr508)/LIMK2(Thr505), LIMK1 (Cell Signaling), or �-tubulin (Sigma).Rabbit polyclonal antibody against SSH1L has been describedpreviously.14 After incubation with horseradish peroxidase-conjugated secondary antibody, proteins were detected by ECLchemiluminescence. The films were analyzed by densitometry withImageJ software (NIH).
Small Interfering RNA and PlasmidTransfection ExperimentsCells were transfected by electroporation using a Nucleofector(Amaxa Biosystems) set to the U25 program with 2 �g of plasmidenhanced green fluorescent protein (pEGFP)-KS (encoding a kinase-dead mutant, which acts as a dominant negative16; a kind gift of DrCondeelis, Albert Einstein College of Medicine, NY) or pEGFP(Clontech) plasmids per 1�106 cells, and/or with 1.5 �g of annealedsmall interfering (si)RNA duplexes for SSH1L or nonsilencingcontrol sequence no. 1 from Qiagen per 1.6�106 cells. siRNAduplexes were synthesized using a previously published sequence17
(sense: 5�-UCG UCA CCC AAG AAA GAU AUU-3�; antisense:5�-UAU CUU UCU UGG GUG ACG AUU-3�). The transfectionwas made three days before a 24-hour serum deprivation before theexperiments.
Real-Time Quantitative RT-PCRTotal RNA was purified from VSMCs using the RNeasy kit(Qiagen), as recommended by the manufacturer. RNA was reverse-transcribed with Superscript II enzyme (Invitrogen) using randomprimers. cDNA samples were additionally purified using microbio-spin 30 columns (Bio-Rad) and amplified in the LightCycler (Roche)real-time thermocycler using recombinant Platinum Taq DNA poly-
merase (Invitrogen) with SYBR green dye. Amplification conditionswere for SSH1L, 150 nmol/L primers (upstream primer, 5�-GATCAAAACCTGCTCAACTCGGAGAACCT-3�; downstreamprimer, 5�-CTGGAGCCTGCTGGTGGTAGGAAC-3�), 4 mmol/LMgCl2, 5% DMSO, annealing at 72°C; for 18S, 50 nmol/L AlternateII 18S rRNA primers (Ambion), 4 mmol/L MgCl2, and annealing at60°C. Standard curves were generated using 10-fold serial dilutionsof a plasmid containing cloned cDNA (either SSH1L or 18S) andmeasured in each run at the same time as unknown samples. Copynumbers were calculated by the software provided with the instru-ment from a linear regression of CT versus Log (copy number).Finally copy numbers of SSH1L were normalized to copy numbersof 18S measured in duplicate samples.
VSMC Migration AssayMigration was assayed using a modified Boyden chamber assay aspreviously described.15 This method has previously been shown tomeasure directed cell migration.18 VSMCs were grown to 85%confluence and then made quiescent in serum-free media for 24hours before migration. VSMCs (5�104 cells/well) were added tothe upper chamber of a Transwell dish on a 6.5-mm insert with acollagen-coated polycarbonate membrane containing 8-�m pores(Costar). VSMCs were then exposed to PDGF (10 ng/mL) in thelower chamber for 3 hours, after which nonmigrated cells wereremoved from the upper chamber using a cotton swab. The cellsremaining on the inserts were fluorescently stained with 4�,6-diamidino-2-phenylindole (DAPI) (1 �g/mL) and visualized using aZeiss Axioskop microscope. Migrated cells per membrane werequantified using ImageJ software.
G-Actin IncorporationIncorporation of G-actin into the cytoskeleton was measured in vivoin permeabilized cells using a previously published protocol19 withslight modifications. VSMCs were seeded in Laboratory-Tek Cham-ber Slides (Nunc) covered with collagen (5 ng/mL). After stimula-tion with PDGF (10 ng/mL) for 30 minutes, cells were incubatedwith permeabilization buffer (20 mmol/L HEPES [pH 7.5],138 mmol/L KCl, 4 mmol/L MgCl2, 3 mmol/L EGTA, 1% BSA)containing 40 �g/mL saponin, 0.45 �mol/L biotinylated nonmuscleG-actin (Cytoskeleton Inc), and 1 mmol/L ATP for 3 minutes. Afterfixation, incorporated G actin was visualized with an anti-biotinantibody coupled to Rhodamine Red-X (The Jackson Laboratory).Actin filaments were stained with phalloidin Alexa-488 (MolecularProbes).
Phosphatase AssayPhosphatase activity was measured in cell lysates or immunoprecipi-tates with a commercial kit (Promega) with modifications. VSMCswere lysed in 20 mmol/L imidazole–HCl (pH 7.2) containing1 mmol/L benzamidine, 1 mmol/L dithiothreitol, 0.05% NP40, andprotease inhibitors and were centrifuged at 100 000g for 1 hour at4°C. The supernatant was passed through a Sephadex G-25 columnand incubated with a phosphopeptide mimicking the N terminus ofcofilin [MAS(PO4)GVA] at a concentration of 100 �mol/L inreaction buffer (50 mmol/L imidazole, 0.02% �-mercaptoethanol,0.1 mg/mL BSA) for 15 minutes at 37°C. The reaction was stoppedwith a molybdate/malachite complex and the absorbance was mea-sured at 600 nm. For immunoprecipitation experiments, VSMCswere lysed in Tris buffer (50 mmol/L Tris [pH 7.5], 150 mmol/LNaCl, 1% Nonidet P-40, 5% glycerol, 1 mmol/L dithiothreitol, 10�g/mL leupeptin, 1 mmol/L PMSF). Lysates (300 �g per reaction)were immunoprecipitated with anti-SSH1L antibody for 2 hours at4°C. Then 20 �L of Protein A/G-Agarose Plus (Santa Cruz Biotech-nology) were added and incubated at 4°C for 1 hour. The pellet wascollected by centrifugation at 3000 rpm for 3 minutes at 4°C andwashed 4 times with lysis buffer. Immunoprecipitates were subjectedto the phosphatase assay described above.
San Martın et al LIMK and SSH1L in PDGF-Induced VSMC Migration 433
by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from
StatisticsResults are expressed as means�SEM. Differences among groupswere analyzed using 1-way ANOVA, with post hoc contrastsadjusted according to the Duncan correction using SPSS 14.0 forWindows. A value of P�0.05 was considered to be statisticallysignificant.
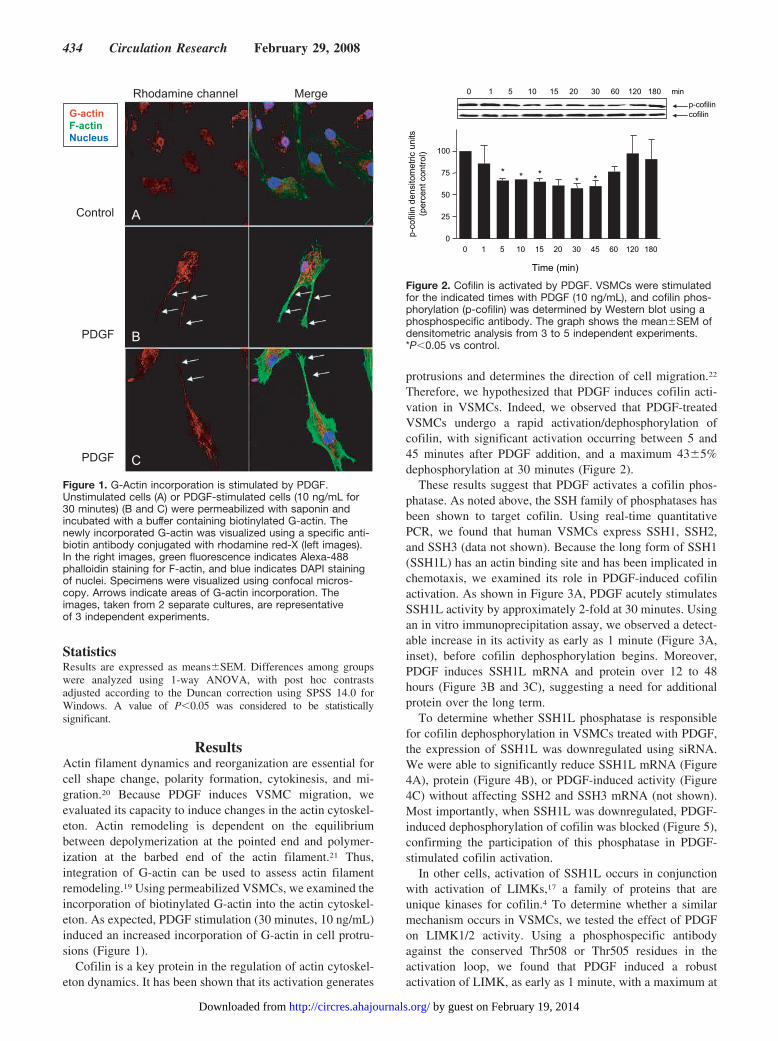
ResultsActin filament dynamics and reorganization are essential forcell shape change, polarity formation, cytokinesis, and mi-gration.20 Because PDGF induces VSMC migration, weevaluated its capacity to induce changes in the actin cytoskel-eton. Actin remodeling is dependent on the equilibriumbetween depolymerization at the pointed end and polymer-ization at the barbed end of the actin filament.21 Thus,integration of G-actin can be used to assess actin filamentremodeling.19 Using permeabilized VSMCs, we examined theincorporation of biotinylated G-actin into the actin cytoskel-eton. As expected, PDGF stimulation (30 minutes, 10 ng/mL)induced an increased incorporation of G-actin in cell protru-sions (Figure 1).
Cofilin is a key protein in the regulation of actin cytoskel-eton dynamics. It has been shown that its activation generates
protrusions and determines the direction of cell migration.22
Therefore, we hypothesized that PDGF induces cofilin acti-vation in VSMCs. Indeed, we observed that PDGF-treatedVSMCs undergo a rapid activation/dephosphorylation ofcofilin, with significant activation occurring between 5 and45 minutes after PDGF addition, and a maximum 43�5%dephosphorylation at 30 minutes (Figure 2).
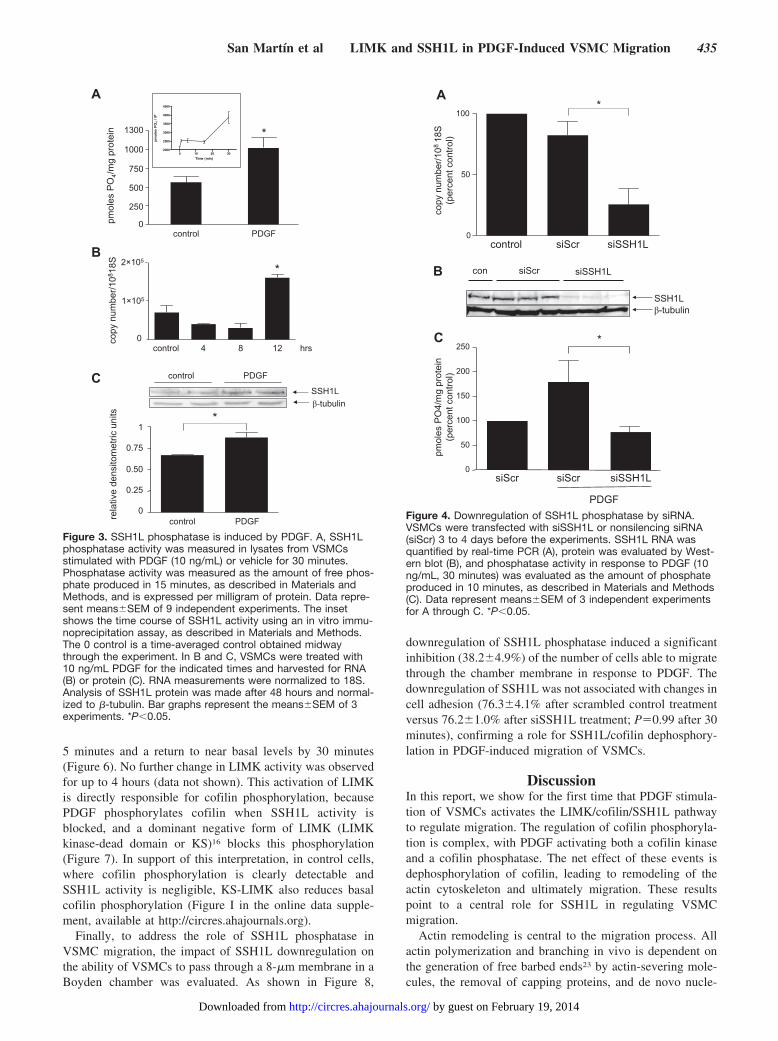
These results suggest that PDGF activates a cofilin phos-phatase. As noted above, the SSH family of phosphatases hasbeen shown to target cofilin. Using real-time quantitativePCR, we found that human VSMCs express SSH1, SSH2,and SSH3 (data not shown). Because the long form of SSH1(SSH1L) has an actin binding site and has been implicated inchemotaxis, we examined its role in PDGF-induced cofilinactivation. As shown in Figure 3A, PDGF acutely stimulatesSSH1L activity by approximately 2-fold at 30 minutes. Usingan in vitro immunoprecipitation assay, we observed a detect-able increase in its activity as early as 1 minute (Figure 3A,inset), before cofilin dephosphorylation begins. Moreover,PDGF induces SSH1L mRNA and protein over 12 to 48hours (Figure 3B and 3C), suggesting a need for additionalprotein over the long term.
To determine whether SSH1L phosphatase is responsiblefor cofilin dephosphorylation in VSMCs treated with PDGF,the expression of SSH1L was downregulated using siRNA.We were able to significantly reduce SSH1L mRNA (Figure4A), protein (Figure 4B), or PDGF-induced activity (Figure4C) without affecting SSH2 and SSH3 mRNA (not shown).Most importantly, when SSH1L was downregulated, PDGF-induced dephosphorylation of cofilin was blocked (Figure 5),confirming the participation of this phosphatase in PDGF-stimulated cofilin activation.
In other cells, activation of SSH1L occurs in conjunctionwith activation of LIMKs,17 a family of proteins that areunique kinases for cofilin.4 To determine whether a similarmechanism occurs in VSMCs, we tested the effect of PDGFon LIMK1/2 activity. Using a phosphospecific antibodyagainst the conserved Thr508 or Thr505 residues in theactivation loop, we found that PDGF induced a robustactivation of LIMK, as early as 1 minute, with a maximum at
Control
PDGF
PDGF
Rhodamine channel Merge
G-actinF-actinNucleus
A
C
B
Figure 1. G-Actin incorporation is stimulated by PDGF.Unstimulated cells (A) or PDGF-stimulated cells (10 ng/mL for30 minutes) (B and C) were permeabilized with saponin andincubated with a buffer containing biotinylated G-actin. Thenewly incorporated G-actin was visualized using a specific anti-biotin antibody conjugated with rhodamine red-X (left images).In the right images, green fluorescence indicates Alexa-488phalloidin staining for F-actin, and blue indicates DAPI stainingof nuclei. Specimens were visualized using confocal micros-copy. Arrows indicate areas of G-actin incorporation. Theimages, taken from 2 separate cultures, are representativeof 3 independent experiments.
0 1 5 10 15 20 30 60 120 180 min
* * ** *
p-co
filin
den
sito
met
ric u
nits
(per
cent
con
trol
)
Time (min)
0 1 5 10 15 20 30 45 60 120 1800
25
50
75
100
p-cofilincofilin
Figure 2. Cofilin is activated by PDGF. VSMCs were stimulatedfor the indicated times with PDGF (10 ng/mL), and cofilin phos-phorylation (p-cofilin) was determined by Western blot using aphosphospecific antibody. The graph shows the mean�SEM ofdensitometric analysis from 3 to 5 independent experiments.*P�0.05 vs control.
434 Circulation Research February 29, 2008
by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from
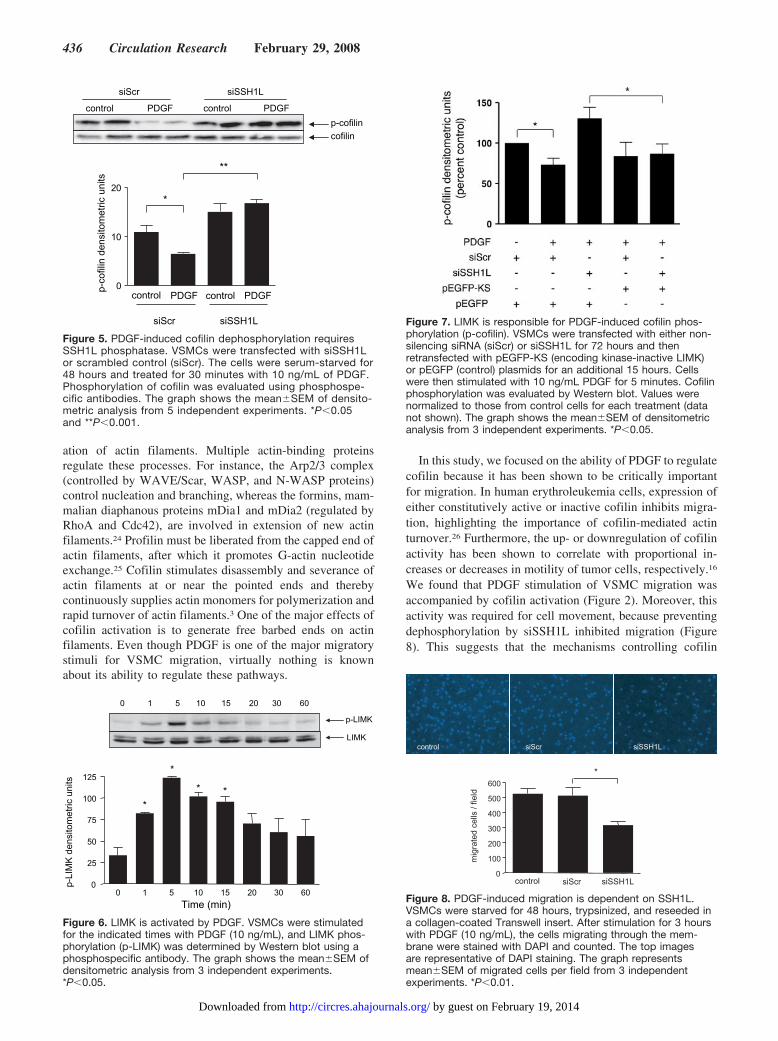
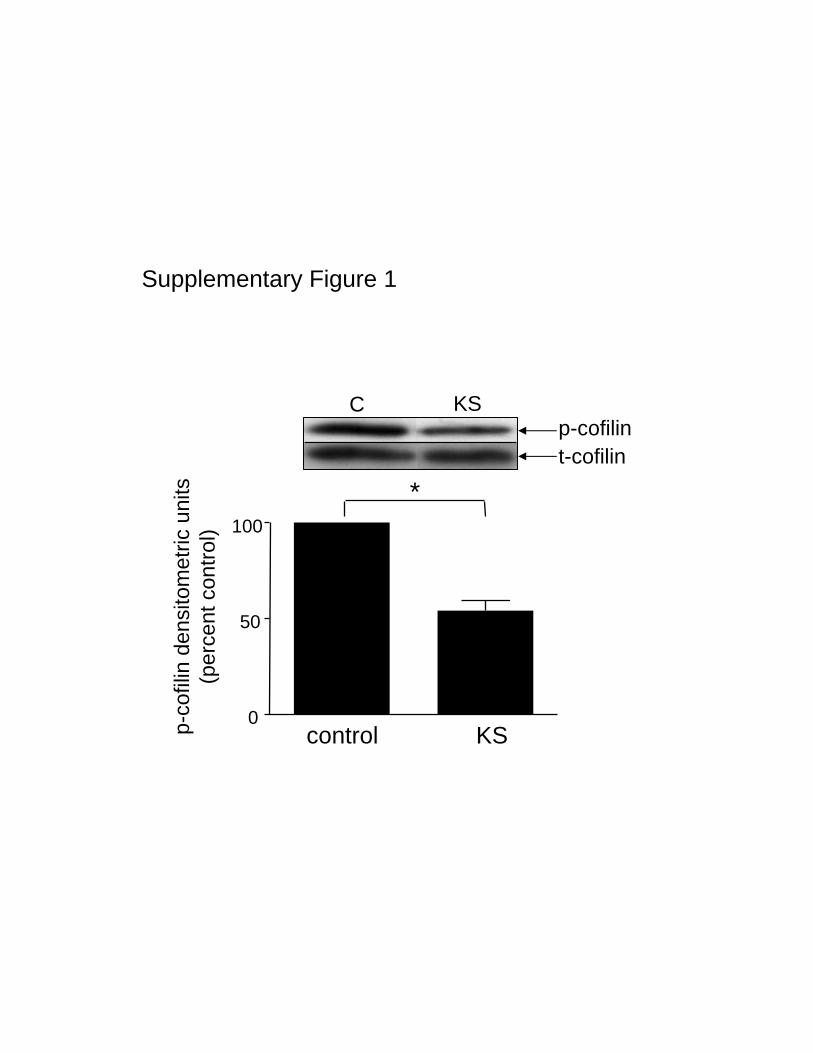
5 minutes and a return to near basal levels by 30 minutes(Figure 6). No further change in LIMK activity was observedfor up to 4 hours (data not shown). This activation of LIMKis directly responsible for cofilin phosphorylation, becausePDGF phosphorylates cofilin when SSH1L activity isblocked, and a dominant negative form of LIMK (LIMKkinase-dead domain or KS)16 blocks this phosphorylation(Figure 7). In support of this interpretation, in control cells,where cofilin phosphorylation is clearly detectable andSSH1L activity is negligible, KS-LIMK also reduces basalcofilin phosphorylation (Figure I in the online data supple-ment, available at http://circres.ahajournals.org).
Finally, to address the role of SSH1L phosphatase inVSMC migration, the impact of SSH1L downregulation onthe ability of VSMCs to pass through a 8-�m membrane in aBoyden chamber was evaluated. As shown in Figure 8,
downregulation of SSH1L phosphatase induced a significantinhibition (38.2�4.9%) of the number of cells able to migratethrough the chamber membrane in response to PDGF. Thedownregulation of SSH1L was not associated with changes incell adhesion (76.3�4.1% after scrambled control treatmentversus 76.2�1.0% after siSSH1L treatment; P�0.99 after 30minutes), confirming a role for SSH1L/cofilin dephosphory-lation in PDGF-induced migration of VSMCs.
DiscussionIn this report, we show for the first time that PDGF stimula-tion of VSMCs activates the LIMK/cofilin/SSH1L pathwayto regulate migration. The regulation of cofilin phosphoryla-tion is complex, with PDGF activating both a cofilin kinaseand a cofilin phosphatase. The net effect of these events isdephosphorylation of cofilin, leading to remodeling of theactin cytoskeleton and ultimately migration. These resultspoint to a central role for SSH1L in regulating VSMCmigration.
Actin remodeling is central to the migration process. Allactin polymerization and branching in vivo is dependent onthe generation of free barbed ends23 by actin-severing mole-cules, the removal of capping proteins, and de novo nucle-
A
B
C
Figure 3. SSH1L phosphatase is induced by PDGF. A, SSH1Lphosphatase activity was measured in lysates from VSMCsstimulated with PDGF (10 ng/mL) or vehicle for 30 minutes.Phosphatase activity was measured as the amount of free phos-phate produced in 15 minutes, as described in Materials andMethods, and is expressed per milligram of protein. Data repre-sent means�SEM of 9 independent experiments. The insetshows the time course of SSH1L activity using an in vitro immu-noprecipitation assay, as described in Materials and Methods.The 0 control is a time-averaged control obtained midwaythrough the experiment. In B and C, VSMCs were treated with10 ng/mL PDGF for the indicated times and harvested for RNA(B) or protein (C). RNA measurements were normalized to 18S.Analysis of SSH1L protein was made after 48 hours and normal-ized to �-tubulin. Bar graphs represent the means�SEM of 3experiments. *P�0.05.
C
B
A
Figure 4. Downregulation of SSH1L phosphatase by siRNA.VSMCs were transfected with siSSH1L or nonsilencing siRNA(siScr) 3 to 4 days before the experiments. SSH1L RNA wasquantified by real-time PCR (A), protein was evaluated by West-ern blot (B), and phosphatase activity in response to PDGF (10ng/mL, 30 minutes) was evaluated as the amount of phosphateproduced in 10 minutes, as described in Materials and Methods(C). Data represent means�SEM of 3 independent experimentsfor A through C. *P�0.05.
San Martın et al LIMK and SSH1L in PDGF-Induced VSMC Migration 435
by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from
ation of actin filaments. Multiple actin-binding proteinsregulate these processes. For instance, the Arp2/3 complex(controlled by WAVE/Scar, WASP, and N-WASP proteins)control nucleation and branching, whereas the formins, mam-malian diaphanous proteins mDia1 and mDia2 (regulated byRhoA and Cdc42), are involved in extension of new actinfilaments.24 Profilin must be liberated from the capped end ofactin filaments, after which it promotes G-actin nucleotideexchange.25 Cofilin stimulates disassembly and severance ofactin filaments at or near the pointed ends and therebycontinuously supplies actin monomers for polymerization andrapid turnover of actin filaments.3 One of the major effects ofcofilin activation is to generate free barbed ends on actinfilaments. Even though PDGF is one of the major migratorystimuli for VSMC migration, virtually nothing is knownabout its ability to regulate these pathways.
In this study, we focused on the ability of PDGF to regulatecofilin because it has been shown to be critically importantfor migration. In human erythroleukemia cells, expression ofeither constitutively active or inactive cofilin inhibits migra-tion, highlighting the importance of cofilin-mediated actinturnover.26 Furthermore, the up- or downregulation of cofilinactivity has been shown to correlate with proportional in-creases or decreases in motility of tumor cells, respectively.16
We found that PDGF stimulation of VSMC migration wasaccompanied by cofilin activation (Figure 2). Moreover, thisactivity was required for cell movement, because preventingdephosphorylation by siSSH1L inhibited migration (Figure8). This suggests that the mechanisms controlling cofilin
p-cofilin
cofilin
control controlPDGF PDGF
siScr siSSH1L
**
*
siScr siSSH1L
PDGF PDGF0
10
20
p-co
filin
den
sito
met
ric u
nits
control control
Figure 5. PDGF-induced cofilin dephosphorylation requiresSSH1L phosphatase. VSMCs were transfected with siSSH1Lor scrambled control (siScr). The cells were serum-starved for48 hours and treated for 30 minutes with 10 ng/mL of PDGF.Phosphorylation of cofilin was evaluated using phosphospe-cific antibodies. The graph shows the mean�SEM of densito-metric analysis from 5 independent experiments. *P�0.05and **P�0.001.
p-LIMK
LIMK
0 1 5 10 15 20 30 60
Time (min)
p-LI
MK
den
sito
met
ric u
nits
*
*
* *
0 1 5 10 15 20 30 600
25
50
75
100
125
Figure 6. LIMK is activated by PDGF. VSMCs were stimulatedfor the indicated times with PDGF (10 ng/mL), and LIMK phos-phorylation (p-LIMK) was determined by Western blot using aphosphospecific antibody. The graph shows the mean�SEM ofdensitometric analysis from 3 independent experiments.*P�0.05.
Figure 7. LIMK is responsible for PDGF-induced cofilin phos-phorylation (p-cofilin). VSMCs were transfected with either non-silencing siRNA (siScr) or siSSH1L for 72 hours and thenretransfected with pEGFP-KS (encoding kinase-inactive LIMK)or pEGFP (control) plasmids for an additional 15 hours. Cellswere then stimulated with 10 ng/mL PDGF for 5 minutes. Cofilinphosphorylation was evaluated by Western blot. Values werenormalized to those from control cells for each treatment (datanot shown). The graph shows the mean�SEM of densitometricanalysis from 3 independent experiments. *P�0.05.
0
100
200
300
400
500
600
mig
rate
d ce
lls /
field
*
control siScr siSSH1L
control siScr siSSH1L
Figure 8. PDGF-induced migration is dependent on SSH1L.VSMCs were starved for 48 hours, trypsinized, and reseeded ina collagen-coated Transwell insert. After stimulation for 3 hourswith PDGF (10 ng/mL), the cells migrating through the mem-brane were stained with DAPI and counted. The top imagesare representative of DAPI staining. The graph representsmean�SEM of migrated cells per field from 3 independentexperiments. *P�0.01.
436 Circulation Research February 29, 2008
by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from

activation are critical to the regulation of PDGF-inducedVSMC motility.
The apparently simultaneous dual regulation of cofilin byLIMK and SSH1L by PDGF in VSMCs may represent afine-tuned control of directional movement. It is surprisingthat we were unable to detect cofilin phosphorylation even atearly times after PDGF stimulation when LIMK was acti-vated. The appearance of cofilin phosphorylation at 5 min-utes, when SSH1L is inhibited, and the ability of dominantnegative LIMK to block this phosphorylation (Figure 7),confirm that LIMK is in fact targeting cofilin. Consistent withthis, at 30 minutes, when LIMK is no longer active (Figure 6),downregulation of SSH1L no longer unmasks increasedcofilin phosphorylation (Figure 5). In Jurkat T cells stimulatedwith stromal cell derived factor-1�, LIMK was found diffuselyin the cytoplasm, whereas SSH1L was localized to lamellipo-dia, but activation of both proteins was necessary for chemo-taxis.17 This suggests that the spatial relationship betweenLIMK/cofilin/SSH1L is crucial for the polarization of cellmovement. Alternatively, it may be the temporal relationshipbetween LIMK and SSH1L activation that dictates the mi-gratory response. After 30 minutes, LIMK activation hasreturned to baseline, whereas SSH1L activity is still robust(Figures 3 and 4). It has been shown that SSH1L candephosphorylate LIMK,27 which may explain the transientnature of LIMK activation, contrasting with a long-termactivation of the phosphatase in PDGF-stimulated cells, as wededuce from the sustained SSH1L upregulation at messengerand protein level. Ultimately, it is the dynamic ratio of activeLIMK to active SSH1L that is important. In VSMCs, thepredominant effect of PDGF appears to be stimulation ofSSH1L activity and dephosphorylation of cofilin.
SSH1L is a member of the slingshot family of phospha-tases that includes SSH1L, SSH2L, and SSH3L. Each ofthese proteins has distinct specific activities and subcellulardistributions.12,14 Our results show that although all 3 SSHisoforms are expressed in VSMCs, SSH1L is activated byPDGF (Figure 3), and, for the first time, clearly demonstratethat it is required for PDGF-mediated migration (Figure 8).The mechanisms that regulate SSH1L activity remain to beelucidated. It has been shown that ATP or histamine-inducedSSH1L activation is mediated by calcium-dependent stimu-lation of calcineurin28 and that dephosphorylation of SSH1Lon Ser978 increases its activity.11 However, calcineurin doesnot mediate PDGF-induced migration in fibroblasts.29 Incontrast, insulin stimulation of SSH1L activity is dependenton phosphatidylinositol 3-kinase,30 a signaling pathway pro-posed to be important in PDGF-induced VSMC migration insome types of smooth muscle cells.31 Determining the up-stream signaling mechanisms responsible for SSH1L activa-tion in migrating VSMC will require further study.
In summary, we find that one of the key factors regulatingPDGF-induced migration is the phosphorylation state ofcofilin. PDGF activates both the cofilin kinase LIMK and thenovel phosphatase SSH1L, the latter of which predominatesto dephosphorylate cofilin. In accordance with these findings,blocking SSH1L significantly inhibits PDGF-induced migra-tion. These results suggest that PDGF-induced migrationrequires fine control of cofilin phosphorylation and that
SSH1L plays a critical role in maintaining cofilin activityduring PDGF-induced migration of VSMCs.
AcknowledgmentWe are grateful to Dr John Condeelis for providing the KS-LIMKplasmid.
Sources of FundingThis work was supported by NIH grants HL38206 and HL058863 (toK.K.G.) and American Heart Association Fellowship 0525465B(to A.S.M.).
DisclosuresNone.
References1. Clowes AW. Regulation of smooth muscle cell proliferation and
migration. Transplant Proc. 1999;31:810–811.2. dos Remedios CG, Chhabra D, Kekic M, Dedova IV, Tsubakihara M,
Berry DA, Nosworthy NJ. Actin binding proteins: regulation ofcytoskeletal microfilaments. Physiol Rev. 2003;83:433–473.
3. Chen H, Bernstein BW, Bamburg JR. Regulating actin-filament dynamicsin vivo. Trends Biochem Sci. 2000;25:19–23.
4. Arber S, Barbayannis FA, Hanser H, Schneider C, Stanyon CA, BernardO, Caroni P. Regulation of actin dynamics through phosphorylation ofcofilin by LIM-kinase. Nature. 1998;393:805–809.
5. Toshima J, Toshima JY, Amano T, Yang N, Narumiya S, Mizuno K.Cofilin phosphorylation by protein kinase testicular protein kinase 1 andits role in integrin-mediated actin reorganization and focal adhesionformation. Mol Biol Cell. 2001;12:1131–1145.
6. Yang N, Higuchi O, Ohashi K, Nagata K, Wada A, Kangawa K, NishidaE, Mizuno K. Cofilin phosphorylation by LIM-kinase 1 and its role inRac-mediated actin reorganization. Nature. 1998;393:809–812.
7. Dawe HR, Minamide LS, Bamburg JR, Cramer LP. ADF/cofilin controlscell polarity during fibroblast migration. Curr Biol. 2003;13:252–257.
8. Zebda N, Bernard O, Bailly M, Welti S, Lawrence DS, Condeelis JS.Phosphorylation of ADF/cofilin abolishes EGF-induced actin nucleationat the leading edge and subsequent lamellipod extension. J Cell Biol.2000;151:1119–1128.
9. Bamburg JR. Proteins of the ADF/cofilin family: essential regulators ofactin dynamics. Annu Rev Cell Dev Biol. 1999;15:185–230.
10. Meberg PJ, Ono S, Minamide LS, Takahashi M, Bamburg JR. Actin depo-lymerizing factor and cofilin phosphorylation dynamics: response to signalsthat regulate neurite extension. Cell Motil Cytoskeleton. 1998;39:172–190.
11. Nagata-Ohashi K, Ohta Y, Goto K, Chiba S, Mori R, Nishita M, OhashiK, Kousaka K, Iwamatsu A, Niwa R, Uemura T, Mizuno K. A pathwayof neuregulin-induced activation of cofilin-phosphatase Slingshot andcofilin in lamellipodia. J Cell Biol. 2004;165:465–471.
12. Ohta Y, Kousaka K, Nagata-Ohashi K, Ohashi K, Muramoto A, Shima Y,Niwa R, Uemura T, Mizuno K. Differential activities, subcellular distri-bution and tissue expression patterns of three members of Slingshotfamily phosphatases that dephosphorylate cofilin. Genes Cells. 2003;8:811–824.
13. Gohla A, Birkenfeld J, Bokoch GM. Chronophin, a novel HAD-typeserine protein phosphatase, regulates cofilin-dependent actin dynamics.Nat Cell Biol. 2005;7:21–29.
14. Kaji N, Ohashi K, Shuin M, Niwa R, Uemura T, Mizuno K. Cellcycle-associated changes in Slingshot phosphatase activity and roles incytokinesis in animal cells. J Biol Chem. 2003;278:33450–33455.
15. Weber DS, Taniyama Y, Rocic P, Seshiah PN, Dechert MA, GerthofferWT, Griendling KK. Phosphoinositide-dependent kinase 1 and p21-acti-vated protein kinase mediate reactive oxygen species-dependent regu-lation of platelet-derived growth factor-induced smooth muscle cellmigration. Circ Res. 2004;94:1219–1226.
16. Wang W, Mouneimne G, Sidani M, Wyckoff J, Chen X, Makris A,Goswami S, Bresnick AR, Condeelis JS. The activity status of cofilin isdirectly related to invasion, intravasation, and metastasis of mammarytumors. J Cell Biol. 2006;173:395–404.
17. Nishita M, Tomizawa C, Yamamoto M, Horita Y, Ohashi K, Mizuno K.Spatial and temporal regulation of cofilin activity by LIM kinase andSlingshot is critical for directional cell migration. J Cell Biol. 2005;171:349–359.
San Martın et al LIMK and SSH1L in PDGF-Induced VSMC Migration 437
by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from

18. Seppa H, Grotendorst G, Seppa S, Schiffmann E, Martin GR. Platelet-derived growth factor in chemotactic for fibroblasts. J Cell Biol. 1982;92:584–588.
19. Lorenz M, DesMarais V, Macaluso F, Singer RH, Condeelis J. Mea-surement of barbed ends, actin polymerization, and motility in live car-cinoma cells after growth factor stimulation. Cell Motil Cytoskeleton.2004;57:207–217.
20. Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, BorisyG, Parsons JT, Horwitz AR. Cell migration: integrating signals from frontto back. Science. 2003;302:1704–1709.
21. DesMarais V, Ghosh M, Eddy R, Condeelis J. Cofilin takes the lead.J Cell Sci. 2005;118:19–26.
22. Ghosh M, Song X, Mouneimne G, Sidani M, Lawrence DS, Condeelis JS.Cofilin promotes actin polymerization and defines the direction of cellmotility. Science. 2004;304:743–746.
23. Wear MA, Schafer DA, Cooper JA. Actin dynamics: assembly anddisassembly of actin networks. Curr Biol. 2000;10:R891–R895.
24. Gerthoffer WT. Mechanisms of vascular smooth muscle cell migration.Circ Res. 2007;100:607–621.
25. Krause M, Dent EW, Bear JE, Loureiro JJ, Gertler FB. Ena/VASPproteins: regulators of the actin cytoskeleton and cell migration. Annu RevCell Dev Biol. 2003;19:541–564.
26. Konakahara S, Ohashi K, Mizuno K, Itoh K, Tsuji T. CD29 integrin- andLIMK1/cofilin-mediated actin reorganization regulates the migration ofhaematopoietic progenitor cells underneath bone marrow stromal cells.Genes Cells. 2004;9:345–358.
27. Soosairajah J, Maiti S, Wiggan O, Sarmiere P, Moussi N, Sarcevic B,Sampath R, Bamburg JR, Bernard O. Interplay between components of anovel LIM kinase-slingshot phosphatase complex regulates cofilin.EMBO J. 2005;24:473–486.
28. Wang Y, Shibasaki F, Mizuno K. Calcium signal-induced cofilin dephos-phorylation is mediated by Slingshot via calcineurin. J Biol Chem. 2005;280:12683–12689.
29. Yang S, Huang XY. Ca2� influx through L-type Ca2� channels controlsthe trailing tail contraction in growth factor-induced fibroblast cellmigration. J Biol Chem. 2005;280:27130–27137.
30. Nishita M, Wang Y, Tomizawa C, Suzuki A, Niwa R, Uemura T, MizunoK. Phosphoinositide 3-kinase-mediated activation of cofilin phosphataseSlingshot and its role for insulin-induced membrane protrusion. J BiolChem. 2004;279:7193–7198.
31. Yamboliev IA, Chen J, Gerthoffer WT. PI 3-kinases and Src kinasesregulate spreading and migration of cultured VSMCs. Am J Physiol CellPhysiol. 2001;281:C709–C718.
438 Circulation Research February 29, 2008
by guest on February 19, 2014http://circres.ahajournals.org/Downloaded from

Online Supplement San Martín et al. Dual regulation of cofilin activity by LIMK and…
Supplementary Figure 1: KS-LIMK indeed reduces basal cofilin phosphorylation.
VSMCs were transfected using electroporation with either pEGFP-KS (encoding kinase
inactive LIMK) or pEGFP (control) plasmids for 15 h. Cofilin phosphorylation was
evaluated by Western blot. Graph shows the mean ± SEM of densitometric analysis from
3 independent experiments. * p<0.05.
C KSp-cofilint-cofilin
0
50
100
control KSp-co
filin
den
sito
met
ricun
its(p
erce
nt c
ontro
l)
*
Supplementary Figure 1
Copyright © 2022 FDOKUMEN




















