
Drivers of β-diversity along latitudinal gradients revisited
12
RESEARCH PAPER Drivers of b-diversity along latitudinal gradients revisited Hong Qian 1 *, Shengbin Chen 2,3 , Lingfeng Mao 4,5 and Zhiyun Ouyang 3 1 Research and Collections Center, Illinois State Museum, 1011 East Ash Street, Springfield, IL 62703, USA; 2 Nanjing Institute of Environmental Sciences Ministry of Environmental Protection, Nanjing, 210042, China; 3 State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China; 4 State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China; 5 Graduate School of Chinese Academy of Sciences, Beijing 100049, China ABSTRACT Aim Ecologists have generally agreed that b-diversity is driven at least in part by ecological processes and mechanisms of community assembly and is a key deter- minant of global patterns of species richness. This idea has been challenged by a recent study based on an individual-based null model approach, which aims to account for the species pool. The goal of the present study is twofold: (1) to analyse data sets from different parts of the world to determine whether there are signifi- cant latitude–b-diversity gradients after accounting for the species pool, and (2) to evaluate the validity of the null model. Location Global. Methods A total of 257 forest plots, each being 0.1 ha in size and having 10 0.01-ha subplots, were used. We conducted four sets of analyses. A modified version of Whittaker’s b-diversity index was used to quantify b-diversity for each forest plot. A randomization procedure was used to determine expected b-diversity. Results The number of individuals per species, which characterizes species abun- dance distribution, alone explains 56.8–84.2% of the variation in observed b-diversity. Species pool (g-diversity) explained only an additional 2.6–15.2% of the variation in observed b-diversity. Latitude explains 18.6% of the variation in raw b deviation in Gentry’s global data set, and explains 11.0–11.6% of the varia- tion in standardized b deviation in the global and three regional analyses. Latitude explains 33.2–46.2% of the variation in the number of individuals per species. Main conclusions Species abundance distribution, rather than species pool size, plays a key role in driving latitude–b-diversity gradients for b-diversity in local forest communities. The individual-based null model is not a valid null model for investigating b-diversity gradients driven by mechanisms of local community assembly because the null model incorporates species abundance distributions, which are driven by mechanisms of local community assembly and in turn generate b-diversity gradients. Keywords Beta diversity, gamma diversity, latitudinal diversity gradient, mechanisms of community assembly, null model, species pool, species turnover, woody plants. *Correspondence: Hong Qian, Research and Collections Center, Illinois State Museum, 1011 East Ash Street, Springfield, IL 62703, USA. E-mail: [email protected] INTRODUCTION The latitudinal diversity gradient – the number of species per unit area decreases from the equator to poles – has been known for over a century (Darwin, 1859; Wallace, 1878), but the causes of this diversity gradient remain poorly resolved (Mittelbach et al., 2007). Species diversity is often described with three components: a-, b- and g-diversity (Whittaker, 1972). The a-diversity measures species richness within local sampling units, g-diversity measures total species richness in a region in which local sampling units are located and b-diversity quantifies the variation in species composition among the sampling units and thus represents the scalar that links a-diversity and g-diversity and unifies local-regional diversity relationships (Ricklefs, 1987). Because both latitude and species diversity are generally related to some environmental factors, particularly Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2012) ••, ••–•• © 2012 Blackwell Publishing Ltd DOI: 10.1111/geb.12020 http://wileyonlinelibrary.com/journal/geb 1
Transcript of Drivers of β-diversity along latitudinal gradients revisited

RESEARCHPAPER
Drivers of b-diversity along latitudinalgradients revisitedHong Qian1*, Shengbin Chen2,3, Lingfeng Mao4,5 and Zhiyun Ouyang3
1Research and Collections Center, Illinois State
Museum, 1011 East Ash Street, Springfield,
IL 62703, USA; 2Nanjing Institute of
Environmental Sciences Ministry of
Environmental Protection, Nanjing, 210042,
China; 3State Key Laboratory of Urban and
Regional Ecology, Research Center for
Eco-Environmental Sciences, Chinese Academy
of Sciences, Beijing 100085, China; 4State Key
Laboratory of Vegetation and Environmental
Change, Institute of Botany, Chinese Academy
of Sciences, Beijing 100093, China; 5Graduate
School of Chinese Academy of Sciences, Beijing
100049, China
ABSTRACT
Aim Ecologists have generally agreed that b-diversity is driven at least in part byecological processes and mechanisms of community assembly and is a key deter-minant of global patterns of species richness. This idea has been challenged by arecent study based on an individual-based null model approach, which aims toaccount for the species pool. The goal of the present study is twofold: (1) to analysedata sets from different parts of the world to determine whether there are signifi-cant latitude–b-diversity gradients after accounting for the species pool, and (2) toevaluate the validity of the null model.
Location Global.
Methods A total of 257 forest plots, each being 0.1 ha in size and having 100.01-ha subplots, were used. We conducted four sets of analyses. A modified versionof Whittaker’s b-diversity index was used to quantify b-diversity for each forestplot. A randomization procedure was used to determine expected b-diversity.
Results The number of individuals per species, which characterizes species abun-dance distribution, alone explains 56.8–84.2% of the variation in observedb-diversity. Species pool (g-diversity) explained only an additional 2.6–15.2% ofthe variation in observed b-diversity. Latitude explains 18.6% of the variation inraw b deviation in Gentry’s global data set, and explains 11.0–11.6% of the varia-tion in standardized b deviation in the global and three regional analyses. Latitudeexplains 33.2–46.2% of the variation in the number of individuals per species.
Main conclusions Species abundance distribution, rather than species pool size,plays a key role in driving latitude–b-diversity gradients for b-diversity in localforest communities. The individual-based null model is not a valid null model forinvestigating b-diversity gradients driven by mechanisms of local communityassembly because the null model incorporates species abundance distributions,which are driven by mechanisms of local community assembly and in turn generateb-diversity gradients.
KeywordsBeta diversity, gamma diversity, latitudinal diversity gradient, mechanisms ofcommunity assembly, null model, species pool, species turnover, woody plants.
*Correspondence: Hong Qian, Research andCollections Center, Illinois State Museum, 1011East Ash Street, Springfield, IL 62703, USA.E-mail: [email protected]
INTRODUCTION
The latitudinal diversity gradient – the number of species per
unit area decreases from the equator to poles – has been known
for over a century (Darwin, 1859; Wallace, 1878), but the causes
of this diversity gradient remain poorly resolved (Mittelbach
et al., 2007). Species diversity is often described with three
components: a-, b- and g-diversity (Whittaker, 1972). The
a-diversity measures species richness within local sampling
units, g-diversity measures total species richness in a region in
which local sampling units are located and b-diversity quantifies
the variation in species composition among the sampling units
and thus represents the scalar that links a-diversity and
g-diversity and unifies local-regional diversity relationships
(Ricklefs, 1987). Because both latitude and species diversity are
generally related to some environmental factors, particularly
bs_bs_banner
Global Ecology and Biogeography, (Global Ecol. Biogeogr.) (2012) ••, ••–••
© 2012 Blackwell Publishing Ltd DOI: 10.1111/geb.12020http://wileyonlinelibrary.com/journal/geb 1

temperature (Pianka, 1966; Rohde, 1992; Gaston, 2000), latitu-
dinal environmental gradients have been proposed as a major
driver of the latitudinal diversity gradient (Rohde, 1992; Allen
et al., 2002).
Several studies have shown that b-diversity is greater at lower
latitudes (e.g. Qian & Ricklefs, 2007, 2012; Buckley & Jetz, 2008;
Qian, 2009; Qian et al., 2009; Lenoir et al., 2010; Chen et al.,
2011) and in areas with higher environmental energy (e.g. Qian
& Xiao, 2012). Tropical species are expected to be specialists
along environmental gradients, which would result in rapid
species turnover (high b-diversity) among localities (Jankowski
et al., 2009). In contrast, species at high latitudes tend to have
wide distribution ranges and wide tolerance to environmental
variation (Stevens, 1989), leading to low species turnover (low
b-diversity) among localities. Ecologists have generally agreed
that b-diversity is a key determinant of global patterns of species
richness (e.g. Stevens, 1989; Koleff et al., 2003; Qian & Ricklefs,
2012). Greater b-diversity has been considered as a partial cause
of higher regional species richness in tropical regions (Koleff
et al., 2003; Qian & Ricklefs, 2007; Qian et al., 2009; Chen et al.,
2011).
Recently, Kraft et al. (2011) have challenged the idea that
differences in the mechanisms of local community assembly
in temperate versus tropical regions may have played a role
in driving the latitudinal gradient of b-diversity. They used a
null model approach to examine the relationship between
b-diversity and latitude in a data set with 197 forest sites from
different continents. Specifically, they used a modified version of
Whittaker’s (1960) b-diversity index to quantify observed
b-diversity for each forest site, used a randomization procedure
to determine expected b-diversity, and related standard differ-
ences between the observed and expected values of b-diversity
(i.e. standardized b deviation) to absolute latitude. Because they
found that standardized b deviation is not linearly correlated
with absolute latitude, they concluded that there is no need to
invoke differences in the mechanisms of local community
assembly in tropical and temperate regions to explain global
patterns of b-diversity. However, as Qian et al. (2012) point out,
there are several major problems with their study.
One major problem with Kraft et al. (2011) is that they
included all sample sites from different continents across the
world in a single analysis to examine the b-diversity and latitude
relationship. It is well known that environmental factors that
drive species diversity gradients are not constant at any given
latitude; rather, they can vary substantially among regions
within and between continents. For example, temperature is
considered as a key driver of latitudinal diversity gradient; the
same temperature can be found more than 30° of latitude apart
within continents and over 40° of latitude apart between conti-
nents (New et al., 1999). Furthermore, at a given latitude, the
Southern Hemisphere is generally warmer than the Northern
Hemisphere (Ahrens, 2007), and b-diversity at the same latitude
is higher in the Southern Hemisphere than in the Northern
Hemisphere (Chen et al., 2011). Therefore, latitude is a poor
surrogate for the underlying environmental drivers of species
diversity in an analysis including sampling units from different
continents and from both Southern and Northern hemispheres.
Consequently, diversity patterns would be obscured in such
analyses. Our first objective in the present study is to use Kraft
et al.’s null model approach to analyse each regional data set
constrained within a relatively narrow longitudinal band.
Forests in eastern Asia generally form an unbroken latitudinal
gradient of forest vegetation, which extends from the tree line at
the Siberian Arctic and Far East in Russia southward to the tip of
the Malay Peninsula near the equator and is one of the longest
latitudinal continua of forest vegetation in the world. However,
forests in this region are poorly represented in Gentry’s data set
(Fig. S1 in Supporting Information). Accordingly, we include a
new data set with 60 forest sites from the eastern part of China
in this study. This region is ideal for investigating the
b-diversity–latitude relationship because it covers a wide range
of latitudes, includes all major types of forest vegetation in
eastern Asia (i.e. tropical, subtropical, temperate and boreal
forests), and has a great variety of climate types, and thus the
latitudinal gradient in the eastern part of China represents a
strong ecological gradient.
Before we ran Kraft et al.’s (2011) R code of their null model
on our data, we first ran the R code on the data for Gentry’s 197
plots in order to make sure that we have correctly understood
and utilized their null model. After comparing our results with
those from Kraft et al. (2011), we found that values of standard-
ized b deviation can differ by more than four times between
their results and ours for the same plots. We checked the data set
that was used in Kraft et al. (2011) (provided by Nathan Kraft)
and found many errors in their data set. For example, the
number of individuals in the Alto de Cuevas site (CUEVAS) in
Colombia is 424 in Kraft et al.’s data set but is only 358 in
Gentry’s original data set; in contrast, the number of individuals
in the Brise Fer site (BRISEFER) in Mauritius is 1010 in Gentry’s
original data set but is only 426 in Kraft et al.’s data set. Accord-
ingly, in addition to analysing regional data sets, our first objec-
tive also includes the reanalysis of the data of the 197 sites using
Gentry’s original data set to determine the degree to which the
errors in Kraft et al.’s data set have biased their conclusion.
Our second objective is to evaluate the validity of Kraft et al.’s
null model for the question that they investigated, i.e. whether
b-diversity is correlated with latitude after accounting for
species pool (g-diversity).
MATERIALS AND METHODS
Data sets
We used two comprehensive data sets of 0.1-ha forest plots: one
includes 197 plots from Gentry’s data set (Fig. S1), and the other
includes 60 plots from the eastern part of China (Fig. S2). Gen-
try’s entire data set includes 226 plots, but 29 plots are either
not 0.1-ha plots or do not contain 10 subplots. The Gentry’s
197 plots were obtained from the website located at http://
www.wlbcenter.org/gentry_data.htm (the same data set is also
available at http://www.mobot.org/MOBOT/Research/gentry/
transect.shtml and salvias.net/Plots/index.php). These 197 plots
H. Qian et al.
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd2

were used in Kraft et al. (2011). Each plot includes 10 0.01-ha
subplots. Woody stems equal to or greater than 2.5 cm diameter
at breast height (d.b.h.; at 1.3 m above ground) were measured
and identified to species or morphospecies (Phillips & Miller,
2002). Gentry’s data set includes the number of individuals and
the number of stems for each species in each subplot. Because
some individuals each have two or more stems, the number of
measured stems in a plot is often greater than the number of
individuals in the plot. Kraft et al.’s null model is an individual-
based randomization approach, and they used individual data in
their study; accordingly, we also used individual data in our
study. Of Gentry’s 197 plots, 158 (80%) are located in the New
World; the remaining 39 plots are scattered in the four conti-
nents of the Old World (Fig. S1), and no meaningful latitudinal
gradients can be assembled from those 39 plots. Thus, we assem-
bled two latitudinal gradients from the New World plots: one
includes all 72 plots located south of the equator and the other
includes 79 plots located north of the equator and east of
100° W longitude (Fig. S1). We excluded a few plots in the
western coast of North America because the same temperature
at a latitude in eastern North America can be found at a latitude
more than 20° further north in western North America, or tem-
perature at the same latitude can differ by over 15 °C between
eastern and western North America (Ahrens, 2007). Informa-
tion for latitude, longitude and elevation of each plot was
obtained from Phillips & Miller (2002). The 72 plots in the New
World (south) cover a latitudinal gradient of 40°, and the 79
plots in the New World (north) cover a latitudinal gradient of
47° (Fig. S1).
The 60 forest plots from China were sampled in 15 nature
reserves (Fig. S2). These nature reserves cover a latitudinal gra-
dient of 35° from tropical rain forests to boreal forests. Four
forest plots were sampled in each nature reserve. Each forest plot
was 20 m ¥ 50 m in size and was divided into 10 0.01-ha sub-
plots (10 m ¥ 10 m). Latitude, longitude and elevation of each
plot were measured with a global positioning system (GPS) unit.
Woody individuals with d.b.h. equal to or greater than 3 cm
were measured and identified to species.
Data analysis
We used the same approach as in Kraft et al. (2011) to measure
a-, b- and g-diversity for each plot. Specifically, we define
a-diversity as the number of species in a single 0.01-ha subplot,
the g-diversity of the plot as the total number of species in the 10
subplots of the plot and b-diversity as b = 1 – a/g, where a is the
average of the 10 a-diversity values of the plot.
We used Kraft et al.’s (2011) R code to calculate expected
b-diversity and standardized b deviation based on a null model.
Details of the null model approach are as follows. Consider a
data matrix with N individuals belonging to M species in a set of
10 subplots within a plot as in Kraft et al. First, b-diversity
among the 10 subplots in the raw data is calculated, and the
resulting value is called observed b-diversity. Second, an
individual-based randomization procedure is used to determine
expected b-diversity. In each randomization, the N individuals
are randomly shuffled and then assigned to the 10 subplots; each
subplot receives the same number of individuals as in the raw
data. In other words, each randomization not only maintains the
total number of individuals and the total number of species for
that plot as in the raw data but also preserves the number of
individuals in each subplot (i.e. maintaining species abundance
distribution patterns among the 10 subplots) for that plot as in
the raw data. The value of b-diversity in each randomized data
set is calculated for each randomization, and the average
b-diversity of 1000 randomizations for each plot is calculated
and is called expected b-diversity. Third, raw b deviation is
calculated as the observed b-diversity minus the mean of the
null distribution of b-diversity values. Fourth, standardized bdeviation, which is simply called b deviation in Kraft et al.
(2011), is calculated as raw b deviation divided by the standard
deviation of the null distribution (Kraft et al., 2011). Kraft et al.
consider standardized b deviation as b-diversity after account-
ing for sampling effects of species pool.
We used correlation coefficients and coefficients of determi-
nation from regressions to assess the relationships among lati-
tude, measures of diversity and species abundance distribution.
Because species abundance distribution is a vector of the abun-
dances of each species in a site rather than a univariate variable,
because the vector of the abundances cannot be directly used in
correlation and regression analyses, and because the number of
individuals per species in a site is a characterization of the rela-
tionship between the number of individuals and the number of
species and thus a characterization of species abundance distri-
bution in a site, we used the number of individuals per species as
a summary value for species abundance distribution in correla-
tion and regression analyses.
Elevation varies greatly among plots within each data set and
there are significant relationships between elevation and latitude
(e.g. r = -0.356, P < 0.05, for the New World north date set). The
average temperature lapse rate, at which the air temperature
decreases with elevation, is about 6.5 °C for every 1000 m rise in
elevation (Ahrens, 2007). An upward shift of 100 m is predicted
to translate into a poleward shift of 100 km in the temperate
zone (Jump et al., 2009; Stephenson & Das, 2011). Accordingly,
in addition to using the original latitude of each plot, we also
used adjusted latitudes based on the converter of 100-m eleva-
tion for 100-km latitude (Qian et al., 2012) in analyses with the
three regional data sets. We didn’t use adjusted latitude in the
analysis with the whole set of Gentry’s 197 plots because the
purpose of this analysis is, as noted above, to repeat Kraft et al.’s
(2011) analysis, which only used the original latitudes of the 197
plots.
RESULTS AND DISCUSSION
Latitudinal gradients of b-diversity and b deviation
For the global data set with Gentry’s 197 plots, values of both
observed and expected b-diversity are negatively and signifi-
cantly correlated with latitude (Fig. 1a). Raw b deviation (i.e.
observed b-diversity minus expected b-diversity) is positively
Latitudinal gradients of b-diversity
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd 3

and significantly correlated with latitude (r = 0.341, P < 0.05;
Fig. 1b), indicating that 11.6% of the variation in b-diversity
was explained by latitude in a linear regression after accounting
for b-diversity generated by the randomization process of the
null model. Standardized b deviation is negatively and signifi-
cantly correlated with latitude (r = -0.175, P < 0.05; Fig. 1c).
This result differs from that of Kraft et al. (2011). We acknowl-
edge that even though we used the same large number of rand-
omizations (n = 1000) in each model run (analysis) as in Kraft
et al., resulting values may vary slightly among different runs
due to the nature of randomization. To examine if the difference
in the results between their and our studies is due to the varia-
tion among different runs of randomizations, we first repeated
the analysis 10 times each with 1000 randomizations and calcu-
lated correlation coefficient between standardized b deviation
and latitude for each run. The 10 correlation coefficients range
from -0.181 to -0.171 (mean � SD, -0.176 � 0.003; P < 0.05 in
all cases). We then ran the data set that Kraft et al. used; we
obtained the same correlation coefficient (r = -0.02) between
standardized b deviation and latitude as in Kraft et al. (2011),
which is far beyond the range of the 10 correlation coefficients
based on our data. All these indicate that the difference in cor-
relation coefficient between their and our analyses is not due to
the variation among different runs of randomizations on the
same data. The relationship between b deviation and latitude
tended to be curvilinear (Fig. 1b, c). Latitude in a second-order
polynomial regression explained 18.6 and 11.0% of the
variation, respectively, in raw b deviation and standardized bdeviation.
The negative relationship between the observed b-diversity
and latitude is significant in all the three regional data sets (i.e.
New World north, New World south and China) regardless of
whether original latitude or adjusted latitude was used (Fig. 2a–
d). The same is true for expected b-diversity (Fig. 2d–f).
Although the linear relationship between b-diversity and origi-
nal latitude tended to be parallel to that between b-diversity and
adjusted latitude, the distance separating the two linear lines of
each region varies among regions. For example, the distance is
larger in China than in the New World. This pattern suggests
that it is necessary to take into account elevations of sampling
units in b-diversity studies, particularly in the case that sampling
units represent local-scale (fine-grain) localities.
Raw b deviation increases with both original and adjusted
latitudes in all three regions, and the positive relationship
between raw b deviation and adjusted latitude is significant (P <0.05) for all regions (Fig. 3a–c). Standardized b deviation sig-
nificantly decreases with original latitude for New World north
and China (Fig. 3d, f). When related to adjusted latitude, stand-
ardized b deviation significantly decreases with latitude for New
World north and China, and significantly increases with latitude
for New World south (Fig. 3). Latitude explained 10.9, 11.6 and
11.2% of the variation in standardized b deviation, respectively,
for New World north, New World south and China. The fact that
the relationship between standardized b deviation and latitude
can be in opposite directions in different regions supports our
notion that latitude-based analyses of biodiversity should be
0 10 20 30 40 50 60
β-di
vers
ity
0.4
0.5
0.6
0.7
0.8
0.9
Absolute latitude (°)
0 10 20 30 40 50 60
Sta
ndar
dize
d β
devi
atio
n
-5
0
5
10
15
20
a)
c)
r = -0.732, P < 0.05
r = -0.175, P < 0.05
r = -0.702, P < 0.05
observed
expected
R2 = 0.110, P < 0.05
0 10 20 30 40 50 60
Raw
β d
evia
tion
-0.05
0.00
0.05
0.10
0.15
0.20
r = 0.341, P < 0.05R2 = 0.186, P < 0.05
b)
Figure 1 (a) Global relations of observed (filled circles andthicker line) and expected (open circles and thinner line)b-diversity of woody plants with latitude, (b) global relation ofraw b deviation with latitude in the first-order (linear regression,grey) and second-order (quadratic regression, black) polynomials,and (c) global relation of standardized b deviation with latitudein the first-order (linear regression, grey) and second-order(quadratic regression, black) polynomials.
H. Qian et al.
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd4

conducted separately for different regions, rather than conduct-
ing a single analysis including different regions across the globe.
Overall, our study showed that significant correlations were
found in all the four analyses examining the relationship
between standardized b deviation and latitude. Latitude
explained 11–12% of the variation in standardized b deviation
in the global and three regional analyses. However, this amount
of explained variation in standardized b deviation only repre-
sents a small fraction of the variation in b-diversity that may be
driven by mechanisms of community assembly and is related to
latitude. A large amount of the variation in b-diversity in the
sampling units is driven by species abundance distribution (i.e.
the number of individuals of each species in a site), and was
removed by the null model from standardized b deviation.
Species abundance distributions result from mechanisms of
local community assembly and are a major driver of b-diversity
and other macroecological patterns such as the species-area
relationship (Rosenzweig, 1995; He & Legendre, 2002; Ricotta
et al., 2002). When observed b-diversity was regressed on the
number of individuals per species with a linear model, the
number of individuals per species explained 81.4, 80.8, 84.2 and
56.8% of the variation in observed b-diversity, respectively, for
the data sets of Gentry’s 197 plots, New World north, New
World south and China. When observed b-diversity was
regressed on both the number of individuals per species and
g-diversity with a linear model, the two variables explained 86.8,
83.4, 91.8 and 72.0% of the variation in observed b-diversity in
the four respective data sets. In other words, adding g-diversity
to the model explained only an additional 2.6–15% the variation
in observed b-diversity. The number of individuals per species
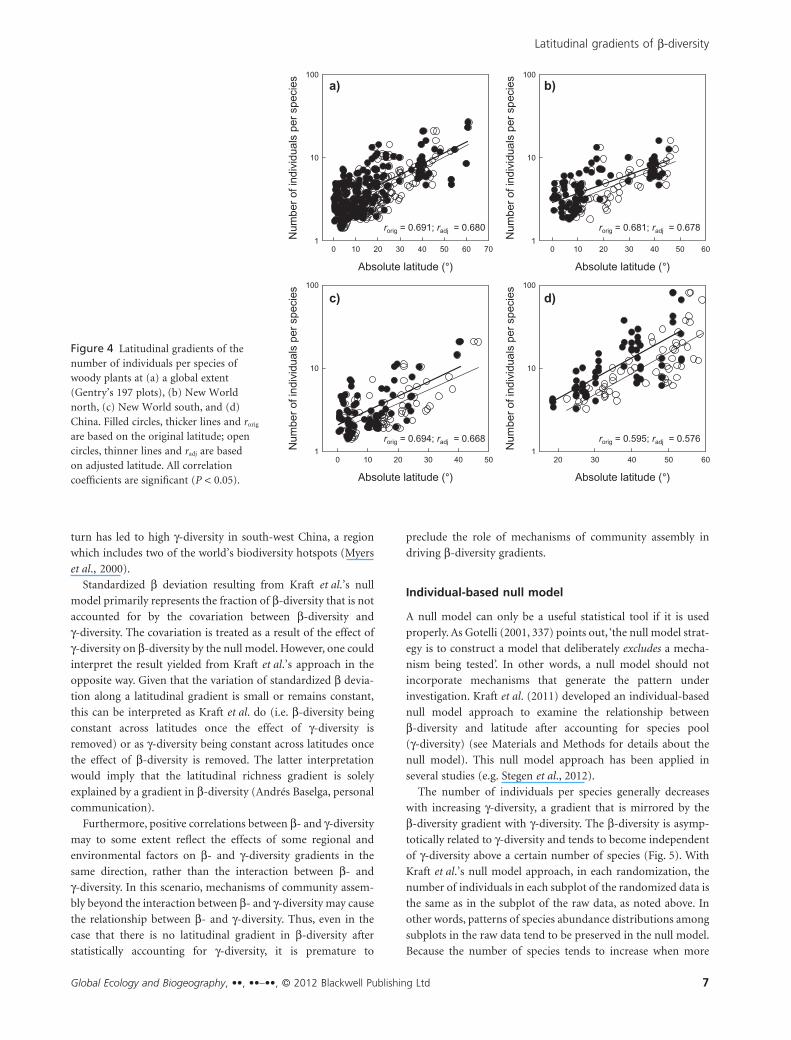
increases with increasing latitude, and latitude explained 33.2–
46.2% of the variation in the number of individuals per species
(Fig. 4). When the variation in observed b-diversity that is
explained by the number of individuals per species was removed
by a second-order polynomial regression, latitude explained
only 0.2–18.6% of the residual of observed b-diversity. This
suggests that the majority of the latitude–b-diversity gradient is
associated with the relationship between b-diversity and species
abundance distributions. Because gradients of species abun-
dance distributions are related to latitude, removing the
b-diversity generated by species abundance distributions
implies that a large part of the relationship between b-diversity
and latitude is removed by the null model (see below for more
discussion). Because standardized b deviation and the number
of individuals per species are largely uncorrelated (e.g. r = 0.017
for the data set with Gentry’s 197 plots), the variation in
b-diversity that would have been explained by latitude would be
much greater than that represented by standardized b deviation
0 10 20 30 40 50
Obs
erve
d β-
dive
rsity
0.4
0.5
0.6
0.7
0.8
0.9
0 10 20 30 40 500.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
20 30 40 50 600.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
a) b) c)
rorig = -0.610, P < 0.05radj = -0.652, P < 0.05
rorig = -0.689, P < 0.05radj = -0.724, P < 0.05
rorig = -0.789, P < 0.05radj = -0.772, P < 0.05
Absolute latitude (°)
0 10 20 30 40 50
Exp
ecte
d β -
dive
rsity
0.4
0.5
0.6
0.7
0.8
0.9
Absolute latitude (°)
0 10 20 30 40 500.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Absolute latitude (°)
20 30 40 50 600.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
rorig = -0.673, P < 0.05radj = -0.706, P < 0.05
rorig = -0.634, P < 0.05radj = -0.672, P < 0.05
rorig = -0.754, P < 0.05radj = -0.796, P < 0.05
d) e) f)
Figure 2 The relations of (a–c) observed and (d–f) expected b-diversity of woody plants with latitude in (a, d) New World north, (b, e)New World south, and (c, f) China. Filled circles, thicker lines and rorig are for the relationship between b-diversity and original latitude;open circles, thinner lines and radj for the relationship between b-diversity and adjusted latitude.
Latitudinal gradients of b-diversity
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd 5

if the relationship between b-diversity and species abundance
distributions had not been removed by the null model. Thus, the
conclusion of our study is contrary to that of Kraft et al. (2011)
on the relationship between b-diversity and latitude after
accounting for species pool.
Relationship between b-diversity and g-diversity
Values of b-diversity and g-diversity are positively correlated (r =0.776 for Gentry’s global dataset, r = 0.746 for China’s dataset,
P < 0.05 in both cases). With Kraft et al.’s null model approach,
such correlations are considered as the effect of g-diversity on
b-diversity. Although it has been hypothesized that the size of a
regional species pool (g-diversity) may be a driving force of
b-diversity (Harrison et al., 2011), this hypothesis has not been
adequately tested and mechanisms of species pools driving
b-diversity are unknown.
Conversely, correlations between b-diversity and g-diversity
may, at least in part, reflect the effect of b-diversity on
g-diversity (Pither & Aarssen, 2005; Meynard et al., 2011; Qian
& Ricklefs, 2012). Indeed, when Robert H. Whittaker initially
proposed the relationships among a-, b- and g-diversity, he
illustrated the role of b-diversity in building larger-scale (g)
diversity patterns (Whittaker, 1960). A larger species pool size
may result from higher b-diversity due to, for example, habitat
heterogeneity (Pither & Aarssen, 2005). For example, if two
contiguous areas each have a large proportion of unique
species, and thus have high b-diversity between them, high
g-diversity of the two areas would be expected. Previous
studies (e.g. Qian et al., 2005) have shown that high g-diversity
of a region results, at least in part, from high b-diversity within
the region. Rodríguez & Arita (2004) also showed that the
higher diversity of non-volant mammals in tropical areas of
North America is a consequence of the increase in b-diversity
towards the tropical areas. Another such example is high
g-diversity in south-west China. The collision of the Indian
subcontinent with the Asian continent during the Eocene
(55–45 million years ago; Sengör & Natal’in, 1996) resulted in
enormous rugged mountains (usually > 6000 m high in the
eastern Himalayas and Hengduanshan regions) separated by
deep valleys. A significant number of species became vicariants
on different mountains during the orogenic processes, which
would favour allopatric speciation (Wu & Wu, 1996; Qian &
Ricklefs, 2000; Baselga et al., 2012). High mountains and deep
valleys in south-west China have become natural barriers pre-
venting species from spreading between mountains within the
region (Li et al., 1999). As a result, each mountain range has a
large number of endemic species, and many species in the
region are shared only by few mountains (H.Q., unpublished
data). This has led to high b-diversity in the region, which in
Absolute latitude (°)
0 10 20 30 40 50
Sta
ndar
dize
d β
devi
atio
n
0
5
10
15
20
Absolute latitude (°)
0 10 20 30 40 50
0
5
10
15
20
Absolute latitude (°)
20 30 40 50 60
0
5
10
15
20rorig = -0.392, P < 0.05radj = -0.330, P < 0.05
rorig = 0.232, P > 0.05radj = 0.340, P < 0.05
rorig = -0.500, P < 0.05radj = -0.335, P < 0.05
d) e) f)
0 10 20 30 40 50
Raw
β d
evia
tion
-0.05
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0 10 20 30 40 50-0.05
0.00
0.05
0.10
0.15
0.20
0.25
0.30
20 30 40 50 60-0.05
0.00
0.05
0.10
0.15
0.20
0.25
0.30
a) b) c)rorig = 0.303, P < 0.05radj = 0.343, P < 0.05
rorig = 0.613, P < 0.05radj = 0.639, P < 0.05
rorig = 0.101, P > 0.05radj = 0.271, P < 0.05
Figure 3 The relations of b deviation of woody plants with latitude in (a, d) New World north, (b, e) New World south, and (c, f) China.Filled circles, thicker lines and rorig are for the relationship between b deviation and original latitude; open circles, thinner lines and radj forthe relationship between b deviation and adjusted latitude.
H. Qian et al.
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd6
turn has led to high g-diversity in south-west China, a region
which includes two of the world’s biodiversity hotspots (Myers
et al., 2000).
Standardized b deviation resulting from Kraft et al.’s null
model primarily represents the fraction of b-diversity that is not
accounted for by the covariation between b-diversity and
g-diversity. The covariation is treated as a result of the effect of
g-diversity on b-diversity by the null model. However, one could
interpret the result yielded from Kraft et al.’s approach in the
opposite way. Given that the variation of standardized b devia-
tion along a latitudinal gradient is small or remains constant,
this can be interpreted as Kraft et al. do (i.e. b-diversity being
constant across latitudes once the effect of g-diversity is
removed) or as g-diversity being constant across latitudes once
the effect of b-diversity is removed. The latter interpretation
would imply that the latitudinal richness gradient is solely
explained by a gradient in b-diversity (Andrés Baselga, personal
communication).
Furthermore, positive correlations between b- and g-diversity
may to some extent reflect the effects of some regional and
environmental factors on b- and g-diversity gradients in the
same direction, rather than the interaction between b- and
g-diversity. In this scenario, mechanisms of community assem-
bly beyond the interaction between b- and g-diversity may cause
the relationship between b- and g-diversity. Thus, even in the
case that there is no latitudinal gradient in b-diversity after
statistically accounting for g-diversity, it is premature to
preclude the role of mechanisms of community assembly in
driving b-diversity gradients.
Individual-based null model
A null model can only be a useful statistical tool if it is used
properly. As Gotelli (2001, 337) points out, ‘the null model strat-
egy is to construct a model that deliberately excludes a mecha-
nism being tested’. In other words, a null model should not
incorporate mechanisms that generate the pattern under
investigation. Kraft et al. (2011) developed an individual-based
null model approach to examine the relationship between
b-diversity and latitude after accounting for species pool
(g-diversity) (see Materials and Methods for details about the
null model). This null model approach has been applied in
several studies (e.g. Stegen et al., 2012).
The number of individuals per species generally decreases
with increasing g-diversity, a gradient that is mirrored by the
b-diversity gradient with g-diversity. The b-diversity is asymp-
totically related to g-diversity and tends to become independent
of g-diversity above a certain number of species (Fig. 5). With
Kraft et al.’s null model approach, in each randomization, the
number of individuals in each subplot of the randomized data is
the same as in the subplot of the raw data, as noted above. In
other words, patterns of species abundance distributions among
subplots in the raw data tend to be preserved in the null model.
Because the number of species tends to increase when more
Absolute latitude (°)
0 10 20 30 40 50 60 70
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
Absolute latitude (°)
0 10 20 30 40 50 60
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
Absolute latitude (°)
0 10 20 30 40 50
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
Absolute latitude (°)
20 30 40 50 60
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
a) b)
c) d)
rorig = 0.691; radj = 0.680 rorig = 0.681; radj = 0.678
rorig = 0.694; radj = 0.668 rorig = 0.595; radj = 0.576
Figure 4 Latitudinal gradients of thenumber of individuals per species ofwoody plants at (a) a global extent(Gentry’s 197 plots), (b) New Worldnorth, (c) New World south, and (d)China. Filled circles, thicker lines and rorig
are based on the original latitude; opencircles, thinner lines and radj are basedon adjusted latitude. All correlationcoefficients are significant (P < 0.05).
Latitudinal gradients of b-diversity
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd 7

individuals are included in a sampling unit (Gotelli & Graves,
1996), as predicted by probability theory (Sokal & Rohlf, 1981)
and observed in many empirical data sets, including our data at
the subplot scale (r = 0.740 for Gentry’s global data set, n = 1970;
r = 0.632 for China’s data set, n = 600; P < 0.05 in both cases) and
at the plot scale (Fig. 5), as a result the expected number of
species in each subplot is strongly forced to approach the
observed number of species in the subplot. As shown clearly in
Fig. 6, the expected and observed a-diversities not only are
strongly and positively correlated (r = 0.962, P < 0.05, n = 1970)
but also tend to approach to the 1 : 1 line where the expected
and observed a-diversities are identical. Because the total
number of species in each plot is set to be equal to that in the raw
data in each randomization, because expected b-diversity is
solely determined by the ratio of the average of expected
a-diversity (i.e. the average of expected numbers of species) in
the subplots of a plot and the total number of species in that
plot, and because the null model is formulated such that
expected a-diversity of each plot approaches the observed
a-diversity of the plot, preserving species abundance distribu-
tions in the null model would naturally lead to maintaining the
mechanisms that generate b-diversity gradients in the null
model. Consequently, values of expected b-diversity would be
strongly correlated with values of observed b-diversity (e.g. r =0.966 for Gentry’s global data set, r = 0.942 for China’s data set,
P < 0.05 in both cases); expected b-diversity resulting from the
null model has actually included b-diversity that results from
mechanisms of community assembly. Clearly, Kraft et al.’s
null model has incorporated the mechanisms that generate
b-diversity patterns and hence is conceptually and methodo-
logically incorrect because the model violates the fundamental
assumption of a null model, i.e. a null model should not incor-
porate mechanisms that generate the pattern under investiga-
tion (Gotelli, 2001).
γ-diversity
0 50 100 150 200 250 300
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
γ-diversity
0 50 100 150 200 250 300
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
γ-diversity
0 50 100 150 200 250 300
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
γ-diversity
0 20 40 60 80
Num
ber
of in
divi
dual
s pe
r sp
ecie
s
1
10
100
β-diversityIndivid. no. per sp.
Obs
erve
d β -
dive
rsity
Obs
erve
d β -
dive
rsity
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Obs
erve
d β-
dive
rsity
Obs
erve
d β-
dive
rsity
a) b)
c) d)
Figure 5 The relations of g-diversity(species pool) with observed b-diversity(filled circles) and the number ofindividuals of each species (log scale) ofwoody plants at (a) a global extent(Gentry’s global data set), (b) New Worldnorth, (c) New World south, and (d)China.
Observed α-diversity
0 25 50 75
Exp
ecte
d α-
dive
rsity
0
25
50
75
r = 0.962, P < 0.05
Figure 6 Relationship between observed and expecteda-diversity (number of species in each 0.01-ha subplot) in the1970 subplots of the 197 plots of Gentry’s global data set.Expected a-diversity was generated based on Kraft et al.’s (2011)null model with 1000 randomizations.
H. Qian et al.
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd8

The number of individuals of each of multiple species in a site
or community produces a species abundance distribution
(Williamson & Gaston, 2005). Species abundance distributions
largely result from deterministic, rather than stochastic, proc-
esses, and have played an important role in the development of
community ecology for over a century (Raunkiaer, 1909; Alonso
et al., 2008) and have been at the heart of the development of
ecological theories, e.g. neutral theory in ecology (Hubbell,
2001) and the more-individuals hypothesis (Hurlbert, 2004;
Evans et al., 2005). It is well known that patterns of species
abundance distributions in biological communities result, at
least in part, from mechanisms of community assembly
(Andrewartha & Birch, 1954; Whittaker, 1965; James &
Rathbun, 1981; Ugland & Gray, 1982; Kolasa & Strayer, 1988;
Tokeshi, 1999; McGill et al., 2007). Species abundance distribu-
tions drive the species–area relationship (Plotkin et al., 2000; He
& Legendre, 2002), and the slope (z-value) of the species–area
relationship has been used as a measure of b-diversity (Cody,
1975; Rosenzweig, 1995; Ricotta et al., 2002). Thus, mechanisms
driving species abundance distributions are closely related to, if
not the same as, the mechanisms driving b-diversity in a local
community. A null model that depends on species abundance
distributions naturally incorporates mechanisms driving
b-diversity.
As discussed above, a large part of the relationship between
b-diversity and latitude is removed from the standardized bdeviation–latitude gradient by the null model. Thus, b-diversity
represented by standardized b deviation may be considered as
the fraction of the b-diversity driven by mechanisms of com-
munity assembly or other processes that is beyond the fraction
of the b-diversity driven by species abundance distributions.
Mechanisms driving b-diversity beyond those driving species
abundance distributions may include, but are not limited to,
niche-based processes (Tokeshi, 1990), habitat filtering (Sven-
ning, 1999) and the priority effect (Young et al., 2001). For forest
communities at a local scale such as 0.1 ha, heterogeneity of
microhabitats, dynamics of forest (light) gaps and biotic inter-
actions within a study site may also be among major factors
influencing b-diversity in local communities. For example, the
behaviour of vertebrate fruit eaters often results in contagious
seed dispersal, in which seeds are deposited in single-species or
multispecies aggregations at sleeping trees or repeatedly used
sites (Schupp et al., 2002). In the case of tree b-diversity at a local
scale, the factors that determine distributions of seeds and seed-
lings around the parent trees (i.e. conspecific tree aggregation)
may also be important in generating patterns of b-diversity.
Clumping of seedlings around parent trees at a scale of metres to
hectares may be expected for species with either limited disper-
sal or specific regeneration requirements (Lambers et al., 2002).
Alternatively, density- and distance-dependent seed and seed-
ling mortality caused by species-specific predators may reduce
conspecific recruitment near the parent trees, as predicted by the
Janzen–Connell hypothesis (Janzen, 1970; Connell, 1971). These
factors may or may not drive b-diversity in the same direction as
that of species abundance distributions. Thus, whether there is a
latitudinal gradient in standardized b deviation resulting from
Kraft et al.’s null model would primarily depend on how well
species abundance distributions can account for the variation in
b-diversity, rather than whether there is a latitudinal gradient of
b-diversity after accounting for species pool.
Another problem with Kraft et al.’s null model is associated
with the method of standardizing raw b deviation in the null
model. The goal of the null model is to determine how much
b-diversity is left after removing the b-diversity that can be
generated by an individual-based randomization approach with
species pool accounted for (i.e. the expected b-diversity) from
observed b-diversity. However, as noted in Materials and
Methods, Kraft et al.’s null model divided raw b deviation (i.e.
the difference between observed and expected b-diversity) by
the standard deviation of the distribution of values of expected
b-diversity (Kraft et al., 2011). Because observed and expected
b-diversity are both in the same unit and both range between 0
and 1, and because the standard deviation of the distribution of
expected b-diversity is strongly correlated with both g-diversity
and latitude (e.g. r = -0.823 for g-diversity and 0.812 for latitude
in the data set with Gentry’s 197 plots; Fig. S3), standardizing
raw b deviation in such a way would introduce a bias in stand-
ardized b deviation (Andrés Baselga, personal communication).
For example, for the data set with Gentry’s 197 plots, latitude
explained 11.6 and 18.6% of the variation in raw b deviation,
respectively, in linear and quadratic regressions (Fig. 1b);
however, when raw b deviation was divided by the standard
deviation of the null distribution of b-diversity value, latitude
explained only 3.1 and 11.0% of the variation in the resulting
values (i.e. standardized b deviation), respectively, in linear and
quadratic regressions (Fig. 1c). Clearly, the standardizing
approach implemented in the null model changes the relation-
ship between b deviation and latitude not only in strength but
also in direction (i.e. changing a positive relationship to a nega-
tive relationship) in most cases (compare Fig. 1b with Fig. 1c
and Fig. 3a–c with Fig. 3d–f).
CONCLUSIONS
We conducted four sets of analyses to examine the relationship
between b-diversity and latitude using Kraft et al.’s (2011) null
model, and we showed that standardized b deviation is signifi-
cantly correlated with latitude in all four sets of analyses. We
demonstrated that Kraft et al.’s null model is not a valid null
model for the study of b-diversity gradient driven by mecha-
nisms of community assembly because the null model incorpo-
rates the mechanisms of community assembly that generate
b-diversity gradients and thus violates a fundamental assump-
tion of the null model. Specifically, expected b-diversity with
Kraft et al.’s null model should not include any part of
b-diversity that is driven by mechanisms of community assem-
bly, but expected b-diversity resulting from the null model in
fact includes the b-diversity that results from species abundance
distributions. Because the majority of the latitude–b-diversity
gradient can be explained by the latitude–species abundance
distribution (represented by the number of individuals per
species) gradient, and because species abundance distributions
Latitudinal gradients of b-diversity
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd 9

result from mechanisms of community assembly (Andrewartha
& Birch, 1954; Whittaker, 1965; James & Rathbun, 1981; Ugland
& Gray, 1982; Kolasa & Strayer, 1988; Tokeshi, 1999; McGill
et al., 2007), we conclude that the latitude–b-diversity gradient
at a local scale is largely driven by mechanisms of community
assembly. Our conclusion is in contrast to that of Kraft et al.
(2011).
ACKNOWLEDGEMENTS
We are grateful to Andrés Baselga and anonymous referees for
helpful comments, Nathan Kraft for the data set of Gentry’s 197
plots that was used in Kraft et al. (2011) and b deviation values,
Alwyn H. Gentry and his co-workers for their plot data, and the
financial support of the National Basic Research Program
(2009CB421105) to Z.O.
REFERENCES
Ahrens, C.D. (2007) Meteorology today: an introduction to
weather, climate, and the environment, 8th edn. Brooks Cole,
Belmont, CA.
Allen, A.P., Gillooly, J.F. & Brown, J.H. (2002) Global biodiver-
sity, biochemical kinetics, and the energetic-equivalence rule.
Science, 297, 1545–1548.
Alonso, D., Ostling, A. & Etienne, R.S. (2008) The implicit
assumption of symmetry and the species abundance distribu-
tion. Ecology Letters, 11, 93–105.
Andrewartha, H.G. & Birch, L.C. (1954) The distribution and
abundance of animals. University of Chicago Press, Chicago,
IL.
Baselga, A., Gómez-Rodríguez, C. & Lobo, J.M. (2012) Histori-
cal legacies in world amphibian diversity revealed by the
turnover and nestedness components of beta diversity. PLoS
ONE, 7, e32341.
Buckley, L.B. & Jetz, W. (2008) Linking global turnover of
species and environments. Proceedings of the National
Academy of Sciences USA, 105, 17836–17841.
Chen, S., Ouyang, Z., Zheng, H., Xiao, Y. & Xu, W. (2011) Lati-
tudinal gradient in beta diversity of forest communities in
America. Acta Ecologica Sinica, 31, 1334–1340.
Cody, M.L. (1975) Toward a theory of continental species diver-
sities: bird distributions over Mediterranean habitat gradi-
ents. Ecology and evolution of communities (ed. by M.L. Cody
and J.M. Diamond), pp. 214–257. Harvard University Press,
Cambridge, MA.
Connell, J.H. (1971) On the role of natural enemies in prevent-
ing competitive exclusion in some marine animals and in rain
forest trees. Dynamics of populations (ed. by P.J. Den Bour and
P.R. Gradwell), pp. 298–312. Center for Agricultural Publish-
ing and Documentation, Wageningen.
Darwin, C. (1859) The origin of species. Reprinted by Penguin
Books, London.
Evans, K.L., Warren, P.H. & Gaston, K.J. (2005) Species–energy
relationships at the macroecological scale: a review of the
mechanism. Biological Reviews, 80, 1–25.
Gaston, K.J. (2000) Global patterns in biodiversity. Nature, 405,
220–227.
Gotelli, N.J. (2001) Research frontiers in null model analysis.
Global Ecology and Biogeography, 10, 337–343.
Gotelli, N.J. & Graves, G.R. (1996) Null models in ecology. Smith-
sonian Institution, Washington, DC.
Harrison, S., Vellend, M. & Damschen, E.I. (2011) ‘Structured’
beta diversity increases with climatic productivity in a classic
dataset. Ecosphere, 2, art11.
He, F. & Legendre, P. (2002) Species-diversity patterns derived
from species-area models. Ecology, 83, 1185–1198.
Hubbell, S.P. (2001) The unified neutral theory of biodiversity and
biogeography. Princeton University Press, Princeton, NJ.
Hurlbert, A.H. (2004) Species–energy relationships and habitat
complexity in bird communities. Ecology Letters, 7, 714–720.
James, C. & Rathbun, S. (1981) Rarefaction, relative abundance,
and diversity of avian communities. Auk, 98, 785–800.
Jankowski, J.E., Ciecka, A.L., Meyer, N.Y. & Rabenold, K.N.
(2009) Beta diversity along environmental gradients: implica-
tions of habitat specialization in tropical montane landscapes.
Journal of Animal Ecology, 78, 315–327.
Janzen, D.H. (1970) Herbivores and the number of tree species
in tropical forests. The American Naturalist, 104, 501–528.
Jump, A.S., Mátyás, C. & Peñuelas, J. (2009) The altitude-for-
latitude disparity in the range retractions of woody species.
Trends in Ecology and Evolution, 24, 694–701.
Kolasa, J. & Strayer, D. (1988) Patterns of the abundance of
species: a comparison of two hierarchical models. Oikos, 53,
235–241.
Koleff, P., Lennon, J.J. & Gaston, K.J. (2003) Are there latitudinal
gradients in species turnover? Global Ecology and Biogeogra-
phy, 12, 483–498.
Kraft, N.J.B., Comita, L.S., Chase, J.M., Sanders, N.J., Swenson,
N.G., Crist, T.O., Stegen, J.C., Vellend, M., Boyle, B., Anderson,
M.J., Cornell, H.V., Davies, K.F., Freestone, A.L., Inouye, B.D.,
Harrison, S.P. & Myers, J.A. (2011) Disentangling the drivers
of b diversity along latitudinal and elevational gradients.
Science, 333, 1755–1758.
Lambers, J.H.R., Clark, J.S. & Beckage, B. (2002) Density-
dependent mortality and the latitudinal gradient in species
diversity. Nature, 417, 732–735.
Lenoir, J., Gégout, J.-C., Guisan, A., Vittoz, P., Wohlgemuth, T.,
Zimmermann, N.E., Dullinger, S., Pauli, H., Willner, W.,
Grytnes, J.-A., Virtanen, R. & Svenning, J.-C. (2010) Cross-
scale analysis of the region effect on vascular plant species
diversity in southern and northern European mountain
ranges. PLoS ONE, 5, e15734.
Li, H., He, D.-M., Bartholomew, B. & Long, C.-L. (1999)
Re-examination of the biological effect of plate movement:
impact of Shan-Malay Plate displacement (the movement of
Burma–Malaya Geoblock) on the biota of Gaoligong Moun-
tains. Acta Botanica Yunnanica, 21, 407–425.
McGill, B.J., Etienne, R.S., Gray, J.S., Alonso, D., Anderson, M.J.,
Benecha, H.K., Dornelas, M., Enquist, B.J., Green, J.L., He, F.,
Hurlbert, A.H., Magurran, A.E., Marquet, P.A., Maurer, B.A.,
Ostling, A., Soykan, C.U., Ugland, K.I. & White, E.P. (2007)
H. Qian et al.
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd10

Species abundance distributions: moving beyond single pre-
diction theories to integration within an ecological frame-
work. Ecology Letters, 10, 995–1015.
Meynard, C.N., Devictor, V., Mouillot, D., Thuiller, W., Jiguet, F.
& Mouquet, N. (2011) Beyond taxonomic diversity patterns:
how do a, b and g components of bird functional and phylo-
genetic diversity respond to environmental gradients across
France? Global Ecology and Biogeography, 20, 893–903.
Mittelbach, G.G., Schemske, D.W., Cornell, H.V. et al. (2007)
Evolution and the latitudinal diversity gradient: speciation,
extinction and biogeography. Ecology Letters, 10, 315–331.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca,
G.A.B. & Kent, J. (2000) Biodiversity hotspots for conserva-
tion priorities. Nature, 403, 853–858.
New, M., Hulme, M. & Jones, P. (1999) Representing twentieth-
century space-time climate variability. Part I: development of
a 1961–90 mean monthly terrestrial climatology. Journal of
Climate, 12, 829–856.
Phillips, O. & Miller, J.S. (2002) Global patterns of plant diversity:
Alwyn H. Gentry’s forest transect data set. Missouri Botanical
Garden Press, St Louis.
Pianka, E.R. (1966) Latitudinal gradients in species diversity:
a review of concepts. The American Naturalist, 100, 33–
46.
Pither, J. & Aarssen, L.W. (2005) The evolutionary species pool
hypothesis and patterns of freshwater diatom diversity along a
pH gradient. Journal of Biogeography, 32, 503–513.
Plotkin, J.B., Potts, M.D., Yu, D.W., Bunyavejchewin, S., Condit,
R., Foster, R., Hubbell, S., LaFrankie, J., Manokaran, N., Lee,
H.-S., Sukumar, R., Nowak, M.A. & Ashton, P.S. (2000) Pre-
dicting species diversity in tropical forests. Proceedings of the
National Academy of Sciences USA, 97, 10850–10854.
Qian, H. (2009) Beta diversity in relation to dispersal ability for
vascular plants in North America. Global Ecology and Bioge-
ography, 18, 327–332.
Qian, H. & Ricklefs, R.E. (2000) Large-scale processes and the
Asian bias in species diversity of temperate plants. Nature,
407, 180–182.
Qian, H. & Ricklefs, R.E. (2007) A latitudinal gradient in large-
scale beta diversity for vascular plants in North America.
Ecology Letters, 10, 737–744.
Qian, H. & Ricklefs, R.E. (2012) Disentangling the effects of
geographic distance and environmental dissimilarity on
global patterns of species turnover. Global Ecology and Bioge-
ography, 21, 341–351.
Qian, H. & Xiao, M. (2012) Global patterns of the beta
diversity–energy relationship in terrestrial vertebrates. Acta
Oecologica, 39, 67–71.
Qian, H., Ricklefs, R.E. & White, P.S. (2005) Beta diversity of
angiosperms in temperate floras of eastern Asia and eastern
North America. Ecology Letters, 8, 15–22.
Qian, H., Badgley, C. & Fox, D.L. (2009) The latitudinal gradient
of beta diversity in relation to climate and topography for
mammals in North America. Global Ecology and Biogeography,
18, 111–122.
Qian, H., Wang, X. & Zhang, Y. (2012) Comment on ‘disentan-
gling the drivers of b diversity along latitudinal and eleva-
tional gradients’. Science, 335, 1573.
Raunkiaer, C. (1909) Formationsuntersögelseog formationssta-
tistik. Botanisk Tidsskrift, 30, 20–132.
Ricklefs, R.E. (1987) Community diversity: relative roles of local
and regional processes. Science, 235, 167–171.
Ricotta, C., Carranza, M.L. & Avena, G. (2002) Computing
b-diversity from species–area curves. Basic and Applied
Ecology, 3, 15–18.
Rodríguez, P. & Arita, H.T. (2004) Beta diversity and latitude in
North American mammals: testing the hypothesis of covaria-
tion. Ecography, 27, 547–556.
Rohde, K. (1992) Latitudinal gradients in species diversity: the
search for the primary cause. Oikos, 65, 514–527.
Rosenzweig, M.L. (1995) Species diversity in space and time.
Cambridge University Press, Cambridge.
Schupp, E.W., Milleron, T. & Russo, S.E. (2002) Dissemination
limitation and the origin and maintenance of species-rich
tropical forests. Seed dispersal and frugivory: ecology, evolution,
and conservation (ed. by D.J. Levey, W.R. Silva and M. Galetti),
pp. 19–33. CAB International, Wallingford, UK.
Sengör, A.M.G. & Natal’in, B.A. (1996) Paleotectonics of Asia:
fragments of a synthesis. The tectonic evolution of Asia (ed. by A.
Yin and T.M. Harrison), pp. 486–640. Cambridge University
Press, Cambridge, UK.
Sokal, R.R. & Rohlf, F.J. (1981) Biometry, 2nd edn. W. H.
Freeman, New York.
Stegen, J.C., Freestone, A.L., Crist, T.O., Anderson, M.J., Chase,
J.M., Comita, L.S., Cornell, H.V., Davies, K.F., Harrison, S.P.,
Hurlbert, A.H., Inouye, B.D., Kraft, N.J.B., Myers, J.A.,
Sanders, N.J., Swenson, N.G. & Vellend, M. (2012) Stochastic
and deterministic drivers of spatial and temporal turnover in
breeding bird communities. Global Ecology and Biogeography,
doi:10.1111/j.1466-8238.2012.00780.x.
Stephenson, N.L. & Das, A.J. (2011) Comment on ‘changes in
climatic water balance drive downhill shifts in plant species’
optimum elevations’. Science, 334, 177.
Stevens, G.C. (1989) The latitudinal gradient in geographical
range: how so many species coexist in the tropics. The Ameri-
can Naturalist, 133, 240–256.
Svenning, J.-C. (1999) Microhabitat specialization in a species-
rich palm community in Amazonian Ecuador. Journal of
Ecology, 87, 55–65.
Tokeshi, M. (1990) Niche apportionment or random assortment
– species abundance patterns revisited. Journal of Animal
Ecology, 59, 1129–1146.
Tokeshi, M. (1999) Species coexistence: ecological and evolution-
ary perspectives. Blackwell Scientific, Oxford.
Ugland, K.I. & Gray, J.S. (1982) Lognormal distributions and
the concept of community equilibrium. Oikos, 39, 171–178.
Wallace, A.R. (1878) Tropical nature and other essays. Macmillan,
New York.
Whittaker, R.H. (1960) Vegetation of the Siskiyou Mountains,
Oregon and California. Ecological Monographs, 30, 279–338.
Latitudinal gradients of b-diversity
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd 11

Whittaker, R.H. (1965) Dominance and diversity in land plant
communities. Science, 147, 250–260.
Whittaker, R.H. (1972) Evolution and measurement of species
diversity. Taxon, 21, 213–251.
Williamson, M. & Gaston, K.J. (2005) The lognormal distribu-
tion is not an appropriate null hypothesis for the species-
abundance distribution. Journal of Animal Ecology, 74, 409–
422.
Wu, C.-Y. & Wu, S.-G. (1996) A proposal for a new floristic
kingdom (realm). Floristic characteristics and diversity of east
Asian plants (ed. by A.-L. Zhang and S.-G. Wu), pp. 3–42.
China Higher Education Press, Beijing.
Young, T.P., Chase, J. & Huddleston, R. (2001) Community suc-
cession and assembly: comparing, contrasting and combining
paradigms in the context of ecological restoration. Ecological
Restoration, 19, 5–18.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online
version of this article:
Figure S1 Distribution of Gentry’s 197 plots used in Kraft et al.
(2011).
Figure S2 Distribution of 15 nature reserves in China, where the
60 forest plots used in the present study were sampled.
Figure S3 Relationships of the standard deviation of the distri-
bution of expected b-diversity with g-diversity and latitude for
Gentry’s 197 plots used in the present study.
BIOSKETCH
Hong Qian’s research is multidisciplinary and
particularly lies at the interface of ecology and
biogeography. His research involves a wide range of
spatial scales (from local to global) and a variety of
taxa (e.g. bryophytes, vascular plants, vertebrates and
invertebrates). In particular, he is interested in
understanding the relative roles of historical and
present-day factors in determining the patterns in
biodiversity.
Editor: Andres Baselga
H. Qian et al.
Global Ecology and Biogeography, ••, ••–••, © 2012 Blackwell Publishing Ltd12




















