
Mitochondrial respiration in B lymphocytes is essential for ...
Upload
independentCategory
view
1download
0
REGULAR ARTICLE
Drivers of increased soil respiration in a poplar coppiceexposed to elevated CO2
Alessandra Lagomarsino & Martin Lukac &
Douglas L. Godbold & Sara Marinari &Paolo De Angelis
Received: 27 July 2011 /Accepted: 13 April 2012 /Published online: 8 May 2012# Springer Science+Business Media B.V. 2012
AbstractBackground and aims The response of soil respiration(SR) to elevated CO2 is driven by a number of pro-cesses and feedbacks. This work aims to i) detect theeffect of elevated CO2 on soil respiration during thesecond rotation of a short rotation forest, at two levelsof N availability; and ii) identify the main driversbehind any changes in soil respiration.Methods A poplar plantation (POP-EUROFACE) wasgrown for two rotations of 3 years under elevated CO2
maintained by a FACE (Free Air CO2 Enrichment)technique. Root biomass, litter production and soilrespiration were followed for two consecutive yearsafter coppice.Results In the plantation, the stimulation of fine rootand litter production under elevated CO2 observed at thebeginning of the rotation declined over time. Soil respi-ration (SR) was continuously stimulated by elevatedCO2, with a much larger enhancement during the grow-ing (up to 111 %) than in the dormant season (40 %).The SR increase at first appeared to be due to theincrease in fine root biomass, but at the end of the 2ndrotation was supported by litter decomposition and theavailability of labile C. Soil respiration increase underelevated CO2 was not affected by N availability.Conclusions The stimulation of SR by elevated CO2
was sustained by the decomposition of above and be-lowground litter and by the greater availability of easilydecomposable substrates into the soil. In the final year aselevated CO2 did not increase C allocation to roots, thehigher SR suggests greater C losses from the soil, thusreducing the potential for C accumulation.
Keywords SRF. FACE . Soil respiration . LabileC . Root biomass . Litter
Introduction
In European forests, about 55 % of photosyntheticallyfixed C finds its way back into the atmosphere via
Plant Soil (2013) 362:93–106DOI 10.1007/s11104-012-1261-0
Responsible Editor: Klaus Butterbach-Bahl.
A. Lagomarsino (*)Agriculture Research Council, Research Centre forAgrobiology and Pedology (CRA-ABP),Piazza M. D’Azeglio 30,50121 Firenze, Italye-mail: [email protected]
M. LukacSchool of Agriculture, Policy and Development,University of Reading,Reading RG6 6AR, UK
D. L. GodboldInstitute of Forest Ecology, Universität für Bodenkultur,Peter Jordan Str 82,1190 Vienna, Austria
S. Marinari : P. De AngelisDIBAF (Department for Innovation in Biological,Agro-Food and Forest Systems), University of Tuscia,Via San Camillo de Lellis,01100 Viterbo, Italy

belowground respiration (Janssens et al. 2001). Due tothe magnitude of this flux and the large stock of Cpresent in soils, any change in soil C emissions inresponse to environmental changes could constitute asignificant feedback on CO2 concentration in the at-mosphere. Moreover, changes in photosynthate allo-cation under elevated CO2 are likely to alter rootbiomass and substrate availability for microbial me-tabolism, potentially affecting soil CO2 efflux (Zak etal. 2000; Hogberg et al. 2001; van Hees et al. 2005).
Soil CO2 efflux is the result of two distinct processes: i)breakdown of root-derived C (root and rhizosphere res-piration) and ii) decomposition of soil-derived C (hetero-trophic respiration of SOM) (Ryan and Law 2005). Thus,soil respiration (SR) results from activity of a multi-organism network of oxidation pathways, where auto-and heterotrophic components may respond to elevatedCO2 in contrasting ways (Trueman and Gonzalez-Meler2005). In terrestrial ecosystems, about 35–80 % of Cfixed through photosynthesis is transferred belowgroundto fuel root activity, mycorrhizal networks and root exu-dates (Raich and Nadelhoffer 1989; Davidson et al. 2002;Giardina et al. 2003; Ryan et al. 2004). Litter and rootproductivity, as well as photosynthetic activity are thusthe main factors controlling belowground C allocation,and therefore the CO2 efflux from soils (Hogberg et al.2001; Kuzyakov and Cheng 2001).
Short rotation forestry practice (SRF), a land-use typelikely to expand in the near future due to increasingdemand for bioenergy, is typical for frequent removal ofaboveground biomass to maximize yield (Sennerby-Forsse 1995). However, suchmanagement induces a verystrong disturbance to the plant-soil system, likely to affectsoil C cycling. The POP-EUROFACE experiment wasestablished in 1999, combining a fast-growing plantationwith a Free Air Carbon dioxide Enrichment (FACE)treatment to test for the interaction between elevatedCO2 and poplar SRF productivity (Scarascia Mugnozzaet al. 2000). After the first 3 year rotation, when poplartrees were grown from hardwood cuttings, the plantationwas coppiced and aboveground woody biomass washarvested. Post-coppice, the trees were allowed to re-grow and a fertilization treatment was added, giving usthe opportunity to assess the impact of elevated CO2 atdifferent levels of N availability. Significant responsesemerged during the first rotation, demonstrating that ele-vated CO2: i) increased fine root biomass and productiv-ity (Lukac et al. 2003); ii) did not change litterproduction, but decreased litter quality (Cotrufo et al.
2005); iii) increased the rate of SR (King et al. 2004),stimulating SOMdecomposition through a priming effectcaused by the new labile C inputs (Hoosbeek et al. 2004;Hoosbeek and Scarascia-Mugnozza 2009).
Since multiyear data from forest FACE experimentssuggest that plant size and productivity are the primarydeterminants of soil CO2 efflux (King et al. 2004), wecontend that the increase in SR under elevated CO2
observed in the first cycle may be sustained in the longrun by a combination of larger root biomass andproductivity (Lukac et al. 2003) and accumulation ofa higher quantity of easily decomposable substratessuch as labile C (Lagomarsino et al. 2006) or litterresidual mass (Cotrufo et al. 2005) over a number ofyears. However, soil C dynamics under the plantationcould be altered by coppicing and the interaction ofelevated CO2 with fertilization. Thus, we hypothesizedthat; i) the stimulating effects of elevated CO2 on SRwould be still evident at the end of the second rotationcycle, ii) SR response to elevated CO2would be affectedby increased N availability in fertilization treatment, andiii) root biomass and labile C availability are the maindrivers behind increased SR under elevated CO2.
Materials and methods
Site description
The POP-EuroFACE experimental plantation (CentralItaly, 42°22’N, 11°48’E, alt. 150 m), was established in1999 over an area of about 9 ha, previously used foragricultural cultivation. The climate is typically Mediter-ranean, with warm and dry summer and mild and humidwinter. The mean annual air temperature is 15 °C, andmean annual precipitation is 676 mm. The soil is aPachic Xerumbrepts (USDA classification), with a deepA horizon, and an O horizon of 2–3 cm, generallydistinct into a litter and a fermentation layer, whichformed after the POP-EuroFACE plantation was estab-lished (Hoosbeek et al. 2004). The layout of the exper-iment consisted of 6 plots of 300 m2 each, divided into 3sectors planted with a different poplar species (Populus xeuramericana Dode (Guinier), genotype I-214, Populusnigra L. (Jean Pourtet) and Populus alba L., genotype2AS11) at a planting density of 10,000 trees ha−1 (spac-ing 1×1 m). Three of the six experimental plots weremaintained at ambient [CO2] (~370 ppm) and three weremaintained at elevated [CO2] (~550 ppm) by the FACE
94 Plant Soil (2013) 362:93–106

technique (Free-Air CO2 Enrichment, Miglietta et al.2001). The management of the plantation consisted of3 years of growth from hardwood cuttings (first rotation),followed by a mid-winter coppice (2001–2002) andfurther 3 years of growth from established root systems(second rotation). A fertilization treatment was started atthe beginning of the second rotation in 2002, applied viathe drip irrigation system to one half of each experimen-tal plot (Liberloo et al. 2006). The total input of N wascalculated on the basis of plant uptake during the firstrotation, with the objective to remove any possible ni-trogen limitation of plant growth. Nitrogen was suppliedin constant weekly amounts with a 4:1 NH4
+:NO3− ratio
in 2002 and 1:1 NH4+:NO3
− ratio in 2003 and in 2004 inproportion to the growth rate. The annual amounts ofnitrogen supplied were 212 kg ha−1 in 2002, 290 kg ha−1
in 2003 and 2004. The whole plantation was drip irri-gated at 6 to 10 mm of water per day during the growingseason. The amount of water was sufficient to avoidwater stress even during the hottest days of a growingseason. Soil water content was measured weekly andwas not limiting throughout the experiment (De Angelis,personal communication).
Root biomass
Standing root biomass was sampled using an 8 cmdiameter corer in June and September of 2003 and2004, and then in January 2005. Root samples weretaken separately from the following 2 soil depths: 0–20 cm and 20–40 cm. Three samples per species perplot were taken from the vicinity of randomly selectedtrees at 30 cm, 50 cm and 70 cm distance from thestem. Care was taken not to sample root system of onetree more than once. Roots were washed free of soil,divided into 2 diameter classes of fine (<2 mm) andcoarse (>2 mm) roots. All samples of roots were driedfor 5 days at 60 °C and weighed. For the purpose ofthis analysis, root biomass from the two depths wasadded to obtain total root biomass, while observationsfrom the three distances were averaged to obtain meanroot biomass per unit land area.
Leaf litter
Leaf litter production was monitored using leaf littertraps. Traps consisted of plastic baskets (0.13m2) placedon the ground in the projected area of a tree or in theintersection of projected areas of four adjacent trees.
Litter was collected with traps twice a month duringthe growing seasons of both years. Three litter traps foreach sector were used, 108 total (3 replicates × 3 plots ×2 CO2 level × 2 fertilization level × 3 species). Aftersampling, leaf litter was dried in an oven at 70 °C for48 h, and dry weight determined. Production data wereexpressed in terms of grams per squaremetre of soil, andannual litter production calculated as the sum of eachperiodic collection.
Soil respiration measurement
SR was measured biweekly during the years 2003–2004. Semi-permanent plastic collars were installedinto the soil surface (up to 2–3 cm depth) to engagethe soil respiration cuvette, ensuring repeated sam-pling of the same soil over time and reducing distur-bance. Measurements were performed in the middle offour stems, three replicates for each sector, 108 total (3replicates × 3 plots × 2 CO2 levels × 2 fertilizationlevels × 3 species). Measurements were made manu-ally with infrared gas analyzers (IRGA) operating inclosed-path mode. The environmental gas monitor(EGM-4, PP systems, UK) was equipped with theSRC-1 SR cuvette, including a metal mesh to preventexcessive ventilation, placed on top of soil collars.Each measurement was made to last double the timeit took for the rate of soil CO2 efflux to stabilize(usually within 2 min). A quadratic model was fittedto the measured data to obtain the rate of CO2 emis-sion. At the time of each measurement, soil tempera-ture was recorded using a manual digital thermometerat a depth of 3–5 cm.
In order to compare the soil CO2 efflux of differentyears at reference temperatures, in the absence ofwater limitation, the following first-order exponentialfunction was used:
SR ¼ a � ekT
where SR is the measured soil CO2 efflux, a is thevalue of SR at 0 °C (intercept), k is the rate constantrepresenting the SR sensitivity to the temperature in-crease and T is the soil temperature in °C.
SR of the dormant and growing season (SRdormant
and SRgrowing) for each year were calculated as meanof SR for the non vegetative (November-first half ofApril) and vegetative (second half of April–October)periods. SRmean was calculated as mean annual values
Plant Soil (2013) 362:93–106 95

of SR for each year. SR’dormant and SR’growing werecalculated for each year from the exponential functionconsidering a mean temperature for both years of 9and 20 °C, respectively for the dormant and growingperiods.
Statistics
Repeated ANOVA was performed to evaluate theeffects of elevated CO2, N fertilization, species andtheir interactions on fine and coarse root biomass, leaflitter and SR (annual, dormant and growing periodsreported as SRmean, SRdormant and SRgrowing) consider-ing the mean annual values of 2003 and 2004. Usingannual data, Fisher’s LSD Post hoc test was performedto evaluate the main effects of elevated CO2 in fertil-ized and unfertilized conditions in each year and toevaluate the significance of the variation between2003 and 2004 in control, FACE, fertilized (high N)and unfertilized (low N) soil. Multiple regression anal-yses (stepwise, forward) were performed separatelyfor the 2 years using the mean SR of the dormantand growing seasons (SRdormant and SRgrowing) as de-pendent variables and other tested parameters as inde-pendent variables. For the regression analyses datapublished in previous manuscripts were also used:Moscatelli et al. 2005 and Lagomarsino et al. 2006for water soluble C (WSC) and microbial biomass C(MBC); Lagomarsino et al. 2008 for inorganic N (assum of NH4
+ and NO3−) and microbial biomass N
(MBN), Lagomarsino et al. 2009 for heterotrophiccomponent of soil respiration. All statistical analyseswere performed with the Systat 12.0 statistical soft-ware package (SPSS Inc.).
Results
In general, the response of the three poplar species toelevated CO2 and fertilization was very similar, wetherefore report an average response across the threepoplar species. When the differences between poplarspecies were statistically significant (e.g. for litterfall),then the single species response is shown.
Root biomass
Fine root biomass showed a marked reduction in 2004with respect to the previous year (Fig. 1a, Table 1).
The decrease in fine root biomass was evident in alltreatments with the exception of the control high Ntreatment (Table 1). In 2003, elevated CO2 significant-ly increased fine root biomass only in high N plots,while in 2004 no significant effects of elevated CO2
were detected (Table 2). Coarse root biomass showeda similar downward trend, although less pronouncedand not statistically significant (Fig. 1b, Table 1). In2003, elevated CO2 increased coarse root biomass,more so in high N sub-plots, and in June and Septem-ber 2004 coarse root biomass was still greater underelevated CO2, although the difference was not statis-tically significant (Table 2).
Fine root biomass correlated significantly (R20
0.711, P<0.001) with water soluble C (data fromLagomarsino et al. 2006), when both years (2003and 2004) were taken into account together (Fig. 2).
Litter
Litter production differed significantly (P<0.001)among poplar species, the lowest values were foundin P. nigra and the highest in P. x euramericana bothin 2003 and 2004 (Fig. 3). Total litter productionshowed a significant decrease in 2004, regardless oftreatment or species (Table 1). In 2003, elevated CO2
induced a significant increase in litter production in allspecies at both N levels, but this effect was no longerpresent in 2004 (Table 2), as also shown by the sig-nificant interaction CO2 × year (Table 1).
Soil respiration
SR rates showed a marked intra and inter-annual varia-tion (Figs. 4 and 5). A seasonal pattern was observed inboth years, strongly driven by ambient temperature. Theexponential function showed on average for the 2 yearsand the treatments 0.3 μmol CO2 m
−2 s−1 as intercept at0 °C (a), and 0.125 as rate constant (k). Elevated CO2
increased the 0 °C intercept in 2003 (+82 % on average)and the slope in 2004 (+46 % on average). k showed asignificant CO2 × year interaction (p<0.001), with asignificant increase in FACE–low N, and a decrease inControl soil in 2004 vs. 2003.
In 2003, elevated CO2 induced a significant in-crease in SR which was similar in both seasons(+57 % on average, Table 2). In 2004, the increase inSR under elevated CO2 was of about 41 % in thedormant season, but much larger in the growing
96 Plant Soil (2013) 362:93–106

season (up to 104 %, Table 2). In fact, the increase inSR in the growing season with respect to the dormantseason was significantly higher under elevated CO2
than in control soils (3.7 vs. 2.5 times respectively,Fig. 5), as also shown by the significant interactionCO2 × year for SRgrowing (Table 1). The observedincrease in SR under elevated CO2 was independentof N availability and poplar species in both years(Table 1). In February and May 2004 (dormant andgrowing seasons), SR at the normalized temperature of18 °C (SR18) was significantly correlated (p<0.001)with heterotrophic respiration (for heterotrophic respi-ration data and details see Lagomarsino et al. 2009),but was more responsive in FACE than in control soil(Fig. 6).
Soil respiration and biotic drivers
From the multiple regression analysis, WSC showedthe strongest relationship with SRdormant and SRgrowing
in 2003 and with SRgrowing in 2004 (Table 3). In 2003,fine and coarse roots biomass was significantly corre-lated with SRgrowing, and litter with SRdormant. In 2004,
both SRdormant and SRgrowing were significantly corre-lated with litter and fine roots produced during theprevious year (Table 3). The conceptual model forthe two study years illustrates processes and poolsaffecting SR (Fig. 7). Fluxes of C from photosyntheticand litter production processes to belowground poolswere affected by elevated CO2 in 2003 (both) and2004 (photosynthesis only). In Fig. 7, the additionalC input from root turnover and litter accumulation onthe forest floor from 2003 to 2004 was drawn. Giventhe lack of elevated CO2 effect on root biomass in2004, C temporally stored in belowground pools wasthen emitted by both autotrophic and heterotrophiccomponents of SRgrowing, with a much larger elevatedCO2 effect in 2004.
Discussion
Impacts of elevated CO2 on biological properties
An increase in fine root biomass under elevated CO2 isa common response widely observed in terrestrial
0
50
100
150
200
250
300
350
fin
e ro
ot
bio
mas
s (g
m-2
) a
b
0
100
200
300
400
500
600
700
Jun-03 Sep-03 Dec-03 Mar-04 Jun-04 Sep-04 Dec-04
coar
se r
oo
t bio
mas
s (g
m-2
)
Control-low N FACE-low N Control-high N FACE-high N
Fig. 1 Fine root biomass(a) and coarse root biomass(b) at 0–40 cm depth, in theyears 2003 and 2004 incontrol (square) and FACE(triangle) low N (open sym-bol) and high N (filled sym-bol). Error bars representstandard errors
Plant Soil (2013) 362:93–106 97

Tab
le1
Meanannu
alvalues
offine
andcoarse
roots,litterprod
uctio
n,SRannu
al,ofthegrow
ingandthedo
rmantperiods
(SRmean,S
RgrowingandSRdorm
ant)of
the2yearsforplotsat
ambientC
O2(con
trol),elevated
CO2(FACE),no
tfertilized
(low
N)andfertilized(highN).For
each
parameterrepeated
ANOVAhasbeen
performed
consideringmeanannu
alvalues
ofthe2years,with
N,C
O2,species
asindepend
entfactors.F
oreach
treatm
ent,thepercentage
annu
alvariationin20
04vs.200
3isalso
repo
rted,w
iththesign
ificance
ofthedifference
(Fisher’sLSDPostH
octest).pvalues
arerepo
rted
foreach
factor
andforinteractions.Interactio
nsno
treportedareno
tsignificant.*p<0.05
,**p<0.01
,***p<0.00
1.Stand
arderroris
repo
rted
inparenthesis
Fineroot
Coarseroot
Litter
SRmean
SRdorm
ant
SRgrowing
gm
−2gm
−2gm
−2μmol
CO2m
−2s−
1μmol
CO2m
−2s−
1μmol
CO2m
−2s−
1
2003
2004
2003
2004
2003
2004
2003
2004
2003
2004
2003
2004
Control-low
N14
8(29)
69(12)
278(39)
288(19)
864(29)
573(29)
1.9(0.1)
1.6(0.1)
0.8(0.07)
0.7(0.06)
2.9(0.2)
2.0(0.2)
Annualvariation(%
)−5
3**
+4
n.s.
−33*
**−1
6n.s.
−9n.s.
−29*
**
Control-highN
73(8)
87(13)
148(24)
217(35)
885(32)
572(32)
1.7(0.1)
1.5(0.09)
0.8(0.07)
0.7(0.04)
2.5(0.2)
1.9(0.1)
Annualvariation(%
)+19
n.s.
+47
n.s.
−36*
**+5n
.s.
−5n.s.
−23*
FACE-low
N17
9(17)
88(13)
412(66)
356(31)
1026
(33)
577(35)
2.9(0.2)
3.0(0.2)
1.4(0.10)
1.0(0.08)
4.4(0.3)
4.2(0.3)
Annualvariation(%
)−5
1**
−14
n.s.
−44*
**−1
0n.s.
−24*
**−6
n.s.
FACE-highN
197(25)
77(12)
303(57)
281(50)
1036
(26)
624(28)
2.6(0.1)
2.9(0.2)
1.2(0.08)
1.1(0.09)
4.0(0.2)
3.9(0.3)
Annualvariation(%
)−6
1***
−7n.s.
−40*
**+12
n.s.
−7n.s.
−3n.s.
RepeatedANOVA
(pvalues)
Nitrogen
n.s.
0.005
n.s.
n.s.
n.s.
n.s.
CO2
0.03
0.00
20.000
0.000
0.000
0.000
Species
n.s.
n.s.
0.000
n.s.
n.s.
n.s.
Year
0.00
0n.s.
0.000
n.s.
0.005
0.000
Y×CO2
0.03
n.s.
0.000
0.006
0.09
0.03
Y×Sp.
n.s.
n.s.
0.046
n.s.
n.s.
n.s.
Y×N
×CO2
0.08
n.s.
n.s.
n.s.
n.s.
n.s.
98 Plant Soil (2013) 362:93–106

ecosystems (Norby et al. 2002; Norby 2004; Iversen etal. 2008), including the in POP-EUROFACE planta-tion in the first cycle (Lukac et al. 2003). However,Brown et al. (2007), Handa et al. (2008) and Bader etal. (2009) found unaltered or even reduced fine rootbiomass after few years of CO2 enrichment. At POP-EUROFACE plantation the positive response of rootbiomass to elevated CO2 declined after coppicing,especially towards the end of the second rotation. Fastgrowing plantations are highly dynamic systems,where large disturbance induced by coppicing andsubsequent re-growth probably affects belowgroundproductivity. Our results demonstrate significantchanges in root biomass, even after a single coppicingevent. Compared to values found at the end of the firstrotation cycle, which were on average 118 and 206 gm−2 for control and FACE soils respectively (Lukac etal. 2003), the decline of fine root biomass occurredfirst in control–high N, and only later in FACE soil.
Liberloo et al. (2009) found that coppicing shifted theinvestment of additional C fixed in elevated CO2 fromroot biomass to aboveground woody biomass pools,possibly in relation to the multistem phase of theplantation. We contend that smaller fine root networkswere sufficient to satisfy N demand in ambient CO2
and fertilized conditions, whereas larger fine root bio-mass was needed for longer under elevated CO2 tosupport faster growth (Liberloo et al. 2005). In parallelwith the root biomass decline in 2004, inorganic Navailability significantly increased, especially in un-fertilized soil. There is evidence to suggest that soilmicroorganisms benefited from the lack of competi-tion from roots, as N immobilized within microbialbiomass doubled during this year (Lagomarsino et al.2008). Changes in the patterns of fine root biomasspreceded similar changes in the patterns of labile C(Lagomarsino et al. 2006, 2009), providing furtherevidence of a strict linkage between fine roots and
Table 2 Percentage effects of elevated CO2 treatment (CO2) atlow and high N levels in the years 2003 and 2004, as meanannual values for fine and coarse root biomass, litter production,
SR annual, of the growing and the dormant periods (SRmean,SRgrowing and SRdormant). Fisher’s LSD Post Hoc test (Systat12.0). #p<0.1, * p<0.05, **p<0.01, ***p<0.001
Fine root Coarse root Litter SRmean SRdormant SRgrowing
2003
CO2–low N +20n.s. +48# +19** +53*** +58*** +54*
CO2–high N +169** +105* +17** +55*** +53** +62**
2004
CO2–low N +30 n.s. +24 n.s. +1 n.s. +91*** +32* +104***
CO2–high N −17 n.s. +29 n.s. +9 n.s. +95*** +50*** +104***
0
10
20
30
40
50
60
0 50 100 150 200 250 300 350
WS
C (µ
g C
g-1
)
fine root biomass (g m-2)
Fig. 2 Regression betweenfine root biomass and watersoluble C (WSC) for 2003(open circles) and 2004(closed circles). The regres-sion function y056.90*(1-e−0.005225x), R200.7108,is significant at p<0.001.WSC data are fromLagomarsino et al. 2006
Plant Soil (2013) 362:93–106 99
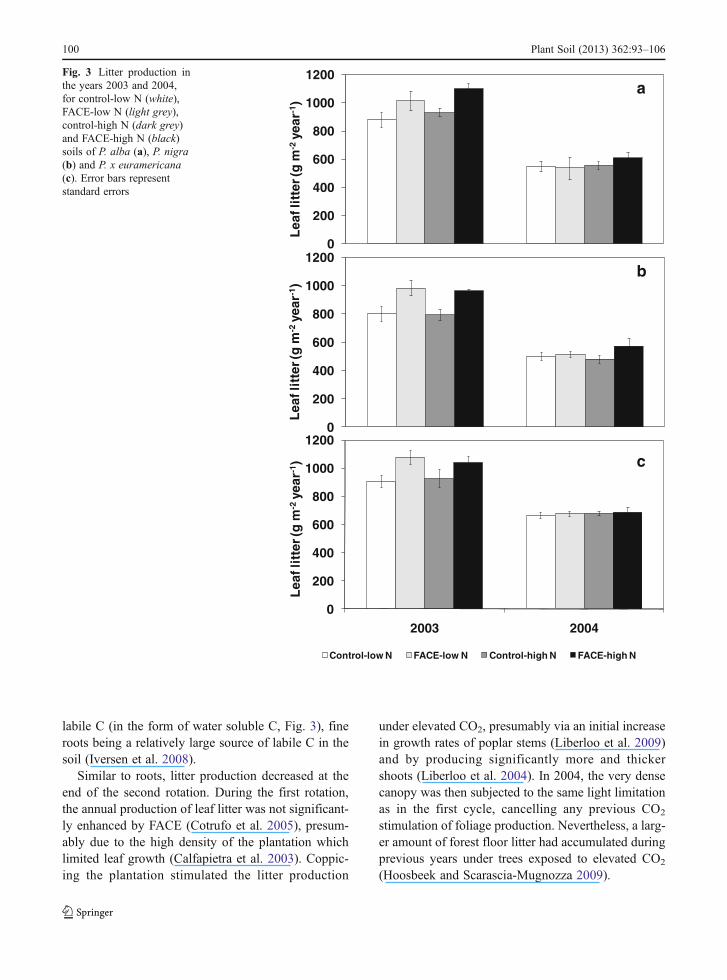
labile C (in the form of water soluble C, Fig. 3), fineroots being a relatively large source of labile C in thesoil (Iversen et al. 2008).
Similar to roots, litter production decreased at theend of the second rotation. During the first rotation,the annual production of leaf litter was not significant-ly enhanced by FACE (Cotrufo et al. 2005), presum-ably due to the high density of the plantation whichlimited leaf growth (Calfapietra et al. 2003). Coppic-ing the plantation stimulated the litter production
under elevated CO2, presumably via an initial increasein growth rates of poplar stems (Liberloo et al. 2009)and by producing significantly more and thickershoots (Liberloo et al. 2004). In 2004, the very densecanopy was then subjected to the same light limitationas in the first cycle, cancelling any previous CO2
stimulation of foliage production. Nevertheless, a larg-er amount of forest floor litter had accumulated duringprevious years under trees exposed to elevated CO2
(Hoosbeek and Scarascia-Mugnozza 2009).
0
200
400
600
800
1000
1200
2003 2004
Lea
f lit
ter (
g m
-2 y
ear-1
)
Control-low N FACE-low N Control-high N FACE-high N
0
200
400
600
800
1000
1200
Lea
f lit
ter (
g m
-2 y
ear-1
)
a
b
c
0
200
400
600
800
1000
1200
Lea
f lit
ter (
g m
-2 y
ear-1
)
Fig. 3 Litter production inthe years 2003 and 2004,for control-low N (white),FACE-low N (light grey),control-high N (dark grey)and FACE-high N (black)soils of P. alba (a), P. nigra(b) and P. x euramericana(c). Error bars representstandard errors
100 Plant Soil (2013) 362:93–106

Mechanisms and drivers increasing soil respiration
An increase in SR under elevated CO2 was observed inthe POP-EUROFACE plantation, as well as in a widerange of other mesocosms (King et al. 2004; Astrid et al.2004; Bernhardt et al. 2006). In addition, the positiveeffect of elevated CO2 on SR in the 2nd rotation (+70 %on average) was larger than that observed by King et al.(2004) in the 1st rotation of this plantation (+39 % onaverage without fertilization).
The strength of SR stimulation under elevated CO2
showed a seasonal and inter-annual pattern. Independentof treatments, the nearly threefold increase in SR in thegrowing compared to dormant season is consistent withother studies which reported three–to fivefold increasesfrom the seasonal low in winter to the seasonal high insummer (Maier and Kress 2000; Epron et al. 2001; King
et al. 2004). In the absence of moisture limitation (DeAngelis, data not shown), the observed pattern correlateswith seasonal variation in temperature, photosyntheticactivity and increased rates of autotrophic (root) respira-tion (Raich and Tufekcioglu 2000; Janssens et al. 2001;Baldocchi et al. 2006). Elevated CO2 induced a steeperslope of the temperature function in the last year of the2nd rotation. Indeed, in the last year of the 2nd rotation, adecrease of SR rates was observed in the control soil, butnot under elevated CO2, with amuch larger stimulation ofSR during the growing than during the dormant season.
Our results suggest that WSC released by roots wasthe primary driver of SR in 2003 (Table 3), explainingmost of the increase observed under elevated CO2. In2004, root biomass was not affected by elevated CO2,not even during the growing season, whereas photo-synthetic rates and the decomposition of above- and
Control-low Ny = 0.2064e0.1359x
R² = 0.7325
FACE-low Ny = 0.4144e0.122x
R² = 0.7852
Control-high Ny = 0.2041e0.1323x
R² = 0.7603
FACE-high Ny = 0.2799e0.1415x
R² = 0.7248
0
1
2
3
4
5
6
7
8
9
10
SR
(µm
ol C
O2
m-2
s-1)
2003
Control-low Ny = 0.335e0.0897x
R² = 0.633
FACE-low Ny = 0.3056e0.1285x
R² = 0.6913
Control-high Ny = 0.2838e0.096x
R² = 0.6949
FACE-high Ny = 0.3483e0.1211x
R² = 0.6314
0
1
2
3
4
5
6
7
8
9
10
0 5 10 15 20 25
SR (µ
mol
CO
2m
-2s-1
)
Temperature - °C
2004
Control-low N FACE-low N Control-high N FACE-high N
Fig. 4 Temperature curvesof SR in the years 2003 and2004 for control (square–grey line) and FACE (dia-mond–black line) low N(open symbol–dotted line)and high N (filled symbol–solid line) soils. Exponentialfunctions and R2 arereported for control-low N,control-high N, FACE-lowN, FACE-high N
Plant Soil (2013) 362:93–106 101

belowground litter accumulated from the previous yearexplained most of the variability in SR. Fine root turn-over is one of themajor pathways of new carbon enteringsoil C pools, besides aboveground litter decomposition.Moreover, several authors suggest that the degree of
rhizosphere respiration by elevated CO2 is much higherthan the enhancement of root biomass (Lekkerkerk et al.1990; Hungate et al. 1997; Cheng and Johnson 1998). AtPOP-EUROFACE light-saturated photosynthesis wasstimulated by 49 % on average across three species at
0
1
2
3
4
5
6
SR (µ
mol
CO
2m
-2s-1
)
2003
0
1
2
3
4
5
6
dormant growing
SR (µ
mol
CO
2m
-2s-1
)2004
Control - low N FACE - low N Control - high N FACE - high N
Fig. 5 Mean values of soilrespiration in the dormantand growing seasons(SR’dormant and SR’growing)calculated for each yearfrom the exponential func-tion considering a meantemperature for both yearsof 9 and 20 °C, respectivelyfor the dormant and growingperiods, for control-low N(white), FACE-low N (lightgrey), control-high N (darkgrey) and FACE-high N(black) soils. Error bars rep-resent standard errors
Controly = 1.1495x + 0.2159
R² = 0.4559
FACEy = 3.1123x - 0.926
R² = 0.67850
1
2
3
4
5
0 1 2 3 4 5
SR
tota
l(µ
mo
l CO
2m
-2s
-1)
SRheterotrophic (µmol CO2 m-2 s-1)
control FACE
Fig. 6 Regression curve be-tween soil and heterotrophicrespiration from P. x eura-mericana soil during thedormant (February) and thegrowing (May) season of2004 (data of SRheterotrophic
are from Lagomarsino et al.2009). Grey square, dottedline: control; black triangle,continuous line: FACE.Both regressions are signifi-cant at p<0.001, functionsare reported for control andFACE soils
102 Plant Soil (2013) 362:93–106

the end of the second rotation without any down-regulation of photosynthetic stimulation under elevatedCO2 (Calfapietra et al. 2005; Liberloo et al. 2007). Fromrecent reviews, about 5 % to 12 % of photosyntheticallyfixed C allocated belowground is thought to be exudatedinto the soil as labile C (Jones et al. 2004, 2009), depend-ing on plant species and age. We therefore contend that
the observed stimulation of SR resulted from larger fluxof fresh photosynthate under elevated CO2 which wasrapidly respired back into the atmosphere. We found agood correlation between total and heterotrophic (includ-ing rhizomicrobial) respiration both in control and FACEsoil (Fig. 6, Lagomarsino et al. 2009). Importantly, thecontribution of root-derived CO2 to the total was
Table 3 Multiple regression analysis (stepwise, forward) forsoil respiration in the dormant and growing periods in 2003and 2004 (SRdormant and SRgrowing). The significance of the
regressions and of independent variables is reported. #p<0.1, *
p<0.05, **p<0.01, ***p<0.001. WSC: water soluble C; MBC:microbial biomass C; MBN: microbial biomass N
Soil respiration 2003 Soil respiration 2004
Dormant season Growing season Dormant season Growing season
r200.690*** r200.66*** r200.526** r200.669***
Independent variables Std. Coefficient p Std. Coefficient p Std. coefficient p Std. Coefficient p
Fine root biomass (g m−2) – n.s. 0.266 # – n.s. – n.s.
Coarse root biomass (g m−2) 0.301 * 0.281 # – n.s. – n.s.
Litter (g Cm−2 y−1) 0.349 * – n.s. – n.s. – n.s.
MBCa (μg Cg−1) – n.s. – n.s. – n.s. – n.s.
WSCa (μg Cg−1) 0.416 ** 0.387 ** – n.s. 0.301 *
MBNb (μg Ng−1) – n.s. – n.s. – n.s. – n.s.
Inorganic Nb (μg N−NH4+ + N−NO3
− g−1) – n.s. – n.s. – n.s. – n.s.
Fine root biomass 2003 (g m−2) 0.416 ** 0.392 **
Litter 2003 (g Cm−2 y−1) 0.238 # 0.303 *
a Moscatelli et al. 2005 and Lagomarsino et al. 2006; b Lagomarsino et al. 2008
Fig. 7 Conceptual model ofprocesses and pools affect-ing SRgrowing in the 2 years,independent of N fertiliza-tion. For each year, process-es (circles) and pools(boxes) are reported withpercentage FACE effects onarrows and within boxes re-spectively. n.s.: no signifi-cant FACE effects; n.d.: notdetermined. C transfer from2003 to 2004 through rootturnover and litter accumu-lation on the forest floor wasdescribed with a broken line
Plant Soil (2013) 362:93–106 103

significantly greater under poplars subjected to CO2
fumigation (Fig. 6). This observation clarifies the impor-tance of the flux of fresh C and its contribution to SR, butit also indicates that larger availability of photosynthateunder elevated CO2 rapidly translates into greater soilCO2 flux, if not fixed in long-lived organs/pools. Thelarger stimulation of SR observed during the growingperiod of 2004 in the absence of the root sink for theextra C fixed under elevated CO2 supports thisstatement.
Currently, several mechanisms of C loss/accumula-tion under elevated CO2 have been discussed, high-lighting the importance of biochemical priming ofSOM decomposition and C stabilization through phys-ical protection (Hoosbeek et al. 2004; Hoosbeek andScarascia-Mugnozza 2009). Lack of consensusderives largely because these processes are thoughtto be related to soil nutrient status. In nutrient poorsoils, a stimulation of microbial activity by greaterorganic matter input may enhance existing SOM de-composition through a priming effect as microbesdecompose old C to obtain nutrients. Nutrient addi-tion, particularly that of N, has led to a reduction ofsuch priming in some studies (Cheng and Coleman1990; Liljeroth et al. 1994). At the POP-EUROFACEsite, the elevated CO2 effect on SR was similar inunfertilized and fertilized soils when nutrient additionwas applied (2002–2004), and N fertilization did notinduce any significant modification on soil CO2
emissions.
Conclusion
An increase in SR under elevated CO2 was observed inthe poplar SRF in both years, but biological propertiesthat induced this stimulation showed a strong inter-annual variation. In 2003, larger fine root biomass andlabile C input drove the increase of SR under elevatedCO2. In 2004, however, the decrease of root biomass incontrol and in FACE lowered SR from control soil only.The larger SR under elevated CO2 was sustained in partby i) the decomposition of above- and belowgroundlitter (higher in elevated CO2 during the previous year)and by ii) the greater availability of easily decomposablesubstrates in the soil. When the larger amount of C fixedthrough photosynthesis under elevated CO2 was notallocated to root biomass, the extra C added to the soilmay have induced higher C losses from the soil during
the growing season, reducing the potential for C accu-mulation. We contend that the switching pattern ofprimary drivers of SR may be typical for the highlydynamic SRF. Understanding belowground processesand feedbacks remain a crucial point to predict soil Closs or accumulation under this type of bio-energyplantations.
Acknowledgments The authors are grateful to Prof. GiuseppeScarascia Mugnozza, coordinator of the EU EUROFACE(EVR1-CT-2002-40027) and MIUR Centre of Excellence “For-ests and climate” projects for allowing the use of POP-EUROFACE experimental station. The authors thanks Mr.Tullio Oro for the technical assistance.
References
Astrid RB, Giesemann A, Anderson TH, Weigel HJ, BuchmannN (2004) Soil respiration under elevated CO2 and its par-titioning into recently assimilated and older carbon sources.Plant Soil 262:85–94
Bader M, Hiltbrunner E, Körner C (2009) Fine root responses ofmature deciduous forest trees to free air carbon dioxideenrichment (FACE). Funct Ecol 23:913–921
Baldocchi D, Tang J, Xu L (2006) How switches and lags inbiophysical regulators affect spatial–temporal variation ofsoil respiration in an oak–grass savanna. J Geophys Res111:GO2008. doi:10.1029/2005JG000063
Bernhardt ES, Barber JJ, Pippen JS, Taneva L, Andrews JA,Schlesinger WH (2006) Long-term effects of Free Air CO2
Enrichment (FACE) on soil respiration. Biogeochem77:91–116
Brown ALP, Day FP, Hungate BA, Drake BG, Hinkle CR(2007) Root biomass and nutrient dynamics in a scrub–oak ecosystem under the influence of elevated atmosphericCO2. Plant Soil 292:219–232
Calfapietra C, Gielen B, Sabatti M, De Angelis P, Miglietta F,Scarascia-Mugnozza GE, Ceulemans R (2003) Do aboveground growth dynamics of poplar change with time underCO2 enrichment? New Phytol 160:305–318
Calfapietra C, Tulva I, Eensalu E, Perez M, De Angelis P,Scarascia-Mugnozza GE, Kull O (2005) Canopy profilesof photosynthetic parameters under elevated CO2 and Nfertilization in a poplar plantation. Environ Pollut 137:525–535
ChengW, ColemanDC (1990) Effect of living roots on soil organicmatter decomposition. Soil Biol Biochem 22:781–787
Cheng W, Johnson DW (1998) Elevated CO2, rhizosphere pro-cesses, and soil organic matter decomposition. Plant Soil202:167–174
Cotrufo MF, De Angelis P, Polle A (2005) Leaf litter productionand decomposition in a poplar short-rotation coppice ex-posed to free air CO2 enrichment (POPFACE). GlobalChange Biol 11:1–12
Davidson EA, Savage K, Bolstad P, Clark DA, Curtis PS, EllsworthDS, Hanson PJ, Law PE, Luo Y, Pregitzer KS, Randolph JC,Zak D (2002) Belowground carbon allocation in forests
104 Plant Soil (2013) 362:93–106

estimates from litterfall and IRGA-based soil respirationmeasurements. Agr Forest Meteorol 113:9–51
Epron D, Le Dantec V, Dufrene E, Granier A (2001) Seasonaldynamics of soil carbon dioxide efflux and simulated rhizo-sphere respiration in a beech forest. Tree Physiol 21:145–152
Giardina CP, Ryan MG, Binkley D, Fownes JH (2003) Primaryproduction and carbon allocation in relation to nutrientsupply in a tropical experiment forest. Global Change Biol9:1438–1450
Handa IT, Hagedorn F, Hättenschwiler S (2008) No stimulationin root production in response to 4 years of in situ CO2
enrichment at the Swiss treeline. Funct Ecol 22:348–358Hogberg P, Nordgren A, Buchmann N, Taylor AFS, Ekblad A,
Hogberg MN, Nyberg G, Ottoson-Lofvenius M, Read DJ(2001) Large-scale forest girdling shows that current pho-tosynthesis drives soil respiration. Nature 411:789–792
Hoosbeek MR, Scarascia-Mugnozza GE (2009) Increased litterbuild up and soil organic matter stabilization in a poplarplantation after 6 years of atmospheric CO2 enrichment(FACE): final results of POP-EUROFACE compared toother forest FACE experiments. Ecosystems 12:220–239.doi:10.1007/s10021-008-9219-z
Hoosbeek MR, Lukac M, van Dam D, Godbold DL, Velthorst EJ,Biondi FA, Peressotti A, CotrufoMF, DeAngelis P, ScarasciaMugnozza G (2004)More new carbon in the mineral soil of apoplar plantation under free air carbon enrichment (POP-FACE): cause of increased priming effect? Global Biogeo-chem Cy 18. doi:10.1029/2003GB002127
Hungate BA, Chapin FS III, Zhong H, Holland EA, Field CB(1997) Stimulation of grassland nitrogen cycling undercarbon dioxide enrichment. Oecologia 109:149–153
Iversen CM, Ledford J, Norby RJ (2008) CO2 enrichmentincreases carbon and nitrogen input from fine roots in adeciduous forest. New Phytol 179:837–847
Janssens IA, Lankreijer H, Matteucci G, Kowalski AS et al(2001) Productivity overshadows temperature in determin-ing soil and ecosystem respiration across European forests.Global Change Biol 7:269–278
Jones DL, Hodge A, Kuzyakov Y (2004) Plant and mycorrhizalregulation of rhizodeposition. New Phytol 163:459–480
Jones DL, Nguyen C, Finlay RD (2009) Carbon flow in therhizosphere: carbon trading at the soil–root interface. PlantSoil 321:5–33
King JS, Hanson PJ, Bernhardt E, De Angelis P, Norby RJ,Pregitzer KS (2004) A multiyear synthesis of soil respirationresponses to elevated atmospheric CO2 from four forestFACE experiments. Global Change Biol 10:1027–1042
Kuzyakov Y, Cheng W (2001) Photosynthesis controls of rhi-zosphere respiration and organic matter decomposition.Soil Biol Biochem 33:1915–1925
Lagomarsino A, Moscatelli MC, De Angelis P, Grego S (2006)Labile substrate quality as the main driving force of micro-bial mineralization activity in a poplar plantation soil underelevated CO2 and nitrogen fertilization. Sci Total Environ372:256–265
Lagomarsino A, Moscatelli MC, Hoosbeek MR, De Angelis P,Grego S (2008) Assessment of soil nitrogen and phospho-rous availability under elevated CO2 and N-fertilization ina short rotation poplar plantation. Plant Soil 308:131–147
Lagomarsino A, De Angelis P, Moscatelli MC, Grego S (2009)The influence of temperature and labile C substrates on
heterotrophic respiration in response to elevated CO2 andnitrogen fertilization. Plant Soil 317:223–234
Lekkerkerk LJA, van De Geijn SC, van Veen J (1990) Effects ofelevated atmospheric CO2 levels on the carbn economy ofa soil planted with wheat. In: Bouwman AF (ed) Soils andthe greenhouse effect. Wiley, USA, pp 423–429
Liberloo M, Gielen B, Calfapietra C, Veys C, Pigliacelli R,Scarascia-Mugnozza GE, Ceulemans R (2004) Growth ofa poplar short rotation coppice under elevated atmosphericCO2 concentrations (EUROFACE) depends on fertilizationand species. Ann For Sci 61:299–307
Liberloo M, Dillen SY, Calfapietra C, Marinari S, Luo ZB, DeAngelis P, Ceulemans R (2005) Elevated CO2 concentra-tion, fertilization and their interaction: growth stimulationin a short-rotation poplar coppice (EUROFACE). TreePhysiol 25:179–189
Liberloo M, Calfapietra C, Lukac M, Godbold D, Luo ZB, PolleA, Hoosbeek M, Kull O, Marek M, Raine CT, Rubino M,Taylor G, Scarascia Mugnozza G, Ceulemans R (2006)Woody biomass production during the second rotation ofa bio-energy Populus plantaion increases in a future highCO2 world. Global Change Biol 12:1094–1106
Liberloo M, Tulva I, Raïm O, Kull O, Ceulemans R (2007)Photosynthetic stimulation under long-term CO2 en-richment and fertilization is sustained across a closedPopulus canopy profile (EUROFACE). New Phytol173:537–549
Liberloo M, Lukac M, Calfapietra C, Hoosbeek MR, Gielen B,Miglietta F, Scarascia-Mugnozza GE, Ceulemans R (2009)Coppicing shifts CO2 stimulation of poplar productivity toabove-ground pools: a synthesis of leaf to stand levelresults from the POP/EUROFACE experiment. New Phy-tol 182:331–346
Liljeroth E, Kuikman P, Van Veen JA (1994) Carbon transloca-tion to the rhizosphere of maize and wheat and influence onthe turnover of native soil organic matter at different soilnitrogen levels. Plant Soil 161:233–240
Lukac M, Calfapietra C, Godbold DL (2003) Root productionand turnover in, pulus grown under elevated CO2 using afree air enrichment system POPFACE. Global Change Biol9:838–848
Maier CA, Kress LW (2000) Soil CO2 evolution and rootrespiration in 11 year-old loblolly pine (Pinus taeda) plan-tations as affected by moisture and nutrient availability.Can J Fort Res 30:347–359
Miglietta F, Peressotti A, Vaccari FP, Zaldei A, De Angelis P,Sarascia Mugnozza G (2001) Free-air CO2 enrichment(FACE) of a poplar plantation: the POPFACE fumigationsystem. New Phytol 150:465–476
Moscatelli MC, Lagomarsino A, Marinari S, De Angelis P, GregoS (2005) Soil microbial indices as bioindicators of environ-mental changes in a poplar plantation. Ecol Ind 5:171–179
Norby RJ (2004) Forest responses to a future CO2-enriched atmo-sphere. In: Steffen W et al (eds) Global change and the earthsystem: a planet under pressure. Springer, Berlin, pp 158–159
Norby RJ, Hanson PJ, O’Neill EG, Tschaplinski TJ, Weltzin JFet al (2002) Net primary productivity of a CO2-enricheddeciduous forest and the implications for carbon storage.Ecol Appl 12:1261–1266
Pregitzer KS, Burton AJ, King JS, Zak DR (2008) Soil respira-tion, root biomass, and root turnover following long-term
Plant Soil (2013) 362:93–106 105

exposure of northern forests to elevated atmospheric CO2
and tropospheric O3. New Phytol 180:153–161Raich JW, Nadelhoffer KJ (1989) Belowground carbon alloca-
tion in forest ecosystems: global trends. Ecology 70:1346–1354
Raich JW, Tufekcioglu A (2000) Vegetation and soil respiration:correlations and controls. Biogeochem 48:71–90
Ryan MG, Law BE (2005) Interpreting, measuring, and model-ing soil respiration. Biogeochem 73:3–27
Ryan MG, Binkley D, Fownes JH, Giardina CP, Senock RS(2004) An experimental test of the causes of forest growthdecline with stand age. Ecol Monogr 74:393–414
Scarascia Mugnozza G, De Angelis P, Sabatti M, Calfapietra C,Ceulemans R, Peressotti A, Miglietta F (2000) A FACEexperiment on short rotation, intensive poplar plantation:objective and experimental set up of POPFACE. In: SuttonMA, et al. (eds) Terrestrial ecosystems research in Europe:
success, challenges and policy. European Communities,Luxemburg, EUR 19375. pp 136–140
Sennerby-Forsse L (1995) Growth processes. Biomass Bioen-ergy 9:35–43
Trueman RJ, Gonzalez-Meler A (2005) Accelerated below-ground C cycling in a managed agriforest acosystem ex-posed to elevated carbon dioxide concentrations. GlobalChange Biol 11:1–14
Van Hees PAW, Jones DL, Finlay R, Godbold DL, LundstromUS (2005) The carbon we do not see—the impact of lowmolecular weight compounds on carbon dynamics andrespiration in forest soils: a review. Soil Biol Biochem37:1–13
Zak DR, Pregitzer KS, King JS, Holmes WE (2000) Elevatedatmospheric CO2, fine roots and the response of soil mi-croorganism: a review and hypotesis. New Phytol147:201–222
106 Plant Soil (2013) 362:93–106
Copyright © 2022 FDOKUMEN




















