
Doubling World production_ Alternative methods
49
DOUBLING WORLD AGRICULTURAL DOUBLING WORLD AGRICULTURAL PRODUCTION PRODUCTION BY ACHIEVING TWO CROPS BY ACHIEVING TWO CROPS SIMULTANEOUSLY SIMULTANEOUSLY WITH ONE SEED WITH ONE SEED CONCEPT
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Doubling World production_ Alternative methods

DOUBLING WORLD AGRICULTURALDOUBLING WORLD AGRICULTURALPRODUCTIONPRODUCTION
BY ACHIEVING TWO CROPSBY ACHIEVING TWO CROPSSIMULTANEOUSLYSIMULTANEOUSLY WITH ONE SEEDWITH ONE SEED
CONCEPT

(https://mrsmatheson.wordpress.com/2012/11/30/higher-genetic-engineering-and-somatic-fusion/)(https://mrsmatheson.wordpress.com/2012/11/30/higher-genetic-engineering-and-somatic-fusion/)
ABSTRACTABSTRACT
Today’s Scenario is more or less like this.Today’s Scenario is more or less like this.
1. Agricultural lands are used for industrial or 1. Agricultural lands are used for industrial or mining or for land filling of Solid Wastes or for mining or for land filling of Solid Wastes or for dwelling thus causing diminishing agricultural lands dwelling thus causing diminishing agricultural lands available.available.
2. Labour is becoming scarce and expensive 2. Labour is becoming scarce and expensive discouraging farmers to grow.discouraging farmers to grow.
3. Wild Animals, Rats and untimely rains, hurricanes,3. Wild Animals, Rats and untimely rains, hurricanes,cyclones etc are damaging agricultural fields and cyclones etc are damaging agricultural fields and thus reducing total available yields.thus reducing total available yields.
4. Pests and insects are becoming more and more 4. Pests and insects are becoming more and more pesticide resistant and reducing the yields.pesticide resistant and reducing the yields.

5. Seasonal fluctuations in market prices sometimes 5. Seasonal fluctuations in market prices sometimes lead to heavy losses to the farmers leading to lead to heavy losses to the farmers leading to suicides in many developing and under-delveloped suicides in many developing and under-delveloped countries where agriculture is the main occupation.countries where agriculture is the main occupation.
6. Younger generations are attracted to more 6. Younger generations are attracted to more remunerative occupations.remunerative occupations.
7. Over generations inherited lands got divided and 7. Over generations inherited lands got divided and became smaller with lesser cultivable land after became smaller with lesser cultivable land after volumes of the boundaries are increased. volumes of the boundaries are increased.
In this context a need arises to solve all these In this context a need arises to solve all these problems. problems. A better proposition is to enable a farmer to use oneA better proposition is to enable a farmer to use oneseed which can yield one crop above the ground and seed which can yield one crop above the ground and simultaneously another crop under the ground like simultaneously another crop under the ground like POMATO which was designed to produce Tomatoes above POMATO which was designed to produce Tomatoes above the ground and Potatoes under the ground; thereby the ground and Potatoes under the ground; thereby farmer will be having a chance to have a better pricefarmer will be having a chance to have a better priceatleast to one of them.atleast to one of them.
In this article efforts are made In this article efforts are made 1.1.to compile the available technologiesto compile the available technologies2.2.to better understand the protocols involved in to better understand the protocols involved in
simpler termssimpler terms3.3.to use improvised versions of the methodologies to to use improvised versions of the methodologies to
augment sustained world food production in the augment sustained world food production in the coming Yearscoming Years. .
MISSION:MISSION:I am looking for associates who can involve with us I am looking for associates who can involve with us in technology, production and marketing on contract in technology, production and marketing on contract basis.basis.

PROLOGUEPROLOGUEFood production must double by 2050 to meet the demand of the world’s growing population and innovative strategies are needed to help combat hunger, which already affects more than 1 billion peoplein the world, several experts today told the Second Committee (Economic and Financial) during a panel discussion on “New cooperation for global food security”.The 2008 food price crisis, the result of decades of insufficient investment in agriculture and food security, swelled the ranks of the poor and undernourished to 1 billion people, and, according to the Food and Agriculture Organization (FAO), an extra 100 million people could go hungry this year as a consequence of the combined negative effects of the food price crisis, the financial and economic crisis, and climate change.Rajul Pandya-Lorch, Chief of Staff and Head of the 2020 Vision for Food, Agriculture, and the Environment Initiative, International Food Policy and Research Institute (IFPRI), said that, despite efforts to improve food security by the international community, thenumber of people going hungry has remained relatively constant over the last half century, which forced the question: “What do we need to do differently?On an inquiry about the tension between supporting smallholder farmers and the need for substantial, larger-scale agricultural investment to improve food security, he said it was an issue of maximum efficiency, and that one way to encourage that could be through support of cooperatives. “Our main job now is to show that,with the available resources, we can get good results during the next few years,” he said.(http://www.un.org/press/en/2009/gaef3242.doc.htm)
HISTORYHISTORYThe concept of grafting related potatoes and tomatoes so that both are produced on the same plant was originally developed in 1977 at the Max Planck Institute for Developmental Biology in Tübingen, Germany, and although healthy, the plant produced neither potatoes nor tomatoes.[2] The Max Planck

Institute for Plant Breeding Research in Köln produced a plantwith fruit in 1994.[2]
As with all grafts, this plant will not occur in nature and cannot be grown from seed, because the two parts of the plant remain genetically separate, and only rely on each other for nourishment and growth. Like most standard types of plant grafting, a small incision is made in the stem of both plants and they are strapped together. Once the cuts have healed and the plants are joined, the leafy top of the potato plant can be cut away and the roots of the tomato can be removed, leaving the leaves of the tomato plant to nourish the roots ofthe potato plant.[3] The rootstock (potato) acts as a stable and healthy root system and the scions (tomato) are chosen fortheir fruit, flowers or leaves. The tomatoes should be ready to harvest after about 12 weeks during the summer months, the potatoes should be ready after the tomato leaves begin to die back, normally in early autumn.[4] Grafting in this way can be used to produce many different related crops from the same plant, for example the growing popularity of 'Fruit salad' trees, which is a single tree that produces multiple types of citrus fruits, or a tree with a variety of fruits with stones (peach, plum etc.). ((http://en.wikipedia.org/wiki/Pomato)
In 1972, Carlson and his associates produced the first inter-specific somatic hybrid between Nicotiana glauca and N. langsdorffii. In 1978, Melchers and his co-workers developed the first inter-genetic somatic hybrids between Solanum tuberosum (potato) and Lycopersicon esculentum (tomato). The hybrids are knownas ‘Pomatoes or Topatoes’(http://www.biotechnology4u.com/plant_biotechnology_applications_cell_tissue_culture.html)
Grafted pomato plants have been launched in the United Kingdom in September 2013 by horticultural mail order company Thompson & Morgan, who sold pre-grafted plants branded as the "TomTato". The Incredible Ediblenursery in New Zealand announced a "DoubleUP Potato Tom" in the same monthGrafting is a difficult process because the tomato and the potato stems have to be the same thickness, and Thompson & Morgan trialled the hybrid for several years before selling it. Production and grafting of tomtatos begins in a specialist laboratory in the

Netherlands, before being shipped back to the UK and grown in greenhouses until they are ready to be sold.(http://en.wikipedia.org/wiki/Pomato)
“In Florida, most orange trees have lemon roots. In California, nearly all lemon trees are grown on orange roots. This sort of thingis not unique with citrus. With the stone fruits, there is a certainlatitude. Plums can be grown on cherry trees and apricots on peach trees, but a one-to-one relationship like that is only the beginningwith citrus. A single citrus tree can be turned into a carnival, with lemons, limes, grapefruit, tangerines, kumquats and oranges allripening on its branches at the same time…Most of the trees on the Ridge [a mountain range in Florida renowned for its orange tree orchards] are growing on Rough Lemon…As a rootstock, it forages withexceptional vigor and, in comparison with others, puts more fruit onthe tree.”Although grafting woody plants, like fruit trees, is an ancient horticultural technique, grafting soft-stemmed vegetables is a much more recent agricultural practice. Perhaps nurseries will soon startselling mixed vegetable shrubs alongside fruit salad trees. Brassica oleracea seems like a particularly good candidate for such an experiment. This one species includes cabbage, broccoli, cauliflower, Brussels sprouts and kale. Yes, all these plants are cultivars of the exact same species—their appearances and characteristics have been altered through artificial selection over the generations, in the same way people have created so many different dog breeds. A broccauliflower sprouts plant sounds particularly delicious. Maybe it’s time for a family reunion.(http://blogs.scientificamerican.com/brainwaves/2012/09/10/the-science-of-pomato-plants-and-fruit-salad-trees/)
Previously NONGWOO BIO developed a new CMS line (NWB-CMS) for radishand the NWB-CMS line indeed provided lots of sub-lines of Radish. And the Biotechnology Institute has developed the new Cruciferae plants by fusing the NWB-CMS cells of Radish to the ordinary cells of Chinese Cabbage and Cabbage.(http://www.nongwoobio.com/nongwoo/html/tech_02.html)
SCIENCESCIENCEThe seed is the mature, fertilized ovule. After fertilization, the haploid cells of the embryo sac disintegrate. The maternally deriveddiploid cells of the ovule develop into the hard, water-resistant outer covering of the seed, called the testa, or seed coat. The diploid zygote develops into the embryo, and the triploid endosperm

cells multiply and provide nutrition. The testa usually shows a scarcalled the hilum where the ovule was originally attached to the funicle. In some seeds a ridge along the testa called the raphe shows where the funicle originally was pressed against the ovule. The micropyle of the ovule usually survives as a small pore in the seed coat that allows passage of water during germination of the seed.
In some species, the funicle develops into a larger structure on theseed called an aril, which is often brightly colored, juicy, and contains sugars that are consumed by animals that may also disperse the seed (as in nutmeg, arrowroot, oxalis, and castor bean). This isdistinct from the fruit, which forms from the ovary itself.
The embryo consists of the cotyledon(s) , epicotyl, and hypocotyl. The cotyledons resemble small leaves, and are usually the first photosynthetic organs of the plant. The portion of the embryo above the cotyledons is the epicotyl, and the portion below is the hypocotyl. The epicotyl is an apical meristem that produces the shoot of the growing plant and the first true leaves after germination. The hypocotyl develops into the root. Often the tip of the hypocotyl, the radicle, is the first indication of germination as it breaks out of the seed. Flowering plants are classified as monocotyledons or dicotyledons (most are now called eudicots ) basedon the number of cotyledons produced in the embryo. Common monocotyledons include grasses, sedges, lilies, irises, and orchids;common dicotyledons include sunflowers, roses, legumes, snapdragons,and all nonconiferous trees.
The endosperm may be consumed by the embryo, as in many legumes, which use the cotyledons as a food source during germination. In other species the endosperm persists until germination, when it is used as a food reserve. In grains such as corn and wheat, the outer layer of the endosperm consists of thick-walled cells called aleurone, which are high in protein.

Anatomy of the reproductive organs in angiosperms. (http://www.biologyreference.com/Re-Se/Reproduction-in-Plants.html#ixzz3T29y3X8n)
SEARCH OF LITERATURESEARCH OF LITERATUREAPPLICATIONS OF SOMATIC HYBRIDIZATION
a) Creation of hybrids with disease resistance - Many disease resistance genes (e.g. tobacco mosaic virus, potato virus X, club rot disease) could be successfully transferred from one species to another. E.g resistance has been introduced in tomato against diseases such as TMV, spotted wilt virus and insect pests.b) Environmental tolerance - using somatic hybridization the genes conferring tolerance for cold, frost and salt were introduced in e.g. in tomato.c) Cytoplasmic male sterility - using cybridization method, it was possible to transfer cytoplasmic male sterility.d) Quality characters - somatic hybrids with selective characteristics have been developed e.g. the production of high nicotine content.
CHROMOSOME NUMBER IN SOMATIC HYBRIDSThe chromosome number in the somatic hybrids is generally more than the total number of both of the parental protoplasts. If the chromosome number in the hybrid is the sum of the chromosomes of thetwo parental protoplasts, the hybrid is said to be symmetric hybrid. Asymmetric hybrids have abnormal or wide variations in the chromosome number than the exact total of two species.(http://www.biotechnology4u.com/plant_biotechnology_applications_cell_tissue_culture.html)

TRADITIONAL METHODSTRADITIONAL METHODSGrafting or graftage[1] is a horticultural technique whereby tissues from oneplant are inserted into those of another so that the two sets of vascular tissues may join together. This vascular joining is called inosculation. The technique is most commonly used in asexual propagation of commercially grown plants for the horticultural and agricultural trades.In most cases, one plant is selected for its roots and this is called the stock orrootstock. The other plant is selected for its stems, leaves, flowers, or fruits and is called the scion or cion.[1] The scion contains the desired genes to be duplicated in future production by the stock/scion plant.
In stem grafting, a common grafting method, a shoot of a selected, desired plantcultivar is grafted onto the stock of another type. In another common form called bud grafting, a dormant side bud is grafted onto the stem of another stock plant, and when it has inosculated successfully, it is encouraged to grow by pruning off the stem of the stock plant just above the newly grafted bud.
For successful grafting to take place, the vascular cambium tissues of the stock and scion plants must be placed in contact with each other. Both tissues must be kept alive until the graft has "taken", usually a period of a few weeks. Successful grafting only requires that a vascular connection take place between the grafted tissues. Joints formed by grafting are not as strong as naturally formed joints, so a physical weak point often still occurs at the graft because only the newly formed tissues inosculate with each other. The existing structural tissue (or wood) of the stock plant does notfuse.(http://en.wikipedia.org/wiki/Grafting)
NOVEL ATERNATIVE- SOMATIC FUSIONNOVEL ATERNATIVE- SOMATIC FUSIONSexual hybridization since time immemorial has been used as a methodfor crop improvement but it has its own limitations as it can onlybe used within members of same species or closely related wildspecies. Thus, this limits the use of sexual hybridization as a

means of producing better varieties. Development of viable cellhybrids by somatic hybridization, therefore, has been considered asan alternative approach for the production of superior hybridsovercoming the species barrier. The technique can facilitatebreeding and gene transfer, bypassing problems associated withconventional sexual crossing such as, interspecific, intergenericincompatibility. This technique of hybrid production via protoplastfusion allows combining somatic cells (whole or partial) fromdifferent cultivars, species or genera resulting in novel geneticcombinations including symmetric somatic hybrids, asymmetric somatichybrids or somatic cybrids.
The most common target using somatic hybridization is the gene ofsymmetric hybrids that contain the complete nuclear genomes alongwith cytoplasmic organelles of both parents. This is unlike sexualreproduction in which organelle genomes are generally contributed bythe maternal parent. On the other hand, somatic cybridization is theprocess of combining the nuclear genome of one parent with themitochondrial and/or chloroplast genome of a second parent. Cybridscan be produced by donor-recipient method or by cytoplast-protoplastfusion. Incomplete asymmetric somatic hybridization also providesopportunities for transfer of fragments of the nuclear genome,including one or more intact chromosomes from one parent (donor)into the intact genome of a second parent (recipient).These methods provide genetic manipulation of plants overcominghurdle of sexual incompatibility, thereby, serving as a method ofbringing together beneficial traits from taxonomically distinctspecies which cannot be achieved by sexual crosses. Severalparameters such as, source tissue, culture medium and environmentalfactors influence ability of a protoplast derived hybrid cells todevelop into a fertile plant. The general steps involved in somatichybridization and cybridization methods are elaborated in Figure13.1 and 13.2 .
Steps involved in somatic hybridization
The somatic fusion process occurs in four steps:The removal of the cell wall of one cell of each type of plant using cellulase enzyme to produce a somatic cell called a protoplast
1. The cells are then fused using electric shock (electrofusion) or chemical treatment to join the cells and fuse together the nuclei. The resulting fused nucleus is called heterokaryon.
2. The somatic hybrid cell then has its cell wall induced to formusing hormones
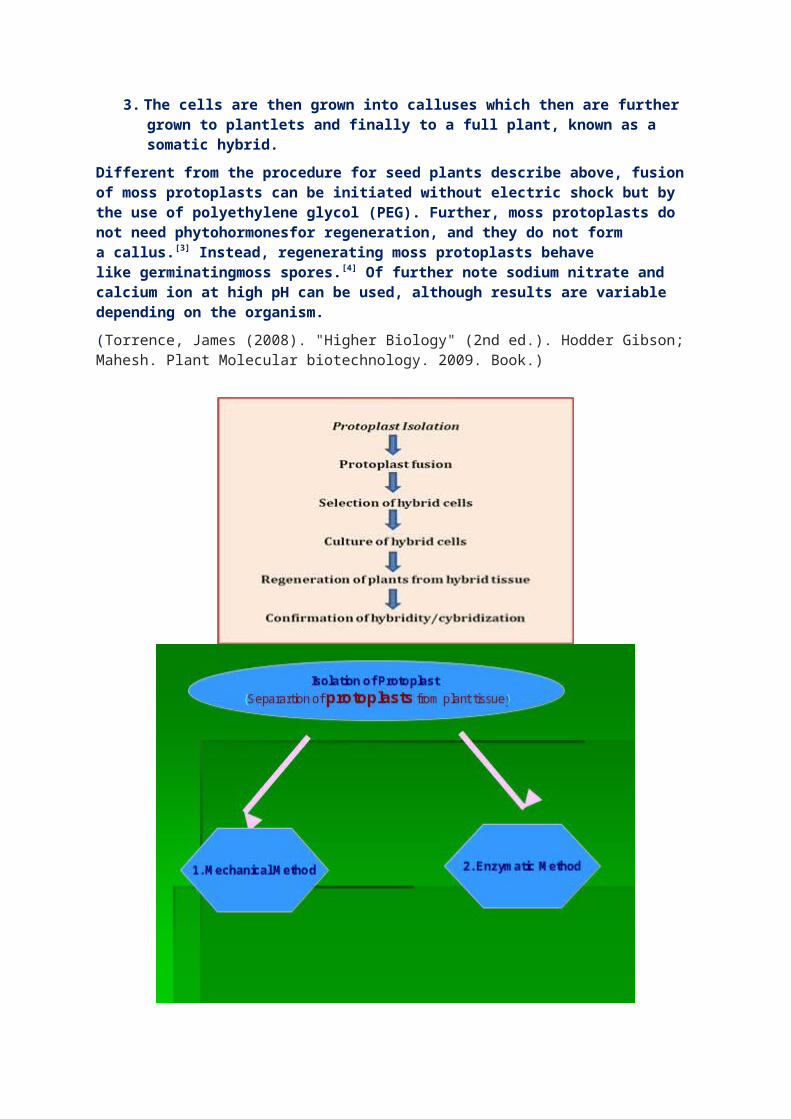
3. The cells are then grown into calluses which then are further grown to plantlets and finally to a full plant, known as a somatic hybrid.
Different from the procedure for seed plants describe above, fusion of moss protoplasts can be initiated without electric shock but by the use of polyethylene glycol (PEG). Further, moss protoplasts do not need phytohormonesfor regeneration, and they do not form a callus.[3] Instead, regenerating moss protoplasts behave like germinatingmoss spores.[4] Of further note sodium nitrate and calcium ion at high pH can be used, although results are variable depending on the organism.(Torrence, James (2008). "Higher Biology" (2nd ed.). Hodder Gibson; Mahesh. Plant Molecular biotechnology. 2009. Book.)

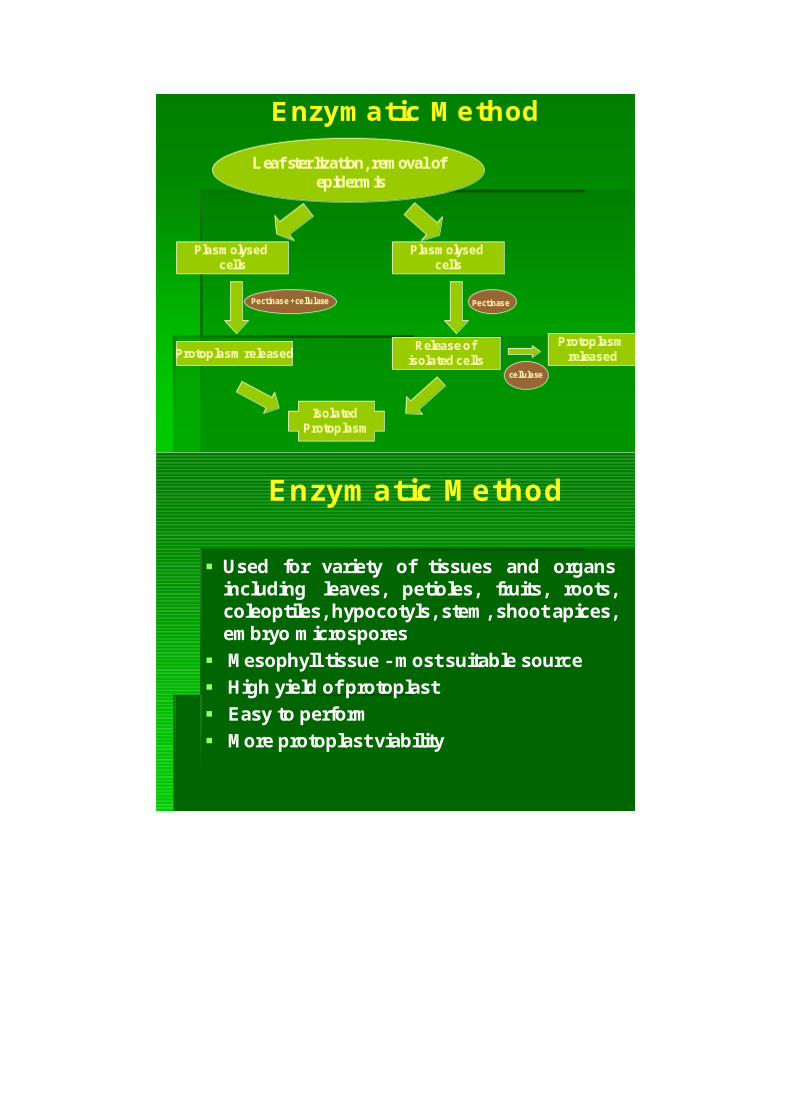
Enzym atic M ethod
Used for variety of tissues and organsincluding leaves, petioles, fruits, roots,coleoptiles,hypocotyls,stem ,shootapices,em bryo m icrospores
M esophylltissue -m ostsuitable source High yield ofprotoplast Easy to perform M ore protoplastviability


(https://www.google.co.in/url?sa=t&rct=j&q=&esrc=s&source=web&cd=10&cad=rja&uact=8&ved=0CFUQFjAJ&url=http
%3A%2F%2Fwww.scirp.org%2Fjournal%2FPaperDownload.aspx%3FpaperID%3D35851&ei=t5PxVI-GI4yjugS-
i4LoBg&usg=AFQjCNEOVeQ2SGm5UNLSRVqlQBxp_Tkecw&bvm=bv.87269000,d.c2E)
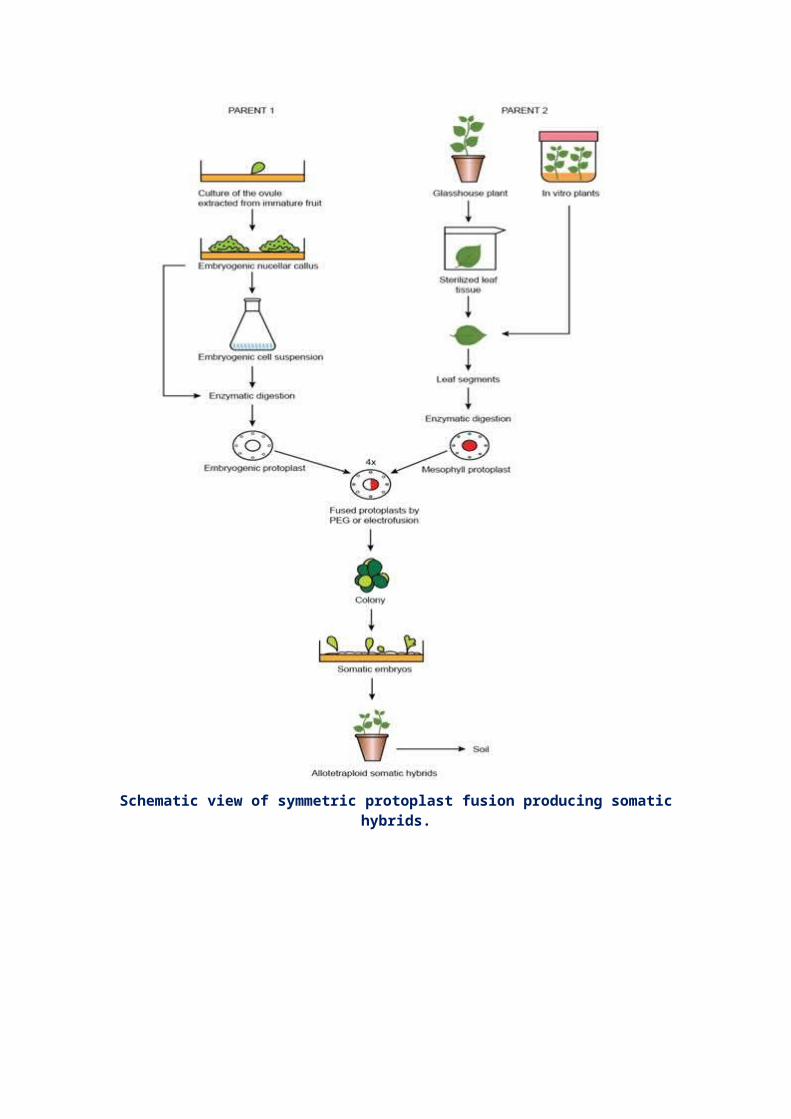
Schematic view of symmetric protoplast fusion producing somatichybrids.
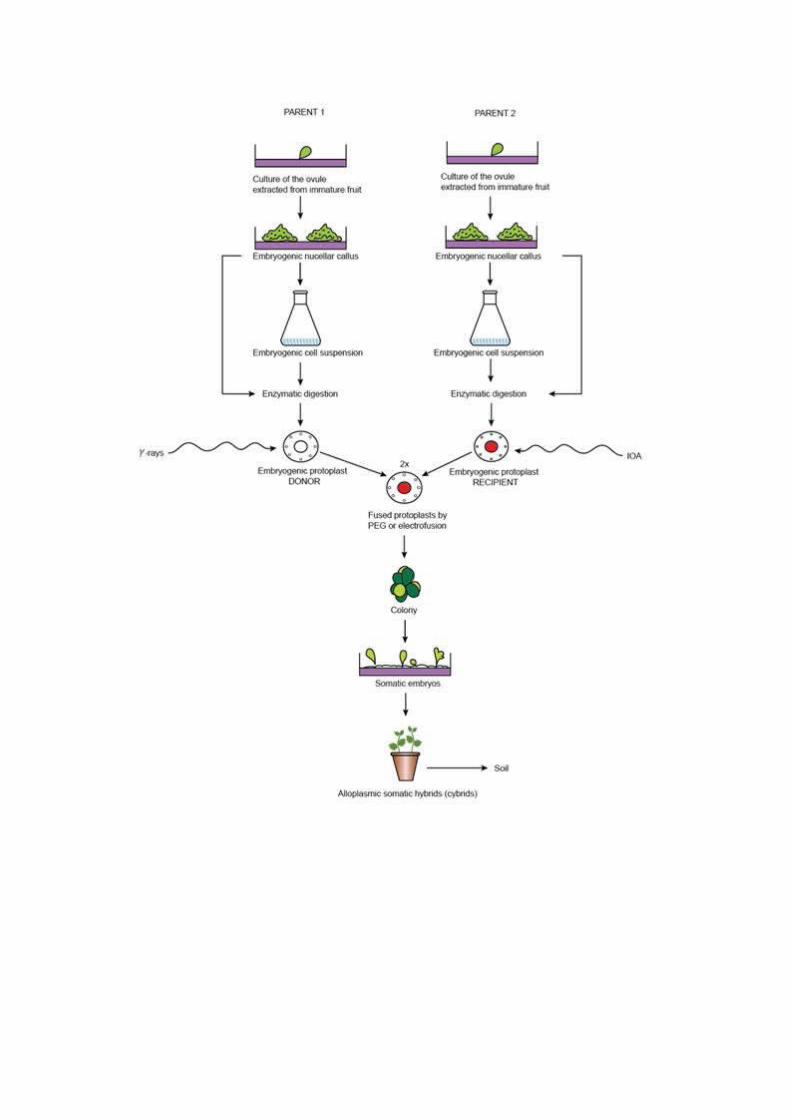
Schematic view of asymmetric protoplast fusion using donor-recipientmethod resulting into creation of alloplasmic somatic hybrid or
cybrids.
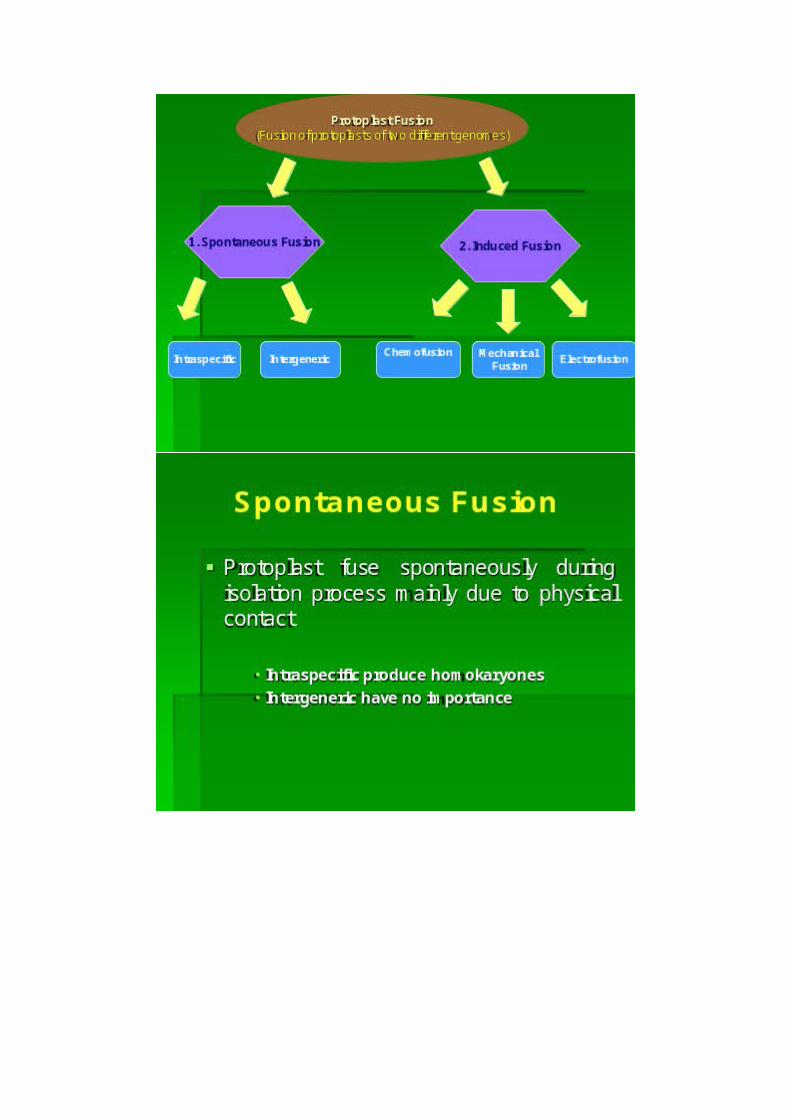
2. Protoplast fusionProtoplast fusion could be spontaneous during isolation ofprotoplast or it can be induced by mechanical, chemical and physicalmeans. During spontaneous process, the adjacent protoplasts fusetogether as a result of enzymatic degradation of cell walls forminghomokaryons or homokaryocytes, each with two to several nuclei. The

occurrence of multinucleate fusion bodies is more frequent when theprotoplasts are prepared from actively dividing callus cells orsuspension cultures. Since the somatic hybridization orcybridization require fusion of protoplasts of different origin, thespontaneous fusion has no value. To achieve induced fusion, asuitable chemical agent (fusogen) like, NaNO3, high Ca2+,polyethylene glycol (PEG), or electric stimulus is needed.i. Fusion by means of NaNO3: It was first demonstrated by Kuster in1909 that the hypotonic solution of NaNO3 induces fusion of isolatedprotoplast forming heterokaryon (hybrid). This method was fullydescribed by Evans and Cocking (1975), however this method has alimitation of generating few no of hybrids, especially when highlyvacuolated mesophyll protoplasts are involved.
ii. High pH and Ca++ treatment: This technique lead to thedevelopment of intra- and interspecific hybrids. It was demonstratedby Keller and Melcher in 1973. The isolated protoplasts from twoplant species are incubated in 0.4 M mannitol solution containinghigh Ca++(50 mM CaCl2.2H2O) with highly alkaline pH of 10.5 at 37°Cfor about 30 min. Aggregation of protoplasts takes place at once andfusion occurs within 10 min.
iii. Polyethylene glycol treatment: Polyethylene glycol (PEG) is themost popularly known fusogen due to ability of forming highfrequency, binucleate heterokaryons with low cytotoxicity. With PEGthe aggregation occurred mostly between two to three protoplastsunlike Ca++ induced fusion which involves large clump formation. Thefreshly isolated protoplasts from two selected parents are mixed inappropriate proportions and treated with 15-45% PEG (1500-6000MW)solution for 15-30 min followed by gradual washing of theprotoplasts to remove PEG. Protoplast fusion occurs during washing.The washing medium may be alkaline (pH 9-10) and contain a high Ca+
+ ion concentration (50 mM). This combined approach of PEG and Ca+
+ is much more efficient than the either of the treatment alone. PEGis negatively charged and may bind to cation like Ca++, which inturn, may bind to the negatively charged molecules present in plasmalemma, they can also bind to cationic molecules of plasma membrane.During the washing process, PEG molecules may pull out the plasmalemma components bound to them. This would disturb plamalemmaorganization and may lead to the fusion of protoplasts located closeto each other (Figure 13.3). The technique is nonselective thus,induce fusion between any two or more protoplasts.

Sequential stages in protoplast fusion. (A) two separateprotoplasts, (B) agglutination of two protoplasts, (C and D)
Membrane fusion at localized site, and (E and F) development ofspherical heterokaryon.
iv. Electrofusion: The chemical fusion of plant protoplast has manydisadvantages – (1) The fusogen are toxic to some cell systems, (2)it produces random, multiple cell aggregates, and (3) must beremoved before culture. Compare to this, electrofusion is rapid,simple, synchronous and more easily controlled. Moreover, thesomatic hybrids produced by this method show much higher fertilitythan those produced by PEG-induced fusion.
Zimmermann and Scheurich (1981) demonstrated that batches ofprotoplasts could be fused by electric fields by devising a protocolwhich is now widely used. This protocol involves a two-step process.First, the protoplasts are introduced into a small fusion chambercontaining parallel wires or plates which serve as electrodes.Second, a low-voltage and rapidly oscillating AC field is applied,which causes protoplasts to become aligned into chains of cellsbetween electrodes. This creates complete cell-to-cell contactwithin a few minutes. Once alignment is complete, the fusion isinduced by application of a brief spell of high-voltage DC pulses(o.125-1 kVcm-1). A high voltage DC pulses induces a reversiblebreakdown of the plasma membrane at the site of cell contact,leading to fusion and consequent membrane reorganization. The entireprocess can be completed within 15 min.
3. Selection of fusion productsThe somatic hybridization by electrofusion of protoplasts allow one-to-one fusion of desired pairs of protoplasts and, therefore, it is

easy to know the fate of fusion products. However, protoplastsuspension recovered after chemical treatments (fusogen) consists ofthe following cell types:
i. unfused protoplasts of the two species/strains
ii. products of fusion between two or more protoplasts of the samespecies (homokaryons), and
iii. ‘hybrid' protoplasts produced by fusion between one (or more)protoplasts of each of the two species (heterokaryons)
The heterokaryons which are the potential source of future hybridsconstitute of a very small (0.5-10%) proportion of the mixture.Therefore, an effective strategy has to be employed for theiridentification and isolation. Various protocols have been proposedand practiced for the effective selection of hybrids, includingmorphological basis, complementation of biochemical and genetictraits of the fusing partners, and manual or electronic sorting ofheterokaryons/hybrid cells.
3.1. Morpho-physiological basis: The whole mixture of theprotoplasts are cultured after fusion treatment and the resultingcalli or regenerants are screened for their hybrid characteristics.Occasionally the hybrid calli outgrow the parental cell colonies andare identified by their intermediate morphology, i.e. green withpurple coloured cells. However, the process is labour intensive andrequires glasshouse facilities. It is limited to certaincombinations showing differences in their regeneration potentialunder specific culture conditions. 3.2. Complementation: In this case complementation or genetic ormetabolic deficiencies of the two fusion partners are utilized toselect the hybrid component. When protoplasts of two parents, (oneparent bearing cytoplasmic albino trait and the other parent bearinggreen trait) each parent carrying a non-allelic genetic or metabolicdefect are fused, it reconstitutes a viable hybrid cell, of wildtype in which both defects are mutually abolished bycomplementation, and the hybrid cells are able to grow on minimalmedium non-permissive to the growth of the parental cells bearinggreen trait. Later, the calli of hybrid nature could be easilydistinguished from the parental type tissue (albino trait) by theirgreen color. The complementation selection can also be applied todominant characters, such as dominant resistance to antibiotics,herbicides or amino acid analogues.

3.3. Isolation of heterokaryons or hybrid cells: The manual orelectronic isolation of heterokaryons or hybrid cells is the mostreliable method. Manual isolation requires that the two parentaltype protoplasts have distinct morphological markers and are easilydistinguishable. For example, green vacuolated, mesophyllprotoplasts from one parent and richly cytoplasmic, non greenprotoplasts from cultured cells of another parent. The dualfluorescence method also helps easy identification of fusionproducts. In this case, the protoplast labeled green by treatmentwith fluorescein diacetate (FDA, 1-20 mgl-1) are fused withprotoplasts emitting a red fluorescence, either from chlorophyllautofluorescence or from exogenously applied rhodamineisothiocyanate (10-20 mgl-1). The labeling can be achieved by addingthe compound into the enzyme mixture. This can be applied even formorphologically indistinguishable protoplasts from two parents.
4. Verification and characterization of somatic hybridsAs no system is foolproof and they have their own advantages anddisadvantages. Therefore, even after selecting the desiredhybrids/cybrids following protoplast fusion, it is required to carryout one or more tests to compare the parent protoplast lines withthe putative hybrids. Some of the techniques that can be tried are: 4.1. Morphology: Somatic hybrids in most of the cases showcharacters intermediate between the two parents such as, shape ofleaves, pigmentation of corolla, plant height, root morphology andother vegetative and floral characters. The method is not muchaccurate as tissue culture conditions may also alter somemorphological characters or the hybrid may show entirely new traitsnot shown by any of the parents. 4.2. Isozyme analysis: Multiple molecular forms of same enzyme whichcatalyses similar or identical reactions are known as isozymes.Electrophoresis is performed to study banding pattern as a check forhybridity. If the two parents exhibit different band patterns for aspecific isozyme the putative hybrid can be easily verified. Theisozymes commonly used for hybrid identification include, acidphosphatase, esterase, peroxidase. 4.3. Cytological analysis: Chromosome counting of the hybrid is aneasier and reliable method to ensure hybridity as it also providesthe information of ploidy level. Cytologically the chromosome countof the hybrid should be sum of number of chromosomes from both theparents. Besides number of chromosomes, the size and structure ofchromosomes can also be monitored. However, the approach is notapplicable to all species, particularly where fusion involves

closely related species or where the chromosomes are very small.Moreover, sometimes the somaclonal variations may also give rise todifferent chromosome number. 4.4. Molecular analysis: Specific restriction pattern of nuclear,mitochondrial and chloroplast DNA characterizes the plastomes ofhybrids and cybrids. Molecular markers such as RFLP, RAPD, ISSR canbe employed to detect variation and similarity in banding pattern offused protoplasts to verify hybrid and cybrid.
5. Cybrids or cytoplasmic hybrids:Sexual hybridization involves fusion of the nuclear genes of boththe parents but somatic hybrids involves even cytoplasm from boththe parental species in hybrid obtained by protoplast fusion.However, in another case somatic hybrids containing nuclear genomeof one parent but cytoplasm from both the parents, are termed ascybrids. The approach is time consuming and require several years ofcrossing plants provides an opportunity to study interparentalmitochondrial, chloroplast fusion giving rise to plants with novelgenomes.
5.1. Methods to produce cybrids: They are produced in variablefrequencies in normal protoplast fusion experiments due to one ofthe following methods:1. Fusion of normal protoplast with an enucleated protoplast. Theenucleated protoplast can be produced by high speed centrifugation(20,000-40,000xg) for 60 min with 5-50% percoll.2. Fusion between a normal protoplast and another protoplast with anon-viable nucleus or suppressed nucleus.3. Elimination of one of the nuclei after heterokaryons formation.4. Selective elimination of chromosomes at a later stage.5. Irradiating (with X-rays or gamma rays) the protoplasts of onespecies prior to fusion in order to inactivate their nuclei.6. By preparing enucleate protoplasts (cytoplasts) of one speciesand fusing them with normal protoplasts of the other species.Cybrids provide the following unique opportunities: (i) transfer ofplasmogenes of one species into the nuclear background of anotherspecies in a single generation, and even in (ii) sexuallyincompatible combinations, (iii) recovery of recombinants betweenthe parental mitochondrial or chloroplast DNAs (genomes), and (iv)production of a wide variety of combinations of the parental andrecombinant chloroplasts with the parental or recombinantmitochondria.
6. Applications of somatic hybridization

1. Novel interspecific and intergeneric crosses which are difficultto produce by conventional methods can be easily obtained.2. Important characters, such as resistance to diseases, ability toundergo abiotic stress and other quality characters, can be obtainedin hybrid plant by the fusion of protoplasts of plant bearingparticular character to the other plant which may be susceptible todiseases.3. Protoplasts of sexually sterile haploid, triploid, aneuploidplants can be fused to obtain fertile diploids and polyploids.4. Studying cytoplasmic genes may be helpful to carry out plantbreeding.5. Most of the agronomically important traits, such as cytoplasmicmale sterility, antibiotic resistance and herbicide resistance, arecytoplasmically encoded, hence can be easily transferred to otherplant.6. Plants in juvenile stage can also be hybridized by means ofsomatic hybridization.7. Somatic hybridization can be used as a method for the productionof autotetraploids.
Limitations of somatic hybridization1. Application of protoplast methodology requires efficient plantregeneration system from isolated protoplasts. Protoplasts from twospecies can be fused, however, production of somatic hybrids is noteasy.2. Lack of a proper selection method for fused products (hybrids)poses a problem.3. The end product of somatic hybridization are often unbalanced(sterile, misformed and unstable)4. Somatic hybridization of two diploids leads to formation ofamphidiploids which is unfavorable.5. It is not sure for a character to completely express aftersomatic hybridization.6. The regeneration products of somatic hybridization are oftenvariable due to somaclonal variation, chromosome elimination,organelle segregation.7. All diverse intergeneric somatic hybrids are sterile and,therefore, have limited chances of development of new varieties.8. To transfer useful genes from wild species to cultivated crop, itis necessary to achieve intergeneric recombination or chromosomesubstitution between parental genomes.(http://nptel.ac.in/courses/102103016/13)
A simple method for the isolation of plant protoplasts

2. Materials and methods 2·1 Source tissue Tissue for protoplast isolation consisted of fully expanded leaves excised from greenhouse-grown plants (Basella, Centella, Withania, Amaranthus, Catharanthus, Phlox, Petunia) as well as from actively-growing shootcultures (tomato cultivars viz·, Arkavikas, pusa Ruby and Punjab Kesari; chilli, Nicotiana rustica, N. glutinosa and N· tabacum cv. Thompson), cotyledons of aseptically-germinated seedlings (sunflowerand niger), placental tissue of fruits (tomato) and friable embryogenic callus (sandalwood). Shoot cultures were stabilized in vitro· The culture conditions and media for the shoot cultures of each species were as follows. Surface sterilized seeds of N. tabacumcv Thompson, N. rustica and N. glutinosa were germinated on half strength MS medium for explanting shoots· Thereafter, shoot cultureswere maintained by routine subculture onto the same medium viz., half strength MS medium. Leaves were obtained for protoplast isolation 3-4 weeks after every subculture· Tomato axillary shoots were obtained from 3–4 weeks old greenhouse grown plants and taken to culture in half strength MS medium. Surface sterilized seeds of sunflower and niger were germinated on half strength MS medium. Seeds were dark-incubated for germination and were shifted to 14 h exposure to light of 200 µEm-2 s- 1 intensity at 25 ± 2°C temperature. Rapidly-growing embryogenic sandalwood callus was obtained from foully developed endosperm as tissue explant. Callus was induced on MS medium incorporated with 2,4-dichlorophenoxyaceticacid (2,4-D) (1 mg/1) and kinetin (0·5 mg/1) and then transferred toMS medium supplemented with 1 mg/1 of 2,4-D for eliciting embryogenic competence.
2.2 Explant preparation prior to enzyme exposure Leaves of field-grown plants were thoroughly washed in running water for about 10 min, cleaned gently with dilute Labolene of Glaxo India Ltd., and quickly rinsed with tap water first and then with deionized water· Sterilization was carried out with 2% sodium hypochlorite for 5 min followed by several rinses with sterile distilled water. Surface decontaminated leaves of greenhouse plants as well as those excised from shoot cultures were chopped into 2-3 mm strips in most cases. But leaves with epidermal layers peeled off were also used except inBasella, Centella, Amaranthus, tomato, and chilli where peeling was not very efficient. Cotyledons of sunflower, niger, chilli seedlingsand placental tissue of tomato fruits were similarly chopped into segments for enzymatic digestion· The embryogenic callus of sandalwood was lightly squashed with a spatula and dropped into protoplast isolation solution. All tissues were weighed in sterilized petriplates for fresh weight determination before they were subjected to enzyme digestion. 2.3 Protoplast isolation, purification and culture The protoplast isolation solution consisted

of digesting enzymes, the osmoticum, a protecting chemical, CaCl2 (10 mM), all dissolved in distilled water. 2.3a Enzymes: The digesting enzymes used viz·, cellulase and pectinase (Celluclast andPectinex/SP249) are a gift from Novo Industri (Denmark)'s Regional Liaison Office, 101, 'Shah Sultan', Cunningham Road, Bangalore. These are liquids and are readily and completely miscible with waterat all concentrations which occur in normal usage. The optimal working conditions with these enzymes viz., temperature and pH are ideally suited for their use in protoplast isolation. 2.3b Osmotic conditions (stabilization): Osmotic stabilization was manipulated byadding to the protoplast isolation and culture solutions, mannitol as the sole osmoticum or mannitol in combination with either sorbitol or magnesium sulphate. For tobacco leaves, niger cotyledonsand sandalwood callus, mannitol alone was finally used as the osmoticum. Balancing of isolation and culture media to protoplast isotonic concentration was carried out for different species and explants individually using a cryoscopic osmometer (Osmomat 030 Gonotec) (tables 1 and 2). 2·3c Incubation for protoplast release: Protoplasts were isolated by incubating the various explants in filter sterilized isolation solution. Incubation was carried out in dark at temperatures of 25 ± 1°C initially. To determine the time required for complete release of protoplasts at incubation temperatures 25 ± 1°C, aliquots were microscopically examined at different time points for all the species/explants studied (table 1). To identify factors that might influence the efficiency in termsof protoplast release, rapidity and viability, incubation of tobaccoleaf tissue was carried out at different temperatures, at pH rangingfrom 5·6 to 6 and with different osmotic stabilizers used either singly or in combination. The quantity and the ratios of enzymes required for protoplast release were also determined.
2.3d Protoplast purification: The suspension of digested tissue in the isolation mixture was sieved through a nylon mesh of appropriatepore size to remove the debris and the filtrate was centrifuged at 100 g for 5 min to further separate the protoplasts from fine debristhat remained floating. The pellet of protoplasts was resuspended in5 ml of washing medium (table 2) and the process was repeated three times. The protoplasts in 2 ml of washing medium were then layered on 9 ml of flotation medium (table 2) in 15 ml capacity glass screw cap vials. The tubes were left undisturbed for 15 min and then centrifuged at 160 g for 10 min. The floating protoplast fraction that formed a clear band at the interphase of washing and flotation medium was recovered with a Pasteur pipette and was mixed with equalvolume of fresh wash medium. Protoplasts at this stage were allowed to equilibrate in the wash medium in dark for about 30 min before they were plated. The density and the yield of protoplasts were

estimated using a hemocytometer and the plating density adjusted. The competence and viability of protoplasts was determined by dye testing (the viability test with fluorescein diacetate (FDA)). Protoplast isolation by this method was compared with that obtained by a standard method (Sankara Rao and Gunasekari 1991). Leaf explants of tobacco and tomato axenic shoot cultures raised under identical conditions were digested by these two methods and the digestion time, and the yield at different temperatures were Determined. 2·3e Protoplast culture: Aliquots of purified protoplasts were plated in 2 ml of modified 8E liquid culture medium(Niedz et al 1985) supplemented with appropriate growth regulators and taken into 60 × 15 mm plastic or glass petriplates (table 2). The osmolarity of the culture medium was adjusted for each species separately by adding appropriate amounts of mannitol. The plating density was adjusted to obtain 2 × 104 protoplasts per ml of medium.The cultures were initially incubated in dark at 25 ± 2°C temperature for two weeks and then shifted to alternating light (of intensity 200 μΕm–2 s –1) and dark regimes of 14 and 10 h respectively. Samples were tested periodically for viability and wall regeneration· Further culturing of protoplasts to obtain cell colonies and plant regeneration was followed for N. tabacum cv Thompson applying the procedure described earlier by Sankara Rao andGunasekari (1991). All media were autoclaved for 15 min at 15 p.s.i.Growth regulators used were filter sterilized. All operations were carried out under aseptic conditions. 3. Results The results obtained from the various protoplast isolation experiments performedare summarized in table 1. With the method described now, tissues ofspecies explanted were most readily acted upon by the enzymes and microscopic examination carried out following digestion showed that the protoplast release was complete. All species investigated yielded protoplasts (figures 1, 2, 5 and 6). These species include herbaceous dicotyledonous plants, woody sandalwood and the mucilage-containing herbaceous vine, Basella. On an average, the incubation time required for complete release of protoplasts varied between 4 and 10 h depending on the species examined. Protoplast release from the explants of N. tabacum, Lycopersicon esculentum cultivars and Capsicum frutescens cultivars, particularly, needed only 4-6 h. Digestion of these explants carried out by another method which was adopted earlier by Sankara Rao and Gunasekari (1991) required a longer time (8-12 h) for protoplast release (table 1). The yield of protoplasts was also comparatively low. The Novo enzymes used in thepresent method maintained protoplast releasing ability at temperatures beyond 25°C. Increase of temperature during tissue incubation using these enzymes, in fact, aided tissue digestion further. The yield of protoplasts also increased slightly in N. tabacum cv Thompson (table 1). More than its beneficial effect on

protoplast yield, increased incubation temperature had reduced the time needed for complete release of protoplasts (table 1). The optimal conditions identified with the protocol were, an incubation temperature range between 28-30°C and a pH between 5·8 and 6. These were also conditions observed to be ideal for protoplast preparationfrom the plant species used in the experiments as the protoplast viability did not suffer the least. Using the present protocol, the yield of protoplasts derived from chopped explants as also from leaves with epidermal layers peeled off were comparable (table 1). Filtration of incubation mixture through nylon mesh reduced the amount of contaminating debris but only incompletely. That isolated protoplasts were free of cell walls was determined by microscopic observation. Virtually a pure population of intact protoplasts was readily isolated by flotation separation on sucrose. The yield data for each of the plant species used is shown in table 1. Washes and sedimentation helped further purification of the protoplast preparation. Protoplasts were most stable if allowed to equilibrate in the wash medium for about 30 min before plating. In aliquots of purified protoplasts tested for viability, a fairly good number werefound positive. Protoplasts plated in modified 8E medium formed cellwall readily (figures 3, 4, 6 and 8). The growth regulators and osmolarity of the culture medium were different for each species/cultivar investigated
Protoplasts of niger, tobacco cultivars, C. frutescens cv Punjab Jwala and L. esculentum cv Arkavikas divided within six to seven days of culture· In tobacco, numerous cell colonies became visible after six weeks of culture
Plants were regenerated for N. tabacum cv Thompson from the cell colonies (figures 11 and 12) as described earlier (Sankara Rao and Gunasekari 1991).
Most protoplast isolation protocols generally aim at higher yields, rapid recovery and regenerability of protoplasts so obtained. But all these objectives are seldom accomplished with any one method as there has been no demonstrated whole plant regeneration from protoplasts obtained by rapid preparations (Moore et al 1988; Hedrich et al 1985; Thirumala Devi et al 1992) and similarly, high-yielding protocols are rarely less simple procedures. Using the method described, we have overcome some of these problems. The protocol provides a simple and an easy-to-handle procedure that ensured satisfactory yields and quick recovery of viable protoplasts.

The method owes its efficiency to elevated digestion temperatures and a set of new enzymes synergistically acting on tissues at these temperatures. The Novo cellulase and pectinase used are stable at higher temperatures. The rate of enzymatic digestion also increased with increase in incubation temperature. The tissue incubation, however, was carried out at temperatures at which protoplast viability remained unaffected. The time required for protoplast release was less than it was observed with methods utilizing other enzymes (Sankara Rao and Gunasekari 1991), tested on the same tissueunder identical conditions. Thus, the extended periods required for enzymes to release protoplasts are avoided by this method and the protoplasts are protected from the deleterious effects of long-term exposure to digestion environment. The protocol is simple in that the various pretreatment steps such as preconditioning of donor plants, presoaking of explanted tissue, shaking during incubation, etc., were totally avoided. Generally different plant species require cell wall degrading enzymes from different sources. In the method presented, the same set of enzymes were found effective on many different and unrelated species equally well. Consistently, higher yields were obtained in all species investigated with an average of 6×l06 protoplasts per gram fresh weight of explanted tissue. Competence of these protoplasts for regeneration was ascertained by their survival and division in culture. Plant regeneration could not be worked out for all the plants investigatedas nutritional and other culture requirements are highly specific for individual species and cultivars. Nevertheless, cell wall regeneration and division were observed with protoplasts of Guizotiaabyssinica (Niger), N. glutinosa, C. frutescens cv Punjab Jwala, andL. esculentum cv Arkavikas, while further development leading to whole plant regeneration was demonstrated for protoplasts of N. tabacum cv Thompson. The method is routinely being used in our laboratory and is extended to isolate protoplasts from other plant species. The release of compounds that inhibit enzymatic cell wall digestion and consequent poor protoplast release from chopped explants (leaves etc.) has been reported (Butt 1985). In our experiments with the digestive enzymes used, chopped explants yielded as many protoplasts as leaves with epidermal layers peeled off would do. The process of wall digestion by these enzymes, obviously, is not impeded by inhibitory compounds exuding from chopped explants. The possibility that the Novo enzymes have a greater penetration ability through multiple layers of tightly packed cells in the explants, could also be a contributing factor tothe overall efficiency of these enzymes. They are supplied in a liquid state and are easily miscible with osmoticum mixture. The tissues are readily attacked by these enzymes and therefore

practices such as vacuum-infiltration or shaking to provide uniform exposure of all cells to the enzyme were totally eliminated.
Taking advantage of the extraordinary lytic properties of the enzymes under study, the exposure time can be optimized for a given species or cultivar to alleviate problems of over- or under-digestion of the protoplasts and higher yields of viable protoplastscan be obtained. We find that the method is simple and relatively rapid· It is advantageous over other methods in the overall ease of the procedure and reproducibility. The ultimate range of applicability of the method, however, has yet to be fully determinedbut may be limited in some plant species(http://www.ias.ac.in/jarch/jbiosci/20/645-655.pdf)
Somatic EmbryoThe process of formation of an embryo is called embryogenesis. Embryogenesis starts from a single embryogenic cell, that can be a zygote (the product of the fusion of an egg and a sperm during fertilization), or an undifferentiated callus cell. Embryos developing from zygotes are called zygotic embryos, while those derived from somatic cellsare called somatic embryos. During the embryonic development, the polar axis of the plant is established, domains that set up the organization of the plant body are defined, and the primary tissue and organ systems are delineated. Somatic embryogenesis is another important way to regenerate new plants in plant tissue culture.
Embryo development occurs through an exceptionally organized sequence of cell division, enlargement and differentiation. Zygotic and somatic embryos share the same gross pattern of development. Both types of embryos develop as passing through typical developmental stages . Embryo development is bipolar, having a shootpole and a radicular pole at opposite ends. Embryos are not organs because they are structurally independent from their parent body (i.e. they do not have a vascular system connecting them with their parent plant body).
Progenitor cell undergoes unequal division.

The physical, observable transition from a nonembryogenic cell to anembryogenic cell in somatic embryogenesis appears to occur when the progenitor cell undergoes an unequal division, resulting in a larger vacuolate cell and a small, densely cytoplasmic (embryogenic)cell . The embryogenic cell then either continues to divide irregularly to form a proembryonal complex or divides in a highly organized manner to form a somatic embryo.
Somatic embryos usually do not mature properly. Instead, due to environmental factors such as constant contact with inducing medium,somatic embryos often deviate from the normal developmental pattern by bypassing embryo maturation producing callus, undergoing direct secondary embryogenesis and/or germinating precociously. Somatic embryos growing from proembryonal complexes tend to develop asynchronously so that several stages are present in culture at any given time.(http://passel.unl.edu/pages/informationmodule.php?idinformationmodule=956783940&topicorder=4&maxto=10)
LIMITATIONS AND SOLUTIONSLIMITATIONS AND SOLUTIONSa) Somatic hybridization does not always produce plants that give fertile and visible seeds.b) There is genetic instability associated with protoplast culture.c) There are limitations in the selection methods of hybrids, as many of them are not efficient.d) Somatic hybridization between two diploids results in the formation of an amphidiploid which is not favourable therefore haploid protoplasts are recommended in somatic hybridization.e) It is not certain that a specific character will get expressed insomatic hybridization.f) Regenerated plants obtained from somatic hybridization are often variable due to somaclonal variations, chromosomal elimination, organelle segregation etc.
Somatic Embryo Chart

g) Protoplast fusion between different species/genus is easy, but the production of viable somatic hybrids is not always possible.(http://www.biotechnology4u.com/plant_biotechnology_applications_cell_tissue_culture.html)
(https://www.google.co.in/url?sa=t&rct=j&q=&esrc=s&source=web&cd=10&cad=rja&uact=8&ved=0CFUQFjAJ&url=http%3A%2F%2Fwww.scirp.org%2Fjournal%2FPaperDownload.aspx%3FpaperID%3D35851&ei=t5PxVI-GI4yjugS-i4LoBg&usg=AFQjCNEOVeQ2SGm5UNLSRVqlQBxp_Tkecw&bvm=bv.87269000,d.c2E)
PLAYERS IN THE FIELDPLAYERS IN THE FIELDPhytowelt (Head office: Kölsumer Weg 33 D-41334 Nettetal P +49.2162.77859 F +49.2162.89215 www.phytowelt.com) has established optimal systems for many plant species, including even difficult ones like grasses, cereals and legumes.Phytowelt already applied the SH technology for many different applications.
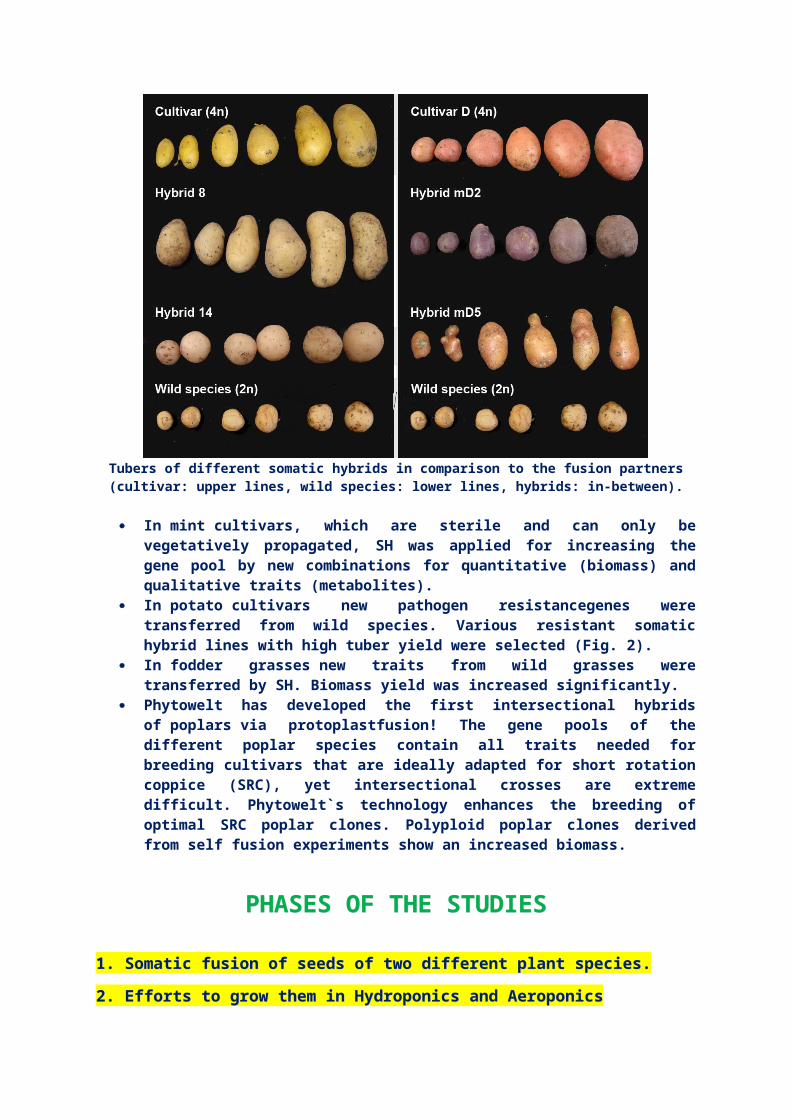
Tubers of different somatic hybrids in comparison to the fusion partners(cultivar: upper lines, wild species: lower lines, hybrids: in-between).
In mint cultivars, which are sterile and can only bevegetatively propagated, SH was applied for increasing thegene pool by new combinations for quantitative (biomass) andqualitative traits (metabolites).
In potato cultivars new pathogen resistancegenes weretransferred from wild species. Various resistant somatichybrid lines with high tuber yield were selected (Fig. 2).
In fodder grasses new traits from wild grasses weretransferred by SH. Biomass yield was increased significantly.
Phytowelt has developed the first intersectional hybridsof poplars via protoplastfusion! The gene pools of thedifferent poplar species contain all traits needed forbreeding cultivars that are ideally adapted for short rotationcoppice (SRC), yet intersectional crosses are extremedifficult. Phytowelt`s technology enhances the breeding ofoptimal SRC poplar clones. Polyploid poplar clones derivedfrom self fusion experiments show an increased biomass.
PHASES OF THE STUDIESPHASES OF THE STUDIES
1. Somatic fusion of seeds of two different plant species.1. Somatic fusion of seeds of two different plant species.
2. Efforts to grow them in Hydroponics and Aeroponics2. Efforts to grow them in Hydroponics and Aeroponics

3. Efforts to instill components to improve factors like 3. Efforts to instill components to improve factors like
colour, flavor, taste, appearance, size, ripening time, colour, flavor, taste, appearance, size, ripening time,
control of vegetative and reproductive phasescontrol of vegetative and reproductive phases
IDENTIFICATION OF PLANTS TO BE MERGEDIDENTIFICATION OF PLANTS TO BE MERGED
Criteria to be followed:Criteria to be followed:
1. Both species may prefer similar climatic conditions.1. Both species may prefer similar climatic conditions.
2. Both species may have same season and/or crop time or 2. Both species may have same season and/or crop time or
cycle.cycle.
3. One species is to give crop above the ground and the second3. One species is to give crop above the ground and the second
one should give crop below the ground.one should give crop below the ground.
4. Coats of the Seeds of both the species either should have 4. Coats of the Seeds of both the species either should have
no ANF and no Antibiotics.no ANF and no Antibiotics.
5. One species should produce yield containing more 5. One species should produce yield containing more
carbohydrates or sugars while the other one should yield othercarbohydrates or sugars while the other one should yield other
nutrients like FAT so a comprehensive nutrition helps to nutrients like FAT so a comprehensive nutrition helps to
achieve better results in both the crops.achieve better results in both the crops.
6. Sizes of the seed should be more accommodative for the 6. Sizes of the seed should be more accommodative for the
somatic fusionsomatic fusion
7. Seed coats should be more accommodative for the somatic 7. Seed coats should be more accommodative for the somatic
fusionfusion
PROBABLE COMBINATIONS THAT MAY BE CHOOSENPROBABLE COMBINATIONS THAT MAY BE CHOOSEN1. Blackgram (1. Blackgram (Vigna MungoVigna Mungo) ) + Peanut ( + Peanut (Arachis hypogaeaArachis hypogaea))
2. Soy (2. Soy (Glycine maxGlycine max ) + Onion ( ) + Onion (AlliumAllium cepacepa))
3. Pudina (3. Pudina (Mentha ArvensisMentha Arvensis ) + Carrot ( ) + Carrot (Daucus carotaDaucus carota))
4. Palak (4. Palak (Spinacia oleracea)Spinacia oleracea) + Muli ( + Muli (RaphanusRaphanus sativus)sativus)

5. Coriander (5. Coriander (Coriandrum sativumCoriandrum sativum ) + Carrot ( ) + Carrot (Daucus carotaDaucus carota))
6. Chana(6. Chana(Cicer arietinumCicer arietinum) ) + Carrot ( + Carrot (Daucus carotaDaucus carota))
7. Guar (7. Guar (Cyamopsis tetragonolobaCyamopsis tetragonoloba ) + Peanut ( ) + Peanut (Arachis hypogaeaArachis hypogaea))
PROTOCOLSPROTOCOLSProtoplast Isolation:Isolation of protoplasts (Fig. 8.9) is readily achieved by treating cells/tissues with a suitable mixture of cell wall degrading enzymes. Usually, a mixture of pectinase or macerozyme (0.1-1.9%) and cellulose (1-2%) is appropriate for most plant materials. Hemicellulase may be necessary for some tissues. Generally crude commercial preparations of enzymes arc used.The pH of enzyme solution is adjusted between 4.7 and 8.0 and the temperature is kept at 25-30°C. The osmotic concentration of enzyme mixture and of subsequent media is elevated (usually, by adding 500-800 m mol l-1 sorbitol or mannitol) to stabilize the protoplasts and to prevent them from bursting. Usually, 50-100 m mol l-1 CaCl2 is added to the osmoticum as it improves plasma membrane stability. Thecells and tissues are incubated in the enzyme mixture for few to several (generally, 16-18) hours; naked protoplasts devoid of cell wall are gradually released in the enzyme mixture.Protoplasts have been isolated from virtually all plant parts, but leaf mesophyll is the most preferred tissue, at least in case of dicots, for this purpose. In general, fully expanded leaves are surface-sterilized, their lower epidermis is peeled off with a pair of forceps and the peeled areas are cut into small (Ca. 1 cm2) pieces with a scalpel and suspended in the enzyme mixture.

When epidermis cannot be peeled, leaf may be cut into Ca. 1 mm2 pieces and treated with the enzyme mixture; vacuum infiltration may be used to facilitate the entry of enzymes into the tissues. After the period of incubation, protoplasts are washed with a suitable washing medium in order to remove the enzymes and the debris.The protoplasts may be cultured in a suitable medium in a variety ofways: (i) Bergmann’s plating technique (in agar medium), (ii) in a thin layer of liquid medium or (iii) in small microdrops of 50-100 µl. Protoplasts readily regenerate cell wall (within 2-4 days) and undergo mitosis to form macroscopic colonies, which can be induced to regenerate whole plants. The conditions for isolation and cultureof protoplasts and regeneration of complete plants have been standardized for a large number of plant species, but cereals still present some problems.Generally, MS and B5 media, and their modifications are used for protoplast culture. The media are supplemented with a suitable osmoticum and, almost always, with an auxin and a cytokinin, their types and concentrations depending mainly on the plant species.After 7-10 days of culture, protoplasts regenerate cell wall, and the osmolarity of medium is gradually reduced to that of normal medium. The macroscopic colonies are transferred onto normal tissue culture media. Protoplasts are very sensitive to light; therefore, they are cultured in diffuse light or dark for the first 4-7 days.
2. Protoplast Fusion:The techniques for protoplast fusion are pretty well refined and highly effective for almost all the systems. A number of strategies have been used to induce fusion between protoplasts of different strains/species; of these the following three (Fig. 8.10) have been relatively more successful.Protoplasts of desired strains/species are mixed in almost equal proportion; generally, they are mixed while still suspended in the enzyme mixture. The protoplast mixture is then subjected to high pH (10.5) and high Ca2+ concentration (50 m mol l-1) at 37°C for about 30min (high pH-high Ca2+ treatment). This technique is quite suitable for some species, while for some others it may be toxic.

Polyethylene glycol (PEG) induced protoplast fusion is the most commonly used as it induces reproducible high frequency fusion accompanied with low toxicity to most cell types. The protoplast mixture is treated with 28-50% PEG (MW 1,500-6,000) for 15-30 min, followed by gradual washing of the protoplasts to remove PEG; protoplast fusion occurs during the washing.The washing medium may be alkaline (pH 9-10) and contain a high Ca2+ ion concentration (50 m mol l-1); this approach is a combination of PEG and high pH-high Ca2+ treatments, and is usually more effective than either treatment alone.PEG is negatively charged and may bind to cation like Ca2+, which, inturn, may bind to the negatively charged molecules present in plasmalemma; they can also bind to cationic molecules of plasma membrane.During the washing process, PEG molecules may pull out the plasma lemma components bound to them. This would disturb plasma lemma organisation and may lead to the fusion of protoplasts located closeto each other (Fig. 8.11).

The above fusion techniques are nonselective in that they induce fusion between any two or more protoplasts. A more selective and less drastic approach is the electrofusion technique, which utilizeslow voltage nonuniform alternating electric current pulses to bring the protoplasts in close contact (Fig. 8.12). Fusion of protoplasts is brought about by a short pulse of high voltage.The duration of high voltage is a few microseconds, and the voltagesranges from 500 to 1,000 V/cm. The high voltage creates transient disturbances in the organisation of plasma lemma, which leads to thefusion of neighbouring protoplasts. The entire operation is carried out manually in specially designed equipment, called electroporator.
Electrofusion has been mostly used with the members of Solanaceae often with very high rate (over 50%) of fusion. This approach induces general fusion among protoplats and there is no control on the type of protoplasts entering fusion. In a modification of electrofusion, protoplast pairs are individually transferred into microfusion chambers with the help of a micromanipulator set up.Thus each microfusion chamber has one pair of protoplasts, which areinduced to fuse by a single or multiple, negative DC-pulse of 800 to1,800 V /cm for 50 microseconds — after mutual dielectrophoresis (1 MHz; 65-80 V/cm). This technique is called microelectrofusion; it leads to highly specific and nearly 100% pairwise fusion. Many workers feel that this fusion technique is more desirable than the others for a number of important reasons.

3. Selection of Hybrid Cells:The protoplast suspension recovered after a treatment with a fusion inducing agent (fusogen) consists of the following cell types: (i) unfused protoplasts of the two species/strains, (ii) products of fusion between two or more protoplasts of the same species (homokaryons), and (iii) ‘hybrid’ protoplasts produced by fusion between one (or more) protoplast(s) of each of the two species (heterokaryons) (Fig. 8.13).
In somatic hybridization experiments, only the heterokaryotic or ‘hybrid’ protoplasts, particularly those resulting from fusion

between one protoplast of each of the two fusion partners, are of interest.However, they form only a small proportion of the population (usually 0.5-10%). Therefore, an effective strategy has to be employed for their identification and isolation. This step is calledthe selection of hybrid cells, is the most critical, and is still anactive area of investigation.A number of strategies have been used for the selection of hybrid protoplasts. (1) Some visual markers, e.g., pigmentation, of the parental protoplasts may be used for the identification of hybrid cells under a microscope; these are then mechanically isolated and cultured.For example, the protoplasts of one species may be green and vacuolated (derived from mesophyll cells), while those of the other may be non-vacuolated and non-green (obtained from cell cultures). Where such features are not available, the protoplasts of two parental species may be separately labelled with different fluorescent agents.
This would allow the microscopic identification of hybrid protoplasts from the parental ones. This approach can even be adapted to automatic cell sorting permitting the recovery of large numbers of heterokaryons in a very short time. This approach is, however, time consuming, and requires considerable skill and effort.
Several workers have attempted to devise systems, which specificallyselect for hybrid cells. In simple words, (2) these systems exploit some properties (usually, deficiencies) of the parental species, which are not expressed in the hybrid cells due to complementation between their genetic systems.These properties may be sensitivity to culture medium constituents, antimetabolites, temperature, etc., inability to produce an essential biochemical (auxotrophic mutants), chlorophyll or some other pigmentation, etc. These properties may be naturally present in the parental species or may be artificially induced through mutagenesis.
The following example should be enough to clearly bring out the essential features of the complementation approach. Protoplasts of Petunia hybrida form calli on the MS medium, while those of P. parodii produce only small colonies.
Further, actinomycin D (1 µg ml-1) inhibits cell division of P. hybrida protoplasts, but it has no effect on those of P. parodii. Thus protoplasts of both these Petunia species fail to produce

macroscopic colonies (calli) on MS medium supplemented with 1 µg ml-
1 actinomycin D.However, their hybrid cells (P. hybrida + P. parodii; it may be noted that somatic hybrids are denoted by a + sign as against the sexual hybrids being designated by a X symbol) divide normally on this medium to produce macroscopic colonies.
This selection strategy exploits those natural properties of the twoparental species, which show complementation in the hybrid cells and, at the same time, permit their selection. These strategies are simple, highly effective and the least demanding, but their application is drastically limited by the nonavailability of suitable properties (both natural and induced) in most of the parental species of interest to the experimenters.
A variation of the complementation approach uses selectable markers.In this approach, (3) different selectable markers like antibiotic and herbicide resistance are engineered into the two fusion partners. The hybrid cells, in such cases, are resistant to both theconcerned selection agents, while the parental cells will be sensitive to one and the other selection agent.
A more general and widely applicable strategy, but demanding more work than the previous approach, is (4) to culture the entire protoplast population without applying any selection for the hybrid cells. All the types of protoplasts form calli; the hybrid calli arelater identified on the basis of callus morphology, chromosome constitution, protein and enzyme banding patterns, etc.In some cases, the identification may be delayed till plants are regenerated. In such an approach, it will be desirable to culture the protoplasts in very low densities since neighbouring colonies are likely to fuse at higher densities; ideally, they should be cultured in microdrops, each drop containing but a single cell. Manyworkers tend to favour this approach since it does not depend the presence of appropriate but difficult to find markers in the parental species.
4. Regeneration of Hybrid Plants:Once hybrid calli are obtained, plants are induced to regenerate from them since this is a prerequisite for their exploitation in crop improvement. Further, the hybrid plants must be at least partially fertile, in addition to having some useful property, to beof any use in breeding schemes. The culture techniques have been refined to a state where plant regeneration has been obtained in a number of somatic hybrids (Table 8.5).

Table 8.5. A list of some distant somatic hybrid plants:
But even today, it has not been possible to recover hybrid plants and/or calli from a number of somatic combinations; this phenomenon is called ‘somatic incompatibility’. The reasons for somatic incompatibility are not clearly understood. The somatic hybrids are of the following two types: (1) symmetric and (2) asymmetric hybrids.
Symmetric Hybrids:Some somatic hybrid plants retain the full or nearly full somatic complements of the two parental species; these are called symmetric hybrids. Such hybrids provide unique opportunities for synthesizing novel species, which may be of theoretical and/or practical interest. Frequently, somatic hybrids between distantly related sexually incompatible species are sterile, precluding their incorporation into a breeding programme.
This may be circumvented by producing 3n somatic hybrids by fusing somatic (2n) cells of one species with haploid (n) cells of the other species; such 3n plants may be expected to be partially fertile. These somatic hybrids can now be used in breeding programmes for limited gene/chromosome introgression from the species contributing the haploid protoplast.
A possible approach to the improvement of apparently useless somatichybrids, e.g., nonflowering somatic hybrid Daucus carota + Aegopodium podagraria, is to fuse protoplasts from the hybrid with those of one of the parental species. The fusion of a somatic hybridprotoplast with that from one of its parents is called somatic back hybridization.When protoplasts from the above somatic hybrid were fused with carrot protoplasts, the resulting somatic hybrid produced flowers. Such hybrids can now be ordered into breeding programmes with the aim of gene/chromosome introgression.

Asymmetric Hybrids:Many somatic hybrids exhibit the full somatic complement of one parental species, while all or nearly all chromosomes of the other parental species are lost during the preceding mitotic divisions; such hybrids are referred to as asymmetric hybrids.
The available evidence suggests that such hybrids are likely to showa limited introgression of chromosome segments from the eliminated genome(s) due to drastically enhanced chromosomal aberrations and/ormitotic crossing over in vitro.
Asymmetric hybrids can be obtained even from those combinations, which normally produce symmetric hybrids by the following approach: protoplasts of one of the parental species are irradiated with a suitable dose of X-rays or gamma-rays to induce extensive chromosomebreakage.In such cases, chromosome segment introgression may be markedly enhanced. It may be pointed out that asymmetric hybrids are essentially cytoplasmic hybrids or cybrids, except for the introgressed genes.
Fate of Plasma-genes:In contrast to sexual hybrid cells, i.e., zygotes, which contain thecytoplasmic genes (plasmon) from the female parent only, somatic hybrid cells, contain cytoplasmic complements from both the parentalspecies.
The cytoplasmic genes (generally studied in terms of chloroplast types or chloroplast DNA, cp-DNA mitochondrial DNA, mtDNA) appear tobe distributed randomly during the mitotic cell divisions. As a result, some cells receive chloroplasts of one parental species, some others of the other species and a small proportion retains the chloroplasts of both the species.
This is reflected in the plants regenerated from these cells. The same applies to mitochondria as well. In addition, the distribution of chloroplasts is independent from that of mitochondria.Therefore, a somatic hybrid plant may contain chloroplasts from one parental species and mitochondria from the other fusion parent. There is considerable evidence that the genomes of both chloroplastsand mitochondria, particularly the latter, undergo recombination in the hybrid cells; this produces recombinant organelles in the progeny.

5. Cybrids:Cybrids or cytoplasmic hybrids are cells or plants containing nucleus of one species but cytoplasm from both the parental species.They are produced in variable frequencies in normal protoplast fusion experiments due to one of the following: (i) fusion of a normal protoplast of one species with an enucleate protoplast or a protoplast having an inactivated nucleus of the other species, (ii) elimination of the nucleus of one species from a normal heterokaryon, or (iii) gradual elimination of the chromosomes of onespecies from a hybrid cell during the subsequent mitotic divisions. Cybrids may be produced in relatively high frequency by (i ) irradiating (with X-rays or gamma-rays) the protoplasts of one species prior to fusion in order to inactivate their nuclei, or (ii)by preparing enucleate protoplasts (cytoplasts) of one species and fusing them with normal protoplasts of the other species.The objective of cybrid production is to combine the cytoplasmic genes of one species with the nuclear and cytoplasmic genes of another species. But the mitotic segregation of plasmagenes, as evidenced by the distribution of chloroplasts, leads to the recoveryof plants having plasmagenes of one or the other species only; only a small proportion of the plants remain ‘cybrid’, which would further segregate into the two parental types.
Cybrids provide the following unique opportunities: (i) transfer of plasmagenes of one species into the nuclear background of another species in a single generation, and even in (ii) sexually incompatible combinations, (iii) recovery of recombinants between the parental mitochondrial or chloroplast DNAs (genomes), and (iv) production of a wide variety of combinations of the parental and recombinant chloroplasts with the parental or recombinant mitochondria. When cybrids are produced by irradiating the protoplasts of one species prior to fusion, they provide the additional opportunity for (v) the recovery of chromosome segment introgressions from the lost genome in combination with variations in the plasmon. The cybrid approach has been used for the transfer of cytoplasmic male sterility from N. tabacum to N. sylvestris, fromP. hybrida to P. axillaris, etc. (vi) In addition, mitochondria fromone parental species may be combined with the chloroplasts of the other parental species.
(http://www.yourarticlelibrary.com/biotechnology/plant-tissues/somatic-hybridization-of-hybrid-plants-explained-with-diagram/33226/

RESULTSRESULTS
DISCUSSIONDISCUSSION
IPIPThe UPOV convention is the basis for legislation on plant breeders’ rights (PBR) in currently 59 single countries and the European Union. The chapter explains - on the basis of the UPOV convention - who is entitled to plant breeders’ rights and the scope of such rights including its conditions, restrictions, and exemptions. Furthermore, the document details keywords of the legislation to enable applicants for plant breeders’ rights to understand the legalprocedure of the granting process. The grant of PBR is independent from regulations on the production or commercialization of a given variety.
EXPLORING MARKETEERSEXPLORING MARKETEERS
STATUTORY REGULATIONS TO BE TAKEN CARESTATUTORY REGULATIONS TO BE TAKEN CAREWhen comparing somatic hybridization to transgenic approaches, somatic hybridization enables broadening of the germplasm base, allows the transfer of uncloned multiple genes and generates products that are not subjected to the same legal regulations as transgenic lines [J. W. Grosser and F. G. Gmitter, “2004 SIVB Congress Symposium Proceedings of Thinking Outside the Cell: Applications of Somatic Hybridization and Cybridization in Crop Improvement, with Citrus as a Model,” In Vitro Cellular & Developmental Biology Plant, Vol. 41, No. 3, 2005, pp. 220-225. doi:10.1079/IVP2004634 J. Grosser and F. Gmitter, “Protoplast Fusion for Production of Tetraploids and Triploids: Applications for Scion and Rootstock Breeding in Citrus,” Plant Cell, Tissue and Organ Culture, Vol. 104,No. 3, 2011, pp. 343-357. doi:10.1007/s11240-010-9823-4
Also, it transfers both mono- and polygenic traits [R. Thieme, U. Darsow, L. Rakosy-Tican, Z. Kang, T. Gavrilenko, O. Antonova, U. Heimbach and T. Thieme, “Use of Somatic Hybridization

to Transfer Resistance to Late Blight and Potato Virus Y (PVY) into Cultivated Potato,” Plant Breeding and Seed Science, Vol. 50, No. 1,2004, pp. 113-118.].
Regulation of GMOs The overseeing of testing, use and commercialisation of GMOs — whether plants, animals or microorganisms — requires a special regulation system. This system establishes the legal and institutional framework for the control of potential negative effects of GMOs on the environment or human and animal health (Băbeanu, 2003). In the US, transgenic plants are only introduced into the environment or on the market following approval from the following governmental agencies responsible for environmental, and human and animal health protection: 1. US Department for Agriculture (USDA), 2. Environmental Protection Agency (EPA), 3. Food and Drug Administration (FDA). In the US and Canada, transgenic plants are grown and used for humanand animal food, and separate storage and labelling are not mandatory. Since 1990, in the European Union, special legislation has been drawn up, enhanced and extended, with the purpose of providing environmental and human health protection, and creating a common market in the field of biotechnology. Thus: - EU Directive No. 219/1990 (amended by Directive No. 81/1998) regulates the contained use of genetically modified microorganisms (for research and commercialisation), - EU Directive No. 220/1990, concerning the deliberate release of genetically modified organisms into the environment, was the main initiative taken by the EU, and was subsequently supplemented by several Commission Decisions (623, 811,812, 813/2003), - EU Directive No. 18/2001 regulates the deliberate release of genetically modified organisms into the environment. ThisDirective repealed Directive No. 220/1990. Having come into force on17 October 2002, Directive 18/2001 both updates and consolidates 15 existing regulations. This Directive also deals with mandatory information to the public, the long-term monitoring of effects, labelling and traceability, in all stages of GMO introduction on themarket. Two other acts have been adopted and published in the Official Journal of the European Communities, with respect to the Community system of GMO traceability, the labelling of genetically modified food and fodder, and the continuous procedure of authorisation or introduction of GMOs in the environment as food or fodder: - EC Regulation No. 1829/2003 (22 September 2003) on genetically modified food and fodder, and - EC Regulation No. 1830/2003 (22 September 2003) on the traceability and labelling of

genetically modified organisms, and GMO-based food and fodder. Theseregulations amended EU Directive No. 18/2001.
WEB REFERENCESWEB REFERENCES
REFERENCESREFERENCES1. Greene, David (27 September 2013). "TomTato Is The Latest
Wonderplant". NPR News. Retrieved 30 September 2013.2. a b Renneberg, Reinhard (2008). Biotechnology for Beginners.
Elsevier. p. 210. ISBN 9780123735812.3. "How to create a pomato plant". Retrieved 29 May 2013.4. "Pomato Plants". The Guru (36). 2010. Retrieved 29 May 2013.5. a b Jabr, Ferris. "The Science of Pomato Plants and Fruit Salad
Trees". Scientific America. Retrieved 29 May 2013.6. "Kenyan farmers produce ‘pomato’ plants to improve land use".
Retrieved 29 May 2013.7. "Prison grows unique ‘pomato’ to fight hunger".
www.businessdailyafrica.com. Retrieved 29 May 2013.8. Jude Gillies. "Potato Tom opens fresh doors". Stuff.co.nz. Retrieved
2013-09-30.9. Hall, John (27 September 2013). "The TomTato: Plant which produces
both potatoes and tomatoes launched in UK". The Independent. Retrieved 30 September 2013.
10. Wilkes, David (25 September 2013). "The TomTato... or how you can make ketchup AND chips from the same plant! |". Mail Online. Retrieved 30 September 2013.
11. Bellincampi D and Morpugo G 1987 Conditioning factor affecting growth in plant cells in culture; Plant Sci. 51 83-91
12. Butt A D 1985 A general method for the high-yielding isolation of mesophyll protoplasts from deciduous tree species; Plant Sci. 42 55-59
13. Cocking Ε C 1960 A method for the isolation of plant protoplasts and vacuoles; Nature (London) 187 927-929
14. d'Utra Vaz F B, Slamet I H, Khatum A, Cocking Ε C and Power J Β1992 Protoplast culture in high molecular oxygen atmosphere; Plant Cell Rep. 11 416-418
15. Hedrich R, Raschke Κ and Stitt Μ 1985 A role for Fructose 2,6-Bisphosphate in regulating carbohydrate metabolism in guard cells; Plant Physiol. 79 977-982
16. Kao Κ Ν and Michayluk Μ Κ 1975 Nutritional requirements for growth of Vinca hajastana cells and protoplasts at a very low population density in liquid media; Planta 126 105-110
17. Kyozuka J, Hayashi Υ and Shinomoto Κ 1987 High frequency plant regeneration from rice protoplasts by novel nurse culture methods; Mol. Gen.Genet. 206 408-413
18. Lee L, Schroll R E, Grimes Η D and Hodges Τ Κ 1990 Plant regeneration from indica rice (Oryza sativa L.) protoplasts; Planta

178 325-333 Maheshwari S C, Gill R, Maheshwari Ν and Gharyal Ρ Κ 1986The isolation and culture of protoplasts; in Differentiation of protoplasts and transformed plant cells (eds) J Reinert and Binding (New York: Springer-Verlag) pp 20-48
19. Mei-Lei Μ C T, Reitveld Ε Μ, Van Marrewijk G A M and Kool A J 1987 Regeneration of leaf mesophyll protoplasts of tomato cultivars (L. esculentum): factors important for efficient protoplast culture and plant regeneration; Plant Cell Rep. 6 172-175
20. Moore Β D, Monson R K, Ku Μ S Β and Edwards G Ε 1988 Activitiesof principal photosynthetic and photorespiratory enzymes in leaf mesophyll and bundle sheath protoplasts from the C3-C4 intermediate Flavaria ramosissima; Plant Cell Physiol. 29 999-1006
21. Niedz R P, Rutter S M, Handley L W and Sink Κ C 1985 Plant regeneration from leaf protoplasts of six tomato cultivars; Plant Sci. 39 199-204
22. Sankara Rao Κ and Srikantha Τ 1986 A simple method for the preparation of mesophyll protoplasts; in Proceedings of the VI International Congress of Plant Tissue and Cell Culture, Minnesota, p37
23. Sankara Rao Κ and Gunasekari Κ 1991 Control of morphogenesis intobacco protoplast cultures: Organogenesis Vs. embryogenesis; Indian J. Biochem. Biophys. 28 467-471
24. Shahin Ε A 1985 Totipotency of tomato protoplasts; Theor. Appl.Genet. 69 235-240 Thirumala Devi M, Vani T, Malla Reddy Μ and Raghavendra A S 1992 Rapid isolation of mesophyll and guard cell protoplasts from pea and maize leaves; Indian J. Exp. Biol. 30 424-428




















