
Doublecortin interacts with the ubiquitin protease DFFRX, which associates with microtubules in...
12
Doublecortin interacts with the ubiquitin protease DFFRX, which associates with microtubules in neuronal processes GaJlle Friocourt, a Caroline Kappeler, a Yoann Saillour, a Fabien Fauchereau, a Manuel S. Rodriguez, b Nadia Bahi, a Marie-Claude Vinet, a Philippe Chafey, a Karine Poirier, a Shinichiro Taya, c Stephen A. Wood, d,e Catherine Dargemont, b Fiona Francis, a, * and Jamel Chelly a a Laboratoire de Ge ´ne ´tique et Physiopathologie des Retards Mentaux, GDPM, Institut Cochin, 75014 Paris, France b Equipe de transport nucle ´ocytoplasmique, Institut Jacques Monod, CNRS UMR 7592, 75005 Paris, France c Department of Cell pharmacology, Graduate school of Medecine, Nagoya University, Nagoya 466-8550, Japan d Child Health Research Institute, North Adelaide SA 5006, Australia e Centre for the Molecular Genetics of Development, University of Adelaide, SA 5005, Australia Received 28 May 2004; revised 7 September 2004; accepted 13 September 2004 Available online 11 November 2004 Doublecortin (DCX) is a microtubule-associated protein involved in neuronal migration, which causes X-linked lissencephaly and subcort- ical laminar heterotopia (SCLH) when mutated. Here we show that DCX interacts with the ubiquitin-specific protease Drosophila fat facets related on X chromosome (DFFRX). This interaction was confirmed by targeted mutagenesis, colocalization, and immunoprecipitation studies. DFFRX is thought to deubiquitinate specific substrates includ- ing B-catenin, preventing their degradation by the proteasome. Interest- ingly, unlike B-catenin, no ubiquitinated forms of DCX could be detected, and indeed we show that DCX interacts with a novel recognition domain in DFFRX, located outside of its catalytic site. We also show that DFFRX associates with microtubules at specific subcellular compartments, including those enriched in DCX. These results thus suggest that in addition to vesicular trafficking, DCX may play a role in the regulation of cell adhesion via its interaction with DFFRX in migrating and differentiating neurons. D 2004 Elsevier Inc. All rights reserved. Introduction The complex laminar structures of the mature cerebral cortex arise through a series of carefully regulated developmental events, including neurogenesis, neuronal migration, and differentiation, as well as the formation of synaptic connections. During cortico- genesis, postmitotic neurons exit the neuroepithelium and migrate to the cortical plate where they eventually form the mature cortical layers. They settle in six layers sequentially formed from the deepest to the most superficial (binside–outQ) so that each new wave of neurons migrates past the previously formed ones (Hatten, 1999). Neuronal migration is thus a key step in cortical layer organization. Neither the molecular events that drive migrating neuronal cells to their final destinations nor the environmental cues responsible for the regulation of their spatial position are fully understood. However, significant contributions to the dissection of several pathways involved in these processes have been provided through the genetic analysis of disorders associated with brain cortical dysgeneses. Such disorders including type-I lissencephaly and subcortical laminar heterotopia (SCLH), which result from muta- tions in the LIS1 (Reiner et al., 1993) and doublecortin genes (also called DBCN or DCX) (des Portes et al., 1998; Gleeson et al., 1998), are characterized by a dramatic disorganization of neocortical layers and gyral abnormalities. In addition to LIS1 and DCX, mutations in cdk5 (Oshima et al., 1996), p35 (Chae et al., 1997), filamin 1 (Fox et al., 1998), reelin (D’Arcangelo et al., 1995; Hong et al., 2000), mDab1 (Howell et al., 1997; Sheldon et al., 1997; Ware et al., 1997), VLDLR and ApoER2 (Trommsdorff et al., 1999), POMGnT1 (Michele et al., 2002; Moore et al., 2002) and POMT1 (Beltran- Valero de Bernabe et al., 2002), and ARX (Kitamura et al., 2002) have been found to lead to specific phenotypes associated with disrupted corticogenesis, either in human or in mouse. One signaling pathway is represented by Reelin, mDab1, ApoER2, and VLDLR, which are potentially involved in the control of the final position of neurons and function through a linear pathway at the plasma membrane (D’Arcangelo et al., 1999; Hiesberger et al., 1999; Trommsdorff et al., 1999). Signaling pathways regulated by Cdk5 and its specific activator p35 were shown to be involved in neuronal migration and several other processes such as neurite outgrowth, vesicular trafficking, and neurodegeneration (Smith and Tsai, 2002; Tan et al., 2003). The Cdk5/p35 kinase has been shown to regulate N-cadherin- mediated adhesion, which also requires h-catenin, isolated as a 1044-7431/$ - see front matter D 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.mcn.2004.09.005 * Corresponding author. Laboratoire de Ge ´ne ´tique et Physiopathologie des retards mentaux, GDPM, Institut Cochin, 24, rue du Faubourg Saint Jacques, 75014 Paris, France. Fax: +33 1 44 41 24 21. E-mail address: [email protected] (F. Francis). Available online on ScienceDirect (www.sciencedirect.com). www.elsevier.com/locate/ymcne Mol. Cell. Neurosci. 28 (2005) 153 – 164
-
Upload
univ-paris5 -
Category
Documents
-
view
1 -
download
0
Transcript of Doublecortin interacts with the ubiquitin protease DFFRX, which associates with microtubules in...

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 28 (2005) 153–164
Doublecortin interacts with the ubiquitin protease DFFRX,
which associates with microtubules in neuronal processes
GaJlle Friocourt,a Caroline Kappeler,a Yoann Saillour,a Fabien Fauchereau,aManuel S. Rodriguez,b
Nadia Bahi,a Marie-Claude Vinet,a Philippe Chafey,a Karine Poirier,a Shinichiro Taya,c
Stephen A. Wood,d,e Catherine Dargemont,b Fiona Francis,a,* and Jamel Chellya
aLaboratoire de Genetique et Physiopathologie des Retards Mentaux, GDPM, Institut Cochin, 75014 Paris, FrancebEquipe de transport nucleocytoplasmique, Institut Jacques Monod, CNRS UMR 7592, 75005 Paris, FrancecDepartment of Cell pharmacology, Graduate school of Medecine, Nagoya University, Nagoya 466-8550, JapandChild Health Research Institute, North Adelaide SA 5006, AustraliaeCentre for the Molecular Genetics of Development, University of Adelaide, SA 5005, Australia
Received 28 May 2004; revised 7 September 2004; accepted 13 September 2004Available online 11 November 2004
Doublecortin (DCX) is a microtubule-associated protein involved in
neuronal migration, which causes X-linked lissencephaly and subcort-
ical laminar heterotopia (SCLH)whenmutated. Here we show that DCX
interacts with the ubiquitin-specific protease Drosophila fat facets
related on X chromosome (DFFRX). This interaction was confirmed
by targeted mutagenesis, colocalization, and immunoprecipitation
studies. DFFRX is thought to deubiquitinate specific substrates includ-
ing B-catenin, preventing their degradation by the proteasome. Interest-
ingly, unlike B-catenin, no ubiquitinated forms of DCX could be
detected, and indeed we show that DCX interacts with a novel
recognition domain in DFFRX, located outside of its catalytic site. We
also show that DFFRX associates with microtubules at specific
subcellular compartments, including those enriched in DCX. These
results thus suggest that in addition to vesicular trafficking, DCX may
play a role in the regulation of cell adhesion via its interaction with
DFFRX in migrating and differentiating neurons.
D 2004 Elsevier Inc. All rights reserved.
Introduction
The complex laminar structures of the mature cerebral cortex
arise through a series of carefully regulated developmental events,
including neurogenesis, neuronal migration, and differentiation, as
well as the formation of synaptic connections. During cortico-
genesis, postmitotic neurons exit the neuroepithelium and migrate to
the cortical plate where they eventually form the mature cortical
layers. They settle in six layers sequentially formed from the deepest
1044-7431/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.mcn.2004.09.005
* Corresponding author. Laboratoire de Genetique et Physiopathologie
des retards mentaux, GDPM, Institut Cochin, 24, rue du Faubourg Saint
Jacques, 75014 Paris, France. Fax: +33 1 44 41 24 21.
E-mail address: [email protected] (F. Francis).
Available online on ScienceDirect (www.sciencedirect.com).
to the most superficial (binside–outQ) so that each new wave of
neurons migrates past the previously formed ones (Hatten, 1999).
Neuronal migration is thus a key step in cortical layer organization.
Neither the molecular events that drive migrating neuronal cells to
their final destinations nor the environmental cues responsible for
the regulation of their spatial position are fully understood.
However, significant contributions to the dissection of several
pathways involved in these processes have been provided through
the genetic analysis of disorders associated with brain cortical
dysgeneses. Such disorders including type-I lissencephaly and
subcortical laminar heterotopia (SCLH), which result from muta-
tions in the LIS1 (Reiner et al., 1993) and doublecortin genes (also
called DBCN or DCX) (des Portes et al., 1998; Gleeson et al., 1998),
are characterized by a dramatic disorganization of neocortical layers
and gyral abnormalities. In addition to LIS1 and DCX, mutations in
cdk5 (Oshima et al., 1996), p35 (Chae et al., 1997), filamin 1 (Fox et
al., 1998), reelin (D’Arcangelo et al., 1995; Hong et al., 2000),
mDab1 (Howell et al., 1997; Sheldon et al., 1997;Ware et al., 1997),
VLDLR and ApoER2 (Trommsdorff et al., 1999), POMGnT1
(Michele et al., 2002; Moore et al., 2002) and POMT1 (Beltran-
Valero de Bernabe et al., 2002), and ARX (Kitamura et al., 2002)
have been found to lead to specific phenotypes associated with
disrupted corticogenesis, either in human or in mouse.
One signaling pathway is represented by Reelin, mDab1,
ApoER2, and VLDLR, which are potentially involved in the
control of the final position of neurons and function through a
linear pathway at the plasma membrane (D’Arcangelo et al.,
1999; Hiesberger et al., 1999; Trommsdorff et al., 1999).
Signaling pathways regulated by Cdk5 and its specific activator
p35 were shown to be involved in neuronal migration and several
other processes such as neurite outgrowth, vesicular trafficking,
and neurodegeneration (Smith and Tsai, 2002; Tan et al., 2003).
The Cdk5/p35 kinase has been shown to regulate N-cadherin-
mediated adhesion, which also requires h-catenin, isolated as a

G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164154
p35-interacting protein in a two-hybrid screen (Kesavapany et al.,
2001; Kwon et al., 2000). More recently this kinase has also been
shown to phosphorylate DCX (Tanaka et al., 2004) and one of
the LIS1-interacting proteins (Niethammer et al., 2000). The
strikingly similar phenotypes of lissencephaly in humans with
mutations in LIS1 or DCX, together with findings demonstrating
that both proteins are microtubule-associated proteins (MAPs;
Francis et al., 1999; Gleeson et al., 1999; Horesh et al., 1999;
Sapir et al., 1997), suggest that LIS1 and DCX may function
through similar mechanisms. Although a recent elegant study
using an RNAi-based approach showed that DCX-deficient
neurons prematurely stop migrating to form subcortical band
heterotopias within the intermediate zone and then white matter
(Bai et al., 2003), basic molecular, biochemical, and biological
processes underlying this premature arrest of migration remain to
be elucidated.
Towards this aim, we carried out a yeast two-hybrid screen to
identify proteins interacting with DCX. Using such a screen, we
previously showed that DCX interacts with the A subunits of the
clathrin adaptor complexes AP-1 and AP-2, known to be
involved in vesicle trafficking associated with the trans-Golgi
network and the plasma membrane, respectively (Friocourt et al.,
2001). We describe here the identification and characterization of
an additional DCX-interacting protein, the ubiquitin protease
Drosophila fat facets related gene on X chromosome (DFFRX),
also called Usp9x), which is involved in cleaving ubiquitin from
protein–ubiquitin conjugates. Besides its role in protein degrada-
tion, ubiquitination has been shown to play an important function
in an increasing number of processes including endocytosis,
signaling, DNA repair, and cell cycle regulation. Monoubiquiti-
nation is in particular associated with the regulation of the
internalization of membrane proteins and their transport to the
lysosomes (Hicke, 2001). Polyubiquitination on the other hand is
necessary for protein degradation by the 26S proteasome, a multi-
subunit proteolytic complex. The deubiquitination process is
catalyzed by proteases called deubiquitinating enzymes, which
fall into four distinct families: ubiquitin C-terminal hydrolases,
ubiquitin-specific processing proteases (USPs or UBPs), out-
domain ubiquitin–aldehyde binding proteins, and Jab1/pad1/
MPN-domain-containing metalloenzymes (Kim et al., 2003).
Among these four families, UBPs represent the most widespread
deubiquitinating enzymes across evolution.
In this study, we found that DCX interacts with a novel
recognition domain in DFFRX, a member of the UBPs family. A
specific lissencephaly causing mutation in DCX leads to the loss of
this interaction. Our data show that the DFFRX mouse orthologue
associates with microtubules in specific subcellular compartments
and is thus likely to play a role in dynamic processes required
during cell migration.
Results
DCX interacts with the C-terminal part of DFFRX in yeast
As the MAP function of DCX does not fully explain the role
of this protein in neuronal migration, we searched for further
DCX-interacting proteins. Using the full-length cDNA of DCX as
bait, we screened a human fetal brain cDNA library using the
two-hybrid system (Fields and Song, 1989). Two successive
screens of this library were performed; the first one identified a
clone that contains the last 255 amino acids of human DFFRX
(DFFRX-Cter, amino acids 2292–2547), whereas the second
isolated a similar but longer C-terminal clone (the last 343 amino
acids). The specificity of the interaction with DCX was confirmed
by testing these two clones with unrelated baits (lamin C and
RAS) (Fig. 1A).
The DFFRX protein contains the typical hallmarks of the UBPs
family of deubiquitinating enzymes, that is, the presence of
cysteine and histidine boxes in a core enzymatic domain (amino
acids 1550–1948) (Jones et al., 1996). Neither the N-terminal part
of DFFRX nor the C-terminus that includes the DCX-interacting
region contain any known functional domains. DCX contains a
microtubule binding domain, composed of two repeats (amino
acids 47–135 and 174–259) (Sapir et al., 2000; Taylor et al., 2000)
and a serine/proline-rich region in its C-terminal part. To map the
domain of interaction with DFFRX in DCX, we tested several
deletion mutants with the clone DFFRX-Cter in a two-hybrid assay
and concluded that the domain of interaction is located between the
amino acids 110 and 214 that contains the proximal part of the
second microtubule binding repeat (Fig. 1B). The A1 subunit of theAP-1 complex was used as a control since we had previously
shown that A1 interacts with a YLPL motif in the C-terminus of
DCX (aa 345–348) (Friocourt et al., 2001). Different mutant clones
of DCX corresponding to missense mutations found in patients
with lissencephaly were similarly tested. Two-hybrid results
showed a loss of interaction between the clone DFFRX-Cter and
the mutant clone R192W (Fig. 1C). The position of this amino acid
and the results obtained with the DCX deletion mutants are
consistent with the defined interacting domain localized between
amino acids 110 and 214. Other mutants, also located in this region
(R186C, R196S, and T203R) still interacted with DFFRX-Cter,
suggesting that the residue R192 is particularly important for this
interaction.
Dffrx is expressed in the developing nervous system
As the pattern of expression of Dffrx, the DFFRX mouse
orthologue, has not been studied in depth in the nervous system,
we performed in situ hybridization studies using mouse embryo
sections at different stages using a 3VUTR probe of Dffrx. A probe
was chosen in the 3VUTR in order to avoid cross hybridization with
other similar UBP family members. Dffrx appears to be expressed
ubiquitously at E12.5 and E14.5 but is interestingly most strongly
expressed in the nervous system in the region of the third, fourth,
and lateral ventricles and in the developing spinal cord (Fig. 2A,
panels A and G). At E14.5, the Dffrx signal is particularly obvious
in the olfactory lobe (Fig. 2A, panels G–I) and in certain ganglia
(trigeminal ganglia, inferior glossopharyngeal ganglia, and dorsal
root ganglia; Fig. 2A, panel G). At E16.5, Dffrx expression is
clearly regionalized to the nervous system. It is strongly expressed
in the developing brain, in the mantle layer of the spinal cord, and
in certain ganglia (Fig. 2A, panels C–F). Some expression was also
detected in the testis. In the developing cortex, Dffrx is expressed
in the neuroblasts of the ventricular zone, as well as in migrating
and differentiating neurons of the intermediate zone and cortical
plate (Fig. 2A, panels J–K). In the eye, the pigment layer of the
retina and the inner neural layer of the optic cup are specifically
labeled (Fig. 2A, panels L–M). Dffrx is still strongly expressed
ubiquitously in mouse newborn brain but its expression seems to
decrease after neonatal stages and it appears generally less
abundant in most regions of the adult brain (Fig. 2B, panels A–

Fig. 1. Two-hybrid analysis. (A) Confirmation of the specificity of the interaction between DCX and DFFRX-Cter (corresponding to the last 255 residues of
DFFRX). Mating was performed between L40 transformants (indicated on left side) and AMR70 (indicated above). X-gal and histidine results are presented.
RAS/RAF: positive control. (B) Definition of DCX-interacting domain. Different constructs of DCX were tested with DFFRX-Cter and with A1 in a two-hybridassay. The identified DCX domain that interacts with DFFRX-Cter spans the residues 110–214. R1 and R2 represent the two repeats present in the microtubule
binding domain. (C) Two-hybrid results obtained with the R192W DCX mutation (identified in a patient with lissencephaly). R192W was the only DCX
mutation that showed a loss of interaction with DFFRX-Cter. Note that this mutant is still capable to interact with the A1 protein.
G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164 155
D), although regions of the hippocampus remain intensely stained
(Fig. 2B, panels C–D). Nevertheless Western blot analysis of
mouse adult brain extracts confirmed that Dffrx is still relatively
ubiquitous (Fig. 2C). These data thus suggest that Dffrx is
important both during neuronal development and in mature
neurons.
Dffrx and DCX interact in vivo
Prior to assessing the interaction between Dffrx and Dcx in
vivo and to further support the two-hybrid results, we further
analyzed the spatial and temporal coexpression and subcellular
colocalization of Dcx and Dffrx. By Western blot analysis, we
observed that Dffrx is expressed in differentiating neurons in
primary culture, in glial and PC12 cells, but is barely detectable
in COS-7 cells (Fig. 3A). Previous studies have shown that
DCX is also expressed in young differentiating neurons, but not
in glial or PC12 cells (Francis et al., 1999; Gleeson et al., 1999;
Horesh et al., 1999). We also performed immunolabelings using
E16.5 mouse embryonic and adult brain sections. The results
obtained with antibodies directed against the catalytic domain
and against the C-terminus of Dffrx were identical and coherent
with those generated by in situ hybridization. Within neuronal
cells, Dffrx seems to be expressed in the soma as well as in the
axons, as shown in the Purkinje cells of the adult cerebellum
(Fig. 3B, panel A). In the embryonic cortex, Dffrx antibody
labels the proliferating cells of the ventricular zone, the
intermediate zone containing neurons in migration and axonal
processes, and the differentiating neurons of the cortical plate
(Fig. 3B, panels B–C). According to previous studies (Francis et
al., 1999; Gleeson et al., 1999), Dcx is present in both the
intermediate zone and the cortical plate regions of the
developing cortex. In the adult brain, Dcx is only expressed
in a limited population of neurons, which migrate from the
subventricular zone to the olfactory bulb (rostral migratory
stream), whereas Dffrx is more widely expressed. A colocaliza-
tion was observed between Dffrx and Dcx in these migrating
neurons (Fig. 3B, panels D–F). Immunolabeling using primary
cultures of cortical neurons confirmed that Dffrx was present in
both the soma and the neurites (Fig. 3C, panel A). It was
strongly expressed in growth cones and at the end of neurites,
where it partially colocalized with Dcx (Fig. 3C, panels D–I).
Double-labeling experiments performed with actin and a-tubulin
showed Dffrx expression in the neurites and the main body of
the growth cone. Growth cones are actin-rich but also contain
microtubules, and indeed Dffrx showed a colocalization with
tubulin rather than with actin, as shown here by the Dcx
labeling (Fig. 3C, panels D–I).
In order to confirm the interaction between Dffrx and Dcx in
vivo, we first performed co-immunoprecipitation experiments in
COS-7 cells transfected with FLAG-tagged DCX and GFP-tagged
DFFRX-Cter (equivalent to the clone isolated in the two-hybrid
assay). GFP-DFFRX-Cter was co-immunoprecipitated with
FLAG-DCX but not with the mutant form FLAG-R192W (Fig.
4A). These results thus confirm those obtained with the yeast two-
hybrid system. As Dffrx is an enzyme, we also looked for an
interaction between its catalytic domain and DCX. A full-length
Dffrx construct tagged with V5, or the catalytic domain of Dffrx
(amino acids 1210–2410) tagged with HA, were expressed with
either GFP-DCX or GFP alone. Immunoprecipitation with anti-
bodies directed against V5 or HA showed that DCX interacted with
full-length Dffrx but was not able to co-immunoprecipitate with the
catalytic domain of Dffrx (Fig. 4B). We then performed co-
immunoprecipitation experiments involving the endogenous pro-
teins using newborn mouse brain extracts. We selected this stage of
development because both Dffrx and Dcx remain well expressed in

Fig. 2. Expression of Dffrx during development. In situ hybridization was performed using mouse sections. (A) Dffrx is ubiquitous at E12.5 (A) and E14.5 (G)
but regionalizes to the nervous system at E16.5 (C–F). These results are likely to be specific since the sense probes show no labeling (B, D, and F). At E14.5,
Dffrx is strongly expressed in the developing olfactory bulb (H–I, I, toluidine blue coloration of the section). In the developing cortex, at E16.5, Dffrx is
expressed in the cortical plate, the intermediate zone, and the ventricular zone (J–K). In the eye of an E16.5 embryo, Dffrx is strongly expressed in the pigment
layer of the retina and in the inner nuclear layer of the optic cup (L–M, M, sense probe showing nonspecific labeling of the lens). drg: dorsal root ganglion, tg:
trigeminal ganglion, cx: cortex, ob: olfactory bulb, cp: cortical plate, iz: intermediate zone, vz: ventricular zone, le: lens, pl: pigment layer, inl: inner neural
layer. (B) Localization of Dffrx in newborn and adult mouse. Dffrx is strongly expressed throughout the newborn brain (coronal sections A–B, B, sense probe).
The absence of signal in a region of the visual cortex is probably an artefact as it was not observed in other experiments. In a coronal section of adult mouse
brain, Dffrx expression appears strongest in regions of the hippocampus (C–D, D, sense probe), although it is still expressed ubiquitously. Sagittal sections of
the adult cerebellum show a strong labeling of the Purkinje cells (E–F, F, sense probe showing nonspecific labeling of the white matter). ML: molecular layer,
GL: granular layer, P: Purkinje cells, WM: white matter. (C) Western blot analysis of protein extracts derived from different regions of the mouse adult brain.
Twenty micrograms of proteins was loaded in each well. Extracts were detected with anti-DFFRX-Cter and anti-actin (control) antibodies.
G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164156
newborn brain. Dffrx was found to co-immunoprecipitate with Dcx
using anti-Dcx antibodies but was not present in anti-GST
precipitates used as a control (Fig. 4C). Dcx was not detected in
anti-Dffrx immunoprecipitates in vivo but the results with tagged
proteins are still in favor of the existence of the interaction. These
results therefore confirm that Dcx and Dffrx can exist in the same
protein complex in mouse brain in vivo and that Dcx probably
interacts only with the C-terminus of Dffrx but not with its catalytic
domain.
Dcx is not ubiquitinated and is not a substrate of Dffrx
To elucidate whether Dcx is a substrate of Dffrx and thus
could be normally regulated by ubiquitination, we first
measured the half-life of Dcx in primary cultures of rat
embryonic cortical neurons and found it to be longer than 12
h (data not shown). We next examined the effect of proteasome
inhibitors on DCX stability. PC12 cells were transfected with a
DCX expression plasmid and then treated with MG132 and
lactacystin. Under these conditions, both proteasome inhibitors
had little effect on the stability of DCX, whereas the amount of
h-catenin, a protein known to be ubiquitinated and degraded by
the proteasome (Aberle et al., 1997), strongly increased with the
presence of several slower migrating species likely resulting
from ubiquitin modifications of this protein in the presence of
MG132 and lactacystin (Fig. 5A). Similar results were obtained
using primary neuronal cultures expressing endogenous Dcx and
h-catenin (data not shown).
To further assess whether Dcx can be ubiquitinated, COS-7
cells were transfected with HA-tagged ubiquitin (HA-Ub) and
GFP-tagged DCX. GFP-p35 was used as a control since this
protein has been previously described to be ubiquitinated and
degraded by the proteasome (Patrick et al., 1998). The
expression levels of GFP-DCX were similar to those obtained
for GFP-p35 in these experiments. High molecular weight
isoforms of p35 likely corresponding to polyubiquitinated
species of this protein clearly accumulated in the presence of
MG132, whereas no modification of GFP-DCX could be
detected under the same conditions (Fig. 5B). To enrich for
further potential ubiquitinated species of p35 and DCX,
corresponding cell lysates were immunoprecipitated with anti-
HA or anti-GFP antibodies (Fig. 5C). High molecular weight

Fig. 3. Coexpression of Dffrx and Dcx during development and subcellular localization of both proteins. (A) Coexpression of Dffrx and Dcx in young
differentiating neurons in culture. Anti-N-term and anti-Dffrx antibodies were used to probe extracts from primary cultures of mouse brain neurons grown for 2,
6, 14, 21, and 30 days as well as glial cells, PC12, and COS-7 cells. (B) Immunohistochemistry performed using mouse sections. Dffrx is expressed in the soma
and in the axons of Purkinje cells in a sagittal section of an adult cerebellum (A). In the developing cortex (coronal section) of an E16.5 embryo, Dffrx is
expressed in the progenitor cells of the ventricular zone (VZ), in the intermediate zone (IZ), and in the cortical plate (CP) (B–C). In the adult brain, cells in the
rostral migratory stream that express Dcx are also stained with Dffrx (D–F), although Dffrx is more widely expressed. (C) Localization of Dffrx in
differentiating neurons derived from E18 fetal rat brain. Dffrx labels both the soma and neurites of neurons in culture, whereas Dcx more specifically labels the
extremities of neurites. Although Dffrx expression seems to extend further into the growth cone than Dcx, it is also present at the end of neurites where it
colocalizes with Dcx. The scale bar equals 25 Am.
G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164 157
species of p35 were immunoprecipitated with both GFP and
anti-HA antibodies confirming that they correspond to p35
ubiquitin conjugates. As expected, in these experiments, treat-
ment with MG132 increased the amount of ubiquitinated p35.
Unlike p35, we observed no higher molecular weight bands
corresponding to DCX in the anti-GFP or anti-HA precipitates
(Fig. 5C). Similarly, polyubiquitinated Dcx forms were not
detected in proteins extracts complemented with proteasome
inhibitors from E15.5 and newborn mouse brains (data not
shown). Though these results cannot totally exclude the absence
of a ubiquitinated fraction of endogenous Dcx, together they
suggest that Dcx is not a major substrate for ubiquitination and
degradation by the proteasome and consequently not a substrate
of Dffrx.

Fig. 4. Interaction between Dffrx and Dcx in vivo. (A) Co-immunoprecipitation of DFFRX-Cter with Dcx but not with the mutant R192W. FLAG-DCX or
R192W were transfected with GFP-DFFRX-Cter in COS-7 cells. After immunoprecipitation of the lysates with DCX anti-N-term or anti-DFFRX-Cter
antibodies, the blots were probed with either goat anti-DCX C18 or anti-GFP. (B) Absence of an interaction between DCX and the catalytic domain of Dffrx.
Full-length Dffrx (V5-Dffrx-FL) or its catalytic domain (HA-Dffrx-CAT) were cotransfected with either GFP-DCX or GFP alone. Lysates were
immunoprecipitated with anti-V5 or anti-HA antibodies and the blots were detected with anti-Dffrx-CAT or with anti-DCX-N-term antibodies. SAP:
supernatant after immunoprecipitation, IP: immunoprecipitation. (C) Immunoprecipitation between Dcx and Dffrx from mouse newborn brain. Extracts were
immunoprecipitated with either anti-DCX N-term, anti-GST, or anti-Dffrx-CAT antibodies and the blot was detected with anti-Dffrx-CAT.
G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164158
The DCX/Dffrx complex is associated with microtubules
Immunoprecipitation experiments and colocalization of Dcx,
Dffrx, and microtubules at the distal part of neurites in differ-
entiating neurons (Fig. 3) led us to hypothesize that the function of
Dffrx may be important when it is localized in protein complexes
that interact with microtubules. The association between endoge-
nous Dffrx and microtubules was thus further investigated in
different cell types. In nondifferentiated proliferating PC12 cells,
Dffrx is concentrated in actin-rich but also microtubule-containing
structures, such as the filopodia and the midbody of dividing cells
during cytokinesis (Figs. 6A–C). In the soma, Dffrx appears to be
most concentrated in a region close to the nucleus. Double labeling
with antibodies directed against g-tubulin suggests that this region
corresponds to the centrosome or to the microtubule-organizing
center (MTOC) (Figs. 6D–F). In dividing cells, Dffrx seems to be
most concentrated at the mitotic spindle (Figs. 6G–I), which
confirms Dffrx association with the MTOC. A similar enrichment
of Dffrx in the region of the MTOC was observed in glial cells
(Figs. 6J–L) and in differentiating neurons in culture (data not
shown). Interestingly, in confluent glial cells, Dffrx showed a
different subcellular localization as it was instead concentrated at
cell–cell junctions, where it colocalized with h-catenin (Figs. 6M–
O), as previously described in MDCKII cells (Taya et al., 1999).
This observation led us to test whether Dcx and h-catenin might be
associated in the same protein complex in vivo. To this end, we
performed co-immunoprecipitation experiments from mouse
embryonic brain using anti-Dcx and anti-Dffrx antibodies and
tested for the presence of h-catenin by Western blot analysis. Fig. 7
shows that h-catenin is co-immunoprecipitated with both Dcx and
Dffrx, suggesting that these three proteins can associate in the same
protein complex in embryonic brain. The different subcellular
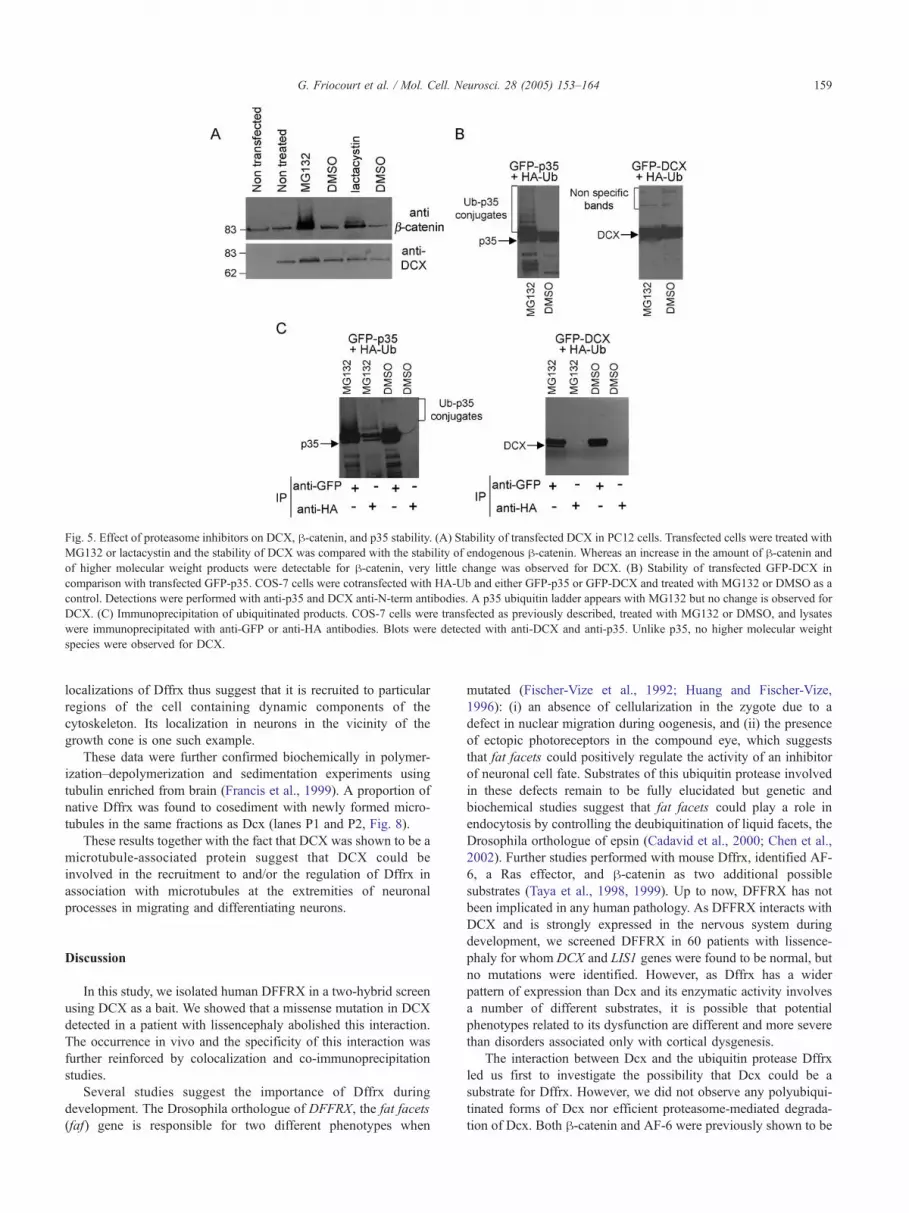
Fig. 5. Effect of proteasome inhibitors on DCX, h-catenin, and p35 stability. (A) Stability of transfected DCX in PC12 cells. Transfected cells were treated with
MG132 or lactacystin and the stability of DCX was compared with the stability of endogenous h-catenin. Whereas an increase in the amount of h-catenin and
of higher molecular weight products were detectable for h-catenin, very little change was observed for DCX. (B) Stability of transfected GFP-DCX in
comparison with transfected GFP-p35. COS-7 cells were cotransfected with HA-Ub and either GFP-p35 or GFP-DCX and treated with MG132 or DMSO as a
control. Detections were performed with anti-p35 and DCX anti-N-term antibodies. A p35 ubiquitin ladder appears with MG132 but no change is observed for
DCX. (C) Immunoprecipitation of ubiquitinated products. COS-7 cells were transfected as previously described, treated with MG132 or DMSO, and lysates
were immunoprecipitated with anti-GFP or anti-HA antibodies. Blots were detected with anti-DCX and anti-p35. Unlike p35, no higher molecular weight
species were observed for DCX.
G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164 159
localizations of Dffrx thus suggest that it is recruited to particular
regions of the cell containing dynamic components of the
cytoskeleton. Its localization in neurons in the vicinity of the
growth cone is one such example.
These data were further confirmed biochemically in polymer-
ization–depolymerization and sedimentation experiments using
tubulin enriched from brain (Francis et al., 1999). A proportion of
native Dffrx was found to cosediment with newly formed micro-
tubules in the same fractions as Dcx (lanes P1 and P2, Fig. 8).
These results together with the fact that DCX was shown to be a
microtubule-associated protein suggest that DCX could be
involved in the recruitment to and/or the regulation of Dffrx in
association with microtubules at the extremities of neuronal
processes in migrating and differentiating neurons.
Discussion
In this study, we isolated human DFFRX in a two-hybrid screen
using DCX as a bait. We showed that a missense mutation in DCX
detected in a patient with lissencephaly abolished this interaction.
The occurrence in vivo and the specificity of this interaction was
further reinforced by colocalization and co-immunoprecipitation
studies.
Several studies suggest the importance of Dffrx during
development. The Drosophila orthologue of DFFRX, the fat facets
(faf) gene is responsible for two different phenotypes when
mutated (Fischer-Vize et al., 1992; Huang and Fischer-Vize,
1996): (i) an absence of cellularization in the zygote due to a
defect in nuclear migration during oogenesis, and (ii) the presence
of ectopic photoreceptors in the compound eye, which suggests
that fat facets could positively regulate the activity of an inhibitor
of neuronal cell fate. Substrates of this ubiquitin protease involved
in these defects remain to be fully elucidated but genetic and
biochemical studies suggest that fat facets could play a role in
endocytosis by controlling the deubiquitination of liquid facets, the
Drosophila orthologue of epsin (Cadavid et al., 2000; Chen et al.,
2002). Further studies performed with mouse Dffrx, identified AF-
6, a Ras effector, and h-catenin as two additional possible
substrates (Taya et al., 1998, 1999). Up to now, DFFRX has not
been implicated in any human pathology. As DFFRX interacts with
DCX and is strongly expressed in the nervous system during
development, we screened DFFRX in 60 patients with lissence-
phaly for whom DCX and LIS1 genes were found to be normal, but
no mutations were identified. However, as Dffrx has a wider
pattern of expression than Dcx and its enzymatic activity involves
a number of different substrates, it is possible that potential
phenotypes related to its dysfunction are different and more severe
than disorders associated only with cortical dysgenesis.
The interaction between Dcx and the ubiquitin protease Dffrx
led us first to investigate the possibility that Dcx could be a
substrate for Dffrx. However, we did not observe any polyubiqui-
tinated forms of Dcx nor efficient proteasome-mediated degrada-
tion of Dcx. Both h-catenin and AF-6 were previously shown to be

Fig. 6. Dffrx expression in PC12 and glial cells. In PC12 cells (A–I), Dffrx is present in cell regions containing microtubules such as the midbody during
cytokinesis (A–C), the centrosome (D–F), and the mitotic spindle in dividing cells (G–I). In nonconfluent glial cells (J–L), Dffrx strongly labels the MTOC
(indicated by an arrow). In confluent cells, Dffrx is strongly colocalized with h-catenin at the cell–cell junctions (M–O). These data thus show a relocalization
of Dffrx depending on the cell state.
G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164160
polyubiquitinated and to interact directly with a region containing
the catalytic domain of Dffrx. In contrast, Dcx interacts with a
novel recognition region in Dffrx involving its C-terminal 250
amino acids, a domain that is highly conserved among Drosophila,
mouse, and human and relatively unique to this protein. Dcx may
therefore help regulate Dffrx localization within the cell. Indeed
Dcx is a microtubule-associated protein with a specific localization
in neuronal cells (Francis et al., 1999; Gleeson et al., 1999; Horesh
et al., 1999), and Dffrx colocalizes to these regions and cosedi-
ments with microtubules. The interaction with Dcx could therefore
anchor Dffrx in specialized compartments of neuronal cells (such
as dynamic structures corresponding to distal segments of neurites)

Fig. 7. Co-immunoprecipitation of endogenous Dcx, h-catenin, and p35.
Mouse E15.5 brain extracts were immunoprecipitated with either anti-Dcx
N-term, anti-DFFRX-Cter, anti-h-catenin, or anti-p35 antibodies. h-Cateninis immunoprecipitated with Dcx, Dffrx, and in small amounts with p35.
Fig. 8. Interaction between Dffrx and microtubules. Native Dffrx is present
in microtubules fractions induced through cycles of polymerization and
depolymerization. Western blots (20 Ag protein/lane) contain total mouse
brain extracts (T), soluble (S0), and insoluble cell components (P0). The
tubulin in the S0 fraction was polymerized to form microtubules and
sedimented, resulting in soluble (S1) and pellet (P1) fractions. Microtubules
in P1 were depolymerized by incubation at 08C for 1 h and resedimented
(PV1), and the resulting soluble fraction was subjected to a further cycle of
polymerization, followed by sedimentation (S2 and P2). Like Dcx, Dffrx
cosediments with newly formed microtubules. Note that in addition to the
band corresponding to the full-length Dffrx protein, anti-Dffrx CAT
antibody detected a product with higher molecular weight than Dffrx. It
remains to be seen whether this protein corresponds to another isoform of
Dffrx or to a cross-reaction of the antibody with an as yet unidentified
protein. p35 was used as a negative control. Unlike Dffrx and DCX, it is not
present in the P2 fraction.
G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164 161
to regulate the activity and/or the stability of relevant proteins in
this region. A similar situation has been evoked for the MID1
microtubule-associated protein, which is likely to target phospha-
tase 2A for degradation (Trockenbacher et al., 2001). It is also
interesting to note that DCLK1C, a protein similar to Dcx, which is
expressed during development and in adult stages in astroglial and
neuronal cells (Burgess et al., 1999; Friocourt et al., 2003), also
interacts with Dffrx (data not shown). It is therefore possible that
DCLK1C could, at relevant times, aid the localization of Dffrx in
certain cells and in cellular compartments where Dcx is not
expressed although this remains to be further investigated.
Besides its potential role anchoring Dffrx to a specific local-
ization, Dcx may also influence the enzymatic activity of Dffrx.
Although it has been reported that human HAUSP is necessary and
sufficient for the in vitro deubiquitination of its specific substrate,
the p53 tumor suppressor (Li et al., 2002), the yeast Ubp3 protein
has been shown to require an additional regulatory subunit, Bre5p,
to insure its function (Cohen et al., 2003a,b). As for the interaction
of Dcx with Dffrx, Bre5p interacts with Ubp3p on a domain
distinct from the catalytic region (M. Cohen and C. Dargemont,
personal data). It is thus possible that DCX acts as a regulatory
subunit of Dffrx, aiding the deubiquitination and stabilization of
specific substrates in the vicinity of the growth cone. Indeed, the
importance of the role of the ubiquitination and protein degradation
in growth cones has been shown for the Netrin-1 signaling
pathway (Campbell and Holt, 2001).
Our co-immunoprecipitation data confirm that Dcx is present in
the same protein complexes as Dffrx and h-catenin in the
developing brain. We observed a colocalization between Dffrx
and h-catenin at cell–cell junctions in confluent glial cells and at
adherens junctions in proliferating cells of the ventricular zone
(data not shown), perhaps suggesting that Dffrx ensures the
stability of h-catenin while homotypic adhesion is required. Dcx
is not expressed in proliferating cells; nevertheless, its influence on
h-catenin stability in postmitotic neurons remains a possibility. It is
interesting to point out that despite the apparent diversity of
ubiquitinated proteins shown to be substrates of Dffrx in cultured
cellular models, all are involved directly or indirectly in cellular
processes such as cell adhesion, endocytosis, and vesicle traffick-
ing. Notably, these functions are extensively required at the
extremities of neuronal processes where Dcx is localized. AF-6,
for example, is implicated in adhesion and has a specific neuronal
isoform (Ikeda et al., 1999; Mandai et al., 1997). In addition, epsin
is involved in endocytosis and vesicle formation (Wendland,
2002). Interestingly, it has been reported that epsin binds to
microtubules in neurons and could play a role in neuritic outgrowth
(Hussain et al., 2003). In addition, a recent study shows the
presence of Dffrx at multiple points of protein trafficking in
epithelial cell lines (Murray et al., 2004). Thus, the known
substrates of Dffrx and possible functions of Faf predicted from
the phenotypes of the Drosophila mutants seem collectively to
suggest the involvement of the Dffrx/Dcx complex in the
regulation of adhesion and membrane turnover, which are essential
processes for neuronal migration and differentiation. In line with
this function are the previously reported data indicating that DCX
interacts with the A1 and A2 subunits of the clathrin adaptor
complexes implicated in vesicle trafficking (Friocourt et al., 2001)
and the phosphorylated form of neurofascin, a protein implicated in
neuronal adhesion (Kizhatil et al., 2002). Determining the relation-
ships between these different partners of DCX will further
elucidate its role in neuronal migration and differentiation.
Experimental methods
Yeast cultures and two-hybrid analysis
The human fetal brain cDNA library cloned in pACT2 was
purchased from CLONTECH laboratories (MATCHMAKER) and
transformed into the L40 yeast strain. A first screen was performed
by transforming the full-length DCX cDNA cloned in pBTM116
into the L40 strain containing the library plasmids (Friocourt et al.,
2001), and the second one was performed by mating between the
L40DGal4 strain transformed with VJL12-DCX and the Y187
strain transformed with the library.
The mutant forms of DCX (S47R, R59L, Y125H, W146C,
R186C, R196S, and T203R) were generated using the Quik
Change Mutagenesis kit (Stratagene). The constructs correspond-

G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164162
ing to the catalytic domain of Dffrx (aa 1210–2410) and to Dffrx
full-length cDNA have been previously described (Murray et al.,
2004; Taya et al., 1998).
Antibodies
We used affinity-purified antibodies directed against the N-
terminal part (anti-N-term; Francis et al., 1999) and the C-terminal
part of DCX (Santa Cruz, C18). A polyclonal antibody directed
against the C-terminal part of DFFRX (anti-DFFRX-Cter, encom-
passing amino acids 2292-2547 according to GenBank X98296)
was generated and affinity purified. Towards this aim, a two-hybrid
clone containing DFFRX-Cter was used to generate a GST fusion
protein in bacteria. The protein obtained was purified by
chromatography using glutathione–sepharose and used to generate
rabbit polyclonal antisera. In this region, the amino acid sequence
is 98% identical between human and mouse. This antibody detects
by immunoblotting a band at the expected size. The specificity of
this band was confirmed by comparison with polyclonal antibodies
directed against the catalytic domain of mouse Dffrx (anti-Dffrx-
CAT) and against the first 20 N-terminal amino acids, gifts from Pr
K. Kaibuchi and S.A. Wood, respectively. The following anti-
bodies were also used: polyclonal anti-p35 (Santa Cruz) and
monoclonal anti-GFP (Roche), anti-actin (Chemicon), anti-HA
(Babco), a-tubulin, and g-tubulin (Sigma); anti-h-catenin anti-
bodies (Transduction Laboratories); and rat anti-HA antibodies
(Roche).
In situ hybridization
Mouse Dffrx 3’UTR sequence (nucleotides 9052–9983,
U67874) was cloned in pBluescript, and sense and antisense
probes were generated by in vitro transcription. Mouse embryos
and adult brains were fixed in 2–4% paraformaldehyde, cryopro-
tected with 30% sucrose in phosphate buffer, frozen with
isopentane, and cut into approximately 20 Am thick sections.
Slides were incubated with a35S-labelled RNA probes in a 50%
formamide solution at 608C. Washes were performed in succes-
sively stringent SSC solutions, with a final wash at 0.1� SSC at
608C. Slides were then dipped in diluted Kodak NTB2 emulsion
and exposed for 24–48 h. Emulsions were developed and sections
counterstained with toluidine blue, mounted in Eukitt, and
examined under light microscopy with an SMZ1000 stereoscopic
microscope (Nikon) and an Axiophot microscope (Zeiss).
Immunohistochemistry and cytochemistry
Mouse embryos and adult brains were frozen with isopentane
and sectioned (approximately 10 Am thick). Sections were fixed at
48C for 20 min in 4% paraformaldehyde solution. Primary
neuronal cultures were prepared from E18 rat brains as previously
described (Berwald-Netter et al., 1981). Neurons grown on
coverslips were fixed for 7 min at �208C in methanol. Immuno-
fluorescence labeling was performed as previously described
(Friocourt et al., 2001).
Primary glial cultures were prepared according to Nowak et al.
(1987) from newborn OF1 mouse cerebral hemispheres. Glial cells
were cultured in Dulbecco’s modified Eagle’s medium (DMEM,
Invitrogen) supplemented with 4.5 mg/ml glucose, 10% fetal
bovine serum (Invitrogen), and 100 units/ml of penicillin and
streptomycin. They were used as secondary cultures containing
more than 95% of GFAP-positive astrocytes. After three washes in
phosphate-buffered saline solution (PBS), the cells were prehybri-
dized for 1 h with TBST containing 2% goat serum or BSA, and
then incubated overnight at 48C with the primary antibody.
Co-immunoprecipitation
COS-7 cells were cultured in DMEM (Invitrogen) supple-
mented with 10% fetal bovine serum. Cells were transfected using
the FUGENE6 transfection reagent (Roche Molecular Biochem-
icals), lysed after 24 h in IP buffer (50 mM Tris–HCl pH 8, 150
mM NaCl, 1% NP40, and protease inhibitors), and centrifuged at
50,000 � g for 30 min at 48C. After a preclearing with protein A-
agarose (Sigma), aliquots of the resulting supernatant were
incubated overnight at 48C with the antibodies of interest. Protein
A-agarose was added and samples were incubated 2 h at 48C.Immunoprecipitates were washed with IP buffer and proteins were
separated on SDS-polyacrylamide gels.
Mouse E15.5 embryonic or newborn brains were crushed in
liquid nitrogen and resuspended in 10 volumes of IP buffer.
Immunoprecipitations were performed as described above.
Ubiquitination studies
PC12 cells were grown in DMEM supplemented with 5% fetal
bovine serum and 10% horse serum and were transfected with
GFP-DCX using GenePorter (gene Therapy System) according to
the manufacturer’s instructions. Twenty hours after transfection,
the proteasome inhibitors lactacystin (Calbiochem) and MG132
(Affiniti) were added at the final concentrations of 12.5 AM for 2 h
and 20 AM for 5 h 30 min, respectively. Cells were harvested and
resuspended in sample buffer.
For immunoprecipitation experiments, COS-7 cells were
transfected with the different constructs. Twenty hours after
transfection, cells were treated with 20 AM of MG132 for 5 h and
lysed in IP buffer complemented with iodoacetic acid and N-
ethylmaleimide (Sigma, final concentrations 10 and 5 mM,
respectively). Immunoprecipitations were performed as described
above.
Sedimentation of microtubules
The temperature-dependent polymerization–depolymerization
experiment was performed as previously described (Francis et al.,
1999). A brain fraction enriched in tubulin was prepared from E20
mouse embryos by homogenization in MEM buffer (50 mM MES
pH 6.8, 2 mM EGTA, 2 mM MgCl2) with protease inhibitors and
ultracentrifugation (150,000 � g at 48C for 1 h). Soluble tubulin
(S0) was incubated in the presence of 1 mM GTP and 25%
glycerol for 40 min at 378C to form microtubules that were
sedimented (150,000 � g) for 60 min at 308C. The resulting pellet
(P1) was depolymerized for 60 min at 08C and gently homogen-
ized. Sedimentation of the insoluble (PV1) from soluble components
was performed. This soluble fraction was polymerized and
sedimented as before giving rise to P2 and S2 fractions.
Acknowledgments
The authors are grateful to K. Kaibuchi, D. Boucher, M.
Bornens, and A. Benmerah for kind gifts of reagents and helpful

G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164 163
discussions, Pierre Billuart, Sebastien L’Hoste, Philippe Couvert,
Evelyne Souil, Jean-Marie Genevard, and Emmanuelle Tisserand
for their technical contribution to the work, and Odette Godard
for her logistic support. This work was supported in part by
grants from INSERM, CNRS, the Fondation Bettencourt-Schuler,
the Fondation pour la Recherche Medicale, the European
Commission (No. QLG3-CT-2002-01810), the Human Frontier
Science Program (ref. RG0283/1999-B), the Ministere de la
Recherche (ACI-1A066G), and the Federation pour la Recherche
sur le Cerveau. G.F. was supported by a PhD fellowship from the
Fondation pour la Recherche Medicale and by the Fondation
Bettencourt-Schuler.
References
Aberle, H., Bauer, A., Stappert, J., Kispert, A., Kemler, R., 1997.
h-catenin is a target for the ubiquitin–proteasome pathway. EMBO
J. 16, 3797–3804.
Bai, J., Ramos, R.L., Ackman, J.B., Thomas, A.M., Lee, R.V., LoTurco,
J.J., 2003. RNAi reveals doublecortin is required for radial migration in
rat neocortex. Nat. Neurosci. 6, 1277–1283.
Beltran-Valero de Bernabe, D., Currier, S., Steinbrecher, A., Celli, J., van
Beusekom, E., van der Zwaag, B., Kayserili, H., Merlini, L., Chitayat,
D., Dobyns, W.B., Cormand, B., Lehesjoki, A.E., Cruces, J., Voit, T.,
Walsh, C.A., van Bokhoven, H., Brunner, H.G., 2002. Mutations in the
O-mannosyltransferase gene POMT1 give rise to the severe neuronal
migration disorder Walker–Warburg syndrome. Am. J. Hum. Genet. 71,
1033–1043.
Berwald-Netter, Y., Martin-Moutot, N., Koulakoff, A., Couraud, F., 1981.
Na+-channel-associated scorpion toxin receptor sites as probes for
neuronal evolution in vivo and in vitro. Proc. Natl. Acad. Sci. U. S. A.
78, 1245–1249.
Burgess, H.A., Martinez, S., Reiner, O., 1999. KIAA0369, doublecortin-
like kinase, is expressed during brain development. J. Neurosci. Res.
58, 567–575.
Cadavid, A.L., Ginzel, A., Fischer, J.A., 2000. The function of the
Drosophila fat facets deubiquitinating enzyme in limiting photoreceptor
cell number is intimately associated with endocytosis. Development
127, 1727–1736.
Campbell, D.S., Holt, C.E., 2001. Chemotropic responses of retinal growth
cones mediated by rapid local protein synthesis and degradation.
Neuron 32, 1013–1026.
Chae, T., Kwon, Y.T., Bronson, R., Dikkes, P., Li, E., Tsai, L.H., 1997.
Mice lacking p35, a neuronal specific activator of cdk5, display
cortical lamination defects, seizures, and adult lethality. Neuron 18,
29–42.
Chen, X., Zhang, B., Fischer, J.A., 2002. A specific protein substrate for a
deubiquitinating enzyme: liquid facets is the substrate of Fat facets.
Genes Dev. 16, 289–294.
Cohen, M., Stutz, F., Belgareh, N., Haguenauer-Tsapis, R., Dargemont, C.,
2003a. Ubp3 requires a cofactor, Bre5, to specifically de-ubiquitinate
the COPII protein, Sec23. Nat. Cell Biol. 5, 661–667.
Cohen, M., Stutz, F., Dargemont, C., 2003b. De-ubiquitination: a new
player in Golgi to ER retrograde transport. J. Biol. Chem. 278,
51989–51992.
D’Arcangelo, G., Miao, G.G., Chen, S.C., Soares, H.D., Morgan, J.I.,
Curran, T., 1995. A protein related to extracellular matrix proteins
deleted in the mouse mutant reeler. Nature 374, 719–723.
D’Arcangelo, G., Homayouni, R., Keshvara, L., Rice, D.S., Sheldon, M.,
Curran, T., 1999. Reelin is a ligand for lipoprotein receptors. Neuron
24, 471–479.
des Portes, V., Pinard, J.M., Billuart, P., Vinet, M.C., Koulakoff, A., Carrie,
A., Gelot, A., Dupuis, E., Motte, J., Berwald-Netter, Y., Catala, M.,
Kahn, A., Beldjord, C., Chelly, J., 1998. A novel CNS gene required for
neuronal migration and involved in X-linked subcortical laminar
heterotopia and lissencephaly syndrome. Cell 92, 51–61.
Fields, S., Song, O., 1989. A novel genetic system to detect protein–protein
interactions. Nature 340, 245–246.
Fischer-Vize, J.A., Rubin, G.M., Lehmann, R., 1992. The fat facets gene is
required for Drosophila eye and embryo development. Development
116, 985–1000.
Fox, J.W., Lamperti, E.D., Eksioglu, Y.Z., Hong, S.E., Feng, Y., Graham,
D.A., Scheffer, I.E., Dobyns, W.B., Hirsch, B.A., Radtke, R.A.,
Berkovic, S.F., Huttenlocher, P.R., Walsh, C.A., 1998. Mutations in
filamin 1 prevent migration of cerebral cortical neurons in human
periventricular heterotopia. Neuron 21, 1315–1325.
Francis, F., Koulakoff, A., Boucher, D., Chafey, P., Schaar, B., Vinet, M.C.,
Friocourt, G., McDonnell, N., Reiner, O., Kahn, A., McConnell, S.K.,
Berwald-Netter, Y., Denoulet, P., Chelly, J., 1999. Doublecortin is a
developmentally regulated, microtubule-associated protein expressed in
migrating and differentiating neurons. Neuron 23, 247–256.
Friocourt, G., Chafey, P., Billuart, P., Koulakoff, A., Vinet, M.C., Schaar,
B., McConnell, S.K., Francis, F., Chelly, J., 2001. Doublecortin
interacts with A subunits of clathrin adaptor complexes in the
developing nervous system. Mol. Cell. Neurosci. 18, 307–319.
Friocourt, G., Koulakoff, A., Chafey, P., Boucher, D., Fauchereau, F.,
Chelly, J., Francis, F., 2003. Doublecortin functions at the extremities of
growing neuronal processes. Cereb. Cortex 13, 620–626.
Gleeson, J.G., Allen, K.M., Fox, J.W., Lamperti, E.D., Berkovic, S.,
Scheffer, I., Cooper, E.C., Dobyns, W.B., Minnerath, S.R., Ross, M.E.,
Walsh, C.A., 1998. Doublecortin, a brain-specific gene mutated in
human X-linked lissencephaly and double cortex syndrome, encodes a
putative signaling protein. Cell 92, 63–72.
Gleeson, J.G., Lin, P.T., Flanagan, L.A., Walsh, C.A., 1999. Doublecortin is
a microtubule-associated protein and is expressed widely by migrating
neurons. Neuron 23, 257–271.
Hatten, M.E., 1999. Central nervous system neuronal migration. Annu.
Rev. Neurosci. 22, 511–539.
Hicke, L., 2001. Protein regulation by monoubiquitination. Nat. Rev., Mol.
Cell Biol. 2, 195–201.
Hiesberger, T., Trommsdorff, M., Howell, B.W., Goffinet, A., Mumby,
M.C., Cooper, J.A., Herz, J., 1999. Direct binding of Reelin to VLDL
receptor and ApoE receptor 2 induces tyrosine phosphorylation of
disabled-1 and modulates tau phosphorylation. Neuron 24, 481–489.
Hong, S.E., Shugart, Y.Y., Huang, D.T., Shahwan, S.A., Grant, P.E.,
Hourihane, J.O., Martin, N.D., Walsh, C.A., 2000. Autosomal recessive
lissencephaly with cerebellar hypoplasia is associated with human
RELN mutations. Nat. Genet. 26, 93–96.
Horesh, D., Sapir, T., Francis, F., Wolf, S.G., Caspi, M., Elbaum, M.,
Chelly, J., Reiner, O., 1999. Doublecortin, a stabilizer of microtubules.
Hum. Mol. Genet. 8, 1599–1610.
Howell, B.W., Hawkes, R., Soriano, P., Cooper, J.A., 1997. Neuronal
position in the developing brain is regulated by mouse disabled-1.
Nature 389, 733–737.
Huang, Y., Fischer-Vize, J.A., 1996. Undifferentiated cells in the develop-
ing Drosophila eye influence facet assembly and require the Fat facets
ubiquitin-specific protease. Development 122, 3207–3216.
Hussain, N.K., Yamabhai, M., Bhakar, A.L., Metzler, M., Ferguson,
S.S., Hayden, M.R., McPherson, P.S., Kay, B.K., 2003. A role for
ENTH/ANTH domains in tubulin binding. J. Biol. Chem. 278,
28823–28830.
Ikeda, W., Nakanishi, H., Miyoshi, J., Mandai, K., Ishizaki, H., Tanaka, M.,
Togawa, A., Takahishi, K., Nishioka, H., Yoshida, H., Mizoguchi, A.,
Nishikawa, S., Takai, Y., 1999. Afadin: a key molecule essential for
structural organization of cell–cell junctions of polarized epithelia
during embryogenesis. J. Cell Biol. 146, 1117–1132.
Jones, M.H., Furlong, R.A., Burkin, H., Chalmers, I.J., Brown, G.M.,
Khwaja, O., Affara, N.A., 1996. The Drosophila developmental gene
fat facets has a human homologue in Xp11.4 which escapes X-
inactivation and has related sequences on Yq11.2. Hum. Mol. Genet. 5,
1695–1701.

G. Friocourt et al. / Mol. Cell. Neurosci. 28 (2005) 153–164164
Kesavapany, S., Lau, K.F., McLoughlin, D.M., Brownlees, J., Ackerley, S.,
Leigh, P.N., Shaw, C.E., Miller, C.C.J., 2001. p35/cdk5 binds and
phosphorylates h-catenin and regulates h-catenin/presenilin-1 interac-
tion. Eur. J. Neurosci. 13, 241–247.
Kim, J.H., Park, K.C., Chung, S.S., Bang, O., Chung, C.H., 2003.
Deubiquitinating enzymes as cellular regulators. J. Biochem. 134, 9–18.
Kitamura, K., Yanazawa, M., Sugiyama, N., Miura, H., Iizuka-Kogo, A.,
Kusaka, M., Omichi, K., Suzuki, R., Kato-Fukui, Y., Kamiirisa, K.,
Matsuo, M., Kamijo, S.I., Kasahara, M., Yoshioka, H., Ogata, T.,
Fukuda, T., Kondo, I., Kato, M., Dobyns, W.B., Yokoyama, M.,
Morohashi, K.I, 2002. Mutation of ARX causes abnormal development
of forebrain and testes in mice and X-linked lissencephaly with
abnormal genitalia in humans. Nat. Genet. 32, 359–369.
Kizhatil, K., Wu, Y., Sen, A., Bennett, V., 2002. A new activity of
Doublecortin in recognition of the phospho-FIGQY tyrosine in the
cytoplasmic domain of Neurofascin. J. Neurosci. 22, 7948–7958.
Kwon, Y.T., Gupta, A., Zhou, Y., Nikolic, M., Tsai, L.H., 2000. Regulation
of N-cadherin-mediated adhesion by the p35-cdk5 kinase. Curr. Biol.
10, 363–372.
Li, M., Chen, D., Shiloh, A., Luo, J., Nikolaev, A.Y., Qin, J., Gu, W., 2002.
Deubiquitination of p53 by HAUSP is an important pathway for p53
stabilization. Nature 416, 648–653.
Mandai, K., Nakanishi, H., Satoh, A., Obaishi, H., Wada, M., Nishioka, H.,
Itoh, M., Mizoguchi, A., Aoki, T., Fujimoto, T., Matsuda, Y., Tsukita,
S., Takai, Y., 1997. Afadin: a novel actin filament-binding protein with
one PDZ domain localized at cadherin-based cell-to-cell adherens
junction. J. Cell Biol. 139, 517–528.
Michele, D.E., Barresi, R., Kanagawa, M., Saito, F., Cohn, R.D., Satz, J.S.,
Dollar, J., Nishino, I., Kelley, R.I., Somer, H., Straub, V., Mathews,
K.D., Moore, S.A., Campbell, K.P., 2002. Post-translational disruption
of dystroglycan-ligand interactions in congenital muscular dystrophies.
Nature 418, 417–422.
Moore, S.A., Saito, F., Chen, J., Michele, D.E., Henry, M.D., Messing,
A., Cohn, R.D., Ross-Barta, S.E., Westra, S., Williamson, R.A.,
Hoshi, T., Campbell, K.P., 2002. Deletion of brain dystroglycan
recapitulates aspects of congenital muscular dystrophy. Nature 418,
422–425.
Murray, R.Z., Jolly, L.A., Wood, S.A., 2004. The FAM deubiquitilating
enzyme localizes to multiple points of protein trafficking in epithelia,
where it associates with E-cadherin and h-catenin. Mol. Biol. Cell 15,
1591–1599.
Niethammer, M., Smith, D.S., Ayala, R., Peng, J., Ko, J., Lee, M.S.,
Morabito, M., Tsai, L.H., 2000. NUDEL is a novel Cdk5
substrate that associates with LIS1 and cytoplasmic dynein. Neuron
28, 697–711.
Nowak, L., Ascher, P., Berwald-Netter, Y., 1987. Ionic channels in mouse
astrocytes in culture. J. Neurosci. 7, 101–109.
Oshima, T., Ward, J.M., Huh, C.G., Longenecker, G., Veeranna, Pant, H.C.,
Brady, R.O., Martin, L.J., Kulkarni, A.B., 1996. Targeted disruption of
the cyclin-dependent kinase 5 gene results in abnormal corticogenesis,
neuronal pathology and perinatal death. Proc. Natl. Acad. Sci. U. S. A.
93, 11173–11178.
Patrick, G.N., Zhou, P., Kwon, Y.T., Howley, P.M., Tsai, L.H., 1998. p35,
the neuronal-specific activator of cyclin-dependent kinase 5 (cdk5) is
degraded by the ubiquitin-proteasome pathway. J. Biol. Chem. 273,
24057–24064.
Reiner, O., Carrozzo, R., Shen, Y., Wehnert, M., Faustinella, F., Dobyns,
W.B., Caskey, C.T., Ledbetter, D.H., 1993. Isolation of a Miller–Dieker
lissencephaly gene containing G protein beta-subunit-like repeats.
Nature 364, 717–721.
Sapir, T., Elbaum, M., Reiner, O., 1997. Reduction of microtubule
catastrophe events by LIS1, platelet-activating factor acetylhydrolase
subunit. EMBO J. 16, 6977–6984.
Sapir, T., Horesh, D., Caspi, M., Atlas, R., Burgess, H.A., Wolf, S.G.,
Francis, F., Chelly, J., Elbaum, M., Pietrokovski, S., Reiner, O., 2000.
Doublecortin mutations cluster in evolutionarily conserved functional
domains. Hum. Mol. Genet. 9, 703–712.
Sheldon, M., Rice, D.S., D’Arcangelo, G., Yoneshima, H., Nakajima, K.,
Mikoshiba, K., Howell, B.W., Cooper, J.A., Goldowitz, D., Curran, T.,
1997. Scrambler and yotari disrupt the disabled gene and produce a
reeler-like phenotype in mice. Nature 389, 730–733.
Smith, D.S., Tsai, L.H., 2002. Cdk5 behind the wheel: a role in trafficking
and transport? Trends Cell Biol. 12, 28–36.
Tan, T.C., Valova, V.A., Malladi, C.S., Graham, M.E., Berven, L.A., Jupp,
O.J., Hansra, G., McClure, S.J., Sarcevic, B., Boadle, R.A., Larsen,
M.R., Cousin, M.A., Robinson, P.J., 2003. Cdk5 is essential for
synaptic vesicle endocytosis. Nat. Cell Biol. 5, 701–710.
Tanaka, T., Serneo, F.F., Tseng, H.C., Kulkarni, A.B., Tsai, L.H., Gleeson,
J.G., 2004. Cdk5 phosphorylation of doublecortin ser297 regulates its
effect on neuronal migration. Neuron 41, 215–227.
Taya, S., Yamamoto, T., Kano, K., Kawano, Y., Iwamatsu, A., Tsuchiya, T.,
Tanaka, K., Kanai-Azuma, M., Wood, S.A., Mattick, J.S., Kaibuchi, K.,
1998. The Ras target AF-6 is a substrate of the fam deubiquitinating
enzyme. J. Cell Biol. 142, 1053–1062.
Taya, S., Yamamoto, T., Kanai-Azuma, M., Wood, S.A., Kaibuchi, K.,
1999. The deubiquitinating enzyme Fam interacts with and stabilizes h-catenin. Genes Cells 4, 757–767.
Taylor, K.R., Holzer, A.K., Bazan, J.F., Walsh, C.A., Gleeson, J.G., 2000.
Patient mutations in doublecortin define a repeated tubulin-binding
domain. J. Biol. Chem. 275, 34442–34450.
Trockenbacher, A., Suckow, V., Foerster, J., Winter, J., Krauss, S., Ropers,
H., Schneider, R., Schweiger, S., 2001. MID1, mutated in Opitz
syndrome encodes an ubiquitin ligase that targets phosphatase 2A for
degradation. Nat. Genet. 29, 287–294.
Trommsdorff, M., Gotthardt, M., Hiesberger, T., Shelton, J., Stockinger, W.,
Nimpf, J., Hammer, R.E., Richardson, J.A., Herz, J., 1999. Reeler/
Disabled-like disruption of neuronal migration in knockout mice
lacking the VLDL receptor and ApoE receptor 2. Cell 97, 689–701.
Ware, M.L., Fox, J.W., Gonzalez, J.L., Davis, N.M., Lambert de Rouvroit,
C., Russo, C.J., Chua, S.C.J., Goffinet, A.M., Walsh, C.A., 1997.
Aberrant splicing of a mouse disabled homolog, mdab1, in the
scrambler mouse. Neuron 19, 239–249.
Wendland, B., 2002. Epsins: adaptors in endocytosis? Nat. Rev., Mol. Cell
Biol. 3, 971–977.




















