
Racial Disparities in Access to Mortgage Credit: Does Governance Matter?
Upload
khangminh22Category
view
0download
0
Does size matter in females? An overview of the impact ofthe high variation in the ovarian reserve on ovarian functionand fertility, utility of anti-Mullerian hormone as adiagnostic marker for fertility and causes of variationin the ovarian reserve in cattle
J. J. IrelandA,D,G. W. SmithB,D. ScheetzA, F. Jimenez-KrasselA, J. K. FolgerA,B,J. L. H. IrelandA, F. MossaC, P. LonerganC and A. C. O. EvansC
AMolecular Reproductive Endocrinology Laboratory, Department of Animal Science,
Michigan State University, East Lansing, MI 48824, USA.BLaboratory of Mammalian Reproductive Biology andGenomics, Department of Animal Science,
Michigan State University, East Lansing, MI 48824, USA.CSchool of Agriculture Food Science and Veterinary Medicine, and the Conway Institute,
College of Life Sciences, University College Dublin, Belfield, Dublin 4, Ireland.DCorresponding author. Email: [email protected]
Abstract. The mechanism whereby the inherently high variation in ovary size and the total number of high-qualityoocytes in ovaries (ovarian reserve) impact on ovarian function and fertility, diagnostics to measure the size of the ovarianreserve and the factors that cause variation in the ovarian reserve are unknown. Our results show that cattle can be
phenotyped reliably based on the number of antral follicles growing during follicular waves (antral follicle count, AFC).Young adult cattle with a consistently low v. a high AFC have smaller gonads, a markedly diminished ovarian reserve andmany other phenotypic characteristics usually associated with ovarian aging and infertility. A powerful new approach
based on a single measurement of serum concentration of anti-Mullerian hormone (AMH) is described to test thelongstanding hypothesis that the size of the ovarian reserve is positively associated with fertility. Also, new evidenceshows that maternal environment has a critical role in regulation of the high variation in the ovarian reserve and perhaps
fertility in offspring. These results support the conclusion that the inherently high variation in the ovarian reserve,potentially caused by alterations in the maternal environment, has a negative impact on ovarian function that may result insuboptimal fertility in young adult cattle, and a single AMH measurement can be used reliably in future studies todetermine if fertility is suboptimal in young adult cattle with low circulating AMH concentrations and a correspondingly
diminished ovarian reserve.
Additional keywords: antral follicle count, endometrial thickness, fetal development, follicle numbers, gonadotrophinsecretion, low progesterone, maternal environment, single measurement of anti-Mullerian hormone, superovulation.
Introduction
Does sizematter in females?While it is well established that sizeof testes is a moderately to highly heritable trait (Bourdon and
Brinks 1986; Smith et al. 1989; Kriese et al. 1991) positivelyassociated with fertility in bulls (Coulter and Foote 1979; Smithet al. 1989; Keeton et al. 1996), it is unknown if the very high
variation in size of ovaries and the ovarian reserve (Erickson1966a, 1966b) is also positively associated with fertility infemale cattle. The idea that the quantity of follicles and oocytes
in ovaries impacts on fertility is a long-held tenet in reproductivebiology (Hunter 1787; Erickson 1966b; te Velde and Pearson
2002). However, this fundamental hypothesis has heretoforenever been tested.
Discovery of a linkage between the size of the ovarian
reserve, alterations in ovarian function and fertility in healthyyoung adult cattle would: (1) provide the foundation andcompelling rationale for new basic experiments to unravel the
mechanisms whereby a relatively small ovarian reserve mayalter ovarian function; (2) lead to new diagnostic methods topredict the size of the ovarian reserve and fertility and thus, for
the first time, provide reproductive biologists with a reliablemethod to monitor the impact of various environmental factors
CSIRO PUBLISHING
www.publish.csiro.au/journals/rfd Reproduction, Fertility and Development, 2011, 23, 1–14
� IETS 2011 10.1071/RD10226 1031-3613/11/010001

(e.g. nutrition, toxins, disease) on the ovarian reserve andfertility; and (3) improve the likelihood that better diagnostic,
selection, therapeutic and perhaps genetic methods be devel-oped to enhance fertility in cattle.
Quantity of follicles and oocytes in ovaries is highlyvariable throughout the reproductive life spanof cattle and other species
Overview
Mammals such as cattle (Erickson 1966a, 1966b), swine (Blackand Erickson 1968), sheep (McNatty et al. 1995; Steckler et al.2005) and human beings (Block 1952) are born with a highly
variable number of morphologically healthy follicles andoocytes in ovaries that dwindle rapidly during aging and arenever replenished. Thus, the quantity of follicles and oocytes is
highly variable throughout the reproductive life spans of adults.For example, the number of healthy follicles and oocytes inovaries of newborn calves ranges from 10 000 to 350 000 at birth
(Erickson 1966b) and from 1 920 to 40 960 in 12-month-oldheifers (Ireland et al. 2008). Moreover, by 1 year of age, cowshave lost ,80% of their original stock of healthy oocytes
(Erickson 1966b). As oocyte quantity rapidly declines duringaging, average conception rate following artificial insemination(AI) of Holstein heifers declines from a peak of,56% at 15–16months of age (297 801 breedings) to 42% at 26–27 months
of age (1754 breedings; Kuhn et al. 2006). Furthermore, inwomen, the decline in fertility ,20 years before menopause(cessation of reproductive cycles and depletion of oocytes from
ovaries) and high variability of age at menopause (range: 40–60 years old) are both hypothesised to be caused by the highvariation in the number of follicles and oocytes in women at
birth (Faddy and Gosden 1996; te Velde and Pearson 2002).Nevertheless, the extent and mechanisms whereby the inher-ently high variation in the quantity of follicles and oocytes inovaries of individuals (Block 1952; Erickson 1966a, 1966b;
Burns et al. 2005; Ireland et al. 2007, 2008, 2009; Jimenez-Krassel et al. 2009) may cause or contribute to the relativelylarge differences in fertility amongst young adults are unknown
and have not been previously directly examined for any species.The reason progress in this fundamentally important area ofreproductive biology has heretofore been severely hindered is
primarily because castration and tedious histological analysis ofovaries are required to determine follicle and oocyte numbersand thousands of animals are required to conduct a statistically
valid fertility trial (Pezzullo 2009). Extensive new results fromour laboratory based on serial ovarian ultrasonography of thecow (Burns et al. 2005; Ireland et al. 2007, 2008, 2009; Jimenez-Krassel et al. 2009;Mossa et al. 2010), however, show that these
obstacles have been overcome.The present overview of recent results from our laboratories
will show the following: (1) the high variation in follicle
numbers growing during follicular waves of oestrous cycles inyoung adult cattle can be used to reliably phenotype cattle basedon antral follicle count (Burns et al. 2005; Ireland et al. 2007);
(2) cattle with a relatively low number of follicles growingduring ovarian follicular waves have numerous phenotypic
characteristics usually associated with infertility (Burns et al.2005; Ireland et al. 2007, 2008, 2009; Jimenez-Krassel et al.
2009; Mossa et al. 2010); (3) a single measurement of anti-Mullerian hormone (AMH) concentration in serum is a usefulbiomarker for the relative size of the ovarian reserve (Ireland
et al. 2008); and (4) maternal nutrition and disease duringpregnancy may contribute to the inherently high variation inthe number of follicles growing during follicular waves and
correspondingly the size of the ovarian reserve in their offspring(Mossa et al. 2009).
Evidence that the high variation in the number of folliclesgrowing during follicular waves of oestrous cyclescan be used to reliably phenotype cattle
In our initial study (Burns et al. 2005), the variation in the
number of antral follicles growing during the two or threefollicular waves of oestrous cycles (Ireland et al. 2000) washypothesised to be high because of the inherently high varia-
tion in the total quantity of healthy follicles and oocytes inovaries (Block 1952; Erickson 1966a, 1966b). This hypothesiswas tested by subjecting cattle to once- or twice-daily ovarian
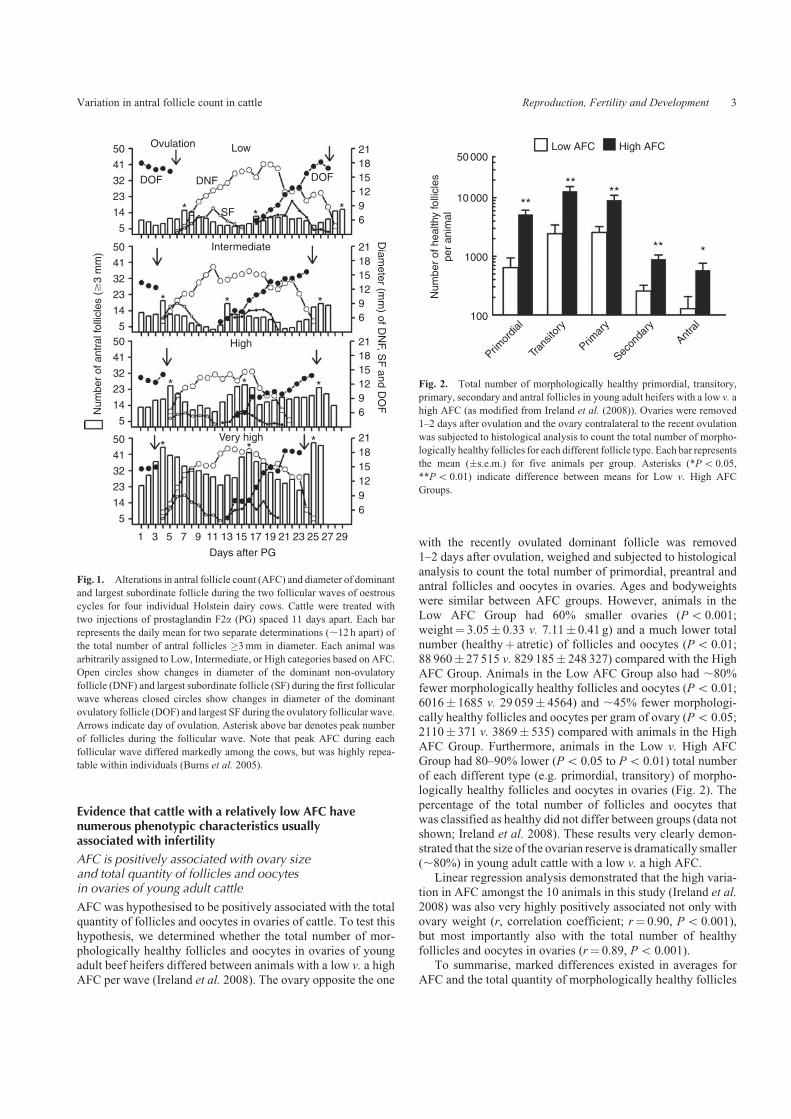
ultrasonography to determine the extent of variation in thenumber of follicles �3mm in diameter growing during thedifferent follicular waves of oestrous cycles. Fig. 1 showsthe results of such analysis for four individual dairy cows.
After analysis of 188 follicular waves for 69 animals in twoseparate studies in Michigan and Ireland (Ireland et al. 2007),we learned that the peak number of follicles growing during
waves (hereafter referred to as the antral follicle count, AFC)varied ,7-fold amongst animals (range: 8–56 follicles perwave), but was surprisingly very highly repeatable (0.84–0.95,
1¼ perfect) within individuals, regardless of (1) their age,breed or country of origin; (2) season or stage of lactation;(3) span of time between AFC measurements in the sameindividuals (up to 15 months); or (4) the ultrasound technician
conducting the ovarian ultrasonography (Burns et al. 2005;Ireland et al. 2007). Repeatability of AFC per wave has beendetermined in over one hundred additional cattle (F. Jimenez
and J. J. Ireland, unpubl. data) with results similar to our firsttwo reports (Burns et al. 2005; Ireland et al. 2007). Thisimportant finding demonstrated for the first time that cattle
could be phenotyped reliably based on AFC. Henceforth, cattlewere arbitrarily classified into three groups based on averageAFC during two to four consecutive follicular waves. Animals
in the Low Group consistently had an AFC of �15 follicles perwave whereas those in the Intermediate Group had 16 to 24follicles per wave and those in the High Group had �25 fol-licles per wave. Typically ,15% to 20% of individuals in a
herd were classified into the Low or High AFC Group whereasthe remainder was in the Intermediate Group (Burns et al.
2005; Ireland et al. 2007, 2008, 2009; Jimenez-Krassel et al.
2009; Mossa et al. 2010). Although initial studies demon-strated that cattle could be phenotyped reliably based on AFC,it was unknown whether the high variation in AFC among
young adult cattle was also associated with the size of theovarian reserve and other potential indices of fertility.
2 Reproduction, Fertility and Development J. J. Ireland et al.
Evidence that cattle with a relatively low AFC havenumerous phenotypic characteristics usuallyassociated with infertility
AFC is positively associated with ovary sizeand total quantity of follicles and oocytesin ovaries of young adult cattle
AFC was hypothesised to be positively associated with the totalquantity of follicles and oocytes in ovaries of cattle. To test this
hypothesis, we determined whether the total number of mor-phologically healthy follicles and oocytes in ovaries of youngadult beef heifers differed between animals with a low v. a high
AFC per wave (Ireland et al. 2008). The ovary opposite the one
with the recently ovulated dominant follicle was removed1–2 days after ovulation, weighed and subjected to histological
analysis to count the total number of primordial, preantral andantral follicles and oocytes in ovaries. Ages and bodyweightswere similar between AFC groups. However, animals in the
Low AFC Group had 60% smaller ovaries (Po 0.001;weight¼ 3.05� 0.33 v. 7.11� 0.41 g) and a much lower totalnumber (healthyþ atretic) of follicles and oocytes (Po 0.01;88 960� 27 515 v. 829 185� 248 327) compared with the High
AFC Group. Animals in the Low AFC Group also had ,80%fewer morphologically healthy follicles and oocytes (Po 0.01;6016� 1685 v. 29 059� 4564) and ,45% fewer morphologi-
cally healthy follicles and oocytes per gram of ovary (Po 0.05;2110� 371 v. 3869� 535) compared with animals in the HighAFC Group. Furthermore, animals in the Low v. High AFC
Group had 80–90% lower (Po 0.05 to Po 0.01) total numberof each different type (e.g. primordial, transitory) of morpho-logically healthy follicles and oocytes in ovaries (Fig. 2). The
percentage of the total number of follicles and oocytes thatwas classified as healthy did not differ between groups (data notshown; Ireland et al. 2008). These results very clearly demon-strated that the size of the ovarian reserve is dramatically smaller
(,80%) in young adult cattle with a low v. a high AFC.Linear regression analysis demonstrated that the high varia-
tion in AFC amongst the 10 animals in this study (Ireland et al.
2008) was also very highly positively associated not only withovary weight (r, correlation coefficient; r¼ 0.90, Po 0.001),but most importantly also with the total number of healthy
follicles and oocytes in ovaries (r¼ 0.89, Po 0.001).To summarise, marked differences existed in averages for
AFC and the total quantity of morphologically healthy follicles
Num
ber
of h
ealth
y fo
llicl
espe
r an
imal
Primor
dial
Trans
itory
Primar
y
Secon
dary
Antra
l100
1000
10 000
50 000Low AFC High AFC
***
****
**
Fig. 2. Total number of morphologically healthy primordial, transitory,
primary, secondary and antral follicles in young adult heifers with a low v. a
high AFC (as modified from Ireland et al. (2008)). Ovaries were removed
1–2 days after ovulation and the ovary contralateral to the recent ovulation
was subjected to histological analysis to count the total number of morpho-
logically healthy follicles for each different follicle type. Each bar represents
the mean (�s.e.m.) for five animals per group. Asterisks (*Po 0.05,
**Po 0.01) indicate difference between means for Low v. High AFC
Groups.
Days after PG
Diam
eter (mm
) of DN
F, SF
and DO
F
6912151821
6912151821
6912151821
6912151821
DOF DOF
***
* * *
***
* * *
DNF
SF
Ovulation Low
Intermediate
High
Very high
5
14
23
32
41
50
5
14
23
32
41
505
14
23
32
41
50
5
14
23
32
41
50
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29
Num
ber
of a
ntra
l fol
licle
s (�
3 m
m)
Fig. 1. Alterations in antral follicle count (AFC) and diameter of dominant
and largest subordinate follicle during the two follicular waves of oestrous
cycles for four individual Holstein dairy cows. Cattle were treated with
two injections of prostaglandin F2a (PG) spaced 11 days apart. Each bar
represents the daily mean for two separate determinations (,12 h apart) of
the total number of antral follicles �3mm in diameter. Each animal was
arbitrarily assigned to Low, Intermediate, or High categories based on AFC.
Open circles show changes in diameter of the dominant non-ovulatory
follicle (DNF) and largest subordinate follicle (SF) during the first follicular
wave whereas closed circles show changes in diameter of the dominant
ovulatory follicle (DOF) and largest SF during the ovulatory follicular wave.
Arrows indicate day of ovulation. Asterisk above bar denotes peak number
of follicles during the follicular wave. Note that peak AFC during each
follicular wave differed markedly among the cows, but was highly repea-
table within individuals (Burns et al. 2005).
Variation in antral follicle count in cattle Reproduction, Fertility and Development 3

and oocytes in ovaries between animals in the Low v.High AFCGroups. Moreover, there was a very high degree of correlation
between these same variables. Taken together, these dataprovided for the first time strong evidence that AFC reflectsreliably the relative quantity of morphologically healthy folli-
cles and oocytes in ovaries of young adult cattle. However,whether the ,15–20% of young adult cattle in a herd thattypically have a relatively lowAFC,much smaller ovaries, and a
very low quantity of healthy follicles and oocytes in ovaries alsohave suboptimal fertility (hereafter defined as�3 inseminationsper conception) compared with their age-matched herdmateswith a higher AFC is unknown.
Young adult cattle with a low v. a high AFCrespond poorly to superovulation
The mechanisms whereby the high variation in the quantity offollicles and oocytes per se may alter ovarian function and
fertility are unknown. Nevertheless, we have generated intri-guing new results (Ireland et al. 2007, 2008, 2009; Jimenez-Krassel et al. 2009) showing that young adult cattle with a low v.
a high AFC also exhibit a variety of additional phenotypiccharacteristics usually associated with impaired ovarian func-tion in older animals, which are reflective of ovarian aging and
suboptimal fertility. For example, responsiveness to super-ovulation and success of IVF and embryo transfer are wellestablished to be negatively associated with aging and the cor-responding reduction in the number of follicles and oocytes in
ovaries and suboptimal fertility in single-ovulating species likecattle (Kawamata 1994; Cushman et al. 1999; Taneja et al.
2000; Singh et al. 2004) and humans (Beckers et al. 2002).
Consequently, we examined whether the high variation in AFCobserved among age-matched young adult cattle also had animpact on their responsiveness to superovulation (Ireland et al.
2007). Beef heifers with a high or a low AFC were subjected tostandard commercial superovulation procedures and artificialinsemination (AI) and the number of ovulations and recovered
oocytes/embryos evaluated (Stringfellow and Seidel 1998).Results showed that animals with a low AFC also had a muchlower number (8.5� 1.1 v. 17.6� 3.6, Po 0.05) of corporalutea (CL), a reduced number (4.7� 0.7 v. 10.6� 2.7,Po 0.05)
of early embryos/unfertilised oocytes recovered on Day 7 and adiminished number (3.8� 0.8 v. 5.4� 1.3, Po 0.05) of trans-ferable embryos per animal compared with animals in the High
AFC Group (Ireland et al. 2007). This observation documentedfor the first time that young adult cattle with a low v. a high AFCalso had a diminished responsiveness to superovulation and
produced significantly fewer high-quality embryos for embryotransfer, similar to previous findings for older, less fertilecompared with younger cattle (Kawamata 1994; Cushman et al.1999; Taneja et al. 2000; Singh et al. 2004). This finding,
coupled with the observation that animals with a relatively lowAFC also had markedly smaller ovaries and a reduced quantityof morphologically healthy follicles and oocytes (Burns et al.
2005), provided the first evidence, albeit indirect, linking lownumbers of follicles and oocytes in ovaries of young adult cattleto suboptimal fertility independent of age.
Young adult cattle with a low v. a high AFC have chronicallyheightened gonadotrophin secretion
Not only does responsiveness to superovulation and success
of embryo transfer decline, but oocyte quantity and quality,embryo development and fertility also decline as circulatingFSH concentrations increase with aging in cattle (Erickson
1966a, 1966b; Erickson et al. 1976; Royal et al. 2000; Lucy2001; Sartori et al. 2002a, 2002b; Washburn et al. 2002; Malhiet al. 2005) and many years before ovulation and reproductive
cycles cease in women (Block 1951, 1952, 1953; Klein et al.
1996; te Velde et al. 1998; Faddy 2000; te Velde and Pearson2002; Ottolenghi et al. 2004). Like FSH, LH also increasesduring aging of women (Ahmed Ebbiary et al. 1994; Reame
et al. 1996). Moreover, for young women born small for theirage that also have relatively small ovaries and uteri, FSH andLHsecretion are higher compared with their age-matched, normal-
sized counterparts (Ibanez et al. 2003). Thus, chronicallyheightened FSH and LH secretion is the physiological hallmarknot only for ovary size, but also for the corresponding age-
related reduction in follicle and oocyte quantity in ovaries andthe onset of suboptimal fertility in single-ovulating species likecattle. However, it was unknown whether the high variation in
AFC, ovary size and follicle and oocyte quantity, as observed inyoung adult cattle in our studies, was also linked to chronicalterations in gonadotrophin secretion independent of aging.
Based on the aforementioned negative association between
follicle numbers and gonadotrophin secretion during aging,circulating gonadotrophin secretion was hypothesised to beassociated inversely with the variation in AFC in young adult
cattle. The results of our studies showed that young adult beefcattle, and late lactation (Burns et al. 2005; Ireland et al. 2007)and culled non-lactatingHolstein dairy cows (Mossa et al. 2010)
with a low AFC also had 15–50% higher circulating FSHconcentrations (Po 0.05) throughout the follicular waves ofoestrous cycles compared with animals with a high AFC. Aninverse relationship between follicle numbers and FSH concen-
trations has also been reported for dairy heifers (Haughian et al.2004) and lactating beef cattle (Singh et al. 2004) even atunknown stages of follicular waves. Also, we recently demon-
strated that young adult beef cattle with a low AFC have a,2-fold higher (Po 0.05 to Po 0.01) basal LH concentrationand episodic LH secretion on Day 11 of the oestrous cycle
compared with animals with a high AFC (Jimenez-Krassel et al.2009). Taken together, these observations show that gonado-trophin secretion is inversely associated with AFC in cattle.
These findings imply that gonadotrophin secretion is chroni-cally heightened in young adult cattle with a relatively lowAFC,smaller ovaries and a diminished quantity of follicles andoocytes in ovaries compared with their age-matched herdmates
with a higher AFC.Chronically heightened gonadotrophin secretion, such as that
observed during follicular waves of cattle with a low v. a high
AFC in our studies (Burns et al. 2005; Ireland et al. 2007;Jimenez-Krassel et al. 2009), likely results in the desensitisationor uncoupling of gonadotrophin receptors from their respective
signalling system in granulosal, thecal and luteal cells (Contiet al. 1976; Amsterdam et al. 2002), which in turn may alter
4 Reproduction, Fertility and Development J. J. Ireland et al.
ovarian function. Disruption of the gonadotrophin receptorsignalling system, therefore, could, in conjunction with a
diminished ovarian reserve, also explain why responsivenessto superovulation is diminished for cattle with a low AFC andchronically high circulating FSH/LH concentrations (Ireland
et al. 2007).Moreover, relatively high FSHconcentrations in thepresence of insulin in rodents (Eppig et al. 1998) heightenedsecretion of FSH in transgenic rodents (McTavish et al. 2007),
and with superovulation of cattle (Lonergan et al. 1994; Blondinet al. 1996) diminished the developmental competence ofoocytes and fertility. Taken together, these findings provide
new insight into potential mechanisms (e.g. desensitisation)whereby the variation in AFC and corresponding alterationsin gonadotrophin secretion may negatively impact on ovarianfunction. Nevertheless, it was unknown whether the high varia-
tion in AFC and gonadotrophin secretion was also associatedwith significant alterations in ovarian function that may, in turn,impact on fertility in cattle.
Young adult cattle with a low v. a high AFC have lowerconcentrations of progesterone and diminishedendometrial growth during oestrous cycles
Progesterone is a key physiological marker of CL function andthus ovarian function, which also has a well established positiverole in uterine function, embryo development and fertility in
single-ovulating species like cattle (Mann and Lamming 1999;Inskeep 2004; Stronge et al. 2005; McNeill et al. 2006; DiskinandMorris 2008). In support of the crucial role for progesteronein successful reproduction, low circulating progesterone con-
centrations are associated with high rates of embryo mortality incattle (Mann and Lamming 1999; Inskeep 2004; Stronge et al.2005; McNeill et al. 2006; Diskin and Morris 2008) and women
(Daily et al. 1994; Elson et al. 2003) and luteal phase defects andinfertility in women (Pritts and Atwood 2002). Therefore, this
study was designed to determine if the high variation in AFC,which is inversely associated with FSH and LH secretion inyoung adult cattle (Burns et al. 2005; Ireland et al. 2007;
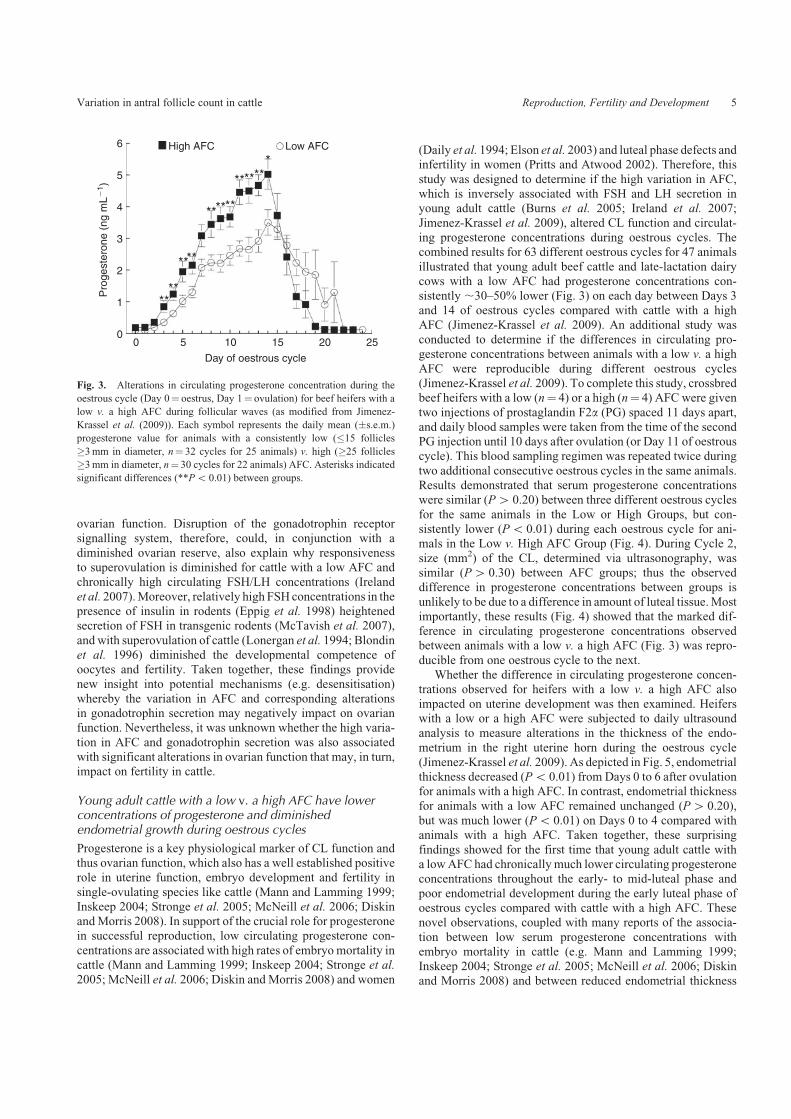
Jimenez-Krassel et al. 2009), altered CL function and circulat-ing progesterone concentrations during oestrous cycles. Thecombined results for 63 different oestrous cycles for 47 animals
illustrated that young adult beef cattle and late-lactation dairycows with a low AFC had progesterone concentrations con-sistently ,30–50% lower (Fig. 3) on each day between Days 3and 14 of oestrous cycles compared with cattle with a high
AFC (Jimenez-Krassel et al. 2009). An additional study wasconducted to determine if the differences in circulating pro-gesterone concentrations between animals with a low v. a high
AFC were reproducible during different oestrous cycles(Jimenez-Krassel et al. 2009). To complete this study, crossbredbeef heifers with a low (n¼ 4) or a high (n¼ 4) AFC were given
two injections of prostaglandin F2a (PG) spaced 11 days apart,and daily blood samples were taken from the time of the secondPG injection until 10 days after ovulation (or Day 11 of oestrouscycle). This blood sampling regimen was repeated twice during
two additional consecutive oestrous cycles in the same animals.Results demonstrated that serum progesterone concentrationswere similar (P4 0.20) between three different oestrous cycles
for the same animals in the Low or High Groups, but con-sistently lower (Po 0.01) during each oestrous cycle for ani-mals in the Low v. High AFC Group (Fig. 4). During Cycle 2,
size (mm2) of the CL, determined via ultrasonography, wassimilar (P4 0.30) between AFC groups; thus the observeddifference in progesterone concentrations between groups is
unlikely to be due to a difference in amount of luteal tissue.Mostimportantly, these results (Fig. 4) showed that the marked dif-ference in circulating progesterone concentrations observedbetween animals with a low v. a high AFC (Fig. 3) was repro-
ducible from one oestrous cycle to the next.Whether the difference in circulating progesterone concen-
trations observed for heifers with a low v. a high AFC also
impacted on uterine development was then examined. Heiferswith a low or a high AFC were subjected to daily ultrasoundanalysis to measure alterations in the thickness of the endo-
metrium in the right uterine horn during the oestrous cycle(Jimenez-Krassel et al. 2009). As depicted in Fig. 5, endometrialthickness decreased (Po 0.01) from Days 0 to 6 after ovulationfor animals with a high AFC. In contrast, endometrial thickness
for animals with a low AFC remained unchanged (P4 0.20),but was much lower (Po 0.01) on Days 0 to 4 compared withanimals with a high AFC. Taken together, these surprising
findings showed for the first time that young adult cattle witha lowAFC had chronically much lower circulating progesteroneconcentrations throughout the early- to mid-luteal phase and
poor endometrial development during the early luteal phase ofoestrous cycles compared with cattle with a high AFC. Thesenovel observations, coupled with many reports of the associa-
tion between low serum progesterone concentrations withembryo mortality in cattle (e.g. Mann and Lamming 1999;Inskeep 2004; Stronge et al. 2005; McNeill et al. 2006; Diskinand Morris 2008) and between reduced endometrial thickness
Pro
gest
eron
e (n
g m
L�1 )
6
5
4
3
2
1
0
Day of oestrous cycle
High AFC
****
**
******
*******
**
Low AFC
0 5 10 15 20 25
Fig. 3. Alterations in circulating progesterone concentration during the
oestrous cycle (Day 0¼ oestrus, Day 1¼ ovulation) for beef heifers with a
low v. a high AFC during follicular waves (as modified from Jimenez-
Krassel et al. (2009)). Each symbol represents the daily mean (�s.e.m.)
progesterone value for animals with a consistently low (�15 follicles
�3mm in diameter, n¼ 32 cycles for 25 animals) v. high (�25 follicles
�3mm in diameter, n¼ 30 cycles for 22 animals) AFC. Asterisks indicated
significant differences (**Po 0.01) between groups.
Variation in antral follicle count in cattle Reproduction, Fertility and Development 5

and infertility in women (Raga et al. 1999; Basir et al. 2002),imply that rates of embryo mortality may be higher in youngadult cattle with a low v. a high AFC and correspondingly
reduced quantity of follicles and oocytes in ovaries. Higherincidences of embryo mortality as a result of chronically lowerprogesterone concentrations during an oestrous cycle, such asthat observed in our studies (Figs 3, 4), provide a reasonable
physiological explanation for the possibility that heifers with arelatively low quantity of follicles and oocytes in ovaries mayalso have suboptimal fertility and thus would require a greater
AI number to conceive compared with their age-matchedherdmates with a higher quantity of follicles and oocytes.
While our studies demonstrated that the chronically heigh-
tened gonadotrophin secretion in young adult cattle with alow v. a high AFC (Burns et al. 2005; Ireland et al. 2007;Jimenez-Krassel et al. 2009; Mossa et al. 2010) may have a
negative impact on CL function (Figs 3, 4), it was unknownwhether the variation in AFC and gonadotrophin secretion also
altered follicular differentiation and function, and perhapsoocyte quality.
Cattle with a low v. a high AFC have a higher abundanceof intrafollicular markers of poor oocyte quality
In our next study (Ireland et al. 2009), the high variation in AFCand the corresponding inverse alterations in circulating gona-dotrophin concentrations were hypothesised to negatively
impact on differentiation and function of follicles growingduring follicular waves and, in turn, on oocyte quality. Toexamine this hypothesis, animals in the Low (n¼ 5 animals) or
High (n¼ 5) AFC Groups were slaughtered at emergence of thefirst follicle wave on Days 2 to 3 of the oestrous cycle (Irelandet al. 2009). Cumulus cells and denuded oocytes were isolated
from the three largest follicles (Bettegowda et al. 2006). Resultsshowed that animals with a low AFC had 2- and 6-fold higherabundance of cathepsin-B (Po 0.01) and cathepsin-S mRNAs(Po 0.10), respectively, in cumulus cells compared with ani-
mals with a high AFC. This finding is significant because recentstudies from our laboratory show that these same mRNA tran-scripts are more abundant in the cumulus cells for adult meta-
phase II bovine oocytes that develop to the blastocyst stage at alow percentage following parthenogenetic activation. More-over, inhibition of cumulus cell cathepsin activity during in vitro
maturation significantly increases the rate of embryonic devel-opment to blastocyst stage in vitro following in vitro fertilisation(Bettegowda et al. 2008). Thus, relatively high expression ofcathepsins in cumulus cells, as observed for animals with low
follicle numbers in the present study, implies that oocyte qualitymay also be compromised. The observation that animals with alow v. a high AFC have much higher expression of the genes for
cathepsins in cumulus cells supports the possibility that the highvariation in follicle and oocyte numbers is also associated withphysiologically significant alterations in follicular differentia-
tion and, in turn, oocyte quality.
180
160
140
120
100
80
60
400
** ****
2
Low High
4 6 8 10
Days after ovulation
End
omet
rial a
rea
(mm
2 )
Fig. 5. Alterations in endometrial thickness for young adult beef heifers
with a low v. a highAFC during follicular waves (asmodified from Jimenez-
Krassel et al. (2009)). Heifers were subjected to daily ultrasound analysis to
measure alterations in the size of the endometrium in the right uterine horn.
Each bar represents the mean (�s.e.m.) for six animals per group. ANOVA
indicated an overall significant effect of group (Po 0.02) and group by
day interaction (Po 0.04). Asterisks indicate significant difference (**Po0.01) between means.
0
400
300
200
100
00 3 6 9 12
3 6 9 12
0 3 6 9 12
0 3 6 9 12
Pro
gest
eron
e (n
g mL�
1 )
Days of oestrous cycle
Cycle 3
Cycle 2
**** **
** **
**
****
****
*
**
**
*
* **
**
CL size (mm2)
Cycle 1
Low High8
6
4
2
0
8
6
4
2
0
8
6
4
2
0
Fig. 4. Reproducibility of circulating progesterone concentrations during
three consecutive oestrous cycles for the same beef heifers with a low (n¼ 4)
v. a high (n¼ 4) AFC (as modified from Jimenez-Krassel et al. (2009)).
Symbols represent mean (�s.e.m.). ANOVA indicated serum progesterone
concentrations were similar (P4 0.20) during the three different oestrous
cycles (Cycles 1, 2, 3) for animals in the Low or High Groups, but differed
(Po 0.05) between groups during each cycle. Asterisks indicate significant
difference (*Po 0.05, **Po 0.01) between groups. The insert in the mid
panel depicts alterations in CL size (mm2) based on results of daily ovarian
ultrasonography of Cycle 2. ANOVA indicated CL size was similar
(P4 0.20) between AFC groups.
6 Reproduction, Fertility and Development J. J. Ireland et al.

Evidence that a single measurement of AMH duringthe oestrous cycle is reflective of AFC, ovary sizeand total quantity of follicles and oocytesin ovaries of young adult cattle
The most reliable and direct means available to determine if
fertility is suboptimal in individual young adult cattlewith a low v.
a high AFC is based on AI number required for pregnancy.However, because of the binary nature of fertility assessment
(pregnant v. not pregnant) after AI, thousands of animals arerequired to ensure that a sufficient number of animals have a lowor a high AFC to firmly establish if a statistically significant
difference exists in fertility (as measured by AI number perpregnancy) between the two groups. Consequently, the use ofintensive serial ovarian ultrasonography to accurately determineAFC, and corresponding quantity of morphologically healthy
follicles and oocytes in ovaries, for the large number of animalsrequired to conduct a statistically valid fertility trial would beimpractical. Moreover, in our experience, large, well managed
cattle operations are unwilling to cooperate in research projectsif the investigator’s requirements for animal handling sig-nificantly disrupt their management practices. To attempt to
circumvent these problems, we examined the possibility thatcollection of a single blood sample during the oestrous cycle andmeasurement of serum anti-Mullerian hormone (AMH) con-centration would be useful as a reliable biomarker reflective of
AFC, ovary size and total quantity of follicles and oocytes inovaries of young adult cattle.
In females, AMH, a member of the TGFß superfamily of
growth factors, is produced exclusively by granulosa cells ofhealthy growing follicles (La Marca and Volpe 2006). Circulat-ing AMH concentrations are positively associated with the total
number of healthy follicles and oocytes in ovaries of mice(Kevenaar et al. 2006) and with follicle numbers in women(van Rooij et al. 2002; Scheffer et al. 2003; Fanchin et al. 2005;
La Marca and Volpe 2006). Furthermore, AMH concentrationsdecrease coincident with the decline in the number of follicles asrodents (Kevenaar et al. 2006) and women (Piltonen et al. 2005;
van Rooij et al. 2005) age. To determine if AMH concentrationswere also positively associated with the high variation in AFCin cattle, a commercially available human AMH kit was used tomeasure serum AMH concentrations in cattle (Ireland et al.
2008). Serial ovarian ultrasonography was used to identifyyoung adult beef cattle with low, intermediate or high AFC(Ireland et al. 2008). Serum samples were obtained daily,
starting 6 to 8 days preceding ovulation, which correspondedwith ovulatory follicular waves, until the day of ovulation andanalysed for AMH. In the left panel of Fig. 6, data for each
animal were aligned relative to day of ovulation for statisticalanalysis, and each point represents the mean (�s.e.m.) for fouror eight animals in the Low (�15 follicles�3mm in diameter),
AM
H (
ng m
L�1 )
0.50
0.40
0.30
0.20
0.10
0.00
0.50
0.40
0.30
0.20
0.10
0.00�8 �7 �6 �5 �4 �3 �2 �1
r � 0.88(n � 16)
High (n � 4)
Intermediate (n � 8)
Low (n � 4)
0
Days before ovulation Antral follicle count5 11 17 23 29 35
Fig. 6. Alterations in circulating AMH concentrations during ovulatory follicular waves
in young adult beef heifers with a consistently low (�5 follicles�3mm in diameter, n¼ 4
animals), intermediate (16–24 follicles, n¼ 8 animals) or high (�25 follicles, n¼ 4
animals) AFC during follicular waves (as modified from Ireland et al. (2008)). PG was
injected twice 11 days apart to synchronise oestrus, and ovarian ultrasonography was used
to confirm that animals were correctly phenotyped based on AFC. Daily blood samples
taken 6 to 8 days preceding ovulation and on the day of ovulation were analysed for AMH
concentration. In the left panel, data for each animal were aligned relative to day of
ovulation and plotted relative to results of linear regression analysis. Each point depicts the
mean (�s.e.m.) for four to eight animals. In the right panel, averageAMHconcentration per
animal was plotted relative to the average peak number of antral follicles per wave during
oestrous cycles for each animal; r¼ correlation coefficient and n¼ total number of animals.
Variation in antral follicle count in cattle Reproduction, Fertility and Development 7
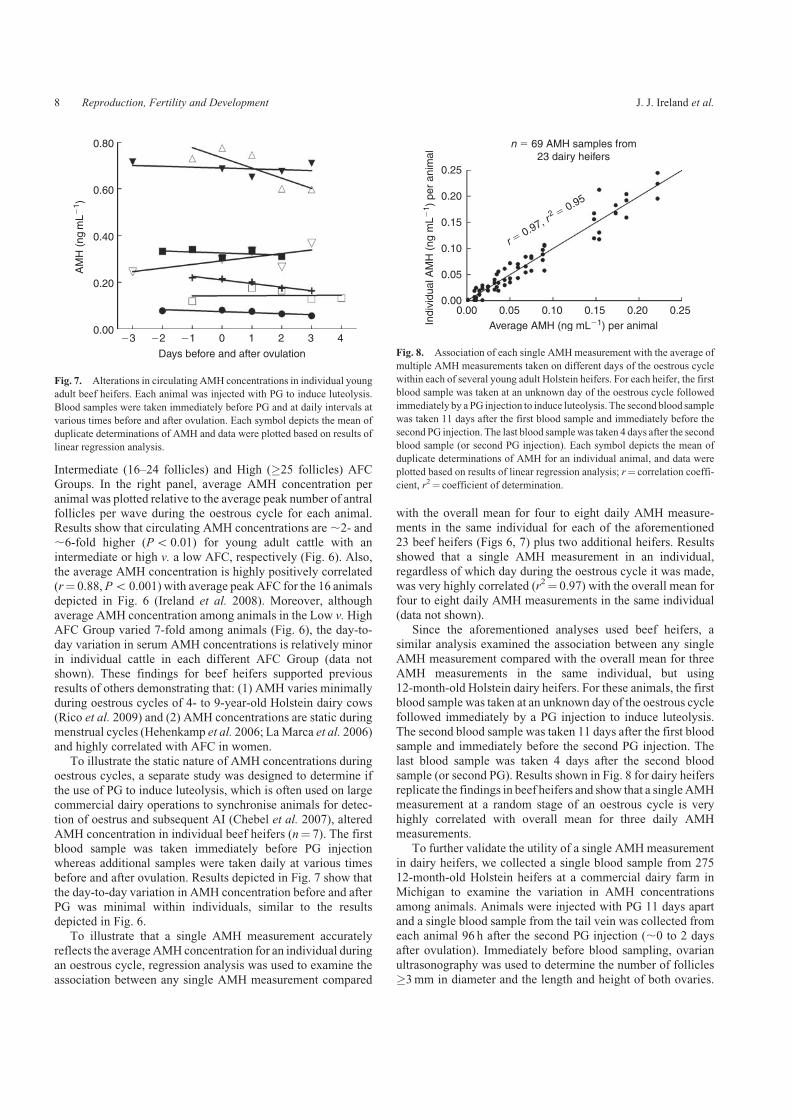
Intermediate (16–24 follicles) and High (�25 follicles) AFCGroups. In the right panel, average AMH concentration per
animal was plotted relative to the average peak number of antralfollicles per wave during the oestrous cycle for each animal.Results show that circulating AMH concentrations are,2- and,6-fold higher (Po 0.01) for young adult cattle with an
intermediate or high v. a low AFC, respectively (Fig. 6). Also,the average AMH concentration is highly positively correlated(r¼ 0.88, Po 0.001) with average peak AFC for the 16 animals
depicted in Fig. 6 (Ireland et al. 2008). Moreover, althoughaverage AMH concentration among animals in the Low v. HighAFC Group varied 7-fold among animals (Fig. 6), the day-to-
day variation in serum AMH concentrations is relatively minorin individual cattle in each different AFC Group (data notshown). These findings for beef heifers supported previous
results of others demonstrating that: (1) AMH varies minimallyduring oestrous cycles of 4- to 9-year-old Holstein dairy cows(Rico et al. 2009) and (2) AMH concentrations are static duringmenstrual cycles (Hehenkamp et al. 2006; LaMarca et al. 2006)
and highly correlated with AFC in women.To illustrate the static nature of AMH concentrations during
oestrous cycles, a separate study was designed to determine if
the use of PG to induce luteolysis, which is often used on largecommercial dairy operations to synchronise animals for detec-tion of oestrus and subsequent AI (Chebel et al. 2007), altered
AMH concentration in individual beef heifers (n¼ 7). The firstblood sample was taken immediately before PG injectionwhereas additional samples were taken daily at various timesbefore and after ovulation. Results depicted in Fig. 7 show that
the day-to-day variation in AMH concentration before and afterPG was minimal within individuals, similar to the resultsdepicted in Fig. 6.
To illustrate that a single AMH measurement accuratelyreflects the averageAMH concentration for an individual duringan oestrous cycle, regression analysis was used to examine the
association between any single AMH measurement compared
with the overall mean for four to eight daily AMH measure-
ments in the same individual for each of the aforementioned23 beef heifers (Figs 6, 7) plus two additional heifers. Resultsshowed that a single AMH measurement in an individual,
regardless of which day during the oestrous cycle it was made,was very highly correlated (r2¼ 0.97) with the overall mean forfour to eight daily AMH measurements in the same individual
(data not shown).Since the aforementioned analyses used beef heifers, a
similar analysis examined the association between any singleAMH measurement compared with the overall mean for three
AMH measurements in the same individual, but using12-month-old Holstein dairy heifers. For these animals, the firstblood sample was taken at an unknown day of the oestrous cycle
followed immediately by a PG injection to induce luteolysis.The second blood sample was taken 11 days after the first bloodsample and immediately before the second PG injection. The
last blood sample was taken 4 days after the second bloodsample (or second PG). Results shown in Fig. 8 for dairy heifersreplicate the findings in beef heifers and show that a singleAMHmeasurement at a random stage of an oestrous cycle is very
highly correlated with overall mean for three daily AMHmeasurements.
To further validate the utility of a single AMHmeasurement
in dairy heifers, we collected a single blood sample from 27512-month-old Holstein heifers at a commercial dairy farm inMichigan to examine the variation in AMH concentrations
among animals. Animals were injected with PG 11 days apartand a single blood sample from the tail vein was collected fromeach animal 96 h after the second PG injection (,0 to 2 days
after ovulation). Immediately before blood sampling, ovarianultrasonography was used to determine the number of follicles�3mm in diameter and the length and height of both ovaries.
AM
H (
ng m
L�1 )
0.80
0.60
0.40
0.20
0.00
Days before and after ovulation
�3 �2 �1 0 1 2 3 4
Fig. 7. Alterations in circulating AMH concentrations in individual young
adult beef heifers. Each animal was injected with PG to induce luteolysis.
Blood samples were taken immediately before PG and at daily intervals at
various times before and after ovulation. Each symbol depicts the mean of
duplicate determinations of AMH and data were plotted based on results of
linear regression analysis.
0.25
r � 0.97, r
2 � 0.950.20
0.15
0.10
0.05
0.000.00 0.05
Average AMH (ng mL�1) per animal
n � 69 AMH samples from23 dairy heifers
Indi
vidu
al A
MH
(ng
mL�
1 ) p
er a
nim
al
0.10 0.15 0.20 0.25
Fig. 8. Association of each single AMHmeasurement with the average of
multiple AMH measurements taken on different days of the oestrous cycle
within each of several young adult Holstein heifers. For each heifer, the first
blood sample was taken at an unknown day of the oestrous cycle followed
immediately by a PG injection to induce luteolysis. The second blood sample
was taken 11 days after the first blood sample and immediately before the
second PG injection. The last blood samplewas taken 4 days after the second
blood sample (or second PG injection). Each symbol depicts the mean of
duplicate determinations of AMH for an individual animal, and data were
plotted based on results of linear regression analysis; r¼ correlation coeffi-
cient, r2¼ coefficient of determination.
8 Reproduction, Fertility and Development J. J. Ireland et al.

The frequency distribution plotted in Fig. 9 shows the numberof animals and their corresponding averages (�s.e.m.) forthe total ovary area (3.1416� (length/2)� (height/2)) and the
number of follicles for the different groups of animals atevery 0.020 ngmL�1 increment of AMH from 0.005 to40.105 ngmL�1. Results showed that AMH levels were
highly variable among dairy heifers and ranged from 0.006to 0.433 ngmL�1 (data not shown), which is well above the0.0015 ngmL�1 limit of sensitivity of the AMH assay. Fre-
quency distribution analysis showed that 20% (n¼ 54) of dairyheifers had AMH levels between 0.005 and 0.025 ngmL�1
while 18% (n¼ 49) had relatively very high (40.105 ngmL�1)
AMH concentrations (Fig. 9), which, as mentioned earlier, issimilar to the proportion of young adult beef heifers in a herdexpected to have a low v. a high AFC and correspondingly arelatively low v. a high AMH concentration (Burns et al. 2005;
Ireland et al. 2007, 2008, 2009). Furthermore, as only a singleultrasound scan of ovaries was performed to count follicles,peak AFC per wave was not determined. Nevertheless, the
heifers with the lowest AMH (0.005–0.025 ngmL�1) also hada ,50% lower average number of follicles (11.8� 0.62 v.
23.8� 1.3, Po 0.0001) and smaller ovaries (457� 20 v.
583� 16mm2, Po 0.0001) compared with animals with thehighest (40.105 ngmL�1) AMH concentrations (Fig. 9). Thesenew results in Holstein heifers (Fig. 9), coupled with ourprevious results in beef and dairy heifers (Figs 6–8), further
corroborate the usefulness of a single AMH measurement topredict relative AFC and size of ovaries and correspondingly the
size of the ovarian reserve in individual dairy heifers (Irelandet al. 2007, 2008, 2009; Jimenez-Krassel et al. 2009).Moreover,
the AMH concentrations were highly variable (e.g. ,72-fold)among the Holstein heifers and correspondingly positivelyassociated with follicle number and ovary size (Fig. 9). Such
variation is more than sufficient to experimentally test thehypothesis that the inherently high variation in AMH concen-tration amongHolstein heifers is positively associated with their
fertility in future fertility trials.To summarise, our previous studies show that a high degree
of correlation exists between AMH, AFC and the number ofhealthy follicles and oocytes in ovaries (Ireland et al. 2008).
Moreover, the day-to-day variation in AMH concentrationsduring reproductive cycles of beef and dairy cattle (Figs 2–4;Ireland et al. 2008; Rico et al. 2009) and women (Hehenkamp
et al. 2006; La Marca et al. 2006) is minimal. Taken together,these results clearly illustrate that a single blood sample taken onany day of the oestrous cycle to measure serum AMH concen-
tration is a reliable phenotypic marker to predict the relativenumber of morphologically healthy follicles and oocytes inovaries and potentially the fertility of individual young adultcattle in future fertility trials.
Evidence that maternal nutrition or disease may contributeto the inherently high variation in the number of folliclesgrowing during follicular waves and corresponding sizeof the ovarian reserve in offspring
Nutrition and disease
The ‘Barker Hypothesis’ states that environmental influencesearly in fetal life impair embryonic development leading toincreased risk for diseases once newborns become adults
(Barker 1992). While most studies have examined the linkagebetween maternal environment during fetal development withonset of cardiovascular disease, obesity and diabetes in off-spring as adults (Gluckman and Hanson 2004; Mcmillen and
Robinson 2005; de Boo and Harding 2006), only a few studieshave examined the impact of maternal environment on repro-duction. For example, although Gardner and colleagues con-
clude that there is only a minimal effect of maternal nutrition onsubsequent reproductive performance in adults (Gardner et al.2009), several studies show that poor fetal growth as a result of
alterations in the maternal diet not only hamper fetal develop-ment, but also suppress gonadotrophin gene expression in thepituitary and the number of ovarian follicles in lambs (Da Silvaet al. 2002) and retards fetal ovarian development (Borwick
et al. 1997). We have recently tested the hypothesis thatalterations in maternal diet inhibit follicular development byrestricting nutrition of beef heifers to 0.6 of their maintenance
energy requirements shortly before conception to the end of thefirst trimester of pregnancy. The first trimester of pregnancycoincides with the peak in the number of follicles and oocytes in
fetal ovaries (Erickson 1966a). Results show that, althoughweight of offspring is unaltered at birth (data not shown), AFCin calves born to nutritionally restricted mothers is 60% lower
(Po 0.01) compared with calves born to control mothers(Mossa et al. 2009; Table 1). Because AFC is positively corre-lated with the size of the ovarian reserve (Ireland et al. 2008),these preliminary findings imply that maternal nutrition may
Ova
ry a
rea
(mm
2 )
Num
ber
of fo
llicl
es(�
3m
m in
dia
met
er)
0.00
5–0.
025
0.02
6–0.
045
0.04
6–0.
065
0.06
6–0.
085
0.08
6–0.
105
�0.
105
200 5
10
15
20
25
30
AMH (ng mL�1)
224
248
272
296
54
***(20%) 61
(22%)52
(19%)42
(15%)17
(6%) 49(18%)
320
Fig. 9. Frequency distribution depicting ovary size and number of follicles
for dairy heifers with different AMH concentrations. AMH concentration in
serum was determined in a single blood sample obtained 96 h after an
injection of PG (0 to 2 days after ovulation) from275, 12-month-oldHolstein
heifers. Just before the PG injection, ovarian ultrasonography was used to
determine the length and height of both ovaries and the total number of
follicles�3mm in diameter. Bars represent the averages (�s.e.m.) for total
ovary area (solid bars) and number of follicles (open bars) for different
groups of animals at each 0.02 ngmL�1 increment of AMH from 0.005
to 40.105 ngmL�1. Number above bar indicates number of animals and
number within parentheses indicates proportion of herd. Asterisks indicate
significant (Po 0.0001) difference compared with the group of cattle with
the highest AMH concentration.
Variation in antral follicle count in cattle Reproduction, Fertility and Development 9

have a previously unrecognised yet important role in the reg-ulation of the size of the ovarian reserve and perhaps fertility of
cattle.We have also examined the effect of disease during preg-
nancy in cattle, which could have a crucial role in fetal devel-
opment and the subsequent health of offspring (Gluckman andHanson 2004), including effects on ovary development andfunction. The purpose of this preliminary study (Ireland et al.
2010) was to determine if a persistent mammary gland infection
and corresponding chronically high somatic cell count (SCC) inmilk of pregnant dairy cows had a negative impact on ovariandevelopment and function of their offspring.
Somatic cells normally present in milk of dairy cows arepredominantly white blood cells. The major factor affectingSCC inmilk is amammary gland infection. SCC inmilk samples
collected from individual dairy cows at various intervals (e.g.monthly) during lactation are commercially determined andrecorded in Dairy Herd Improvement (DHI) records for many
dairy herds in the USA. Consequently, high SCC in milk (e.g.4200 000) is useful as an index for potential previous or currentudder infections or inflammation that may have been caused bymastitis or injury (Caraviello et al. 2005). It is well known that
infections such asmastitis reducemilk production (Bartlett et al.1990; Caraviello et al. 2005), and cows with chronically veryhigh SCC (e.g.4500 000) should probably be culled to enhance
herd profitability (Lehenbauer and Oltjen 1998; Caraviello et al.2005).
Important new preliminary data from our laboratory show
for the first time that dairy cows with a high number of SCCmeasurements exceeding 200 000 may produce daughters withdiminished ovarian function and suboptimal fertility as adults.In our recent study on a commercial dairy farm in Michigan,
serum concentrations of AMH were determined in 192,12-month-old Holstein heifers. We tested whether serum con-centration of AMH and correspondingly ovary size, total num-
ber of follicles and oocytes in ovaries and ovarian function ofeach heifer were correlated with the number of SCC measure-ments 4200 000 for each heifer’s dam. As previously men-
tioned, the number of SCC measurements per individual cowis an accurate index of recurrent udder infections, injury orinflammation (Green et al. 2004; Caraviello et al. 2005), and a
total of 5–7measurements of SCCweremade for each dairy cowbeginning 2 months before and during her pregnancy in thepresent study. The number of SCCmeasurements4200 000 percow ranged from 0 to 5, and cows were separated into groups
based on the number of their SCC measurements 4200 000.
The number of cows and percentage of the herd for eachdifferent number of SCC measurements 4200 000 per animal(0–5) are depicted above each bar in Fig. 10. Cows with 4 or 5
SCC measurements 4200 000 were ,1 year older (Po 0.01,5.5 v. 4.5 years old) and tended (Po 0.10) to produce less milkcompared with cows that had lower SCC measurements (data
not shown). Results of the statistically significant (Po 0.02)polynomial regression analysis, depicted by the black line inFig. 10, show that the daughters of cows with 4 and 5 SCC
measurements 4200 000 also had markedly lower (Po 0.02)AMH concentrations, and correspondingly much smaller ovar-ies, a diminished number of high-quality eggs in ovaries andreduced ovarian function (Burns et al. 2005; Ireland et al. 2008,
2009; Jimenez-Krassel et al. 2009) as young adults comparedwith daughters of the cows with 0–3 SCC measurements4200 000.
These new, albeit preliminary data imply that a chronicmammary infection or injury during pregnancy (as predictedby a high number of SCCmeasurements4200 000), or perhaps
the antibiotic treatments used to alleviate the mammary glandinfection, may also have a negative impact not only on milkproduction, but also subsequent reproductive performance (fer-tility and reproductive lifespan) of the potentially chronically
infected cow’s daughters. If these preliminary findings areconfirmed using larger numbers of animals, then culling ofcows with chronically highly elevated SCC during pregnancy or
postpartum and (possibly) their daughters could improve notonly overall herd health and milk production, but also fertility.Results would also further illustrate the critical importance of a
Table 1. Mean number of follicles $3mm in diameter (antral follicle
count) during folliclewaves in heifer calves born tomothers fed a control
diet (Control) or who were nutritionally restricted (Restricted) for the
first 110 days of gestation
From Mossa et al. (2009)
Age (weeks) Antral follicle count
Control (n¼ 13) Restricted (n¼ 10) P
7 23.9� 2.1 14.1� 0.9 o0.01
18 26.1� 2.9 16.2� 1.1 o0.01
35 23.9� 2.2 16.6� 1.2 o0.01
0 1 2 3 4 5
Number of SCC �200 000 per cow
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
AM
H (
ng m
L�1 )
in d
augh
ters
n � 3
(29%)n � 56
(9%)n � 18
(2%)
(17%)n � 32
(6%)
n � 12
(37%)n � 71
(P � 0.02)
Fig. 10. Association of the number of somatic cell count (SCC) measure-
ments in milk of 4200 000 in dams with AMH concentrations in their
female offspring. A total of 5–7 measurements of SCC were made for each
dairy cow beginning 2months before and during her pregnancy. The number
of SCC measurements 4200 000 was used as an index of recurrent udder
infection, and the number of SCC measurements4200 000 per cow ranged
from 0 to 5. For analysis, cows were separated into groups based on the
number of their SCC measurements 4200 000. Bars indicate mean
(�s.e.m.) for AMH concentrations in daughters of dairy cows with 0–5
SCCmeasurements4200 000 permL.Above bar, n¼ number of heifers and
number within parentheses indicates percentage of herd. The bold line
depicts results of polynomial regression analysis (Po 0.02).
10 Reproduction, Fertility and Development J. J. Ireland et al.

well managed udder health care system on dairy farms foroptimal long-term herd performance and profitability.
Summary, practical application and conclusion
Our new results (Burns et al. 2005; Ireland et al. 2007, 2008,
2009; Jimenez-Krassel et al. 2009) provide strong evidence thatthe high variation in the quantity of follicles and oocytes inovaries has a negative impact on ovarian function that may,
in turn, result in suboptimal fertility in young adult cattle.This evidence included the novel findings that young adult cattlewith a low v. a high AFC also had: (1) much smaller ovaries;(2) markedly lower total number of morphologically healthy
follicles and oocytes in ovaries; (3) a reduced responsiveness tosuperovulation and diminished number of transferable embryos;(4) chronically heightened gonadotrophin secretion, but much
lower circulating AMH and progesterone concentrations duringoestrous cycles; (5) reduced endometrial thickness; and(6) higher amounts of cumulus cell markers for diminished
oocyte quality. Many of these phenotypic differences betweenyoung adult cattle with a low v. a high AFC have also beenreported for older, less fertile cattle (Erickson 1966a, 1966b;Erickson et al. 1976; Royal et al. 2000; Lucy 2001; Sartori et al.
2002a, 2002b; Washburn et al. 2002; Malhi et al. 2005) andwomen (Block 1951, 1952, 1953; te Velde et al. 1998; Faddy2000; te Velde and Pearson 2002; Ottolenghi et al. 2004)
compared with their younger counterparts. The results of ourstudies also demonstrated that a single measurement of AMHconcentration is a reliablemethod to determine relativeAFC and
quantity of follicles and oocytes in ovaries. Thus, our combinedresults provide a strong morphological, physiological andmechanistic rationale to test the hypothesis in future studies
that AMH concentration, and correspondingly AFC, ovary size,quantity of morphologically healthy follicles and oocytes inovaries, and the associated alterations in gonadotrophin secre-tion, ovarian function, and endometrial thickness, impact on
fertility in cattle.A positive outcome of future fertility trials would conclu-
sively establish that young adult cattle with a low v. a highAMH
concentration and a correspondingly diminished quantity ofmorphologically healthy oocytes require a greater AI numberto achieve pregnancy, thus providing new insights into the
potential factors that may cause or contribute to suboptimalfertility in heifers (e.g. low progesterone or poor oocyte quality,as previously observed for animals with a lowAFC and quantityof oocytes (Burns et al. 2005; Ireland et al. 2007, 2008, 2009;
Jimenez-Krassel et al. 2009)). A positive outcome will alsodemonstrate that the use of a simple AMH blood test is areliable method to not only identify young adult cattle with
suboptimal fertility, but also, for example, to monitor the impactof various environmental factors, such as nutrition and disease,on fertility.
Reliable methods to predict fertility and decrease infertilityin female cattle have heretofore not been developed. Indeed, it iswell recognised that inheritance of fertility, which is a trait
regulated by many genetic and environmental factors, is verylow (5%) in cattle (Philipsson 1981; Freeman 1984;Moore et al.1990). However, the variation for reproductive traits in cattle isvery large (Philipsson 1981). Consequently, discovery of a trait
contributing to fertility that is highly repeatable within indivi-duals and also potentially highly heritable and correlated with
fertility would be very useful not only as a new genetic tool toenhance selection of high-fertility females for AI with semenfrom high-genetic merit bulls, but also as a juvenile or adult
phenotypic marker to identify high-fertility females in the herd.We recognise that even if high-fertility heifers could be
identified reliably, culling of potentially inferior heifers is not
currently extensively practiced during reproductive manage-ment of dairy cattle, primarily because most heifers are neededas herd replacements to maintain relatively high culling rates of34% for existing cows in commercial herds (Weigel et al. 2003;
Caraviello et al. 2006). Nevertheless, sex-sorted semen (con-taining primarily spermatozoa with an X chromosome) fromhigh-geneticmerit bulls continues to becomemore available and
economical to use during AI to enhance the number of femalesborn in a herd (Hohenboken 1999;Weigel 2004; Cerchiaro et al.2007; De Vries et al. 2008). Consequently, the economic
benefits of using sexed semen from high-genetic merit bulls toenhance the number of female offspring for herd replacements,especially in the dairy industry (Hohenboken 1999; Cerchiaroet al. 2007; De Vries et al. 2008), could be further optimised
once newmethods are available to reliably identify the heifers ina herd with inferior or suboptimal fertility.
We also presented preliminary evidence that maternal nutri-
tion and disease could have profound negative impacts on theovarian reserve and potentially on the fertility (Mossa et al.
2009; Ireland et al. 2010) and reproductive longevity of off-
spring. This burgeoning area of research into the importance ofthe maternal environment on embryo development and adultonset of disease is predicted to yield significant new insights into
infertility, especially in dairy cows, and also lead to novelprenatal therapeutic methods to enhance fertility and perhapsthe health of offspring in cattle.
In conclusion, the inherently high variation in the ovarian
reserve, potentially caused by alterations in the maternal envir-onment, has a negative impact on ovarian function that mayresult in suboptimal fertility in young adult cattle, and a single
AMHmeasurement can be used reliably to determine if fertilityis suboptimal, as young adult cattle with low circulating AMHconcentrations have a correspondingly diminished ovarian
reserve.
Acknowledgements
Theworkwas supported byNational Research Initiative CompetitiveGrants
2001–35203–10910, 2004–35203–14781 and 2007–35203–18178 from the
USDA National Institute of Food and Agriculture to J.J.I. and by the Irish
Department of Agriculture, Fisheries and Food Research Stimulus Fund
program 2006 (RSF 06–328) to A.C.O.E.
References
Ahmed Ebbiary, N. A., Lenton, E. A., and Cooke, I. D. (1994).
Hypothalamic–pituitary ageing: progressive increase in FSH and LH
concentrations throughout the reproductive life in regularly
menstruating women. Clin. Endocrinol. 41(2), 199–206. doi:10.1111/
J.1365-2265.1994.TB02530.X
Amsterdam,A., Hanoch, T., Dantes, A., Tajima,K., Strauss, J. F., and Seger,
R. (2002). Mechanisms of gonadotrophin desensitization. Mol. Cell.
Endocrinol. 187(1–2), 69–74. doi:10.1016/S0303-7207(01)00701-8
Variation in antral follicle count in cattle Reproduction, Fertility and Development 11

Barker, D. J. (1992). The effect of nutrition of the fetus and neonate on
cardiovascular disease in adult life. Proc. Nutr. Soc. 51, 135–144.
doi:10.1079/PNS19920023
Bartlett, P. C., Miller, G. Y., Anderson, C. R., and Kirk, J. H. (1990). Milk
production and somatic cell count in Michigan dairy herds. J. Dairy Sci.
73(10), 2794–2800. doi:10.3168/JDS.S0022-0302(90)78966-7
Basir, G. S., O, W.-S., So, W. W. K., Ng, E. H. Y., and Ho, P. C. (2002).
Evaluation of cycle-to-cycle variation of endometrial responsiveness
using transvaginal sonography in women undergoing assisted reproduc-
tion. Ultrasound Obstet. Gynecol. 19(5), 484–489. doi:10.1046/J.1469-
0705.2002.00685.X
Beckers, N. G. M., Macklon, N. S., Eijkemans, M. J. C., and Fauser,
B. C. J. M. (2002). Women with regular menstrual cycles and a poor
response to ovarian hyperstimulation for in vitro fertilization exhibit
follicular phase characteristics suggestive of ovarian aging. Fertil.
Steril. 78, 291–297. doi:10.1016/S0015-0282(02)03227-2
Bettegowda, A., Patel, O. V., Ireland, J. J., and Smith, G. W. (2006).
Quantitative analysis of messenger RNA abundance for ribosomal
protein L-15, cyclophilin-A, phosphoglycerokinase, glucuronidase,
glyceraldehyde 3-phosphate dehydrogenase, b-actin and histone H2A
during bovine oocyte maturation and early embryogenesis in vitro.Mol.
Reprod. Dev. 73, 267–278. doi:10.1002/MRD.20333
Bettegowda, A., Patel, O. V., Lee, K.-B., Park, K.-E., Salem, M., Yao, J.,
Ireland, J. J., and Smith, G. W. (2008). Identification of novel bovine
cumulus cell molecular markers predictive of oocyte competence:
functional and diagnostic implications. Biol. Reprod. 79, 301–309.
doi:10.1095/BIOLREPROD.107.067223
Black, J. L., and Erickson, B. H. (1968).Oogenesis and ovarian development
in the prenatal pig. Anat. Rec. 161, 45–55. doi:10.1002/AR.1091610105
Block, E. (1951). Quantitativemorphological investigations of the follicular
system in women. Acta Endocrinol. 8, 33–54.
Block, E. (1952). Quantitative morphological investigation of the follicular
systen in women. Acta Anat. 14, 108–123. doi:10.1159/000140595
Block, E. (1953). A quantitative morphological investigation of the
follicular system in newborn female infants. Acta Anat. 17, 201–206.
doi:10.1159/000140805
Blondin, P., Coenen, K., Guilbault, L. A., and Sirard, M.-A. (1996).
Superovulation can reduce the developmental competence of bovine
embryos.Theriogenology 46, 1191–1203. doi:10.1016/S0093-691X(96)
00290-7
Borwick, S. C., Rhind, S. M., McMillen, S. R., and Racey, P. A. (1997).
Effect of undernutrition of ewes from the time of mating on fetal ovarian
development in mid-gestation. Reprod. Fertil. Dev. 9, 711–715.
doi:10.1071/R97011
Bourdon, R. M., and Brinks, J. S. (1986). Scrotal circumference in yearling
Hereford bulls: adjustment factors, heritabilities and genetic, environ-
mental and phenotypic relationships with growth traits. J. Anim. Sci. 62,
958–967.
Burns, D. S., Jimenez-Krassel, F. J., Ireland, J. L. H., Knight, P. G., and
Ireland, J. J. (2005). Numbers of antral follicles during follicular waves
in cattle: evidence for high variation among animals, very high repeat-
ability in individuals, and an inverse association with serum follicle-
stimulating hormone concentrations. Biol. Reprod. 73, 54–62.
doi:10.1095/BIOLREPROD.104.036277
Caraviello, D. Z., Weigel, K. A., Shook, G. E., and Ruegg, P. L. (2005).
Assessment of the impact of somatic cell count on functional longevity in
Holstein and Jersey cattle using survival analysis methodology. J. Dairy
Sci. 88(2), 804–811. doi:10.3168/JDS.S0022-0302(05)72745-4
Caraviello, D. Z., Weigel, K. A., Fricke, P. M., Wiltbank, M. C., Florent,
M. J., Cook, N. B., Nordlund, K. V., Zwald, N. R., and Rawson, C. L.
(2006). Survey of management practices on reproductive performance
of dairy cattle on large US commercial farms. J. Dairy Sci. 89(12),
4723–4735. doi:10.3168/JDS.S0022-0302(06)72522-X
Cerchiaro, I., Cassandro,M., Dal Zotto, R., Carnier, P., andGallo, L. (2007).
A field study on fertility and purity of sex-sorted cattle sperm. J. Dairy
Sci. 90(5), 2538–2542. doi:10.3168/JDS.2006-694
Chebel, R. C., Braga, F. A., and Dalton, J. C. (2007). Factors affecting
reproductive performance of Holstein heifers. Anim. Reprod. Sci.
101(3–4), 208–224. doi:10.1016/J.ANIREPROSCI.2006.09.010
Conti, M., Harwood, J. P., Hsueh, A. J., Dufau, M. L., and Catt, K. J. (1976).
Gonadotrophin-induced loss of hormone receptors and desensitization of
adenylate cyclase in the ovary. J. Biol. Chem. 251(23), 7729–7731.
Coulter, G. H., and Foote, R. H. (1979). Bovine testicular measurements as
indicators of reproductive performance and their relationship to produc-
tive traits in cattle: a review. Theriogenology 11, 297. doi:10.1016/0093-
691X(79)90072-4
Cushman, R. A., DeSouza, J. C., Hedgpeth, V. S., and Britt, J. H. (1999).
Superovulatory response of one ovary is related to the micro- and
macroscopic population of follicles in the contralateral ovary of the cow.
Biol. Reprod. 60(2), 349–354. doi:10.1095/BIOLREPROD60.2.349
Daily, C. A., Laurent, S. L., and Nunley, W. C., Jr (1994). The prognostic
value of serum progesterone and quantitative b-human chorionic gona-
dotrophin in early human pregnancy. Am. J. Obstet. Gynecol. 171,
380–384.
Da Silva, P., Aitken, R. P., Rhind, S. M., Racey, P. A., and Wallace, J. M.
(2002). Impact of maternal nutrition during pregnancy on pituitary
gonadotrophin gene expression and ovarian development in growth-
restricted and normally grown late gestation sheep fetuses.Reproduction
123, 769–777. doi:10.1530/REP.0.1230769
de Boo, H. A., and Harding, J. E. (2006). The developmental origins of adult
disease (Barker) hypothesis.Aust. N. Z. J. Obstet. Gynaecol. 46(1), 4–14.
doi:10.1111/J.1479-828X.2006.00506.X
De Vries, A., Overton, M., Fetrow, J., Leslie, K., Eicker, S., and Rogers, G.
(2008). Exploring the impact of sexed semen on the structure of the dairy
industry. J. Dairy Sci. 91(2), 847–856. doi:10.3168/JDS.2007-0536
Diskin, M. G., and Morris, D. G. (2008). Embryonic and early foetal losses
in cattle and other ruminants. Reprod. Domest. Anim. 43(Suppl. 2),
260–267. doi:10.1111/J.1439-0531.2008.01171.X
Elson, J., Salim, R., Tailor, A., Banerjee, S., Zosmer, N., and Jurkovic, D.
(2003). Prediction of early pregnancy viability in the absence of an
ultrasonically detectable embryo. Ultrasound Obstet. Gynecol. 21,
57–61. doi:10.1002/UOG.1
Eppig, J. J., O’Brien, M. J., Pendola, F. L., andWatanabe, S. (1998). Factors
affecting the developmental competence of mouse oocytes grown
in vitro: follicle-stimulating hormone and insulin. Biol. Reprod. 59(6),
1445–1453. doi:10.1095/BIOLREPROD59.6.1445
Erickson, B. H. (1966a). Development and radio-response of the prenatal
bovine ovary. J. Reprod. Fertil. 11, 97–105. doi:10.1530/JRF.0.0110097
Erickson, B. H. (1966b). Development and senescence of the postnatal
bovine ovary. J. Anim. Sci. 25, 800–805.
Erickson, B. H., Reynolds, R. A., and Murphree, R. L. (1976). Ovarian
characteristics and reproductive performance of the aged cows. Biol.
Reprod. 15, 555–560. doi:10.1095/BIOLREPROD15.4.555
Faddy, M. J. (2000). Follicle dynamics during ovarian ageing. Mol. Cell.
Endocrinol. 163, 43–48. doi:10.1016/S0303-7207(99)00238-5
Faddy, M. J., and Gosden, R. G. (1996). A model conforming the decline in
follicle numbers to the age of menopause in women. Hum. Reprod. 11,
1484–1486.
Fanchin, R., Taieb, J., Lozano, D. H. M., Ducot, B., Frydman, R., and
Bouyer, J. (2005). High reproducibility of serum anti-Mullerian hor-
mone measurements suggests a multi-staged follicular secretion and
strengthens its role in the assessment of ovarian follicular status. Hum.
Reprod. 20(4), 923–927. doi:10.1093/HUMREP/DEH688
Freeman,A. E. (1984). Secondary traits: sire evaluation and the reproductive
complex. J. Dairy Sci. 67, 449–458. doi:10.3168/JDS.S0022-0302(84)
81324-7
12 Reproduction, Fertility and Development J. J. Ireland et al.

Gardner, D. S., Ozanne, S. E., and Sinclair, K. D. (2009). Effect of the early-
life nutritional environment on fecundity and fertility of mammals.
Philos. Trans. R. Soc. Lond. B. Biol. Sci. 364(1534), 3419–3427.
doi:10.1098/RSTB.2009.0121
Gluckman, P. D., and Hanson,M. A. (2004). Living with the past: evolution,
development and patterns of disease. Science 305(5691), 1733–1736.
doi:10.1126/SCIENCE.1095292
Green, M. J., Green, L. E., Schukken, Y. H., Bradley, A. J., Peeler, E. J.,
Barkema, H. W., de Haas, Y., Collis, V. J., and Medley, G. F. (2004).
Somatic cell count distributions during lactation predict clinical mastitis.
J. Dairy Sci. 87(5), 1256–1264. doi:10.3168/JDS.S0022-0302(04)
73276-2
Haughian, J. M., Ginther, O. J., Kot, K., and Wiltbank, M. C. (2004).
Relationship between FSH patterns and follicular dynamics and the
temporal associations among hormones in natural and GnRH-induced
gonadotrophin surges in heifers. Reproduction 127, 23–33. doi:10.1530/
REP.1.00030
Hehenkamp,W. J. K., Looman, C.W.N., Themmen, A. P. N., de Jong, F. H.,
teVelde, E. R., andBroekmans, F. J.M. (2006).Anti-Mullerian hormone
levels in the spontaneous menstrual cycle do not show substantial
fluctuation. J. Clin. Endocrinol. Metab. 91(10), 4057–4063.
doi:10.1210/JC.2006-0331
Hohenboken, W. D. (1999). Applications of sexed semen in cattle produc-
tion. Theriogenology 52(8), 1421–1433. doi:10.1016/S0093-691X(99)
00227-7
Hunter, J. (1787). An experiment to determine the effect of extirpating one
ovarium upon the number of young produced. Philos. Trans. R. Soc.
Lond. A 77, 233–239. doi:10.1098/RSTL.1787.0021
Ibanez, L., Potau, N., Enriquez, G., Marcos, M. V., and de Zegher, F.
(2003). Hypergonadotrophinaemia with reduced uterine and ovarian
size in women born small-for-gestational-age. Hum. Reprod. 18(8),
1565–1569. doi:10.1093/HUMREP/DEG351
Inskeep, E. K. (2004). Preovulatory, postovulatory, and postmaternal
recognition effects of concentrations of progesterone on embryonic
survival in the cow. J. Anim. Sci. 82(13_Suppl.), E24–E39.
Ireland, J. J., Mihm, M., Austin, E., Diskin, M. G., and Roche, J. F. (2000).
Historical perspective of turnover of dominant follicles during the
bovine oestrous cycle: key concepts, studies, advancements and terms.
J. Dairy Sci. 83, 1648–1658. doi:10.3168/JDS.S0022-0302(00)75033-8
Ireland, J. J., Ward, F., Jimenez-Krassel, F., Ireland, J. L. H., Smith, G. W.,
Lonergan, P., and Evans, A. C. O. (2007). Follicle numbers are highly
repeatable within individual animals but are inversely correlated with
FSH concentrations and the proportion of good-quality embryos after
ovarian stimulation in cattle.Hum. Reprod. 22, 1687–1695. doi:10.1093/
HUMREP/DEM071
Ireland, J.L.H., Scheetz,D., Jimenez-Krassel, F.,Themmen,A.P.N.,Ward,F.,
Lonergan, P., Smith, G. W., Perez, G. I., Evans, A. C. O., and Ireland, J. J.
(2008). Antral follicle count reliably predicts number of morphologically
healthy oocytes and follicles in ovaries of young adult cattle.Biol. Reprod.
79, 1219–1225. doi:10.1095/BIOLREPROD.108.071670
Ireland, J. J., Zielak, A. E., Jimenez-Krassel, F., Folger, J., Bettegowda, A.,
et al. (2009). Variation in the ovarian reserve is linked to alterations in
intrafollicular oestradiol production and ovarian biomarkers of follicular
differentiation and oocyte quality in cattle. Biol. Reprod. 80, 954–964.
doi:10.1095/BIOLREPROD.108.073791
Ireland, J. J., Scheetz, D., Jimenez, F., Folger, J. K., Smith, G.W.,Mossa, F.,
and Evans, A. C. O. (2010). Evidence that mammary gland infection/
injury during pregnancy in dairy cows may have a negative impact on
size of the ovarian reserve in their daughters In ‘43rd Annual Meeting
for the Society for Study of Reproduction’. (Society for Study of
Reproduction/HighWire Press: Milwaukee, USA.)
Jimenez-Krassel, F., Folger, J., Ireland, J. L. H., Smith, G. W., Hou, X.,
Davis, J. S., Lonergan, P., Evans, A. C. O., and Ireland, J. J. (2009).
Evidence that high variation in ovarian reserves of healthy young adults
has a negative impact on the corpus luteum and endometrium during
reproductive cycles of single-ovulating species. Biol. Reprod. 80,
1272–1281. doi:10.1095/BIOLREPROD.108.075093
Kawamata, M. (1994). Relationship between the number of small follicles
prior to superovulatory treatment and superovulatory response in Hol-
stein cows. J. Vet. Med. Sci. 56, 965–967.
Keeton, L. L., Green, R. D., Golden, B. L., and Anderson, K. J. (1996).
Estimation of variance components and prediction of breeding values for
scrotal circumference and weaning weights in Limousin cattle. J. Anim.
Sci. 74, 31–36.
Kevenaar, M. E., Meerasahib, M. F., Kramer, P., van de Lang-Born,
B. M. N., de Jong, F. H., Groome, N. P., Themmen, A. P. N., and Visser,
J. A. (2006). Serum AMH levels reflect the size of the primordial
follicle pool in mice. Endocrinology 147, 3228–3234. doi:10.1210/
EN.2005-1588
Klein, N. A., Battaglia, D. E., Clifron, D. K., Bremner, W. J., and Soules,
M. R. (1996). The gonadotrophin secretion pattern in normal women of
advanced reproductive age in relation to themonotropic FSH rise. J. Soc.
Gynecol. Investig. 3(1), 27–32. doi:10.1016/1071-5576(95)00040-2
Kriese, L. A., Bertrand, J. K., and Benyshek, L. L. (1991). Age adjustment
factors, heritabilities and genetic correlations for scrotal circumference
and related growth traits in Hereford and Brangus bulls. J. Anim. Sci. 69,
478–489.
Kuhn, M. T., Hutchison, J. L., and Wiggans, G. R. (2006). Characterization
of Holstein heifer fertility in the United States. J. Dairy Sci. 89(12),
4907–4920. doi:10.3168/JDS.S0022-0302(06)72541-3
La Marca, A., and Volpe, A. (2006). Anti-Mullerian hormone (AMH)
in female reproduction: is measurement of circulating AMH a useful
tool? Clin. Endocrinol. 64(6), 603–610. doi:10.1111/J.1365-2265.2006.
02533.X
La Marca, A., Stabile, G., Artenisio, A. C., and Volpe, A. (2006). Serum
anti-Mullerian hormone throughout the human menstrual cycle. Hum.
Reprod. 21(12), 3103–3107. doi:10.1093/HUMREP/DEL291
Lehenbauer, T. W., and Oltjen, J. W. (1998). Dairy cow culling strategies:
making economical culling decisions. J. Dairy Sci. 81(1), 264–271.
doi:10.3168/JDS.S0022-0302(98)75575-4
Lonergan, P., Monaghan, P., Rizos, D., Boland, M., and Gordon, I. (1994).
Effect of follicle size on bovine oocyte quality and developmental
competence following maturation, fertilization and culture in vitro.
Mol. Reprod. Dev. 37, 48–53. doi:10.1002/MRD.1080370107
Lucy,M. C. (2001). Reproductive loss in high-producing dairy cattle: where
will it end? J. Dairy Sci. 84, 1277–1293. doi:10.3168/JDS.S0022-0302
(01)70158-0
Malhi, P. S., Adams, G. P., and Singh, J. (2005). Bovine model for the study
of reproductive aging in women: follicular, luteal and endocrine char-
acteristics. Biol. Reprod. 73(1), 45–53. doi:10.1095/BIOLREPROD.104.
038745
Mann, G. E., and Lamming, G. E. (1999). The influence of progesterone
during early pregnancy in cattle. Reprod. Domest. Anim. 34(3–4),
269–274. doi:10.1111/J.1439-0531.1999.TB01250.X
Mcmillen, I. C., and Robinson, J. S. (2005). Developmental origins of the
metabolic syndrome: prediction, plasticity, and programming. Physiol.
Rev. 85(2), 571–633. doi:10.1152/PHYSREV.00053.2003
McNatty, K. P., Smith, P., Hudson, N. L., Heath, D. A., Tisdall, D. J., O,W.-S.,
and Braw-Tal, R. (1995). Development of the sheep ovary during fetal and
early neonatal life and the effect of fecundity genes. J. Reprod. Fertil. 49,
123–135.
McNeill, R. E., Diskin, M. G., Sreenan, J. M., and Morris, D. G. (2006).
Associations betweenmilk progesterone concentration on different days
and with embryo survival during the early luteal phase in dairy cows.
Theriogenology 65(7), 1435–1441. doi:10.1016/J.THERIOGENOL
OGY.2005.08.015
Variation in antral follicle count in cattle Reproduction, Fertility and Development 13

McTavish, K. J., Jimenez, M., Walters, K. A., Spaliviero, J., Groome, N. P.,
Themmen, A. P., Visser, J. A., Handelsman, D. J., and Allan, C. M.
(2007). Rising follicle-stimulating hormone levels with age accelerate
female reproductive failure. Endocrinology 148(9), 4432–4439.
doi:10.1210/EN.2007-0046
Moore, R. K., Kennedy, B. W., Schaeffer, L. R., and Moxley, J. E. (1990).
Relationships between reproduction traits, age and body weight at
calving, and days dry in first lactation Ayrshires and Holsteins. J. Dairy
Sci. 73, 835–842. doi:10.3168/JDS.S0022-0302(90)78737-1
Mossa, F., Kenny, D., Jimenez-Krassel, F., Smith, G. W., Berry, D., Butler,
S., Fair, T., Lonergan, P., Ireland, J. J., and Evans, A. C. O. (2009).
Undernutrition of heifers during the first trimester of pregnancy
diminishes size of the ovarian reserve in female offspring. In ‘42nd
Annual Meeting of the Society for Study of Reproduction. Vol. Abstract
135, 81’. p. 135. (Society for Study of Reproduction/HighWire Press:
Pittsburg, USA.)
Mossa, F., Jimenez-Krassel, F., Walsh, S., Berry, D. P., Butler, S., et al.
(2010). The inherent capacity of the pituitary gland to produce gonado-
trophins is not influenced by the number of ovarian follicles .3mm in
diameter in cattle. Reprod. Fertil. Dev. 22, 550–557. doi:10.1071/
RD09100
Ottolenghi, C., Uda, M., Hamatani, T., Crisponi, L., Garcia, J.-E., Ko, M.,
Pilia, G., Sforza, C., Schlessinger, D., and Forabosco, A. (2004). Aging
of oocyte, ovary and human reproduction. Ann. N. Y. Acad. Sci. 1034(1),
117–131. doi:10.1196/ANNALS.1335.015
Pezzullo, J. C. (2009). Web site for Statistical Power Analysis: http://
statpages.org/proppowr.html Accessed 4 December 2008.
Philipsson, J. (1981). Genetic aspects of female fertility in dairy cattle.
Livest. Prod. Sci. 8(4), 307–319. doi:10.1016/0301-6226(81)90049-X
Piltonen, T., Morin-Papunen, L., Koivunen, R., Perheentupa, A., Ruokonen,
A., and Tapanainen, J. S. (2005). Serum anti-Mullerian hormone levels
remain high until late reproductive age and decrease during metformin
therapy in womenwith polycystic ovary syndrome.Hum. Reprod. 20(7),
1820–1826. doi:10.1093/HUMREP/DEH850
Pritts, E. A., and Atwood, A. K. (2002). Luteal phase support in infertility
treatment: a meta-analysis of the randomized trials.Hum. Reprod. 17(9),
2287–2299. doi:10.1093/HUMREP/17.9.2287
Raga, F., Bonilla-Musoles, F., Casan, E. M., Klein, O., and Bonilla, F.
(1999). Assessment of endometrial volume by three-dimensional ultra-
sound prior to embryo transfer: clues to endometrial receptivity. Hum.
Reprod. 14(11), 2851–2854. doi:10.1093/HUMREP/14.11.2851
Reame, N. E., Kelch, R. P., Beitins, I. Z., Yu, M.-Y., Zawacki, C. M., and
Padmanabhan, V. (1996). Age effects on follicle-stimulating hormone
and pulsatile luteinizing hormone secretion across the menstrual cycle
of premenopausal women. J. Clin. Endocrinol. Metab. 81, 1512–1518.
doi:10.1210/JC.81.4.1512
Rico, C., Fabre, S., Medigue, C., Clemente, N. d., Clement, F., et al.
(2009). Anti-Mullerian hormone is an endocrine marker of ovarian
gonadotrophin-responsive follicles and can help to predict superovul-
atory responses in the cow. Biol. Reprod. 80(1), 50–59. doi:10.1095/
BIOLREPROD.108.072157
Royal, M., Mann, G. E., and Flint, A. P. F. (2000). Strategies for reversing
the trend towards subfertility in dairy cattle. Vet. J. 160, 53–60.
doi:10.1053/TVJL.1999.0450
Sartori, R., Rosa, G. J. M., and Wiltbank, M. C. (2002a). Ovarian structures
and circulating steroids in heifers and lactating cows in summer and
lactating and dry cows in winter. J. Dairy Sci. 85(11), 2813–2822.
doi:10.3168/JDS.S0022-0302(02)74368-3
Sartori, R., Sartor-Bergfelt, R., Mertens, S. A., Guenther, J. N., Parrish,
J. J., and Wiltbank, M. C. (2002b). Fertilization and early embryonic
development in heifers and lactating cows in summer and lactating and
dry cows in winter. J. Dairy Sci. 85(11), 2803–2812. doi:10.3168/JDS.
S0022-0302(02)74367-1
Scheffer, G. J., Broekmans, F. J. M., Looman, C. W. N., Blankenstein, M.,
Fauser, B. C. J. M., De Jong, F. H., and te Velde, E. R. (2003). The
number of antral follicles in normal women with proven fertility
is the best reflection of reproductive age. Hum. Reprod. 18, 700–706.
doi:10.1093/HUMREP/DEG135
Singh, J., Dominguez, M., Jaiswal, R., and Adams, G. P. (2004). A simple
ultrasound test to predict the superstimulatory response in cattle.
Theriogenology 62, 227–243. doi:10.1016/J.THERIOGENOLOGY.
2003.09.020
Smith, B. A., Brinks, J. S., and Richardson, G. V. (1989). Estimation of
genetic parameters among breeding soundness examination components
and growth traits in yearling bulls. J. Anim. Sci. 67, 2892–2896.
Steckler, T.,Wang, J., Bartol, F. F., Roy, S. K., and Padmanabhan,V. (2005).
Fetal programming: prenatal testosterone treatment causes intrauterine
growth retardation, reduces ovarian reserve and increases ovarian
follicular recruitment. Endocrinology 146(7), 3185–3193. doi:10.1210/
EN.2004-1444
Stringfellow, D. A., and Seidel, S. M. (Eds) (1998). ‘Manual of the
International Embryo Transfer Society (IETS).’ 3rd edn. (International
Embryo Transfer Society: Savoy, USA.)
Stronge, A. J. H., Sreenan, J. M., Diskin, M. G., Mee, J. F., Kenny, D. A.,
and Morris, D. G. (2005). Post-insemination milk progesterone con-
centration and embryo survival in dairy cows. Theriogenology 64(5),
1212–1224. doi:10.1016/J.THERIOGENOLOGY.2005.02.007
Taneja, M., Bols, P. E., Van de Velde, A., Ju, J. C., Schreiber, D., Tripp,
M. W., Levine, H., Echelard, Y., Riesen, J., and Yang, X. (2000).
Developmental competence of juvenile calf oocytes in vitro and in vivo:
influence of donor animal variation and repeated gonadotrophin
stimulation. Biol. Reprod. 62(1), 206–213. doi:10.1095/BIOLREPROD
62.1.206
te Velde, E. R., and Pearson, P. L. (2002). The variability of female
reproductive ageing.Hum. Reprod. Update 8(2), 141–154. doi:10.1093/
HUMUPD/8.2.141
te Velde, E. R., Dorland,M., andBroekmans, F. J. (1998). Age at menopause
as amarker of reproductive ageing.Maturitas 30, 119–125. doi:10.1016/
S0378-5122(98)00067-X
van Rooij, I. A., Broekmans, F. J., te Velde, E. R., Fauser, B. C., Bancsi,
L. F., de Jong, F. H., and Themmen, A. P. (2002). Serum anti-Mullerian
hormone levels: a novel measure of ovarian reserve. Hum. Reprod. 17,
3065–3071. doi:10.1093/HUMREP/17.12.3065
van Rooij, I. A. J., Broekmans, F. J. M., Scheffer, G. J., Looman, C. W. N.,
Habbema, J. D. F., de Jong, F.H., Fauser, B. J. C.M., Themmen,A. P. N.,
and te Velde, E. R. (2005). Serum anti-Mullerian hormone levels best
reflect the reproductive decline with age in normal women with proven
fertility: a longitudinal study.Fertil. Steril. 83(4), 979–987. doi:10.1016/
J.FERTNSTERT.2004.11.029
Washburn, S. P., Silvia, W. J., Brown, C. H., McDaniel, B. T., and
McAllister, A. J. (2002). Trends in reproductive performance in south-
eastern Holstein and Jersey DHI herds. J. Dairy Sci. 85, 244–251.
doi:10.3168/JDS.S0022-0302(02)74073-3
Weigel, K. A. (2004). Exploring the role of sexed semen in dairy production
systems. J. Dairy Sci. 87(13_Suppl.), E120–E130. doi:10.3168/JDS.
S0022-0302(04)70067-3
Weigel, K. A., Palmer, R. W., and Caraviello, D. Z. (2003). Investigation
of factors affecting voluntary and involuntary culling in expanding
dairy herds in Wisconsin using survival analysis. J. Dairy Sci. 86(4),
1482–1486. doi:10.3168/JDS.S0022-0302(03)73733-3
http://www.publish.csiro.au/journals/rfd
14 Reproduction, Fertility and Development J. J. Ireland et al.
Copyright © 2022 FDOKUMEN




















