
Diversity of epiphytic lichens in boreo-nemoral forests on the North-Estonian limestone escarpment:...
16
Diversity of epiphytic lichens in boreo-nemoral forests on the North-Estonian limestone escarpment: the effect of tree level factors and local environmental conditions Inga JÜRIADO, Jaan LIIRA and Jaanus PAAL Abstract: The species richness and composition of lichens was quantified for four temperate broad-leaved tree species (Acer platanoides, Fraxinus excelsior, Tilia cordata and Ulmus glabra) in boreo-nemoral forests on the talus slope of the North-Estonian limestone escarpment (North- Estonian Klint). Thirteen study sites were distributed along the klint on a west to east gradient. The effects of tree and stand characteristics and geographical location of a stand on composition and diversity of epiphytic lichens were evaluated by multivariate analyses (DCA, CCA, pCCA) and by general linear mixed models (GLMM). Tree level variables (e.g. bark pH, bryophytes cover and host tree species) explained the largest fraction of the variation in lichen species composition. However, species richness and composition were significantly influenced also by the unique habitat conditions of klint forest (length of the forest fragment, proximity of the stand to the sea and height of the escarpment). A significant correlation between stand locality and lichen diversity on the tree bole was found which is most likely related to local air pollution gradients caused by alkaline cement dust and acid pollutants in the north-eastern part of Estonia. Key words: Acer platanoides, air pollution, bark pH, bryophytes, cement dust, forest fragment size, Fraxinus excelsior, temperate broad-leaved trees, trunk circumference, Tilia cordata, Ulmus glabra Introduction In northern Europe, deciduous forests with temperate broad-leaved trees, such as small- leaved lime (Tilia cordata Mill.), Norway maple (Acer platanoides L.), common ash (Fraxinus excelsior L.) and wych elm (Ulmus glabra Huds.) are distributed mainly in the hemiboreal subzone of the boreal forest zone and are restricted to areas with soils of high fertility (Valk & Eilart 1974; Diekmann 1996). Anthropogenic pressure on broad- leaved forests on fertile soils has been long- term; stands have been cut for valuable timber and forest land has been used for agriculture (Laasimer 1965; Hansson 1997). Now, as most original forests have been replaced by young or middle-aged secondary forests, old-growth deciduous forests are par- ticularly scarce in northern Europe (Nilsson 1992; Esseen et al. 1997; Lõhmus et al. 2004). Fragmentation and degradation of old- growth deciduous forests have exerted negative effects on persistence of many organisms, including lichens growing on temperate broad-leaved trees (Thor 1998; Berg et al. 2002). Although epiphytic lichens depend primarily upon the host tree species and its properties (Barkman 1958; Brodo 1973), stand age and spatial-temporal con- tinuity are considered key factors for the development of highly complex lichen assemblages in forests (Boudreault et al. 2000; Price & Hochachka 2001; Snäll et al. 2003; Ellis & Coppins 2007a). This can be explained by the fact that many epiphytic forest-dwelling lichens have a long gener- ation time as well as poor dispersal and colonization ability (Sillett et al. 2000; Scheidegger & Goward 2002). Lichen assemblages on broadleaved trees, which are dominant in forests of tem- perate regions (e.g. oak, ash, lime), have been I. Ju ¨ riado, J. Liira and J. Paal: Institute of Ecology and Earth Sciences, University of Tartu 40 Lai st., Tartu 51005, Estonia. Email: [email protected] The Lichenologist 41(1): 81–96 (2009) © 2009 British Lichen Society doi:10.1017/S0024282909007889 Printed in the United Kingdom
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Diversity of epiphytic lichens in boreo-nemoral forests on the North-Estonian limestone escarpment:...

Diversity of epiphytic lichens in boreo-nemoral forests on theNorth-Estonian limestone escarpment: the effect of tree level
factors and local environmental conditions
Inga JÜRIADO, Jaan LIIRA and Jaanus PAAL
Abstract: The species richness and composition of lichens was quantified for four temperatebroad-leaved tree species (Acer platanoides, Fraxinus excelsior, Tilia cordata and Ulmus glabra) inboreo-nemoral forests on the talus slope of the North-Estonian limestone escarpment (North-Estonian Klint). Thirteen study sites were distributed along the klint on a west to east gradient. Theeffects of tree and stand characteristics and geographical location of a stand on composition anddiversity of epiphytic lichens were evaluated by multivariate analyses (DCA, CCA, pCCA) and bygeneral linear mixed models (GLMM). Tree level variables (e.g. bark pH, bryophytes cover and hosttree species) explained the largest fraction of the variation in lichen species composition. However,species richness and composition were significantly influenced also by the unique habitat conditions ofklint forest (length of the forest fragment, proximity of the stand to the sea and height of theescarpment). A significant correlation between stand locality and lichen diversity on the tree bole wasfound which is most likely related to local air pollution gradients caused by alkaline cement dust andacid pollutants in the north-eastern part of Estonia.
Key words: Acer platanoides, air pollution, bark pH, bryophytes, cement dust, forest fragment size,Fraxinus excelsior, temperate broad-leaved trees, trunk circumference, Tilia cordata, Ulmus glabra
Introduction
In northern Europe, deciduous forests withtemperate broad-leaved trees, such as small-leaved lime (Tilia cordata Mill.), Norwaymaple (Acer platanoides L.), common ash(Fraxinus excelsior L.) and wych elm (Ulmusglabra Huds.) are distributed mainly in thehemiboreal subzone of the boreal forest zoneand are restricted to areas with soils of highfertility (Valk & Eilart 1974; Diekmann1996). Anthropogenic pressure on broad-leaved forests on fertile soils has been long-term; stands have been cut for valuabletimber and forest land has been used foragriculture (Laasimer 1965; Hansson 1997).Now, as most original forests have beenreplaced by young or middle-aged secondaryforests, old-growth deciduous forests are par-ticularly scarce in northern Europe (Nilsson
1992; Esseen et al. 1997; Lõhmus et al.2004).
Fragmentation and degradation of old-growth deciduous forests have exertednegative effects on persistence of manyorganisms, including lichens growing ontemperate broad-leaved trees (Thor 1998;Berg et al. 2002). Although epiphytic lichensdepend primarily upon the host tree speciesand its properties (Barkman 1958; Brodo1973), stand age and spatial-temporal con-tinuity are considered key factors for thedevelopment of highly complex lichenassemblages in forests (Boudreault et al.2000; Price & Hochachka 2001; Snäll et al.2003; Ellis & Coppins 2007a). This can beexplained by the fact that many epiphyticforest-dwelling lichens have a long gener-ation time as well as poor dispersal andcolonization ability (Sillett et al. 2000;Scheidegger & Goward 2002).
Lichen assemblages on broadleavedtrees, which are dominant in forests of tem-perate regions (e.g. oak, ash, lime), have been
I. Juriado, J. Liira and J. Paal: Institute of Ecology andEarth Sciences, University of Tartu 40 Lai st., Tartu51005, Estonia. Email: [email protected]
The Lichenologist 41(1): 81–96 (2009) © 2009 British Lichen Societydoi:10.1017/S0024282909007889 Printed in the United Kingdom

intensively studied in central Europe (e.g.Barkman 1958; Cieslinski et al. 1996) butmore rarely at the boundary of their dispersalrange in northern Europe (Löbel et al. 2006;Jüriado et al. 2008; Mežaka et al. 2008). Thecomposition of lichen communities coulddepend upon the location of the host tree inrelation to its natural distribution range, asthe number of epiphyte species tends todecrease on trees growing near the limits oftheir natural distribution area (Barkman1958; Jüriado et al. 2003). Moreover, tocover the full spectrum of habitat conditionssuitable for different threatened species, astudy of sites with similar substrata, but withdifferent soil moisture and nutrient con-ditions, is needed (Berg et al. 2002). In thecontext of long-term anthropogenic impacton the boreal landscape it is also important toidentify remaining habitats with potentiallyhigh biodiversity values (Gustafsson et al.1999).
The so-called klint forest of Estonia is anunusual habitat for temperate broad-leavedtree species in northern Europe. It occursas a narrow stripe on the talus slopes and atthe base of the limestone escarpment alongthe northern coastline of Estonia (North-Estonian Klint) (Paal 1998). In places, dueto inaccessibility, the forests are thought tobe among the very few primary forest standssurviving in Estonia (Kalda 1962). The his-torical continuity of the forests, mild climaticconditions in proximity of the sea, and shadeprovided by the cliffs, render the forests onthe talus slope of the North-Estonian Klintunique among habitats in the boreal forestzone (Paal 1998). According to the criteriaof the EU Habitat Directive (EC 1992),these forests belong to the north-easternmostvariety of type 9180 ‘Tilio-Acerion forests ofslopes, screes and ravines’, and have in thecontext of biodiversity protection the rank ofhigh priority for all Europe. These forests arecharacterized by high richness of vascularplant species and bryophytes, among themmany red-listed species (Paal 2001; Ingerpuuet al. 2003). In this study, we quantify therichness and composition of epiphytic lichenspecies on four temperate broad-leaved treespecies (Acer platanoides, Fraxinus excelsior,
Tilia cordata and Ulmus glabra) in theEstonian klint forests, and examine multipleenvironmental drivers influencing epiphyticlichen communities.
Materials and Methods
Study area
The North-Estonian Klint forms part of the largerBaltic Klint, creating a conspicuous geographicalboundary in northen Europe (Raukas & Teedumäe1997). The line of the klint follows the northern andnorth-western boundaries of the Ordovician limestoneoutcrop (Raukas & Teedumäe 1997); the upper part ofthe klint exposes calcareous rocks and the lower layersconsist of clays, silts, sandstones and argillites (Miidel1997). In Estonia, the klint is distributed across morethan 200 km (Miidel 1997). The height of the escarp-ment increases from west to east, varying between 24and 67 m a.s.1.; its slope varies from almost vertical(90°) to gently sloping (<20°), with one to five terraces.In many places the klint falls almost directly to the sea,but more often its base and the sea are separated by acoastal plain of variable width (Suuroja 2005). Thedominant tree species in the forests on the talus slopeof the escarpment are Fraxinus excelsior, Tilia cordata,Ulmus glabra and Acer platanoides and the characteristicspecies of the field layer are Lunaria rediviva L. andMercurialis perennis L. (Paal 1997; Paal et al. 2001).
A large expanse of klint forests is located in north-eastern Estonia, the area most severely affected by thesimultaneous presence of alkaline cement dust and oilshale fly ash, as well as by acid gaseous pollutants(mainly SO2 but also volatile organic compounds,ammonia and hydrogen sulphide) (Mandre 1995; Liblik2007). The pollutants originate from several electricitypower plants fired by oil shale and from chemical indus-try plants in north-eastern Estonia (Fig. 1) and also fromRussia (Haapala et al. 1996). The simultaneous pres-ence of alkaline emissions and acid gaseous pollutantshas the potential to neutralize the effects of acidification(Haapala et al. 1996; Liblik & Pensa 2001). The concen-trations of pollution in north-east Estonia were ex-tremely high up until 1990, since then emissions havebeen progressively decreasing (Liblik 2007).
Data collection
Field data were collected in 2002 as part of a projectdescribing the typology and soils of the Estonian klintforests (Paal et al. 2001; Paal 2001, 2007). On the basisof the composition of tree species recorded in stands, 13localities along the talus slope of the escarpment fromwest to east were selected for this study. For data analy-sis, the stands studied were grouped into four studyregions according to their geographic location in Estonia(Fig. 1; Table 1). As the studied forests occur in a line onthe talus slopes and at the base of the limestone escarp-ment along the northern coastline, the size of each
82 THE LICHENOLOGIST Vol. 41

locality was determined by the length of an unbrokenforest fragment (km) measured using a digital ruler onthe Internet-serviced map of Estonia (Web map server ofEstonian Land Board 2007). The same map was alsoused to measure the distance of the stand from thesea. The approximate height of the escarpment in eachlocality was provided by Suuroja (2005) (Table 1).
In western Estonia (study regions 1 and 2), twosample trees of four temperate broad-leaved species,Acer platanoides, Fraxinus excelsior, Tilia cordata andUlmus glabra, were studied in each locality. In easternEstonia, Tilia cordata was generally absent, and in onelocality only lichens on Acer platanoides were studied.Altogether 86 tree trunks were analysed (Table 1).
F. 1. Location of the study sites (�) in four study regions (Reg.) along the North-Estonian Klint. = Pollutionsources (cement plant in study region 3 and electricity power plants in the vicinity of study region 4).
T 1. Explanatory variables employed in the data analyses. The average values and minimum-maximum range of thevariables are presented in the brackets
Variables Comments
Tree level variablesBark pH Bark pH of sample quadrat on the bole (6·0; 4·2–8·0)Bryophyte cover Total bryophyte cover in a 20 × 20 sample quadrat on the bole (19·3%; 0-95%)Tree aspect The quadrat was set on two aspects of the trunk: (1) the aspect facing the escarpment
and (2) the aspect facing away from the escarpmentCircumference Circumference of a sample tree at a height of 1·3 m above ground (128·7 cm; 77–279
cm)Tree species Tree species studied: Acer platanoides (n=24), Fraxinus excelsior (n=24), Tilia cordata
(n=14), and Ulmus glabra (n=24)Stand level variables
Canopy cover Canopy coverage on the scale from zero to one near each sample tree (0·7; 0·5–0·9)Forest length* Length of an unbroken forest fragment (0·75–25 km)Height of klint** Height of the escarpment at each study site (15–55 m)Distance to sea* Distance of study stand from the coastline (0·07–7·7 km)
Geographic variablesStudy region The studied stands were grouped into four study regions according to their geographical
location along the northern coastline
*The value of the parameter was obtained from the digital map of Estonia (Web map server of Estonian Land Board2007).**The variable was provided by Suuroja (2005).
2009 Epiphytic lichens on the North-Estonian limestone escarpment—Jüriado et al. 83

Owing to the poor accessibility of the trees on the steepslope of the escarpment, sample trees (>20 cm diam.)were chosen subjectively within the study localities es-tablished by Paal et al. (2001). The circumference ofeach sample tree was measured at breast height (1·3 mabove ground level) and light conditions (% canopycover) were estimated visually near each sample tree onthe scale from zero to one (Table 1).
The epiphytic lichen communities on the tree trunkswere sampled using a 20 × 20 cm quadrat, at a height of1·3 m above ground level. As the trees on the talus slopeof the escarpment were in many cases leaning, the quad-rat was placed on two aspects of the trunk (one facing,the second facing away from the escarpment). To esti-mate the % cover of lichen species and the total cover ofbryophytes, the sample quadrat was divided into 100subquadrats. For measurement of bark pH, samplesof the bark were removed with a knife from each 20 ×20 cm sample quadrat on both sides of the tree trunk.Bark samples were air dried and stored in paper bagsuntil laboratory analysis. The bark (1 g) was incubatedin distilled water (1·5 ml g−1) for 24 hours and the pH ofthe solution was then measured with a standard pHmeter (E6121) (Table 1).
The specimens which were not identified in the fieldwere studied in the laboratory, using a stereomicro-scope, a light microscope, ‘spot tests’, UV light andstandardized thin-layer chromatography (TLC). Owingto their difficult taxonomy, species of Arthopyreniawere treated at the generic level. Reference materialsare deposited in the lichen herbarium at the NaturalHistory Museum of the University of Tartu (TU). Thenomenclature of the lichens follows Randlane et al.(2007). Data on species frequency in Estonia are derivedfrom Randlane and Saag (1999) and updated accordingto the Database of Estonian Lichens (2008). The list ofprotected lichen species is presented according to legaldocuments (Keskkonnaministri määrus nr 51, 2004;Vabariigi Valitsuse määrus nr 195, 2004) and red-listedlichen species are presented following Randlane et al.(2008).
Data analysis
Before the data analysis, some explanatory variableswere transformed to improve the assumptions of homo-scedacity and to render units of the variables compar-able. The cover values of bryophytes were square-roottransformed and the variables ‘circumference’, ‘forestlength’ and ‘distance to sea’ were log-transformed. Thecover values of lichen species were also square-roottransformed, to lessen the disproportionate effects of thedominant species (McCune & Grace 2002).
The main gradient structure of the species data wasexamined using Detrended Correspondence Analysis(DCA) (Hill & Gauch 1980) implemented in theprogram package PC-Ord version 4.25 (McCune &Mefford 1999). The axes were rescaled using 26 seg-ments. To reduce noise, the species appearing only in 1or 2 sample quadrats were removed from the data setprior to ordination. The proportion of variance repre-sented by ordination axes was estimated by the after-the-
fact method using relative Euclidean distance (McCune& Mefford 1999). Spearman rank order correlations(implemented in the program package Statistica 7.1;Statsoft Inc. 2005) were estimated among the environ-mental variables and the ordination axes.
Canonical Correspondence Analysis (CCA) (terBraak 1986) implemented in the program packageCANOCO ver. 4.5 (ter Braak & Šmilauer 2002) wasused to evaluate the descriptive power of environmentalvariables on species composition. The variables weregrouped into three subsets: tree level, stand level andgeographic variables (Table 1). To evaluate the explana-tory power of each set of the environmental variables,singly and in combination, the variation partitioningapproach (Borcard et al. 1992) in partial correspondenceanalysis (pCCA) (ter Braak 1986), was used. The for-ward selection procedure with randomization tests(Monte-Carlo permutation test, 1000 unrestricted per-mutations) was employed to select the most importantenvironmental variables influencing species compos-ition, retaining the variables with an independent signifi-cant contribution at the P<0·05 level. The statisticallysignificant environmental variables had the value ofinflation factors less than 10·0, i.e. below the suggestedlimit value of 20 (ter Braak & Šmilauer 2002).
The response of species richness to the studied en-vironmental variables was tested using a general linearmixed model (GLMM) (Littell et al. 1996) with thestepwise selection procedure implemented in the pro-gram package SAS ver. 8.2 (proc MIXED) (SASInstitute Inc. 1989). The response variable ‘number oflichen species’ and the predictor variable ‘circumfer-ence’ were log-transformed; and the variable ‘bryophytecover’ was square-root transformed to obtain homo-scedacity of model residual variation and normality ofresiduals. The locality of each stand studied was consid-ered the fixed factor and the factor ‘tree aspect’ wastreated as the result of repeated observations per sampletree. We tested interactions of the factor ‘tree species’with the continuous factors ‘bryophyte cover’, ‘circum-ference’ and ‘bark pH’. Backward elimination was usedto select the final model for species richness. At each stepthe variable with the highest P value was removed, andall variables at P<0·05 were retained in the final model.Akaike’s information criterion (AIC) (Akaike 1973) andthe significance of the factors were used to identifyoptimal parameterization of the models (Shao 1997).For multiple comparisons between tree species and dif-ferent aspects of the tree trunk, the Tukey-Krameradjustment was used. The GLMM analysis was alsoapplied to evaluate the influence of ‘bryophyte cover’,‘circumference’ and ‘study region’ on bark pH using thesame model settings as in the models described above.
Results
Bark pH of temperate broad-leavedtrees
The mean values of bark pH were signifi-cantly different among the study regions
84 THE LICHENOLOGIST Vol. 41

(Table 2). The bark pH of trees in region 3differed from that of trees in all other studyareas in having subneutral mean values (6·69±0·12). The mean bark pH of region 2 wasthe lowest (5·59±0·10) being marginally sig-nificantly different from the correspondingvalue for region 1 (5·94±0·07, Table 2).
Bark pH varied significantly between thetemperate broad-leaved tree species studied.The bark of Acer platanoides was less acid(mean pH = 6·48 ±0·08) than the bark of theother broad-leaved tree species (mean pHvalues for Fraxinus excelsior, Tilia cordata andUlmus glabra being 5·91±0·08, 5·63±0·11and 6·11±0·09, respectively) (Table 2). Thevalue of the bark pH of Tilia cordata differedsignificantly from that of Ulmus glabra, thebark of T. cordata being more acid. In ad-dition, there was an overall positive relation-ship observed between cover of bryophytesand bark pH (Table 2).
Composition of lichen community
A total of 74 lichen species was found onthe trunks of the four broad-leaved treespecies. The highest number was recordedon Fraxinus excelsior with 46 species, followedby Tilia cordata with 42 species, Ulmus glabrawith 39 species, and Acer platanoides with 38species (Appendix 1). Most of the recorded
lichens (92%) were crustose, while only sixfoliose or fruticose species (Cladonia fim-briata, Leptogium lichenoides, Melanelia fuligi-nosa, M. subaurifera, Phaeophyscia orbicularisand Ramalina farinacea) were recorded. Themost frequent lichen species on the temper-ate broad-leaved trees were Graphis scripta,Opegrapha rufescens and Lecidella elaeo-chroma, occurring in more than 40% of thesample quadrats. Out of the total of 74species, 20% of the taxa (15 species) areeither rare in Estonia (having less than tenlocalities), or are red-listed or protected bylaw (e.g. Arthonia byssacea, A. didyma, Bia-toridium monasteriense, Lecidea erythrophaea,Opegrapha ochrocheila and Pyrenula nitidella)(Appendix 1).
In the DCA ordination, the first two axesdescribed 36% of variance in the compos-ition of the lichen communities, (25% foraxis 1 and 11% for axis 2). Since axis 3 didnot show a significant contribution to theresults of ordination (3·1%), the results arepresented only for the first two axes. Thesample quadrats from regions 1 and 2 arescattered all over the DCA ordination plot;the other two study regions are moreclustered. The quadrats from region 3 aremostly located on the right side of the ordi-nation plot and the quadrats from region 4form a compact cluster in the centre of the
T 2. The results of general linear mixed model analysis (GLMM) for bark pH on temperate broad-leaved trees
Effect df P Mean (± SE) Slope
Study region 3; 79 <0·0001Region 1 5·94±0·07b
Region 2 5·59±0.10c
Region 3 6·69±0·12a
Region 4 5·91±0·08b,c
Tree species 3; 79 <0·0001Acer platanoides 6·48±0·08a
Fraxinus excelsior 5·91±0·08b,c
Tilia cordata 5·63±0·11c
Ulmus glabra 6·11±0·09b
Bryophyte cover 1; 77 <0·0001 0·050***
Explanations of the explanatory variables are given in Table 1. Within-group mean values (mean) with standarderrors (± SE) are presented for categorical variables and slope estimates (slope) are presented for continuousvariables. Letter labels denote homogeneity groups according to the results of the Tukey-Kramer multiplecomparison test. The significance test for the slope estimate:***P<0·001.
2009 Epiphytic lichens on the North-Estonian limestone escarpment—Jüriado et al. 85
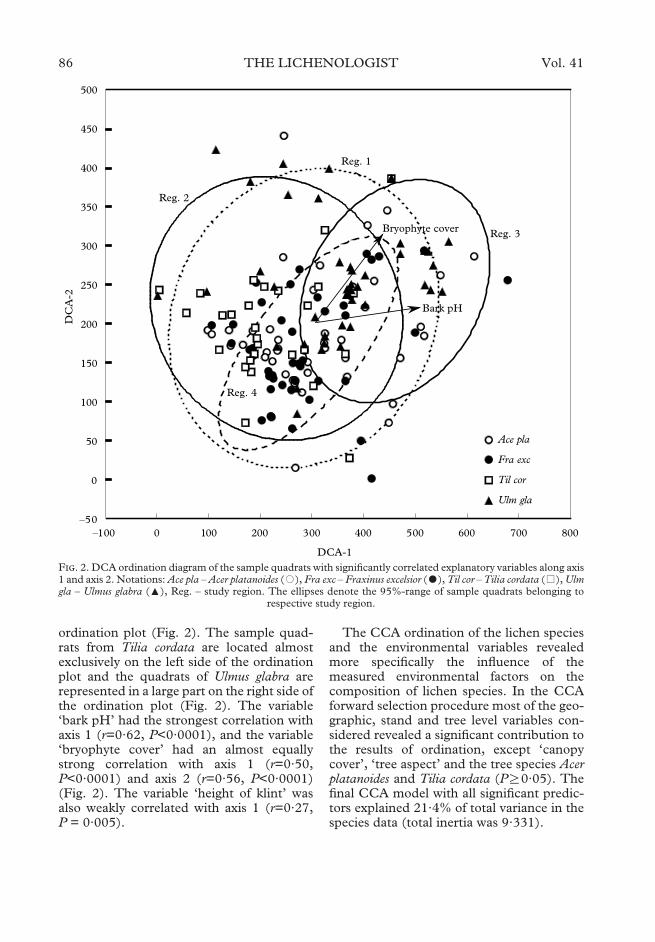
ordination plot (Fig. 2). The sample quad-rats from Tilia cordata are located almostexclusively on the left side of the ordinationplot and the quadrats of Ulmus glabra arerepresented in a large part on the right side ofthe ordination plot (Fig. 2). The variable‘bark pH’ had the strongest correlation withaxis 1 (r=0·62, P<0·0001), and the variable‘bryophyte cover’ had an almost equallystrong correlation with axis 1 (r=0·50,P<0·0001) and axis 2 (r=0·56, P<0·0001)(Fig. 2). The variable ‘height of klint’ wasalso weakly correlated with axis 1 (r=0·27,P = 0·005).
The CCA ordination of the lichen speciesand the environmental variables revealedmore specifically the influence of themeasured environmental factors on thecomposition of lichen species. In the CCAforward selection procedure most of the geo-graphic, stand and tree level variables con-sidered revealed a significant contribution tothe results of ordination, except ‘canopycover’, ‘tree aspect’ and the tree species Acerplatanoides and Tilia cordata (PR0·05). Thefinal CCA model with all significant predic-tors explained 21·4% of total variance in thespecies data (total inertia was 9·331).
F. 2. DCA ordination diagram of the sample quadrats with significantly correlated explanatory variables along axis1 and axis 2. Notations: Ace pla – Acer platanoides (�), Fra exc – Fraxinus excelsior (�), Til cor – Tilia cordata (�), Ulmgla – Ulmus glabra (:), Reg. – study region. The ellipses denote the 95%-range of sample quadrats belonging to
respective study region.
86 THE LICHENOLOGIST Vol. 41
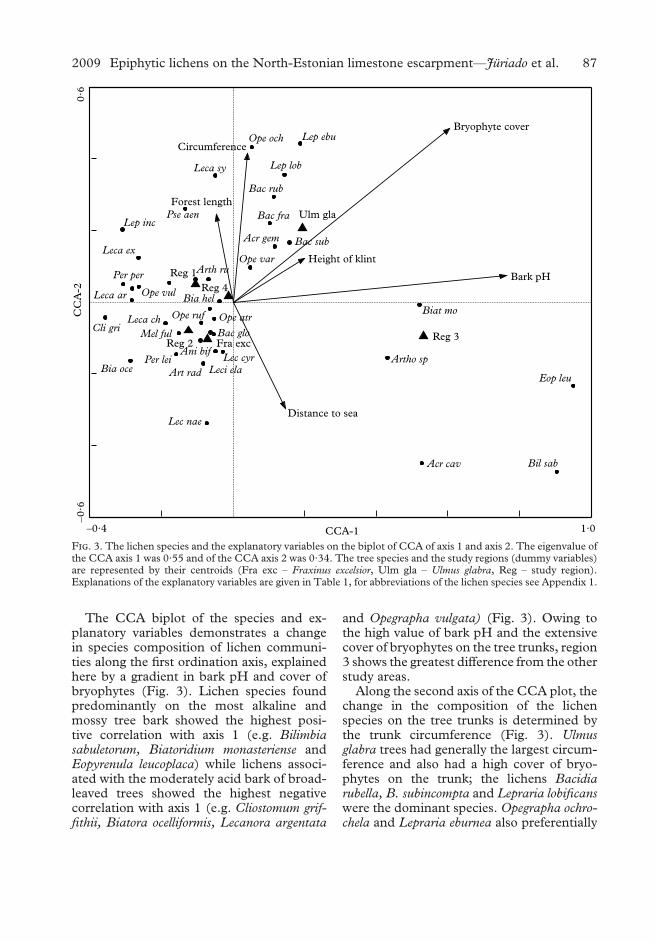
The CCA biplot of the species and ex-planatory variables demonstrates a changein species composition of lichen communi-ties along the first ordination axis, explainedhere by a gradient in bark pH and cover ofbryophytes (Fig. 3). Lichen species foundpredominantly on the most alkaline andmossy tree bark showed the highest posi-tive correlation with axis 1 (e.g. Bilimbiasabuletorum, Biatoridium monasteriense andEopyrenula leucoplaca) while lichens associ-ated with the moderately acid bark of broad-leaved trees showed the highest negativecorrelation with axis 1 (e.g. Cliostomum grif-fithii, Biatora ocelliformis, Lecanora argentata
and Opegrapha vulgata) (Fig. 3). Owing tothe high value of bark pH and the extensivecover of bryophytes on the tree trunks, region3 shows the greatest difference from the otherstudy areas.
Along the second axis of the CCA plot, thechange in the composition of the lichenspecies on the tree trunks is determined bythe trunk circumference (Fig. 3). Ulmusglabra trees had generally the largest circum-ference and also had a high cover of bryo-phytes on the trunk; the lichens Bacidiarubella, B. subincompta and Lepraria lobificanswere the dominant species. Opegrapha ochro-chela and Lepraria eburnea also preferentially
F. 3. The lichen species and the explanatory variables on the biplot of CCA of axis 1 and axis 2. The eigenvalue ofthe CCA axis 1 was 0·55 and of the CCA axis 2 was 0·34. The tree species and the study regions (dummy variables)are represented by their centroids (Fra exc – Fraxinus excelsior, Ulm gla – Ulmus glabra, Reg – study region).Explanations of the explanatory variables are given in Table 1, for abbreviations of the lichen species see Appendix 1.
2009 Epiphytic lichens on the North-Estonian limestone escarpment—Jüriado et al. 87

occurred on larger trees, while Lecanianaegelii, L. cyrtella and Lecidella elaeochromawere predominantly found on smaller trees.
The third axis of CCA is explained by theenvironmental conditions of klint forests andseparates the sites of region 4 from the otherstudy sites (Fig. 4). Stands in region 4 com-prised the longest fragment of the klint forest,located close to the coastline and on the talusslope of the highest part of the escarpment.Lichens characteristic of stands in region 4are those located on the upper left side ofthe ordination plot (e.g. Opegrapha atra,O. ochrochela and Chaenotheca brachypoda)(Fig. 4).
The partial CCA model indicated that thelargest amount of variance in the compos-ition of lichen communities on the tree bolewas explained by the set of tree level variables[11·2% of TVE (total variation explained)],followed by the set of geographic variables(6·9% of TVE) and the set of stand levelvariables (6·8% of TVE) (Table 3). Eachsubset of variables explained an independentcomponent of variation, while the fraction ofshared variation was marginal (Table 3).Tree level variables alone (‘bark pH’, ‘bryo-phyte cover’, ‘circumference’, ‘Ulmus glabra’and ‘Fraxinus excelsior’) explained 8·5% ofTVE when the other two environmental data
F. 4. The lichen species and the explanatory variables on the biplot of CCA of axis 1 and axis 3. The eigenvalue ofthe CCA axis 1 was 0·55 and of the CCA axis 3 was 0·30. Notations as in Fig. 3, explanations of the variables are
given in Table 1 and for abbreviations of the lichen species see Appendix 1.
88 THE LICHENOLOGIST Vol. 41

sets (geographic and stand level variables)were used as the covariables. The uniquevariation component of the stand level vari-ables (‘forest length’, ‘height of klint’ and‘distance to sea’) accounted for 5·2% of TVE(tree level and geographic variables wereused as the covariables) and that of the geo-graphic variables (Regions 1, 2, 3 and 4)accounted for 4·6% of TVE (tree and standlevel variables were used as the covariables)(Table 3).
Richness of lichen species
The stepwise building of the general linearmodel suggested that several environmentalvariables have a significant influence on thespecies richness of epiphytic lichen commu-nities (Table 4). In regions 1 and 4 the meannumber of lichen species differed signifi-cantly, being higher and lower, respectively,than those of regions 3 and 4. The significanteffect of tree species indicates that on aver-age, there was one more lichen species in thesample quadrats from Acer platanoides com-pared with those from Ulmus glabra (Table4). There were also significant differences
in species richness related to the aspect ofthe tree trunk relative to the escarpment. Thenumber of lichen species was lower on thesample quadrats facing the escarpment com-pared to the quadrats facing away from theescarpment (Table 4).
Species richness of lichens had an overallnegative correlation with bark pH (Table 4).The effect of bryophytes on species richnessof lichen communities was dependent upontree species. On Tilia cordata, lichen speciesrichness increased with increasing cover ofbryophytes, while the number of lichenspecies decreased with increasing moss coveron Acer platanoides (Table 4, Fig. 5). Thelength of the fragment of klint forest wasa significant positive predictor of speciesrichness of epiphytic lichen communities(Table 4).
Discussion
Species richness and composition of lichenson temperate broad-leaved trees on the talusslope of the limestone escarpment of theNorth-East Estonian Klint is substantiallyinfluenced both by tree level variables,unique habitat conditions of klint forest(length of the forest fragment, proximity ofthe stand to the sea and height of the escarp-ment) as well as geographical location of thestand. However, the tree level variables ex-plained the largest fraction of variation inrichness or composition of lichen species, ashas also been reported in other studies wherelichen diversity was evaluated at the tree levelwithin a limited study area (Aude & Poulsen2000; Löbel et al. 2006; Belinchón et al.2007).
As in a study of lichen communities ontemperate broad-leaved trees in floodplainforests (Jüriado et al. 2008), changes in lichenassemblages on broad-leaved trees in klintforest are mainly caused by the co-variationof the bryophyte cover with bark pH andtrunk circumference (Figs 3 & 4). As themean bark pH of the studied trees was usu-ally subneutral, a decrease of species rich-ness with increasing bark pH, as well as thenegative effect of bryophyte cover on species
T 3. Partitioning of variation in epiphytic lichencommunities using three sets of explanatory variables in
variation partitioning analysis (pCCA)
Unique components* EV (IU)† FTVE(%)
TgSgG 1·990 21·4T 1·042 11·2S 0·631 6·8G 0·644 6·9T|SgG 0·483 5·2G|TgS 0·426 4·6ThS|G 0·065 0·7ThG|S 0·136 1·5ShG|T 0·040 0·4
*T = tree level environmental variables (‘bark pH’,‘bryophyte cover’, ‘circumference’, ‘Ulmus glabra?’ and‘Fraxinus excelsior’); S = stand level variables (‘forestlength’, ‘height of klint’ and ‘distance to sea’); G =geographical variables (Regions 1, 2, 3 and 4) (see Table1). Symbol ‘|’ means covarying out variable sets, ‘g’refers to union of variable sets, ‘h’ refers to intersectionof variable sets.†EV (the sum of all canonical eigenvalues) is given ininertia units (IU; total inertia 9.331 TU) as well as thefraction of the total variation explained (FTVE; %).
2009 Epiphytic lichens on the North-Estonian limestone escarpment—Jüriado et al. 89

richness of lichen communities, were ex-pected (Jüriado et al. 2008). In this study, wealso detected a reverse effect of moss cover onlichen species richness on Tilia cordata. Tiliahad a comparatively low cover of bryophyteson the tree bole compared with the other treespecies studied (Fig. 5); increasing bryophytecover appears to promote lichen species rich-ness on this tree species. This relationshipcould be attributed to a situation in whichboth lichens and bryophytes have enoughspace on the tree bole and optimal conditionsfor growth.
In the case of mature or older broad-leavedtrees, the effect of trunk size on lichen speciesrichness on the tree bole has proved to benegative (Jüriado et al. 2008), or is of minorimportance when compared to other factors(Löbel et al. 2006; Johansson et al. 2007).However, changes in the composition oflichen specie communities with increasingbole size have been documented in manyother studies (Burgaz et al. 1994; Moe &
Botnen 1997; Belinchón et al. 2007). Altera-tions in the physical and chemical propertiesof tree bark with tree ageing may induce atransition from a crustose community, domi-nated by sexually reproducing species, to anassembly dominated by asexually reproduc-ing species (Ellis & Coppins 2007b).
Owing to the steep slope of the escarp-ment, the trunks of the trees growing on it areleaning which can explain the difference inthe species richness of lichens between thedifferent aspects of the trunk. On the side ofthe trunk facing the escarpment, bryophytesare probably favoured by the larger amountof stem flow as well as by increased shade(Gustafsson & Eriksson 1995; Moe &Botnen 2000). The side of the trunk facingaway from the escarpment is apparently drierand better illuminated, which has led to thedominance of lichen species.
The extraordinary environmental con-ditions of klint forests have a unique effecton the composition and richness of lichen
T 4. The results of general linear mixed model analysis (GLMM) for number of lichen species (log-transformed) ontemperate broad-leaved trees
Effect df P Mean Slope
Study region 3; 78 0·009Region 1 5·6a
Region 2 4·4Region 3 5·4Region 4 3·4b
Tree species 3; 78 0·008Acer platanoides 5·2a
Fraxinus excelsior 4·5Tilia cordata 4·9Ulmus glabra 4·0b
Bryophyte cover 1; 72 0·773Bryophyte cover*Tree species 3; 72 0·004
Acer platanoides 0·014 −0·049*Fraxinus excelsior 0·275 −0·022Tilia cordata 0·005 0·107**Ulmus glabra 0·361 −0·019
Tree aspect 1; 72 0·0481 (facing the escarpment) 4·3a
2 (facing away from the escarpment) 4·9b
Bark pH 1; 72 <0·0001 −0·310***Forest length 1; 78 0·0004 0·507**
Explanations of the explanatory variables are given in Table 1, notations as in Table 2. The significance test for theslope estimates:*P%0.05, **P<0.01, ***P<0-001.
90 THE LICHENOLOGIST Vol. 41

communities. Stands on the longest stretchof the klint forest, close to the sea, and onthe highest slope of the escarpment supportlichens that are not found, or are rare, inother regions (e.g. Opegrapha atra, O. ochro-chela and Pyrenula nitidella). For dispersal ofepiphytic lichens (Löbel et al. 2006; Ellis &Coppins 2007a), the connectivity of narrowforest fragments along the talus slope of theescarpment may be important whereas inmore extensive landscapes such stands areusually distinctly isolated from other forestareas.
In general, habitat area has proved tobe positively correlated with number ofspecies (Gotelli & Graves 1996; Berglund &Jonsson 2001; Jüriado et al. 2006). Wefound that the fragment length of klint forestis a significant positive predictor of speciesrichness of lichen communities on the tem-perate broad-leaved trees (Table 4). How-ever, previous studies on the diversity of
vascular plants and bryophytes in Estonianklint forests have not found a significant re-lationship between length of the forest frag-ment and species richness (Ingerpuu et al.2003). The difference between the resultsof our study and those reported by Ingerpuuet al. (2003) might be due to the differencein the treatment of respective variables indata analysis. Ingerpuu et al. (2003) dividedforest fragments into three size classes, whilein the present study the absolute dimensionsof forest fragments were used.
We found a significant effect of standlocality on the composition and richness oflichen communities on the tree bole, which ismost likely explained by a gradient in local airpollution. Owing to the extraordinarily highlevel of alkaline pollutants originating from alocal cement plant during the past decades(Mandre 1995), the epiphytic lichen com-munities have changed near study region 3(Martin & Nilson 1992; Nilson 1995). In
F. 5. Relationship between number of lichen species and cover of bryophytes according to general linear mixedmodel analysis (GLMM) for different broad-leaved tree species (see Table 4). A, Acer platanoides; B, Fraxinusexcelsior, C, Tilia cordata, D, Ulmus glabra. The scale of ‘Number of lichen species’ is log-transformed and the scale
of ‘Bryophyte cover’ is square-root transformed
2009 Epiphytic lichens on the North-Estonian limestone escarpment—Jüriado et al. 91

this area of klint forests, bark is alkaline andunder the shelter of the closed tree canopy,the tree trunks are covered with an extensivemat of bryophytes accompanied by bryo-philous lichens (e.g. Bilimbia sabuletorum),or lichen species favouring high bark pH (e.g.Biatoridium monasteriense and Eopyrenulaleucoplaca) (Figs 3 & 4).
In the easternmost part of the klint forest(study region 4) increasing acidificationcould explain the lower species richness oflichens compared with the klint forests in thewestern part of the country (Table 4). Inrecent decades, the proportion of SO2 in theratio of alkaline dust to acid components inthe north-eastern part of Estonia (Fig. 1) hasincreased, which might indicate that SO2emissions have reached a level affectinglichen and bryophyte species (Liblik et al.2000; Liblik & Pensa 2001).
Paal et al. (2001) and Ingerpuu et al.(2003) have emphasized the high con-servation value of Estonian klint forests forvascular plants and bryophytes and the needfor the strict protection of these habitatsthroughout their range in Estonia. We foundthat klint forests support lichen species re-stricted to only a few localities in Estonia(e.g. Anisomeridium biforme, Arthopyreniaspp., Lecidea albohyalina) or not found inother forest types here (Eopyrenula leuco-placa) (Database of Estonian Lichens 2008).In addition, klint forests account for a sub-stantial proportion of the national localitiesof red-listed or protected lichens (Appendix1). In spite of the presumably detrimentaleffect of air pollution on epiphytic lichens insome places, we consider that klint forestsform a valuable habitat from the lichenologi-cal perspective.The authors are grateful to A. Jüriado, K. and Kr.Jürgens, and R. Vaske for assistance in fieldwork. Specialthanks belong to L. Saag who carried out TLC analysesand identified some specimens. We also thank A. Suijaand two anonymous referees for their helpful commentson the manuscript. E. Jaigma is acknowledged for revis-ing the English text. Financial support was received fromthe Estonian Ministry of Education and Research (tar-geted financing Nos. SF0182639s04, SF0180153s08and SF0180098sO8). This research was also supportedby the European Union through the European RegionalDevelopment Fund and by the Archimedes Foundation(grant RLOOMTIPP).
REFERENCES
Akaike, H. (1973) Information theory and an extensionof the maximum likelihood principle. In SecondInternational Symposium on Information Theory(B. N. Petrov & F. Csáki, eds): 267–281. Budapest:Akadémiai Kiadó.
Aude, E. & Poulsen, R. S. (2000) Influence of manage-ment on the species composition of epiphytic cryp-togams in Danish Fagus forests. Applied VegetationScience 3: 81–88.
Barkman, J. J. (1958) Phytosociology and Ecology ofCryptogamic Epiphytes. Assen: Van Gorcum &Comp. N. V.
Belinchón, R., Martinéz, I., Escudero, A., Aragón, G. &Valladares, F. (2007) Edge effects on epiphyticcommunities in a Mediterranean Quercus pyrenaicaforest. Journal of Vegetation Science 18: 81–90.
Berg, Å., Gärdenfors, U., Hallingbäck, T. & Nordén,M. (2002) Habitat preferences of red-listed fungiand bryophytes in woodland key habitats in south-ern Sweden – analyses of data from a national sur-vey. Biodiversity and Conservation 11: 1479–1503.
Berglund, H. & Jonsson, B. G. (2001) Predictability ofplant and fungal species richness of old-growthboreal forest islands. Journal of Vegetation Science 12:857–866.
Borcard, D., Legendre, P. & Drapeau, P. (1992) Par-tialling out the spatial component of ecological vari-ation. Ecology 73: 1045–1055.
Boudreault, C., Gauthier, S. & Bergeron, Y. (2000)Epiphytic lichens and bryophytes on Populus tremu-loides along a chronosequence in the southwesternboreal forest of Québec, Canada. Bryologist 103:725–738.
Brodo, I. M. (1973) Substrate ecology. In The Lichens(V. Ahmadjian & M. E. Hale, eds): 401–441. NewYork and London: Academic Press.
Burgaz, A. R., Fuertes, E. & Escudero, A. (1994) Ecol-ogy of cryptogamic epiphytes and their communi-ties in deciduous forests in mediterranean Spain.Vegetatio 112: 73–86.
Cieslinski, S., Czyzewska, K., Klama, H. & Zarnowiec,J. (1996) Epiphytes and epiphytism. Phytocoenosis 8:15–35.
Database of Estonian Lichens (2008) eSamba – TartuÜlikooli Loodusmuuseumi (TU) Eesti samblikeandmebaas. http://www.ut.ee/ial5/lich/baasid/esamba.html. Cited 10 Aug 2008. [In Estonian].
Diekmann, M. (1996) Ecological behaviour of decidu-ous hardwood trees in Boreo-nemoral Sweden inrelation to light and soil conditions. Forest Ecologyand Management 86: 1–14.
EC (1992) Council directive 92/43/EEC of 21 May1992 on the conservation of natural habitats and ofwild fauna and flora. Official Journal of EuropeanCommunity L 206: 7–50.
Ellis, C. J. & Coppins, B. J. (2007a) 19th centurywoodland structure controls stand-scale epiphytediversity in present-day Scotland. Diversity andDistributions 13: 84–91.
92 THE LICHENOLOGIST Vol. 41

Ellis, C. J. & Coppins, B. J. (2007b) Reproductive strat-egy and the compositional dynamics of crustoselichen communities on aspen (Populus tremula L.) inScotland. Lichenologist 39: 377–391.
Esseen, P.-A., Ehnström, B., Ericson, L. & Sjöberg, K.(1997) Boreal forests. Ecological Bulletins 46: 16–47.
Gotelli, N. & Graves, G. (1996) Null Models in Ecology.Washington and London: Smithsonian InstitutionPress.
Gustafsson, L. & Eriksson, I. (1995) Factors of impor-tance for the epiphytic vegetation of aspen Populustremula with special emphasis on bark chemistry andsoil chemistry. Journal of Applied Ecology 32: 412–424.
Gustafsson, L., de Jong, J. & Norén, M. (1999) Evalu-ation of Swedish woodland key habitats using red-listed bryophytes and lichens. Biodiversity andConservation 8: 1101–1114.
Haapala, H., Goltsova, N., Seppälä, R., Huttunen, S.,Kouki, J., Lamppu, J. & Popovichev, B. (1996)Ecological condition of forests around the easternpart of the Gulf of Finland. Environmental Pollution91: 253–265.
Hansson, L. (ed) (1997) Boreal ecosystems and land-scapes; structures processes and conservation ofbiodiversity. Ecological Bulletins 46: 1–203.
Hill, M. O. & Gauch, H. G. J. (1980) Detrended corre-spondence analysis: an improved ordination tech-nique. Vegetatio 42: 47–58.
Ingerpuu, N., Vellak, K., Liira, J. & Pärtel, M. (2003)Relationships between species richness patterns indeciduous forests at the north Estonian limestoneescarpment. Journal of Vegetation Science 14: 773–780.
Johansson, P., Rydin, H. & Thor, G. (2007) Tree agerelationships with epiphytic lichen diversity andlichen life history traits on ash in southern Sweden.Ecoscience 14: 81–91.
Jüriado, I., Liira, J. & Paal, J. (2003) Epiphytic andepixylic lichen species diversity in Estonian naturalforests. Biodiversity and Conservation 12: 1587–1607.
Jüriado, I., Suija, A. & Liira, J. (2006) Biogeographicaldeterminants of lichen species diversity on isletsin the West-Estonian Archipelago. Journal of Veg-etation Science 17: 125–134.
Jüriado, I., Liira, J., Paal, J. & Suija, A. (2008) Tree andstand level variables influencing diversity of lichenson temperate broad-leaved trees in boreo-nemoralfloodplain forests. Biodiversity and Conservation(DOI 10.1007/s10531-008-9460-y).
Kalda, A. (1962) Die Edellaubwälder Estlands. In Fra-gen der Bewirtschaftung der Hainwälder (H. Rebane,ed): 129–135. Tartu: Teaduste Akadeemia [InEstonian with German summary].
Keskkonnaministri määrus nr 51 (2004) III kaitsekate-gooria liikide kaitse ala võtmine (Regulation of theMinister of the Environment No. 51. 19.05.2004).Riigi Teataja Lisa 27.05.2004, 69: 1134.
Laasimer, L. (1965) Vegetation of the Estonian S.S.R.Tallinn: Valgus [In Estonian with English sum-mary].
Liblik, V. (2007) Temporal changes in atmospheric airpollution in Ida-Viru county and impact of pollut-ants on the nature. A review of investigations inthe years 1990–2005. In Problems of ContemporaryEnvironmental Studies (J.-M. Punning, ed): 173–208. Tallinn: Tallinn University, Institute of Ecol-ogy [In Estonian with English summary].
Liblik, V. & Pensa, M. (2001) Specifics and temporalchanges in air pollution in areas affected by emis-sions from oil shale industry, Estonia. Water, Air,and Soil Pollution 130: 1787–1792.
Liblik, V., Pensa, M. & Kundel, H. (2000) Temporalchanges in atmospheric air pollution in industrialareas of Ida- and Lääne-Viru counties. ForestryStudies 33: 17–36 [In Estonian with English sum-mary].
Littell, R. C., Milliken, G. A., Stroup, W. W. &Wolfinger, R. D. (1996) SAS System for MixedModels. Cary: SAS Institute Inc.
Lõhmus, A., Kohv, K., Palo, A. & Viilma, K. (2004)Loss of old-growth, and the minimum need forstrictly protected forests in Estonia. Ecological Bul-letins 51: 401–411.
Löbel, S., Snäil, T. & Rydin, H. (2006) Species richnesspatterns and metapopulation processes – evidencefrom epiphyte communities in boreo-nemoral for-ests. Ecography 29: 169–182.
Mandre, M. (ed) (1995) Dust Pollution and Forest Eco-systems. A Study of Conifers in an Alkalized Environ-ment. Tallinn: Institute of Ecology.
Martin, L. & Nilson, E. (1992) Impact of the Kundacement plant (North-East Estonia) emission on thedistribution of epiphytic lichens. Proceedings of theEstonian Academy of Sciences, Ecology 2: 181–185.
McCune, B. & Grace, J. B. (2002) Analysis of EcologicalCommunities. Oregon: Glenden Beach.
McCune, B. & Mefford, M. J. (1999) PC-ORD.Multivariate Analysis of Ecological Data, Version 4.Oregon: Gleneden Beach.
Mežaka, A., Brumelis, G. & Piterans, A. (2008) Thedistribution of epiphytic bryophyte and lichenspecies in relation to phorophyte characters inLatvian natural old-growth broad leaved forests.Folia Cryptogamica Estonica 44: 89–99.
Miidel, A. (1997) Escarpments and waterfalls. InGeology and Mineral Resources of Estonia (A. Raukas& A. Teedumäe. eds): 391–395. Tallinn: EstonianAcademy Publishers.
Moe, B. & Botnen, A. (1997) A quantitative study of theepiphytic vegetation on pollarded trunks of Fraxinusexcelsior at Havrå Osterøy, western Norway. PlantEcology 129: 157–177.
Moe, B. & Botnen, A. (2000) Epiphytic vegetation onpollarded trunks of Fraxinus excelsior in four differ-ent habitats at Grinde, Leikanger, western Norway.Plant Ecology 151: 143–159.
Nilson, E. (1995) Species composition and structure ofepiphytic lichen assemblages on Scots pine aroundthe Kunda cement plant. In Dust Pollution and ForestEcosystems. A Study of Conifers in an AlkalizedEnvironment (M. Mandre, ed): 134–140. Tallinn:Institute of Ecology.
2009 Epiphytic lichens on the North-Estonian limestone escarpment—Jüriado et al. 93

Nilsson, S. G. (1992) Forests in the temperate–borealtransition–natural and man-made features. In Eco-logical Principles of Nature Conservation. Applicationsin Temperate and Boreal Environments (L. Hansson,ed): 373–393. London and New York: ElsevierApplied Science.
Paal, J. (1997) Classification of Estonian Vegetation SiteTypes. Tallinn: Estonian Environment InformationCentre [In Estonian].
Paal, J. (1998) Rare and threatened plant communitiesof Estonia. Biodiversity and Conservation 7: 1027–1049.
Paal, J. (2001) The North-Estonian klint: soil and veg-etation diversity. Publicationes Instituti GeographiciUniversitatis Tartuensis 92: 761–764.
Paal, J. (2007) Eesti pangametsade tüpoloogia. In XXXEstonian Naturalists’ Congress. Nature of Klint Areas.June 30 – July 1, 2007 (I. Puura, S. Pihu & L. Amon,eds): 55–64. Nõva, Estonia [In Estonian].
Paal, J., Vellak, K. & Ingerpuu, N. (2001) Speciescontent of the Estonian klint forests, their classifica-tion structure and their correlation with the mainsoil parameters. Forestry Studies 35: 104–132 [InEstonian with English summary].
Price, K. & Hochachka, G. (2001) Epiphytic lichenabundance: effects of stand age and composition incoastal British Columbia. Ecological Applications 11:904–913.
Randlane, T. & Saag, A. (eds) (1999) Second checklistof lichenized, lichenicolous and allied fungi ofEstonia. Folia Cryptogamica Estonica 35: 1–132.
Randlane, T., Jüriado, I., Suija, A., Lóhmus, P. &Leppik, E. (2008) Lichens in the new Red List ofEstonia. Folia Cryptogamica Estonica 44: 113–120.
Randlane, T., Saag, A. & Suija, A. (2007) Lichenized,lichenicolous and allied fungi of Estonia. http://www.ut.ee/lichens/fce.html. Cited 11 Nov 2007.
Raukas, A. & Teedumäe, A. (eds) (1997) Geology andMineral Resources of Estonia. Tallinn: EstonianAcademy Publishers.
SAS Institute Inc. (1989) SAS/STAT® User’s Guide.Version 6. Fourth Edition. Cary: SAS Institute Inc.
Scheidegger, C. & Goward, T. (2002) Monitoring li-chens for conservation: red lists and conservationaction plans. In: Monitoring with Lichens – Monitor-ing Lichens. (P. L. Nimis, C. Scheidegger & P. A.Wolseley, eds): 163–181. Dordrecht: Kluwer Aca-demic Publishers.
Sillett, S. C., McCune, B., Peck, J. E., Rambo, T. R. &Ruchty, A. (2000) Dispersal limitations of epiphyticlichens result in species dependent on old-growthforests. Ecological Applications 10: 789–799.
Shao, J. (1997) An asymptotic theory for linear modelselection. Statistica Sinica 7: 221–264.
Snäll, T., Ribeiro, P. J. & Rydin, H. (2003) Spatialoccurrence and colonisations in patch-trackingmetapopulations: local conditions versus dispersal.Oikos 103: 566–578.
StatSoft, Inc. (2005) Statistica for Windows, Ver 7.1.Tulsa: Statsoft, Inc.
Suuroja, K. (2005) Põhja-Eesti klint. Tallinn: TallinnaRaamatutrükikoda [In Estonian].
ter Braak, C. J. F. (1986) Canonical correspondenceanalysis: a new eigenvector technique for multi-variate direct gradient analysis. Ecology 67: 1167–1179.
ter Braak, C. J. F. & Šmilauer, P. (2002) CANOCOReference Manual and CanoDrawfor Windows User’sGuide: Software for Canonical Community Ordination(version 4.5). Ithaca, New York: MicrocomputerPower.
Thor, G. (1998) Red-listed lichens in Sweden: habitats,threats, protection, and indicator value in borealconiferous forests. Biodiversity and Conservation 7:59–72.
Vabariigi Valitsuse määrus nr 195. (2004) I ja II kait-sekategooriana kaitse ala võetavate liikide loetelu(Regulation of the Government of Estonia No. 195.20.05.2004). Riigi Teataja I 21.05.2004, 44: 313.
Valk, U. & Eilart, J. (1974) Estonian Forests. Tallinn:Valgus [In Estonian].
Web map server of Estonian Land Board (2007) X-GISsystem. http://xgis.maaamet.ee Cited 10 Oct 2007.
Accepted for publication 14 October 2008
94 THE LICHENOLOGIST Vol. 41

Appendix 1. List of the recorded species and their occurrence on fourbroad-leaved tree species in boreo-nemoral forest on the talus slope of the
North-Estonian limestone escarpment
Species (the abbreviations of species’ names are presented in the brackets) A. p. F.e. T.c. U.g.
Acrocordia cavata (Ach.) R. C. Harris (Acr cav) + + +A. gemmata (Ach.) A. Massal. (Acr gem) + + + +Amandinea punctata (Hoffm.) Coppins. & Scheid +Anisomeridium biforme (Borrer & Sowerb.) R. C. Harris (Ani bif)* + + +Arthonia byssacea (Weigel) Almq.** +A. didyma Körb. (Art did)NT + + + +A. mediella Nyl. +A. radiata (Pers.) Ach. (Art rad) + + + +A. vinosa Leight. + +Arthopyrenia spp. (Artho sp) + + +Arthothelium ruanum (A. Massal.) Körb. (Arth ru) + + + +Bacidia beckhausii Körb. + +B. fraxinea Lönnr. (Bac fra) + + +B. globulosa (Flörke) Hafellner & V. Wirth (Bac glo) + + +B. rubella (Hoffm.) A. Massal. (Bac rub) + + + +B. subincompta (Nyl.) Arnold (Bac sub) + + +Biatora efflorescens (Hedl.) Räsänen +B. helvola Körb. +B. ocelliformis (Nyl.) Arnold (Bia oce) + + +Biatoridium monasteriense J. Lahm ex Körb. (Biat mo)**,NT + + +Bilimbia sabuletorum (Schreb.) Arnold (Bil sab) + + +Buellia disciformis (Fr.) Mudd. + +B. griseovirens (Turner & Borrer ex Sm.) Almb. +Chaenotheca brachypoda (Ach.) Tibell (Cha bra) + +C. furfuracea (L.) Tibell +C. xyloxena Nádv. +Chaenothecopsis pusilla (Ach.) A. F. W. Schmidt +Cladonia fimbriata (L.) Fr. +Cladonia sp. +Cliostomum griffithii (Sm.) Coppins (Cli gri) + + +Dimerella pineti (Ach.) Vezda +Diplotomma pharcidium (Ach.) M. Choisy* +Eopyrenula leucoplaca (Wallr.) R. C. Harris (Eop leu)*,EN + + +Graphis scripta (L.) Ach. (Gra scr) + + + +Gyalecta truncigena (Ach.) Hepp + +Lecania cyrtella (Ach.) Th. Fr. (Lec cyr) + + +L naegelii (Hepp) Diederich & Van den Boom (Lec nae) +Lecanora argentata (Ach.) Malme (Leca ar) + + +L. carpinea (L.) Vain. +L. chlarotera Nyl. (Leca ch) + + + +L. conizaeoides Nyl. ex Cromb. +L. expallens Ach. (Leca ex) +L. intumescens (Rebent.) Rabenh.* +L. leptyrodes (Nyl.) Degel. + +L. symmicta (Ach.) Ach. (Leca sy) + + +Lecidea albohyalina (Nyl.) Th. Fr.* +L. erythrophaea Flörke ex Sommerf.**,NT + +Lecidella elaeochroma (Ach.) M. Choisy (Leci ela) + + + +Lepraria eburnea J. R. Laundon (Lep ebu) + + +L. elobata Tønsberg* +L. incana (L.) Ach. (Lep inc) + + +
2009 Epiphytic lichens on the North-Estonian limestone escarpment—Jüriado et al. 95

Appendix 1. continued.
Species (the abbreviations of species’ names are presented in the brackets) A. p. F.e. T.c. U.g.
L. lobificans Nyl. (Lep lob) + + + +Leptogium lichenodes (L.) Zahlbr. +Melanelia fuliginosa (Fr. ex Duby) Essl. (Mel ful) + +M. subaurifera (Nyl.) Essl. + + +Ochrolechia androgyna (Hoffm.) Arnold +Opegrapha atra Pers. (Ope atr)NT + + + +O. ochrocheila Nyl. (Ope och)*,VU + + +O. rufescens Pers. (Ope ruf) + + + +O. varia Pers. (Ope var) + + + +O. viridis (Pers. ex Ach.) Behlen & Desberg*,VU +O. vulgata Ach. (Ope vul) + + + +Pertusaria amara (Ach.) Nyl. + +P. leioplaca DC. (Per lei) + + + +P. pertusa (Weigel) Tuck. (Per per) + +Phaeophyscia orbicularis (Neck.) Moberg +Phlyctis agelaea (Ach.) Flot. +P. argena (Spreng.) Flot. (Phl arg) + + + +Placynthiella icmalea (Ach.) Coppins & P. James +Pseudosagedia aenea (Wallr.) Hafellner & Kalb. (Pse aen)* + +Pyrenula nitidella (Flörke ex Schraer.) Müll. Arg.**,VU +Ramalina farinacea (L.) Ach. + +Schismatomma pericleum (Ach.) Branth & Rostr. +
A. p. – Acer platanoides, F. e. – Fraxinus excelsior, T. c. – Tilia cordata and U. g. – Ulmus glabra. Rare species in Estoniaare marked with an asterisk (*) and species under protection are marked with two asterisks (**). Red-listed lichenspecies are assigned the abbrevation of red-list category: EN – endagered, NT – near threatened, VU – vulnerable.
96 THE LICHENOLOGIST Vol. 41




















