
Diversity of culturable actinobacteria from Qinghai–Tibet plateau, China
11
ORIGINAL PAPER Diversity of culturable actinobacteria from Qinghai–Tibet plateau, China Yu-Qin Zhang • Hong-Yu Liu • Jie Chen • Li-Jie Yuan • Wei Sun • Li-Xin Zhang • Yue-Qin Zhang • Li-Yan Yu • Wen-Jun Li Received: 3 December 2009 / Accepted: 22 March 2010 / Published online: 2 April 2010 Ó Springer Science+Business Media B.V. 2010 Abstract To investigate the diversity of culturable actinobacteria and further screen for microbial pharma- ceutics, seven different media were chosen to isolate actinobacteria from 87 soil samples collected from Qinghai–Tibet plateau. A total of 1930 strains was isolated and identified to belong to 11 suborders, i.e., Actinopolysporineae, Corynebacterineae, Frankineae, Glycomycineae, Kineosporiineae, Micrococcineae, Micromonosporineae, Propionibacterineae, Pseudono- cardineae, Streptomycineae and Streptosporangineae, and 16 families, i.e., Nocardioidaceae, Actinopolyspor- aceae, Actinosynnemataceae, Dermacoccaceae, Geo- dermatophilaceae, Glycomycetaceae, Kineosporiaceae, Microbacteriaceae, Micromonosporaceae, Nocardia- ceae, Promicromonosporaceae, Propionibacteriaceae, Pseudonocardiaceae, Streptomycetaceae, Streptospo- rangiaceae and Thermomonosporaceae. A primary taxonomic study showed that at least 22 genera of actinobacteria were identified from the soil samples, among which ten isolates represented hitherto unknown species. The results showed that there was abundant actinobacterial species diversity in the soil samples from the Qinghai–Tibet plateau. Electronic supplementary material The online version of this article (doi:10.1007/s10482-010-9434-4) contains supple- mentary material, which is available to authorized users. Y.-Q. Zhang H.-Y. Liu J. Chen L.-J. Yuan W. Sun Y.-Q. Zhang L.-Y. Yu (&) Institute of Medicinal Biotechnology, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing 100050, People’s Republic of China e-mail: [email protected] Y.-Q. Zhang e-mail: [email protected] W.-J. Li (&) The Key Laboratory for Microbial Resources of the Ministry of Education, Kunming, People’s Republic of China e-mail: [email protected] W.-J. Li Laboratory for Conservation and Utilization of Bio- Resources, Yunnan Institute of Microbiology, Yunnan University, Kunming 650091, People’s Republic of China L.-X. Zhang W.-J. Li Guangdong Key Laboratory of Marine Materia Medica, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, People’s Republic of China L.-X. Zhang W.-J. Li Institute of Microbiology, Chinese Academy of Sciences, 100101 Beijing, People’s Republic of China 123 Antonie van Leeuwenhoek (2010) 98:213–223 DOI 10.1007/s10482-010-9434-4
Transcript of Diversity of culturable actinobacteria from Qinghai–Tibet plateau, China

ORIGINAL PAPER
Diversity of culturable actinobacteria from Qinghai–Tibetplateau, China
Yu-Qin Zhang • Hong-Yu Liu • Jie Chen •
Li-Jie Yuan • Wei Sun • Li-Xin Zhang •
Yue-Qin Zhang • Li-Yan Yu • Wen-Jun Li
Received: 3 December 2009 / Accepted: 22 March 2010 / Published online: 2 April 2010
� Springer Science+Business Media B.V. 2010
Abstract To investigate the diversity of culturable
actinobacteria and further screen for microbial pharma-
ceutics, seven different media were chosen to isolate
actinobacteria from 87 soil samples collected from
Qinghai–Tibet plateau. A total of 1930 strains was
isolated and identified to belong to 11 suborders, i.e.,
Actinopolysporineae, Corynebacterineae, Frankineae,
Glycomycineae, Kineosporiineae, Micrococcineae,
Micromonosporineae, Propionibacterineae, Pseudono-
cardineae, Streptomycineae and Streptosporangineae,
and 16 families, i.e., Nocardioidaceae, Actinopolyspor-
aceae, Actinosynnemataceae, Dermacoccaceae, Geo-
dermatophilaceae, Glycomycetaceae, Kineosporiaceae,
Microbacteriaceae, Micromonosporaceae, Nocardia-
ceae, Promicromonosporaceae, Propionibacteriaceae,
Pseudonocardiaceae, Streptomycetaceae, Streptospo-
rangiaceae and Thermomonosporaceae. A primary
taxonomic study showed that at least 22 genera of
actinobacteria were identified from the soil samples,
among which ten isolates represented hitherto
unknown species. The results showed that there was
abundant actinobacterial species diversity in the soil
samples from the Qinghai–Tibet plateau.Electronic supplementary material The online version ofthis article (doi:10.1007/s10482-010-9434-4) contains supple-mentary material, which is available to authorized users.
Y.-Q. Zhang � H.-Y. Liu � J. Chen � L.-J. Yuan �W. Sun � Y.-Q. Zhang � L.-Y. Yu (&)
Institute of Medicinal Biotechnology, Chinese Academy
of Medical Sciences and Peking Union Medical College,
Beijing 100050, People’s Republic of China
e-mail: [email protected]
Y.-Q. Zhang
e-mail: [email protected]
W.-J. Li (&)
The Key Laboratory for Microbial Resources of the
Ministry of Education, Kunming, People’s Republic of
China
e-mail: [email protected]
W.-J. Li
Laboratory for Conservation and Utilization of Bio-
Resources, Yunnan Institute of Microbiology, Yunnan
University, Kunming 650091, People’s Republic of China
L.-X. Zhang � W.-J. Li
Guangdong Key Laboratory of Marine Materia Medica,
South China Sea Institute of Oceanology, Chinese
Academy of Sciences, Guangzhou 510301, People’s
Republic of China
L.-X. Zhang � W.-J. Li
Institute of Microbiology, Chinese Academy of Sciences,
100101 Beijing, People’s Republic of China
123
Antonie van Leeuwenhoek (2010) 98:213–223
DOI 10.1007/s10482-010-9434-4

Keywords Actinobacteria � 16S rRNA �Isolation � Identification
Introduction
It is becoming increasingly clear that un- and under-
explored habitats are a rich source of novel actinobac-
teria which have the capacity to produce interesting new
bioactive compounds, including antibiotics (Bull et al.
2005; Bull and Stach 2007; Okoro et al. 2008). The
Qinghai–Tibet plateau is the largest plateau in China
(about 2,400,000 km2) and the highest plateau in the
world (the average altitude is above 4000 m), encom-
passing Qinghai province, the Tibet Autonomous
Region, the western part of Sichuan and parts of Gansu
and Yunnan provinces. The complex climatic factors
and geographical topography of the Qinghai–Tibet
plateau promote the formation of a unique ecological
environment. In recent studies, several new taxa of
bacteria (Li et al. 2005a, 2006; Zhang et al. 2008) and
actinobacteria (Li et al. 2005b; Zhang et al. 2005, 2007)
were reported to have been isolated from the Qingha–
Tibet plateau, which show that such habitats are a
promising source of microorganisms. This has encour-
aged further studies on the biodiversity of actinomycetes
from the Qinghai–Tibet plateau. The present study
focused on the isolation and identification of Actino-
bacteria to reveal the diversity of culturable actinobac-
teria from Qinghai–Tibet plateau.
Materials and methods
Soil samples
Soil samples collected from different locales on
Qinghai–Tibet plateau were taken from a depth of
10–15 cm and kept in sterilized paper bags at 4�C
before processing. The details of the 87 samples used
for the isolation of actinobacteria are: 58 samples (No.
1–58) were from Qinghai Province; most of the
sampling locations were in hyper-arid areas (latitude
36�350–37�360N, longitude 99�520–101�460E, eleva-
tion 3000–3900 m). Seven samples were collected
from the Xinjiang Uygur Autonomous Region (latitude
40�420–43�530N, longitude 88�080–89�220E, elevation
700–1900 m), 15 samples from Deqen in Yunnan
Province (latitude 28�080–28�280N, longitude 103�540–
104�280E, elevation 800–1920 m), and seven samples
were collected at Ganzi Tibetan Autonomous Prefecture
in Sichuan Province (latitude 27�300–29�580N, longi-
tude 102�180–108�110E, elevation 1200–2000 m). The
pH value for most soil samples is around 7.0–8.0
(neutral to alkalescent), and few samples are low to 6.5
or above to 9.0, thus the pH values of the isolation media
were designed to be as 7.2–7.5.
Pretreatment of soil samples
Soil samples were air-dried at room temperature for
about 2 weeks before isolation. They were subse-
quently ground and passed through a 2 mm-mesh
sieve and then dry-heated at 80�C for 1 h. After
cooling, 2 g soil samples were put into 18 ml mod-
ified Ringer’s solution (g/l; NaCl 7, KCl 0.1, CaCl21.1, NaHCO3 0.2, Na4P2O7 1.0, pH 7.0) and shaken
(about 150 rpm) for 1 h at 28�C. Then 1 ml of each
suspension was added to 9 ml of phenol (1% w/v),
pH 7.0, then shaken (about 100 rpm) for 30 min at
30�C. The samples were finally diluted to 1-in-10 in
0.1% (w/v) sodium pyrophosphate solution (pH 7.0).
Isolation and maintenance of putative
actinobacteria
Seven isolation media were employed in this study
(Table 1). M5 and M6 media preparation followed
the references (Hayakawa and Nonomura 1987;
Hsu and Lockwood 1975) with little modification
(Table 1). Gellan Gum was used as a setting agent
instead of agar in all media. The pH was adjusted
to 7.2–7.5 using 1 N NaOH and/or 1 N HCl. In
addition, betaine (0.125% w/v) and sodium pyruvate
(0.125% w/v) were added to every medium to
facilitate the isolation of strains that are difficult to
culture (Yue et al. 2006). Nystatin, rifampicin, nali-
dixic acid, aztreonam and novobiocin were added to
the media in groups to diminish the growth of fungi
and non-actinobacteria (Table 1). 0.2–0.3 ml pre-
pared soil sample suspension was spread on the
isolation media and then incubated at 28�C for
3–4 weeks. The pure cultures were maintained on
yeast extract-malt extract agar (ISP medium 2;
Shirling and Gottlieb 1966) slants at 4�C and as
20% (v/v) glycerol suspensions at -80�C. Biomass
for molecular systematic and chemotaxonomic stud-
ies was obtained by cultivation in shake flasks (about
214 Antonie van Leeuwenhoek (2010) 98:213–223
123

150 rpm) of ISP 2 medium or Tryptic Soy Broth
(TSB; Difco) at 28�C for 5–7 days.
Morphological observations
ISP 2, ISP 3, ISP 4, ISP 5 (Shirling and Gottlieb
1966), Czapek solution agar (Waksman 1961), nutri-
ent agar (Difco), and tomato paste-oat-meal agar
(Waksman 1961) seeded with the isolates were used
for observation of the colony and cultural character-
istics. Cultures were incubated at 28�C and properties
were recorded after 4, 7, 14, 21, and 28 days. For cell
morphology, the coverslip technique was used on ISP
2 agar, as described by Kawato and Shinobu (1959).
Light microscopy with a model BH-2 microscope
(Olympus) was employed to observe the morpholog-
ical properties. Some gold-coated dehydrated speci-
mens of 7–14 day cultures were examined by
scanning electron microscopy (SEM) (Quanta; FEI).
Metabolic properties
Carbohydrate utilization tests were carried out using
API 50 CH test kits (bioMerieux) and Biolog GEN III
MicroPlates (Biolog Inc.) according to the manufac-
turer’s instructions, and enzyme activities were
determined using API ZYM test kits (bioMerieux),
as recommended by the manufacturer. Growth was
tested at 4, 10, 20, 28–37 (at intervals of 1�C), 40
and 45�C on ISP 2 agar medium incubated for
15–30 days. The ability of the strains to grow at
different NaCl concentrations (0, 1, 3, 5–20%, w/v)
(at intervals of 0.5%) was determined according
to Wang et al. (2001). PH tolerances (5.0–11.0)
(at intervals of 0.5 pH units) were examined with TSB
medium. Other physiological and biochemical tests
were performed according to the established methods of
Williams et al. (1983) and Kampfer et al. (1991).
Chemotaxonomic analyses
Gram stains were performed according to the modified
Hucker method (Hucker 1921) and confirmed by using
3% KOH (Buck 1982). Whole-cell sugar compositions
and diagnostic isomers of diaminopimelic acid were
analysed by TLC as described by Lechevalier and
Lechevalier (1980). Purified peptidoglycan preparations
were obtained by the method of Schleifer and Kandler
(1972). The detailed procedure for isolation of pure
peptidoglycan was summarized as follows. After
collection the cultures from the agar free medium, the
pellet was washed three times with distilled water.
Table 1 The isolation media and their compositions
Medium M1 M2 M3 M4 M5 M6 M7
Composition Starch 5, Histidine 0.1, Amorphous Xylan 5, Humic acid
1.0,
Colloidal chitin
2,
Sodium
propionate 2,
(g l-1) Glycerol 5, Raffinose 1.0, Cellulose 5, Casein 0.3, Na2HPO4 0.5, K2HPO4 0.7 g, Casein
hydrolysate
0.3,
Proline 1, Na2HPO4 0.5, KH2PO4 0.5, KNO3 0.2, KCl 1.7, KH2PO4 0.3, KNO3 0.1.
(NH4)2SO4 1, KCl 1.7, KNO3 0.2, KH2PO4 0.5, MgSO4�7H2O
0.05,
MgSO4�7H2O
0.5,
NaCl 1, MgSO4�4H2O
0.05,
KCl 0.1, KCl 0.1, FeSO4�7H2O
0.01,
FeSO4 0.01 g,
CaCl2 2, FeSO4�7H2O
0.01,
MgSO4�7H2O
0.3,
MgSO4�7H2O
0.3,
CaCl2 1 ZnSO4 0.001,
K2HPO4 1, CaCO3 0.02 NaCl 0.1, NaCl 0.1, MnCl2 0.001
MgSO4�7H2O
1
FeCl3 0.01, FeCl3 0.01,
NaNO3 2.5 NaNO3 2.5
Antibiotics
(mg l-1)
Nystatin 50,
aztreonam
25
Novobiocin 50,
rifampicin 20,
nalidixic acid
50
Nystatin 50,
nalidixic acid
50,
aztreonam 20
Nystatin 50,
nalidixic
acid 50,
Nystatin 50,
aztreonam
50
Novobiocin 50,
rifampicin 20,
nalidixic acid
50
Nystatin 50,
nalidixic acid
50,
aztreonam 20
pH = 7.2–7.5
Antonie van Leeuwenhoek (2010) 98:213–223 215
123

About 3 g cells were used for cell wall preparation.
Firstly, cells were grinded with the help of the liquid
nitrogen, and then suspended in 10 ml solution (con-
taining 1 mg proteinase K and 2 mg SDS). This
suspension was incubated at 55�C on a shaker for about
1 h and was centrifuged, and the pellet was washed two
times with distilled water. Then the sediment was
re-suspended in 10 ml potassium hydrate solution
(containing 0.2 g potassium hydrate solution) and was
incubated at 37�C for 20 h. The suspension was then
centrifuged and the pellet was washed two times with
ethanol 95% and 3 times with distilled water. The pellet
was suspended in 10 ml trypsin–phosphate buffer
(2 mg trypsin in 0.1 M phosphate buffer, pH 7.9) and
was incubated at 37�C on a shaker for about 2 h and was
centrifuged, and the pellet washed two times with
diluted hydrochloric (0.2 mol HCl in 10 ml water) and
three times with distilled water, at last was washed once
with chloroform. The final pellet was dried and the cell
wall was harvested. About 2 mg the cell wall prepara-
tion was hydrolyzed with 4 N HCl at 100�C for 16 h to
determine the quantitative amino acid content, and
another 2 mg cell wall was hydrolyzed with 4 N HCl in
a boiling-water bath for 45–60 min to determine the
peptide compared with those of known peptidoglycan
types.
Amino acids and peptides in cell-wall hydrolysates
were analysed by two-dimensional ascending TLC on
Table 2 16S rRNA gene
information of the 30
isolates studied in detail
Strain number Accession number Genus Similarity with the closest
described strain in GenBank (%)
6014 EU438912 Streptosporangium 96.1
03-9939 EU438907 Actinokineospora 96.8
CPCC202692 FJ529705 Actinokineospora 96.6
06-2230 EU438909 Actinopolymorpha 99.5
CPCC202699 FJ529702 Amycolatopsis 96.9
CPCC202698 FJ529703 Amycolatopsis 97.0
CPCC202695 FJ529717 Agromyces 97.0
06-3147 EU438911 Kineosporia 99.3
CPCC100076 FJ529700 Microlunatus 95.4
6015 EU438903 Saccharopolyspora 99.6
CPCC201357 EU438906 Actinomadura 99.7
CPCC202697 FJ529710 Actinomadura 98.2
6192 FJ529711 Actinomadura 98.9
06-2658 EU438910 Glycomyces 99.2
CPCC202694 FJ529715 Pseudonocardia 98.7
CPCC202696 FJ529716 Pseudonocardia 98.8
CPCC202724 FJ529719 Actinopolyspora 99.1
06-2143 EU531459 Micromonospora 98.9
07-1838 EU531461 Micromonospora 99.6
07-01839 EU531462 Micromonospora 99.8
06-1231-1 EU438908 Micromonospora 99.8
CPCC202691 FJ529699 Nonomuraea 97.5
CPCC202725 FJ529707 Nocardia 98.7
CPCC202726 FJ529708 Nocardia 98.6
CPCC202701 FJ529709 Nocardia 99.3
CPCC202693 FJ529714 Saccharothrix 97.9
6016 EU438904 Dermacoccus 95.9
CPCC201356 EU438905 Geodermatophilus 97.7
CPCC100074 FJ529713 Citricoccus 98.1
CPCC100077 FJ529706 Promicromonospora 97.3
216 Antonie van Leeuwenhoek (2010) 98:213–223
123

cellulose plates using the solvent systems of Schleifer
and Kandler (1972). The N-terminal amino acid of the
interpeptide bridge was determined by dinitropheny-
lation, as described by Schleifer (1985). Molar
ratios of amino acids were determined by GC and
GC-mass spectrometry of N-heptafluorobutyryl amino
acid isobutyl esters (MacKenzie 1987). Analysis of
enantiomers of peptidoglycan amino acids was per-
formed by GC of N-pentafluoropropionyl amino acid
isopropyl esters (Frank et al. 1980) on an L-Chirasil–
Val column (Macherey–Nagel) as described by Groth
et al. (1997). Menaquinones were extracted by using
the method of Collins (1985) and analyzed by HPLC
(Groth et al. 1997). Polar lipids were extracted
according to the method of Minnikin et al. (1984)
and were then identified by separating with two-
dimensional TLC and spraying the chromatogram with
appropriate detection reagents (Minnikin et al. (1984).
The resultant fatty acids were prepared and analyzed
following the instructions of the standard Sherlock
MIDI (Microbial Identification) system (Sasser 1990;
Kampfer and Kroppenstedt 1996).
Molecular analyses
Extraction of genomic DNA and amplification of 23S
rRNA and 16S rRNA genes were done as described by
Xu et al. (2003). The products of amplification of the
23S rRNA gene sequence of some of the isolates,
comprising 380 bp, indicated that these strains were
high-G ? C-DNA Gram-positive bacteria, i.e., actino-
bacteria (Stackebrandt et al. 1997; Yu et al. 2001). The
G ? C content of the genomic DNA was determined
using the thermal denaturation method (Marmur and
Doty 1962) with Streptomyces griseus ATCC 23345T
as a control. Multiple alignments with sequences of
validly described members of related taxa and calcu-
lation of levels of sequence similarity were carried out
using the EzTaxon server (Chun et al. 2007). Phylo-
genetic trees and distance matrices were reconstructed
using the neighbour-joining method (Saitou and Nei
1987) from Knuc values (Kimura 1980, 1983) using
MEGA version 4.0 (Tamura et al. 2007). The topology
of the phylogenetic tree was evaluated by the bootstrap
resampling method of Felsenstein (1985) with 1,000
replicates. Taxon-specific 16S rRNA signature nucle-
otides analysis was carried out using the CLUSTAL W
tool in MEGA version 4.0 (Tamura et al. 2007).
Results
Isolation and primary identification results
A total of 1800 mycelium-forming actinobacterial
strains and 130 non-mycelium forming strains were
purified from the isolation plates. After Gram
staining (confirmed by the KOH method), 97 isolates
were identified as Gram positive bacteria among the
130 non-mycelium forming isolates. Amplification
of 23S rRNA genes of the strains was carried out on
the 97 Gram positive bacteria. As a result, the
amplification of the 23S rRNA from the 97 Gram
positive bacteria showed that 63 strains produced the
expected 380 bp-fragment, which indicated that
these 63 strains were actinobacteria (Stackebrandt
et al. 1997; Yu et al. 2001). From an examination of
the colonies and cells of the Gram positive bacteria,
35 of them looked like Rhodococcus (most colony
color was pink or orange, and amino acids in cell-
wall hydrolysates contained meso-DAP as isomers)
strains and 24 of them looked like Micrococcaceae
(most colony color was pale yellow or orange–
yellow, while amino acids in cell-wall hydroly-
sates did not contain meso-DAP or LL-DAP as
diaminopimelic acid isomers) strains. The remaining
four unclassified bacterial strains, namely 6016,
CPCC 201356, CPCC 10074 and CPCC10077 were
selected to be subjected to a taxonomic study.
Among the 1800 actinomycetes, 817 isolates
formed extensively branched substrate mycelia that
carried abundant aerial hyphae that differentiated into
chains of spores on ISP 2 and ISP 4 plates. Another
634 isolates gave orange or black colonies on ISP 2
and ISP 4 plates. The isomer of diaminopimelic acid
in the cell hydrolysates indicated that the 817 strains
mainly contained LL-diaminopimelic acid and the
634 strains contained meso-diaminopimelic acid.
Combining the data on the isomers of diaminopimelic
acid with the cultural characteristics, the 817 strains
were assigned to the genus Streptomyces and the 634
isolates were assigned to the genus Micromonospora
(Williams et al. 1989). The colonies and cultural
characteristics of the remaining 349 actinomycetes
were further observed on ISP 3, ISP 5, Czapek
solution agar, nutrient agar (Difco), and tomato paste-
oat-meal agar. Twenty-six mycelium-forming strains
were chosen for further 16S rRNA gene sequencing.
16S rRNA gene sequence analysis showed that these
Antonie van Leeuwenhoek (2010) 98:213–223 217
123

Ta
ble
3C
hem
ota
xo
no
mic
pro
per
ties
of
the
30
stu
die
dst
rain
s
Str
ain
nu
mb
erP
epti
do
gly
can
typ
eW
ho
le-c
ell
sug
arp
atte
rnP
ho
sph
oli
pid
sP
red
om
inan
t
men
aqu
ino
nes
Maj
or
fatt
yac
ids
([1
0%
)G
eno
mic
DN
A
G?
Cco
nte
nt
(mo
l.%
)
60
14
mes
o-D
AP
Ara
,G
al,
Rib
DP
G,
PE
,P
IM
K-9
(H6),
MK
-10
C14:0
,is
o-C
15:0
,is
o-C
16:0
,
C16:0
,C
16:1
10
met
hl,
71
.0
03
-99
39
mes
o-D
AP
Gal
,R
ibD
PG
,P
G,
PC
MK
-9(H
4)
iso
-C16:0
,is
o-C
16:1
H6
8.2
CP
CC
20
26
92
mes
o-D
AP
Gal
,R
ibD
PG
,P
G,
PC
MK
-9(H
4)
iso
-C16:0
,C
16:1
H,
C17:1
cis9
67
.1
06
-22
30
LL
-DA
PG
luP
I,P
G,
PIM
MK
-9(H
4)
iso
-C15:
0,
iso
-C16:
0,
iso
-C16:
1H
69
.3
CP
CC
20
26
99
mes
o-D
AP
Ara
,G
alP
E,
PM
E,
PC
MK
-9(H
4)
iso
-C16:0
,is
o-C
15:0
,is
o-C
14:0
,
C16:1
cis9
,C
17:1
cis9
65
.9
CP
CC
20
26
98
mes
o-D
AP
Ara
,G
alP
E,
PM
E,
PC
MK
-9(H
4)
iso
-C16:0
,is
o-C
15:0
,is
o-C
14:0
,
C16:1
cis9
,C
17:1
cis9
66
.2
CP
CC
20
26
95
B2c:
{G
ly}
[L-D
ab]
D-G
lu-D
-Dab
Gal
,M
an,
Rh
aD
PG
,P
GM
K-1
2,
MK
-11
ante
iso
-C17:0
,an
teis
o-C
15:0
,
iso
-C16:0
.
68
.9
06
-31
47
mes
o-D
AP
(mai
nly
);
LL
-DA
P(m
ino
r)
Gal
,G
lu,
Man
,
Rh
a,R
ib
PC
,P
G,
DP
G,
PI
MK
-9(H
4)
C18:1
,C
16:0
69
.8
CP
CC
10
00
76
A3c0
LL
-DA
P—
Gly
Ara
,G
luP
G,
DP
G,
PI
MK
-9(H
4)
ante
iso
-C15:0
,is
o-C
16:0
72
.3
60
15
mes
o-D
AP
Ara
,G
alD
PG
,P
C,
PE
,P
IM
K-9
(H4)
iso
-C15:0
,is
o-C
16:0
,is
o-C
17:0
70
.5
CP
CC
20
13
57
mes
o-D
AP
Ara
,G
al,
Mad
DP
G,
PC
,P
E,
PI
MK
-9(H
6),
K-9
(H8)
iso
-C15:0
,is
o-C
16:0
,is
o-C
17:0
70
.1
CP
CC
20
26
97
mes
o-D
AP
Gal
,A
ra,
Mad
DP
G,
PC
,P
E,
PI
MK
-9(H
6),
K-9
(H8),
iso
-C15:0
,is
o-C
16:0
,is
o-C
17:0
71
.3
61
92
mes
o-D
AP
Gal
,G
lu,
Mad
,M
an,
Rib
DP
G,
PC
,P
E,
PI
MK
-9(H
6),
K-9
(H8),
MK
-9(H
4)
iso
-C16:0
,C
17:0
,C
16:0
,C
17:1x
8c
73
.9
06
-26
58
mes
o-D
AP
Gal
,G
lu,
Rib
,X
yl
DP
G,
PG
,P
IM,
PI
MK
-10
(H4)
C15:0
,is
o-
C16:0
,an
teis
o-C
17:0
,
iso
-C
15:0
72
.2
CP
CC
20
26
94
mes
o-D
AP
Ara
,G
alP
E,
PC
,P
ME
,D
PG
MK
-8(H
4)
iso
-C16:0
,C
16:1
,ci
s9,
iso
-C16:1
H,
C17:1
cis9
70
.4
CP
CC
20
26
96
mes
o-D
AP
Ara
,G
alP
E,
PC
,P
ME
,D
PG
MK
-8(H
4)
iso
-C16:0
,C
17:1
cis9
,is
o-C
15:0
72
.0
CP
CC
20
27
24
mes
o-D
AP
Ara
,G
al,
Rib
PE
,P
C,
PM
E,
DP
GM
K-8
(H4)
iso
-C16:0
,C
17:1
cis9
,is
o-C
15:0
71
.7
06
-21
43
mes
o-D
AP
(mai
nly
);
LL
-DA
P(m
ino
r)
Ara
,G
lu,
Man
,R
ha,
Rib
,X
yl
PC
,PG
,DP
GM
K-9
(H4)
iso
-C16:0
,is
o-C
15:0
,C
17:1x
8c,
ante
iso
-C15:0
66
.1
07
-18
38
mes
o-D
AP
Ara
,G
lu,
Man
,R
ha,
Rib
,X
yl
DP
G,
PE
,P
IM
K-1
0(H
4)
iso
-C16:0
,is
o-C
15:0
,C
17:1x
8c,
ante
iso
-C15:0
72
.3
07
-01
83
9m
eso-D
AP
Ara
,G
lu,
Man
,R
ha,
Rib
,X
yl
DP
G,
PE
,P
IM
K-1
0(H
4)
iso
-C1
6:0
,is
o-C
15:0
,C
17:
1x
8c,
ante
iso
-C15:0
72
.1
06
-12
31
-1m
eso-D
AP
Ara
,G
lu,
Man
,
Rh
a,R
ib,
Xy
l
DP
G,
PE
,P
IM
K-1
0(H
4)
iso
-C16:0
,is
o-C
15:0
,C
17:
1x
8c,
ante
iso
-C15:0
72
.6
218 Antonie van Leeuwenhoek (2010) 98:213–223
123
Ta
ble
3co
nti
nu
ed
Str
ain
nu
mb
erP
epti
do
gly
can
typ
eW
ho
le-c
ell
sug
arp
atte
rnP
ho
sph
oli
pid
sP
red
om
inan
t
men
aqu
ino
nes
Maj
or
fatt
yac
ids
([1
0%
)G
eno
mic
DN
A
G?
Cco
nte
nt
(mo
l.%
)
CP
CC
20
26
91
mes
o-D
AP
Gal
,G
lu,
Mad
,M
anD
PG
,P
I,P
E,
PM
EM
K-9
(H4),
MK
-9(H
2)
iso
-C16:0
,C
16:0
75
.9
CP
CC
20
27
25
mes
o-D
AP
Ara
,G
aLD
PG
,P
E,
PI,
PIM
MK
-8(H
4)
C16:1x
6c,
C16:0
,C
18:1x
9c,
C18:0
10
-met
hy
l
67
.7
CP
CC
20
27
26
mes
o-D
AP
Ara
,G
aLD
PG
,P
E,
PI,
PIM
MK
-8(H
4)
C16:1x
6c,
C16:
0,
C18:1x
9c,
C18:0
10
-met
hy
l
69
.2
CP
CC
20
27
01
mes
o-D
AP
Ara
,G
aLD
PG
,P
E,
PI,
PIM
MK
-8(H
4)
C16:1x
6c,
C16:0
,C
18:1x
9C
,
C18:0
10
-met
hy
l
68
.9
CP
CC
20
26
93
mes
o-D
AP
Gal
,M
anP
Ean
dD
PG
MK
-10
(H4)
iso
-C16:0
,an
teis
o-C
15:0
71
.0
60
16
A4a
L-L
ys–
L-S
er–
D-G
luA
ra,
Gal
,R
ibD
PG
,P
G,
PE
,P
IM
K-8
(H4)
iso
-C15:0
,C
17:0
,C
18:0
,C
18:1
65
.5
CP
CC
20
13
56
mes
o-D
AP
Ara
,G
alD
PG
,P
EM
K-9
(H4),
MK
-9is
o-C
16:
0,
iso
-C1
5:
0,
C17:1
CIS
9c
72
.8
CP
CC
10
00
74
A4a
L-L
ys–
Gly
–D
-Glu
Glu
,M
anD
PG
,P
G,
PI
MK
-9(H
2),
MK
-8(H
2)
ante
iso
-C15:0
,an
teis
o-C
17:0
,is
o-C
15:0
68
.0
CP
CC
10
00
77
A4a
L-L
ys/
D-G
luG
al,
Glu
,R
ha,
Rib
DP
G,
PG
MK
-9(H
4)
iso
-C15:0
,an
teis
o-C
15:0
,is
o-C
16:0
,
ante
iso
-C17:0
72
.0
DA
Pd
iam
ino
pim
elic
acid
,D
PG
dip
ho
sph
atid
yl
gly
cero
l,P
Ep
ho
sph
atid
yl
eth
ano
lam
ine,
PG
ph
osp
hat
idy
lg
lyce
rol,
PI
ph
osp
hat
idy
lin
osi
tol,
PIM
ph
osp
hat
idy
lin
osi
tol
man
no
sid
es,
PM
Ep
ho
sph
atid
yl
met
hy
leth
ano
lam
ine,
PC
ph
osp
hat
idy
lch
oli
ne.
Ara
arab
ino
se,
Ga
lg
alac
tose
,M
an
man
no
se,
Rh
arh
amn
ose
,R
ibri
bo
se,
Glu
glu
cose
,M
ad
mad
uro
se,
Xyl
xy
lose
Antonie van Leeuwenhoek (2010) 98:213–223 219
123

26 strains belonged to 16 different genera (Table 2).
Together with the four non-mycelium forming
actinobacteria, 30 isolates (Table 2) were selected
for detailed taxonomic characterization.
Taxonomic results
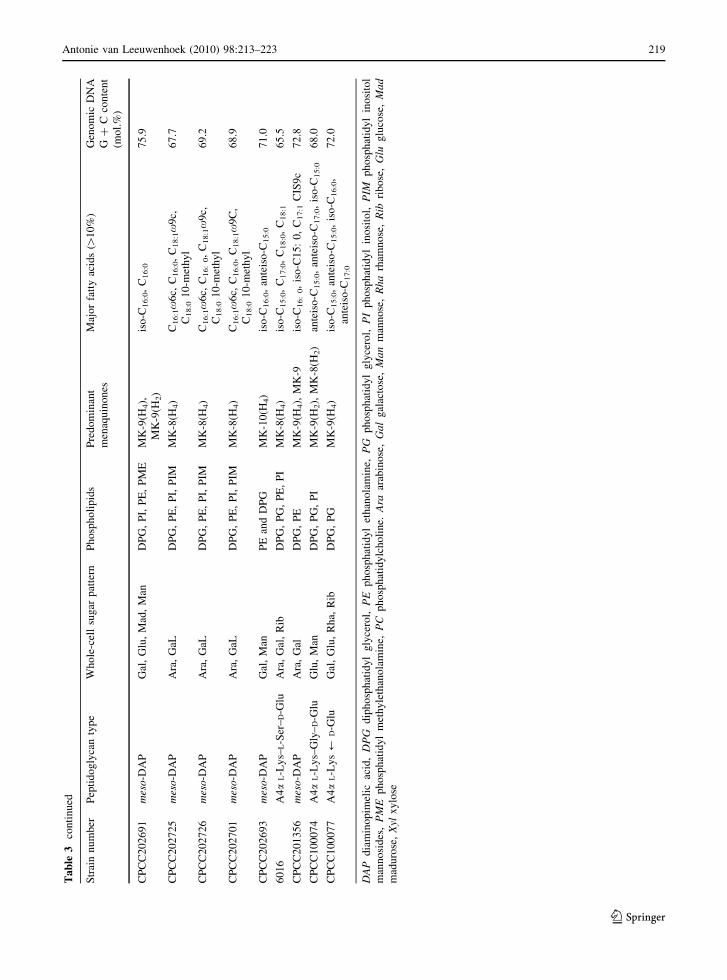
The chemotaxonomic properties, including cell-wall
peptidoglycan diamino acid, predominant menaqui-
nones, genomic DNA G ? C contents, phospholipids
and fatty acid profiles for the selected 30 test strains
are listed in Table 3. Combined with the results of
phylogenetic analyses and physiological tests (data
not shown), ten novel taxa are proposed in this study.
The 16S rRNA signature nucleotide pattern anal-
ysis indicated that strain 6014 possessed the unique
signature nucleotides 1116 (G), 1137 (C), 1436–1465
(C–U) instead of 1116 (G), 1137 (U), 1436–1465
(G–C) of the family Streptosporangiaceae of the sub-
order Streptosporangineae (this study and Stackebrandt
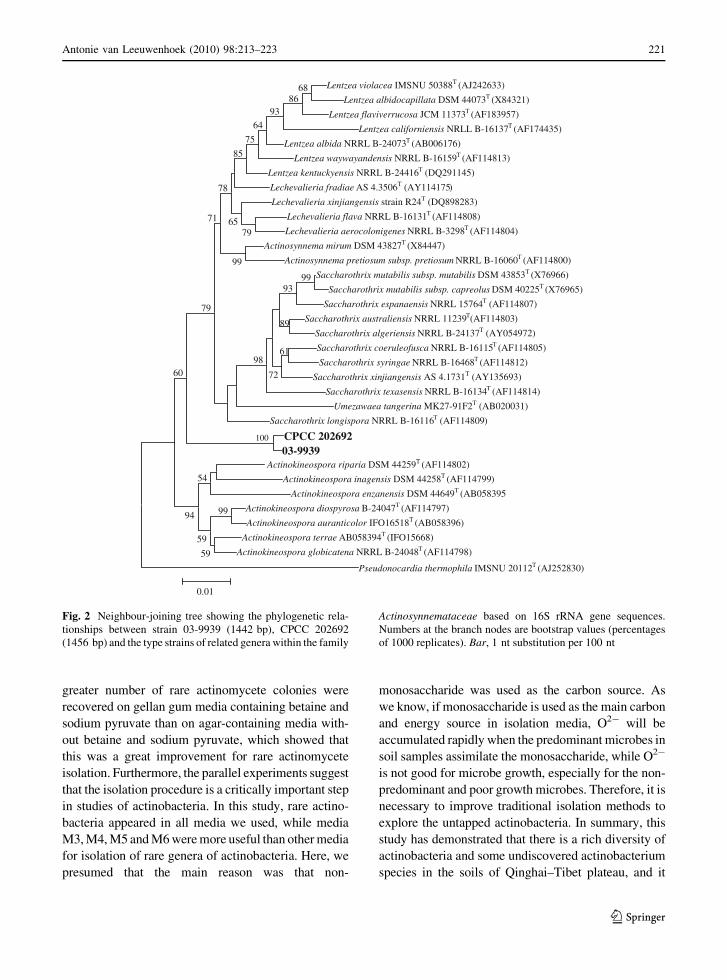
et al. 1997), and strain 03-9939 possessed unique 16S
rRNA signature nucleotides compared to other genera
of the family Actinosynnemataceae, namely, 603–635
(C–G), 617–623 (U–C), 619 (U) (this study and
Labeda and Kroppenstedt 2007). In the phylogenetic
tree, strain 6014 (Fig. 1) and 03-9939 (Fig. 2) formed
a distinct lineage, respectively. Therefore, these two
strains may represent two new genera of the above
two families. The detailed taxonomic study results
are going to be published in another paper. According
to the morphological, physiological and biochemical,
chemotaxonomic and phylogenetic data, strain
06-2230 could be identified as a new member of the
genus Actinopolymorpha (Supplementary Fig. S1);
strain CPCC202699 represents a new species of
the genus Amycolatopsis (Supplementary Fig. S2);
strain CPCC100076 represents a new species of the
genus Microlunatus (Supplementary Fig. S3); strain
CPCC202695 represents a new species of the genus
Agromyces (Supplementary Fig. S4); strain 6016
represents a new species of the genus Demetria
(Supplementary Fig. S5); strain CPCC201356 repre-
sents a new species of the genus Geodermatophilus
(Supplementary Fig. S6); strain CPCC100074 repre-
sents a new species of the genus Citricoccus (Sup-
plementary Fig. S7); strain CPCC100077 represents a
new species of the genus Promicromonospora (Sup-
plementary Fig. S8). It will be necessary to carry out
further experiments (including DNA–DNA hybrid-
ization analysis) to determine whether the remaining
20 strains belong to recognized species.
Conclusion
In this study, gellan gum was used as a setting agent
instead of agar, and betaine and sodium pyruvate were
added to the isolation media. This study showed that a
Planotetraspora mira IFO 15435T (D85496)Acrocarpospora pleiomorpha R-31T ( AB006174) Acrocarpospora pleiomorpha IFO 16266T (AB025318)
Acrocarpospora corrugata DSM 43316T (X89941)
Thermopolyspora flexuosa strain DSM 43186T (AY039253) Planobispora longispora IFO 13918T (D85494)
NonomuraeaPlanomonospora parontospora IFO 13880T (D85495)
Streptosporangium
Streptosporangium violaceochromogenes DSM 43849T (X89951)
6014 Streptomyces megasporus DSM 41476T (Z68100)
100
93
64
95
60
100
7688
64
80
74
79
0.01
Microbispora
HerbisporaMicrotetraspora
Sphaerisporangium
Nonomuraea
Streptosporangium
Fig. 1 Neighbour-joining tree showing the phylogenetic rela-
tionships between strain 6014 (1497 bp) and the type strains of
related genera within the suborder Streptosporangineae based on
16S rRNA gene sequences. Numbers at the branch nodes are
bootstrap values (percentages of 1000 replicates). Bar, 1 nt
substitution per 100 nt
220 Antonie van Leeuwenhoek (2010) 98:213–223
123
greater number of rare actinomycete colonies were
recovered on gellan gum media containing betaine and
sodium pyruvate than on agar-containing media with-
out betaine and sodium pyruvate, which showed that
this was a great improvement for rare actinomycete
isolation. Furthermore, the parallel experiments suggest
that the isolation procedure is a critically important step
in studies of actinobacteria. In this study, rare actino-
bacteria appeared in all media we used, while media
M3, M4, M5 and M6 were more useful than other media
for isolation of rare genera of actinobacteria. Here, we
presumed that the main reason was that non-
monosaccharide was used as the carbon source. As
we know, if monosaccharide is used as the main carbon
and energy source in isolation media, O2- will be
accumulated rapidly when the predominant microbes in
soil samples assimilate the monosaccharide, while O2-
is not good for microbe growth, especially for the non-
predominant and poor growth microbes. Therefore, it is
necessary to improve traditional isolation methods to
explore the untapped actinobacteria. In summary, this
study has demonstrated that there is a rich diversity of
actinobacteria and some undiscovered actinobacterium
species in the soils of Qinghai–Tibet plateau, and it
Lentzea violacea IMSNU 50388T (AJ242633)
Lentzea albidocapillata DSM 44073T (X84321)
Lentzea flaviverrucosa JCM 11373T (AF183957)
Lentzea californiensis NRLL B-16137T (AF174435)
Lentzea albida NRRL B-24073T (AB006176)
Lentzea waywayandensis NRRL B-16159T (AF114813)
Lentzea kentuckyensis NRRL B-24416T (DQ291145)
Lechevalieria fradiae AS 4.3506T (AY114175)
Lechevalieria xinjiangensis strain R24T (DQ898283)
Lechevalieria flava NRRL B-16131T (AF114808)
Lechevalieria aerocolonigenes NRRL B-3298T (AF114804)
Actinosynnema mirum DSM 43827T (X84447)
Actinosynnema pretiosum subsp. pretiosum NRRL B-16060T (AF114800)
Saccharothrix mutabilis subsp. mutabilis DSM 43853T (X76966)
Saccharothrix mutabilis subsp. capreolus DSM 40225T (X76965)
Saccharothrix espanaensis NRRL 15764T (AF114807)
Saccharothrix australiensis NRRL 11239T(AF114803)
Saccharothrix algeriensis NRRL B-24137T (AY054972)
Saccharothrix coeruleofusca NRRL B-16115T (AF114805)
Saccharothrix syringae NRRL B-16468T (AF114812)
Saccharothrix xinjiangensis AS 4.1731T (AY135693)
Saccharothrix texasensis NRRL B-16134T (AF114814)
Umezawaea tangerina MK27-91F2T (AB020031)
Saccharothrix longispora NRRL B-16116T (AF114809)
CPCC 202692 03-9939
Actinokineospora riparia DSM 44259T (AF114802)
Actinokineospora inagensis DSM 44258T (AF114799)
Actinokineospora enzanensis DSM 44649T (AB058395
Actinokineospora diospyrosa B-24047T (AF114797)
Actinokineospora auranticolor IFO16518T (AB058396)
Actinokineospora terrae AB058394T (IFO15668)
Actinokineospora globicatena NRRL B-24048T (AF114798)
Pseudonocardia thermophila IMSNU 20112T (AJ252830)
100
99
99
68 86
93
59
54
59
94
93
89
61
72
98 60
99
64
75
79
85
79
71 65
78
0.01
Fig. 2 Neighbour-joining tree showing the phylogenetic rela-
tionships between strain 03-9939 (1442 bp), CPCC 202692
(1456 bp) and the type strains of related genera within the family
Actinosynnemataceae based on 16S rRNA gene sequences.
Numbers at the branch nodes are bootstrap values (percentages
of 1000 replicates). Bar, 1 nt substitution per 100 nt
Antonie van Leeuwenhoek (2010) 98:213–223 221
123

suggests that these strains might represent a valuable
source of new taxa for further microbial development
and utilization. Additionally, this study indicates that
the biogeographic distribution of actinobacteria in
Qinghai–Tibet plateau is an interesting topic for further
study.
Acknowledgments The authors are grateful to the two
anonymous reviewers and Miss Cao Yan-Ru from Yunnan
University for their critical helping to improve the quality of
this manuscript. This research was supported by National
Facilities and Information Infrastructure for Science and
Technology (2005DKA21203), National Basic Research
Program of China (2010CB833800), the National Natural
Science Foundation of China (nos. 30970008 and 30970038),
Key Projects of International Cooperation (2007DFB31620),
State-level Public Welfare Scientific Research Institutes for
Basic R&D special fund (no. IMBF200915) and Yunnan
Provincial Natural Science Foundation (Nos. 2009AC017 and
2009DA002).
References
Buck JD (1982) Non-staining (KOH) method for determination
of Gram reactions of marine bacteria. Appl Environ
Microbio l44:992–993
Bull AT, Stach JE (2007) Marine actinobacteria: new oppor-
tunities for natural product search and discovery. Trends
Microbiol 15:491–499
Bull AT, Stach JEM, Ward AC, Goodfellow M (2005) Marine
actinobacteria: perspectives, challenges, future directions.
Antonie van Leeuwenhoek 87:259–276
Chun J, Lee JH, Jung Y, Kim M, Kim S, Kim BK, Lim YW
(2007) EzTaxon: a web-based tool for the identification of
prokaryotes based on 16S ribosomal RNA gene sequen-
ces. Int J Syst Evol Microbiol 57:2259–2261
Collins MD (1985) Isoprenoid quinone analysis in classifica-
tion and identification. In: Goodfellow M, Minnikin DE
(eds) Chemical methods in bacterial systematics. Aca-
demic Press, London, pp 267–287
Felsenstein J (1985) Conference limits on phylogenies: an
approach using the bootstrap. Evolution 39:783–789
Frank H, Rettenmeier A, Weicker H, Nicholson GJ, Bayer E
(1980) A new gas chromatographic method for determi-
nation of amino acid levels in human serum. Clin Chim
Acta 105:201–211
Groth I, Schumann P, Rainey FA, Martin K, Schuetze B,
Augsten K (1997) Demetria terragena gen. nov., sp. nov.,
a new genus of actinomycetes isolated from compost soil.
Int J Syst Bacteriol 47:1129–1133
Hayakawa M, Nonomura H (1987) Humic acid-vitamin agar, a
new medium for the selective isolation of soil actinomy-
cetes. J Ferment Technol 65:501–509
Hsu SC, Lockwood JL (1975) Powdered chitin agar as a
selective medium for enumeration of actinomycetes in
water and soil. Appl Microbiol 29:422–426
Hucker GJ (1921) A new modification and application of the
Gram stain. J Bacteriol 6:396–397
Kampfer P, Kroppenstedt RM (1996) Numerical analysis of
fatty acid patterns of coryneform bacteria and related taxa.
Can J Microbiol 42:989–1005
Kampfer P, Kroppenstedt RM, Dott W (1991) A numerical
classification of the genera Streptomyces and Streptover-ticillium using miniaturized physiological tests. J Gen
Microbiol 137:1831–1891
Kawato M, Shinobu R (1959) On Streptomyces herbaricolor sp.
nov., supplement: a single technique for microscopical
observation. Mem Osaka Unit Lib Arts Educ B Nat Sci
8:114–119
Kimura M (1980) A simple method for estimating evolutionary
rates of base substitutions through comparative studies of
nucleotide sequence. J Mol Evol 16:111–120
Kimura M (1983) The neutral theory of molecular evolution.
Cambridge University Press, Cambridge
Labeda DP, Kroppenstedt RM (2007) Proposal of Umezawaeagen. nov., a new genus of the Actinosynnemataceaerelated to Saccharothrix, and transfer of Saccharothrixtangerinus Kinoshita et al. 2000 as Umezawaea tangerinagen. nov. comb. nov. Int J Syst Evol Microbiol 57:2758–
2761
Lechevalier MP, Lechevalier HA (1980) The chemotaxonomy
of actinomycetes. In: Dietz A, Thayer DW (eds) Actino-
mycete taxonomy. SIM special publication no. 6. Society
for Industrial Microbiology, Fairfax, VA, pp 227–291
Li WJ, Schumann P, Zhang YQ, Chen GZ, Tian XP, Xu LH,
Stackebrandt E, Jiang CL (2005a) Marinococcus halo-tolerans sp. nov., isolated from Qinghai, north-west
China. Int J Syst Evol Microbiol 55:1801–1804
Li WJ, Schumann P, Zhang YQ, Xu P, Chen GZ, Xu LH,
Stackebrandt E, Jiang CL (2005b) Proposal of Yaniaceaefam. nov. and Yania flava sp. nov. and emended
description of the genus Yania. Int J Syst Evol Microbiol
55:1933–1938
Li WJ, Zhang YQ, Schumann P, Tian XP, Zhang YQ, Xu LH,
Jiang CL (2006) Sinococcus qinghaiensis gen. nov., sp.
nov., a novel member of the order Bacillales from a saline
soil in China. Int J Syst Evol Microbiol 56:1189–1192
MacKenzie SL (1987) Gas chromatographic analysis of amino
acids as the N-heptafluorobutyryl isobutyl esters. J Assoc
Off Anal Chem 70:151–160
Marmur J, Doty P (1962) Determination of base composition
of deoxyribonucleic acid from its denaturation tempera-
ture. J Mol Biol 5:109–118
Minnikin DE, O’Donnell AG, Goodfellow M, Alderson G,
Athalye M, Schaal A, Parlett JH (1984) An integrated
procedure for the extraction of isoprenoid quinones and
polar-lipids. J Microbiol Methods 2:233–241
Okoro CK, Brown R, Jones AL, Andrews BA, Asenjo JA,
Goodfellow M, Bull AT (2008) Diversity of culturable
actinomycetes in hyper-arid soils of the Atacama Desert,
Chile. Antonie van Leeuwenhoek 95:121–133
Saitou N, Nei M (1987) The neighbor-joining method: a new
method for reconstructing phylogenetic tree. Mol Biol
Evol 4:406–425
Sasser M (1990) Identification of bacteria by gas chromatog-
raphy of cellular fatty acids. Technical note 101. Micro-
bial ID, Newark, DE
222 Antonie van Leeuwenhoek (2010) 98:213–223
123

Schleifer KH (1985) Analysis of the chemical composition and
primary structure of murein. Methods Microbiol 18:123–156
Schleifer KH, Kandler O (1972) Peptidoglycan types of bac-
terial cell walls and their taxonomic implications. Bacte-
riol Rev 36:407–477
Shirling EB, Gottlieb D (1966) Methods for characterization of
Streptomyces species. Int J Syst Bacteriol 16:313–340
Stackebrandt E, Rainey FA, Ward-Rainey NL (1997) Proposal
for a new hierarchic classification system, Actinobacteria
classis nov. Int J Syst Bacteriol 47:479–491
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4:
molecular evolutionary genetics analysis (MEGA) soft-
ware version 4.0. Mol Biol Evol 24:1596–1599
Waksman SA (1961) The actinomycetes, vol 2. The Williams
& Wilkins Co, Baltimore
Wang YM, Zhang ZH, Xu XL, Ruan JS (2001) Actinopoly-morpha singaporensis gen. nov., sp. nov., a novel acti-
nomycete tropical rainforest of Singapore. Int J Syst Evol
Microbiol 51:467–473
Williams ST, Goodfellow M, Alderson G, Wellington EMH,
Sneath PHA, Sackin MJ (1983) Numerical classification
of Streptomyces and related genera. J Gen Microbiol 129:
1743–1813
Williams ST, Goodfellow M, Alderson G (1989) In: Williams
ST, Sharpe ME, Holt JG (eds) Bergey’s manual of
systematic bacteriology, vol 4. Williams & Wilkins,
Baltimore, pp 2452–2492
Xu P, Li WJ, Xu LH, Jiang CL (2003) A microwave-based
method for genomic DNA extraction from actinomycetes.
Microbiology 30:73–75 (in Chinese)
Yu LY, Li QP, Yao TJ (2001) Comparison of two methods for
identifying Actinobacteria. Chin J Antibiot 26:10–14
Yue XJ, Yu LY, Li QP, Wei YZ, Guan Y, Zhang YQ (2006)
Study of methods to isolate viable but non-culturable
microorganisms from natural environment. Microbiology
33:77–81 (in Chinese)
Zhang YQ, Schumann P, Li WJ, Chen GZ, Tian XP, Stacke-
brandt E, Xu LH, Jiang CL (2005) Isoptericola halotol-erans sp. nov., a novel actinobacterium isolated from
saline soil from Qinghai Province, north-west China. Int J
Syst Evol Microbiol 55:1867–1870
Zhang YQ, Schumann P, Yu LY, Liu HY, Zhang YQ, Xu LH,
Stackebrandt E, Jiang CL, Li WJ (2007) Zhihengliuellahalotolerans gen. nov., sp. nov., a novel member of the
family Micrococcaceae. Int J Syst Evol Microbiol 57:
1018–1023
Zhang YQ, Yu LY, Wang D, Liu HY, Sun CH, Jiang W, Zhang
YQ, Li WJ (2008) Roseomonas vinacea sp. nov., a Gram-
negative coccobacillus isolated from a soil sample. Int J
Syst Evol Microbiol 58:2070–2074
Antonie van Leeuwenhoek (2010) 98:213–223 223
123




















