
Diversity, Epidemiology, and Genetics of Class D -Lactamases
15
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 2010, p. 24–38 Vol. 54, No. 1 0066-4804/10/$12.00 doi:10.1128/AAC.01512-08 Copyright © 2010, American Society for Microbiology. All Rights Reserved. MINIREVIEW Diversity, Epidemiology, and Genetics of Class D -Lactamases Laurent Poirel, Thierry Naas, and Patrice Nordmann* Service de Bacte ´riologie-Virologie, INSERM U914 Emerging Resistance to Antibiotics, Ho ˆpital de Bice ˆtre, Faculte ´ de Me ´decine et Universite ´ Paris-Sud, 94275 Le Kremlin-Bice ˆtre, France Class D -lactamase-mediated resistance to -lactams has been increasingly reported during the last decade. Those enzymes also known as oxacillinases or OXAs are widely distributed among Gram negatives. Genes encoding class D -lactamases are known to be intrinsic in many Gram-negative rods, including Acinetobacter baumannii and Pseudomonas aeruginosa, but play a minor role in natural resistance phenotypes. The OXAs (ca. 150 variants reported so far) are characterized by an important genetic diversity and a great heterogeneity in terms of -lactam hydrolysis spectrum. The acquired OXAs possess either a narrow spectrum or an expanded spectrum of hydrolysis, including carbapenems in several instances. Acquired class D -lac- tamase genes are mostly associated to class 1 integron or to insertion sequences. Class D -lactamases, also known as oxacillinases or OXA- type -lactamases (OXAs), are active-serine-site enzymes like Ambler class A and class C -lactamases, differing from class A and C enzymes in amino acid structure, whereas class B -lac- tamases are metalloenzymes with a Zn 2 ion(s) in the active site (4, 71, 78). Even though class D includes mostly enzymes with higher hydrolysis rates for cloxacillin and oxacillin than for benzylpenicillin (hence the name oxacillinases), not all class D -lactamases have this characteristic. Most of the class D enzymes belong to group 2d of the Bush functional classifica- tion scheme for -lactamases (23). Among the four -lacta- mase molecular classes, class D -lactamases are the most diverse enzymes (107). This diversity is observed at both the genetic and biochemical levels, with enzymes possessing either a narrow or expanded spectrum of hydrolysis. In addition, several class D -lactamases have an expanded spectrum of activity resulting from point mutations. Although many class D -lactamase genes are embedded into class 1 integrons, recent reports indicated that other spe- cific genetic structures, including insertion sequences and transposons, may be associated with class D -lactamase genes. Numerous class D -lactamase genes have been identified as a source of acquired resistance in gram-negative bacteria, but recent studies have shown that class D -lactamases are also naturally produced in clinically significant pathogens and en- vironmental species (107). This review focuses on the diversity and substrate profiles of class D -lactamases, their sources, and the genetics of acqui- sition of the corresponding genes. All the class D -lactamases for which a sequence is available in the GenBank databases are listed in Table 1. GENERAL PROPERTIES OXAs are -lactamases belonging to class D according to the molecular nomenclature based on amino acid identity com- parison. Similar to class A and C -lactamases, they possess an active-site serine which is located at position 70 in the class D -lactamase numbering system (the DBL numbering system) (72, 97) (Fig. 1). The DBL numbering system has been estab- lished to analyze the molecular structures of the class D -lac- tamases, in analogy to the numbering system adopted for class A enzymes (5, 32). The two groups of enzymes share the serine and lysine residues which are part of the highly conserved motif S-T-F-K found at positions 70 to 73 (79). The Y-G-N motif (positions 144 to 146) and the K-T-G motif (positions 216 to 218) are highly conserved in class D -lactamases, but the F-G-N motif may replace Y-G-N in several OXAs (Fig. 1). Class D -lactamases are usually not inhibited by clavulanic acid, tazobactam, and sulbactam, whereas their activities may be inhibited in vitro by sodium chloride (NaCl). This property is not shared by -lactamases of other classes, thus defining it as a useful characteristic for in vitro identification. NaCl at a concentration of 100 mM inhibits totally the activities of most class D -lactamases (9, 54, 65, 127, 130). This property is not clearly explained, but it has been related at least to the pres- ence of a Tyr residue at position 144. In vitro mutagenesis showed that the replacement of a Tyr residue by a Phe at that position makes a mutant resistant to NaCl inhibition (65). Class D -lactamases were first defined as -lactamases hy- drolyzing cloxacillin and oxacillin faster than benzylpenicillin (23). This definition does not seem valuable anymore since recently described enzymes actually inactivate cloxacillin and oxacillin poorly, even sometimes sparing those substrates. However, all class D -lactamases significantly hydrolyze amino- and carboxypenicillins. Although all the OXAs first characterized corresponded to acquired genes, it has been demonstrated lately that many gram-negative species naturally possess genes in their genomes coding for OXAs, which are therefore considered to be resi- dent enzymes. * Corresponding author. Mailing address: Service de Bacte ´riologie- Virologie, Ho ˆpital de Bice ˆtre, 78 rue du Ge ´ne ´ral Leclerc, 94275 Le Kremlin-Bice ˆtre Cedex, France. Phone: 33-1-45-21-36-32. Fax: 33-1- 45-21-63-40. E-mail: [email protected]. Published ahead of print on 31 August 2009. 24
Transcript of Diversity, Epidemiology, and Genetics of Class D -Lactamases

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 2010, p. 24–38 Vol. 54, No. 10066-4804/10/$12.00 doi:10.1128/AAC.01512-08Copyright © 2010, American Society for Microbiology. All Rights Reserved.
MINIREVIEW
Diversity, Epidemiology, and Genetics of Class D �-Lactamases�
Laurent Poirel, Thierry Naas, and Patrice Nordmann*Service de Bacteriologie-Virologie, INSERM U914 Emerging Resistance to Antibiotics, Hopital de Bicetre, Faculte de
Medecine et Universite Paris-Sud, 94275 Le Kremlin-Bicetre, France
Class D �-lactamase-mediated resistance to �-lactams has been increasingly reported during the lastdecade. Those enzymes also known as oxacillinases or OXAs are widely distributed among Gram negatives.Genes encoding class D �-lactamases are known to be intrinsic in many Gram-negative rods, includingAcinetobacter baumannii and Pseudomonas aeruginosa, but play a minor role in natural resistance phenotypes.The OXAs (ca. 150 variants reported so far) are characterized by an important genetic diversity and a greatheterogeneity in terms of �-lactam hydrolysis spectrum. The acquired OXAs possess either a narrow spectrumor an expanded spectrum of hydrolysis, including carbapenems in several instances. Acquired class D �-lac-tamase genes are mostly associated to class 1 integron or to insertion sequences.
Class D �-lactamases, also known as oxacillinases or OXA-type �-lactamases (OXAs), are active-serine-site enzymes likeAmbler class A and class C �-lactamases, differing from class Aand C enzymes in amino acid structure, whereas class B �-lac-tamases are metalloenzymes with a Zn2� ion(s) in the activesite (4, 71, 78). Even though class D includes mostly enzymeswith higher hydrolysis rates for cloxacillin and oxacillin thanfor benzylpenicillin (hence the name oxacillinases), not all classD �-lactamases have this characteristic. Most of the class Denzymes belong to group 2d of the Bush functional classifica-tion scheme for �-lactamases (23). Among the four �-lacta-mase molecular classes, class D �-lactamases are the mostdiverse enzymes (107). This diversity is observed at both thegenetic and biochemical levels, with enzymes possessing eithera narrow or expanded spectrum of hydrolysis. In addition, severalclass D �-lactamases have an expanded spectrum of activityresulting from point mutations.
Although many class D �-lactamase genes are embeddedinto class 1 integrons, recent reports indicated that other spe-cific genetic structures, including insertion sequences andtransposons, may be associated with class D �-lactamase genes.Numerous class D �-lactamase genes have been identified asa source of acquired resistance in gram-negative bacteria, butrecent studies have shown that class D �-lactamases are alsonaturally produced in clinically significant pathogens and en-vironmental species (107).
This review focuses on the diversity and substrate profiles ofclass D �-lactamases, their sources, and the genetics of acqui-sition of the corresponding genes. All the class D �-lactamasesfor which a sequence is available in the GenBank databases arelisted in Table 1.
GENERAL PROPERTIES
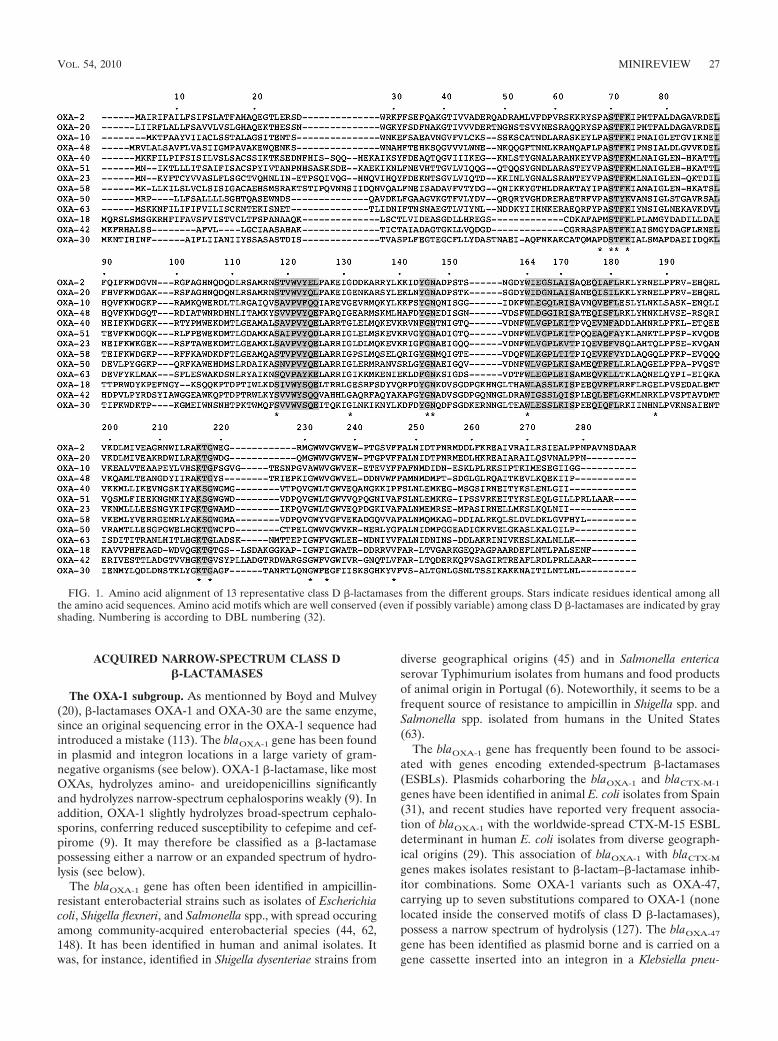
OXAs are �-lactamases belonging to class D according tothe molecular nomenclature based on amino acid identity com-parison. Similar to class A and C �-lactamases, they possess anactive-site serine which is located at position 70 in the class D�-lactamase numbering system (the DBL numbering system)(72, 97) (Fig. 1). The DBL numbering system has been estab-lished to analyze the molecular structures of the class D �-lac-tamases, in analogy to the numbering system adopted for classA enzymes (5, 32). The two groups of enzymes share the serineand lysine residues which are part of the highly conservedmotif S-T-F-K found at positions 70 to 73 (79). The Y-G-Nmotif (positions 144 to 146) and the K-T-G motif (positions216 to 218) are highly conserved in class D �-lactamases, butthe F-G-N motif may replace Y-G-N in several OXAs (Fig. 1).
Class D �-lactamases are usually not inhibited by clavulanicacid, tazobactam, and sulbactam, whereas their activities maybe inhibited in vitro by sodium chloride (NaCl). This propertyis not shared by �-lactamases of other classes, thus defining itas a useful characteristic for in vitro identification. NaCl at aconcentration of 100 mM inhibits totally the activities of mostclass D �-lactamases (9, 54, 65, 127, 130). This property is notclearly explained, but it has been related at least to the pres-ence of a Tyr residue at position 144. In vitro mutagenesisshowed that the replacement of a Tyr residue by a Phe at thatposition makes a mutant resistant to NaCl inhibition (65).
Class D �-lactamases were first defined as �-lactamases hy-drolyzing cloxacillin and oxacillin faster than benzylpenicillin(23). This definition does not seem valuable anymore sincerecently described enzymes actually inactivate cloxacillin andoxacillin poorly, even sometimes sparing those substrates.However, all class D �-lactamases significantly hydrolyze amino-and carboxypenicillins.
Although all the OXAs first characterized corresponded toacquired genes, it has been demonstrated lately that manygram-negative species naturally possess genes in their genomescoding for OXAs, which are therefore considered to be resi-dent enzymes.
* Corresponding author. Mailing address: Service de Bacteriologie-Virologie, Hopital de Bicetre, 78 rue du General Leclerc, 94275 LeKremlin-Bicetre Cedex, France. Phone: 33-1-45-21-36-32. Fax: 33-1-45-21-63-40. E-mail: [email protected].
� Published ahead of print on 31 August 2009.
24

TABLE 1. Features of oxacillinases
Namea Alternatename
OXAgroup Type Original host A or
Nb
Associated mobile elementGene GCcontent
(%)
Isoelectric pointd
GenBankaccession
no.eReferencefTransposon or
insertionsequence
Integronc Exptl Theoretical
OXA-1 OXA-30 Narrow spectrum E. coli A Tn2603 � 34.4 7.4 7.7 J02967 113OXA-2 OXA-2 Narrow spectrum S. Typhimurium A � 50 7.7 9.1 X07260 97OXA-3 OXA-2 Narrow spectrum K. pneumoniae A Tn1411 � 50 7.1 8.1 L07945 142OXA-4 OXA-35 Narrow spectrum E. coli A Tn1409 � 7.5 AY162283 142OXA-5 Narrow spectrum P. aeruginosa A Tn1406 � 40.2 7.6 8.4 X58272 32OXA-6 Narrow spectrum P. aeruginosa A 7.7 UPOXA-7 OXA-10 Narrow spectrum E. coli A � 40.6 7.7 9.3 X75562 145OXA-8OXA-9 Narrow spectrum K. pneumoniae A Tn1331 � 49.5 6.9 7.1 M55547 155LCR-1 Narrow spectrum P. aeruginosa A Tn1412 � 52 6.5 7.1 X56809 32OXA-10 Narrow spectrum P. aeruginosa A Tn1404 � 42.1 6.1 7.0 U37105 70OXA-11 OXA-10 ES-OXA P. aeruginosa A � 42 6.4 6.3 Z22590 60OXA-12 Narrow spectrum A. jandaei N � 62.3 8.6 8.4 U10251 139AmpS Narrow spectrum A. hydrophila N � 63 7.9 7.1 X80276 165OXA-13 OXA-10 Narrow spectrum P. aeruginosa A � 41.2 8.0 8.7 U59183 99OXA-14 OXA-10 ES-OXA P. aeruginosa A � 42.1 6.2 6.3 L38523 38OXA-15 OXA-2 ES-OXA P. aeruginosa A � 50 8.7 9.3 U63835 36OXA-16 OXA-10 ES-OXA P. aeruginosa A � 42.1 6.2 6.3 AF043100 39OXA-17 OXA-10 ES-OXA P. aeruginosa A � 42.1 6.1 7.0 AF060206 37OXA-18 ES-OXA P. aeruginosa A ISCR19 � 61.2 5.5 5.9 U85514 118OXA-19 OXA-13 ES-OXA P. aeruginosa A � 41.2 7.6 8.4 AF043381 98OXA-20 Narrow spectrum P. aeruginosa A � 45.1 7.4 9.0 AF024602 108OXA-21 OXA-3 Narrow spectrum A. baumannii A � 50.1 7.0 8.1 Y10693 162OXA-22 Narrow spectrum R. pickettii N � 65.5 7.0 6.4 AF064820 112OXA-23 CHDL A. baumannii A Tn2006/Tn2007 � 38 6.7 7.0 AJ132105 42OXA-24 OXA-40 CHDL A. baumannii A � 34.4 8.6 9.0 AJ239129 19OXA-25 OXA-40 CHDL A. baumannii A � 34.4 8.0 8.5 AF201826 1OXA-26 OXA-40 CHDL A. baumannii A � 34.4 7.9 9.0 AF201827 1OXA-27 OXA-23 CHDL A. baumannii A � 38 6.8 8.0 AF201828 1OXA-28 OXA-13 ES-OXA P. aeruginosa A � 41.2 8.1 8.7 AF231133 125OXA-29 Narrow spectrum L. gormanii N � 36.6 9.0 9.4 AJ400619 49OXA-30 OXA-1 Narrow spectrum E. coli A � 34.4 7.3 6.8 AF255921 148OXA-31 OXA-1 ES-OXA P. aeruginosa A � 34.4 7.5 6.8 AF294653 9OXA-32 OXA-2 ES-OXA P. aeruginosa A � 50 7.7 9.0 AF315351 124OXA-33 OXA-40 CHDL A. baumannii A � 34.4 8.6 9.0 AY008291OXA-34 OXA-2 ES-OXA P. aeruginosa A � 50 8.9 AF350424 UPOXA-35 OXA-10 ES-OXA P. aeruginosa A � 41.2 8.0 8.7 AF315786 8OXA-36 OXA-2 ES-OXA P. aeruginosa A � 49.4 9.2 AF300985 UPOXA-37 OXA-20 Narrow spectrum A. baumannii A � 44.8 7.4 8.9 AY007784 109OXA-38OXA-39OXA-40 CHDL A. baumannii A � 34.4 8.6 9.0 AF509241 65OXA-41OXA-42 Narrow spectrum B. pseudomallei N � 66.3 9.2 9.3 AJ488302 111OXA-43 Narrow spectrum B. pseudomallei N � 65.9 9.2 9.3 AJ488303 111OXA-44OXA-45 ES-OXA P. aeruginosa A ISCR5 � 61.8 8.8 9.4 AJ519683 153OXA-46 Narrow spectrum P. aeruginosa A � 47.1 7.8 8.7 AF317511 57OXA-47 OXA-1 Narrow spectrum K. pneumoniae A � 34.1 7.4 6.8 AY237830 127OXA-48 CHDL K. pneumoniae A Tn1999 � 44.5 7.2 8.0 AY236073 127OXA-49 OXA-23 CHDL A. baumannii A � 38 6.0 AY288523 UPOXA-50 Narrow spectrum P. aeruginosa N � 64.8 8.6 9.0 AY306130 54OXA-51 OXA-Ab1 CHDL A. baumannii A � 39.3 7.0 8.0 AJ309734 22OXA-52OXA-53 OXA-2 ES-OXA S. Agona A � 50.2 6.9 7.2 AY289608 103OXA-54 CHDL S. oneidensis N � 46.6 6.8 6.7 AY500137 126OXA-55 CHDL S. algae N � 53.8 8.6 8.6 AY343493 68OXA-56 Narrow spectrum P. aeruginosa A � 40.7 6.5 8.7 AY445080 25OXA-57 Narrow spectrum B. pseudomallei N � 66 9.3 AJ631966 73OXA-58 CHDL A. baumannii A � 37.4 7.2 7.2 AY665723 130OXA-59 Narrow spectrum B. pseudomallei N � 65.9 9.3 AJ632249 73OXA-60 Narrow spectrum R. pickettii N � 64.9 5.1 5.4 AF525303 55OXA-61 Narrow spectrum C. jejuni N � 27.4 9.1 AY587956 2OXA-62 CHDL P. pnomenusa N � 65.3 �9.0 9.5 AY423074 144OXA-63 Narrow spectrum B. pilosicoli N � 24.9 6.0 AY619003 94OXA-64 OXA-Ab2 OXA-51 CHDL A. baumannii N � 39.6 8.0 AY750907 21OXA-65 OXA-Ab3 OXA-51 CHDL A. baumannii N � 39.2 8.8 AY750908 21OXA-66 OXA-Ab4 OXA-51 CHDL A. baumannii N � 39.4 9.0 AY750909 21OXA-67 OXA-Ab5 OXA-51 CHDL A. baumannii N � 39 8.0 DQ491200 UPOXA-68 OXA-Ab6 OXA-51 CHDL A. baumannii N � 39 7.1 AY750910 21OXA-69 OXA-Ab7 OXA-51 CHDL A. baumannii N � 39.3 8.4 8.6 AY750911 66OXA-70 OXA-Ab8 OXA-51 CHDL A. baumannii N � 39.3 9.0 AY750912 21OXA-71 OXA-Ab9 OXA-51 CHDL A. baumannii N � 39.7 8.0 AY750913 21OXA-72 OXA-40 CHDL A. baumannii A � 36.4 8.8 EF534256 166OXA-73 OXA-23 CHDL K. pneumoniae A � 37.6 8.0 AY762325 UPOXA-74 OXA-10 Unknown P. aeruginosa A � 41.9 6.5 7.0 AJ854182 46OXA-75 OXA-Ab10 OXA-51 CHDL A. baumannii N � 38.7 8.6 AY859529 66OXA-76 OXA-Ab11 OXA-51 CHDL A. baumannii N � 39.3 9.2 AY949203 66OXA-77 OXA-Ab12 OXA-51 CHDL A. baumannii N � 39.2 8.6 AY949202 66OXA-78 OXA-Ab13 OXA-51 CHDL A. baumannii N � 39.2 8.9 AY862132 UP
Continued on following page
VOL. 54, 2010 MINIREVIEW 25

TABLE 1—Continued
Namea Alternatename
OXAgroup Type Original host A or
Nb
Associated mobile elementGene GCcontent
(%)
Isoelectric pointd
GenBankaccession
no.ReferenceeTransposon or
insertionsequence
Integronc Exptl Theoretical
OXA-79 OXA-Ab14 OXA-51 CHDL A. baumannii N � 39.5 9.0 EU019534 47OXA-80 OXA-Ab15 OXA-51 CHDL A. baumannii N � 39.3 9.0 EU019535 47OXA-81OXA-82 OXA-Ab16 OXA-51 CHDL A. baumannii N � 39.4 9.0 EU019536 158OXA-83 OXA-Ab17 OXA-51 CHDL A. baumannii N � 39.5 9.0 DQ309277 158OXA-84 OXA-Ab18 OXA-51 CHDL A. baumannii N � 39.4 9.0 DQ309276 158OXA-85 Narrow spectrum F. nucleatum N � 24.6 5.3 6.1 AY227054 164OXA-86 OXA-Ab19 OXA-51 CHDL A. baumannii N � 38.8 8.0 DQ149247 159OXA-87 OXA-Ab20 OXA-51 CHDL A. baumannii N � 38.9 8.0 DQ348075 159OXA-88 OXA-Ab21 OXA-51 CHDL A. baumannii N � 39.2 9.2 DQ392963 75OXA-89 OXA-Ab22 OXA-51 CHDL A. baumannii N � 38.4 7.0 8.6 DQ445683 94OXA-90 OXA-Ab23 OXA-51 CHDL A. baumannii N � 39.2 8.6 EU433382 UPOXA-91 OXA-Ab24 OXA-51 CHDL A. baumannii N � 39 8.0 DQ519083 75OXA-92 OXA-Ab25 OXA-51 CHDL A. baumannii N � 39.3 8.6 DQ335566 156OXA-93 OXA-Ab26 OXA-51 CHDL A. baumannii N � 39.3 8.0 DQ519087 75OXA-94 OXA-Ab27 OXA-51 CHDL A. baumannii N � 39.3 8.9 DQ519088 75OXA-95 OXA-Ab28 OXA-51 CHDL A. baumannii N � 39.5 8.6 DQ519089 75OXA-96 OXA-58 CHDL A. baumannii A � 37.5 7.2 DQ519090 75OXA-97 OXA-58 CHDL A. baumannii A � 37.8 7.2 EF102240 129OXA-98 OXA-Ab29 OXA-51 CHDL A. baumannii N � 39.2 8.6 AM279652 UPOXA-99 OXA-Ab30 OXA-51 CHDL A. baumannii N � 39.4 8.0 DQ888718 UPOXA-100OXA-101 OXA-10 Unknown C. freundii A � 40.7 8.8 AM412777 UPOXA-102 OXA-23 CHDL A. radioresistens N � 38 5.8 Unknown 123OXA-103 OXA-23 CHDL A. radioresistens N � 38 5.8 Unknown 123OXA-104 OXA-Ab31 OXA-51 CHDL A. baumannii N � 39.3 8.6 EF581285 47OXA-105 OXA-23 CHDL A. radioresistens N � 38 7.0 Unknown UPOXA-106 OXA-Ab32 OXA-51 CHDL A. baumannii N � 39.3 8.9 EF650032 47OXA-107 OXA-Ab33 OXA-51 CHDL A. baumannii N � 39.3 8.6 EF650033 47OXA-108 OXA-Ab34 OXA-51 CHDL A. baumannii N � 39 8.5 EF650034 47OXA-109 OXA-Ab35 OXA-51 CHDL A. baumannii N � 39.3 9.0 EF650035 47OXA-110 OXA-Ab36 OXA-51 CHDL A. baumannii N � 39.3 8.6 EF650036 47OXA-111 OXA-Ab37 OXA-51 CHDL A. baumannii N � 39.4 7.1 EF650037 47OXA-112 OXA-Ab38 OXA-51 CHDL A. baumannii N � 39.4 8.6 EF650038 47OXA-113 OXA-Ab39 OXA-51 CHDL A. baumannii N � 39.3 8.0 EF653400 106OXA-114 Narrow spectrum A. xylosoxidans N � 70.4 8.6 9.0 EU188842 41OXA-115 OXA-Ab40 OXA-51 CHDL A. baumannii N � 39.3 9.0 EU029998 UPOXA-116 OXA-Ab41 OXA-51 A. baumannii N � 39.3 8.6 EU220744 UPOXA-117 OXA-Ab42 OXA-51 A. baumannii N � 39.2 8.6 EU220745 UPOXA-118 Narrow spectrum B. cepacia A � 49.3 7.3 AF371964 33OXA-119 Narrow spectrum Uncultured
bacteriumA � 49.4 6.7 AY139598 150
OXA-120OXA-121OXA-122OXA-123OXA-124OXA-125OXA-126OXA-127OXA-128 OXA-10 CHDL A. baumannii N � 39.1 8.0 EU375515 52aOXA-129 OXA-Ab43 OXA-5 Unknown S. Bredeney A � 39.9 9.1 AM932669 95OXA-130 OXA-Ab44 OXA-51 A. baumannii N � 39.1 8.5 EU547445 UPOXA-131 OXA-Ab45 OXA-51 A. baumannii N � 39.4 9.0 EU547446 UPOXA-132 OXA-Ab46 OXA-51 A. baumannii N � 39.3 8.0 EU547447 UPOXA-133 OXA-23 CHDL A. radioresistens N � 39.3 6.1 EU571228 123OXA-134 CHDL A. lwoffii N � 46.2 5.3 UPOXA-135OXA-136 OXA-63 Narrow spectrum B. pilosicoli N � 25.1 5.3 EU086830 96OXA-137 OXA-63 Narrow spectrum B. pilosicoli N � 24.9 5.7 EU086834 96OXA-138OXA-139OXA-140OXA-141 OXA-2 ES-OXA P. aeruginosa A � 49.9 9.1 EF552405 UPOXA-142 OXA-10 ES-OXA P. aeruginosa A � 42 6.3 EU358785 UPOXA-143 CHDL A. baumannii A � 34.4 8.7 UPOXA-144OXA-145 OXA-10 ES-OXA P. aeruginosa A � 41.1 8.7 FJ790516 UPOXA-146OXA-147 OXA-10 ES-OXA P. aeruginosa A 41 8.1 FJ848783 UP
a The nomenclature is in accordance with that provided by G. Jacoby on the Lahey website (http://www.lahey.org/Studies/other.asp#table1). Lacking variants(in boldface) are those for which a number has been assigned on this website but for which no information is yet available.
b A, acquired; N, natural.c �, the oxacillinase gene was found to be associated with an integron-borne gene cassette; �, the gene is not associated with an integron-borne gene cassette.d Experimentally obtained pI values (when available) versus calculated values. Theoretical values were calculated using software found at the ExPASy proteomics
tools website (http://www.expasy.ch/tools/) and the amino acid sequences of the mature proteins only. Peptide cleavage site identification was performed with SignalP(http://www.cbs.dtu.dk/services/SignalP/), and pI computing was performed with the Compute pI/Mw tool (http://www.expasy.ch/tools/pi_tool.html).
e UP, unpublished.
26 MINIREVIEW ANTIMICROB. AGENTS CHEMOTHER.
ACQUIRED NARROW-SPECTRUM CLASS D�-LACTAMASES
The OXA-1 subgroup. As mentionned by Boyd and Mulvey(20), �-lactamases OXA-1 and OXA-30 are the same enzyme,since an original sequencing error in the OXA-1 sequence hadintroduced a mistake (113). The blaOXA-1 gene has been foundin plasmid and integron locations in a large variety of gram-negative organisms (see below). OXA-1 �-lactamase, like mostOXAs, hydrolyzes amino- and ureidopenicillins significantlyand hydrolyzes narrow-spectrum cephalosporins weakly (9). Inaddition, OXA-1 slightly hydrolyzes broad-spectrum cephalo-sporins, conferring reduced susceptibility to cefepime and cef-pirome (9). It may therefore be classified as a �-lactamasepossessing either a narrow or an expanded spectrum of hydro-lysis (see below).
The blaOXA-1 gene has often been identified in ampicillin-resistant enterobacterial strains such as isolates of Escherichiacoli, Shigella flexneri, and Salmonella spp., with spread occuringamong community-acquired enterobacterial species (44, 62,148). It has been identified in human and animal isolates. Itwas, for instance, identified in Shigella dysenteriae strains from
diverse geographical origins (45) and in Salmonella entericaserovar Typhimurium isolates from humans and food productsof animal origin in Portugal (6). Noteworthily, it seems to be afrequent source of resistance to ampicillin in Shigella spp. andSalmonella spp. isolated from humans in the United States(63).
The blaOXA-1 gene has frequently been found to be associ-ated with genes encoding extended-spectrum �-lactamases(ESBLs). Plasmids coharboring the blaOXA-1 and blaCTX-M-1
genes have been identified in animal E. coli isolates from Spain(31), and recent studies have reported very frequent associa-tion of blaOXA-1 with the worldwide-spread CTX-M-15 ESBLdeterminant in human E. coli isolates from diverse geograph-ical origins (29). This association of blaOXA-1 with blaCTX-M
genes makes isolates resistant to �-lactam–�-lactamase inhib-itor combinations. Some OXA-1 variants such as OXA-47,carrying up to seven substitutions compared to OXA-1 (nonelocated inside the conserved motifs of class D �-lactamases),possess a narrow spectrum of hydrolysis (127). The blaOXA-47
gene has been identified as plasmid borne and is carried on agene cassette inserted into an integron in a Klebsiella pneu-
FIG. 1. Amino acid alignment of 13 representative class D �-lactamases from the different groups. Stars indicate residues identical among allthe amino acid sequences. Amino acid motifs which are well conserved (even if possibly variable) among class D �-lactamases are indicated by grayshading. Numbering is according to DBL numbering (32).
VOL. 54, 2010 MINIREVIEW 27

moniae isolate from Turkey coproducing OXA-48 (123) (seebelow).
Among the OXA-1 derivatives that have been identified,no known variant possesses the ability to hydrolyze ceftazi-dime. OXA-1 possesses the ability to hydrolyze cefepime andcefpirome slightly, and OXA-31, which differs from OXA-1 bytwo amino acid substitutions and has been identified in aPseudomonas aeruginosa isolate from France (9), also pos-sesses that property. We have demonstrated previously that atransfer of the natural plasmid harboring the blaOXA-31 geneinto a P. aeruginosa recipient strain confers resistance tocefepime and cefpirome but that susceptibility to ceftazidime isnot modified. By comparing the MICs of �-lactams for E. colirecombinant clones producing OXA-1 or OXA-31, as well thekinetic parameters, we showed that OXA-1 and OXA-31 pos-sess similar hydrolysis profiles, including the ability to hydro-lyze cefepime and cefpirome (9). Therefore, OXA-1 andOXA-31 may be considered to be class D �-lactamases charac-terized by expanded-spectrum profiles with significant impactson MICs for bacterial species that possess high-level intrinsicimpermeability (such as P. aeruginosa) and not on those forbacterial species with low-level intrinsic impermeability (suchas E. coli).
All the genes for the OXA-1-like �-lactamases describedabove have been identified in the form of gene cassettes in-serted into class 1 integrons (9, 44, 107, 127).
The OXA-2 subgroup. OXA-2 �-lactamase is another nar-row-spectrum �-lactamase that shares ca. 30% amino acididentity with OXA-1 (77). It constitutes, together with itsderivatives OXA-3, OXA-15, OXA-21, OXA-32, OXA-34,OXA-36, and OXA-53, a distinct genetic cluster (Fig. 2). TheblaOXA-2 gene has been identified often in P. aeruginosa and S.Typhimurium isolates producing the ESBL PER-1 (46), as hasthe variant OXA-3 (142). Recently, blaOXA-2 has been identi-fied in P. aeruginosa isolates from Serbia and Hungary that alsoproduced the ESBL PER-1 (81). In addition, it has been iden-tified in other species such as Morganella morganii in Argentina(135), K. pneumoniae in Uruguay (161), and E. coli in France(GenBank accession no. CAJ13583). Surprisingly, it has alsobeen identified in distantly related species such as Bordetellabronchiseptica (GenBank accession no. ABD63309) and Aero-monas hydrophila (GenBank accession no. ABF69297) andeven the gram-positive species Corynebacterium amycolatum(GenBank accession no. CAI40608) (unpublished data). TheOXA-21 variant, a point mutation derivative of the OXA-3 en-zyme, has been identified once in Acinetobacter baumannii(163). The genes for these �-lactamases have been also iden-tified in the form of gene cassettes inserted into class 1 inte-grons.
The OXA-10 subgroup. OXA-10 �-lactamase (formerlyknown as PSE-2) also possesses the ability to hydrolyze ceph-alosporins, hydrolyzing cefotaxime, ceftriaxone, and aztreo-nam at low levels but sparing ceftazidime, cephamycins, andcarbapenems (70). The blaOXA-10 gene is encountered in alarge variety of gram-negative species, being the blaOXA geneidentified most frequently in P. aeruginosa (8, 25, 37–39, 46, 60,99, 125).
There are some point mutation derivatives of OXA-10, suchas OXA-11, OXA-13, OXA-16, OXA-28, OXA-35, and OXA-74, possessing increased activities toward expanded-spectrum
cephalosporins (see below) (Fig. 2). Very few OXA-10-likeenzymes possessing a narrow spectrum of hydrolysis have beendescribed, one of which is OXA-7, encoded by a gene reportedto be plasmid borne in E. coli. OXA-7 has 95% amino acididentity to OXA-10 (91, 145).
Other subgroups. Among the narrow-spectrum OXAs, somevariants are not, or are weakly, related to the OXA-1, OXA-2,and OXA-10 subgroups (Fig. 2). This is the case for OXA-9,which shares 45 and 54% amino acid identity with the mostclosely related enzymes, OXA-12 and OXA-18 (155). OXA-9has the property, uncommon for an OXA enzyme, of beinginhibited by clavulanic acid and cloxacillin but not by NaCl(16). Its hydrolytic profile remains to be precisely determined,even if OXA-9 would likely correspond to a narrow-spectrum�-lactamase (our unpublished data). The blaOXA-9 gene wasfirst identified on a plasmid in a K. pneumoniae isolate (155).The blaOXA-9 gene has been identified in a Pseudomonasputida isolate from France coexpressing the metallo-�-lacta-mase VIM-2 (122), in an Enterobacter cloacae isolate fromCanada (119), and in an Enterobacter aerogenes isolate fromFrance (120). Recently, it has also been identified in a carbap-enem-resistant E. cloacae strain originating in the UntiedStates and coproducing the class A carbapenemase KPC-3 (43)and in a carbapenem-resistant K. pneumoniae isolate in Turkeycoproducing the class D carbapenemase OXA-48 (127).
Another narrow-spectrum class D �-lactamase is LCR-1,which was one of the very first class D �-lactamases to beidentified on the basis of biochemical properties only (146),was recovered from P. aeruginosa, and hydrolyzes only penicil-lins, nitrocefin, and oxacillin (168). Its recently determinedamino acid sequence (32) shows that it belongs to a separatecluster among class D enzymes (Fig. 2), sharing 40% or lessamino acid identity with other OXAs (40% with OXA-55 and37% with OXA-48). NPS-1 is another narrow-spectrum en-zyme which is related to LCR-1 (carrying eight amino aciddifferences) (114). The blaNPS-1 gene was identified on a con-jugative plasmid from P. aeruginosa isolates in the UnitedKingdom (83).
The product of the blaOXA-5 gene represents another sub-group of narrow-spectrum class D �-lactamases (Fig. 2). Afterthe gene was phenotypically described as plasmid borne in a P.aeruginosa isolate recovered in the United Kingdom (91), itssequence was determined (32). The OXA-5 determinantshares less than 81% identity with OXA-10 derivatives. TheblaOXA-5 gene was found as a gene cassette inserted into a class1 integron and was later found in association with the blaGES-2
gene, encoding a peculiar ESBL with carbapenemase proper-ties, in carbapenem-resistant P. aeruginosa isolates from SouthAfrica during an outbreak period (134). The hydrolysis spec-trum of OXA-5 is limited to narrow-spectrum penicillins, ox-acillin, and cephalothin (cefalotin). Its activity is inhibited byNaCl but not by clavulanate (91). Recently, OXA-129 wasidentified in S. enterica subsp. enterica serovar Bredeney por-cine isolates from Brazil (95). It differs from OXA-5 by 12amino acids.
OXA-20 is another narrow-spectrum class D �-lactamasethat has weak amino acid identity to other class D �-lactamases(108). It shares less than 75% identity with the most closelyrelated enzyme, OXA-2. OXA-20 hydrolyzes penicillins,cephalothin, and cephaloridine but spares expanded-spec-
28 MINIREVIEW ANTIMICROB. AGENTS CHEMOTHER.

trum cephalosporins. Interestingly, OXA-20 is inhibited byclavulanic acid (108). The blaOXA-20 gene was identified in a P.aeruginosa isolate from Sicily, Italy, that coexpressed OXA-18,the latter OXA possessing an expanded-spectrum profile (118)(see below). Noteworthily, the blaOXA-20 gene was also iden-tified in A. baumannii isolates from France and Italy and wasalso located inside integron structures (53, 58, 121, 169). Apoint mutation derivative of OXA-20, OXA-37, has been iden-tified in an A. baumannii isolate from Spain (109).
OXA-46 was identified in a P. aeruginosa isolate from Italy.In fact, this determinant was identified in a multidrug-resistantisolate that coexpressed the metallo-�-lactamase VIM-1 (57).
OXA-46 shares 78% amino acid identity with OXA-2 andOXA-53, the most closely related enzymes. It hydrolyzes mostlypenicillins, oxacillin, and narrow-spectrum cephalosporins. In-terestingly, it is inactivated by tazobactam and carbapenemsbut exhibits low susceptibility to NaCl (57).
ACQUIRED ES-OXAs
Whereas most of the previously mentioned class D �-lacta-mases exhibit a substrate profile limited mostly to penicillins,nitrocefin, oxacillin, and cloxacillin, class D �-lactamases ableto hydrolyze expanded-spectrum cephalosporins have also
FIG. 2. Dendrogram obtained for 73 class D �-lactamases by using Phylip (ClustalW). Branch lengths are to scale and proportional to thenumber of amino acid changes. The distance along the vertical axis has no significance. The different clusters identified allowed the identificationof nine main groups, considering that proteins from the same group have more than 80% amino acid identity.
VOL. 54, 2010 MINIREVIEW 29

been identified (though their distribution remains quite lim-ited). There are two main types of expanded-spectrum class D�-lactamases (ES-OXAs) (Table 1). Some ES-OXAs are justpoint mutation derivatives of narrow-spectrum class D �-lac-tamases. Others may be completely different enzymes withweak amino acid sequence identity. Interestingly, most of theES-OXAs identified so far are from P. aeruginosa.
ES-OXAs as point mutation derivatives of narrow-spectrumOXAs. The first variant described to possess an extended-spectrum profile was OXA-15, a point mutation derivative ofOXA-2 (with a change from Asp to Gly at position 150 in theDBL numbering system). The blaOXA-15 gene was identifiedinside a class 1 integron structure in a P. aeruginosa isolatefrom Turkey (36). OXA-15 has increased ability to hydrolyzeceftazidime compared to that of OXA-2, but it also hydrolyzescefepime and aztreonam at a lower level than OXA-2. OXA-32 isanother OXA-2 derivative possessing an expanded-spectrumhydrolysis profile, identified in a ceftazidime-resistant P.aeruginosa isolate recovered from a patient originating fromGuadeloupe in the French West Indies (124). The blaOXA-32
gene was integron borne and located on a 250-kb conjugativeplasmid. OXA-32 differs from OXA-2 by a Leu169Ile substi-tution (DBL numbering) that extends its hydrolysis spectrumtoward ceftazidime but not cefotaxime (124). Interestingly, thissubstitution also modifies the behavior of the class D �-lacta-mase OXA-32 toward inhibitors, since imipenem and cefoxitinwere found to significantly inhibit the activity of OXA-32 com-pared to that of OXA-2.
A large number of class D �-lactamases with expanded-spectrum activity have been identified among OXA-10 vari-ants. OXA-11, identified in a P. aeruginosa isolate from aTurkish patient, was found to be encoded by a plasmid (60).Compared to OXA-10, it possesses two substitutions(Asn146Ser and Gly167Asp [DBL numbering]) that extend itsability to hydrolyze ceftazidime. Other OXA-10-like ES-OXAshave been identified in P. aeruginosa strains from Turkey, in-cluding OXA-14 (carrying a Gly167Asp change) (38), OXA-16(with Ala114Thr and Gly167Asp changes) (39), and OXA-17,the latter having an Asn76Ser substitution conferring an in-creased hydrolysis rate for cefotaxime and a decreased rate forceftazidime (37). OXA-35, which has been identified in a P.aeruginosa isolate from France (8), is a point mutation deriv-ative of both OXA-19 (95) and OXA-28 (125). OXA-35 pos-sesses hydrolytic activity toward penicillins and cefsulodin butspares narrow- and expanded-spectrum cephalosporins. In con-trast, OXA-19 and OXA-28, differing from OXA-10 by 9 and10 amino acid substitutions, respectively, confer resistance toceftazidime (98, 125).
Other ES-OXAs. Several ES-OXAs are not structurally re-lated to narrow-spectrum OXAs. OXA-18 was the first enzymeof this type to be identified, in a P. aeruginosa isolate obtainedin 1995 in Paris from a patient previously hospitalized in Sicily(118). OXA-18 shares less than 50% amino acid identity withthe other OXAs. It is peculiar since it confers high-level resis-tance to expanded-spectrum cephalosporins and is surprisinglywell inhibited (for a class D �-lactamase) by clavulanic acid(118). Its substrate profile does not include cephamycins andcarbapenems. OXA-18 can therefore be considered a typicalESBL. The blaOXA-18 gene is very likely chromosomally lo-cated. Other OXA-18-producing P. aeruginosa isolates have
been recovered recently in Belgium, as sources of outbreaks(105). The structures surrounding the blaOXA-18 gene in thosestrains were identical to those discovered in the OXA-18-producing Italian isolate (105).
OXA-45 is another ES-OXA, identified in a multidrug-re-sistant Texan P. aeruginosa isolate coexpressing the class B�-lactamase VIM-7 (153). OXA-45 shows the highest degreesof identity to OXA-18 (66%), OXA-9 (43%), and OXA-22(40%). The blaOXA-45 gene, located on a 24-kb plasmid, wasnot in the form of a gene cassette but was associated with twocopies of an ISCR5-like element. OXA-45 includes in its sub-strate profile expanded-spectrum cephalosporins, such asceftazidime, cefotaxime, and aztreonam, but spares carbapen-ems and cephamycins. The activity of OXA-45, like that ofOXA-18, is inhibited by clavulanic acid (153).
Finally, OXA-53 is another ES-OXA that has been identi-fied recently in an S. enterica serovar Agona isolate from Brazil(103). It shares 90% amino acid identity with the narrow-spectrum OXA-2 but exhibits unusual properties since it is wellinhibited by clavulanic acid and confers reduced susceptibilityto ceftazidime. The blaOXA-53 gene was plasmid and integronborne (103).
ACQUIRED CHDLs
Some acquired class D �-lactamases may also hydrolyzecarbapenems. None of these carbapenem-hydrolyzing class D�-lactamases (CHDLs) (131, 133) significantly hydrolyze ex-panded-spectrum cephalosporins, therefore indicating thatcurrently known class D �-lactamases are unable to combineextended-spectrum and carbapenem-hydrolyzing properties.Most of these enzymes are from Acinetobacter spp.
As noticed by Queenan and Bush (137), the level of hydro-lysis of carbapenems by CHDLs remains low, due to poorturnover of these �-lactams. In general, the hydrolysis of imi-penem, even if slow, is faster than that of meropenem. Usually,the Km values for imipenem are low, indicating that CHDLshave very high apparent affinities for that substrate (65, 130).Due to these peculiar properties, the exact contribution of theseenzymes to phenotypic resistance has been debated. However,it has been demonstrated recently using knockout or comple-mentation experiments that acquired CHDLs such as OXA-23,OXA-40, and OXA-58 identified in A. baumannii significantlycontribute to carbapenem resistance (67).
The first reported acquired class D �-lactamase with carbap-enemase activity was OXA-23 (also known as ARI-1 [43, 115]),detected in an A. baumannii isolate from Scotland and foundto be plasmid mediated after its transfer to Acinetobacter junii(143). The ability of OXA-23 to hydrolyze carbapenems wasweak, since hydrolytic measurements showed only low-levelactivity against imipenem. However, compared to the activitiesof other class D �-lactamases, this hydrolysis was significantand was allowed to define the novel subgroup of OXAs cur-rently named CHDLs (133). OXA-23, weakly related to otherclass D �-lactamases with only 36% amino acid identity toOXA-5 and OXA-10, belongs to the first group of CHDLs.Although kinetic parameters of hydrolysis have been deter-mined with crude extracts of OXA-23 (115), precise parame-ters are still not available. The other member of the first groupis OXA-27, which possesses two substitutions (Thr86Ala and
30 MINIREVIEW ANTIMICROB. AGENTS CHEMOTHER.

Asn250Lys [DBL numbering]) compared to OXA-23 (1). Thecorresponding blaOXA-27 gene, identified in a carbapenem-resistant A. baumannii isolate from Singapore, was likely chro-mosomally located (1). Whereas blaOXA-27 has been identifiedin only a single isolate, blaOXA-23 is widespread and identifiedonly in Acinetobacter spp., with a single exception, since thisgene was also identified as chromosomally carried in a carbap-enem-resistant Proteus mirabilis isolate from France (17). Re-cent data indicate that OXA-23-positive A. baumannii strainshave spread throughout the world, to locations includingFrance (30), Bulgaria (149), Iran (48), the United Arab Emir-ates (101), Tunisia (86), Brazil (35), and Australia (160). Inaddition, OXA-23 producers have been at the origin of hospi-tal outbreaks in French Polynesia (104), Colombia (163), theUnited Kingdom (28), Turkey (92), China (166, 170), andKorea (74). OXA-23-positive A. baumannii isolates and one A.junii strain have been also identified in Romania (87).
A second group of CHDLs is made up of OXA-25, OXA-26,OXA-40, and OXA-72 (an original sequencing error in OXA-24makes it now OXA-40 [see the Lahey website at http://www.lahey.org/Studies/]). These enzymes differ by a few amino acidsubstitutions only. OXA-24/OXA-40 was originally identifiedas chromosomally encoded in a carbapenem-resistant A. bau-mannii isolate recovered from Spain (18, 19). The blaOXA-40
gene has since been identified in different areas, especially inPortugal and Spain (40, 116, 138, 141), but also in the UntiedStates (84, 136). Interestingly, the blaOXA-40 gene has beenidentified as either chromosomally carried or plasmid borne(138). Surprisingly, it has been very recenty identified as plas-mid located in two carbapenem-resistant P. aeruginosa isolatesin Spain (146). OXA-40 is not inhibited by NaCl (65). OXA-25and OXA-26 are point mutation derivatives of OXA-40 thathave been identified in carbapenem-resistant A. baumanniiisolates recovered from Spain and Belgium, respectively (1).OXA-72 has been identified in A. baumannii isolates fromChina, South Korea, Taiwan, and Bahrain (80, 85, 102, 166).
A third identified group of CHDLs is represented by OXA-58, which has also been detected only in Acinetobacter spp. sofar. OXA-58 was first identified in a multidrug-resistant A.baumannii isolate from France (130), that strain being at theorigin of an outbreak in the local hospital (64). OXA-58 shares35, 33, and 18% amino acid identity with OXA-5, OXA-10, andOXA-1, respectively. OXA-58 hydrolyzes penicillins and car-bapenems at low levels. Whereas weak hydrolysis of cefpiromewas detected, hydrolysis of cefepime, ceftazidime, and cefo-taxime was not. The rate of hydrolysis of imipenem was 10-foldlower, and that of meropenem was 100-fold lower, than that ofbenzylpenicillin (130). The blaOXA-58 gene was found to beplasmid borne in A. baumannii MAD (130), as well as in mostof the OXA-58-producing A. baumannii isolates reportedworldwide, including those in Europe, Argentina, Australia,and the United States (26–28, 69, 87, 117); OXA-58 is oftenassociated with hospital outbreaks and has been involved insuch outbreaks in France, Belgium, Italy, Turkey, Greece, andthe United States (12, 15, 53, 64, 69, 128, 159). Noteworthily,OXA-58 has also been identified in other Acinetobacter spe-cies, such as A. junii in Romania (87) and Australia (117),Acinetobacter genomospecies 3 in Spain (88), and Acinetobacterphenon 6/ct13TU in Spain (89). OXA-97 is a point mutationvariant of OXA-58 that shares the same hydrolytic properties
and has been recently identified in A. baumannii isolates fromTunisia (129). The blaOXA-97 gene was plasmid carried andidentified in a clonal strain at the origin of an outbreak at theSalhoul Hospital in Sousse, Tunisia. Another point mutationderivative is OXA-96, identified in A. baumannii in Singapore(75).
Very recently, the novel CHDL OXA-143 was identified ina clinical A. baumannii isolate that had been recovered inBrazil (68a). It shares 88% amino acid identity with OXA-40,63% with OXA-23, and 52% with OXA-58. It may thereforecorrespond to a new subgroup of CHDLs encountered in A.baumannii and involved in carbapenem resistance in that spe-cies. Its substrate profile was similar to those of other CHDLs,and the corresponding gene was not integron or transposoncarried but was likely acquired by a homologous recombinationprocess.
Another unrelated CHDL is OXA-48, which has been iden-tified in a carbapenem-resistant K. pneumoniae isolate fromIstanbul, Turkey (127), thus, in a member of the Enterobacte-riaceae. That isolate coexpressed several �-lactamases, includ-ing the class A ESBL SHV-2a and the narrow-spectrum �-lac-tamases TEM-1 and OXA-47. OXA-48 �-lactamase hydrolyzespenicillins and, at a lower level, imipenem but is not activeagainst expanded-spectrum cephalosporins (127). This enzymeis the class D �-lactamase with the highest known catalyticefficiency for imipenem (having a kcat value of 2 s�1) (127). Itshares less than 46% amino acid identity with other class D�-lactamases and has weak identity to OXA-23 and OXA-40(36 and 32%, respectively). The blaOXA-48 gene was found tobe plasmid located in K. pneumoniae 11978 (127). Very re-cently, we reported an important outbreak of infection withOXA-48-producing K. pneumoniae that occurred in Istanbulfrom May 2006 to January 2007 (24). Two distinct clones dif-fering from the first OXA-48 producer and producing differentESBL determinants (SHV-12 and CTX-M-15) were identifiedin the same hospital. In addition, the blaOXA-48 gene has beenidentified in E. coli and Citrobacter freundii strains in Turkey(59, 110). The spread of the blaOXA-48 gene may be much moreimportant than expected. Indeed, the gene has been found inK. pneumoniae isolates from Lebanon and from Belgium (34,90). Its detection may be difficult since the level of acquiredresistance to carbapenems may remain quite low.
Overall, the hydrolysis spectra of these class D �-lactamasesare quite consistent, all enzymes hydrolyzing imipenem at lowlevels but not broad-spectrum cephalosporins and aztreonam.
NATURALLY OCCURRING CLASS D �-LACTAMASES
Recent studies have shown that many bacterial species pos-sess chromosomally located and naturally occurring class D�-lactamase genes. The first such OXA gene to be identifiedwas that from Aeromonas jandaei (formerly A. sobria). Thisspecies naturally produces the class D �-lactamase OXA-12,which has strong activity against cloxacillin and oxacillin andwhose expression is inducible (3, 139). Another class D �-lac-tamase, AmpS, lacking significant activity against cloxacillinand oxacillin but hydrolyzing penicillins, was also detected inA. jandaei (A. sobria) (165). AmpS shares 96% amino acididentity with OXA-12. However, analysis using current GenBankdatabases indicates that AmpS likely corresponds to the intrin-
VOL. 54, 2010 MINIREVIEW 31

sic class D �-lactamase of another Aeromonas species whosegenome sequences have been released, namely, A. hy-drophila (GenBank accession no. YP_858675). �-Lactama-ses OXA-12 and AmpS both possess the property, peculiarfor a class D enzyme, of being inhibited by clavulanic acid(139, 165).
Ralstonia pickettii is naturally resistant to penicillins, narrow-spectrum cephalosporins, ceftazidime, and aztreonam or showsintermediate susceptibilities to these agents. Two types of classD �-lactamases have been identified in that species. OXA-22exhibits narrow-spectrum �-lactam hydrolysis activity that doesnot explain resistance to monobactams in its natural host(112). The gene encoding OXA-22 is chromosomally located,and induction experiments demonstrated that its expression isinducible (linked to a peculiar open reading frame [ORF]located upstream of blaOXA-22) but also revealed the overex-pression of another �-lactamase gene. Further cloning exper-iments identified a second class D �-lactamase gene, blaOXA-60,which is also located on the chromosome of R. pickettii (55).PCR screening of multiple R. pickettii isolates identified vari-ants of both blaOXA-22 and blaOXA-60 genes. OXA-60 possessesonly 19% amino acid identity to OXA-22 and is weakly relatedto the other OXAs. It exhibits a narrow-spectrum hydrolysisprofile, including activity against penicillins and nitrocefin andweak activity against carbapenems (55). The molecular basis ofthe induction process requires the presence of ORF RP3,located 190 bp upstream of blaOXA-60 and divergently tran-scribed. Disruption of ORF RP3 abolishes induction of both�-lactamases, suggesting that ORF RP3 may be a global reg-ulator, although the 532-amino-acid-long product displays noobvious sequence homology to known regulatory proteins (56).
The blaOXA-61 gene was identified on the chromosome ofCampylobacter jejuni and encodes a narrow-spectrum �-lacta-mase not inhibited by clavulanic acid (2). Considering theMICs of �-lactams for the original C. jejuni isolates, it seemslikely that OXA-61-like enzymes are poorly expressed in theirnatural host and consequently play a minor role in susceptibil-ity to �-lactams (2).
The blaOXA-62 gene was identified on the chromosome of acarbapenem-resistant Pandoarea pnomenusa isolate recoveredfrom a cystic fibrosis patient (145). Pandoraea spp. are gram-negative, non-glucose-fermenting rods. The blaOXA-62 gene,which possesses an unusually high GC content of 65.3%, en-codes a CHDL that is well inhibited by NaCl and hydrolyzespenicillins and oxacillin, as well as imipenem and meropenemat low levels, but does not hydrolyze expanded-spectrum ceph-alosporins. OXA-62 shares weak amino acid identity withother class D �-lactamases, the highest degree of similaritybeing that to OXA-50 (43% amino acid identity). RelatedblaOXA-62 genes have been identified in other P. pnomenusaisolates but not in other Pandoraea species (144).
The causative agent of melioidosis, Burkholderia pseudoma-llei, naturally produces OXA-42 (111) and derivatives, such asOXA-43 (111) and OXA-57 and OXA-59 (73). OXA-57, whichhas also been identified in Bulkholderia thailandensis, is theonly enzyme among these variants that has been studied bio-chemically, and it hydrolyzes penicillins, oxacillin, cephalothin,and nitrocefin but spares ceftazidime. It is uninhibited by cla-vulanate and uninhibited by NaCl, the latter property beingquite uncommon (73).
The blaOXA-63 gene was identified in a clinical isolate of theanaerobic spirochete species Brachyspira pilosicoli (94). It en-codes a narrow-spectrum �-lactamase that possesses the un-usual property of being well inhibited by clavulanic acid andresistant to NaCl inhibition. OXA-63 shows the closest identity(53%) to FUS-1 (OXA-85) (see below). Very recently, Mor-timer-Jones and colleagues (96) analyzed B. pilosicoli srainsrecovered from humans and pigs in Australia and Papua NewGuinea. They identified genes encoding OXA-136 and OXA-137, sharing 94 to 95% amino acid identity with OXA-63 andconferring similar patterns of resistance once the genes werecloned and expressed in E. coli. Interestingly, in that studyblaOXA-63-like genes were identified in penicillin-resistantstrains only and not in penicillin-susceptible isolates (96).
FUS-1 (also named OXA-85) corresponds to the naturallyoccurring class D �-lactamase of Fusobacterium nucleatumsubsp. polymorphum, a gram-negative anaerobic rod (164).OXA-85 shares only 25 to 44% identity with other class D�-lactamases. It also possesses a narrow spectrum of �-lactamhydrolysis and is not inhibited by NaCl or by clavulanate.
The blaOXA-29 gene was identified in a Legionella (Fluori-bacter) gormanii strain recovered from soil (52). OXA-29shares less than 50% amino acid identity with other OXAenzymes (44% with OXA-18) and hydrolyzes oxacillin, peni-cillins, cefazolin, and at lower levels, cefuroxime, cefotaxime,ceftazidime, and aztreonam, sparing carbapenems. OXA-29activity is inhibited by tazobactam but not by clavulanic acidand is also surprisingly resistant to NaCl (52).
The blaOXA-114 gene was cloned from the genome of Ach-romobacter xylosoxidans (formerly Alcaligenes denitrificanssubsp. xylosoxydans) (41). It encodes OXA-114, which shares56% amino acid identity with the naturally occurring class D�-lactamase of Burkholderia cepacia (as determined by GenBankin silico analysis) and 42% with �-lactamases OXA-9 andOXA-18. OXA-114 has a narrow-spectrum hydrolysis profile,although it shows a very low level of activity against imipenem.Several blaOXA-114-like genes have been identified in all fiveisolates of A. xylosoxidans tested, making such genes a featureof that species (41). In contrast to the expression of some othernatural blaOXA genes, the expression of blaOXA-114 is not in-ducible (41).
Interestingly, we recently identified Acinetobacter radioresis-tens, a nonpathogenic and environmental Acinetobacter spe-cies, as the progenitor of the acquired CHDL-encoding geneblaOXA-23 (122). Similar blaOXA genes identified in other A.radioresistens isolates encode OXA-23 variants (namely, OXA-102, OXA-103, and OXA-105) exhibiting no more than sixamino acid substitutions compared to OXA-23 and conferringsimilar levels of resistance to carbapenems once expressed inE. coli (123). Since both A. baumannii and A. radioresistensnaturally possess class D �-lactamase genes, we believe thatother Acinetobacter species may act as reservoirs for otherclinically relevant class D �-lactamase genes. Preliminary in-vestigations identified the blaOXA-134 gene in the chromosomeof Acinetobacter lwoffii (51). blaOXA-134 encodes a CHDL thatshares 63, 58, 57, and 53% amino acid identity with OXA-23,OXA-51, OXA-24, and OXA-58, respectively. The substrateprofile of OXA-134, including penicillins and carbapenems, issimilar to those of other CHDLs.
Expansion of the knowledge of entire genome sequences
32 MINIREVIEW ANTIMICROB. AGENTS CHEMOTHER.

leads to the identification of putative ORFs that may encodeclass D �-lactamases. Some of these ORFs have subsequentlybeen shown to encode functional �-lactamases, while the prod-ucts of others are not �-lactamases even if such designationsare falsely used in the databases, constituting a source of con-fusion. Some other products are good candidates for �-lacta-mases on the basis of in silico analysis and require furtherinvestigations, in particular accurate enzymatic analyses.
Sequencing analysis of the entire genome of A. baumanniiisolate AYE revealed a chromosomal gene encoding OXA-69that was further characterized (66). OXA-69 shares 62 and56% amino acid identity with OXA-40 and OXA-23, respec-tively, and 97% identity with OXA-51, identified in severalunrelated A. baumannii isolates from Argentina (22). OXA-69hydrolyzes penicillins and, at low levels, carbapenems. OXA-51/OXA-69 enzymes do not contribute significantly to the nat-ural resistance pattern observed in A. baumannii. The blaOXA
genes for these enzymes are neither inducible nor regulated.Genes encoding OXA-51/OXA-69 derivatives were identified indifferent collections of A. baumannii isolates recovered fromwidespread geographical areas (66, 93). Some of these variantshad also been identified in carbapenem-resistant A. baumanniiisolates disseminated worldwide (21, 47, 158). Up to 45 OXA-51/OXA-69 variants are now known (Table 1), and most if not allprobably have identical biochemical properties. Due to the verylarge and increasing number of OXA-51/OXA-69 variants iden-tified, we propose here to define an alternative nomenclature forthese class D �-lactamases specific for A. baumannii, which mayinclude OXA-AB (for OXA-A. baumannii) followed by a numberand which would start with OXA-51’s being consequently re-named OXA-Ab1, with the next enzyme being called OXA-Ab2,etc. (this nomenclature is presented in Table 1). Interestingly,even though the role of OXA-51-like �-lactamases in conferringthe natural resistance pattern is small, they may be at least par-tially the source of acquired resistance. Indeed, it has been re-ported previously that insertion sequences ISAba1 and ISAba9may sometimes be located upstream of the blaOXA-51-like gene incarbapenem-resistant A. baumannii isolates, providing promotersequences for high-level expression of OXA-51-like enzymes (49,50, 157).
In silico analysis has identified a putative class D �-lacta-mase gene in the chromosome of P. aeruginosa. The gene,blaOXA-50, was cloned and expressed in E. coli (54). OXA-50(also referred to as PoxB [76]) hydrolyzes �-lactams at a verylow level (54) and plays only a minor role in the phenotype ofnatural resistance to �-lactams in P. aeruginosa. OXA-50 vari-ants have been identified in diverse P. aeruginosa clinical iso-lates recovered from different geographical areas (54). How-ever, the expression of blaOXA-50 in P. aeruginosa is not inducible,as opposed to that of the blaOXA-22 gene in R. pickettii (56).
In silico analysis revealed that the chromosome ofShewanella oneidensis strain MR-1, an environmental gram-negative isolate from lake sediment, harbored a class D �-lac-tamase gene (126). This blaOXA-54 gene was cloned and ex-pressed in E. coli. OXA-54 shares 92% amino acid identitywith OXA-48, the gene for which was identified in K. pneu-moniae (see above). OXA-54 possesses enzymatic propertiessimilar to those of OXA-48, significantly hydrolyzing imipenem(with even a higher kcat/Km ratio due to better affinity for thissubstrate) and not being inhibited by clavulanic acid. Thus, S.
oneidensis may be considered a reservoir of CHDLs that areable to spread among Enterobacteriaceae (126).
More generally, the Shewanella genus may be considered tobe a progenitor of other related enzymes, since OXA-55 (with55% amino acid identity to OXA-54) was further identified inShewanella algae and possesses biochemical properties similarto those of OXA-54, in particular the ability to hydrolyzecarbapenems (68). In addition, with the recent release of manygenome sequences, it appears after in silico analysis that Sh-ewanella-related species are often (if not always) potentialsources of class D �-lactamase genes, as observed for Sh-ewanella pealeana, Shewanella loihica, Shewanella baltica, andShewanella putrefaciens (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
In addition, in silico analyses of several other genomes ofvery unrelated species, such as Limnobacter sp. (Burkholderi-aceae family), Idiomarina loihiensis (Idiomarinaceae family),Bradyrhizobium japonicum and Rhodopseudomonas palustris(Bradyrhizobiaceae family), Delftia acidovorans (Comamona-daceae family), Geobacter uraniireducens (Geobacteraceae fam-ily), Agrobacterium tumefaciens (Rhizobiaceae family), and Cy-tophaga hutchinsonii (“Flexibacteraceae” family), reveal someORFs that may correspond to class D �-lactamases. However,due to the lack of adequate cloning experiments, the ability ofall these putative proteins to act as �-lactamases has not yetbeen demonstrated. These observations, however, suggest thatsuch ORFs may play a physiological but still unidentified rolein many gram-negative organisms.
GENETIC PLATFORMS FOR ACQUIRED CLASS D�-LACTAMASE GENES
Most of the class D �-lactamase genes identified in Entero-bacteriaceae and Pseudomonas species are located on large,transferable plasmids. Some class D �-lactamase genes foundin P. aeruginosa, such as those encoding OXA-13, OXA-17,OXA-18, and OXA-20, have been found to be chromosomallylocated (37, 99, 108, 118). However, negative results frommating-out assays and the lack of plasmid detection in P.aeruginosa do not rule out plasmid origins for a class D �-lac-tamase gene.
Most plasmid-located class D �-lactamase genes have beenfound in association with genetic vehicles responsible for theirmobilization. These vehicles are mobile elements, often trans-posons belonging to the Tn3 family (10, 82). These transposonsenable integrons (which are not self-mobile elements) and,consequently, their associated resistance gene cassettes to dif-fuse and spread. Integrons are genetic structures capable ofintegrating individual genes in the form of gene cassettes (61).
Among class D �-lactamase genes which have been identi-fied in the form of gene cassettes (140), the most common arethe blaOXA-1-like, blaOXA-2-like, and blaOXA-10-like genes(107). There are peculiar features associated with the blaOXA-9
gene identified in a plasmid from a K. pneumoniae isolate(155), since the blaOXA-9 gene was in the form of a genecassette but, instead of being associated with an integron, waspart of the Tn3-type transposon Tn1331 also carrying theblaTEM-1 gene and lacking an integrase gene.
Besides all these blaOXA gene cassettes, there are otherblaOXA genes which have been identified in other genetic en-vironments. The blaOXA-23 gene originating from the chromo-
VOL. 54, 2010 MINIREVIEW 33

some of A. radioresistens may be mobilized onto plasmids dif-fusing into A. baumannii through different genetic structures,such as the composite transposon Tn2006, formed by twoISAba1 elements (30, 100). Alternatively, ISAba4 was foundupstream of the blaOXA-23 gene in a single copy (14, 30).
The blaOXA-48 gene has been identified in association withinsertion sequence IS1999 in K. pneumoniae (127). TheblaOXA-48 gene was part of composite transposon Tn1999,made of two copies of IS1999, with the copy located upstreamof blaOXA-48 providing promoter sequences for blaOXA-48 ex-pression (7).
The blaOXA-58 gene, which has been identified only in Acin-etobacter sp. isolates, has been associated with a variety ofdifferent genetic structures. By analyzing a series of OXA-58-positive A. baumannii isolates recovered in different countries(France, Spain, Romania, Greece, and Turkey), 11 distinctstructures were identified (128, 130, 132). Nevertheless, thereare always common features, since blaOXA-58 is always brack-eted by two ISAba3 elements. The ISAba3 copy upstream ofblaOXA-58, as well as other insertion sequences, such as IS18,ISAba1, and ISAba2, may provide promoter sequences playinga role in blaOXA-58 expression (132). For the A. baumanniiisolate MAD, which has been at the origin of a clonal outbreakin a French hospital (64, 130), we demonstrated that the entirestructure encompassing the ISAba2-blaOXA-58-ISAba3 frag-ment had very likely integrated into the plasmid backbone by arecombination process (132). In addition to being dependenton IS elements, the level of expression of the blaOXA-58 genemay be related to the gene copy number, as exemplified in oneItalian clone exhibiting various levels of resistance to imi-penem as a consequence of the blaOXA-58 copy number (13).
Finally, another mechanism of acquisition of class D �-lac-tamase genes has been identified recently. It is mobilizationby peculiar IS elements named ISCRs, which mobilize adjacentsequences by rolling-circle (RC) transposition (151, 152).blaOXA-18 gene acquisition is likely the result of RC transpo-sition mediated by the ISCR19 element (105), and the aquisi-tion of the blaOXA-45 gene is likely mediated by ISCR5 (154).
DETECTION OF CLASS D �-LACTAMASE GENES
One of the major concerns for controlling the spread of classD �-lactamase producers is the absence of phenotypic teststhat could contribute to their easy recognition. Although toolsare available for the detection of class A ESBLs (clavulanicacid-based synergy tests), metallo-�-lactamases (EDTA-basedapproaches), and AmpC enzymes (cloxacillin-based tests),there is no phenotypic tool allowing rapid and easy detection ofclass D �-lactamase producers. However, some class D �-lac-tamases possess peculiar properties that may facilitate theirdetection. For example, OXA-13 and its extended-spectrumvariant OXA-19 both possess the ability to be inhibited byimipenem (98). Once the enzyme is produced in P. aeruginosa,it is possible to demonstrate this characteristic by placing a diskof imipenem next to one of cefsulodin, a substrate well hydro-lyzed by OXA-13 (99). We have observed that this specificproperty is shared by other class D �-lactamases, such as thosebelonging to the OXA-10 subgroup (unpublished data). Also,the production of some OXAs whose activity is inhibited byclavulanate or tazobactam, such as OXA-12 from A. jandaei
(139) and OXA-18, OXA-45, and OXA-46 from P. aeruginosa(57, 118, 153), may be identified by synergy tests using clavu-lanic acid-containing disks. However, the result of these syn-ergy tests may be misinterpretated and the corresponding iso-lates may be considered to be class A ESBL producers.
One alternative approach that may be performed in special-ized laboratories is spectrophometric analysis. By using crudeextracts and UV spectrophotometry, it is possible to evaluatethe abilities of �-lactamase extracts to hydrolyze oxacillin. It isalso possible to use the NaCl inhibition property with a refer-enced substrate such as benzylpenicillin. However, as indicatedabove, there are some limits in this approach since (i) someclass D �-lactamases do not hydrolyze oxacillin, (ii) the in vitroinhibition of their activities by NaCl is not always observed,and (iii) OXA-positive clinical isolates often express additionalnon-class D �-lactamases. In addition, many CHDLs hydrolyzecarbapenems at a low level, interfering negatively with thesensitivity of the technique. PCR-based methods remain,therefore, the “gold standard” for the identification of class D�-lactamases (133, 167).
CONCLUSIONS
OXA �-lactamases are distributed in a large variety of gram-negative species. Whereas their acquisition is detected mostlyin Enterobacteriaceae, Pseudomonas spp., and Acinetobacterspp., naturally occurring chromosomal genes are increasinglydiscovered in gram-negative organisms, such as Aeromonas,Legionella, Shewanella, and Campylobacter species. The iden-tification of the progenitor of a clinically significant OXA is anexciting perspective, since it may help to better control thesources of emerging class D �-lactamases, to better evaluatetheir dissemination, and eventually to better control theirspread.
According to Barlow and Hall (11), this diversity of class D�-lactamases, at least those whose genes have been incorpo-rated onto plasmids, is the result of ancient events that oc-curred millions of years ago. That hypothesis is debatable,since for example, the widely distributed OXA-23 determinanthas been found with almost perfect identity in the chromosomeof an A. radioresistens strain which corresponds to its naturalreservoir. It is very likely that the diversity of plasmid-borneblaOXA genes simply reflects the diversity of their natural pro-genitors.
It is likely that antibiotic selective pressure is a driving forcefor the spread of several blaOXA genes. The blaOXA genesidentified as having been aquired are often expressed at highlevels, and in contrast, the genes are very often silent in theirnatural progenitors. Mobilization elements (e.g., integrons andIS elements) contribute to the spread and expression of thesegenes by providing strong promoters. Future work may bedirected toward the role of antibiotic concentrations in givenenvironments as a source of enhancement of the spread ofblaOXA genes from their reservoirs to secondary hosts.
From a clinical point of view, these OXA �-lactamasesshould be considered a threat similar to �-lactamases of theother Ambler classes, since many of them possess an expandedspectrum of activity. Currently, CHDLs in A. baumannii, ES-OXAs in P. aeruginosa, and the OXA-48 CHDL in Enterobac-teriaceae represent potential sources of clinical failure for many
34 MINIREVIEW ANTIMICROB. AGENTS CHEMOTHER.

�-lactams (70a). Lack of detection may enhance their hiddenand rapid spread among clinical isolates.
ACKNOWLEDGMENTS
The work performed in our unit and acknowledged in the review hasbeen funded partly by a grant from the Ministere de l’EducationNationale et de la Recherche (UPRES-EA3539), Universite Paris XI,Paris, France, and mostly by grants from the European Community(DRESP2 grant no. LSHM-CT-2005-018705 and TROCAR grant no.HEALTH-F3-2008-223031) and the INSERM (Paris, France).
REFERENCES
1. Afzal-Shah, M., N. Woodford, and D. Livermore. 2001. Characterization ofOXA-25, OXA-26, and OXA-27, molecular class D �-lactamases associ-ated with carbapenem resistance in clinical isolates of Acinetobacter bau-mannii. Antimicrob. Agents Chemother. 45:583–588.
2. Alfredson, D. A., and V. Korolik. 2005. Isolation and expression of a novelmolecular class D �-lactamase, OXA-61, from Campylobacter jejuni. Anti-microb. Agents Chemother. 49:2515–2518.
3. Alksne, L. E., and B. A. Rasmussen. 1997. Expression of the AsbA1,OXA-12, and AsbM1 �-lactamases in Aeromonas jandaei AER 14 is coor-dinated by a two-component regulon. J. Bacteriol. 179:2006–2013.
4. Ambler, R. P. 1980. The structure of �-lactamases. Phil. Trans. R. Soc.London B Biol. Sci. 289:321–331.
5. Ambler, R. P., A. F. Coulson, J. M. Frere, J. M. Ghuysen, B. Joris, M.Forsman, R. C. Levesque, G. Tiraby, and S. G. Waley. 1991. A standardnumbering scheme for the class A �-lactamases. Biochem. J. 276:269–272.
6. Antunes, P., J. Machado, J. C. Sousa, and L. Peixe. 2004. Disseminationamongst humans and food products of animal origin of a Salmonella typhi-murium clone expressing an integron-borne OXA-30 �-lactamase. J. Anti-microb. Chemother. 54:429–434.
7. Aubert, D., T. Naas, C. Heritier, L. Poirel, and P. Nordmann. 2006. Func-tional characterization of IS1999, an IS4 family element involved in mobi-lization and expression of �-lactam resistance genes. J. Bacteriol. 188:6506–6514.
8. Aubert, D., L. Poirel, A. Ben Ali, F. W. Goldstein, and P. Nordmann. 2001.OXA-35 is an OXA-10-related �-lactamase from Pseudomonas aeruginosa.J. Antimicrob. Chemother. 48:717–721.
9. Aubert, D., L. Poirel, J. Chevalier, S. Leotard, J.-M. Pages, and P. Nord-mann. 2001. Oxacillinase-mediated resistance to cefepime and susceptibil-ity to ceftazidime in Pseudomonas aeruginosa. Antimicrob. Agents Che-mother. 45:1615–1620.
10. Avila, P., and F. de la Cruz. 1991. Site-specific recombination and shufflingof resistance genes in transposon Tn21. Res. Microbiol. 142:701–704.
11. Barlow, M., and B. G. Hall. 2002. Phylogenetic analysis shows that the OXA�-lactamase genes have been on plasmids for millions of years. J. Mol. Evol.55:314–321.
12. Bertini, A., A. Giordano, P. Varesi, L. Villa, C. Mancini, and A. Carattoli.2006. First report of the carbapenem-hydrolyzing oxacillinase OXA-58 inAcinetobacter baumannii isolates in Italy. Antimicrob. Agents Chemother.50:2268–2269.
13. Bertini, A., L. Poirel, S. Bernabeu, D. Fortini, L. Villa, P. Nordmann, andA. Carattoli. 2007. Multicopy blaOXA-58 gene as a source of high-levelresistance to carbapenems in Acinetobacter baumannii. Antimicrob. AgentsChemother. 51:2324–2328.
14. Bogaerts, P., G. Cuzon, T. Naas, C. Bauraing, A. Deplano, B. Lissoir, P.Nordmann, and Y. Glupczynski. 2008. Carbapenem-resistant Acinetobacterbaumannii isolates expressing the blaOXA-23 gene associated with ISAba4 inBelgium. Antimicrob. Agents Chemother. 52:4205–4206.
15. Bogaerts, P., T. Naas, I. Wybo, C. Bauraing, O. Soetens, D. Pierard, P.Nordmann, and Y. Glupczynski. 2006. Outbreak of infection by carbap-enem-resistant Acinetobacter baumannii producing the carbapenemaseOXA-58 in Belgium. J. Clin. Microbiol. 44:4189–4192.
16. Bojorquez, D., M. Belei, S. F. Delira, S. Sholly, J. Mead, and M. E. Tol-masky. 1998. Characterization of OXA-9, a �-lactamase encoded by themultiresistance transposon Tn1331. Cell. Mol. Biol. (Noisy-le-Grand) 44:483–491.
17. Bonnet, R., H. Marchandin, C. Chanal, D. Sirot, R. Labia, C. De Champs,E. Jumas-Bilak, and J. Sirot. 2002. Chromosome-encoded class D �-lacta-mase OXA-23 in Proteus mirabilis. Antimicrob. Agents Chemother. 46:2004–2006.
18. Bou, G., G. Cervero, M. Angeles Dominguez, C. Quereda, and J. Martinez-Beltran. 2000. Characterization of a nosocomial outbreak caused by amultiresistant Acinetobacter baumannii strain with a carbapenem-hydrolyz-ing enzyme: high-level carbapenem resistance in A. baumannii is not duesolely to the presence of �-lactamases. J. Clin. Microbiol. 38:3299–3305.
19. Bou, G., A. Oliver, and A. Martinez-Beltran. 2000. OXA-24, a novel class D�-lactamase with carbapenemase activity in an Acinetobacter baumanniiclinical strain. Antimicrob. Agents Chemother. 44:1556–1561.
20. Boyd, D. A., and M. R. Mulvey. 2006. OXA-1 is OXA-30 is OXA-1. J.Antimicrob. Chemother. 58:224–225.
21. Brown, S., and S. G. Amyes. 2005. The sequences of seven class D �-lac-tamases isolated from carbapenem-resistant Acinetobacter baumannii fromfour continents. Clin. Microbiol. Infect. 11:326–329.
22. Brown, S., H. K. Young, and S. G. Amyes. 2005. Characterisation of OXA-51, a novel class D carbapenemase found in genetically unrelated clinicalstrains of Acinetobacter baumannii from Argentina. Clin. Microbiol. Infect.11:15–23.
23. Bush, K., G. A. Jacoby, and A. A. Medeiros. 1995. A functional classificationscheme for �-lactamases and its correlation with molecular structure. An-timicrob. Agents Chemother. 39:1211–1233.
24. Carrer, A., L. Poirel, H. Eraksoy, A. A. Cagatay, S. Badur, and P. Nord-mann. 2008. Spread of OXA-48-positive carbapenem-resistant Klebsiellapneumoniae isolates in Istanbul, Turkey. Antimicrob. Agents Chemother.52:2950–2954.
25. Carvalho, A. P., R. M. Albano, D. N. de Oliveira, D. A. Cidade, L. M.Teixeira, and A. Marques Ede. 2006. Characterization of an epidemiccarbapenem-resistant Pseudomonas aeruginosa producing SPM-1 metallo-�-lactamase in a hospital located in Rio de Janeiro, Brazil. Microb. DrugResist. 12:103–108.
26. Castanheira, M., A. Wanger, M. Kruzel, L. M. Deshpande, and R. N. Jones.2008. Emergence and clonal dissemination of OXA-24- and OXA-58-pro-ducing Acinetobacter baumannii strains in Houston, Texas: report from theSENTRY Antimicrobial Surveillance Program. J. Clin. Microbiol. 46:3179–3180.
27. Coelho, J. M., J. F. Turton, M. E. Kaufmann, J. Glover, N. Woodford, M.Warner, M.-F. Palepou, R. Pike, T. L. Pitt, B. C. Patel, and D. M. Liver-more. 2006. Occurrence of carbapenem-resistant Acinetobacter baumanniiclones at multiple hospitals in London and Southeast England. J. Clin.Microbiol. 44:3623–3627.
28. Coelho, J., N. Woodford, M. Afzal-Shah, and D. M. Livermore. 2006.Occurrence of OXA-58-like carbapenemases in Acinetobacter spp. collectedover 10 years in three continents. Antimicrob. Agents Chemother. 50:756–758.
29. Coque, T. M., A. Novais, A. Carattoli, L. Poirel, J. D. Pitout, L. Peixe, F.Baquero, R. Canton, and P. Nordmann. 2008. International disseminationof clonally related Escherichia coli strains expressing the extended-spec-trum �-lactamase CTX-M-15. Emerg. Infect. Dis. 14:195–200.
30. Corvec, S., L. Poirel, T. Naas, H. Drugeon, and P. Nordmann. 2007. Ge-netics and expression of the carbapenem-hydrolyzing oxacillinase geneblaOXA-23 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 51:1530–1533.
31. Costa, D., P. Poeta, Y. Saenz, A. C. Coelho, M. Matos, L. Vinue, J. Ro-drigues, and C. Torres. 2007. Prevalence of antimicrobial resistance andresistance genes in faecal Escherichia coli isolates recovered from healthypets. Vet. Microbiol. 127:97–105.
32. Couture, F., J. Lachapelle, and R. C. Levesque. 1992. Phylogeny of LCR-1and OXA-5 with class A and class D �-lactamases. Mol. Microbiol. 6:1693–1705.
33. Crowley, D., M. Daly, B. Lucey, P. Shine, J. J. Collins, B. Cryan, J. E.Moore, P. Murphy, G. Buckley, and S. Fanning. 2002. Molecular epidemi-ology of cystic fibrosis-linked Burkholderia cepacia complex isolates fromthree national referral centres in Ireland. J. Appl. Microbiol. 92:992–1004.
34. Cuzon, G., T. Naas, P. Bogaerts, Y. Glupczynski, T. D. Huang, and P.Nordmann. 2008. Plasmid-encoded carbapenem-hydrolyzing �-lactamaseOXA-48 in an imipenem-susceptible Klebsiella pneumoniae strain fromBelgium. Antimicrob. Agents Chemother. 52:3463–3464.
35. Dalla-Costa, L. M., J. M. Coelho, H. A. P. H. M. Souza, M. E. S. Castro,C. J. N. Stier, K. L. Bragagnolo, A. Rea-Neto, S. R. Penteado-Filho, D. M.Livermore, and N. Woodford. 2003. Outbreak of carbapenem-resistantAcinetobacter baumannii producing the OXA-23 enzyme in Curitiba, Brazil.J. Clin. Microbiol. 41:3403–3406.
36. Danel, F., L. M. Hall, D. Gur, and D. M. Livermore. 1997. OXA-15, anextended-spectrum variant of OXA-2 �-lactamase, isolated from a Pseudo-monas aeruginosa strain. Antimicrob. Agents Chemother. 41:785–790.
37. Danel, F., L. M. Hall, B. Duke, D. Gur, and D. M. Livermore. 1999.OXA-17, a further extended-spectrum variant of OXA-10 �-lactamase,isolated from Pseudomonas aeruginosa. Antimicrob. Agents Chemother.43:1362–1366.
38. Danel, F., L. M. Hall, D. Gur, and D. M. Livermore. 1995. OXA-14, anotherextended-spectrum variant of OXA-10 (PSE-2) �-lactamase from Pseudo-monas aeruginosa. Antimicrob. Agents Chemother. 39:1881–1884.
39. Danel, F., L. M. Hall, D. Gur, and D. M. Livermore. 1998. OXA-16, afurther extended-spectrum variant of OXA-10 �-lactamase, from twoPseudomonas aeruginosa isolates. Antimicrob. Agents Chemother. 42:3117–3122.
40. Da Silva, G. J., S. Quinteira, E. Bertolo, J. C. Sousa, L. Gallego, A. Duarte,and L. Peixe on behalf of the Portuguese Resistance Study Group. 2004.Long-term dissemination of an OXA-40 carbapenemase-producing Acin-etobacter baumannii clone in the Iberian Peninsula. J. Antimicrob. Che-mother. 54:255–258.
VOL. 54, 2010 MINIREVIEW 35

41. Doi, Y., L. Poirel, D. L. Paterson, and P. Nordmann. 2008. Characterizationof a naturally occurring class D �-lactamase from Achromobacter xylosoxi-dans. Antimicrob. Agents Chemother. 52:1952–1956.
42. Donald, H. M., W. Scaife, S. G. Amyes, and H. K. Young. 2000. Sequenceanalysis of ARI-1, a novel OXA �-lactamase, responsible for imipenemresistance in Acinetobacter baumannii 6B92. Antimicrob. Agents Che-mother. 44:196–199.
43. Dortet, L., I. Radu, V. Gautier, F. Blot, E. Chachaty, and G. Arlet. 2008.Intercontinental travels of patients and dissemination of plasmid-mediatedcarbapenemase KPC-3 associated with OXA-9 and TEM-1. J. Antimicrob.Chemother. 61:455–457.
44. Dubois, V., C. Arpin, C. Quentin, J. Texier-Maugein, L. Poirel, and P.Nordmann. 2003. Decreased susceptibility to cefepime in a clinical strain ofEscherichia coli related to plasmid- and integron-encoded OXA-30 �-lac-tamase. Antimicrob. Agents Chemother. 47:2380–2381.
45. Dubois, V., M. P. Parizano, C. Arpin, L. Coulange, M. C. Bezian, and C.Quentin. 2007. High genetic stability of integrons in clinical isolates ofShigella spp. of worldwide origin. Antimicrob. Agents Chemother. 51:1333–1340.
46. Empel, J., K. Filczak, A. Mrowka, W. Hryniewicz, D. M. Livermore, and M.Gniadkowski. 2007. Outbreak of Pseudomonas aeruginosa infections withPER-1 extended-spectrum �-lactamase in Warsaw, Poland: further evi-dence for an international clonal complex. J. Clin. Microbiol. 45:2829–2834.
47. Evans, B. A., S. Brown, A. Hamouda, J. Findlay, and S. G. Amyes. 2007.Eleven novel OXA-51-like enzymes from clinical isolates of Acinetobacterbaumannii. Clin. Microbiol. Infect. 13:1137–1138.
48. Feizabadi, M. M., B. Fathollahzadeh, M. Taherikalani, M. Rasoolinejad, N.Sadeghifard, M. Aligholi, S. Soroush, and S. Mohammadi-Yegane. 2008.Antimicrobial susceptibility patterns and distribution of blaOXA genesamong Acinetobacter spp. isolated from patients at Tehran hospitals. Jpn.J. Infect. Dis. 61:274–278.
49. Figueiredo, S., L. Poirel, J. Croize, C. Recule, and P. Nordmann. 2009. Invivo selection of reduced susceptibility to carbapenems in Acinetobacterbaumannii related to ISAba1-mediated overexpression of the naturalblaOXA-66 oxacillinase gene. Antimicrob. Agents Chemother. 53:2657–2659.
50. Figueiredo, S., L. Poirel, A. Papa, V. Koulourida, and P. Nordmann. 29June 2009. Overexpression of the naturally occurring blaOXA-51 gene inAcinetobacter baumannii mediated by novel insertion sequence ISAba9.Antimicrob. Agents Chemother. doi:10.1128/AAC.00292-09.
51. Figueiredo, S., L. Poirel, H. Seifert, and P. Nordmann. 2008. Abstr. 48thIntersci. Conf. Antimicrob. Agents Chemother., abstr. C1-1038.
52. Franceschini, N., L. Boschi, S. Pollini, R. Herman, M. Perilli, M. Galleni,J. M. Frere, G. Amicosante, and G. M. Rossolini. 2001. Characterization ofOXA-29 from Legionella (Fluoribacter) gormanii: molecular class D �-lac-tamase with unusual properties. Antimicrob. Agents Chemother. 45:3509–3516.
52a.Giannouli, M., F. Tomasone, A. Agodi, H. Vahaboglu, Z. Daoud, M. Triassi,A. Tsakris, and R. Zarrilli. 2009. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii strains in inteansive care units of multipleMediterranean hospitals. J. Antimicrob. Chemother. 63:828–830.
53. Giordano, A., P. Varesi, A. Bertini, L. Villa, A. M. Dionisi, M. Venditti, P.Carfagna, I. Luzzi, C. Mancini, and A. Carattoli. 2007. Outbreak of Acin-etobacter baumannii producing the carbapenem-hydrolyzing oxacillinaseOXA-58 in Rome, Italy. Microb. Drug Resist. 13:37–43.
54. Girlich, D., T. Naas, and P. Nordmann. 2004. Biochemical characterizationof the naturally occurring oxacillinase OXA-50 of Pseudomonas aeruginosa.Antimicrob. Agents Chemother. 48:2043–2048.
55. Girlich, D., T. Naas, and P. Nordmann. 2004. OXA-60, a chromosomal,inducible, and imipenem-hydrolyzing class D �-lactamase from Ralstoniapickettii. Antimicrob. Agents Chemother. 48:4217–4225.
56. Girlich, D., T. Naas, and P. Nordmann. 2006. Regulation of class D �-lac-tamase gene expression in Ralstonia pickettii. Microbiology 152:2661–2672.
57. Giuliani, F., J.-D. Docquier, M. L. Riccio, L. Pagani, and G. M. Rossolini.2005. OXA-46, a new class D �-lactamase of narrow substrate specificityencoded by a blaVIM-1-containing integron from a Pseudomonas aeruginosaclinical isolate. Antimicrob. Agents Chemother. 49:1973–1980.
58. Gombac, F., M. L. Riccio, G. M. Rossolini, C. Lagatolla, E. Tonin, C.Monti-Bragadin, A. Lavenia, and L. Dolzani. 2002. Molecular character-ization of integrons in epidemiologically unrelated clinical isolates of Acin-etobacter baumannii from Italian hospitals reveals a limited diversity of genecassette arrays. Antimicrob. Agents Chemother. 46:3665–3668.
59. Gulmez, D., N. Woodford, M.-F. Palepou, S. Mushtaq, G. Metan, Y. Yaku-pogullari, S. Kocagoz, O. Uzun, G. Hascelik, and D. M. Livermore. 2008.Carbapenem-resistant Escherichia coli and Klebsiella pneumoniae isolatesfrom Turkey with OXA-48-like carbapenemases and outer membrane pro-tein loss. Int. J. Antimicrob. Agents 31:523–526.
60. Hall, L. M., D. M. Livermore, D. Gur, M. Akova, and H. E. Akalin. 1993.OXA-11, an extended-spectrum variant of OXA-10 (PSE-2) �-lactamasefrom Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 37:1637–1644.
61. Hall, R. M., and C. M. Collis. 1995. Mobile gene cassettes and integrons:
capture and spread of genes by site-specific recombination. Mol. Microbiol.15:593–600.
62. Hanson, N. D., E. S. Moland, S. G. Hong, K. Propst, D. J. Novak, and S. J.Cavalieri. 2008. Surveillance of community-based reservoirs reveals thepresence of CTX-M, imported AmpC, and OXA-30 �-lactamases in urineisolates of Klebsiella pneumoniae and Escherichia coli in a U.S. community.Antimicrob. Agents Chemother. 52:3814–3816.
63. Hanson, N. D., E. S. Moland, A. Hossain, S. A. Neville, I. B. Gosbell, andK. S. Thomson. 2002. Unusual Salmonella enterica serotype Typhimuriumisolate producing CMY-7, SHV-9 and OXA-30 �-lactamases. J. Antimi-crob. Chemother. 49:1011–1014.
64. Heritier, C., A. Dubouix, L. Poirel, N. Marty, and P. Nordmann. 2005. Anosocomial outbreak of Acinetobacter baumannii isolates expressing thecarbapenem-hydrolysing oxacillinase OXA-58. J. Antimicrob. Chemother.55:115–118.
65. Heritier, C., L. Poirel, D. Aubert, and P. Nordmann. 2003. Genetic andfunctional analysis of the chromosome-encoded carbapenem-hydrolyzingoxacillinase OXA-40 of Acinetobacter baumannii. Antimicrob. Agents Che-mother. 47:268–273.
66. Heritier, C., L. Poirel, P. E. Fournier, J. M. Claverie, D. Raoult, and P.Nordmann. 2005. Characterization of the naturally occurring oxacillinase ofAcinetobacter baumannii. Antimicrob. Agents Chemother. 49:4174–4179.
67. Heritier, C., L. Poirel, T. Lambert, and P. Nordmann. 2005. Contributionof acquired carbapenem-hydrolyzing oxacillinases to carbapenem resis-tance in Acinetobacter baumannii. Antimicrob. Agents Chemother.49:3198–3202.
68. Heritier, C., L. Poirel, and P. Nordmann. 2004. Genetic and biochemicalcharacterization of a chromosome-encoded carbapenem-hydrolyzing Am-bler class D �-lactamase from Shewanella algae. Antimicrob. Agents Che-mother. 48:1670–1675.
68a.Higgins, P. G., L. Poirel, M. Lehmann, P. Nordmann, and H. Seifert. 2009.OXA-143, a novel carbapenem-hydrolyzing class D �-lactamase in Acineto-bacter baumannii. Antimicrob. Agents Chemother. 53:5035–5038.
69. Hujer, K. M., A. M. Hujer, E. A. Hulten, S. Bajaksouzian, J. M. Adams,C. J. Donskey, D. J. Ecker, C. Massire, M. W. Eshoo, R. Sampath, J. M.Thomson, P. N. Rather, D. W. Craft, J. T. Fishbain, A. J. Ewell, M. R.Jacobs, D. L. Paterson, and R. A. Bonomo. 2006. Analysis of antibioticresistance genes in multidrug-resistant Acinetobacter sp. isolates from mil-itary and civilian patients treated at the Walter Reed Army Medical Center.Antimicrob. Agents Chemother. 50:4114–4123.
70. Huovinen, P., S. Huovinen, and G. A. Jacoby. 1988. Sequence of PSE-2�-lactamase. Antimicrob. Agents Chemother. 32:134–136.
70a.Jacoby, G. A., and L. S. Munoz-Price. 2005. The new �-lactamases. N. Engl.J. Med. 352:380–391.
71. Jaurin, B., and T. Grundstrom. 1981. ampC cephalosporinase of Esche-richia coli K-12 has a different evolutionary origin from that of �-lactamasesof the penicillinase type. Proc. Natl. Acad. Sci. U. S. A. 78:4897–4901.
72. Joris, B., P. Ledent, O. Dideberg, E. Fonze, J. Lamotte-Brasseur, J. A.Kelly, J. M. Ghuysen, and J.-M. Frere. 1991. Comparison of the sequencesof class A �-lactamases and of the secondary structure elements of peni-cillin-recognizing proteins. Antimicrob. Agents Chemother. 35:2294–2301.
73. Keith, K. E., P. C. Oyston, B. Crossett, N. F. Fairweather, R. W. Titball,T. R. Walsh, and K. A. Brown. 2005. Functional characterization of OXA-57, a class D �-lactamase from Burkholderia pseudomallei. Antimicrob.Agents Chemother. 49:1639–1641.
74. Kim, J. W., S. T. Heo, J. S. Jin, C. H. Choi, Y. C. Lee, Y. G. Jeong, S.-J. Kim,and J. C. Lee. 2008. Characterization of Acinetobacter baumannii carryingblaOXA-23, blaPER-1 and armA in a Korean hospital. Clin. Microbiol. Infect.14:716–718.
75. Koh, T. H., L. H. Sng, G. C. Wang, L. Y. Hsu, and Y. Zhao. 2007. IMP-4 andOXA �-lactamases in Acinetobacter baumannii from Singapore. J. Antimi-crob. Chemother. 59:627–632.
76. Kong, K. F., S. R. Jayawardena, A. Del Puerto, L. Wiehlmann, U. Laabs, B.Tummler, and K. Mathee. 2005. Characterization of poxB, a chromosomal-encoded Pseudomonas aeruginosa oxacillinase. Gene 358:82–92.
77. Kratz, J., F. Schmidt, and B. Wiedemann. 1983. Transposition of a geneencoding OXA-2 �-lactamase. J. Gen. Microbiol. 129:2951–2957.
78. Lamotte-Brasseur, J., J. Knox, J. A. Kelly, P. Charlier, E. Fonze, O. Dide-berg, and J.-M. Frere. 1994. The structures and catalytic mechanisms ofactive-site serine �-lactamases. Biotechnol. Genet. Eng. Rev. 12:189–230.
79. Ledent, P., X. Raquet, B. Joris, J. Van Beeumen, and J.-M. Frere. 1993. Acomparative study of class-D �-lactamases. Biochem. J. 292:555–562.
80. Lee, K., M. N. Kim, T. Y. Choi, S. E. Cho, S. Lee, D. H. Whang, D. Yong,Y. Chong, N. Woodford, D. M. Livermore, and the KONSAR Group. 2009.Wide dissemination of OXA-type carbapenemases in clinical Acinetobacterspp. isolates from South Korea. Int. J. Antimicrob. Agents 33:520–524.
81. Libisch, B., L. Poirel, Z. Lepsanovic, V. Mirovic, B. Balogh, J. Paszti, Z.Hunyadi, A. Dobak, M. Fuzi, and P. Nordmann. 2008. Identification ofPER-1 extended-spectrum �-lactamase producing Pseudomonas aeruginosaclinical isolates of the international clonal complex CC11 from Hungaryand Serbia. FEMS Immunol. Med. Microbiol. 54:330–338.
36 MINIREVIEW ANTIMICROB. AGENTS CHEMOTHER.

82. Liebert, C. A., R. M. Hall, and A. O. Summers. 1999. Transposon Tn21,flagship of the floating genome. Microbiol. Mol. Biol. Rev. 63:507–522.
83. Livermore, D. M., and C. S. Jones. 1986. Characterization of NPS-1, a novelplasmid-mediated �-lactamase, from two Pseudomonas aeruginosa isolates.Antimicrob. Agents Chemother. 29:99–103.
84. Lolans, K., T. W. Rice, L. S. Munoz-Price, and J. P. Quinn. 2006. Multicityoutbreak of carbapenem-resistant Acinetobacter baumannii isolates produc-ing the carbapenemase OXA-40. Antimicrob. Agents Chemother. 50:2941–2945.
85. Lu, P. L., M. Doumith, D. M. Livermore, T. P. Chen, and N. Woodford.2009. Diversity of carbapenem resistance mechanisms in Acinetobacter bau-mannii from a Taiwan hospital: spread of plasmid-borne OXA-72 carbap-enemase. J. Antimicrob. Chemother. 63:641–647.
86. Mansour, W., L. Poirel, D. Bettaieb, O. Bouallegue, N. Boujaafar, and P.Nordmann. 2008. Dissemination of OXA-23-producing and carbapenem-resistant Acinetobacter baumannii in a University Hospital in Tunisia. Mi-crob. Drug Resist. 14:289–292.
87. Marque, S., L. Poirel, C. Heritier, S. Brisse, M. D. Blasco, R. Filip, G.Coman, T. Naas, and P. Nordmann. 2005. Regional occurrence of plasmid-mediated carbapenem-hydrolyzing oxacillinase OXA-58 in Acinetobacterspp. in Europe. J. Clin. Microbiol. 43:4885–4888.
88. Marti, S., J. Sanchez-Cespedes, M. D. Blasco, M. Ruiz, P. Espinal, V. Alba,F. Fernandez-Cuenca, A. Pascual, and J. Vila. 2008. Characterization of thecarbapenem-hydrolyzing oxacillinase OXA-58 in an Acinetobacter genospe-cies 3 clinical isolate. Antimicrob. Agents Chemother. 52:2966–2968.
89. Marti, S., J. Sanchez-Cespedes, M. D. Blasco, P. Espinal, M. Ruiz, V. Alba,and J. Vila. 2008. Characterization of the carbapenem-hydrolyzing oxacil-linase OXA-58 in an Acinetobacter phenon 6/ct13TU clinical isolate. Diagn.Microbiol. Infect. Dis. 61:468–470.
90. Matar, G. M., G. Cuzon, G. F. Araj, T. Naas, J. Corkill, M. M. Kattar, andP. Nordmann. 2008. Oxacillinase-mediated resistance to carbapenems inKlebsiella pneumoniae from Lebanon. Clin. Microbiol. Infect. 14:887–888.
91. Medeiros, A. A., M. Cohenford, and G. A. Jacoby. 1985. Five novel plasmid-determined �-lactamases. Antimicrob. Agents Chemother. 27:715–719.
92. Meric, M., M. Kasap, G. Gacar, F. Budak, D. Dundar, F. Kolayli, C.Eroglu, and H. Vahaboglu. 2008. Emergence and spread of carbapenem-resistant Acinetobacter baumannii in a tertiary care hospital in Turkey.FEMS Microbiol. Lett. 282:214–218.
93. Merkier, A. K., and D. Centron. 2006. blaOXA-51-type �-lactamase genes areubiquitous and vary within a strain in Acinetobacter baumannii. Int. J.Antimicrob. Agents 28:110–113.
94. Meziane-Cherif, D., T. Lambert, M. Dupechez, P. Courvalin, and M. Gali-mand. 2008. Genetic and biochemical characterization of OXA-63, a newclass D �-lactamase from Brachyspira pilosicoli BM4442. Antimicrob.Agents Chemother. 52:1264–1268.
95. Michael, G. B., M. Cardoso, and S. Schwarz. 2008. Molecular analysis ofmultiresistant porcine Salmonella enterica subsp. enterica serovar Bredeneyisolates from Southern Brazil: identification of resistance genes, integronsand a group II intron. Int. J. Antimicrob. Agents 32:120–129.
96. Mortimer-Jones, S. M., N. D. Phillips, T. La, R. Naresh, and D. J. Hamp-son. 2008. Penicillin resistance in the intestinal spirochaete Brachyspirapilosicoli associated with OXA-136 and OXA-137, two new variants of theclass D �-lactamase OXA-63. J. Med. Microbiol. 57:1122–1128.
97. Mossakowska, D., N. A. Ali, and J. W. Dale. 1989. Oxacillin-hydrolysing�-lactamases. A comparative analysis at nucleotide and amino acid se-quence levels. Eur. J. Biochem. 180:309–318.
98. Mugnier, P., I. Casin, A.-T. Bouthors, and E. Collatz. 1998. Novel OXA-10-derived extended-spectrum �-lactamases selected in vivo or in vitro.Antimicrob. Agents Chemother. 42:3113–3116.
99. Mugnier, P., I. Podglajen, F. W. Goldstein, and E. Collatz. 1998. Carbap-enems as inhibitors of OXA-13, a novel, integron-encoded �-lactamase inPseudomonas aeruginosa. Microbiology 144:1021–1031.
100. Mugnier, P. D., L. Poirel, and P. Nordmann. 2009. Functional analysis ofinsertion sequence ISAba1, responsible for genomic plasticity of Acineto-bacter baumannii. J. Bacteriol. 191:2414–2418.
101. Mugnier, P., L. Poirel, M. Pitout, and P. Nordmann. 2008. Carbapenem-resistant and OXA-23-producing Acinetobacter baumannii isolates in theUnited Arab Emirates. Clin. Microbiol. Infect. 14:879–882.
102. Mugnier, P. D., K. M. Bindayna, L. Poirel, and P. Nordmann. 2009. Di-versity of plasmid-mediated carbapenem-hydrolysing oxacillinases amongcarbapenem-resistant Acinetobacter baumannii isolates from Kingdom ofBahrain. J. Antimicrob. Chemother. 63:1071–1073.
103. Mulvey, M. R., D. A. Boyd, L. Baker, O. Mykytczuk, E. M. Reis, M. D.Asensi, D. P. Rodrigues, and L. K. Ng. 2004. Characterization of a Salmo-nella enterica serovar Agona strain harbouring a class 1 integron containingnovel OXA-type �-lactamase (blaOXA-53) and 6�-N-aminoglycoside acetyl-transferase genes [aac(6�)-I30]. J. Antimicrob. Chemother. 54:354–359.
104. Naas, T., M. Levy, C. Hirschauer, H. Marchandin, and P. Nordmann. 2005.Outbreak of carbapenem-resistant Acinetobacter baumannii producing thecarbapenemase OXA-23 in a tertiary care hospital of Papeete, FrenchPolynesia. J. Clin. Microbiol. 43:4826–4829.
105. Naas, T., F. Namdari, P. Bogaerts, T. D. Huang, Y. Glupczynski, and P.
Nordmann. 2008. Genetic structure associated with blaOXA-18 gene, encod-ing a clavulanic acid-inhibited extended-spectrum oxacillinase. Antimicrob.Agents Chemother. 52:3898–3904.
106. Naas, T., F. Namdari, H. Reglier-Poupet, C. Poyart, and P. Nordmann.2007. Panresistant extended-spectrum �-lactamase SHV-5-producing Acin-etobacter baumannii from New York City. J. Antimicrob. Chemother. 60:1174–1176.
107. Naas, T., and P. Nordmann. 1999. OXA-type �-lactamases. Curr. Pharm.Des. 5:865–879.
108. Naas, T., W. Sougakoff, A. Casetta, and P. Nordmann. 1998. Molecularcharacterization of OXA-20, a novel class D �-lactamase, and its integronfrom Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 42:2074–2083.
109. Navia, M. M., J. Ruiz, and J. Vila. 2002. Characterization of an integroncarrying a new class D �-lactamase (OXA-37) in Acinetobacter baumannii.Microb. Drug Resist. 8:261–265.
110. Nazic, H., L. Poirel, and P. Nordmann. 2005. Further identification ofplasmid-mediated quinolone resistance determinant in Enterobacteriaceaein Turkey. Antimicrob. Agents Chemother. 49:2146–2147.
111. Niumsup, P., and V. Wuthiekanun. 2002. Cloning of the class D �-lacta-mase gene from Burkholderia pseudomallei and studies on its expression inceftazidime-susceptible and -resistant strains. J. Antimicrob. Chemother.50:445–455.
112. Nordmann, P., L. Poirel, M. Kubina, A. Casetta, and T. Naas. 2000. Bio-chemical-genetic characterization and distribution of OXA-22, a chromo-somal and inducible class D �-lactamase from Ralstonia (Pseudomonas)pickettii. Antimicrob. Agents Chemother. 44:2201–2204.
113. Ouellette, M., L. Bissonnette, and P. H. Roy. 1987. Precise insertion ofantibiotic resistance determinants into Tn21-like transposons: nucleotidesequence of the OXA-1 �-lactamase gene. Proc. Natl. Acad. Sci. U. S. A.84:7378–7382.
114. Pai, H., and G. A. Jacoby. 2001. Sequences of the NPS-1 and TLE-1�-lactamase genes. Antimicrob. Agents Chemother. 45:2947–2948.
115. Paton, R., R. S. Miles, J. Hood, and S. G. B. Amyes. 1993. ARI-1: �-lacta-mase-mediated imipenem resistance in Acinetobacter baumannii. Int. J.Antimicrob. Agents 2:81–88.
116. Peixe, L. 1999. Emergence of carbapenem-hydrolyzing enzymes in Acineto-bacter baumannii clinical isolates. J. Clin. Microbiol. 37:2109–2110.
117. Peleg, A. Y., C. Franklin, L. J. Walters, J. M. Bell, and D. W. Spelman.2006. OXA-58 and IMP-4 carbapenem-hydrolyzing �-lactamases in anAcinetobacter junii blood culture isolate from Australia. Antimicrob. AgentsChemother. 50:399–400.
118. Philippon, L. N., T. Naas, A.-T. Bouthors, V. Barakett, and P. Nordmann.1997. OXA-18, a class D clavulanic acid-inhibited extended-spectrum �-lac-tamase from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 41:2188–2195.
119. Plante, I., D. Centron, and P. H. Roy. 2003. Direct sequencing and PCRmapping of integrons reveals multiple class 1 integrons in the multiresistantstrain Enterobacter cloacae. FEMS Microbiol. Lett. 221:59–62.
120. Ploy, M. C., P. Courvalin, and T. Lambert. 1998. Characterization of In40of Enterobacter aerogenes BM2688, a class 1 integron with two new genecassettes, cmlA2 and qacF. Antimicrob. Agents Chemother. 42:2557–2563.
121. Ploy, M. C., F. Denis, P. Courvalin, and T. Lambert. 2000. Molecularcharacterization of integrons in Acinetobacter baumannii: description of ahybrid class 2 integron. Antimicrob. Agents Chemother. 44:2684–2688.
122. Poirel, L., L. Cabanne, L. Collet, and P. Nordmann. 2006. Class II trans-poson-borne structure harboring metallo-�-lactamase gene blaVIM-2 inPseudomonas putida. Antimicrob. Agents Chemother. 50:2889–2891.
123. Poirel, L., S. Figueiredo, V. Cattoir, A. Carattoli, and P. Nordmann. 2008.Acinetobacter radioresistens as a silent source of carbapenem resistance forAcinetobacter spp. Antimicrob. Agents Chemother. 52:1252–1256.
124. Poirel, L., P. Gerome, C. De Champs, J. Stephanazzi, T. Naas, and P.Nordmann. 2002. Integron-located oxa-32 gene cassette encoding an ex-tended-spectrum variant of OXA-2 �-lactamase from Pseudomonas aerugi-nosa. Antimicrob. Agents Chemother. 46:566–569.
125. Poirel, L., D. Girlich, T. Naas, and P. Nordmann. 2001. OXA-28, anextended-spectrum variant of OXA-10 �-lactamase from Pseudomonasaeruginosa and its plasmid- and integron-located gene. Antimicrob. AgentsChemother. 45:447–453.
126. Poirel, L., C. Heritier, and P. Nordmann. 2004. Chromosome-encodedAmbler class D �-lactamase of Shewanella oneidensis as a progenitor ofcarbapenem-hydrolyzing oxacillinase. Antimicrob. Agents Chemother. 48:348–351.
127. Poirel, L., C. Heritier, V. Tolun, and P. Nordmann. 2004. Emergence ofoxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. An-timicrob. Agents Chemother. 48:15–22.
128. Poirel, L., E. Lebessi, C. Heritier, A. Patsoura, M. Foustoukou, and P.Nordmann. 2006. Nosocomial spread of OXA-58-positive carbapenem-resistant Acinetobacter baumannii isolates in a pediatric hospital in Greece.Clin. Micobiol. Infect. 12:1138–1140.
129. Poirel, L., W. Mansour, O. Bouallegue, and P. Nordmann. 2008. Carbap-enem-resistant Acinetobacter baumannii isolates from Tunisia producing
VOL. 54, 2010 MINIREVIEW 37

the OXA-58-like carbapenem-hydrolyzing oxacillinase OXA-97. Antimi-crob. Agents Chemother. 52:1613–1617.
130. Poirel, L., S. Marque, C. Heritier, C. Segonds, G. Chabanon, and P. Nor-dmann. 2005. OXA-58, a novel class D �-lactamase involved in resistanceto carbapenems in Acinetobacter baumannii. Antimicrob. Agents Che-mother. 49:202–208.
131. Poirel, L., J. D. Pitout, and P. Nordmann. 2007. Carbapenemases: molec-ular diversity and clinical consequences. Future Microbiol. 2:501–512.
132. Poirel, L., and P. Nordmann. 2006. Genetic structures at the origin ofacquisition and expression of the carbapenem-hydrolyzing oxacillinase geneblaOXA-58 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 50:1442–1448.
133. Poirel, L., and P. Nordmann. 2006. Carbapenem resistance in Acinetobacterbaumannii: mechanisms and epidemiology. Clin. Microbiol. Infect. 12:826–836.
134. Poirel, L., G. F. Weldhagen, C. De Champs, and P. Nordmann. 2002. Anosocomial outbreak of Pseudomonas aeruginosa isolates expressing theextended-spectrum �-lactamase GES-2 in South Africa. J. Antimicrob.Chemother. 49:561–565.
135. Power, P., M. Galleni, J. Di Conza, J. A. Ayala, and G. Gutkind. 2005.Description of In116, the first blaCTX-M-2-containing complex class 1 inte-gron found in Morganella morganii isolates from Buenos Aires, Argentina.J. Antimicrob. Chemother. 55:461–465.
136. Qi, C., M. Malczynski, M. Parker, and M. H. Scheetz. 2008. Characteriza-tion of genetic diversity of carbapenem-resistant Acinetobacter baumanniiclinical strains collected from 2004 to 2007. J. Clin. Microbiol. 46:1106–1109.
137. Queenan, A. M., and K. Bush. 2007. Carbapenemases: the versatile �-lac-tamases. Clin. Microbiol. Rev. 20:440–458.
138. Quinteira, S., F. Grosso, H. Ramos, and L. Peixe. 2007. Molecular epide-miology of imipenem-resistant Acinetobacter haemolyticus and Acineto-bacter baumannii isolates carrying plasmid-mediated OXA-40 from a Por-tuguese hospital. Antimicrob. Agents Chemother. 51:3465–3466.
139. Rasmussen, B. A., D. Keeney, Y. Yang, and K. Bush. 1994. Cloning andexpression of a cloxacillin-hydrolyzing enzyme and a cephalosporinase fromAeromonas sobria AER 14M in Escherichia coli: requirement for an E. colichromosomal mutation for efficient expression of the class D enzyme.Antimicrob. Agents Chemother. 38:2078–2085.
140. Recchia, G. D., and R. M. Hall. 1995. Gene cassettes: a new class of mobileelement. Microbiology 141:3015–3027.
141. Ruiz, M., S. Marti, F. Fernandez-Cuenca, A. Pascual, and J. Vila. 2007.High prevalence of carbapenem-hydrolysing oxacillinases in epidemiologi-cally related and unrelated Acinetobacter baumannii clinical isolates inSpain. Clin. Microbiol. Infect. 13:1192–1198.
142. Sanschagrin, F., F. Couture, and R. C. Levesque. 1995. Primary structure ofOXA-3 and phylogeny of oxacillin-hydrolyzing class D �-lactamases. Anti-microb. Agents Chemother. 39:887–893.
143. Scaife, W., H.-K. Young, R. Paton, and S. G. B. Amyes. 1995. Transferableimipenem-resistance in Acinetobacter species from a clinical source. J. An-timicrob. Chemother. 36:585–586.
144. Schneider, I., A. M. Queenan, and A. Bauernfeind. 2006. Novel carbap-enem-hydrolyzing oxacillinase OXA-62 from Pandoraea pnomenusa. Anti-microb. Agents Chemother. 50:1330–1335.
145. Scoulica, E., A. Aransay, and Y. Tselentis. 1995. Molecular characterizationof the OXA-7 �-lactamase gene. Antimicrob. Agents Chemother. 39:1379–1382.
146. Sevillano, E., L. Gallego, and J. M. García-Lobo. 2009. First detection ofthe OXA-40 carbapenemase in P. aeruginosa isolates, located on a plasmidalso found in A. baumannii. Pathol. Biol. (Paris) 57:493–495.
147. Reference deleted.148. Siu, L. K., J. Y. Lo, K. Y. Yuen, P. Y. Chau, M. H. Ng, and P. L. Ho. 2000.
�-Lactamases in Shigella flexneri isolates from Hong Kong and Shanghaiand a novel OXA-1-like �-lactamase, OXA-30. Antimicrob. Agents Che-mother. 44:2034–2038.
149. Stoeva, T., P. G. Higgins, K. Bojkova, and H. Seifert. 2008. Clonal spreadof carbapenem-resistant OXA-23-positive Acinetobacter baumannii in aBulgarian university hospital. Clin. Microbiol. Infect. 14:723–727.
150. Tennstedt, T., R. Szczepanowski, S. Braun, A. Puehler, and A. Schlueter.2003. Occurrence of integron-associated resistance gene cassettes locatedon antibiotic resistance plasmids isolated from a wastewater treatmentplant. FEMS Microbiol. Ecol. 45:239–252.
151. Toleman, M. A., P. M. Bennett, and T. R. Walsh. 2006. ISCR elements:
novel gene capturing systems of the 21st century. Mol. Microbiol. Rev.70:296–316.
152. Toleman, M. A., P. M. Bennett, and T. R. Walsh. 2006. Common regionse.g. orf513 and antibiotic resistance: IS91-like elements evolving complexclass 1 integrons. J. Antimicrob. Chemother. 58:1–6.
153. Toleman, M. A., K. Rolston, R. N. Jones, and T. R. Walsh. 2003. Molecularand biochemical characterization of OXA-45, an extended-spectrum class2d� �-lactamase in Pseudomonas aeruginosa. Antimicrob. Agents Che-mother. 47:2859–2863.
154. Toleman, M. A., and T. R. Walsh. 2008. Evolution of the ISCR3 group ofISCR elements. Antimicrob. Agents Chemother. 52:3789–3791.
155. Tolmasky, M. E., and J. H. Crosa. 1993. Genetic organization of antibioticresistance genes (aac(6�)-Ib, aadA, and oxa9) in the multiresistance trans-poson Tn1331. Plasmid 29:31–40.
156. Tsakris, A., A. Ikonomidis, N. Spanakis, S. Pournaras, and K. Bethimouti.2007. Identification of a novel blaOXA-51 variant, blaOXA-92, from a clinicalisolate of Acinetobacter baumannii. Clin. Microbiol. Infect. 13:348–349.
157. Turton, J. F., M. E. Ward, N. Woodford, M. E. Kaufmann, R. Pike, D. M.Livermore, and T. L. Pitt. 2006. The role of ISAba1 in expression of OXAcarbapenemase genes in Acinetobacter baumannii. FEMS Microbiol. Lett.258:72–77.
158. Turton, J. F., N. Woodford, J. Glover, S. Yarde, M. E. Kaufmann, and T. L.Pitt. 2006. Identification of Acinetobacter baumannii by detection of theblaOXA-51-like carbapenemase gene intrinsic to this species. J. Clin. Micro-biol. 44:2974–2976.
159. Vahaboglu, H., F. Budak, M. Kasap, G. Gacar, S. Torol, A. Karadenizli, F.Kolayli, and C. Eroglu. 2006. High prevalence of OXA-51-type class D�-lactamases among ceftazidime-resistant clinical isolates of Acinetobacterspp.: co-existence with OXA-58 in multiple centres. J. Antimicrob. Che-mother. 58:537–542.
160. Valenzuela, J. K., L. Thomas, S. R. Partridge, T. van der Reijden, L.Dijkshoorn, and J. Iredell. 2007. Horizontal gene transfer in a polyclonaloutbreak of carbapenem-resistant Acinetobacter baumannii. J. Clin. Micro-biol. 45:453–460.
161. Vignoli, R., N. Cordeiro, V. Seija, F. Schelotto, M. Radice, J. Ayala, P.Power, and G. Gutkind. 2006. Genetic environment of CTX-M-2 in Kleb-siella pneumoniae isolates from hospitalized patients in Uruguay. Rev. Ar-gent. Microbiol. 38:84–88. (In Spanish.)
162. Vila, J., M. Navia, J. Ruiz, and C. Casals. 1997. Cloning and nucleotidesequence analysis of a gene encoding an OXA-derived �-lactamase inAcinetobacter baumannii. Antimicrob. Agents Chemother. 41:2757–2759.
163. Villegas, M. V., J. N. Kattan, A. Correa, K. Lolans, A. M. Guzman, N.Woodford, D. Livermore, and J. P. Quinn. 2007. Dissemination of Acineto-bacter baumannii clones with OXA-23 carbapenemase in Colombian hos-pitals. Antimicrob. Agents Chemother. 51:2001–2004.
164. Voha, C., J.-D. Docquier, G. M. Rossolini, and T. Fosse. 2006. Genetic andbiochemical characterization of FUS-1 (OXA-85), a narrow-spectrum classD �-lactamase from Fusobacterium nucleatum subsp. polymorphum. Anti-microb. Agents Chemother. 50:2673–2679.
165. Walsh, T. R., L. Hall, A. P. MacGowan, and P. M. Bennett. 1995. Sequenceanalysis of two chromosomally mediated inducible �-lactamases from Aero-monas sobria, strain 163a, one a class D penicillinase, the other an AmpCcephalosporinase. J. Antimicrob. Chemother. 36:41–52.
166. Wang, H., P. Guo, H. Sun, H. Wang, Q. Yang, M. Chen, Y. Xu, and Y. Zhu.2007. Molecular epidemiology of clinical isolates of carbapenem-resistantAcinetobacter spp. from Chinese hospitals. Antimicrob. Agents Chemother.51:4022–4028.
167. Woodford, N., M. J. Ellington, J. M. Coelho, J. F. Turton, M. E. Ward, S.Brown, S. G. Amyes, and D. M. Livermore. 2006. Multiplex PCR for genesencoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. An-timicrob. Agents 27:351–353.
168. Yang, Y., and K. Bush. 1995. Oxacillin hydrolysis by the LCR-1 �-lacta-mase. Antimicrob. Agents Chemother. 39:1209.
169. Zarrilli, R., R. Casillo, A. Di Popolo, M. F. Tripodi, M. Bagattini, S.Cuccurullo, V. Crivaro, E. Ragone, A. Mattei, N. Galdieri, M. Triassi, andR. Utili. 2007. Molecular epidemiology of a clonal outbreak of multidrug-resistant Acinetobacter baumannii in a university hospital in Italy. Clin.Microbiol. Infect. 13:481–489.
170. Zong, Z., X. Lu, J. K. Valenzuela, S. R. Partridge, and J. Iredell. 2008. Anoutbreak of carbapenem-resistant Acinetobacter baumannii producingOXA-23 carbapenemase in western China. Int. J. Antimicrob. Agents 31:50–54.
38 MINIREVIEW ANTIMICROB. AGENTS CHEMOTHER.




















