
Diversity and abundance of nematode-trapping fungi from decaying litter in terrestrial, freshwater...
20
ORIGINAL PAPER Diversity and abundance of nematode-trapping fungi from decaying litter in terrestrial, freshwater and mangrove habitats Aung Swe Rajesh Jeewon Stephen B. Pointing Kevin D. Hyde Received: 12 August 2008 / Accepted: 25 November 2008 / Published online: 11 December 2008 Ó Springer Science+Business Media B.V. 2008 Abstract Nematode-trapping fungi are ubiquitous in terrestrial habitats in dung, soils, litter and woody debris and they also occur in freshwater, but only one species has been found in marine habitats. The purpose of this study was therefore to investigate whether nematode-trapping fungi occurred in mangrove habitats. To achieve this we assessed the diversity of nematode-trapping fungi on decaying litter from mangroves, freshwater and terrestrial habitats (22 sites) in Hong Kong. Composite samples (n = 1,320) of decaying litter (wood and leaves) were examined and a total of 31 species of nematode-trapping fungi belonging to four genera, Arthrobotrys, Monacrosporium, and Dactylella were recorded. Twenty-nine species reported in this study are new records for Hong Kong and 16 species are new records from mangrove habitats worldwide. Nematode trapping fungi are therefore present in marine environments. Commonly encountered taxa were Arthro- botrys oligospora and Monacrosporium thaumasium which are abundant in all habitats. A. oligospora, M. thaumasium and Arthrobotrys musiformis were frequent (F [ 10%). Twenty-six species were rare (0.16–9.32%). Species richness and diversity was higher in terrestrial than in freshwater and mangrove habitats (ANOVA, P \ 0.001). A higher mean diversity was observed on decaying leaves as compared to decaying wood in all habitats (P \ 0.001). Based on Shannon diversity index, it was also observed that taxa charac- terized by adhesive nets were more frequent in all habitats. This can be explained by the fact that these taxa may have a better competitive saprotrophic ability which would allow them to compete favourably in nutrient limited environments. Abiotic factors that could be A. Swe (&) R. Jeewon S. B. Pointing Division of Microbiology, School of Biological Sciences, Faculty of Science, The University of Hong Kong, Pokfulam Road, Hong Kong SAR, People’s Republic of China e-mail: [email protected] K. D. Hyde International Fungal Research & Development Centre, The Research Institute of Resource Insects, Chinese Academy of Forestry, 650224 Kunming, People’s Republic of China K. D. Hyde School of Science, Mae Fah Luang University, Chiang Rai, Thailand 123 Biodivers Conserv (2009) 18:1695–1714 DOI 10.1007/s10531-008-9553-7
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Diversity and abundance of nematode-trapping fungi from decaying litter in terrestrial, freshwater...

ORI GIN AL PA PER
Diversity and abundance of nematode-trapping fungifrom decaying litter in terrestrial, freshwater andmangrove habitats
Aung Swe Æ Rajesh Jeewon Æ Stephen B. Pointing Æ Kevin D. Hyde
Received: 12 August 2008 / Accepted: 25 November 2008 / Published online: 11 December 2008� Springer Science+Business Media B.V. 2008
Abstract Nematode-trapping fungi are ubiquitous in terrestrial habitats in dung, soils,
litter and woody debris and they also occur in freshwater, but only one species has been
found in marine habitats. The purpose of this study was therefore to investigate whether
nematode-trapping fungi occurred in mangrove habitats. To achieve this we assessed the
diversity of nematode-trapping fungi on decaying litter from mangroves, freshwater and
terrestrial habitats (22 sites) in Hong Kong. Composite samples (n = 1,320) of decaying
litter (wood and leaves) were examined and a total of 31 species of nematode-trapping
fungi belonging to four genera, Arthrobotrys, Monacrosporium, and Dactylella were
recorded. Twenty-nine species reported in this study are new records for Hong Kong and
16 species are new records from mangrove habitats worldwide. Nematode trapping fungi
are therefore present in marine environments. Commonly encountered taxa were Arthro-botrys oligospora and Monacrosporium thaumasium which are abundant in all habitats. A.oligospora, M. thaumasium and Arthrobotrys musiformis were frequent (F [ 10%).
Twenty-six species were rare (0.16–9.32%). Species richness and diversity was higher in
terrestrial than in freshwater and mangrove habitats (ANOVA, P \ 0.001). A higher mean
diversity was observed on decaying leaves as compared to decaying wood in all habitats
(P \ 0.001). Based on Shannon diversity index, it was also observed that taxa charac-
terized by adhesive nets were more frequent in all habitats. This can be explained by the
fact that these taxa may have a better competitive saprotrophic ability which would allow
them to compete favourably in nutrient limited environments. Abiotic factors that could be
A. Swe (&) � R. Jeewon � S. B. PointingDivision of Microbiology, School of Biological Sciences, Faculty of Science, The Universityof Hong Kong, Pokfulam Road, Hong Kong SAR, People’s Republic of Chinae-mail: [email protected]
K. D. HydeInternational Fungal Research & Development Centre, The Research Institute of Resource Insects,Chinese Academy of Forestry, 650224 Kunming, People’s Republic of China
K. D. HydeSchool of Science, Mae Fah Luang University, Chiang Rai, Thailand
123
Biodivers Conserv (2009) 18:1695–1714DOI 10.1007/s10531-008-9553-7

linked to differences in species diversity between decaying wood and leaves are also
discussed.
Keywords Biodiversity � Ecology � Nematode-trapping fungi � Mangrove fungi �Marine fungi
Introduction
Nematode-trapping fungi are hyphomycetous predators of nematodes (Kerry 1987) and
have been used in agriculture and animal husbandry as biological control agents (Larsen
et al. 1997; Kerry 2000). Nematode-trapping fungi are ubiquitous in a variety of habitats
such as agricultural, horticultural and forest soils, but have rarely been reported from
aquatic and especially marine environments (Gray 1983; Persmark and Jansson 1997;
Jansson and Lopez-Llorca 2001). Currently 35 species of nematode-trapping fungi have
been recorded from aquatic environments (Ingold 1944; Peach 1950, 1952, 1954; Johnson
and Autery 1961; Anastasiou 1964; Hao et al. 2004), however, Arthrobotrys dactyloides is
the only a species that has been isolated from brackish water (Johnson and Autery 1961).
Nematodes are populous in marine and mangrove habitats (Tietjen and Alongi 1990;
Somerfield et al. 2002; Zhang and Zhou 2003; Chinnadurai and Fernando 2007). We might
therefore expect nematode-trapping fungi to be ubiquitous in saline habitats.
Thorn and Barron (1984) noted that wood-decomposing, nematode-trapping fungi
resemble carnivorous plants. The plants and fungi grow in nitrogen poor environments;
plants capture animals to support photosynthesis; whereas fungi may capture nematodes to
support saprotrophism. Pfister (1994, 1997) Pfister and Liftik (1995) showed that the
telemorphs of Arthrobotrys oligospora var. oligospora and certain other nematode-trap-
ping fungi (Orbilia species) are saprotrophs on dead wood and bark revealing a connection
between wood and nematode-trapping fungi. Similar findings have been reported (Webster
et al. 1998; Liu et al. 2005; Mo et al. 2005; Yu et al. 2007a, b) and it is probably that the
saprotrophic Orbilia species gain extra nutrition, especially nitrogen through trapping
nematodes (via their anamorphic stage). The teleomorph Orbilia, however, has not been
recorded from marine habitats and therefore other fungal genera may have taken over
this role.
Fungi are considered to be the most important agents involved in wood and leaf decay
in terrestrial, freshwater and mangroves habitats (Sin et al. 2002; Seena et al. 2008). They
release nutrients for other organisms to survive on these substrates (Hyde 1989, 1990;
Hyde and Lee 1995; Whitford 1996; Hyde et al. 1998; Wong et al. 1998; Boddy 2001).
Numerous studies have focused on fungal diversity of decomposing leaf and woody litter
(Hyde and Alias 2000; Tsui et al. 2000; Wai et al. 2001; Berg et al. 2002; Cai et al. 2003;
Ananda and Sridhar 2004; Gopal and Chauhan 2006; O’dor et al. 2006; Gulis et al. 2008;
Lonsdale et al. 2008). These studies, however, provided no information on the diversity of
nematode-trapping fungi. This is because detection of nematode-trapping fungi requires
specific techniques, thus they are usually overlooked by mycologists unless they are
nematode-trapping fungi specialists.
The objectives of this study were (1) to assess whether nematode trapping fungi occur in
marine habitats and (2) to investigate the biodiversity of nematode-trapping fungi in ter-
restrial, freshwater and mangrove habitats to establish if there were differences in species
composition in different substrates and habitats.
1696 Biodivers Conserv (2009) 18:1695–1714
123

Materials and methods
Location and study sites
In this study, 22 sites situated in Hong Kong with seasonal tropical climate were selected,
nine from terrestrial five from freshwater (lotic) and eight from mangrove habitats,
respectively (Fig. 1; Table 1). The terrestrial, freshwater and mangrove sites were in close
proximity. The collection time was between October and April in 2005.
Sampling design and process
A composite sampling method (Rohde 1976) was used. Ten sampling areas were randomly
selected at each site and 50 decaying leaves and 20 decaying wood samples were collected.
All the samples were placed individually in Zip-lock plastic bags and maintained at 4�C
before treatment. Each sample (50 leaves or 20 wood samples) were coarsely ground and
homogenously mixed with a blender. The slurry (3 g) as a composite sample was sprinkled
onto corn meal agar media (three replicates), containing 1% streptomycin to inhibit bac-
teria growth. Five hundred (±32, n = 3) healthy Panagrellus redivivus (free living
nematodes from soil) were added to each plate as nematode baits. Plates were sealed and
incubated for 4 weeks at room temperature (26�C). Following incubation plates were
observed under a dissecting microscope and the nematode-trapping fungi present were
Fig. 1 A map of Hong Kong showing the study sites in three habitats: T terrestrial (j), F freshwater ( ), Mmangrove (d). (Draft)
Biodivers Conserv (2009) 18:1695–1714 1697
123

recorded. A total of 1,320 composite samples were examined. A single spore was trans-
ferred to a new CMA plate with a sterilized toothpick. Then a plug (2 9 2 cm) of media
was removed adjacent to the spore to provide a well and plates were incubated on CMA at
26�C until the mycelium grew out into the well. Living nematodes were placed in the well
in order to stimulate fungi to form trapping devices. Types of trapping device were
checked under a dissecting microscope after 24 h. The isolation method is modified in this
study based on the method of Liu and Zhang (2003) and Hao et al. (2004). Taxa were
identified using the keys of Li et al. (2000) for Arthrobotrys and Monacrosporium and
Chen et al. (2007) for Dactylella.
Data analysis
Total numbers of taxa, frequency of occurrence of each species and abundance (total
occurrence of all taxa) were recorded and calculated for each sampling site at each habitat.
In order to quantify nematode-trapping fungi, we followed the method of Mo et al. (2006).
The individual number of a species was counted as one occurrence of a species if it was
isolated from any of the three replicates.
The species diversity of each sampling site was calculated using Shannon’s diversity
index, H0 (Shannon and Weaver 1963) and the Simpson index, D (Simpson 1949).
Table 1 One location and study sites of the study
Habitats SiteNo.
Location
Terrestrial T1 Tai Mo Shan Country Park, New Territories, 22�25 35.210 0 N 114�10 33.560 0 E
T2 Tai Mo Shan Country Park, New Territories, 22�250 24.900 0 N 114�100 25.110 0 E
T3 Tai Mo Shan Country Park, New Territories, 22�250 28.400 0 N 114�100 39.100 0 E
T4 Tai Lam Country Park, New Territories, 22�240 30.270 0 N 114�030 31.180 0 E
T5 Tai Lam Country Park, New Territories, 22�240 15.970 0 N 114�030 29.090 0 E
T6 Tai Po Kau Natural Reserve, New Territories, 22�250 30.720 0 N 114�100 37.460 0 E
T7 Tai Po Kau, New Territories, 22�240 42.320 0 N 114�070 00.730 0 E
T8 Lantau North Country Park, Mui Wo, Lantau, 22�160 39.570 0 N 113�590 45.000 0 E
T9 Lung Fu Shan Country Park, Hong Kong, 22�160 53.740 0 N 114�080 14.090 0 E
Freshwater FW1 Shing Mu Reservoir, New Territories, 22�230 51.890 0 N 114�020 48.780 0
FW2 Forest Stream, Tai Po Kau, New Territories 22�240 59.080 0 N 114�070 01.620 0 E
FW3 Tai Shing Stream, Tai Mo Shan Country Park, New Territories 22�230 34.010 0 N114�080 3.320 0 E
FW4 Lantau North Contry Park, Mui Wo, Lantau, 22�160 24.170 0 N 113�590 45.000 0 E
FW5 Forest Stream, Pat Sin Leng Country Park, 22�290 28.240 0 N 114�100 53.440 0
Mangroves M1 Mai Po Nature Reserve, Mai Po, New Territories, 22�290 11.000 0 N114�020 24.350 0 E
M2 Shuen Wang, Sai Kung, New Territories, 22�270 49.740 0 N 114�120 33.530 0 E
M3 Ting Kok, Sai Kung, New Territories, 22�280 08.160 0 N 114�130 01.970 0 E
M4 Ha Pak Na, Deep Bay, New Territories, 22�280 22.120 0 N 113�590 10.710 0 E
M5 Kei Ling Ha Lo Wai, Sai Kung, 22�240 49.320 0 N 114�160 21.470 0 E
M6 Sham Chung, Sai Kung, New Territories, 22�260 35.190 0 N 114�170 01.360 0 E
M7 Hoi Ha Wan, Sai Kung, New Territories, 22�280 10.580 0 N 114�200 07.340 0 E
M8 Pak Sha Wan, Sai Kung, New Territories, 22�220 02.910 0 N 114�150 38.330 0
Total 22
1698 Biodivers Conserv (2009) 18:1695–1714
123

H0 ¼ �Xn
i¼1
Pi loge Pi where, Pi ¼Ni
Nð1Þ
D ¼ �Xn
i¼1
P2i ð2Þ
where Ni is individual number of i species and N is individual number of all species: Pi is
the proportion of i species and n is the number of species at the site. The occurrence
frequencies of each species (F) were calculated based on total number of all species by
using following formula:
F ¼ Individual number of species
Individual number of all species� 100 ð3Þ
To compare the similarity of the species composition among different habitats,
Sørensen’s index of similarity (S0) was applied (Magurran 1988).
S0 ¼ 2C
Aþ Bð Þ ð4Þ
where A and B are the number of species at site A and B, respectively and C is the number
of species common to both collections.
Evenness indices were estimated to establish the closeness of equability of species
present (Gotelli and Colwell 2001). All statistical analyses were performed using SPSS
[Release 14.0.0 (2005), SPSS Inc., Chicago, IL, USA] and PRIMER [version 6 (2005),
Primer-E Ltd, Plymouth, UK]. Species diversity between habitats and substrates were
compared using two-way ANOVA followed by Tukey multiple comparison tests (HSD).
ANOSIM was also performed to compare the ranks of species similarities between habi-
tats. Statistical significance was defined at a = 0.05.
Results
Biodiversity of nematode-trapping fungi
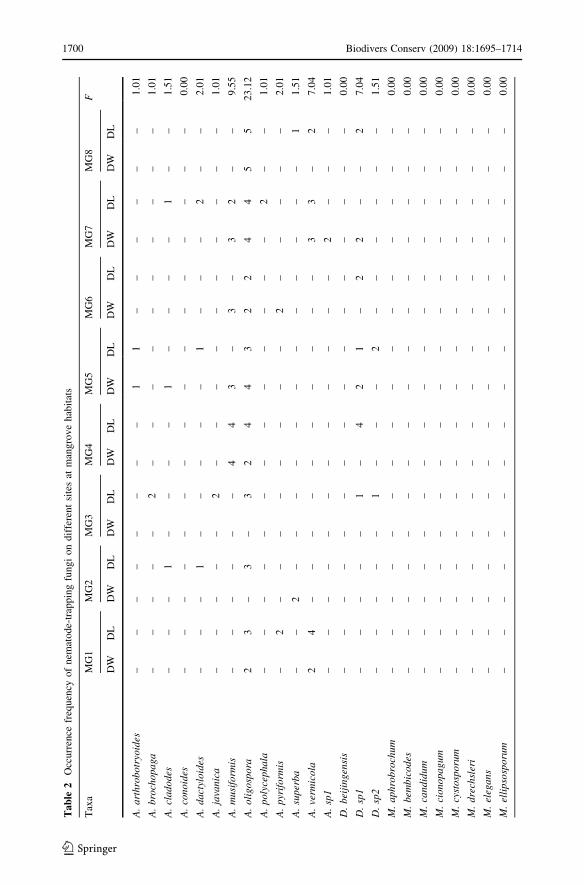
Mangrove
Seventeen nematode-trapping fungal species were recorded from 480 composite samples at
eight different of mangrove sites (Table 2). The most common species in mangroves were
Monacrosporium thaumasium (F = 24.62%), A. oligospora (F = 23.12%), and Mon-acrosporium eudermatum (F = 10.55%). A higher diversity was found on decaying leaves
at Site M3 (H0DL = 1.99; DDL = 0.853; SDW = 8), follow by M7 (H0DL = 1.932;
DDL = 0.833; SDL = 8) whereas the lowest diversity was observed at Site M2
(H0DL = 1.28; DDL = 0.694; SDL = 4) (Table 2; Fig. 2).
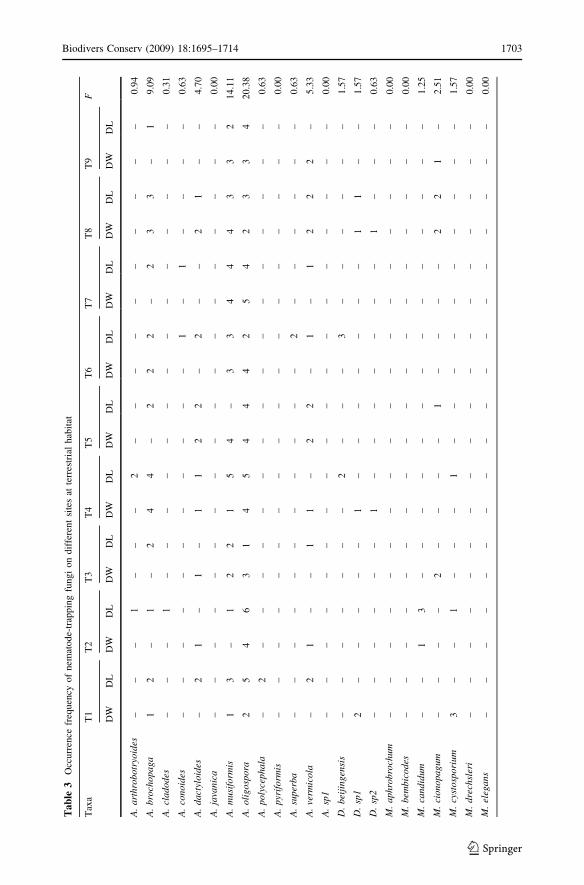
Terrestrial
Twenty-four nematode-trapping fungal species were recorded from 540 composite samples
collected at nine different terrestrial sites (Table 3). The most common species were
A. oligospora (F = 20.38%), Arthrobotrys musiformis (F = 14.11%), and M. thaumasium(F = 12.85%). Species diversity (H0 and D) varied between sample sites and substrates
Biodivers Conserv (2009) 18:1695–1714 1699
123
Ta
ble
2O
ccu
rren
cefr
equen
cyo
fn
emat
od
e-tr
app
ing
fun
gi
on
dif
fere
nt
site
sat
man
gro
ve
hab
itat
s
Tax
aM
G1
MG
2M
G3
MG
4M
G5
MG
6M
G7
MG
8F
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
A.
art
hro
botr
yoid
es–
––
––
––
–1
1–
––
––
–1
.01
A.
bro
cho
pa
ga
––
––
–2
––
––
––
––
––
1.0
1
A.
cla
do
des
––
–1
––
––
1–
––
–1
––
1.5
1
A.
con
oid
es–
––
––
––
––
––
––
––
–0
.00
A.
da
ctyl
oid
es–
––
1–
––
––
1–
––
2–
–2
.01
A.
java
nic
a–
––
––
2–
––
––
––
––
–1
.01
A.
mu
sifo
rmis
––
––
––
44
3–
3–
32
––
9.5
5
A.
oli
go
spo
ra2
3–
3–
32
44
32
24
45
52
3.1
2
A.
po
lyce
ph
ala
––
––
––
––
––
––
–2
––
1.0
1
A.
pyr
iform
is–
2–
––
––
––
–2
––
––
–2
.01
A.
sup
erba
––
2–
––
––
––
––
––
–1
1.5
1
A.
verm
ico
la2
4–
––
––
––
––
–3
3–
27
.04
A.
sp1
––
––
––
––
––
––
2–
––
1.0
1
D.
bei
jin
gen
sis
––
––
––
––
––
––
––
––
0.0
0
D.
sp1
––
––
–1
–4
21
–2
2–
–2
7.0
4
D.
sp2
––
––
–1
––
–2
––
––
––
1.5
1
M.
ap
hro
bro
chu
m–
––
––
––
––
––
––
––
–0
.00
M.
bem
bic
od
es–
––
––
––
––
––
––
––
–0
.00
M.
can
did
um
––
––
––
––
––
––
––
––
0.0
0
M.
cio
nop
ag
um
––
––
––
––
––
––
––
––
0.0
0
M.
cyst
osp
oru
m–
––
––
––
––
––
––
––
–0
.00
M.
dre
chsl
eri
––
––
––
––
––
––
––
––
0.0
0
M.
eleg
ans
––
––
––
––
––
––
––
––
0.0
0
M.
elli
pso
spor
um
––
––
––
––
––
––
––
––
0.0
0
1700 Biodivers Conserv (2009) 18:1695–1714
123

Ta
ble
2co
nti
nu
ed
Tax
aM
G1
MG
2M
G3
MG
4M
G5
MG
6M
G7
MG
8F
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
M.
eud
erm
atu
m–
2–
–1
31
42
2–
–3
3–
–1
0.5
5
M.
gep
hyr
opagu
m–
––
––
––
––
––
––
––
–0
.00
M.
ha
pto
tylu
m–
––
––
––
––
––
––
––
–0
.00
M.
hu
isu
nia
na
24
––
–3
––
––
––
––
––
4.5
2
M.
meg
alo
spor
um
––
––
––
––
––
––
––
––
0.0
0
M.
mic
rosc
apho
ides
––
––
––
––
––
––
––
––
0.0
0
M.
thau
ma
siu
m2
41
24
4–
4–
33
46
73
22
4.6
2
N(i
ndiv
idual
no
of
all
spec
ies)
819
37
51
97
20
13
13
10
82
32
48
12
S(s
pec
ies
no
.re
cord
ed)
46
24
28
35
67
43
78
25
H0
(Shan
non–W
einer
index
)1.3
86
1.7
49
0.6
37
1.2
77
0.5
00
1.9
86
0.9
56
1.6
09
1.6
72
1.8
45
1.3
66
1.0
40
1.8
77
1.9
32
0.6
62
1.4
68
Sim
pso
n_
1-D
0.7
50
0.8
20
0.4
44
0.6
94
0.3
20
0.8
53
0.5
71
0.8
00
0.7
93
0.8
28
0.7
40
0.6
25
0.8
36
0.8
33
0.4
69
0.7
36
Ev
ennes
s_e^
H/S
1.0
00
0.9
59
0.9
45
0.8
97
0.8
25
0.9
11
0.8
67
1.0
00
0.8
87
0.9
04
0.9
80
0.9
43
0.9
33
0.8
63
0.9
69
0.8
68
A=
Art
hro
botr
ys,
D=
Da
ctyl
ella
,M
=M
on
acr
osp
ori
um
Biodivers Conserv (2009) 18:1695–1714 1701
123
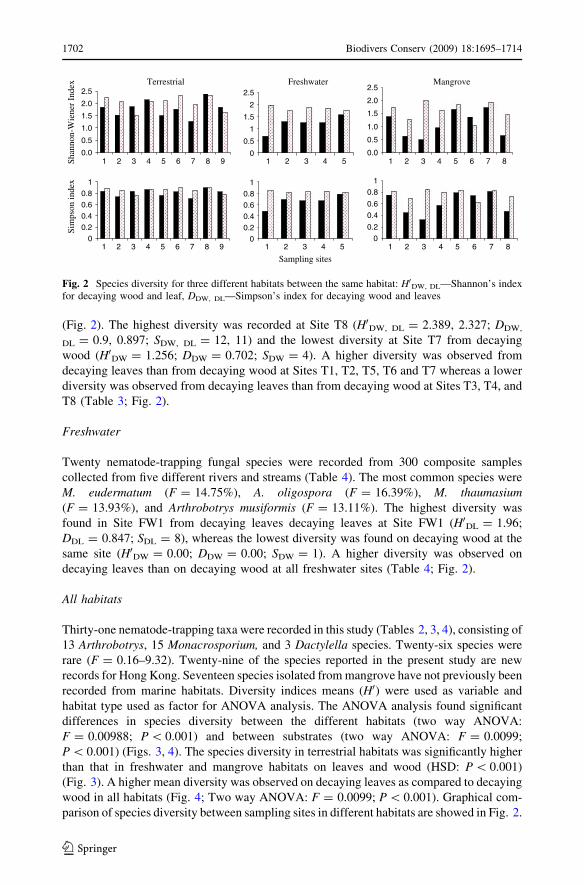
(Fig. 2). The highest diversity was recorded at Site T8 (H0DW, DL = 2.389, 2.327; DDW,
DL = 0.9, 0.897; SDW, DL = 12, 11) and the lowest diversity at Site T7 from decaying
wood (H0DW = 1.256; DDW = 0.702; SDW = 4). A higher diversity was observed from
decaying leaves than from decaying wood at Sites T1, T2, T5, T6 and T7 whereas a lower
diversity was observed from decaying leaves than from decaying wood at Sites T3, T4, and
T8 (Table 3; Fig. 2).
Freshwater
Twenty nematode-trapping fungal species were recorded from 300 composite samples
collected from five different rivers and streams (Table 4). The most common species were
M. eudermatum (F = 14.75%), A. oligospora (F = 16.39%), M. thaumasium(F = 13.93%), and Arthrobotrys musiformis (F = 13.11%). The highest diversity was
found in Site FW1 from decaying leaves decaying leaves at Site FW1 (H0DL = 1.96;
DDL = 0.847; SDL = 8), whereas the lowest diversity was found on decaying wood at the
same site (H0DW = 0.00; DDW = 0.00; SDW = 1). A higher diversity was observed on
decaying leaves than on decaying wood at all freshwater sites (Table 4; Fig. 2).
All habitats
Thirty-one nematode-trapping taxa were recorded in this study (Tables 2, 3, 4), consisting of
13 Arthrobotrys, 15 Monacrosporium, and 3 Dactylella species. Twenty-six species were
rare (F = 0.16–9.32). Twenty-nine of the species reported in the present study are new
records for Hong Kong. Seventeen species isolated from mangrove have not previously been
recorded from marine habitats. Diversity indices means (H0) were used as variable and
habitat type used as factor for ANOVA analysis. The ANOVA analysis found significant
differences in species diversity between the different habitats (two way ANOVA:
F = 0.00988; P \ 0.001) and between substrates (two way ANOVA: F = 0.0099;
P \ 0.001) (Figs. 3, 4). The species diversity in terrestrial habitats was significantly higher
than that in freshwater and mangrove habitats on leaves and wood (HSD: P \ 0.001)
(Fig. 3). A higher mean diversity was observed on decaying leaves as compared to decaying
wood in all habitats (Fig. 4; Two way ANOVA: F = 0.0099; P \ 0.001). Graphical com-
parison of species diversity between sampling sites in different habitats are showed in Fig. 2.
00.2
0.4
0.6
0.8
1
0
0.5
1
1.5
2
2.5
00.2
0.4
0.6
0.8
1
0.0
0.5
1.0
1.5
2.0
2.5
0
0.2
0.4
0.6
0.8
1
0.0
0.5
1.0
1.5
2.0
2.5
1 2 3 4 5 6 7 8 9
1 2 3 4 5
1 2 3 4 5
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 8 9Shan
non-
Wie
ner
Inde
x Si
mps
on in
dex
MangroveFreshwaterTerrestrial
Sampling sites
Fig. 2 Species diversity for three different habitats between the same habitat: H0DW, DL—Shannon’s indexfor decaying wood and leaf, DDW, DL—Simpson’s index for decaying wood and leaves
1702 Biodivers Conserv (2009) 18:1695–1714
123
Tab
le3
Occ
urr
ence
freq
uen
cyof
nem
atode-
trap
pin
gfu
ngi
on
dif
fere
nt
site
sat
terr
estr
ial
hab
itat
Tax
aT
1T
2T
3T
4T
5T
6T
7T
8T
9F
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
A.
art
hro
botr
yoid
es–
––
1–
––
2–
––
––
––
––
–0.9
4
A.
bro
chopaga
12
–1
–2
44
–2
22
–2
33
–1
9.0
9
A.
cladodes
––
–1
––
––
––
––
––
––
––
0.3
1
A.
conoid
es–
––
––
––
––
––
1–
1–
––
–0.6
3
A.
dact
yloid
es–
21
–1
–1
12
2–
2–
–2
1–
–4.7
0
A.
java
nic
a–
––
––
––
––
––
––
––
––
–0.0
0
A.
musi
form
is1
3–
12
21
54
–3
34
44
33
214.1
1
A.
oli
gosp
ora
25
46
31
45
44
42
54
23
34
20.3
8
A.
poly
cephala
–2
––
––
––
––
––
––
––
––
0.6
3
A.
pyr
iform
is–
––
––
––
––
––
––
––
––
–0.0
0
A.
super
ba
––
––
––
––
––
–2
––
––
––
0.6
3
A.
verm
icola
–2
1–
–1
1–
22
–1
–1
22
2–
5.3
3
A.
sp1
––
––
––
––
––
––
––
––
––
0.0
0
D.
bei
jingen
sis
––
––
––
–2
––
–3
––
––
––
1.5
7
D.
sp1
2–
––
––
1–
––
––
––
11
––
1.5
7
D.
sp2
––
––
––
1–
––
––
––
1–
––
0.6
3
M.
aphro
bro
chum
––
––
––
––
––
––
––
––
––
0.0
0
M.
bem
bic
odes
––
––
––
––
––
––
––
––
––
0.0
0
M.
candid
um
––
13
––
––
––
––
––
––
––
1.2
5
M.
cionopagum
––
––
2–
––
–1
––
––
22
1–
2.5
1
M.
cyst
osp
ori
um
3–
–1
––
–1
––
––
––
––
––
1.5
7
M.
dre
chsl
eri
––
––
––
––
––
––
––
––
––
0.0
0
M.
eleg
ans
––
––
––
––
––
––
––
––
––
0.0
0
Biodivers Conserv (2009) 18:1695–1714 1703
123

Ta
ble
3co
nti
nu
ed
Tax
aT
1T
2T
3T
4T
5T
6T
7T
8T
9F
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
DW
DL
M.
elli
pso
sporu
m1
–1
––
––
––
–2
2–
–2
2–
–3.1
3
M.
euder
matu
m–
3–
––
–2
––
13
31
1–
–1
–4.7
0
M.
gep
hyr
opagum
–3
–1
––
––
––
––
––
22
–2
3.1
3
M.
hapto
tylu
m–
––
––
––
––
3–
––
––
––
–0.9
4
M.
huis
unia
na
––
–3
–3
33
–4
22
–2
––
1–
7.2
1
M.
meg
alo
sporu
m–
2–
–1
––
––
––
––
––
––
–0.9
4
M.
mic
rosc
aphoid
es–
––
–1
––
––
––
––
–1
1–
11.2
5
M.
thaum
asi
um
34
53
2–
33
12
––
53
12
31
12.8
5
N(i
ndiv
idual
no
of
all
spec
ies)
13
28
13
21
12
921
26
13
21
16
23
15
18
23
22
14
11
S(s
pec
ies
no.
reco
rded
)7
10
610
75
10
95
96
11
48
12
11
76
H0
(Shan
non–W
einer
index
)1.8
45
2.2
46
1.5
19
2.0
62
1.8
64
1.5
23
2.1
37
2.0
66
1.4
99
2.0
95
1.7
54
2.3
44
1.2
65
1.9
37
2.3
89
2.3
27
1.8
34
1.6
42
Sim
pso
n_1-D
0.8
28
0.8
88
0.7
34
0.8
44
0.8
33
0.7
65
0.8
66
0.8
61
0.7
57
0.8
66
0.8
20
0.9
00
0.7
02
0.8
40
0.9
00
0.8
97
0.8
27
0.7
77
Even
nes
s_e^
H/S
0.9
04
0.9
45
0.7
62
0.7
86
0.9
21
0.9
17
0.8
47
0.8
77
0.8
95
0.9
03
0.9
63
0.9
48
0.8
86
0.8
67
0.9
09
0.9
31
0.8
94
0.8
61
A=
Art
hro
botr
ys,
D=
Da
ctyl
ella
,M
=M
on
acro
spo
rium
1704 Biodivers Conserv (2009) 18:1695–1714
123
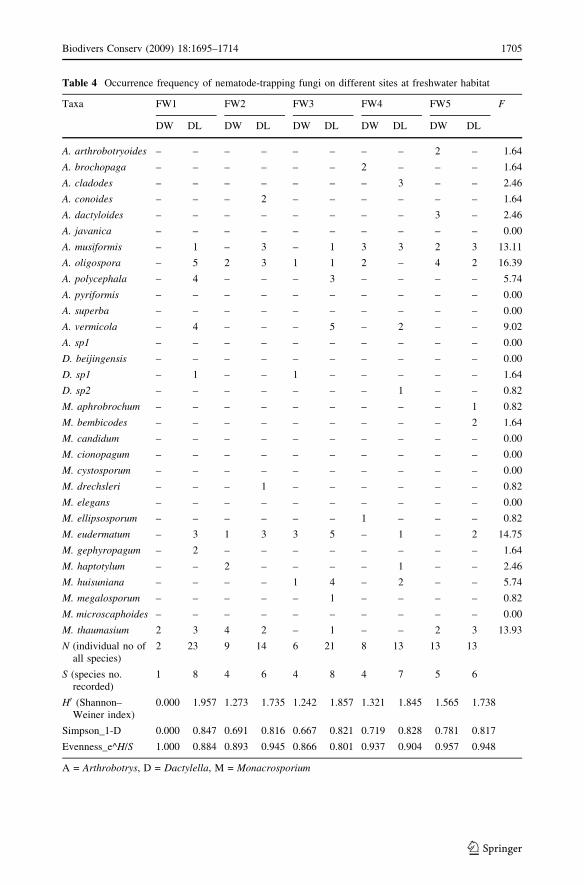
Table 4 Occurrence frequency of nematode-trapping fungi on different sites at freshwater habitat
Taxa FW1 FW2 FW3 FW4 FW5 F
DW DL DW DL DW DL DW DL DW DL
A. arthrobotryoides – – – – – – – – 2 – 1.64
A. brochopaga – – – – – – 2 – – – 1.64
A. cladodes – – – – – – – 3 – – 2.46
A. conoides – – – 2 – – – – – – 1.64
A. dactyloides – – – – – – – – 3 – 2.46
A. javanica – – – – – – – – – – 0.00
A. musiformis – 1 – 3 – 1 3 3 2 3 13.11
A. oligospora – 5 2 3 1 1 2 – 4 2 16.39
A. polycephala – 4 – – – 3 – – – – 5.74
A. pyriformis – – – – – – – – – – 0.00
A. superba – – – – – – – – – – 0.00
A. vermicola – 4 – – – 5 – 2 – – 9.02
A. sp1 – – – – – – – – – – 0.00
D. beijingensis – – – – – – – – – – 0.00
D. sp1 – 1 – – 1 – – – – – 1.64
D. sp2 – – – – – – – 1 – – 0.82
M. aphrobrochum – – – – – – – – – 1 0.82
M. bembicodes – – – – – – – – – 2 1.64
M. candidum – – – – – – – – – – 0.00
M. cionopagum – – – – – – – – – – 0.00
M. cystosporum – – – – – – – – – – 0.00
M. drechsleri – – – 1 – – – – – – 0.82
M. elegans – – – – – – – – – – 0.00
M. ellipsosporum – – – – – – 1 – – – 0.82
M. eudermatum – 3 1 3 3 5 – 1 – 2 14.75
M. gephyropagum – 2 – – – – – – – – 1.64
M. haptotylum – – 2 – – – – 1 – – 2.46
M. huisuniana – – – – 1 4 – 2 – – 5.74
M. megalosporum – – – – – 1 – – – – 0.82
M. microscaphoides – – – – – – – – – – 0.00
M. thaumasium 2 3 4 2 – 1 – – 2 3 13.93
N (individual no ofall species)
2 23 9 14 6 21 8 13 13 13
S (species no.recorded)
1 8 4 6 4 8 4 7 5 6
H0 (Shannon–Weiner index)
0.000 1.957 1.273 1.735 1.242 1.857 1.321 1.845 1.565 1.738
Simpson_1-D 0.000 0.847 0.691 0.816 0.667 0.821 0.719 0.828 0.781 0.817
Evenness_e^H/S 1.000 0.884 0.893 0.945 0.866 0.801 0.937 0.904 0.957 0.948
A = Arthrobotrys, D = Dactylella, M = Monacrosporium
Biodivers Conserv (2009) 18:1695–1714 1705
123
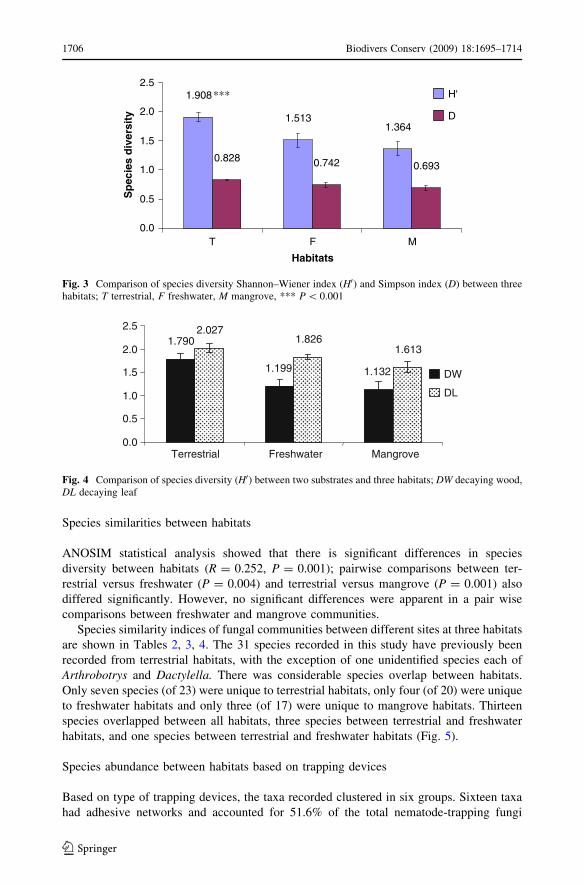
Species similarities between habitats
ANOSIM statistical analysis showed that there is significant differences in species
diversity between habitats (R = 0.252, P = 0.001); pairwise comparisons between ter-
restrial versus freshwater (P = 0.004) and terrestrial versus mangrove (P = 0.001) also
differed significantly. However, no significant differences were apparent in a pair wise
comparisons between freshwater and mangrove communities.
Species similarity indices of fungal communities between different sites at three habitats
are shown in Tables 2, 3, 4. The 31 species recorded in this study have previously been
recorded from terrestrial habitats, with the exception of one unidentified species each of
Arthrobotrys and Dactylella. There was considerable species overlap between habitats.
Only seven species (of 23) were unique to terrestrial habitats, only four (of 20) were unique
to freshwater habitats and only three (of 17) were unique to mangrove habitats. Thirteen
species overlapped between all habitats, three species between terrestrial and freshwater
habitats, and one species between terrestrial and freshwater habitats (Fig. 5).
Species abundance between habitats based on trapping devices
Based on type of trapping devices, the taxa recorded clustered in six groups. Sixteen taxa
had adhesive networks and accounted for 51.6% of the total nematode-trapping fungi
1.908
1.5131.364
0.828 0.742 0.693
0.0
0.5
1.0
1.5
2.0
2.5
T F M
Habitats
Sp
ecie
s d
iver
sity
H'
D
***
Fig. 3 Comparison of species diversity Shannon–Wiener index (H0) and Simpson index (D) between threehabitats; T terrestrial, F freshwater, M mangrove, *** P \ 0.001
1.790
1.199 1.132
2.0271.826
1.613
0.0
0.5
1.0
1.5
2.0
2.5
Terrestrial Freshwater Mangrove
DW
DL
Fig. 4 Comparison of species diversity (H0) between two substrates and three habitats; DW decaying wood,DL decaying leaf
1706 Biodivers Conserv (2009) 18:1695–1714
123
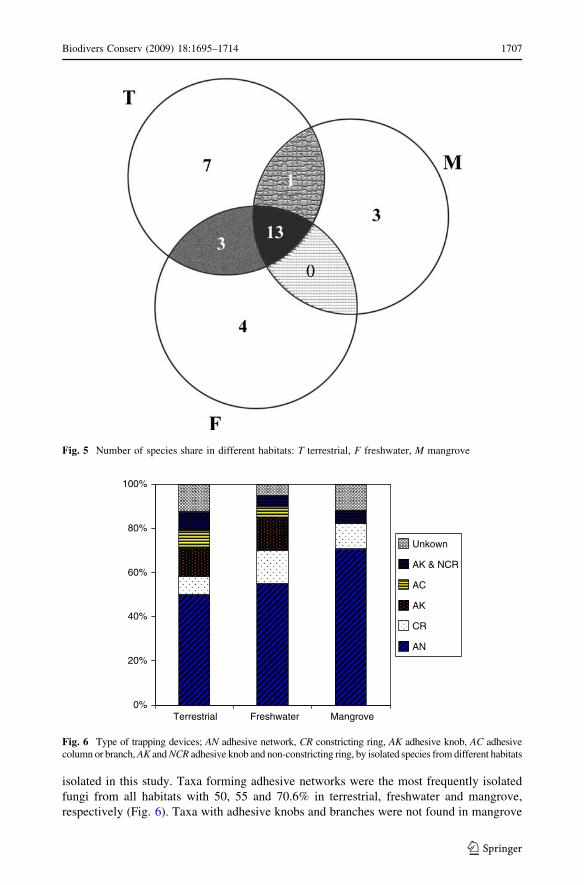
isolated in this study. Taxa forming adhesive networks were the most frequently isolated
fungi from all habitats with 50, 55 and 70.6% in terrestrial, freshwater and mangrove,
respectively (Fig. 6). Taxa with adhesive knobs and branches were not found in mangrove
0%
20%
40%
60%
80%
100%
Terrestrial Freshwater Mangrove
Unkown
AK & NCR
AC
AK
CR
AN
Fig. 6 Type of trapping devices; AN adhesive network, CR constricting ring, AK adhesive knob, AC adhesivecolumn or branch, AK and NCR adhesive knob and non-constricting ring, by isolated species from different habitats
Fig. 5 Number of species share in different habitats: T terrestrial, F freshwater, M mangrove
Biodivers Conserv (2009) 18:1695–1714 1707
123

habitats, whilst all types of trapping devices were isolated from terrestrial and freshwater
habitats.
Discussion
Do nematode trapping fungi occur in marine habitats?
Considering the high diversity and large numbers of nematodes that have been shown to
occur in marine and mangrove habitats (Bongers and Ferris 1999; Lambshead and Boucher
2003), it is surprising that only one species of nematode trapping fungi, A. dactyloides, has
been recorded. It is not clear whether this is due to the fact that saline habitats have never
been studied for nematode-trapping fungi or because they do not exist in nature. The main
purpose of this study was therefore to establish whether nematode-trapping fungi occurred
in marine habitats, and if they did, whether they were typical of or identical to the
terrestrial and freshwater species, or comprised a completely different group.
This study revealed 17 nematode-trapping fungal species from the mangrove habitat.
All species could be accommodated in previously described species, however, their
characteristics differed to a lesser or greater extent and none were identical. This variation
in characters will be discussed in next paper. The nematode-trapping fungi were found to
be typical of the freshwater and terrestrial species found in Hong Kong. One species was
new to science and an unusual hyphomycete which we could not place in any genus (not
shown here) appeared to have sticky knob-like nematode trapping spores.
Although no Orbilia species have been recorded in marine habitats we confirm here that
Orbilia anamorphs are the dominant nematode trapping fungi found in marine environ-
ments. It is interesting that the teleomorphs have not been found in mangroves in nature.
Hyde made extensive collections in mangroves systems in Brunei, Indonesia and the
Seychelles and never found any Orbilia species (Hyde 1988, 1990; Hyde and Goh 1998). It
would be interesting to establish the reason for this.
Comparison of nematode trapping fungi with previous studies
Communities of nematode-trapping fungi among terrestrial, freshwater and mangrove
habitats and substrates in Hong Kong have been compared. There is no similar published
study that we are aware of in any other country, thus direct comparisons with other studies
cannot be made. We can, however, compare our results with those from other specific
habitats. Nematode-trapping fungi were abundant in all the habitats examined, with over
65% of the samples containing nematode-trapping fungi. With the exception of two taxa, a
unidentified species and Arthrobotrys conoides (Poon and Hyde 1998), nematode-trapping
fungal species recorded in this study have not been recorded from Hong Kong. In a survey
of nematode-trapping of lead polluted soil in Kunming, China, Mo et al. (2006) recorded
28 taxa, the most common species being M. thaumasium, A. oligospora and Monacros-porium phymatopagum. It is interesting to note that M. phymatopagum was absent in Hong
Kong, although this species was common in Kunming. In similar survey on diversity of
nematode-trapping fungi from freshwater in Kunming, China, Hao et al. (2005) recorded
35 nematode-trapping species all of which have previously been recorded from terrestrial
habitats. Twenty of the nematode-trapping species isolated from freshwater habitats in this
study are the same as those reported from freshwater in Kunming (Hao et al. 2005). The 16
nematode-trapping species recorded from mangroves in Hong Kong are new records for
1708 Biodivers Conserv (2009) 18:1695–1714
123

mangrove habitats worldwide. However, these species have previously been reported from
terrestrial and freshwater habitats (Peterson and Katznelson 1964; Gray 1983; Jaffee et al.
1996; Elshafie et al. 2003; Hao et al. 2005; Mo et al. 2006).
Many fungal genera (e.g. Anthostomella, Aniptodera, Didymella, Massarina, Lophios-toma, Phomatospora, Saccardoella and Savoryella have species reported from freshwater,
marine and terrestrial habitats Vijaykrishna et al. 2006). However, the species are mostly
unique to terrestrial, freshwater or marine habitats. Aniptodera chesapeakensis and Sa-voryella lignicola are exception as they are know from freshwater and marine habitats
(Alias and Jones 2000; Cai et al. 2002; Tsui et al. 2003). In this study, 13 species of
nematode-trapping fungi were overlapped in terrestrial, freshwater and mangrove habitats
of Hong Kong. Cai et al. (2006) noted that fungal species identified from bamboo culms
submerged in freshwater overlapped with those on terrestrial bamboo culms samples—
however, the overlap was low (18.3%). Vijaykrishna et al. (2006) proposed that freshwater
and marine lignicolous ascomcyetes had evolved from terrestrial fungi. There are little to
no distinct morphologically differences between the species of genera Massarina and
Lophiostoma that occur in different habitats (Vijaykrishna et al. 2006). Byrne and Jones
(1975) also pointed out that the conidia carried from freshwater regions to brackish water
may also be able to germinate and colonize the leaf detritus in the brackish water.
Additionally, our observation suggests that the ability of a fungal species to survive and
propagate in wider ecological-niches is a considerable advantageous trait. It is, never-
theless, equally probable that different populations of a fungal species are restricted to
different ecological niches and habitats.
Fungal diversity and similarities between habitats
Biotic and abiotic factors (e.g. altitude, habitat, soil moisture, pH, and nematode density)
affect the occurrence and species diversity of terrestrial nematode-trapping fungi (Gray
1987). In this study we observed that species diversity in terrestrial habitats was signifi-
cantly higher than in freshwater and mangrove habitats, whereas no significant differences
in species diversity were observed between freshwater and mangrove habitats. Shearer
et al. (2007) stated that there is lower diversity of fungi in general in freshwater or marine
habitats as compared to terrestrial habitats. This is thought to be because (a) basidiomy-
cetes, zygomycetes and lichens are uncommon in aquatic habitats (b) of low plant species
diversity in aquatic habitats, and (c) of physiological constraints of growth when sub-
merged in water; especially seawater (Shearer et al. 2007). Some studies, however, have
reported contrasting results; a higher species richness was found on submerged decaying
bamboo culms as compared to terrestrial culms (Cai et al. 2006). Similarly higher diversity
was reported from submerged versus terrestrial wood samples (Fryar et al. 2004).
Accounting for differences in species diversity in decaying wood and leaves
The species diversity (H0) values clearly show that decaying leaves support a significantly
higher nematode-trapping fungal species diversity than decaying wood in all habitats
(Fig. 4). Coarse woody debris provides vital macro habitats for a variety of organisms,
such as fungi, bryophytes, lichens, invertebrates, and amphibians (Maser and Trappe 1984;
Harmon et al. 1986; Esseen et al. 1997; O’dor and Standovar 2002). Generally with
decaying microorganisms, there is lower diversity in larger substrates than smaller sub-
strates due to the high C/N ratio and available oxygen. Moreover, the abundance of
nematodes in the litter is significantly affected by litter quality (low quality: high C/N ratio,
Biodivers Conserv (2009) 18:1695–1714 1709
123

high quality: low C/N ratio); nematode density was significantly higher in the high quality
litter than low quality litter (Ilieva-Makulec et al. 2006). A possible explanation could be
that leaves break down faster than wood because of favourable C/N ratios and relatively
rapid release of nutrients. This would enhance bacteria growth which in turn will feed
bacterivorous nematodes, which eventually feed the nematode-trapping fungi.
When considering species to surface area relationships, we might expect that if a
substrate has a larger surface area it is more likely to harbour a greater number of species.
However, a large item of course woody debris may provide more ‘‘space’’ for larger
individuals to grow, rather than providing more space for a wider range of species
(Lonsdale et al. 2008). Heilmann-Clausen and Christensen (2004) also found that although
the number of wood-inhabiting fungal species increases with coarse woody debris size, the
number of fungi per unit decreases. These findings are supported by our results. Moreover,
annual fungal mass production in wood and leaves per basis has been calculated and
compared by Gulis et al. (2008). The annual fungal mass production from wood was 4.3–
5.5 g cm-2 per year, which is considerably lower than that from leaves (15.8–33.1 g cm-2
per year). Fungal mass production and microbial respiration per gram of detrital C were 3–
13 times lower in wood than in leaves. Fungal growth rates were also higher on leaves than
on wood (Gulis et al. 2008). It is, however, unclear whether these factors correlate with
fungal species members. Nevertheless, species diversity data does not support the higher
species to larger surface area assumption. There should also be different nematode-trap-
ping species communities on the two substrates because the composition of fungal
communities largely depends on environmental factors such as nutrients, C/N ratio, lignin
concentration, size and thickness of substrates, available oxygen, residence time in nature
and biotic factors such as nematode density (Gray 1987; Das et al. 2007; Li et al. 2007;
Lonsdale et al. 2008; Wakelin et al. 2008). The fungal communities on leaves and wood
should therefore differ.
Why adhesive trappers are more abundant than others?
Nematode-trapping fungal species forming adhesive networks were isolated more fre-
quently in all habitats than other trapping types and agrees with previous findings (Gray
1987; Persmark and Jansson 1997; Hao et al. 2005; Mo et al. 2006). Although several
environmental factors may affect the occurrence and diversity of fungi, the frequent
occurrence of adhesive networks might be explained by growth rate and competitive
saprotrophic ability of the fungi. Nematode-trapping fungi with adhesive networks grow
most rapidly, while those with other trapping devices grow 2–3 times more slowly (Cooke
1963). Network trapping fungi appeared to be able to compete successfully with the soil
mycota, whereas those with constricting rings are less competitive (Cooke 1963). Adhesive
network trapping fungi are largely independent of soil fertility and can be found in very
low nutrients soil (Gray 1988). It seems likely that adhesive network trapping fungi have a
good saprotrophic ability and a rapid growth rate which will allow them to compete
favourably with other organisms for limited nutrients. However, whether those abilities are
correlated with a predacious ability in nature is unknown.
Is fungal diversity data technique dependent?
In total 20 nematode-trapping fungal taxa were recorded from freshwater habitats in this
study. Tsui et al. (2000) isolated 92 anamorphic hyphomycetes on decaying wood from
freshwater habitats in Hong Kong. Ho et al. (2002) also recorded 51 hyphomycetes from
1710 Biodivers Conserv (2009) 18:1695–1714
123

the Tai Po Kau forest stream. It is interesting to note that, no nematode-trapping fungi were
isolated from decaying wood in these studies considering that our sample sites cover the
Shing Mun Reservoir stream and Tai Po Kau forest stream. Moreover, numerous studies
have focused on fungal diversity of decomposing leaf and woody litter (Tsui et al. 2000;
Wai et al. 2001; Cai et al. 2003; Ananda and Sridhar 2004; Gopal and Chauhan 2006;
O’dor et al. 2006; Gulis et al. 2008; Lonsdale et al. 2008; Kodsueb et al. 2008; Duong et al.
2008). These studies, however, provided no information on the diversity of nematode-
trapping fungi. These results clearly support the observation that the type of fungi collected
from nature is technique dependent (Shearer et al. 2007).
Nematode-trapping fungi are usually difficult to identify even to genus level without
having information on the type of trapping device. It is a serious flaw of the classical
method, especially in quantifying the frequency of occurrence for nematode-trapping
fungi. Future studies should combine classical methods with other methods (especially
environmental molecular sampling) to measure the taxonomic diversity of nematode-
trapping fungi, e.g. denaturing gradient gel electrophoresis (Anderson and Cairney 2004;
Duong et al. 2006; Kataoka et al. 2008) are encouraged. Now that we have established that
nematode-trapping fungi can occur in mangroves the study should be expanded to involve
other marine habitats (sandy beaches, oceans, deep sea). We also need to establish the role
of NTF in marine habitats as we have found that their ability to produce trapping devices
diminishes with increasing salinity in vitro.
Acknowledgments We would like to thank Dr. Lau Chun Pong for his generosity in sharing knowledge instatistics. Helen Leung is thanked for technical assistance. The University of Hong Kong is thanked forproviding fund, a doctoral studentship.
References
Alias SA, Jones EBG (2000) Colonization of mangrove wood by marine fungi at Kuala Selangor mangrovestand, Malaysia. Fungal Divers 5:9–21
Ananda K, Sridhar KR (2004) Diversity of filamentous fungi on decomposing leaf and woody litter ofmangrove forests in the southwest coast of India. Curr Sci 87:1431–1435
Anastasiou CJ (1964) Some aquatic fungi imperfecti from Hawaii. Pac Sci 18:202–206Anderson IC, Cairney JWG (2004) Diversity and ecology of soil fungal communities: increased under-
standing through the application of molecular techniques. Environ Microbiol 6:769–779. doi:10.1111/j.1462-2920.2004.00675.x
Berg A, Gardenfors U, Hallingback T, Noren M (2002) Habitat preferences of red-listed fungi and bryo-phytes in woodland key habitats in southern Sweden—analyses of data from a national survey.Biodivers Conserv 11:1479–1503. doi:10.1023/A:1016271823892
Boddy L (2001) Fungal community ecology and wood decomposition processes in angiosperms, fromstanding tree to complete decay of course woody debris. Ecol Bull 49:43–56
Bongers T, Ferris H (1999) Nematode community structure as a bioindicator in environmental monitoring.Trends Ecol Evol 14:224–228. doi:10.1016/S0169-5347(98)01583-3
Byrne PJ, Jones EBG (1975) Effect of salinity on spore germination of terrestrial and marine fungi. Trans BrMycol Soc 64:497–503
Cai L, Tsui CKM, Zhang K, Hyde KD (2002) Aquatic fungi from Lake Fuxian, Yunnan, China. FungalDivers 9:57–70
Cai L, Zhang KQ, McKenzie EHC, Hyde KD (2003) Freshwater fungi from bamboo and wood submergedin the Liput River in the Philippines. Fungal Divers 13:1–12
Cai L, Ji KF, Hyde K (2006) Variation between freshwater and terrestrial fungal communities on decayingbamboo culms. Antonie Van Leeuwenhoek 89:293–301. doi:10.1007/s10482-005-9030-1
Chen J, Xu LL, Liu B, Liu XZ (2007) Taxonomy of Dactylella complex and Vermispora. I. Genericconcepts based on morphology and ITS sequences data. Fungal Divers 26:73–83
Chinnadurai G, Fernando OJ (2007) Meiofauna of mangroves of the southeast coast of India with specialreference to the free-living marine nematode assemblage. Estuar Coast Shelf Sci 72:329–336.doi:10.1016/j.ecss.2006.11.004
Biodivers Conserv (2009) 18:1695–1714 1711
123

Cooke RC (1963) Ecological characteristics of nematode-trapping hyphomycetes I. Preliminary studies.Ann Appl Biol 52:431–437. doi:10.1111/j.1744-7348.1963.tb03767.x
Das M, Royer TV, Leff LG (2007) Diversity of fungi, bacteria, and actinomycetes on leaves decomposing ina stream. Appl Environ Microbiol 73:756–767. doi:10.1128/AEM.01170-06
Duong LM, Jeewon R, Lumyong S, Hyde KD (2006) DGGE coupled with ribosomal DNA gene phylogeniesreveal uncharacterized fungal phylotypes. Fungal Divers 23:121–138
Duong LM, McKenzie EHC, Lumyong S, Hyde KD (2008) Fungal succession on senescent leaves ofCastanopsis diversifolia in Doi Suthep-Pui National Park, Thailand. Fungal Divers 30:23–36
Elshafie AE, Al-Bahry SN, Ba-Omar T (2003) Nematophagous fungi isolated from soil in Oman. Sydowia55:18–32
Esseen PA, Ehnstrom B, Ericson L, Sjoberg K (1997) Boreal forests. Ecol Bull 46:16–47Fryar SC, Booth W, Davies J, Hodgkiss IJ, Hyde KD (2004) Distribution of fungi on wood in the Tutong
River, Brunei. Fungal Divers 17:17–38Gopal B, Chauhan M (2006) Biodiversity and its conservation in the Sundarban mangrove ecosystem. Aquat
Sci 68:338–354Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and
comparison of species richness. Ecol Lett 4:379–391Gray NF (1983) Ecology of nematophagous fungi: distribution and habitat. Ann Appl Biol 102:501–509.
doi:10.1111/j.1744-7348.1983.tb02721.xGray NF (1987) Nematophagous fungi with particular reference to their ecology. Biol Rev Camb Philos Soc
62:245–304. doi:10.1111/j.1469-185X.1987.tb00665.xGray NF (1988) Ecology of nematophagous fungi: effect of the soil nutrients N, P and K, and seven major
metals on distribution. Plant Soil 108:286–290. doi:10.1007/BF02375661Gulis V, Suberkropp K, Rosemond AD (2008) Comparison of fungal activities on wood and leaf litter in
unaltered and nutrient-enriched headwater streams. Appl Environ Microbiol 74:1094–1101. doi:10.1128/AEM.01903-07
Hao Y, Luo J, Zhang K (2004) A new aquatic nematode-trapping hyphomycete. Mycotaxon 89:235–239Hao Y, Mo M, Su H, Zhang KQ (2005) Ecology of aquatic nematode-trapping hyphomycetes in south-
western China. Aquat Microb Ecol 40:175–181. doi:10.3354/ame040175Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV, Lattin JD et al (1986) Ecology of coarse
woody debris in temperate ecosystems. Adv Ecol Res 15:133–276. doi:10.1016/S0065-2504(08)60121-X
Heilmann-Clausen J, Christensen M (2004) Does size matter? On the importance of various dead woodfractions for fungal diversity in Danish beech forests. For Ecol Manag 201:105–117
Ho WH, Yanna, Hyde KD, Hodgkiss IJ (2002) Seasonality and sequential occurrence of fungi on woodsubmerged in Tai Po Kau Forest Stream, Hong Kong. Fungal Divers 10:21–43
Hyde KD (1988) Observations on the vertical distribution of marine fungi on Rhizophora spp., at KampongDanau mangrove Brunei. Brunei Asian Mar 5:77–81
Hyde KD (1989) Ecology of tropical marine fungi. Hydrobiologia 178:199–208. doi:10.1007/BF00006027Hyde KD (1990) A study of vertical zonation of intertidal fungi on Rhizophora apiculata at Kampong
Kapok mangrove, Brunei. Aquat Bot 36:255–262. doi:10.1016/0304-3770(90)90039-NHyde KD, Alias SA (2000) Biodiversity and distribution of fungi associated with decomposing Nypa
fruticans. Biodivers Conserv 9:393–402. doi:10.1023/A:1008911121774Hyde KD, Goh TK (1998) Fungi on submerge wood in the Riviere St Marie-Louis, The Seychelles. S Afr J
Bot 64:330–336Hyde KD, Lee SY (1995) Ecology of mangrove fungi and their role in nutrient cycling: what gaps occur in
our knowledge? Hydrobiologia 295:107–118. doi:10.1007/BF00029117Hyde KD, Jones EBG, Leano E, Pointing SB, Poonyth AD, Vrijmoed LLP (1998) Role of fungi in marine
ecosystems. Biodivers Conserv 7:1147–1161. doi:10.1023/A:1008823515157Ilieva-Makulec K, Olejniczak I, Szanser M (2006) Response of soil micro- and mesofauna to diversity and
quality of plant litter. Eur J Soil Biol 42:S244–S249. doi:10.1016/j.ejsobi.2006.07.030Ingold CT (1944) Some new aquatic hyphomycetes. Trans Br Mycol Soc 27:45–46Jaffee BA, Strong DR, Muldoon AE (1996) Nematode-trapping fungi of a natural scrubland: tests for food
chain involvement. Mycologia 88:554–564. doi:10.2307/3761149Jansson HB, Lopez-Llorca LV (2001) Biology of nematophagous fungi. In: Misra JK, Horn BW (eds)
Trichomycetes and other fungal groups, 1st edn. Science, Plymouth, pp 145–172Johnson TW, Autery CL (1961) An Arthrobotrys from brackish water. Mycologia 53:432–433. doi:10.2307/
3756586
1712 Biodivers Conserv (2009) 18:1695–1714
123

Kataoka R, Taniguchi T, Ooshima H, Futai K (2008) Comparison of the bacterial communities establishedon the mycorrhizae formed on Pinus thunbergii root tips by eight species of fungi. Plant Soil 304:267–275. doi:10.1007/s11104-008-9548-x
Kerry BR (1987) Biological control. In: Brown RH, Kerry BR (eds) Principles and practice of nematodecontrol in crops. Academic Press, New York, pp 233–263
Kerry BR (2000) Rhizosphere interactions and the exploitation of microbial agents for the biological controlof plant-parasitic nematodes. Annu Rev Phytopathol 38:423–441. doi:10.1146/annurev.phyto.38.1.423
Kodsueb R, McKenzie EHC, Lumyong P, Hyde KD (2008) Fungal succession on leaf litter of Mangolialiliifera (Magnoliaceae). Fungal Divers 30:55–72
Lambshead PJD, Boucher G (2003) Marine nematode deep-sea biodiversity-hyperdiverse or hype? J Bio-geogr 30:475–485
Larsen M, Nansen P, Gronvold J, Wolstrup J, Henriksen SA (1997) Biological control of gastro-intestinalnematodes; facts, future, or fiction? Vet Parasitol 72:479–492. doi:10.1016/S0304-4017(97)00112-X
Li TF, Zhang KQ, Liu XZ (2000) Taxonomy of nematophagous fungi, 1st edn. Science, BeijingLi LF, Li T, Zhao ZW (2007) Differences of arbuscular mycorrhizal fungal diversity and community
between a cultivated land, an old field, and a never-cultivated field in a hot and arid ecosystem ofsouthwest China. Mycorrhiza 17:655–665. doi:10.1007/s00572-007-0143-4
Liu XF, Zhang KQ (2003) Dactylella shizishanna sp. nov., from Shizi Mountain, China. Fungal Divers14:103–107
Liu B, Liu XZ, Zhuang WY (2005) Orbilia querci sp. nov. and its knob-forming nematophagous anamorph.FEMS Microbiol Lett 245:99–105. doi:10.1016/j.femsle.2005.02.027
Lonsdale D, Pautasso M, Holdenrieder O (2008) Wood-decaying fungi in the forest: conservation needs andmanagement options. Eur J For Res 127:1–22. doi:10.1007/s10342-007-0182-6
Magurran AE (1988) Ecological diversity and its measurement. Croom Helm, LondonMaser C, Trappe JM (1984) The seen and unseen world of the fallen tree. Gen. Tech. Rep. PNW-164. US
Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station, 56 pMo MH, Huang X, Zhou W, Huang Y, Hao YE, Zhang KQ (2005) Arthrobotrys yunnanensis sp. nov., the
fourth anamorph of Orbilia auricolor. Fungal Divers 18:107–115Mo MH, Chen WM, Su HY, Zhang KQ, Duan CQ, He DM (2006) Heavy metal tolerance of nematode-
trapping fungi in lead-polluted soils. Appl Soil Ecol 31:11–19O’dor P, Standovar T (2002) Substrate specificity and community structure of bryophyte vegetation in a
near-natural montane beech forest. Community Ecol 3:39–49. doi:10.1556/ComEc.3.2002.1.5O’dor P, Heilmann-Clausen J, Christensen M, Aude E, van Dort KW, Piltaver A, Siller I, Veerkamp MT,
Walleyn R, Standovar T, van Hees AFM, Kosec J, Matocec N, Kraigher H, Grebenc T (2006) Diversityof dead wood inhabiting fungi and bryophytes in semi-natural beech forests in Europe. Biol Conserv131:58–71. doi:10.1016/j.biocon.2006.02.004
Peach M (1950) Aquatic predacious fungi I. Trans Br Mycol Soc 33:148–153Peach M (1952) Aquatic predacious fungi. II. Trans Br Mycol Soc 35:19–23Peach M (1954) Aquatic predaceous fungi III. Trans Br Mycol Soc 37:240Persmark L, Jansson H-B (1997) Nematophagous fungi in the rhizosphere of agricultural crops. FEMS
Microbiol Ecol 22:303–312. doi:10.1111/j.1574-6941.1997.tb00382.xPeterson EA, Katznelson H (1964) Occurence of nematode-trapping fungi in the rhizosphere. Nature
204:111–112. doi:10.1038/2041111b0Pfister DH (1994) Orbilia fimicola, a nematophagous discomycete and its Arthrobotrys anamorph. Myco-
logia 86:451–453. doi:10.2307/3760578Pfister DH (1997) Castor, Pollux and life histories of fungi. Mycologia 89:1–23. doi:10.2307/3761168Pfister DH, Liftik ME (1995) Two Arthrobotrys anamorphs from Orbilia auricolor. Mycologia 87:684–688.
doi:10.2307/3760812Poon MOK, Hyde KD (1998) Biodiversity of intertidal estuarine fungi on Phragmites at Mai Po marshes,
Hong Kong. Bot Mar 41:141–155Rohde CA (1976) Composite sampling. Biometrics 32:273–282. doi:10.2307/2529498Seena S, Wynberg N, Barlocher F (2008) Fungal diversity during the leaf decomposition in a stream
assessed through clone libraries. Fungal Divers 30:1–14Shannon CE, Weaver W (1963) The mathematical theory of communications, 1st edn. University of Il-
lionois Press, UrbanaShearer CA, Descals E, Kohlmeyer B, Kohlmeyer J, Marvanova L, Padgett D et al (2007) Fungal biodi-
versity in aquatic habitats. Biodivers Conserv 16:49–67. doi:10.1007/s10531-006-9120-zSimpson EH (1949) Measurement of diversity. Nature 163:688. doi:10.1038/163688a0Sin KW, Hyde KD, Pointing SB (2002) Comparative enzyme production by fungi from diverse lignocel-
lulosic substrates. J Microbiol Korea 40:241–244
Biodivers Conserv (2009) 18:1695–1714 1713
123

Somerfield PJ, Yodnarasri S, Aryuthaka C (2002) Relationships between seagrass biodiversity and infaunalcommunities: implications for studies of biodiversity effects. Mar Ecol Prog Ser 237:97–109. doi:10.3354/meps237097
Thorn RG, Barron GL (1984) Carnivorous mushrooms. Science 224:76–78. doi:10.1126/science.224.4644.76Tietjen JH, Alongi DM (1990) Population growth and effects of nematodes on nutrient regeneration and
bacteria associated with mangrove detritus from northeastern Queensland (Australia). Mar Ecol ProgSer 68:169–179. doi:10.3354/meps068169
Tsui CKM, Hyde KD, Hodgkiss IJ (2000) Biodiversity of fungi on submerged wood in Hong Kong streams.Aquat Microb Ecol 21:289–298. doi:10.3354/ame021289
Tsui CKM, Hyde KD, Fukushima K (2003) Fungi on submerged wood in the Koito River, Japan. Mycosci44:55–59
Vijaykrishna D, Jeewon R, Hyde KD (2006) Molecular taxonomy, origins and evolution of freshwaterascomycetes. Fungal Divers 23:351–390
Wai HH, Hyde KD, Hodgkiss IJ, Yanna (2001) Fungal communities on submerged wood from streams inBrunei, Hong Kong, and Malaysia. Mycol Res 105:1492–1501. doi:10.1017/S095375620100507X
Wakelin SA, Macdonald LM, Rogers SL, Gregg AL, Bolger TP, Baldock JA (2008) Habitat selective factorsinfluencing the structural composition and functional capacity of microbial communities in agriculturalsoils. Soil Biol Biochem 40:803–813. doi:10.1016/j.soilbio.2007.10.015
Webster J, Henrici A, Spooner B (1998) Orbilia fimicoloides sp. nov., the teleomorph of Dactylella cf.oxyspora. Mycol Res 102:99–102. doi:10.1017/S0953756297004747
Whitford WG (1996) The importance of the biodiversity of soil biota in arid ecosystems. Biodivers Conserv5:185–195. doi:10.1007/BF00055829
Wong MKM, Goh TK, Hodgkiss IJ, Hyde KD, Ranghoo VM, Tsui CKM et al (1998) Role of fungi infreshwater ecosystems. Biodivers Conserv 7:1187–1206. doi:10.1023/A:1008883716975
Yu Z, Qiao M, Zhang Y, Baral HO, Zhang KQ (2007a) Orbilia vermiformis sp. nov. and its anamorph.Mycotaxon 99:271–278
Yu ZF, Zhang Y, Qiao M, Zhang KQ (2007b) Orbilia dorsalia sp nov., the teleomorph of Dactylelladorsalia sp nov. Cryptogam Mycol 28:55–63
Zhang ZN, Zhou H (2003) The systematics of free-living marine nematodes. J Ocean Univ Qingdao 33:891–900
1714 Biodivers Conserv (2009) 18:1695–1714
123




















