
Leaf-litter breakdown in streams of East Malaysia (Borneo) along an altitudinal gradient: initial...
12
Leaf-litter breakdown in streams of East Malaysia (Borneo) along an altitudinal gradient: initial nitrogen content of litter limits shredder feeding Author(s): Tajang Jinggut and Catherine M. Yule Source: Freshwater Science, (-Not available-), p. 000 Published by: The University of Chicago Press on behalf of Society for Freshwater Science Stable URL: http://www.jstor.org/stable/10.1086/681256 . Accessed: 26/03/2015 02:41 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . The University of Chicago Press and Society for Freshwater Science are collaborating with JSTOR to digitize, preserve and extend access to Freshwater Science. http://www.jstor.org This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AM All use subject to JSTOR Terms and Conditions
-
Upload
umonash-my -
Category
Documents
-
view
0 -
download
0
Transcript of Leaf-litter breakdown in streams of East Malaysia (Borneo) along an altitudinal gradient: initial...

Leaf-litter breakdown in streams of East Malaysia (Borneo) along an altitudinal gradient:initial nitrogen content of litter limits shredder feedingAuthor(s): Tajang Jinggut and Catherine M. YuleSource: Freshwater Science, (-Not available-), p. 000Published by: The University of Chicago Press on behalf of Society for Freshwater ScienceStable URL: http://www.jstor.org/stable/10.1086/681256 .
Accessed: 26/03/2015 02:41
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press and Society for Freshwater Science are collaborating with JSTOR to digitize,preserve and extend access to Freshwater Science.
http://www.jstor.org
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

TROPICAL STREAMS
Leaf-litter breakdown in streams of East Malaysia(Borneo) along an altitudinal gradient: initial nitrogencontent of litter limits shredder feeding
Tajang Jinggut1,2 and Catherine M. Yule1,3
1Monash University Malaysia, Jalan Lagoon Selatan, 47500 Bandar Sunway, Selangor Darul Ehsan, Malaysia
Abstract: Breakdown of litter in streams is an important terrestrial–aquatic C pathway in forests. Macroinver-tebrate detritivore shredders that feed on litter may be more abundant in temperate than tropical streams becausethe feeding guild is generally restricted to cool waters and because temperate streams may have more high-qualityresources. However, rich shredder communities have been found in some high-altitude tropical streams. Weinvestigated litter breakdown and the variation in resource quality of native riparian vegetation in tropicalstreams at different altitudes (86–3393 m) in eastern Malaysia (Borneo). At each stream, we collected 3 nativelocal leaf species, analyzed their initial resource quality, and incubated them for 56 d in coarse- (shredder andmicrobial colonization) and fine- (only microbial colonization) mesh bags. We incubated Macaranga tanariuslitter in all streams to compare breakdown rates directly. Shredder-mediated breakdown of native litter was de-tected in only 5 of 24 cases and was not accompanied by altitudinal trends, whereas Macaranga shredder-mediated breakdown rates increased with elevation when rates were normalized for temperature (/degree-day).Regressions of resource quality and breakdown rates revealed consistent effects of initial N concentration inpredicting breakdown rates that transcended decomposer and temperature effects. Shredder abundance, but notdiversity, increased with altitude. The shredder guild may be important in tropical streams, particularly streamsat higher altitudes and when litter N is not limiting.Key words: tropical, altitude, Borneo, litter decomposition, macroinvertebrates, resource quality, latitude
Litter breakdown is a significant pathway for terrestrialplant primary production, and >90% of net primary produc-tion ends up as detritus (Fisher and Likens 1973, Cebrian1999). In forested streams, litter breakdown is a C drain-age system that channels terrestrially derived C out of thegreater catchment and downstream where it is assimilated,sequestered, or mineralized (Webster andMeyer 1997). Theagents of litter breakdown in low-order temperate streamsare microbial decomposers (mainly aquatic hyphomycetes)and leaf-shreddingmacroinvertebrates (shredders). In tropi-cal streams, this process is generally attributed to microbialaction and physical fragmentation (Irons et al. 1994, Boyeroet al. 2011b) because of a reported scarcity of shredders intropical streams (e.g., Papua New Guinea: Yule 1996, HongKong: Dudgeon and Wu 1999, Kenya: Dobson et al. 2002,Brazil: Gonçalves et al. 2006). Shredders are thought to bemore diverse and abundant in temperate than in tropicalstreams because they are evolutionary products of cool run-
ning waters and the accumulation of high-quality leaf litterin pulses caused by seasonality (Irons et al. 1994, Dobsonet al. 2002).
Authors of some surveys have reported that shreddersmay be common and abundant in some tropical streams.In the absence of shredders common in the temperatezone, other taxa—crayfish in Australia (Cheshire et al.2005), snails, crabs, and cockroaches in peninsular Malay-sia (Yule et al. 2009), and mayflies in Papua New Guinea(Yule 1996)—have filled the role, whereas some high-altitude tropical streams support abundant and diverse shred-der communities similar to those of temperate streams (Ecua-dor: Jacobsen et al. 1997, Australian tropics: Cheshire et al.2005, Camacho et al. 2009, peninsular Malaysia: Yule et al.2009, Kenya: Masese et al. 2014). These contrasting re-ports on shredder distribution in the tropics raise ques-tions about their functional importance in tropical streamsand challenge the general perception that litter breakdown
E-mail addresses: [email protected]; [email protected]
DOI: 10.1086/681256. Received 29 August 2014; Accepted 29 January 2015; Published online 20 March 2015.Freshwater Science. 2015. 34(2):000–000. © 2015 by The Society for Freshwater Science. 000
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

in tropical streams is mainly driven by microbes and frag-mentation (Mathuriau and Chauvet 2002, Gonçalves et al.2006, Rincón and Santelloco 2009).
If shredders are temperate-adapted taxa, then a recipro-cal functional trend may exist at altitudinal and latitudinalgradients because tropical streams begin to loosely resem-ble temperate streams with increasing altitude (Yule et al.2009), where they are characterized by low temperature,low riparian diversity, and the presence of temperate planttaxa (Kitayama 1992). At altitudinal–latitudinal scales, tem-perature is pivotal in controlling breakdown rates via micro-bial metabolism (Ferreira and Chauvet 2011), with possiblecompensation effects on breakdown via shredder feeding incooler streams, where this feeding group is more prevalent(Boyero et al. 2011a). A study in France revealed trends inlitter breakdown with altitude (690–1300 m) that were afunction of temperature (Taylor and Chauvet 2014). Down-stream reaches had faster microbially mediated breakdownrates, whereas further upstream, shredder contribution tobreakdown was greater when rates were corrected for tem-perature (Taylor and Chauvet 2014). These results weresimilar to patterns in breakdown rates with distance fromthe equator (Boyero et al. 2011b).
The effects of shredders on breakdown also depend onthe functional traits of riparian plant species (Cornwell et al.2008). The general hypothesized trends across latitudesare an increase in structural and secondary defensive com-pounds of leaves closer to the equator as an adaptation tointensified herbivory (Coley and Barone 1996). Makkonenet al. (2012) studied the terrestrial breakdown of temperateand tropical litter across the globe in a fully reciprocal studyand found that breakdown rates were influenced moreby litter traits than local factors because the same rank ofdecomposability was detected regardless of climate/biome(subarctic–tropical). Tropical leaves consistently broke downmore slowly than leaves from other regions. These trendsmay not reciprocate with altitude because foliar and litternutrients typically decrease with increasing elevation as aconsequence of nutrient-poor soils, which lead to low-qualitylitter (Tanner et al. 1998) at higher altitudes. However, dataare lacking to confirm whether these patterns are reflectedduring litter breakdown (Kitayama and Aiba 2002). If par-allel latitudinal–altitudinal plant functional trends do exist,larger shredder communities and better resource qualitymay have additive effects on litter breakdown, with shred-ders acting as important agents of breakdown in highlandstreams and microbial decomposers and fragmentation tak-ing precedence in lowland streams (Irons et al. 1994, Ca-macho et al. 2009, Yule et al. 2009).
Understanding the interactions between decomposercommunities, temperature, and plant traits in controllingbreakdown is crucial in predicting effects of climate changeon key ecosystem processes, particularly if the functionalpartitioning of microbial and invertebrate communities onbreakdown depends on temperature regimes (Boyero et al.
2011b, Martínez et al. 2014). We examined the relative im-portance of shredders in facilitating litter breakdown withaltitude in streams across East Malaysia (Borneo) by mea-suring litter breakdown rates and initial leaf chemistry with2 approaches: 1) use of a common leaf species (Macarangatanarius) across altitudes to compare breakdown rates di-rectly with the varying effect of litter quality removed fromaltitudinal comparisons, and 2) use of the 3 most abundantleaf species from the riparian zones at each study site to testfor reciprocal trends in resource quality and altitude. Weselected Macaranga tanarius, a secondary-forest gap spe-cies, as the common leaf species because it is not foundin any riparian zone in our study and is of high resourcequality (high initial N and low lignin content). We did notselect sites along an altitudinal transect per se, but ratherfrom contrasting forest types that occur at different alti-tudes (86–3393 m asl). Altitudinal patterns in local speciesquality could account for possible positive or negative ef-fects of resource quality interacting with shredder commu-nities and breakdown rates. We hypothesized that shredder-mediated breakdown of litter is not negligible and increaseswith altitude when the effects of varying resource qualityand temperature are removed. Previous investigators whotested altitudinal effects on shredders focused on their dis-tribution. To our knowledge, ours is the first study in whichlitter breakdown in tropical streams was examined alongan altitudinal gradient.
METHODSStudy sites
We sampled eight 1st- to 2nd-order streams flowingthrough different forest types and altitudes in 4 nationalparks across eastern Malaysia, Borneo: alluvial (86 m),heath (102 m), limestone (120 m), dipterocarp (180 m), hilldipterocarp (503 m), lower montane (1580 m), montane(2200 m), and subalpine forests (3393 m) (Appendix S1).We conducted our study over 4 occasions, one national parkat a time, to accommodate logistical issues. Borneo lies onthe equator and has a wet tropical climate. We avoided run-ning experiments during the monsoon season when rainfallis the greatest to avoid flash floods (December–February;Appendix S1).
The heath forest stream in Bako National Park was alowland stream characterized by low-nutrient sandy sub-strate. Dacrydium (family Podocarpaceae: conifers), Gym-nostoma (family Casuarinaceae), and a few dipterocarps(e.g., Cotylelobium burckii, endemic to Borneo) typicallyare found in Malaysian heath forests (Ashton 1971, Beckeret al. 1999).
The lowland dipterocarp stream was in Kubah NationalPark and was characterized by red–yellow, nutrient-poorpodzolic soils and mainly consisted of lowland dipterocarpforest on steep terrain. Dipterocarpaceae and Euphorbia-ceae are the dominant families in lowland dipterocarp for-ests of Borneo (22 and 12% of all trees). Widespread genera
000 | Tropical altitudinal litter-breakdown rates T. Jinggut and C. M. Yule
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

include Shorea (Dipterocarpaceae), Syzygium (Myrtaceae),and Dipterocarpus (Dipterocarpaceae) (Iwata et al. 2003,Slik et al. 2003).
Streams flowing through alluvial, limestone, and hilldipterocarp forests were in Mulu National Park, wherethe underlying rock type exists as a gradient along an alti-tudinal scale. Lowland (<250 m) alluvium (clay and sand)and terrace (clay, sand, and gravel) formations are followedby limestone and shale/sandstone that range from 250 to2376 m and give rise to a succession of forest types. Dip-terocarps predominate on limestone and in alluvial forests.Species, such as Hopea andersonii and Shorea multiflora,are common on limestone formations, whereas trees withstilt roots (e.g., Lauraceae) are more prevalent in low-lyingalluvial forests.
The lower montane, montane, and subalpine streamswere situated within Kinabalu National Park (4095 m sum-mit). In a quantitative transect study, Kitayama (1992) clas-sified the mountain into 4 distinct forest zones based onsoil types and floristic zones: lowland (<1200 m), lowermontane (1200–2350 m), upper montane (2000–2800 m),and subalpine (2800 m–tree line). Tree diversity and spe-cies dispersal decreased with altitude, and the distributionof dipterocarps was limited to the lower montane belt,whereas oaks (Fagaceae: Quercus) and conifers were widelydistributed from the lower montane to subalpine forests(Kitayama 1992).
We measured water temperature, pH, conductivity, anddissolved O2 (DO) at each site in situ with a multiprobemeter (Cyberscan PCD 650; Eutech, Waltham, Massachu-setts) on a weekly basis for 5 wk (Table 1). We took 5 depthmeasurements along each of 5 transects along a 50-mreach. We measured foliage density at each transect as leafarea index (LAI) with a digital canopy imager (CI-110; CIDBio-Science, Camas,Washington).
Litter-breakdown experimentsWe studied a total of 25 leaf species. We collected
leaves for the common leaf-species treatment, M. tanarius,adjacent to Ampang Forest Reserve in peninsular Malaysia.We collected fallen leaves of local species along the riparianzones of the study streams from five 2- × 2-m quadrats, andbrought them back to the laboratory. We sorted litter mor-phospecies, and identified them to the best of our abilitybased on knowledge of local park officials and reaffirmedthese identifications with keys (Appendix S2). For each for-est type, we selected the 3 most abundant leaf species for thelitter-breakdown study and measured initial leaf chemistryas proximate lignin and cellulose, total polyphenolic andtannin content, total Kjeldahl N, and P content (Graça et al.2005).
No oven was available at the field sites, so we air-driedleaves for a minimum of 24 h, stored them in a heated dryroom (7–10 d), and weighed them into portions of 6 ±
0.01 g of air-dried leaves. We placed leaves in individual20- × 20-cm coarse- (10-mm aperture) and fine-mesh(0.5-mm aperture) bags. We established 8 treatments perstream: M. tanarius leaves and leaves of 3 local speciesin 2 mesh sizes. We prepared and deployed a total of160 bags (192 bags for subalpine streams) for each studysite. We secured leaf bags to the stream bed with ropesand tied them down using available substrate (rocks andexposed roots).
We incubated leaf bags in the streams and retrieved4 replicates of each treatment type after 0, 7, 14, 28, and56 d, with the exception of the subalpine streams, wherean additional set was incubated for 150 d to account for theeffect of extremely low temperatures on breakdown rates.We used day-0 leaf bags (n = 4/mesh size) to correct initialmass for handling losses, measurement of initial ash-freedry mass (AFDM), and conversion factors. During retrieval,we collected leaf bags from the streams with a fine-meshnet and placed them in separate plastic bags. In the labora-tory, we removed leaves carefully from the bags and rinsedthem gently to remove sediments before oven-drying themat 50°C for 48 h. We weighed the dried leaves, ignited themat 500°C for 4 h, and reweighed them to measure AFDM.For each sampling date, remaining leaf mass was calculatedas remaining AFDM/corrected initial AFDM.
Shredder communitiesTo substantiate potential altitudinal trends in shredder-
mediated breakdown with invertebrate data, we exploreddata from an earlier macroinvertebrate survey of the samestreams for altitudinal trends in shredder assemblages. In-vertebrates were sampled haphazardly from 20- × 20-cmsamples of submerged leaf-litter packs from 5 riffles and5 pools at each stream along a 50-m reach using nets(250-μm mesh). In the field, invertebrates were washedgently away from large leaves, and the remaining sam-pled materials of invertebrates and fine debris were pre-served in 70% ethanol for immediate transport back to thelaboratory. In the laboratory, invertebrates were sortedfrom the debris under a stereomicroscope, preserved in70% ethanol, and identified to the lowest possible taxo-nomic level using keys published by Yule and Yong (2004).Classification into functional feeding groups was done withreference to Yule et al. (2009) and confirmed with gut anal-ysis. Gut contents of specimens were removed from inver-tebrates under 40× magnification onto a glass slide beforebeing mounted with polyvinyl alcohol–lactophenol and ob-served at 200 and 400× magnification under a compoundmicroscope.
Statistical analysisWe compared environmental variables (water-chemistry
and physical variables) among sites with analysis of var-iance (ANOVA) and Tukey’s post hoc test for differences
Volume 34 June 2015 | 000
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

Table
1.Geographiclocation
andmean(±1SE
)values
forenvironm
entalcharacteristicsof
thestud
ystream
s.Tem
perature,p
H,con
ductivity,dissolvedO
2,and
leaf
area
index(LAI)werecomparedusinganalysisof
variance.M
eans
withthesameletter
areno
tsign
ificantly
different.
Site
Altitud
e(m
asl)
Latitud
e,Lon
gitude
Order
Width
(m)
Depth
(cm)
Tem
perature
(°C)
pHCon
ductivity
(μS/cm
)Dissolved
O2
(mg/L)
LAI
Allu
vial
864°02′32.4″,114°48′49.7″
1st
1.05
–1.30
5.9–
30.2
24.3a±0.1
6.16
b±0.01
54.42a
±4.50
3.520a
±0.115
2.10
a±0.04
Heath
102
1°43
′24.1″,110°27′30.3″
2nd
0.45
–1.32
6.4–
70.6
25.3b±0.2
4.18
c±0.70
25.27b
±2.37
5.406b
±1.317
1.59
a±0.04
Lim
estone
120
4°01
′29.2″,114°49′09.0″
1st
0.92
–1.39
8.8–
67.1
24.4a±0.1
7.05
a±0.19
116.30
c±23.65
6.00
b±1.576
2.29
a±0.14
Dipterocarp
180
1°36
′33.5″,110°11′12.2″
1st
0.35
–1.24
4.3–
17.4
24.3a±0.1
6.90
a±0.15
42.44a
±3.10
7.459c
±0.753
2.35
abc±0.24
Hill
Dipterocarp
503
4°02′20.15″,114°52
′25.06″
2nd
0.92
–2.00
5.8–
11.2
22.8c±0.1
6.10
b±0.23
7.51
d±0.25
7.892c
±0.179
1.84
ab±0.19
Low
erMon
tane
1580
6°00
′59.8″,116°2′39.0″
1st
0.64
–0.87
5.9–
8.8
16.8d±0.4
6.62
ab±0.54
17.91b
d±2.03
9.897d
±0.704
2.08
ab±0.02
Mon
tane
2200
6°02
′85.5″,116°34′74.7″
1st
0.90
–1.35
4.4–
16.4
13.6e±0.2
7.22
a±1.05
12.09d
±0.75
9.54
d±0.544
1.93
bc±0.65
Subalpine
3393
6°03
′70.5″,116°34′01.1″
1st
0.07
–0.53
2.7–
5.9
6.9f±0.4
7.25
a±0.27
5.80
d±1.76
7.667c
±0.124
1.44
c±0.07
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

between forest types. We tested data for normality and ho-mogeneity of variances and transformed them to meetANOVA assumptions (log10[x]-transformations).
We estimated breakdown rates with exponential decaymodels. We regressed ln(proportion remaining AFDM)against incubation period (d), and used the slopes of theseregressions as the breakdown rate.We also estimated break-down rates per degree-day, which normalizes for the effectof temperature and assumes no biological activity at tem-peratures below freezing. Temperature was integrated withincubation time based on the average water temperaturemeasured during the experiment. First, we explored for sig-nificant regressions of mass remaining over days or degree-days with simple linear regressions. This step also was aprerequisite for comparing the variation in breakdown ratesacross treatments. We then explored for significant effectsof altitude by regressing breakdown rates against altitudewith simple linear regression. We used analysis of covari-ance (ANCOVA) to compare mass loss between mesh sizesand among forest types separately for Macaranga treat-ments and local leaf species. We used proportion of AFDMremaining as the response variable, mesh size and foresttype as treatments, and time (d) or degree-days as covari-ates. Faster breakdown rates in coarse- than in fine-meshbags indicated greater shredder-mediated breakdown. Priorto analysis, data were checked for normality and homoge-neity of variances, and ln(x)-transformed to meet assump-tions of the ANCOVA.
We visualized the variation in interspecific litter traitswith principal component analysis (PCA). We explored cor-relations between initial leaf chemistry (expressed as mini-mum, maximum, and mean concentrations) and altitudewith nonparametric Spearman’s rank correlation. We ex-plored the relationship between litter traits and breakdownrates across species with Akaike’s information criteria (AIC)and forward-stepwise multiple linear regressions (Makko-nen et al. 2012). We first ran simple linear regressions ofall litter traits and breakdown rates and strengthened lin-ear relationships with log10(x)-transformation wheneverrequired. After significant multiple linear models were se-lected, we investigated collinearity between significant pre-dictors based on the variation inflation factor (VIF < 4indicated no collinearity) and removed colinear traits thatdecreased the fit of the model. We also ran simple linearregressions of breakdown rates against PCA scores to de-tect relationships as an aggregate of plant functional traitsand breakdown rate. We regressed shredder abundance(number of individuals [ind]/0.04 m2), relative abundance(% shredders), and diversity (number of species or mor-phospecies/0.04 m2) against altitude, and explored non-significant regressions with an ANOVA between streams(Yule et al. 2009).
RESULTSRegressions for Macaranga mass remaining against
time (d) were significant at all sites except in the heath
forest stream (simple linear regression, p > 0.05). Maca-ranga breakdown rates (/d) were not predicted by altitude(coarse: F = 0.46, p = 0.53; fine: F = 1.73, p = 0.25; n = 8).Instead, coarse-mesh breakdown rates were faster at thelower montane stream than at the dipterocarp stream, andbreakdown rates for all other forest types were interme-diate (ANCOVA, F6,107 = 3.56, p = 0.003). Fine-meshbreakdown rates were fastest in the alluvial, limestone,and lower montane streams, followed by hill dipterocarpand montane stream. The slowest rates were recorded indipterocarp and subalpine streams (ANCOVA, F6,110 =21.5, p < 0.0001). Shredder-mediated leaf breakdown (indi-cated by the significant effect of mesh size on breakdown)was detected at the dipterocarp, hill dipterocarp, lowermontane, montane, and subalpine streams (ANCOVA, p <0.05).
Temperature-corrected Macaranga breakdown rates(/degree-day [dd]) revealed an increasing trend in coarse-mesh breakdown rates with altitude across the forest types(simple linear regression, y = [1.147 × 10–6]x + 0.000812,r2 = 0.87, p = 0.0007; Fig. 1). The differences betweencoarse- and fine-mesh breakdown rates increased with el-evation (range: 1.7–4.2×), whereas the regression of fine-mesh breakdown rates against altitude was not significant(F = 0.03, p = 0.58). ANCOVA on breakdown rates (/dd)separated coarse-mesh rates into 2 groups. The lower allu-vial, limestone, dipterocarp, and hill dipterocarp streamshad slower breakdown rates than did the lower montane,montane, and subalpine streams (ANCOVA, F6,107 = 8.3,p < 0.0001). Fine-mesh breakdown rates (/dd) were fas-test for the lower montane and limestone streams, fol-lowed by alluvial, hill dipterocarp, montane, and subalpine,with dipterocarp forest having the slowest breakdown rates(ANCOVA, F6,107 = 8.3, p < 0.0001).
Figure 1. Breakdown rates expressed per degree-day (/dd) ofMacaranga tanarius incubated at different altitudes in coarse-and fine-mesh bags. Coarse-mesh breakdown rates and altitudewere positively related (y = [1.147 × 10–6]x + 0.000812, r 2 =0.87, p = 0.0007), whereas fine-mesh breakdown rates were notpredicted by altitude (p = 0.58).
Volume 34 June 2015 | 000
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

For the local-species treatments, faster breakdown ratesin coarse- than in fine-mesh bags were detected in only5 cases: leaves incubated in the dipterocarp stream (Koom-passia malaccensis), lower montane stream (Lithocarpussp.), montane stream (Quercus kinabaluensis), and subal-pine stream (Polyosma hookeri, Eugenia kinabaluensis)(ANCOVA between mesh sizes with time as the covariate,p < 0.05; Table 2). Coarse-mesh breakdown rates were notpredicted by altitude (F = 3.7, p = 0.07, n = 24), whereasfine-mesh breakdown rates were negatively correlated withaltitude (F = 7.0, p = 0.01, r2 = 0.24, n = 24). Temperature-corrected breakdown rates were not predicted by altitudefor either mesh size (coarse: F = 0.0004, p = 0.98; fine: F =2.6, p = 0.12; n = 24).
Not all local-species regressions of mass remainingagainst time were significant. Mass remaining of some leafspecies incubated in coarse- (alluvial forest: Macarangahosei, Bhesa paniculata) and fine-mesh bags (alluvial: Ca-narium denticulatum, limestone: Palaquium elegans, sub-alpine: Schima brevifolia, Polyosma hookeri, Eugenia kina-
baluensis) were not significantly predicted by time (sim-ple linear regressions of ln[AFDM remaining] vs time, p >0.05; Table 2).
Leaf chemistry varied among the species studied (Ta-ble S1). Leaf chemistry of the 24 local plant species ex-plained 77.1% of the variation in a PCA (PC1: 56.3%, PC2:20.8%; Fig. 2). PC1 was negatively related to initial P con-tent: total P (component loading = –0.31), tannin ∶P (0.32),polyphenolics ∶P (0.33), and N ∶P (0.33). PC2 was positivelyrelated to initial N content: total N (0.31), lignin ∶N (–0.38),tannin ∶N (–0.50), and polyphenolics ∶N (–0.54). Maximumand mean %lignin were negatively correlated with altitude(Spearman’s correlation: maximum %lignin: σ = –0.76, p =0.028; mean %lignin, σ = –0.81, p = 0.015), whereas initialpolyphenolic concentrations was positively correlated withaltitude (σ = 0.71, p = 0.04). Initial P concentrations of high-land litter (>1500 m; 0.01–0.03% AFDM) were lower thanthat of lowland litter (0.20–0.23% AFDM; p < 0.01).
Simple linear regressions revealed a positive relation-ship between breakdown rates (/dd) with increasing ini-
Table 2. Leaf breakdown rates (/d) of the 3 most abundant local tree species in coarse- and fine-mesh bags in streams flowingthrough their native forest type. * = p < 0.05, ** = p < 0.01, *** = p < 0.001. A = regressions differ between coarse- and fine-mesh bags(ANCOVA, p < 0.05).
Forest type Leaf species Coarse SE r2 Fine SE r2
Heath Ploiarium alternifolium 0.0109 0.0018 0.74*** 0.0122 0.0014 0.78***
Elaeocarpus chrysophyllum 0.0039 0.0014 0.39** 0.0044 0.0014 0.44**
Ixonanthes reticulata 0.0040 0.0014 0.38* 0.0030 0.0014 0.27*
Alluvial Macaranga hosei 0.0013 0.0062 0.00 0.0067 0.0031 0.28
Bhesa paniculata 0.0089 0.0061 0.15 0.0088 0.0036 0.33*
Canarium denticulatum 0.0149 0.0031 0.70*** 0.0087 0.0044 0.28
Limestone Hopea nutans 0.0173 0.0073 0.34* 0.0199 0.0042 0.68***
Shorea patoiensis 0.0337 0.0115 0.42* 0.0296 0.0065 0.63***
Palaquium elegans 0.0173 0.0057 0.44* 0.0001 0.0153 0.00
Dipterocarp Dryobalanops beccarii 0.0110 0.0020 0.71*** 0.0159 0.0044 0.51**
Koompassia malaccensis 0.0112A 0.0041 0.37* 0.0264 0.0045 0.72***
Shorea species 1 0.0129 0.0031 0.57** 0.0089 0.0030 0.40**
Hill Dipterocarp Hopea species 1 0.0124 0.0022 0.72*** 0.0116 0.0038 0.41**
Shorea parvistipulata 0.0144 0.0020 0.80*** 0.0122 0.0022 0.70***
Melanorrhoea wallichii 0.0240 0.0042 0.71*** 0.0194 0.0021 0.86***
Lower Montane Lithocarpus species 1 0.0241A 0.0045 0.70*** 0.0075 0.0013 0.72***
Lauraceae species 1 0.0091 0.0019 0.72*** 0.0072 0.0011 0.82***
Eugenia valdevenosa 0.0085 0.0020 0.67** 0.0078 0.0011 0.84***
Montane Eugenia subdecussata 0.0058 0.0009 0.72*** 0.0039 0.0011 0.41**
Quercus kinabaluensis 0.0154A 0.0011 0.92*** 0.0118 0.0008 0.92***
Eugenia houttuynii 0.0094 0.0007 0.91*** 0.0090 0.0008 0.87***
Subalpine Schima brevifolia 0.0007 0.0007 0.06 0.0004 0.0004 0.04
Polyosma hookeri 0.0024A 0.0007 0.40** 0.0001 0.0004 0.00
Eugenia kinabaluensis 0.0038A 0.0008 0.54*** 0.0005 0.0004 0.06
000 | Tropical altitudinal litter-breakdown rates T. Jinggut and C. M. Yule
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

tial % N and negative relationships with lignin ∶N, poly-phenolics ∶N, and tannin ∶N of initial litter chemistry withbreakdown rates (Fig. S1). Stepwise multiple linear re-gressions showed that the inclusion of initial N, P, and lig-nin concentrations in a single model provided the bestsolution that explained the variability between breakdownrates for both mesh-size treatments (Table 3). PC2 scores,but not PC1 scores, were significant predictors of break-down rates across species for both mesh sizes (simple lin-ear regressions, coarse mesh: r2 = 0.30, p = 0.0011, n = 24;finemesh: r2 = 0.44, p= 0.0064, n= 24). In all cases, initial Nconcentration predicted breakdown rates.
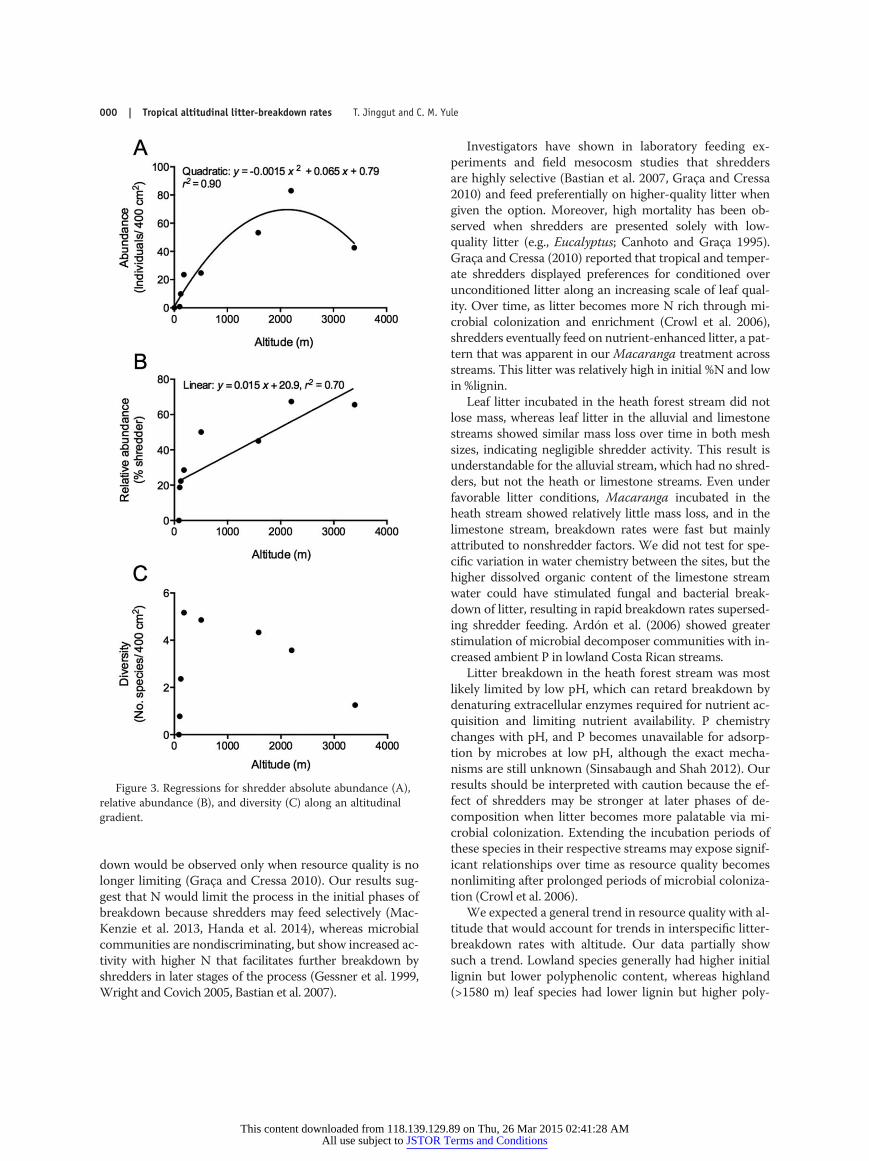
A total of 4270 invertebrates were collected in 80 litterpacks from all sites, and 1920 of the invertebrates wereclassified as shredders. No shredders were found in thelowest stream (alluvial, 86 m). Shredders were representedby 14 species classified into 5 orders across 10 families:Blattodea, Coleoptera (1), Diptera (2), Plecoptera (3), andTrichoptera (3). Shredder abundance (ind/0.04 m2) andrelative abundance (% shredders) were correlated with al-titude, but the relationships were described by differentfunctions. Shredder abundance was described by a qua-dratic relationship (y = –0.0015x2 + 0.065x + 0.79, r2 =0.90, p = 0.0058; Fig. 3A), whereas relative abundance wasdescribed by a linear function (y = 0.015x + 20.9, r2 = 0.70,p = 0.01; Fig. 3B). Shredder diversity was not correlatedwith altitude (F1,6 = 0.04, p = 0.86, n = 8), and the highestdiversities were observed in the dipterocarp (180 m) andhill dipterocarp (503 m) streams, whereas the lowest di-
versities were in the limestone (120 m), heath (102 m), andsubalpine streams (3393 m) with the other streams inter-mediate (ANOVA, F4,56 = 4.15, p = 0.0085; Fig. 3C).
DISCUSSIONTrends in shredder-mediated breakdown with altitude
were not apparent when we used local litter but were evi-dent when direct comparisons were made withM. tanarius.Our use of 2 approaches revealed effects of interspecificvariation in resource quality that naturally controls litterbreakdown and increasing shredder involvement in break-down with altitude when the effect of resource quality andtemperature were removed. Interspecific variation in break-down rates is consistent with results of previous studiesthat detected low or negligible shredder involvement inthe breakdown of litter in tropical streams (e.g., Dudgeon1982, Mathuriau and Chauvet 2002, Gonçalves et al. 2006,Rincón and Santelloco 2009). However, at some sites, lit-ter breakdown was much faster in coarse- than in fine-mesh bags, indicating faster breakdown rates when shred-ders were not excluded (Cowan et al. 1983).
Shredder abundance increased with altitude, andtemperature-corrected M. tanarius breakdown rates re-vealed increasing shredder-mediated breakdown with alti-tude. Our findings are consistent with those of Taylor andChauvet (2014) and support the hypothesis of greater shred-der involvement in breakdown with increasing elevation thatparallels litter-breakdown processes in streams away fromthe equator (Irons et al. 1994, Boyero et al. 2011b). We pro-pose that the shredder feeding group becomes functionallymore important with altitude, but the effect of varyingshredder communities may be measurable only when high-quality resources are used, i.e., shredder-mediated break-
Figure 2. Principal component analysis (PCA) ordination ofresource quality (N, P, cellulose, lignin, polyphenolic content(Phenol), tannic content (Tannin), N ∶P, lignin ∶N, lignin ∶ P,polyphenolics ∶N, polyphenolics ∶P) of the 24 litter species. Thetraits were centered and standardized prior to ordination.
Table 3. Results of multiple linear regressions of breakdown rate(k) (/degree-day [dd]) of 24 litter species, with the litter traits asindependent variables for different mesh sizes using the forwardstepwise method.
Variable Estimate t ratio p-value r2Adjusted
r2
Log10(k/dd) fine
Model 0.0023 0.63 0.55
N 0.35 3.46 0.0038
P 3.39 3.22 0.0061
Lignin –0.01 –2.17 0.0475
Log10(k/dd) coarse
Model 0.0042 0.53 0.45
N 0.30 3.08 0.0067
P 2.99 2.73 0.0142
Lignin –0.01 –2.36 0.0303
Volume 34 June 2015 | 000
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions
down would be observed only when resource quality is nolonger limiting (Graça and Cressa 2010). Our results sug-gest that N would limit the process in the initial phases ofbreakdown because shredders may feed selectively (Mac-Kenzie et al. 2013, Handa et al. 2014), whereas microbialcommunities are nondiscriminating, but show increased ac-tivity with higher N that facilitates further breakdown byshredders in later stages of the process (Gessner et al. 1999,Wright and Covich 2005, Bastian et al. 2007).
Investigators have shown in laboratory feeding ex-periments and field mesocosm studies that shreddersare highly selective (Bastian et al. 2007, Graça and Cressa2010) and feed preferentially on higher-quality litter whengiven the option. Moreover, high mortality has been ob-served when shredders are presented solely with low-quality litter (e.g., Eucalyptus; Canhoto and Graça 1995).Graça and Cressa (2010) reported that tropical and temper-ate shredders displayed preferences for conditioned overunconditioned litter along an increasing scale of leaf qual-ity. Over time, as litter becomes more N rich through mi-crobial colonization and enrichment (Crowl et al. 2006),shredders eventually feed on nutrient-enhanced litter, a pat-tern that was apparent in our Macaranga treatment acrossstreams. This litter was relatively high in initial %N and lowin %lignin.
Leaf litter incubated in the heath forest stream did notlose mass, whereas leaf litter in the alluvial and limestonestreams showed similar mass loss over time in both meshsizes, indicating negligible shredder activity. This result isunderstandable for the alluvial stream, which had no shred-ders, but not the heath or limestone streams. Even underfavorable litter conditions, Macaranga incubated in theheath stream showed relatively little mass loss, and in thelimestone stream, breakdown rates were fast but mainlyattributed to nonshredder factors. We did not test for spe-cific variation in water chemistry between the sites, but thehigher dissolved organic content of the limestone streamwater could have stimulated fungal and bacterial break-down of litter, resulting in rapid breakdown rates supersed-ing shredder feeding. Ardón et al. (2006) showed greaterstimulation of microbial decomposer communities with in-creased ambient P in lowland Costa Rican streams.
Litter breakdown in the heath forest stream was mostlikely limited by low pH, which can retard breakdown bydenaturing extracellular enzymes required for nutrient ac-quisition and limiting nutrient availability. P chemistrychanges with pH, and P becomes unavailable for adsorp-tion by microbes at low pH, although the exact mecha-nisms are still unknown (Sinsabaugh and Shah 2012). Ourresults should be interpreted with caution because the ef-fect of shredders may be stronger at later phases of de-composition when litter becomes more palatable via mi-crobial colonization. Extending the incubation periods ofthese species in their respective streams may expose signif-icant relationships over time as resource quality becomesnonlimiting after prolonged periods of microbial coloniza-tion (Crowl et al. 2006).
We expected a general trend in resource quality with al-titude that would account for trends in interspecific litter-breakdown rates with altitude. Our data partially showsuch a trend. Lowland species generally had higher initiallignin but lower polyphenolic content, whereas highland(>1580 m) leaf species had lower lignin but higher poly-
Figure 3. Regressions for shredder absolute abundance (A),relative abundance (B), and diversity (C) along an altitudinalgradient.
000 | Tropical altitudinal litter-breakdown rates T. Jinggut and C. M. Yule
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

phenolic content. The leaf species from highland and low-land heath streams had higher polyphenolic and tanninconcentrations but lower N and P content relative to leavesfrom other forest types. This pattern reflects the loss of soilN and P from extreme weathering, a common trait in high-altitude forests (Kitayama 1992) and the sandy soils ofheath forests (Becker et al. 1999). This effect was coupledwith increased accumulation of polyphenolics and tanninsby leaves in response to environmental stress, i.e., high ul-traviolet radiation and low temperatures in highlands andhigh temperatures, low nutrients, and extended dry peri-ods in coastal heath forests (Becker et al. 1999, Kurokawaand Nakashizuka 2008).
Tropical litter is highly diverse and heterogeneous, andup to 100 different species can be found in a single hectare(Bastian et al. 2007), resulting in chemically divergent re-sources at small and large scales (Reice and Cummins1974, Covich 1988, Bastian et al. 2007). Thus, litter chemis-try would have greater control over breakdown in tropicalthan in less diverse temperate forests (Aerts 1997), and ourresults showed a consistent effect of initial N concentrationin predicting breakdown rates. We initially expected thatparallel latitudinal–altitudinal breakdown trends would beexplained by the initial concentrations of lignin and sec-ondary compounds, which have been proposed as deter-mining limiting factors in tropical streams (Coley and Ba-rone 1996, Wantzen et al. 2008, Zhang et al. 2008). Handaet al. (2014) showed the greater importance of initial N inmodulating the breakdown of litter mixtures (across ter-restrial and aquatic biomes) by increasing the overall qual-ity of mixtures. Their results implied that preferences forhigher-quality resources could account for greater break-down by larger detritivore faunas (Ostrofsky 1997, Graçaet al. 2001, Hieber and Gessner 2002, Leroy and Marks2006, Talbot and Treseder 2012). The mechanism under-lying the positive effect of high N content on breakdownis relief from nutrient limitation, which promotes con-sumer productivity (Sinsabaugh et al. 1994, Gessner andChauvet 1994, Mathuriau and Chauvet 2002, Crowl et al.2006).
Shredder total and relative abundance, but not diversity,increased with altitude. In both peninsular Malaysia (Yuleet al. 2009) and the Australian wet tropics (Camacho et al.2009), positive linear relationships were detected betweenaltitude and shredder abundance, whereas in Panama, anegative relationship was detected between shredder abun-dance and altitude, but not between shredder diversity andaltitude (Camacho et al. 2009). These variations are not un-common, as Boyero et al. (2012) highlighted in their globalsynthesis of shredder diversity patterns across the globe.Diversity is highly variable across tropical streams, i.e., αdiversity was low and variable across tropical regions, sothese streams had low shredder diversity and were taxo-nomically variable. Our results suggest that trends in shred-
der communities with altitude can exist, but may not beuniversal across the tropics. Gradients in local environmen-tal factors also may vary greatly among tropical regions,giving rise to variable observations of shredder communi-ties at local and regional scales. Therefore, interpretationsof distributional gradients should be limited to local factorsover broad latitudinal–longitudinal gradients when com-paring tropical streams.
In conclusion, shredder abundance increased with al-titude, but this increase did not directly translate into aparallel functional trend of increased shredder-mediatedbreakdown of local litter. Instead, initial N concentrationof litter was a major and consistent predictor of breakdownrates, whereas no obvious trends were detected amongfunctional traits, breakdown rates, and altitude. We pro-pose that the effect of shredders on breakdown is not neg-ligible when N is not limiting (MacKenzie et al. 2013,Handa et al. 2014), and the relative contribution of shred-ders to the process is greater in cooler waters where theseinvertebrates are prevalent, similar to trends reported inother altitudinal (Taylor and Chauvet 2014) and latitudinal(Boyero et al. 2011b) studies. In light of global climate-change predictions of a warmer world (IPCC 2007), thebreakdown of litter could shift from a shredder-mediatedprocess to a microbially driven process because shreddersmay be climate sensitive (Yule et al. 2009, Boyero et al.2012), with highland tropical streams potentially experi-encing greater shifts than lowland streams, leading to pos-sible alterations to the flow of C through these catchmentswith consequences for downstream systems.
ACKNOWLEDGEMENTSLim Tse Yuen, Ray Lean, Cheryl Ong, Chou Lee Yiung, Lee Jo
Kien, and Sonja Raub assisted with fieldwork and laboratory anal-ysis. We thank Sarawak Forestry and Sabah Parks for providingaccess to the study sites. We also thank the park officials andrangers for their help and support during the study. The manu-script greatly benefitted from constructive comments by Associ-ate Editor M. A. S. Graça and 2 anonymous referees. This workwas part of TJ’s PhD project funded by Monash University.
LITERATURE CITEDAerts, R. 1997. Climate, leaf litter chemistry and leaf litter de-
composition in terrestrial ecosystems: a triangular relation-ship. Oikos 79:439–449
Ardón, M., L. A. Stallcup, and C. M. Pringle. 2006. Does leafquality mediate the stimulation of leaf breakdown by phos-phorus in Neotropical streams? Freshwater Biology 51:618–633.
Ashton, P. S. 1971. The plants and vegetation of Bako NationalPark. Malayan Nature Journal 24:151–162.
Bastian, M., L. Boyero, B. R. Jackes, and R. G. Pearson. 2007.Leaf litter diversity and shredder preferences in an Austra-lian tropical rain-forest stream. Journal of Tropical Ecology23:219–229.
Volume 34 June 2015 | 000
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

Becker, P., S. J. Davies, M. Z. H. I. Moksin, and P. M. Simanjuntak.1999. Leaf size distributions of understory plants in mixed dip-terocarp forest and heath forests of Brunei. Journal of Tropi-cal Ecology 15:123–128.
Boyero, L., R. G. Pearson, D. Dudgeon, V. Ferreira, M. A. S.Graça, M. O. Gessner, A. J. Boulton, E. Chauvet, C. M. Yule,R. J. Albariño, A. Ramírez, J. E. Helson, M. Callisto, M.Arunachalam, J. Chará, R. Figueroa, J. M. Mathooko, J. F.Gonçalves, M. S. Moretti, A. M. Chará-Serná, J. N. Davies,A. C. Encalada, S. Lamothe, L. M. Buria, J. Castela, A.Cornejo, A. O. Y. Li, C. M’Erimba, V. D. Villanueva, M. C.Zúñiga, C. M. Swan, and L. A. Barmuta. 2012. Global pat-terns of stream detritivore distribution: implications for bio-diversity loss in changing climates. Global Ecology and Bio-geography 21:134–141.
Boyero, L., R. G. Pearson, D. Dudgeon, M. A. S. Graça, M. O.Gessner, R. J. Albariño, V. Ferreira, C. M. Yule, A. J. Boulton,M. Arunachalam, M. Callisto, E. Chauvet, A. Ramírez, J. Chará,M. S. Moretti, J. F. Gonçalves, J. E. Helson, A. M. Chará-Serna,A. C. Encalada, J. N. Davies, S. Lamothe, A. Cornejo, A. O. Y.Li, L. M. Buria, V. D. Villanueva, M. C. Zúñiga, and C. M.Pringle. 2011a. Global distribution of a key trophic guild con-trasts with common latitudinal diversity patterns. Ecology 92:1839–1848.
Boyero, L., R. G. Pearson, M. O. Gessner, L. A. Barmuta, V. Ferreira,M. A. S. Graça, D. Dudgeon, A. J. Boulton, M. Callisto, E.Chauvet, J. E. Helson, A. Bruder, R. J. Albariño, C. M. Yule,M. Arunachalam, J. N. Davies, R. Figueroa, A. S. Flecker, A.Ramírez, R. G. Death, T. Iwata, J. M. Mathooko, C. Mathuriau,J. F. Gonçalves, M. S. Moretti, T. Jinggut, S. Lamothe, C.M’Erimba, L. Ratnarajah, M. H. Schindler, J. Castela, L. M. Buria,A. Cornejo, V. D. Villanueva, and D. C. West. 2011b. A globalexperiment suggests climate warming will not accelerate litterdecomposition in streams but might reduce carbon sequestra-tion. Ecology Letters 14:289–294.
Camacho, R., L. Boyero, A. Cornejo, A. Ibañez, and R. G. Pear-son. 2009. Local variation in shredder distribution can ex-plain their oversight in tropical streams. Biotropica 41:625–632.
Canhoto, C., and M. A. S. Graça. 1995. Food value of intro-duced eucalypt leaves for a Mediterranean stream detritivore:Tipula lateralis. Freshwater Biology 34:209–214.
Cebrian, J. 1999. Patterns in the fate of production in plantcommunities. American Naturalist 154:449–468.
Cheshire, K., L. Boyero, and R. G. Pearson. 2005. Food webs intropical Australian streams: shredders are not scarce. Fresh-water Biology 50:748–769.
Coley, P. D., and J. A. Barone. 1996. Herbivory and plant de-fenses in tropical forests. Annual Review of Ecology and Sys-tematics 27:305–335.
Cornwell, W. K., J. H. C. Cornelissen, K. Amatangelo, E. Dorrepaal,V. T. Eviner, O. Godoy, S. E. Hobbie, B. Hoorens, H. Kurokawa,N. Pérez-Harguindeguy, H. N. Quested, L. S. Santiago, D. A.Wardle, I. J. Wright, R. Aerts, S. D. Allison, P. Van Bodegom,V. Brovkin, A. Chatain, T. V. Callaghan, S. Díaz, E. Garnier,D. E. Gurvich, E. Kazakou, J. A. Klein, J. Read, P. B. Reich,N. A. Soudzilovskaia, M. V. Vaieretti, and M. Westoby. 2008.Plant species traits are the predominant control on litter de-
composition rates within biomes worldwide. Ecology Letters11:1065–1071.
Covich, A. P. 1988. Geographical and historical comparisons ofneotropical streams: biotic diversity and detrital processingin highly variable habitats. Journal of the North AmericanBenthological Society 7:361–386.
Cowan, C. A., M. W. Oswood, C. A. Buttimore, and P. W. Flan-agan. 1983. Processing and macroinvertebrate colonizationof detritus in an Alaskan subarctic stream. Holarctic Ecology6:340–348.
Crowl, T. A., V. Welsh, T. Heartsill-Scalley, and A. P. Covich.2006. Effects of different types of conditioning on rates ofleaf-litter shredding by Xiphocaris elongata, a Neotropicalfreshwater shrimp. Journal of the North American Benthol-ogical Society 25:198–208.
Dobson, M., A. Magana, J. M. Mathooko, and F. K. Ndegwa.2002. Detritivores in Kenyan highland streams: more evi-dence for the paucity of shredders in the tropics? Freshwa-ter Biology 47:909–919.
Dudgeon, D. 1982. An investigation of physical and biologicalprocessing of two species of leaf litter in Tai Po Kau foreststream, New Territories, Hong Kong. Archiv für Hydrobio-logie 96:1–32.
Dudgeon, D., and K. K. Y. Wu. 1999. Leaf litter in a tropicalstream: food or substrate for macroinvertebrates? Archiv fürHydrobiologie 146:65–82.
Ferreira, V., and E. Chauvet. 2011. Future increase in tempera-ture more than decrease in litter quality can affect microbiallitter decomposition in streams. Oecologia (Berlin) 167:279–291.
Fisher, S. G., and G. E. Likens. 1973. Energy flow in BearBrook, New Hampshire: an integrative approach to streamecosystem metabolism. Ecological Monographs 43:421–439.
Gessner, M. O., and E. Chauvet. 1994. Importance of streammicrofungi in controlling breakdown rates of leaf litter. Ecol-ogy 75:1807–1817.
Gessner, M. O., E. Chauvet, and M. Dobson. 1999. A perspec-tive on leaf litter breakdown in streams. Oikos 85:377–384.
Gonçalves, J. F., M. A. S. Graça, and M. Callisto. 2006. Leaf-litter breakdown in 3 streams in temperate, Mediterranean,and tropical Cerrado climates. Journal of the North Ameri-can Benthological Society 25:344–355.
Graça, M. A. S., F. Bärlocher, and M. O. Gessner. 2005. Methodsto study litter decomposition: a practical guide. Springer, Dor-drecht, The Netherlands.
Graça, M. A. S., and C. Cressa. 2010. Leaf quality of some tropi-cal and temperate tree species as food resource for streamshredders. International Review of Hydrobiology 95:27–41.
Graça, M. A. S., C. Cressa, M. O. Gessner, M. J. Feio, K. A. Callies,and C. Barrios. 2001. Food quality, feeding preferences, sur-vival and growth of shredders from temperate and tropicalsteams. Freshwater Biology 46:947–957.
Handa, I. T., R. Aerts, F. Berendse, M. P. Berg, A. Bruder,O. Butenschoen, E. Chauvet, M. O. Gessner, J. Jabiol, M. Mak-konen, B. G. McKie, B. Malmqvist, E. Peeters, S. Scheu,B. Schmidt, J. V. Ruijven, V. C. A. Vos, and S. Hättenschwiler.2014. Consequences of biodiversity loss for litter decomposi-tion across biomes. Nature 509:218–221.
000 | Tropical altitudinal litter-breakdown rates T. Jinggut and C. M. Yule
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions

Hieber, M., and M. O. Gessner. 2002. Contribution of streamdetritivores, fungi and bacteria to leaf breakdown based onbiomass estimates. Ecology 83:1026–1038.
IPCC (International Panel on Climate Change). 2007. Climatechange 2007: the physical science basis. Cambridge Univer-sity Press, Cambridge, UK.
Irons, J. G., M. W. Oswood, R. J. Stout, and C. M. Pringle.1994. Latitudinal patterns in leaf litter breakdown: is tem-perature really important? Freshwater Biology 32:401–411.
Iwata, T., S. Nakano, and M. Inoue. 2003. Impacts of past ri-parian deforestation on stream communities in a tropicalrain forest in Borneo. Ecological Applications 13:461–473.
Jacobsen, D., R. Schultz, and A. Encalada. 1997. Structure anddiversity of stream invertebrate assemblages: the influenceof temperature with altitude and latitude. Freshwater Biol-ogy 38:247–261.
Kitayama, K. 1992. An altitudinal transect study of the vegeta-tion on Mount Kinabalu, Borneo. Vegetatio 102:149–171.
Kitayama, K., and S. Aiba. 2002. Ecosystem structure and pro-ductivity of tropical rain forests along altitudinal gradientswith contrasting soil phosphorus pools of Mount Kinabalu,Borneo. Journal of Ecology 90:37–51.
Kurokawa, H., and T. Nakashizuka. 2008. Leaf herbivory anddecomposability in a Malaysian tropical rain forest. Ecology89:2645–2656.
Leroy, C. J., and J. C. Marks. 2006. Litter quality, stream char-acteristics and litter diversity influence decomposition ratesand macroinvertebrates. Freshwater Biology 51:605–617.
MacKenzie, R. A., T. N. Wiegner, F. Kinslow, N. Cormier,and A. M. Strauch. 2013. Leaf-litter inputs from an invasivenitrogen-fixing tree influence organic-matter dynamics andnitrogen inputs in a Hawaiian river. Freshwater Science 32:1036–1052.
Makkonen, M., M. P. Berg, I. T. Handa, S. Hättenschwiler, J. vanRuijven, P. M. van Bodegom, and R. Aerts. 2012. Highly con-sistent effects of plant litter identity and functional traits ondecomposition across a latitudinal gradient. Ecology Letters15:1033–1041.
Martínez, A., A. Larrañaga, J. Pérez, E. Descals, and J. Pozo.2014. Temperature affects leaf litter decomposition in loworder forest streams: field and microcosm approaches. FEMSMicrobiology Ecology 87:257–267.
Masese, F. O., N. Kitaka, J. Kipkemboi, G. M. Gettel, K. Irvine,and M. E. McClain. 2014. Macroinvertebrate functionalfeeding groups in Kenyan highland streams: evidence for adiverse shredder guild. Freshwater Science 33:435–450.
Mathuriau, C., and E. Chauvet. 2002. Breakdown of leaf litterin a Neotropical stream. Journal of the North American Ben-thological Society 21:384–396.
Ostrofsky, M. L. 1997. Relationship between chemical charac-teristics of autumn-shed leaves and aquatic processing rates.Journal of the North American Benthological Society 16:750–759.
Reice, R. C., and K. W. Cummins. 1974. Leaf processing in awoodland stream. Archiv für Hydrobiologie 116:1271–1281.
Rincón, N. J., and R. Santelloco. 2009. Aquatic fungi associatedwith decomposing Ficus sp. leaf litter in a Neotropical stream.Journal of the North American Benthological Society 28:416–425.
Sinsabaugh, R. L., D. L. Moorhead, and A. E. Linkins. 1994.The enzymic basis of plant litter decomposition: emergenceof an ecological process. Applied Soil Ecology 1:97–111.
Sinsabaugh, R. L., and J. J. F. Shah. 2012. Ecoenzymatic stoichi-ometry and ecological theory. Annual Review of Ecology,Evolution, and Systematics 43:313–343.
Slik, J. W. F., A. D. Poulsen, P. S. Ashton, C. H. Cannon,K. A. O. Kartawinata, I. Lanniari, H. Nagamasu, M. Nakagawa,M. G. L. Van Nieuwstadt, J. Payne, A. P. Saridan, K. Sidiyasa,R. W. Verburg, C. O. Webb, and P. Wilkie. 2003. A floristicanalysis of the lowland dipterocarp forests of Borneo. Journalof Biogeography 30:1517–1531.
Talbot, J. M., and K. K. Treseder. 2012. Interactions amonglignin, cellulose, and nitrogen drive litter chemistry–decayrelationships. Ecology 93:345–354.
Tanner, E. V. J., P. M. Vitousek, and E. Cuevas. 1998. Experi-mental investigation of nutrient limitation of forest growthon wet tropical mountains. Ecology 79:10–22.
Taylor, B. R., and E. Chauvet. 2014. Relative influence of shred-ders and fungi on leaf litter decomposition along a riveraltitudinal gradient. Hydrobiologia 721:239–250.
Wantzen, K. M., C. M. Yule, J. M. Mathooko, and C. M. Pringle.2008. Organic matter processing in tropical streams. Pages 43–63 in D. Dudgeon (editor). Tropical stream ecology. Elsevier,Dordrecht, The Netherlands.
Webster, J. R., and J. L. Meyer. 1997. Stream organic matterbudgets. Journal of the North American Benthological Soci-ety 16:3–161.
Wright, M. S., and A. P. Covich. 2005. Relative importance ofbacteria and fungi in a tropical headwater stream: leaf de-composition and invertebrate feeding preference. MicrobialEcology 49:536–546.
Yule, C. M. 1996. Trophic relationships and food webs of thebenthic invertebrate fauna of two aseasonal streams on Bou-gainville Island, Papua New Guinea. Journal of Tropical Ecol-ogy 12:517–534.
Yule, C. M., M. Y. Leong, K. C. Liew, L. Ratnarajah, K.Schmidt, H. M. Wong, R. G. Pearson, and L. Boyero. 2009.Shredders in Malaysia: abundance and richness are higherin cool upland tropical streams. Journal of the North Amer-ican Benthological Society 28:404–415.
Yule, C. M., and H. S. Yong (editors). 2004. Freshwater inverte-brates of the Malaysian region. Academy of Sciences, KualaLumpur, Malaysia.
Zhang, D., D. Hui, Y. Luo, and G. Zhou. 2008. Rates of litterdecomposition in terrestrial ecosystems: global patterns andcontrolling factors. Journal of Plant Ecology 1:85–93.
Volume 34 June 2015 | 000
This content downloaded from 118.139.129.89 on Thu, 26 Mar 2015 02:41:28 AMAll use subject to JSTOR Terms and Conditions




















