
Diverse communities of arbuscular mycorrhizal fungi inhabit sites with very high altitude in Tibet...
15
RESEARCH ARTICLE Diverse communities of arbuscular mycorrhizal fungi inhabit sites with very high altitude inTibet Plateau Yongjun Liu 1,2 , Junxia He 1 , Guoxi Shi 1 , Lizhe An 1 , Maarja O ¨ pik 3 & Huyuan Feng 1 1 Key Laboratory of Arid and Grassland Ecology of Ministry of Education, School of Life Sciences, Lanzhou University, Lanzhou, China; 2 College of Life Science and Engineering, Northwest University for Nationalities, Lanzhou, China; and 3 Department of Botany, Institute of Ecology and Earth Sciences, University of Tartu, Tartu, Estonia Correspondence: Huyuan Feng, Key Laboratory of Arid and Grassland Ecology of Ministry of Education, School of Life Sciences, Lanzhou University, Room 322, Lanzhou 730000, China. Tel: 186 931 891 2560; fax: 186 931 891 2537; e-mail: [email protected] Received 24 January 2011; revised 6 June 2011; accepted 19 June 2011. Final version published online 18 July 2011. DOI:10.1111/j.1574-6941.2011.01163.x Editor: Philippe Lemanceau Keywords arbuscular mycorrhiza; Dracocephalum heterophyllum; pioneer species; cold- dominated ecosystem; high altitude; Tibet Plateau. Abstract Diversity of arbuscular mycorrhizal fungi (AMF) is well studied in many ecosystems, but little is known about AMF in cold-dominated regions with very high altitude. Here, we examined AMF communities associated with two plant species in the Tibet Plateau. Roots and rhizosphere soils of Dracocephalum heterophyllum (pioneer species) and Astragalus polycladus (late-successional spe- cies) were sampled at five sites with altitude from 4500 to 4800 m a.s.l. A total of 21 AMF phylotypes were identified from roots and spores following cloning and sequencing of 18S rRNA gene, including eight new phylotypes and one new family-like clade. More AMF phylotypes colonized root samples of D. hetero- phyllum (5.4 0.49) than of A. polycladus (1.93 0.25). Vegetation coverage was the most important factor influencing AMF community composition in roots. Globally infrequent phylotype Glo-B2 in Glomus group B was the most dominant in roots, followed by globally frequent phylotype Glo-A2 related to Glomus fasciculatum/intraradices group. Our findings suggest that a diverse AMF flora is present in the Tibet Plateau, comprising both potentially habitat-selective and generalist fungi. Introduction The vast majority of terrestrial plants form mutualistic associations with arbuscular mycorrhizal fungi (AMF), which belong to the phylum Glomeromycota with about 200 described species (Smith & Read, 2008). AMF can increase nutrient uptake as well as improve the stress tolerance of their hosts (Smith et al., 2003; Aroca et al., 2007). AMF also exert strong effects on the plant communities (van der Heijden et al., 1998; Vogelsang et al., 2006). In particular, the diversity of AMF and plants can be inter-related (Johnson et al., 2004; Landis et al., 2004). A high number of AMF taxa has been found in tropical and boreal forests (Husband et al., 2002; O ¨ pik et al., 2008, 2009), whereas the AMF richness is often low in disturbed and degraded habitats (O ¨ pik et al., 2006; Tian et al., 2009). Even where richness remains high, environmental factors such as pH have been shown to determine community composition even at relatively small spatial scales (Dumbrell et al., 2010). At a much smaller, within sample site scale, studies indicate that AMF have different levels of host preference or specifi- city (e.g. Husband et al., 2002; Vandenkoornhuyse et al., 2003), suggesting that the host plant is also an important factor in assembling the AMF community. Thus, it is likely that the diversity and distribution of AMF are codetermined by the biotic and abiotic factors, and that the balance between the two depends on spatial scale and local factors. Diversity of AMF has been well studied in grasslands, forests, and some other ecosystems of mainly the temperate climatic zone (O ¨ pik et al., 2010). Little is known about AMF diversity in regions with very high altitude (above 3500 m a.s.l.; but see Gai et al., 2009 and some studies in montane sites with altitude below 2000 m a.s.l.: for example Sy ´korov´ a et al., 2007; Wu et al., 2007), where the habitats are often cold-dominated. Kyt¨ oviita & Ruotsalainen (2007) suggested that AMF could not improve nutrient capture of host plants in low temperature habitats, but this does not imply that AMF in such habitats cannot persist as plant symbionts. On the contrary, a high diversity of AMF spores FEMS Microbiol Ecol 78 (2011) 355–365 c 2011 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved MICROBIOLOGY ECOLOGY
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Diverse communities of arbuscular mycorrhizal fungi inhabit sites with very high altitude in Tibet...

R E S E A R C H A R T I C L E
Diverse communities ofarbuscularmycorrhizal fungi inhabit siteswithveryhighaltitude inTibetPlateauYongjun Liu1,2, Junxia He1, Guoxi Shi1, Lizhe An1, Maarja Opik3 & Huyuan Feng1
1Key Laboratory of Arid and Grassland Ecology of Ministry of Education, School of Life Sciences, Lanzhou University, Lanzhou, China; 2College of Life
Science and Engineering, Northwest University for Nationalities, Lanzhou, China; and 3Department of Botany, Institute of Ecology and Earth Sciences,
University of Tartu, Tartu, Estonia
Correspondence: Huyuan Feng, Key
Laboratory of Arid and Grassland Ecology of
Ministry of Education, School of Life Sciences,
Lanzhou University, Room 322, Lanzhou
730000, China. Tel: 186 931 891 2560;
fax: 186 931 891 2537;
e-mail: [email protected]
Received 24 January 2011; revised 6 June 2011;
accepted 19 June 2011.
Final version published online 18 July 2011.
DOI:10.1111/j.1574-6941.2011.01163.x
Editor: Philippe Lemanceau
Keywords
arbuscular mycorrhiza; Dracocephalum
heterophyllum; pioneer species; cold-
dominated ecosystem; high altitude;
Tibet Plateau.
Abstract
Diversity of arbuscular mycorrhizal fungi (AMF) is well studied in many
ecosystems, but little is known about AMF in cold-dominated regions with very
high altitude. Here, we examined AMF communities associated with two plant
species in the Tibet Plateau. Roots and rhizosphere soils of Dracocephalum
heterophyllum (pioneer species) and Astragalus polycladus (late-successional spe-
cies) were sampled at five sites with altitude from 4500 to 4800 m a.s.l. A total of 21
AMF phylotypes were identified from roots and spores following cloning and
sequencing of 18S rRNA gene, including eight new phylotypes and one new
family-like clade. More AMF phylotypes colonized root samples of D. hetero-
phyllum (5.4� 0.49) than of A. polycladus (1.93� 0.25). Vegetation coverage was
the most important factor influencing AMF community composition in roots.
Globally infrequent phylotype Glo-B2 in Glomus group B was the most dominant
in roots, followed by globally frequent phylotype Glo-A2 related to Glomus
fasciculatum/intraradices group. Our findings suggest that a diverse AMF flora is
present in the Tibet Plateau, comprising both potentially habitat-selective and
generalist fungi.
Introduction
The vast majority of terrestrial plants form mutualistic
associations with arbuscular mycorrhizal fungi (AMF),
which belong to the phylum Glomeromycota with about 200
described species (Smith & Read, 2008). AMF can increase
nutrient uptake as well as improve the stress tolerance of
their hosts (Smith et al., 2003; Aroca et al., 2007). AMF also
exert strong effects on the plant communities (van der
Heijden et al., 1998; Vogelsang et al., 2006). In particular,
the diversity of AMF and plants can be inter-related
(Johnson et al., 2004; Landis et al., 2004). A high number
of AMF taxa has been found in tropical and boreal forests
(Husband et al., 2002; Opik et al., 2008, 2009), whereas the
AMF richness is often low in disturbed and degraded
habitats (Opik et al., 2006; Tian et al., 2009). Even where
richness remains high, environmental factors such as pH
have been shown to determine community composition
even at relatively small spatial scales (Dumbrell et al., 2010).
At a much smaller, within sample site scale, studies indicate
that AMF have different levels of host preference or specifi-
city (e.g. Husband et al., 2002; Vandenkoornhuyse et al.,
2003), suggesting that the host plant is also an important
factor in assembling the AMF community. Thus, it is likely
that the diversity and distribution of AMF are codetermined
by the biotic and abiotic factors, and that the balance
between the two depends on spatial scale and local factors.
Diversity of AMF has been well studied in grasslands,
forests, and some other ecosystems of mainly the temperate
climatic zone (Opik et al., 2010). Little is known about AMF
diversity in regions with very high altitude (above
3500 m a.s.l.; but see Gai et al., 2009 and some studies in
montane sites with altitude below 2000 m a.s.l.: for example
Sykorova et al., 2007; Wu et al., 2007), where the habitats are
often cold-dominated. Kytoviita & Ruotsalainen (2007)
suggested that AMF could not improve nutrient capture of
host plants in low temperature habitats, but this does not
imply that AMF in such habitats cannot persist as plant
symbionts. On the contrary, a high diversity of AMF spores
FEMS Microbiol Ecol 78 (2011) 355–365 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
MIC
ROBI
OLO
GY
EC
OLO
GY

and phylotypes has been reported in sites with altitude
ranging from 3500 to 5200 m a.s.l. (Gai et al., 2009) and in
a low-Arctic meadow (Pietikainen et al., 2007), respectively.
Nonetheless, it is widely accepted that the major plant–fun-
gal associations in high-altitude and polar regions are
ectomycorrhiza and dark septate endophytes rather than
AM (Gardes & Dahlberg, 1996; Newsham et al., 2009; Gao &
Yang, 2010); this perhaps suggests that most plants in these
regions are not good hosts for AMF, and that AMF should
have a different life-history strategy in these regions. For
example, these fungi would select suitable host plants
(Gardes & Dahlberg, 1996) and the spores might be dor-
mant for a long time (Pietikainen et al., 2007). Even if
diverse AMF communities could be present in the regions
with very high altitude, which AMF species can persist and
which host plants they colonize is still unclear. The evolution
of AMF traits may be highly dependent on abiotic factors
such as soil temperature and hypoxia (Helgason & Fitter,
2009). Specific AMF have been found in extreme habitats
such as geothermal and gypsum soils (Appoloni et al., 2008;
Alguacil et al., 2009).
Therefore, we predict that AMF species and their traits in
regions with very high altitude may be different from those
in ecosystems with more moderate abiotic conditions. To
better understand the biodiversity of AMF and its determi-
nants in the regions with very high altitude, we chose to
study AMF associated with two plant species with different
life-history strategies in the Tibet Plateau of China (mean
altitude over 4500 m a.s.l.). We used molecular techniques to
describe AMF communities to address the following ques-
tions: (1) what is the AMF diversity in the roots of a pioneer
and a late-successional plant species in the central Tibet
Plateau? (2) Which environmental factors determine the
richness and community composition of AMF associated
with these two plant species? (3) Are the AMF taxa in this
region different from those in other ecosystems?
Materials and methods
Study sites and sampling
This study was conducted in the central Tibet Plateau, where
the average altitude is above 4500 m a.s.l. and the mean
annual temperature ranges between � 3 and � 7 1C. Five
study sites were chosen along the Qinghai-Tibet highway
from Xidatan to Amdo, in a line running approximately
north-east to south-west (Table 1, Supporting information,
Fig. S1). In each site, a pioneer plant species Dracocephalum
heterophyllum (Lamiaceae) was dominant in the region
nearby the highway (disturbed once due to the construction
of highway 30 years ago, o 50 m from highway), whereas
the natural plant communities (late-successional stage;
4 60 m from highway) were dominated by Cyperaceae,
Caryophyllaceae or leguminous plants such as Astragalus
polycladus (Table 1). Because the plants of Cyperaceae
and Caryophyllaceae are traditionally regarded as little
AM-dependent (Wang & Qiu, 2006; Smith & Read, 2008),
we chose a leguminous species, A. polycladus, as the
Table 1. Sampling sites and their vegetation details
Early-successional plant community Late-successional plant community
Study site
Latitude,
longitude
Altitude
(m a.s.l.)
Vegetation
coverage
Species
richness
Dominant plant
species (coverage)
Vegetation
coverage
Species
richness
Dominant plant
species (coverage)
Xidatan (X)� 351 420N
941 070E
4524 c. 30% 2 D. heterophyllum (c.
25%), Aster alpinus
(c.5%)
c. 45% 10 Androsace tapete (c.15%),
Astragalus polycladus
(c.10%), Arenaria kansuensis
(c.8%)w
Kunlunshan
(K)
351 370N
941 040E
4746 c. 70% 4 D. heterophyllum (c.55%),
Avena fatua (c. 5%),
Poa sp. (c. 5%)
c. 65% 9 Kobresia pygmaea (c.40%)w,
Astragalus polycladus (c.10%)
Wudaoliang
(W)
351 140N
931 050E
4568 c. 30% 2 D. heterophyllum
(c. 20%), Saussurea
wellbyl (c.10%)
c. 15% 1 Astragalus polycladus (c.15%)
Tongtianhe
(T)
331 480N
921 180E
4603 c. 25% 1 D. heterophyllum (c. 25%) c. 65% 10 K. pygmaea (c.30%)w, Stipa
aliena (c.10%), Astragalus
polycladus (c.8%)
Amdo (A) 321 100N
911 410E
4773 c. 75% 6 D. heterophyllum (c.50%),
Potentilla bifurca (c.7%),
Elymus nutans (c.5%)
c. 75% 11 K. pygmaea (c.35%)w,
Potentilla parvifolia (c.8%),
Arenaria kansuensis (c.5%)w,
Astragalus polycladus (c.4%)
�Site codes are shown in parentheses.wPotentially non-AM species of Caryophyllaceae or Cyperaceae (Smith & Read, 2008).
FEMS Microbiol Ecol 78 (2011) 355–365c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
356 Y. Liu et al.

representative of late-successional plant species. Before
sampling, the vegetation coverage, plant species coverage
and species richness were estimated on three plots of 1� 1 m
at each plant community using Sutherland’s method
(Sutherland, 1996). Vegetation coverage and plant species
richness varied in different sites, with low vegetation cover-
age in sites of Xidatan and Wudaoliang due to serious
permafrost degradation and rodent damage. More details
of the sampling sites are presented in Table 1.
Sampling was conducted in the beginning of August 2008.
In each site, three individuals of D. heterophyllum and
A. polycladus were randomly sampled (30 samples in total:
2 plant species� 3 plant individuals� 5 sampling sites).
Plants together with their rhizosphere soil were carefully
excavated and placed in plastic bags. Roots were separated
from each plant individual, washed with tap water and cut
into approximately 1-cm-long fragments. Due to the limited
quantity of roots, we randomly picked about 10 root
fragments from samples of the same plant species and
pooled these in order to verify the AM status of the study
plant species following the method of McGonigle et al.
(1990). The remaining roots (30 samples) were stored at
� 20 1C until DNA extraction. Soils (30 samples) were air-
dried and used for AMF spore extraction and soil chemical
analyses.
Spore extraction and soil chemical analysis
AMF spores were extracted from 30 soil samples of 100 g by
wet sieving (a pair of sieves: 750 and 38-mm mesh) and
sucrose centrifugation (Brundrett et al., 1994), and the spore
densities were counted. Spores from soil samples of the same
plant species in each site were pooled (resulting in 10 pooled
spore samples) and then grouped according to spore mor-
phology and color using a dissecting microscope. Spore
number of each morphotype in each pooled sample was
counted, and the spore morphotypes were further identified
using the molecular method as described below.
Soil total nitrogen, organic carbon and available (Olsen)
phosphorus contents were analyzed using the methods
described previously (Liu et al., 2009). Soil pH was mea-
sured in 1 M KCl (10 g soil in a 50 mL solution).
DNA extraction and amplification
For each root sample, about 30 root fragments (c. l-cm
lengths) were randomly picked and washed four times with
sterilized distilled water. Root DNA was extracted using
Plant DNA Extraction Kit (Tiangen Biotech, China) follow-
ing the manufacturer’s instructions, and then diluted 1 : 10
with ddH2O to be used as PCR template. For spore samples,
up to three clean and healthy-looking spores of each
morphotype per pooled sample were transferred into micro-
centrifuge tubes and vortexed at maximum speed. Spores
were further rinsed four times with sterilized distilled water.
Single spores were transferred into tubes with 10 mL ddH2O,
and crushed with forceps under the dissecting microscope.
A total of 120 single spores were incubated at 65 1C for
30 min and the liquid used as DNA template in PCR.
All root- and spore-derived DNA samples were subjected
to nested PCR. The first PCR reaction was performed with
the universal fungal primers GeoA2 and Geo11 to amplify a
c. 1.8 kb fragment of the 18S rRNA gene (Schwarzott &
Schußler, 2001). PCR reactions were carried out in a final
volume of 25 mL with 2 mL template and 0.5mM of each
primer using the Pfu PCR mastermix system (Tiangen
Biotech) with the following cycling conditions: 94 1C for
120 s; 30� (94 1C for 30 s; 59 1C for 60 s and 72 1C for
150 s); and 72 1C for 600 s. Successful products of the first
amplification were diluted 1 : 100 and 2 mL of this dilution
was used as a template in the second PCR with universal
eukaryotic primer NS31 (Simon et al., 1992) and AMF-
specific primer AML2 (Lee et al., 2008). Most first PCR
products of spores could not be detected on agarose gel, and
thus 1 mL of the first PCR product was used as template for
the second PCR. The NS31-AML2 primer combination was
used because amplification using AML1-AML2 (Lee et al.,
2008) was unsuccessful in our lab. PCR reactions were run
with the same conditions as described above using the
following cycling conditions: 94 1C for 120 s; 30� (94 1C
for 30 s; 58 1C for 60 s and 72 1C for 80 s); and 72 1C for
600 s. All the PCR reactions were run on a GeneAmps PCR
system 2700 (Applied Biosystems). PCR products were
examined on a 1.5% (w/v) agarose gel with ethidium
bromide staining.
Cloning, restriction fragment lengthpolymorphism (RFLP) typing and sequenceanalysis
All root-derived PCR products were used for constructing
clone libraries (30 in total). PCR products of expected
length (c. 560 bp) were purified using the Gel and PCR clear
up System (Promega). After an A-tailing procedure, the
DNA products were ligated into pGEM-T vector (Promega)
and cloned into Escherichia coli DH5a (Tiangen Biotech)
according to the manufacturer’s instructions. For each clone
library, 50 randomly selected white clones were immersed in
30 mL ddH2O and subjected to three cycles of freezing and
thawing for preparation of plasmid templates. Inserts were
reamplified with primers NS31/AML2 with the same PCR
conditions as above and double-digested with the restriction
enzymes HinfI and Hin1II (MBI, Lithuania). Digested
products were examined on a 2.5% (w/v) agarose gel.
One representative of each root-derived RFLP type was
sequenced with primer T7 by Major Biotech Company
(Shanghai, China).
FEMS Microbiol Ecol 78 (2011) 355–365 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
357Diverse AM fungi in Tibet Plateau

The spore-derived PCR products were directly subjected
to RFLP typing as described above. One representative of
each spore-derived RFLP type was cloned as described
above. One confirmed positive clone was sequenced from
each spore-derived clone library with primer T7 as above.
Sequences were edited using the CONTIGEXPRESS module of
VECTOR NTI Suite 6.0 (InforMax Inc., MD), and submitted to
BLAST search against public nucleotide sequence databases
(Altschul et al., 1997). All AMF sequences have been
submitted to GenBank database under the accession num-
bers GU238334–GU238408 and HQ610612–HQ610619.
The representative sequences obtained in this study, the
most closely related sequences from GenBank and the
representative sequences of major families of Glomero-
mycota were used in the phylogenetic analysis. Sequences
were aligned using CLUSTALX (Thompson et al., 1997) and
manually edited using GENEDOC (Nicholas et al., 1997).
Bayesian (GTR1G1I model) and neighbor-joining
(Kimura 2-parameter model with 1000 bootstrap replica-
tions) phylogenetic analyses were performed using MRBAYES
3.1 (Ronquist & Huelsenbeck, 2003) and MEGA 4.0 (Tamura
et al., 2007), respectively. Bayesian analysis was performed
with two independent runs of Markov chain Monte Carlo
with 106 generations (sampling frequency = 10), and a
conservative burn-in of 250 000 generations (25%) was
chosen.
Statistical data analysis
Due to limited identification success of AMF spores, the
molecular spore identification data are presented, but the
community composition of AMF spores was not analyzed
further. Root-colonizing AMF community composition was
analyzed on the basis of number of clones of each phylotype
in a root sample. The effects of host plant and sampling site
(as independent factors) on AMF richness per root sample
and spore density per soil sample were analyzed using two-
way ANOVA. Differences in AMF richness, spore density and
soil variables between samples were tested using Fisher’s
least significant difference at the 5% level after one-way
ANOVA. All above statistical analyses were performed using
the SPSS 13.0 (SPSS Inc., IL). Sampling effort curves of root-
colonizing AMF richness (Mao Tau) were computed using
ESTIMATES 8.0 (Colwell, 2006).
Before analysis of AMF community composition, raw
data of clone counts were square root transformed in order
to down-weigh the importance of abundant phylotypes. The
dissimilarities of AMF communities among samples were
computed by nonmetric multidimensional scaling (NMDS)
with Bray–Curtis distance using the function ‘isoMDS’ in
the R PACKAGE VEGAN version 1.17–8 (R Development Core
Team, 2010). To explore the relationships between environ-
mental variables and AMF community composition, the
environmental variables (including data of soil properties,
altitude, latitude, longitude, plant species richness and
vegetation coverage; all data except the latitude and long-
itude were square root transformed) were fitted as vectors
onto the NMDS plot using the function ‘envfit’ in the R
PACKAGE VEGAN version 1.17-8 (R Development Core Team,
2010).
We used the online database MaarjAM (http://maarjam.
botany.ut.ee; Opik et al., 2010; status of October 20, 2010) to
compare the AMF phylotypes obtained in this study with
the AMF detected in other ecosystems. Because the
MaarjAM database comprises AMF 18S rRNA gene se-
quences derived from 105 published papers of different
ecosystems, we could determine the detection frequencies
of our phylotypes in different ecosystems. We first used our
sequences to BLAST against the MaarjAM database and
grouped our sequences into the corresponding molecular
virtual taxa with the sequence identity Z98%, and then
searched how many studies had detected the corresponding
molecular virtual taxa.
Results
Spore density and molecular identification ofspores
AMF spore density varied in soil samples, with the highest
and lowest spore density being present in rhizosphere soils
of D. heterophyllum (1007� 77.4 spores per 100 g soil;
mean� SE) and A. polycladus (46.7� 4.1) in Xidatan
(Table 2). Spore density was different between sampling
sites (F = 11.45, Po 0.001), but not between host plant
species (F = 3.14, P = 0.09); site and plant species had a
significant interaction (F = 36.24, Po 0.001).
Amplification of spore-derived DNA was unsuccessful for
most, but not all spore morphotypes (Table 2, Table S1). In
total, 56 out of 120 single spores that were subjected to
molecular analysis (46.7%) were successfully amplified and
subjected to RFLP typing (Table 2). A total of 17 RFLP types
were detected, and one representative of each spore-derived
RFLP type was sequenced. Most spores of the same mor-
photype shared the same RFLP pattern (data not shown), so
that the remaining spores were classified by RFLP typing. A
BLAST search showed that eight sequences belonged to
Glomeromycota (Fig. 1), and nine sequences were related to
Ascomycota, possibly representing sequences of parasitizing
ascomycetes in spores (Hijri et al., 2002).
Molecular identification of AMF in roots
Before molecular analysis, the AM status of the plant species
studied was confirmed by root staining and microscopy,
showing high levels of root AM colonization: 53.5%
and 58.9% of root length was colonized by AMF for
FEMS Microbiol Ecol 78 (2011) 355–365c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
358 Y. Liu et al.

D. heterophyllum and A. polycladus, respectively. Both
arbuscules (12.8%) and vesicles (10.6%) were observed
in the roots of D. heterophyllum; only vesicles (15.7%),
but no arbuscules were observed in the roots of A.
polycladus.
All 30 root samples yielded PCR products of expected
length in the first (c. 1.8 kb) and second PCR reactions
(c. 560 bp). A total of 1500 clones were screened, and 1464
clones containing insert of correct size were submitted to
RFLP typing. One representative clone of each root-derived
RFLP type of each sample group (same plant species in same
site) was sequenced, yielding a total of 117 sequences. The
remaining clones were classified by RFLP typing. BLAST
search showed that 75 sequences had high homology to
members of Glomeromycota, 17 sequences were related to
Ascomycota, one sequence to Basidiomycota, nine sequences
to Metazoa and Cercozoa, and 13 sequences to plants.
Two sequences appeared to be chimeric. Altogether, 822
AMF clones/sequences (56%) were retained in the further
analyses.
Phylogenetic analysis
Both Bayesian and neighbor-joining phylogenetic analyses
of 44 representative AMF sequences from roots and eight
spore-derived sequences revealed 21 sequence groups with
sequence identity Z98% (Fig. 1), hereafter referred to as
phylotypes. Most of the phylotypes had high credibility
values and bootstrap support (4 80%). Seven phylotypes
were related to sequences of described species, six to
uncultured AMF and eight phylotypes (Glo-A1, Glo-A6,
Glo-A8, Aca-1, Clade-1, Clade-2, Clade-3 and Amb-2) were
undescribed previously ( � 97% identity with published
sequences; Fig. 1). Twenty phylotypes belonged to six
families and three orders of Glomeromycota (Fig. 1): nine
Glomeraceae A (Glomus group A), two Glomeraceae B
(Glomus group B), five Diversisporaceae, one Acaulospora-
ceae, one Pacisporaceae and two Ambisporaceae phylotypes.
The remaining phylotype Clade-1 within order Diversispor-
ales could not be placed in the known families and should be
considered as a new family-like clade (Fig. 1). Clade-2 and
Clade-3 were highly divergent from the genus Diversispora,
but were placed with high credibility values in the family
Diversisporaceae (Fig. 1), possibly representing two genera;
Clade-2 and Glomus fulvum (AM418543) belong to the
recently described genus Redeckera (Schußler & Walker,
2010); Clade-3 appears to be a new genus. Phylotypes of
Glomerales were most abundant in roots, representing
approximately 84% of the clones, followed by Diversisporales
(13%) and Archaeosporales (3%). Two phylotypes, Glo-B2
(33% of all AMF clones) and Glo-A2 (30%), were most
abundant in both plant species and all sampling sites.
Specific phylotypes (occurred in one host-site combination)
such as Glo-A3, Glo-A4 and Glo-A5 were often rare (Table
S2). A total of seven phylotypes were detected from
spores, but only two of them (Glo-B1 and Div-2) were also
detected in roots (Table 3). Most frequent phylotypes
among spores were not found in roots: Pac-1 was detected
at three sites mostly from rhizospheres of A. polycladus
(Xidatan, Kunlunshan, Amdo) and Div-3 in three sites of
D. heterophyllum (Kunlunshan, Wudaoliang, Amdo), re-
spectively (Table S1).
Table 2. Spore density and success rate of molecular identification of single spores in rhizosphere soils of each plant species per site
Sampling sites Plant species
Spore density
(spores per 100 g soil)
No. of morphotypes in
each pooled sample
No. of analyzed
single spores
No. of successfully
amplified
single spores�No. of AMF
phylotypes
X DH 1007.3�77.4 a 5 13 9 1
AP 46.7�4.1 f 4 10 5 0
K DH 400.7�82.2 c 6 17 5 3
AP 232.7�43.3 d 7 19 6 1
W DH 86.7�5.3 ef 4 7 4 0
AP 786�36.1 b 3 8 4 2
T DH 61.3�12.7 f 4 8 1 0
AP 205.3�48.6 de 5 10 7 1
A DH 213.3�55.5 de 6 13 7 3
AP 203.3�14.7 de 6 15 8 1
�If the second PCR reaction yielded a c. 560-bp product, it was considered as a successful amplification regardless of whether the PCR production was
an AMF or a non-AMF sequence.
Spore densities are shown with mean� SE (n = 3). Significant differences of spore densities between samples were tested using Fisher’s least significant
difference at the 5% level and indicated by different letters. Please note that spore morphotypes were analyzed following pooling of three replicate
samples per site (in total of 10 samples; see Materials and methods). Site codes are as in Table 1. DH, Dracocephalum heterophyllum; AP, Astragalus
polycladus.
FEMS Microbiol Ecol 78 (2011) 355–365 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
359Diverse AM fungi in Tibet Plateau
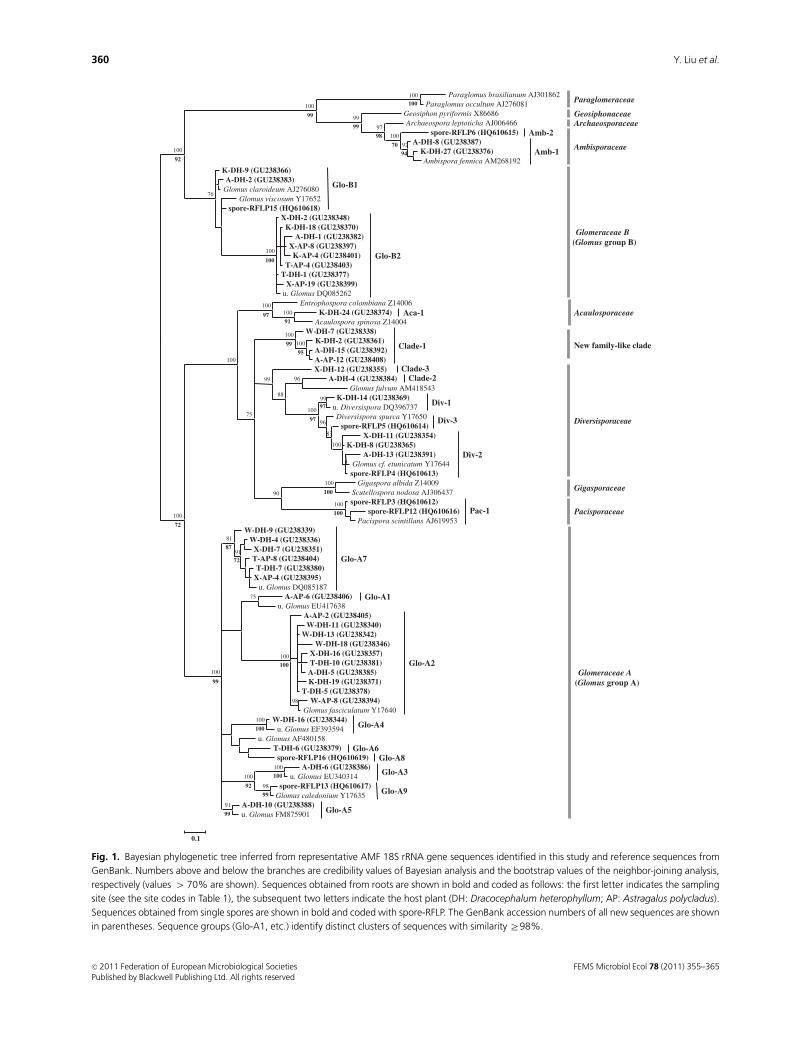
Fig. 1. Bayesian phylogenetic tree inferred from representative AMF 18S rRNA gene sequences identified in this study and reference sequences from
GenBank. Numbers above and below the branches are credibility values of Bayesian analysis and the bootstrap values of the neighbor-joining analysis,
respectively (values 4 70% are shown). Sequences obtained from roots are shown in bold and coded as follows: the first letter indicates the sampling
site (see the site codes in Table 1), the subsequent two letters indicate the host plant (DH: Dracocephalum heterophyllum; AP: Astragalus polycladus).
Sequences obtained from single spores are shown in bold and coded with spore-RFLP. The GenBank accession numbers of all new sequences are shown
in parentheses. Sequence groups (Glo-A1, etc.) identify distinct clusters of sequences with similarity Z98%.
FEMS Microbiol Ecol 78 (2011) 355–365c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
360 Y. Liu et al.

Search against the MaarjAM database
Thirteen out of 21 AMF phylotypes detected in the present
study could be grouped into the corresponding molecular
virtual taxa of the MaarjAM database (Table 3). Eight
phylotypes showed low similarity (93–97%) to the
sequences of this database and those in GenBank
( � 97%). The most dominant phylotype in the present
study (Glo-B2) has been detected by seven studies in the
MaarjAM database (as VT00056, no related species is
known, Table 3), but it was often rare in these studies (e.g.
Opik et al., 2009). Phylotype Glo-A2, second most domi-
nant in this study, has been earlier detected in 37 studies in
three climatic zones (as VT00113 related to Glomus fascicu-
latum group), followed by Glo-A9 (VT00065 related to
Glomus geosporum group, 15 studies) and Div-2 (VT00062,
14 studies) that were infrequent in our study.
Composition and diversity of AMF communitiesin roots
Sampling effort curves clearly indicate that a large propor-
tion of the total AMF diversity colonizing roots was cap-
tured for both plant species (Fig. 2). Fifteen AMF phylotypes
were detected in the roots of D. heterophyllum, whereas only
five phylotypes were detected in the roots of A. polycladus
(Fig. 2, Table S2). Both host plant (F = 386.29, Po 0.001)
and sampling site (F = 36.07, Po 0.001) showed significant
effects on AMF richness, with significant site and host plant
interaction (F = 28.07, Po 0.001). The mean number of
AMF phylotypes per root sample of D. heterophyllum
(5.4� 0.49; mean� SE) was higher than that of A. poly-
cladus (1.93� 0.25). Higher AMF richness was detected in
D. heterophyllum roots at Amdo (8� 0.58) and Kunlunshan
(7� 0) as compared with the other three sites (Fig. 3). The
AMF richness in A. polycladus roots followed a different
pattern: it was highest in Amdo and Tongtianhe (3� 0),
medium in Xidatan (1.67� 0.3) and lowest in Wudaoliang
Table 3. Number of publications in the MaarjAM database (http://maarjam.botany.ut.ee/, Opik et al., 2010) that have earlier reported the AMF
phylotypes of this study
Clones in
roots/from spores
Molecular
virtual taxon� Related species
Temperate
zone
Subtropical
zone
Tropical
zone
Subarctic
zone Total
Glo-A1 18/- – – – – – – –
Glo-A2 250/- VT00113 Glomus fasciculatum 27 8 2 – 37
Glo-A3 5/- VT00064 Glomus constrictum 5 6 1 – 12
Glo-A4 3/- VT00198 Glomus hoi 6 1 0 – 7
Glo-A5 3/- VT00301 – 0 1 0 – 1
Glo-A6 8/- – – – – – – –
Glo-A7 83/- VT00143 – 2 2 1 – 5
Glo-A8 -/1 – – – – – –
Glo-A9 -/1 VT00065 Glomus geosporum 14 1 0 – 15
Glo-B1 49/1 VT00193 Glomus claroideum, G. etunicatum 11 2 0 – 13
Glo-B2 272/- VT00056 – 3 2 2 – 7
Pac-1 -/7 VT00284 Pacispora scintillans 1 1 0 – 2
Aca-1 7/- – – – – – – –
Clade-1 47/- – – – – – – –
Clade-2 18/- – – – – – – –
Clade-3 11/- – – – – – – –
Div-1 8/- VT00060 Diversispora celata, Glomus eburneum 5 1 1 – 7
Div-2 16/2 VT00062 - 8 5 1 – 14
Div-3 -/4 VT00263 Diversispora spurca 1 0 0 – 1
Amb-1 27/- VT00283 Ambispora fennica 1 0 1 1 3
Amb-2 -/1 – – – – – –
�Codes of molecular virtual taxa (VT) in the MaarjAM database (http://maarjam.botany.ut.ee/, Opik et al., 2010).
Fig. 2. Sampling effort curves (Mao Tau) for AMF phylotypes detected in
roots of Dracocephalum heterophyllum and Astragalus polycladus.
FEMS Microbiol Ecol 78 (2011) 355–365 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
361Diverse AM fungi in Tibet Plateau

and Kunlunshan (1� 0) (Fig. 3). Taken as a whole, the total
number of detected AMF phylotypes in roots was highest in
Amdo (11 phylotypes), and lowest in Wudaoliang and
Tongtianhe (both four, Table S2).
NMDS ordination of AMF communities showed a low
stress level (0.09), indicating good representation of phylo-
type composition. NMDS ordination yielded sample group-
ings according to host plant (Fig. 4). There was also a trend
for samples from each site to cluster together regardless of
host plant, except for Kunlunshan and Amdo samples from
D. heterophyllum grouping together, and A. polycladus
samples grouping with samples from the same host from
other sites (Fig. 4). Ten environmental factors fitted as
vectors onto the NMDS plot showed that eight factors were
significantly correlated with the AMF community (Fig. 4);
of these, vegetation coverage was most strongly related to
the AMF community composition (r2 = 0.539, Po 0.001).
Environmental factors
Soil properties varied among samples (Table S3). In parti-
cular, soil available P content, ranging from 0.59 to
2.2 mg kg�1 (Table S3), was affected by sampling site
(F = 120.69, Po 0.001), but not by host plant species
(F = 2.95, P = 0.101), with significant interaction between
them (F = 19.40, Po 0.001). Soil available P (r =� 0.37,
P = 0.05; Pearson’s correlation) and C/N ratio (r =� 0.44,
P = 0.02) were negatively correlated with the vegetation
coverage. None of soil variables were correlated with altitude
(data not shown). Vegetation coverage showed a positive
relationship with plant richness (r = 0.77, Po 0.001) and
altitude (r = 0.63, Po 0.001), suggesting that the vegetation
coverage was higher in high-altitude sites than in low-
altitude sites of this study.
Discussion
Diverse and novel AMF are present in the TibetPlateau
Our results show that a relatively high AMF diversity was
present in the high-altitude alpine environment of the
central Tibet Plateau. A total of 21 AMF phylotypes (from
roots and spores) detected in five study sites of this study is
close to the number of 23 morphospecies detected in the
southern Tibet Plateau (Gai et al., 2009), where the climate
is less extreme than at the sites of this study. Although
comparison of the morphospecies detected by Gai et al.
(2009) and the phylotypes detected in this study is difficult,
a clear difference is the lack of Scutellospora in our study, but
found by Gai and colleagues. This suggests that there is
variation in AMF community composition among locations
in the central and southern Tibet Plateau, also seen among
sampling sites of this study, indicating even higher numbers
of AMF species in this region. Four to 13 AMF phylotypes
were detected per study site, which is comparable to those
known in temperate natural ecosystems (Opik et al., 2006,
2010). Our results are also consistent with a study showing
that at the level of individual plants, the mean number of
AMF taxa detected from plant roots in the low-arctic
meadow habitat was not lower than that in temperate
ecosystems (Pietikainen et al., 2007).
Eight out of 21 AMF phylotypes detected in this study are
not well affiliated to known AMF species or phylotypes.
Fig. 3. The richness of AMF phylotypes detected in root samples in each
site. Mean richness� SE (n = 3) are shown. Abbreviations of sample sites
are as in Table 1. Bars with different letters are significantly different at
P � 0.05.
–1.0 –0.5 0.0 0.5 1.0
–0.5
0.0
0.5
1.0
NMDS axis-1
NM
DS
axis
-2
XKWTA
Fig. 4. Joint plot of NMDS ordination of AMF communities colonizing
roots of different host plant species and the vectors of significant
(P � 0.05) environmental variables across sites. The length of the arrow
is proportional to the strength of correlation between environmental
variable and community dissimilarities. Filled symbols: Dracocephalum
heterophyllum; open symbols: Astragalus polycladus. Abbreviations of
sample sites are as in Table 1.
FEMS Microbiol Ecol 78 (2011) 355–365c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
362 Y. Liu et al.

Among these, Clade-1 and Clade-3 are sufficiently divergent
that they may represent a new family and a new genus
within Diversisporales (Fig. 1). A high proportion of new
AMF taxa detected in this study confirms the view that there
may be many AMF taxa awaiting discovery (Helgason et al.,
2002; Fitter, 2005), in particular in less-studied geographic
regions and biomes (Opik et al., 2010). The most dominant
phylotype (Glo-B2) in this study has been infrequently
detected in other ecosystems (Table 3), suggesting that this
AMF perhaps prefers habitats like those in the present study.
On the other hand, the second most abundant phylotype
was a globally widespread taxon related to G. fasciculatum/
intraradices group (Glo-A2; Table 3; Opik et al., 2006, 2010).
These findings suggest that the AMF communities in cold-
dominated environments in the Tibet Plateau include some
unique taxa and have also some taxa in common with more
moderate habitats. The unique taxa in this system may
include AMF specialized to this extreme habitat with
possibly specific functional properties. For example, Ambis-
pora fennica (Amb-1; Fig. 1), detected in two sites in this
study, may prefer cold habitats because this species was first
isolated from subarctic region in Finland (621300N; Walker
et al., 2007) and has been rarely detected in plant roots in
other climatic regions (Table 3).
AMF richness patterns
Our study system revealed a unique pattern: more AMF
phylotypes colonized roots of early-successional D. hetero-
phyllum in sites disturbed 30 years ago than roots of
A. polycladus in undisturbed vegetation (16 and five phylo-
types, respectively). The same trend was apparent at the level
of individual root samples, whereby the phylotype number
was higher for D. heterophyllum (5.4� 0.49 per sample)
than for A. polycladus (1.93� 0.25). It is generally held that
disturbance affects AMF diversity negatively (e.g. Opik et al.,
2006; Tian et al., 2009). Unfortunately, our study system
does not allow disentangling the effects of disturbance/
successional stage and host plant, because D. heterophyllum
occurs almost exclusively in formerly disturbed vegetation,
while A. polycladus occurs only in late-successional vegeta-
tion. Therefore, we cannot make firm conclusions about the
disturbance effect on the AMF richness. We can still say that
there were abundant arbuscules in the roots of D. hetero-
phyllum, but not A. polycladus, suggesting active symbiosis
in the first case, but not the latter. Spore density and
richness, however, did not differ between vegetation of
different disturbance histories, suggesting that there is no
effect of differing vegetation composition or disturbance
history. At the same time, only two of the AMF phylotypes
were detected both from spores and (rarely) in roots, which
might mean that different factors influence sporulating and
root-colonizing fractions of AMF communities, resulting in
different richness estimates. Clearly, more data are needed,
using both molecular and morphological methods about
ecosystems of different successional stages in this region,
and only then can we fully understand the ecological
importance and traits of AMF in this unique ecosystem.
In fact, natural communities in central Tibet Plateau are
dominated by Kobresia sedges that are known to host diverse
communities of ectomycorrhizal fungi in Himalayan mea-
dows (Gao & Yang, 2010), but can also exhibit AMF
colonization (Gai et al., 2006). Sedges can show quite high
AM colonization levels and positive growth response to
AMF in extreme habitats such as ultramafic soils (Lagrange
et al., 2011). Whether ectomycorrhizal fungi are dominant
over AMF in late-successional plant species in these habitats,
and AMF predominate in plants of early succession, remains
to be clarified in future studies.
We found that vegetation coverage was an important
factor influencing the AMF community composition. The
observed higher AMF richness in sites with higher vegeta-
tion coverage suggests that decrease in vegetation coverage
could decrease AMF diversity (Tian et al., 2009). Also, other
soil factors such as available P and organic C were related
with AMF community composition (Fig. 4), in concordance
with previous studies, showing that abiotic factors may
influence the patterns of AMF communities (Lekberg et al.,
2007; Dumbrell et al., 2010). Nonetheless, it is difficult to
separate environmental selection from effect of host plants
and plant communities on AMF (Schechter & Bruns, 2008),
because these organisms are always codetermined by biotic
and abiotic factors (Bever et al., 2001).
In conclusion, the present study is the first investigation
of the molecular diversity of AMF in the Tibet Plateau of
China. A high AMF diversity with many new taxa detected
in this region suggests that there are unique AMF in cold
ecosystems with very high altitude as well as generalist fungi
common in many types of ecosystems. AMF communities
colonizing roots varied in different plant species, with more
AMF phylotypes inhabiting the roots of pioneer plant
species than late-successional plant, suggesting that the
pioneer plant may be a better host for AMF in the Tibet
Plateau. Because the vegetation area in the Tibet Plateau has
decreased sharply in recent decades due to anthropogenic
disturbances and climate change (Harris, 2010), evaluation
of the AMF diversity patterns as well as the ecological traits
of AMF in this region is necessary if protection and
restoration of degraded regions is to be effective.
Acknowledgements
We are grateful to Prof. Tuo Chen and Dr Gaosen Zhang
from Cold and Arid Regions Environmental and Engineering
Research Institute, Chinese Academy of Sciences, and
Prof. Shiweng Li from Lanzhou Jiaotong Unviersity for
FEMS Microbiol Ecol 78 (2011) 355–365 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
363Diverse AM fungi in Tibet Plateau

facilitating the sample collection. We owe particular grati-
tude to Dr Thorunn Helgason at University of York for
constructive comments and valuable suggestions on the
manuscript. This research was supported by National Nat-
ural foundation of China (30870438, 40930533), The
Major Project of Cultivating New Varieties of Transgenic
Organisms (2009ZX08009-029B), State Key Laboratory of
Frozen Soil Engineering, Chinese Academy of Sciences
(SKLFSE200901), PhD Programs Foundation of Ministry
of Education of China (2010021111002), Estonian Science
Foundation (7738, SF0180098s08), 7th European Commu-
nity Framework Programme (PERG03-GA-2008-231034)
and the European Regional Development Fund (Centre of
Excellence FIBIR).
References
Alguacil MM, Roldan A & Torres MP (2009) Assessing the
diversity of AM fungi in arid gypsophilous plant communities.
Environ Microbiol 11: 2649–2659.
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W
& Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucleic Acids
Res 25: 3389–3402.
Appoloni S, Lekberg Y, Tercek MT, Zabinski CA & Redecker D
(2008) Molecular community analysis of arbuscular
mycorrhizal fungi in roots of geothermal soils in Yellowstone
National Park (USA). Microb Ecol 56: 649–659.
Aroca R, Porcel R & Ruiz-Lozano JM (2007) How does
arbuscular mycorrhizal symbiosis regulate root hydraulic
properties and plasma membrane aquaporins in Phaseolus
vulgaris under drought, cold or salinity stresses? New Phytol
173: 808–816.
Bever JD, Schultz PA, Pringle A & Morton JB (2001) Arbuscular
mycorrhizal fungi: more diverse than meets the eye, and the
ecological tale of why. Bioscience 51: 923–931.
Brundrett M, Melville L & Peterson L (1994) Practical Methods in
Mycorrhiza Research. Mycologue Publication, University of
Guelph, Canada.
Colwell RK (2006) EstimateS: statistical estimation of species
richness and shared species from samples. Version 8. Available
at purl.oclc.org/estimates.
Dumbrell AJ, Nelson M, Helgason T, Dytham C & Fitter AH
(2010) Relative roles of niche and neutral processes in
structuring a soil microbial community. ISME J 4: 337–345.
Fitter AH (2005) Darkness visible: reflections on underground
ecology. J Ecol 93: 231–243.
Gai JP, Cai XB, Feng G, Christie P & Li XL (2006) Arbuscular
mycorrhizal fungi associated with sedges on the Tibetan
plateau. Mycorrhiza 16: 151–157.
Gai JP, Christie P, Cai XB, Fan JQ, Zhang JL, Feng G & Li XL
(2009) Occurrence and distribution of arbuscular mycorrhizal
fungal species in three types of grassland community of the
Tibetan Plateau. Ecol Res 24: 1345–1350.
Gao Q & Yang ZL (2010) Ectomycorrhizal fungi associated with
two species of Kobresia in an alpine meadow in the eastern
Himalaya. Mycorrhiza 20: 281–287.
Gardes M & Dahlberg A (1996) Mycorrhizal diversity in arctic
and alpine tundra: an open question. New Phytol 133:
147–157.
Harris RB (2010) Rangeland degradation on the Qinghai-Tibetan
plateau: a review of the evidence of its magnitude and causes.
J Arid Environ 74: 1–12.
Helgason T & Fitter AH (2009) Natural selection and the
evolutionary ecology of the arbuscular mycorrhizal fungi
(Phylum Glomeromycota). J Exp Bot 60: 2465–2480.
Helgason T, Merryweather JW, Denison J, Wilson P, Young JPW
& Fitter AH (2002) Selectivity and functional diversity in
arbuscular mycorrhizas of co-occurring fungi and plants from
a temperate deciduous woodland. J Ecol 90: 371–384.
Hijri M, Redecker D, Petetot JAM, Voigt K, Wostemeyer J &
Sanders IR (2002) Identification and isolation of two
ascomycete fungi from spores of the arbuscular mycorrhizal
fungus Scutellospora castanea. Appl Environ Microb 68:
4567–4573.
Husband R, Herre EA, Turner SL, Gallery R & Young JPW (2002)
Molecular diversity of arbuscular mycorrhizal fungi and
patterns of host association over time and space in a tropical
forest. Mol Ecol 11: 2669–2678.
Johnson D, Vandenkoornhuyse PJ, Leake JR, Gilbert L, Booth RE,
Grime JP, Young JPW & Read DJ (2004) Plant communities
affect arbuscular mycorrhizal fungal diversity and community
composition in grassland microcosms. New Phytol 161:
503–515.
Kytoviita MM & Ruotsalainen AD (2007) Mycorrhizal benefit in
two low arctic herbs increases with increasing temperature. Am
J Bot 94: 1309–1315.
Lagrange A, Ducousso M, Jourand P, Majorel C & Amir H (2011)
New insights into the mycorrhizal status of Cyperaceae from
ultrmafic soils in New Caledonia. Can J Microbiol 57: 21–28.
Landis FC, Gargas A & Givnish TJ (2004) Relationships among
arbuscular mycorrhizal fungi, vascular plants and environ-
mental conditions in oak savannas. New Phytol 164:
493–504.
Lee J, Lee S & Young JPW (2008) Improved PCR primers for the
detection and identification of arbuscular mycorrhizal fungi.
FEMS Microbiol Ecol 65: 339–349.
Lekberg Y, Koide RT, Rohr JR, Aldrich-Wolfe L & Morton JB
(2007) Role of niche restrictions and dispersal in the
composition of arbuscular mycorrhizal fungal communities.
J Ecol 95: 95–105.
Liu Y, He L, An LZ, Helgason T & Feng HY (2009) Arbuscular
mycorrhizal dynamics in a chronosequence of Caragana
korshinskii plantations. FEMS Microbiol Ecol 67: 81–92.
McGonigle TP, Miller MH, Evans DG, Fairchild GL & Swan JA
(1990) A new method which gives an objective measure of
colonization of roots by vesicular-arbuscular mycorrhizal
fungi. New Phytol 115: 495–501.
FEMS Microbiol Ecol 78 (2011) 355–365c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
364 Y. Liu et al.

Newsham KK, Upson R & Read DJ (2009) Mycorrhizas and dark
septate root endophytes in polar regions. Fungal Ecol 2: 10–20.
Nicholas KB, Nicholas HBJ & Deerfield DW (1997) GeneDoc:
analysis and visualization of genetic variation. Embnew News
4: 14.
Opik M, Moora M, Liira J & Zobel M (2006) Composition of
root-colonizing arbuscular mycorrhizal fungal communities in
different ecosystems around the globe. J Ecol 94: 778–790.
Opik M, Moora M, Zobel M, Saks U, Wheatley R, Wright F &
Daniell T (2008) High diversity of arbuscular mycorrhizal
fungi in a boreal herb-rich coniferous forest. New Phytol 179:
867–876.
Opik M, Metsis M, Daniell TJ, Zobel M & Moora M (2009)
Large-scale parallel 454 sequencing reveals host ecological
group specificity of arbuscular mycorrhizal fungi in a
boreonemoral forest. New Phytol 184: 424–437.
Opik M, Vanatoa A, Vanatoa E, Moora M, Davison J, Kalwij JM,
Reiner U & Zobel M (2010) The online database MaarjAM
reveals global and ecosystemic distribution patterns in
arbuscular mycorrhizal fungi (Glomeromycota). New Phytol
188: 223–241.
Pietikainen A, Kytoviita MM, Husband R & Young JPW (2007)
Diversity and persistence of arbuscular mycorrhizas in a low-
Arctic meadow habitat. New Phytol 176: 691–698.
R Development Core Team (2010) R: A Language and
Environment for Statistical Computing. R Foundation for
Statistical Computing, Vienna, Austria.
Ronquist F & Huelsenbeck JP (2003) MRBAYES 3: Bayesian
phylogenetic inference under mixed models. Bioinformatics 19:
1572–1574.
Schechter SP & Bruns TD (2008) Serpentine and non-serpentine
ecotypes of Collinsia sparsiflora associate with distinct
arbuscular mycorrhizal fungal assemblages. Mol Ecol 17:
3198–3210.
Schußler A &Walker C (2010) The Glomeromycota: A species list
with new families and new genera. Available at http://www.
amf-phylogeny.com.
Schwarzott D & Schußler A (2001) A simple and reliable method
for SSU rRNA gene DNA extraction, amplification, and
cloning from single AM fungal spores. Mycorrhiza 10:
203–207.
Simon L, Lalonde M & Bruns TD (1992) Specific amplification of
18S fungal ribosomal genes from vesicular-arbuscular
endomycorrhizal fungi colonizing roots. Appl Environ Microb
58: 291–295.
Smith SE & Read DJ (2008) Mycorrhizal Symbiosis. Academic
Press, Amsterdam, The Netherlands.
Smith SE, Smith FA & Jakobsen I (2003) Mycorrhizal fungi can
dominate phosphate supply to plants irrespective of growth
responses. Plant Physiol 133: 16–20.
Sutherland W (1996) Ecological Census Techniques: A Handbook.
Cambridge University Press, Cambridge, UK.
Sykorova Z, Wiemken A & Redecker D (2007) Co-occurring
Gentiana verna and Gentiana acaulis and their neighboring
plants in two Swiss upper montane meadows harbor distinct
arbuscular mycorrhizal fungal communities. Appl Environ
Microb 73: 5426–5434.
Tamura K, Dudley J, Nei M & Kumar S (2007) MEGA4:
molecular evolutionary genetics analysis (MEGA) software
version 4.0. Mol Biol Evol 24: 1596–1599.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F & Higgins
DG (1997) The CLUSTAL_X Windows interface: flexible
strategies for multiple sequence alignment aided by quality
analysis tools. Nucleic Acids Res 25: 4876–4882.
Tian H, Gai JP, Zhang JL, Christie P & Li XL (2009) Arbuscular
mycorrhizal fungi in degraded typical steppe of Inner
Mongolia. Land Degrad Dev 20: 41–54.
Vandenkoornhuyse P, Ridgway KP, Watson IJ, Fitter AH & Young
JPW (2003) Co-existing grass species have distinctive
arbuscular mycorrhizal communities. Mol Ecol 12: 3085–3095.
van der Heijden MGA, Klironomos JN, Ursic M, Moutoglis P,
Streitwolf-Engel R, Boller T, Wiemken A & Sanders IR (1998)
Mycorrhizal fungal diversity determines plant biodiversity,
ecosystem variability and productivity. Nature 396: 69–72.
Vogelsang KM, Reynolds HL & Bever JD (2006) Mycorrhizal
fungal identity and richness determine the diversity and
productivity of a tallgrass prairie system. New Phytol 172:
554–562.
Walker C, Vestberg M, Demircik F, Demircik F, Stockinger H,
Saito M, Sawaki H, Nishmura I & Schußler A (2007)
Molecular phylogeny and new taxa in the Archaeosporales
(Glomeromycota): Ambispora fennica gen. sp nov.,
Ambisporaceae fam. nov., and emendation of Archaeospora and
Archaeosporaceae. Mycol Res 111: 137–153.
Wang B & Qiu YL (2006) Phylogenetic distribution and evolution
of mycorrhizas in land plants. Mycorrhiza 16: 299–363.
Wu B, Hogetsu T, Isobe K & Ishii R (2007) Community structure
of arbuscular mycorrhizal fungi in a primary successional
volcanic desert on the southeast slope of Mount Fuji.
Mycorrhiza 17: 495–506.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Fig. S1. Geographical location of study sites.
Table S1. Number of spores of AMF phylotypes detected
from rhizospheres of two plant species at five study sites.
Table S2. Number of clones of AMF phylotypes detected in
the roots of two plant species at five study sites.
Table S3. The characteristics of rhizosphere soils of each
plant species in each site.
Please note: Wiley-Blackwell is not responsible for the
content or functionality of any supporting materials sup-
plied by the authors. Any queries (other than missing
material) should be directed to the corresponding author
for the article.
FEMS Microbiol Ecol 78 (2011) 355–365 c� 2011 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
365Diverse AM fungi in Tibet Plateau

Supporting information
Table S1 Number of spores of AMF phylotypes detected from rhizospheres of two plant
species at five study sites. First, number of spores of each morphotype was counted; then at
least one spore per morphotype was subjected to molecular identification. Site codes are as in
Table 1.
Dracocephalum heterophyllum Astragalus polycladus
X K W T A Total X K W T A Total
Glo-A8 - 240 - - - 240 - - - - - 0
Glo-A9 - - - - - 0 - - 26 - - 26
Glo-B1 - 97 - - - 97 - - - - - 0
Div-2 - - - - - 0 - - - 179 - 179
Div-3 16 444 - - 27 487 - - - - - 0
Amb-2 - - - - 12 12 - - - - - 0
Pac-1 - - - - 108 108 - 92 536 - 8 636
Total richness 1 3 0 0 3 5 0 1 2 1 1 3
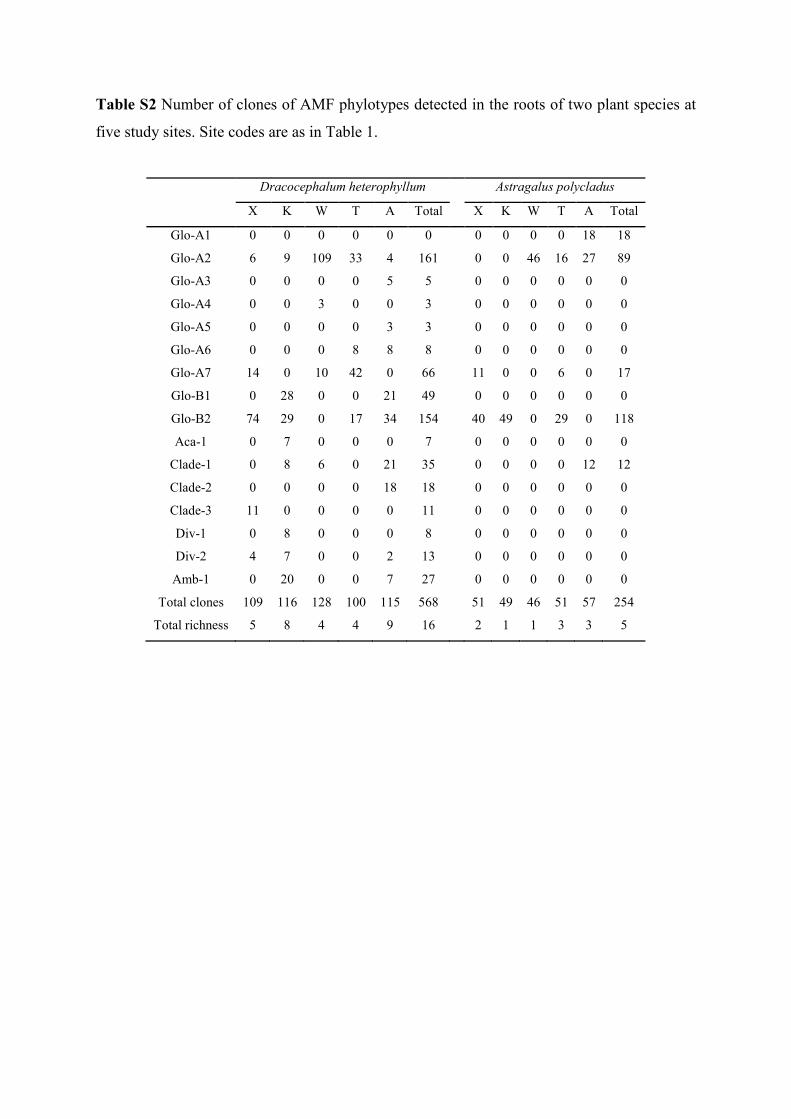
Table S2 Number of clones of AMF phylotypes detected in the roots of two plant species at
five study sites. Site codes are as in Table 1.
Dracocephalum heterophyllum Astragalus polycladus
X K W T A Total X K W T A Total
Glo-A1 0 0 0 0 0 0 0 0 0 0 18 18
Glo-A2 6 9 109 33 4 161 0 0 46 16 27 89
Glo-A3 0 0 0 0 5 5 0 0 0 0 0 0
Glo-A4 0 0 3 0 0 3 0 0 0 0 0 0
Glo-A5 0 0 0 0 3 3 0 0 0 0 0 0
Glo-A6 0 0 0 8 8 8 0 0 0 0 0 0
Glo-A7 14 0 10 42 0 66 11 0 0 6 0 17
Glo-B1 0 28 0 0 21 49 0 0 0 0 0 0
Glo-B2 74 29 0 17 34 154 40 49 0 29 0 118
Aca-1 0 7 0 0 0 7 0 0 0 0 0 0
Clade-1 0 8 6 0 21 35 0 0 0 0 12 12
Clade-2 0 0 0 0 18 18 0 0 0 0 0 0
Clade-3 11 0 0 0 0 11 0 0 0 0 0 0
Div-1 0 8 0 0 0 8 0 0 0 0 0 0
Div-2 4 7 0 0 2 13 0 0 0 0 0 0
Amb-1 0 20 0 0 7 27 0 0 0 0 0 0
Total clones 109 116 128 100 115 568 51 49 46 51 57 254
Total richness 5 8 4 4 9 16 2 1 1 3 3 5

Table S3 The characteristics of rhizosphere soils of each plant species in each site. Data are
shown with mean ± SE (n=3). Significant differences of each soil variable between samples
were determined using Fisher’s Least Significant Difference at the 5% level after one-way
ANOVA and indicated by different letters. Site codes are as in Table 1. DH: Dracocephalum
heterophyllum; AP: Astragalus polycladus.
Sampling
sites
Plant
species
Total N
(g kg-1)
Organic C
(g kg-1) C/N ratio
Available P
(mg kg-1) pH
X DH 0.72±0.06 c 0.89±0.03 g 1.23±0.06 i 1.32±0.13 d 7.38±0.02 b
AP 0.5±0.02 f 0.79±0.07 h 1.58±0.11 g 1.37±0.05 cd 7.59±0.02 a
K DH 0.53±0.04 e 0.98±0.04 f 1.83±0.09 f 0.59±0.28 g 7.64±0.05 a
AP 0.42±0.03 g 0.96±0.03 f 2.27±0.17 e 0.83±0.2 f 7.56±0.01 a
W DH 0.36±0.02 h 1.75±0.07 c 4.82±0.23 a 2.20±0.11 a 7.57±0.06 a
AP 0.43±0.02 g 1.19±0.15 e 2.76±0.16 c 1.55±0.19 bc 7.38±0.01 b
T DH 0.57±0.09 d 1.85±0.07 b 3.26±0.13 b 0.97±0.17 ef 7.05±0.09 e
AP 0.78±0.05 b 2.02±0.05 a 2.59±0.19 d 1.08±0.08 e 7.31±0.03 bc
A DH 0.87±0.03 a 1.32±0.02 d 1.52±0.04 h 1.29±0.13 cd 7.23±0.02 cd
AP 0.87±0.05 a 1.37±0.05 d 1.57±0.09 g 1.68±0.06 b 7.16±0.06 d

Fig. S1 Geographical location of study sites.




















