
Effect of physical, chemical and environmental characteristics on arbuscular mycorrhizal fungi in...
10
ORIGINAL ARTICLE Effect of physical, chemical and environmental characteristics on arbuscular mycorrhizal fungi in Brachiaria decumbens (Stapf) pastures R.H. Posada 1,2 , L.A. Franco 2 , C. Ramos 3 , L.S. Plazas 2 , J.C. Sua ´ rez 2 and F. A ´ lvarez 2 1 Jardı´n Bota ´ nico Jose ´ Celestino Mutis, Bogota ´ , Colombia 2 Departamento de Biologı´a, Facultad de Ciencias, Universidad de la Amazonı´a, Florencia, Caqueta ´ , Colombia 3 Lab. Ecologı´a terrestre, Depto. de Ciencias Ecolo ´ gicas, Universidad de Chile, Santiago, Chile Introduction Arbuscular mycorrhizal fungi (AMF) are important sym- bionts for most terrestrial plants. AMF take up nutrients, especially phosphorus from the soil, and these nutrients are then exchanged for carbon from the host plant. In forests, more than 80% of pioneer and early successional plant species have AMF symbiosis (Fontenla et al. 1998; Siqueira et al. 1998; Andrade et al. 2000; Siqueira and Saggin-Ju ´ nior 2001; Gehring and Connell 2006). AMF have therefore an important role in the recuperation and restoration of both deforested zones and gaps, influencing the establishment ability and persistence of the pioneer plants, and affecting the plant succession. Soil characteristics, plant species and climate may regu- late the AMF community (Escudero and Mendoza 2005). The occurrence of mycorrhiza is influenced by the land slope and the geographic and topographic locality (Dick- inson 1974). High temperatures result in a greater extent of infection by AMF (Diederich and Moawad 1993); under specific conditions, the spore density correlates with fluctuations in temperature (Koske 1987). In grasses, low moisture levels lead to increases in root colonizations and decrements of the spore production by AMF (Simp- son and Daft 1990; Rickerl et al. 1994; Camargo-Ricalde and Espero ´n-Rodrı´guez 2005). However, both very dry and flooded soils decrease colonization by AMF (Lodge 1989; Miller and Bever 1999; Miller 2000). In general, Keywords Amazonian foothill, arbuscular mycorrhizal fungi, Brachiaria decumbens, pastures. Correspondence Rau ´ l Hernando Posada, Calle 63 A , No 32-09 Piso 2, Bogota ´ , Colombia. E-mail: [email protected] 2007 ⁄ 1275: received 11 September 2006, revised 24 April 2007 and accepted 4 July 2007 doi:10.1111/j.1365-2672.2007.03533.x Abstract Aim: To evaluate the effects of soil physical and chemical factors (pH, conduc- tivity, humidity, available phosphorus and organic matter) and environmental factors (temperature, relative air humidity, altitude and atmospheric pressure) on arbuscular mycorrhizal fungi (AMF)–Brachiaria decumbens grass relation- ship. Furthermore to establish patterns of microbiological responses that allow to differentiate the study sites in two relief types. Methods and Results: Mycorrhizal characteristics (spore density, external hyphae and root colonizations by hyphae, vesicles and arbuscules), physical and chemical factors in soil and environmental factors were measured. Conclusions: The effect of physical, chemical and environmental factors on microbiological variables was related to the type of relief ‘valley and hilly ter- rain’; the AMF behaviour was affected only over narrower ranges of evaluated variables. Similarly, the colonization of B. decumbens roots by AMF hyphae, vesicles and the mycorrhizal spore density follow different patterns according to the relief type. Significance and Impact of the Study: The type of relief is one of the factors to be taken into consideration to evaluate the AMF inoculum and root coloniza- tion of these pastures, because of the influence of slope – as physical property of soil – on AMF. Journal of Applied Microbiology ISSN 1364-5072 132 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140 ª 2007 The Authors
Transcript of Effect of physical, chemical and environmental characteristics on arbuscular mycorrhizal fungi in...

ORIGINAL ARTICLE
Effect of physical, chemical and environmentalcharacteristics on arbuscular mycorrhizal fungi in Brachiariadecumbens (Stapf) pasturesR.H. Posada1,2, L.A. Franco2, C. Ramos3, L.S. Plazas2, J.C. Suarez2 and F. Alvarez2
1 Jardın Botanico Jose Celestino Mutis, Bogota, Colombia
2 Departamento de Biologıa, Facultad de Ciencias, Universidad de la Amazonıa, Florencia, Caqueta, Colombia
3 Lab. Ecologıa terrestre, Depto. de Ciencias Ecologicas, Universidad de Chile, Santiago, Chile
Introduction
Arbuscular mycorrhizal fungi (AMF) are important sym-
bionts for most terrestrial plants. AMF take up nutrients,
especially phosphorus from the soil, and these nutrients
are then exchanged for carbon from the host plant. In
forests, more than 80% of pioneer and early successional
plant species have AMF symbiosis (Fontenla et al. 1998;
Siqueira et al. 1998; Andrade et al. 2000; Siqueira and
Saggin-Junior 2001; Gehring and Connell 2006). AMF
have therefore an important role in the recuperation and
restoration of both deforested zones and gaps, influencing
the establishment ability and persistence of the pioneer
plants, and affecting the plant succession.
Soil characteristics, plant species and climate may regu-
late the AMF community (Escudero and Mendoza 2005).
The occurrence of mycorrhiza is influenced by the land
slope and the geographic and topographic locality (Dick-
inson 1974). High temperatures result in a greater extent
of infection by AMF (Diederich and Moawad 1993);
under specific conditions, the spore density correlates
with fluctuations in temperature (Koske 1987). In grasses,
low moisture levels lead to increases in root colonizations
and decrements of the spore production by AMF (Simp-
son and Daft 1990; Rickerl et al. 1994; Camargo-Ricalde
and Esperon-Rodrıguez 2005). However, both very dry
and flooded soils decrease colonization by AMF (Lodge
1989; Miller and Bever 1999; Miller 2000). In general,
Keywords
Amazonian foothill, arbuscular mycorrhizal
fungi, Brachiaria decumbens, pastures.
Correspondence
Raul Hernando Posada, Calle 63A, No 32-09
Piso 2, Bogota, Colombia.
E-mail: [email protected]
2007 ⁄ 1275: received 11 September 2006,
revised 24 April 2007 and accepted 4 July
2007
doi:10.1111/j.1365-2672.2007.03533.x
Abstract
Aim: To evaluate the effects of soil physical and chemical factors (pH, conduc-
tivity, humidity, available phosphorus and organic matter) and environmental
factors (temperature, relative air humidity, altitude and atmospheric pressure)
on arbuscular mycorrhizal fungi (AMF)–Brachiaria decumbens grass relation-
ship. Furthermore to establish patterns of microbiological responses that allow
to differentiate the study sites in two relief types.
Methods and Results: Mycorrhizal characteristics (spore density, external
hyphae and root colonizations by hyphae, vesicles and arbuscules), physical
and chemical factors in soil and environmental factors were measured.
Conclusions: The effect of physical, chemical and environmental factors on
microbiological variables was related to the type of relief ‘valley and hilly ter-
rain’; the AMF behaviour was affected only over narrower ranges of evaluated
variables. Similarly, the colonization of B. decumbens roots by AMF hyphae,
vesicles and the mycorrhizal spore density follow different patterns according
to the relief type.
Significance and Impact of the Study: The type of relief is one of the factors to
be taken into consideration to evaluate the AMF inoculum and root coloniza-
tion of these pastures, because of the influence of slope – as physical property
of soil – on AMF.
Journal of Applied Microbiology ISSN 1364-5072
132 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140
ª 2007 The Authors

vesicle colonization (Stevens and Peterson 1996) and the
external hyphae (Schack-Kirchner et al. 2000) are not typ-
ically affected by the water gradient.
The pH affects the distribution and abundance of dif-
ferent fungal species (Read et al. 1976; Porter et al.
1987a,b; Wang et al. 1993). Small increases in pH are
associated with greater root colonization by AMF in acid
soils with low phosphorus availability (Soedarjo and
Habte 1995; Heijne et al. 1996).
Besides the difficulty in separating the influences of
host plant species and soil characteristics on root coloni-
zation and the inoculum (external hyphae and spore
density) density (Escudero and Mendoza 2005), there is
not a clear separation between plant and soil factors.
There is growing evidence that diversity and distribution
of AMF depend on the community structure and char-
acteristics of the ecosystem (Van der Heijden and Sand-
ers 2002).
Brachiaria decumbens (Stapf) is a grass species intro-
duced to the Amazonian foothills in Colombia in 1970 as
a forage pasture to improve the cattle productivity
(Cuesta 1978; Siqueira et al. 1990; Velasquez and Cuesta
1990). Since then, extensive monocultures of this grass
have replaced the forest coverage. Although B. decumbens
has root colonizations of 75% in Brazilian soils with high
phosphorus content (Oliveira et al. 1997), continuity of
these pastures can affect the AMF populations, resulting
in the reduction of soil biological and productive capaci-
ties (Robertson et al. 1997). Commonly, the mycorrhizal
fungi follow heterogeneous distribution patterns, accord-
ing to the soil and environmental characteristics where
the plant species are (Camargo-Ricalde and Esperon-Rod-
rıguez 2005); the study of this association with just one
dependent plant species helps to take the decision about
the requirement or not of previous soil enrichment with
mycorrhizal supply when sowing the seedlings and sap-
lings – of pioneer trees to restoration programmes. The
objectives to study on B. decumbens pastures were to:
(i) determine the most influential soil and environmental
variables, and how these variables affect AMF behaviour,
and (ii) establish patterns of microbiological responses
that allow to differentiate the study sites in two relief
types.
Materials and methods
This study was conducted at 26 sites located in the Amazo-
nian foothills of Caqueta, Colombia, in the area between
1�28¢50Æ3¢¢–1�26¢41¢¢N and 75�40¢14¢¢–75�28¢26¢¢W. This is
situated in a zone of confluence between the Andean
Cordillera and the Amazonian forest, with mean annual
temperature of 28�C, mean annual rainfall of 3500 mm,
and mean air relative humidity of 87%. The sites were
pastures resulting from clearing of tropical rain forest,
and comprised two types of relief – hilly terrain and fertile
valley with a floodplain.
Twenty-six sites of 1000 m2 in 18 farms were selected
to take samples in the hilly areas -slope 20–35� from the
horizontal (APY, ARY, FBY, JMY, UAY, AAY, ABYA,
FMY, GSY, DCY, ESYA, ESYB, FGY, NAY, ABYB), and
in the valley plains (slope 0–11� from the horizontal
(CNX, ESX, JAX1, JAX2, ACX, GSX, VPX, FGX, NAXA,
NAXB, VAX), with B. decumbens (Stapf) as pasture, and
the samples were collected between January and April of
2003 (dry season). A cylinder of 38 mm diameter and
250 mm length was used to take soil samples of 0–0Æ2 m
deep – at 0–0Æ1 m from the plant, completing 10 ran-
domly selected samples per site (260 in total). All samples
were thoroughly homogenized. Two subsamples (each
200 g) were used to evaluate spore density and external
mycelia; one root subsample (1 g) was used to evaluate
colonization, and one soil subsample (100 g) for physical
and chemical determinations. Environmental conditions
were measured with a field KONUS digital thermo-
hygrometer and an altimeter-barometer. The labelled
samples were separated into roots and soil in the labora-
tory, and were stored at 2�C for later assessment.
The AMF spore density was determined according to
Sieverding (1983), by wet sieving with 45, 120 and
500 lm sieves and decanting, followed by centrifugal flo-
tation (500 g l–1 sucrose). The results were expressed as
spore number 10 g–1 dry soil. Coenocytic extra-radical
hyphae or external hyphae were extracted according to
Herrera et al. (1986). Air-dried samples were added to
H2O2 (0Æ2 l l–1 H2O2), blended for 30 s, rinsed on the
45 lm sieve, air-dried for 48 h and weighed. A further
sample of 0Æ02 g was mixed with two drops of glycerin
(100%) on a microscope slide. The number of coenocytic
hyphae of AMF that intersected four squared transects
(two horizontal and two vertical, separated by 5 mm) on
each slide were counted at 100· magnification using a
compound microscope. The results were expressed in
meters of external hyphae (m g–1 soil).
Roots were cleared and stained by the Philips and Hay-
man (1970) modified method. Cleared roots were acidi-
fied with HCl (10 g l–1 HCl) for 300–900 s and stained in
acid Trypan blue (0Æ5 g l–1 Trypan blue); the roots were
mounted on microscope slides for assessment by the mag-
nified intercept method as described by Pabon (2000),
Posada (2001) and Aristizabal et al. (2004). Colonization
of roots (% aseptate hyphae colonization, % vesicle colo-
nization, % arbuscule colonization and % septate hyphae
colonization) was estimated as the number of colonized
intersections divided by the total number of observed
intersections; septate hyphae colonization was measured
as an indicative of fungal competition against AMF and
R.H. Posada et al. Arbuscular mycorrhiza in pastures
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140 133

non-AMF in roots and as ecological behaviour of plant
species.
Soil humidity (%) determination was made by drying
samples at 80�C for 72 h, the electric conductivity (EC)
(lS cm–1) by direct measurement in conductivity-meter
and the pH in a 1 : 1 soil:water slurry; the organic matter
determination was realized by Wilkley–Black method (%
carbon 100 g–1 soil) and the available phosphorus was mea-
sured employing the Bray II method (lg g–1) (IGAC 1999).
Physical and chemical variables in soil (conductivity,
soil relative moisture, organic matter, available phospho-
rus, pH), environmental variables (altitude, air relative
humidity, atmospheric pressure, temperature), and micro-
biological variables (spore density, external hyphae, colo-
nization by hyphae, colonization by vesicles, colonization
by arbuscules and colonization by septate fungi) were
summarized in a matrix by sites. Colonization variables
and conductivity were –log10(x + 1) normalized and the
remainder variables were log10 normalized before the
analysis. We used one-way anova to evaluate differences
in the variables, between types of relief; and Spearman’s
rank correlation coefficients were computed for all pair-
wise combination of variables measured by relief. The
microbiological, physical, chemical and environmental
variables with higher influence on the variability between
sites were determined by principal component analysis.
Similarity between sites by microbiological characteristics
was detected clustering the microbiological variables in a
correlation matrix by sites and the results were expressed
in dendrograms. Canonical correlations were used to
determinate the effect of physical, chemical and environ-
mental variables on the AMF behaviour. Statistical analy-
sis was performed using the sas program (SAS Institute
Inc., Cary, NC, USA).
Results
The measured characteristics (physical and chemical of
soil, environmental and microbiological), were highly var-
iable (Table 1). Three characteristics differed significantly
from one type of relief to another; the phosphorus avail-
ability and pH were larger in the valley than in the hilly
terrain; but the spore density was larger in the hilly ter-
rain than in the valley.
In the hilly terrain, there were highly positive correla-
tions between conductivity and colonization by arbuscules
(r = 0Æ672, P = 0Æ0176), between soil humidity and exter-
nal hyphae (r = 0Æ646, P = 0Æ0192) and between the coloni-
zation by hyphae and colonization by vesicles (r = 0Æ950,
P < 0Æ0001); there were negative correlations between
spore density and altitude (r = –0Æ660, P = 0Æ0183). In
contrast, in the valley there were highly positive correla-
tions between colonization by hyphae and colonization
by vesicles (r = 0Æ952, P < 0Æ0001), between colonization
by hyphae and colonization by arbuscules (r = 0Æ768,
P = 0Æ0057), between colonization by vesicles and coloniza-
tion by arbuscules (r = 0Æ823, P = 0Æ0021); and negative
correlations between spore density and colonization by
hyphae (r = –0Æ618, P = 0Æ0416), between organic mat-
ter and soil humidity (r = –0Æ840, P = 0Æ0012), and
between available phosphorus and altitude (r = –0Æ659,
P = 0Æ0184).
The microbiological characteristics more sensitive to
the variability between sites were the root colonizations
by hyphae and vesicles (explained variability: 39%); and
the inoculum (explained variability: 26%) (Fig. 1). The
samples of ABYB showed responses different from other
sites (dotted line right and bottom); while the characteris-
tic least sensitive to the variability was the colonization by
arbuscules.
Among physical and chemical variables in the soil, the
first principal component (30Æ8%) joined pH, soil humid-
ity and organic matter; the second principal component
(27Æ4%) was the phosphorus availability, and the third
one represented the EC (20Æ8%). The valley showed a dif-
ferentiation of the sites in function from the axes 1 and 2
bigger to the differentiation of sites in the hilly terrain.
The samples in the valley had the lower conductivities,
and the samples in the hilly terrain had the lower phos-
phorus availabilities. Samples from ESYA were different
Table 1 Mean (±SD) of physical, chemical, environmental and micro-
biological measures in the sampling sites, according to the type of
relief (fertile valley with a floodplain and hilly terrain), obtained by
one-way ANOVA
Characteristics
Type of relief
Fertile valley Hilly terrain
Soil Physical and Chemical
Available phosphorus (lg g–1)* 22Æ5 ± 12Æ8 11Æ7 ± 7Æ44
pH* 5Æ05 ± 0Æ46 4Æ59 ± 0Æ27
Organic matter (%) 3Æ51 ± 0Æ98 3Æ26 ± 0Æ56
Conductivity (lS cm–1) 0Æ160 ± 0Æ009 0Æ172 ± 0Æ027
Soil humidity (%) 32Æ8 ± 5Æ33 31Æ6 ± 4Æ22
Environmental
Air relative humidity (%) 61Æ0 ± 6Æ09 62Æ8 ± 6Æ61
Altitude (m a.s.l.) 267 ± 37Æ5 334 ± 90Æ7
Pressure (MPa) 98Æ3 ± 0Æ87 98Æ7 ± 1Æ61
Temperature (�C) 30Æ3 ± 2Æ43 30Æ6 ± 2Æ21
Microbiological
Colonization by hyphae (%) 30Æ5 ± 29Æ8 30Æ7 ± 24Æ3
Colonization by vesicles (%) 14Æ5 ± 19Æ7 14Æ7 ± 15Æ7
Colonization by arbuscules (%) 1Æ16 ± 2Æ12 0Æ64 ± 1Æ10
Colonization by septate fungi (%) 37Æ3 ± 20Æ9 39Æ5 ± 21Æ7
External hyphae (m g–1) 44Æ8 ± 27Æ4 47Æ5 ± 47Æ4
Spore density no.* 183 ± 78Æ4 242 ± 113Æ5
*Significant at P £ 0Æ05.
Arbuscular mycorrhiza in pastures R.H. Posada et al.
134 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140
ª 2007 The Authors
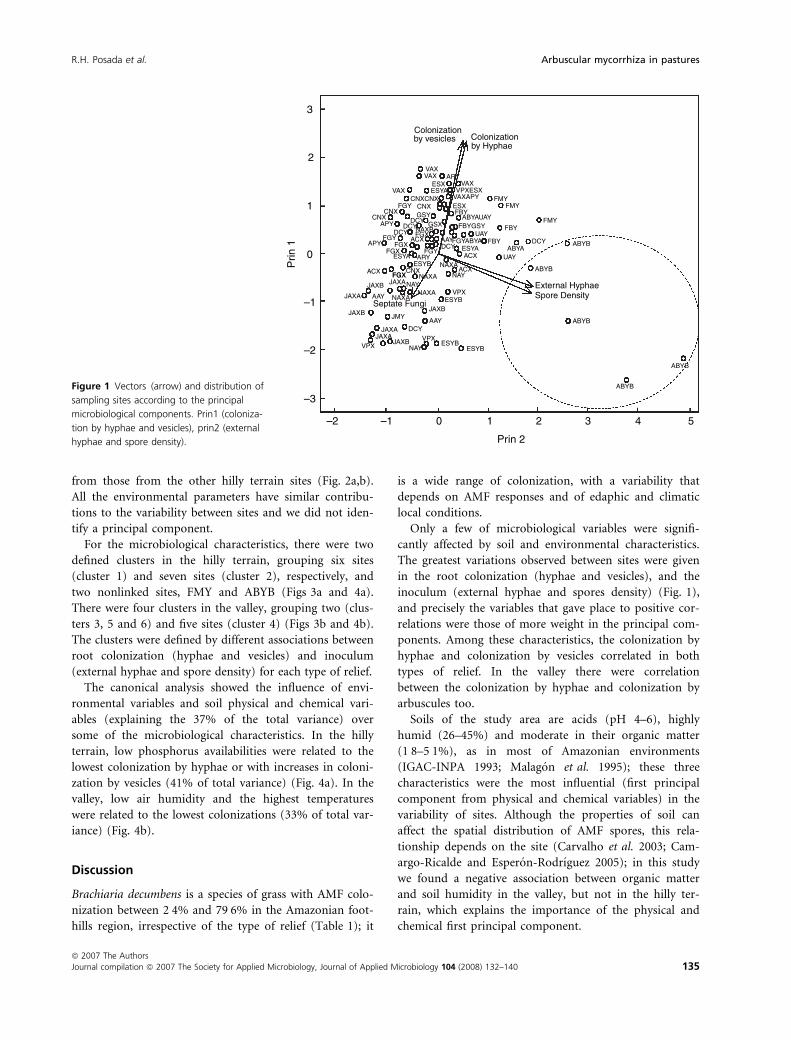
from those from the other hilly terrain sites (Fig. 2a,b).
All the environmental parameters have similar contribu-
tions to the variability between sites and we did not iden-
tify a principal component.
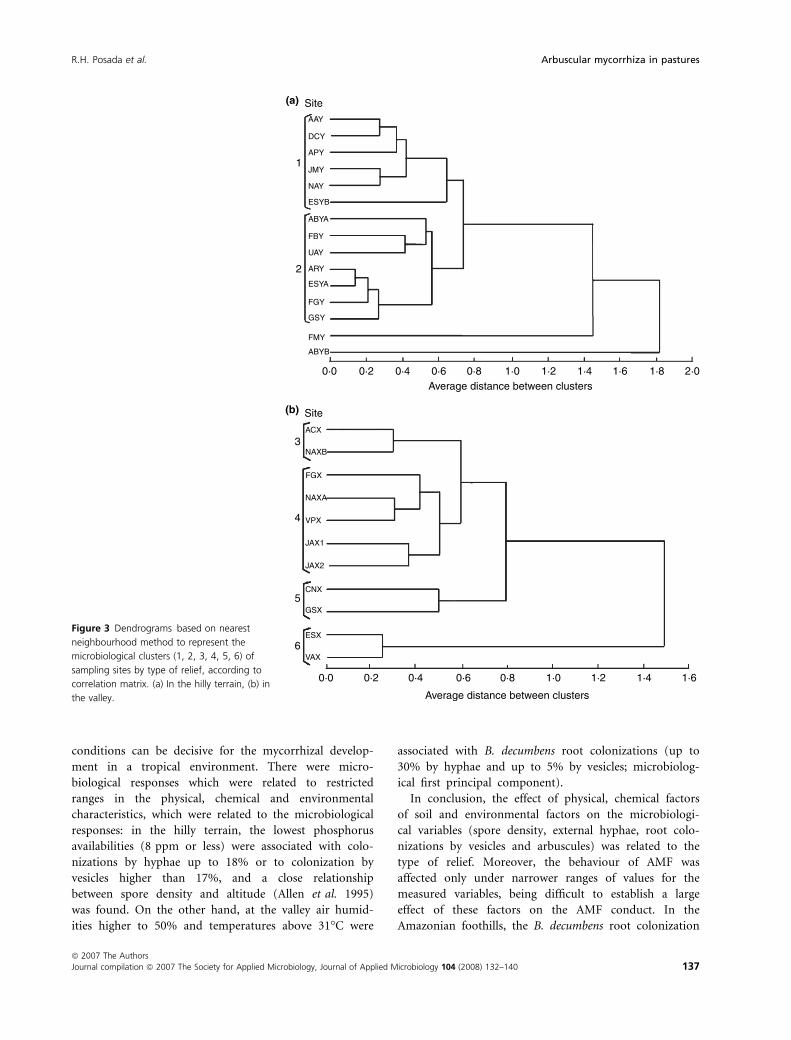
For the microbiological characteristics, there were two
defined clusters in the hilly terrain, grouping six sites
(cluster 1) and seven sites (cluster 2), respectively, and
two nonlinked sites, FMY and ABYB (Figs 3a and 4a).
There were four clusters in the valley, grouping two (clus-
ters 3, 5 and 6) and five sites (cluster 4) (Figs 3b and 4b).
The clusters were defined by different associations between
root colonization (hyphae and vesicles) and inoculum
(external hyphae and spore density) for each type of relief.
The canonical analysis showed the influence of envi-
ronmental variables and soil physical and chemical vari-
ables (explaining the 37% of the total variance) over
some of the microbiological characteristics. In the hilly
terrain, low phosphorus availabilities were related to the
lowest colonization by hyphae or with increases in coloni-
zation by vesicles (41% of total variance) (Fig. 4a). In the
valley, low air humidity and the highest temperatures
were related to the lowest colonizations (33% of total var-
iance) (Fig. 4b).
Discussion
Brachiaria decumbens is a species of grass with AMF colo-
nization between 2Æ4% and 79Æ6% in the Amazonian foot-
hills region, irrespective of the type of relief (Table 1); it
is a wide range of colonization, with a variability that
depends on AMF responses and of edaphic and climatic
local conditions.
Only a few of microbiological variables were signifi-
cantly affected by soil and environmental characteristics.
The greatest variations observed between sites were given
in the root colonization (hyphae and vesicles), and the
inoculum (external hyphae and spores density) (Fig. 1),
and precisely the variables that gave place to positive cor-
relations were those of more weight in the principal com-
ponents. Among these characteristics, the colonization by
hyphae and colonization by vesicles correlated in both
types of relief. In the valley there were correlation
between the colonization by hyphae and colonization by
arbuscules too.
Soils of the study area are acids (pH 4–6), highly
humid (26–45%) and moderate in their organic matter
(1Æ8–5Æ1%), as in most of Amazonian environments
(IGAC-INPA 1993; Malagon et al. 1995); these three
characteristics were the most influential (first principal
component from physical and chemical variables) in the
variability of sites. Although the properties of soil can
affect the spatial distribution of AMF spores, this rela-
tionship depends on the site (Carvalho et al. 2003; Cam-
argo-Ricalde and Esperon-Rodrıguez 2005); in this study
we found a negative association between organic matter
and soil humidity in the valley, but not in the hilly ter-
rain, which explains the importance of the physical and
chemical first principal component.
3
2
1
0
–1
–2
–3
Prin
1
–2 –1 0 1 2 3 4 5
Prin 2
ABYB
ABYB
ABYB
ABYB
ABYB
External HyphaeSpore Density
ESYBESYB
ESYB
ESYB
ESYAESYA
JAXA
JAXA
JAXAJAXA
JAXB
JAXB
JAXB
JAXB
VPX
JMY
VPX
VPX
NAY
NAYNAY
NAXANAXA
NAXA
NAXA
AAY
AAY
DCY
DCY
DCYDCY
AAY
ACX
ACXACX
ACX
Septate Fungi
FGXCNX
VAX
VAXVAX
VAXVPXESX
ESX
ESXCNX
CNXAPY
APYFGX
FGX
FGX
FGX
FGY
FGY
FMYFMY
FBY
FBYFMY
ColonizationColonizationby vesiclesby Hyphae
ARY
DCY ABYAUAY
FBYGSYABYAUAY
UAYFBYFGYABYA DCY
ARY
CNXCNXCNX
VAXAPYFGY
GSYGSX
NAXB
ESYA
Figure 1 Vectors (arrow) and distribution of
sampling sites according to the principal
microbiological components. Prin1 (coloniza-
tion by hyphae and vesicles), prin2 (external
hyphae and spore density).
R.H. Posada et al. Arbuscular mycorrhiza in pastures
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140 135
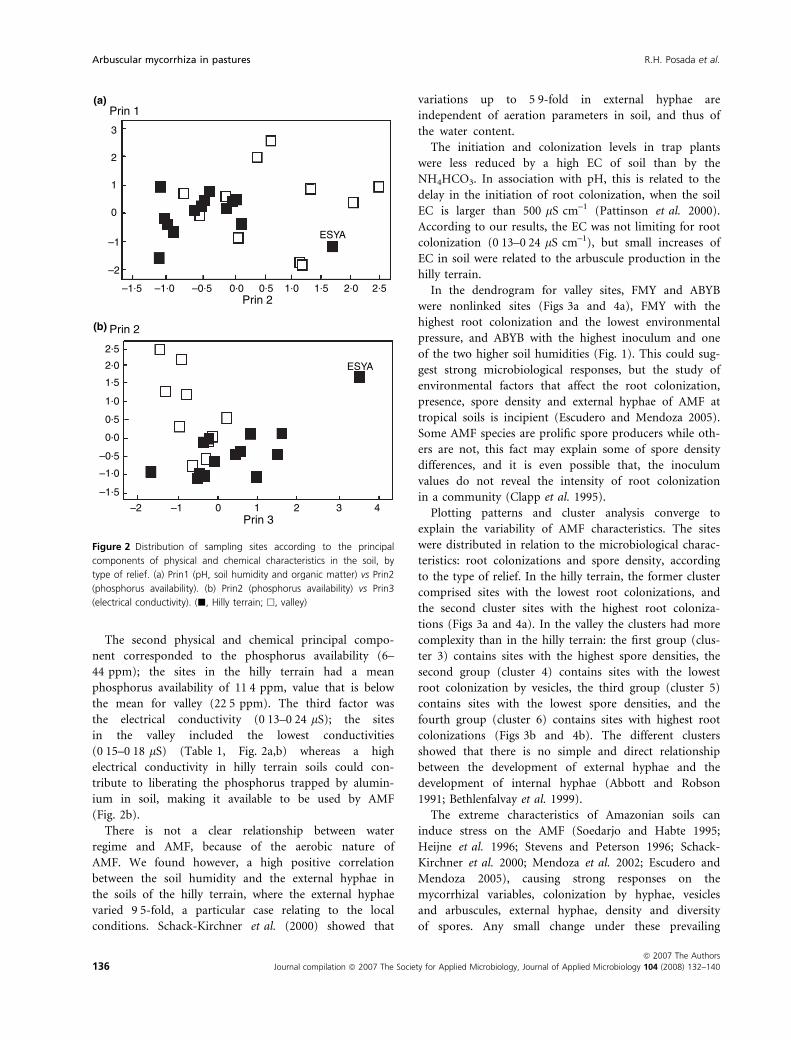
The second physical and chemical principal compo-
nent corresponded to the phosphorus availability (6–
44 ppm); the sites in the hilly terrain had a mean
phosphorus availability of 11Æ4 ppm, value that is below
the mean for valley (22Æ5 ppm). The third factor was
the electrical conductivity (0Æ13–0Æ24 lS); the sites
in the valley included the lowest conductivities
(0Æ15–0Æ18 lS) (Table 1, Fig. 2a,b) whereas a high
electrical conductivity in hilly terrain soils could con-
tribute to liberating the phosphorus trapped by alumin-
ium in soil, making it available to be used by AMF
(Fig. 2b).
There is not a clear relationship between water
regime and AMF, because of the aerobic nature of
AMF. We found however, a high positive correlation
between the soil humidity and the external hyphae in
the soils of the hilly terrain, where the external hyphae
varied 9Æ5-fold, a particular case relating to the local
conditions. Schack-Kirchner et al. (2000) showed that
variations up to 5Æ9-fold in external hyphae are
independent of aeration parameters in soil, and thus of
the water content.
The initiation and colonization levels in trap plants
were less reduced by a high EC of soil than by the
NH4HCO3. In association with pH, this is related to the
delay in the initiation of root colonization, when the soil
EC is larger than 500 lS cm–1 (Pattinson et al. 2000).
According to our results, the EC was not limiting for root
colonization (0Æ13–0Æ24 lS cm–1), but small increases of
EC in soil were related to the arbuscule production in the
hilly terrain.
In the dendrogram for valley sites, FMY and ABYB
were nonlinked sites (Figs 3a and 4a), FMY with the
highest root colonization and the lowest environmental
pressure, and ABYB with the highest inoculum and one
of the two higher soil humidities (Fig. 1). This could sug-
gest strong microbiological responses, but the study of
environmental factors that affect the root colonization,
presence, spore density and external hyphae of AMF at
tropical soils is incipient (Escudero and Mendoza 2005).
Some AMF species are prolific spore producers while oth-
ers are not, this fact may explain some of spore density
differences, and it is even possible that, the inoculum
values do not reveal the intensity of root colonization
in a community (Clapp et al. 1995).
Plotting patterns and cluster analysis converge to
explain the variability of AMF characteristics. The sites
were distributed in relation to the microbiological charac-
teristics: root colonizations and spore density, according
to the type of relief. In the hilly terrain, the former cluster
comprised sites with the lowest root colonizations, and
the second cluster sites with the highest root coloniza-
tions (Figs 3a and 4a). In the valley the clusters had more
complexity than in the hilly terrain: the first group (clus-
ter 3) contains sites with the highest spore densities, the
second group (cluster 4) contains sites with the lowest
root colonization by vesicles, the third group (cluster 5)
contains sites with the lowest spore densities, and the
fourth group (cluster 6) contains sites with highest root
colonizations (Figs 3b and 4b). The different clusters
showed that there is no simple and direct relationship
between the development of external hyphae and the
development of internal hyphae (Abbott and Robson
1991; Bethlenfalvay et al. 1999).
The extreme characteristics of Amazonian soils can
induce stress on the AMF (Soedarjo and Habte 1995;
Heijne et al. 1996; Stevens and Peterson 1996; Schack-
Kirchner et al. 2000; Mendoza et al. 2002; Escudero and
Mendoza 2005), causing strong responses on the
mycorrhizal variables, colonization by hyphae, vesicles
and arbuscules, external hyphae, density and diversity
of spores. Any small change under these prevailing
Prin 1
3
2
2 3 4
1
1
0
0
–1
–1
–2
–2
–1.5
–1.5
–1.0
–1.0
–0.5
–0.5
0.0
0.0
0.5
0.5
1.0
1.0
1.5
1.5
2.0
ESYA
ESYA2.0
2.5
2.5
Prin 2
Prin 2
Prin 3
(a)
(b)
Figure 2 Distribution of sampling sites according to the principal
components of physical and chemical characteristics in the soil, by
type of relief. (a) Prin1 (pH, soil humidity and organic matter) vs Prin2
(phosphorus availability). (b) Prin2 (phosphorus availability) vs Prin3
(electrical conductivity). (j, Hilly terrain; h, valley)
Arbuscular mycorrhiza in pastures R.H. Posada et al.
136 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140
ª 2007 The Authors
conditions can be decisive for the mycorrhizal develop-
ment in a tropical environment. There were micro-
biological responses which were related to restricted
ranges in the physical, chemical and environmental
characteristics, which were related to the microbiological
responses: in the hilly terrain, the lowest phosphorus
availabilities (8 ppm or less) were associated with colo-
nizations by hyphae up to 18% or to colonization by
vesicles higher than 17%, and a close relationship
between spore density and altitude (Allen et al. 1995)
was found. On the other hand, at the valley air humid-
ities higher to 50% and temperatures above 31�C were
associated with B. decumbens root colonizations (up to
30% by hyphae and up to 5% by vesicles; microbiolog-
ical first principal component).
In conclusion, the effect of physical, chemical factors
of soil and environmental factors on the microbiologi-
cal variables (spore density, external hyphae, root colo-
nizations by vesicles and arbuscules) was related to the
type of relief. Moreover, the behaviour of AMF was
affected only under narrower ranges of values for the
measured variables, being difficult to establish a large
effect of these factors on the AMF conduct. In the
Amazonian foothills, the B. decumbens root colonization
SiteAAY
DCY
APY
JMY
NAY
ESYB
ABYA
FBY
UAY
ARY
ESYA
FGY
GSY
FMY
ABYB
Site
ACX
NAXB
NAXA
VPX
JAX1
JAX2
CNX
GSX
ESX
VAX
Average distance between clusters
Average distance between clusters
0·0 0·2 0·4 0·6 0·8 1·2 1·41·0 1·6
1
2
3
4
5
6
FGX
0·0 0·2 0·4 0·6 0·8 1·0 1·2 1·4 1·6 1·8 2·0
(a)
(b)
Figure 3 Dendrograms based on nearest
neighbourhood method to represent the
microbiological clusters (1, 2, 3, 4, 5, 6) of
sampling sites by type of relief, according to
correlation matrix. (a) In the hilly terrain, (b) in
the valley.
R.H. Posada et al. Arbuscular mycorrhiza in pastures
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140 137

by AMF hyphae and vesicles, and the mycorrhizal spore
density follow different response patterns according to
the type of relief. Some authors suggest that the plant
factors are more important than soil factors (Koomenn
et al. 1987; Mendoza et al. 2002), but this does not
help to understand their significance in the community
structure.
The restoration of zones previously dominated by
forest and currently with pastures of B. decumbens
requires an understanding of the mycorrhizal condition
on soil and their variability. To determine what sites
could favour a fast forest recovery, it is necessary to
consider the availability of the mycorrhizal inoculum,
and to employ this knowledge for selection of short-
cycle species with favourable mycorrhizal responses. The
type of relief is a key factor for determination of
inoculum and root colonization of these pastures,
because of their influence on physical and chemical soil
characteristics.
Acknowledgements
The authors thank the Amazonia University for financial
support and the use of installations; CORPOICA Titaitata
for soil analyses; Geovany Lara Zuluaga, Andres Olaya
Montes, Carlos Alberto Rodrıguez, Wilson Sanchez
Chacon, Adriana Patricia Sanchez Figueroa, Edith Medina
Giron of the Symbiotic Microorganisms Investigations
Team for their work and collaboration during its develop-
ment.
References
Abbott, L.K. and Robson, A.D. (1991) Factors influencing the
occurrence of vesicular-arbuscular mycorrhizas. Agric Eco-
syst Environ 35, 121–150.
Allen, E.B., Allen, M.F., Helm, D.J., Trappe, J.M., Molina, R.
and Rincon, E. (1995) Patterns and regulation of mycor-
rhizal plant and fungal diversity. Plant Soil 170, 47–62.
Andrade, A.S., Queiroz, M.H., Hermes, R.A. and Oliveira, V.L.
(2000) Mycorrhizal status of some plants of the Araucaria
forestand the Atlantic rainforest in Santa Catarina, Brazil.
Mycorrhiza 10, 131–136.
Aristizabal, C., Rivera, E.L. and Janos, D.P. (2004) Arbuscular
mycorrhizal fungi colonize decomposing leaves of Myrica
parvifolia, M. pubescens and Paepalanthus sp. Mycorrhiza
14, 221–228.
Bethlenfalvay, G.J., Cantrell, I.C., Mihara, K.L. and Schreiner,
R.P. (1999) Relationships between soil aggregation and
mycorrhizae as influenced by soil biota and nitrogen nutri-
tion. Biol Fertil Soils 28, 356–363.
Camargo-Ricalde, S.L. and Esperon-Rodrıguez, M. (2005) Efec-
to de la heterogeneidad espacial y estacional del suelo
sobre la abundancia de esporas de hongos micorrizogenos
arbusculares en el valle semiarido de Tehuacan-Cuicatlan,
Mexico. Rev Biol Trop 53, 339–352.
Carvalho, L.M., Correia, P.M., Ryel, R.J. and Martins-Loucao,
M.A. (2003) Spatial variability of arbuscular mycorrhizal
fungal spores in two natural plant communities. Plant Soil
251, 227–236.
Clapp, J.P., Young, J.P.W., Merryweather, J.W. and Fitter, A.H.
(1995) Diversity of fungal symbiosis on arbuscular mycor-
rhizas from a natural community. New Phytol 13, 259–265.
Cuesta, P.A. (1978) Algunas especies forrajeras promisorias en
el Piedemonte del Caqueta. ICA Informa 12, 12–13.
Dickinson, C.H. (1974) Decomposition of litter in soil. In Bio-
logy of Plant Litter Decomposition, Vol. 2, pp. 633–658. ed.
Dickinson, C.H. and Pugh, G.J. London and New York:
Academic Press.
Diederich, C. and Moawad, A.M. (1993) The potential of VA
mycorrhizae for plant nutrition in the tropics. Angew Bot
67, 91–96.
Escudero, V. and Mendoza, R. (2005) Seasonal variation of
arbuscular mycorrhizal fungi in temperate grasslands along
a wide hydrologic gradient. Mycorrhiza 15, 291–299.
V12·067
1·116
0·165
–0·787
–1·738
ABYBFMY
Non-linked SitesGSY
JMY
ESYB
Cluster 1 and 2
APY
ARY
AAYESYA
ABYA
DCYNAY
FBYFGY
UAY
–1·648 –0·696 0·256 1·208 2·160
W1
V12·025
2·025
1·102
1·102
0·179
0·179
–0·744
–1·666–1·666
W1–0·744
NAXBNAXA
VAX
VPX
Cluster 5 Cluster 4
Cluster 6
CNXGSX
ESX FGX
JAX1
JAX2
ACX
(a)
(b)
Figure 4 Canonical relationships between the environmental and soil
characteristics (W) and the microbiological variables (V). (a) In the hilly
terrain. Cluster 1 in grey, cluster 2 in black. (b) in the valley.
Arbuscular mycorrhiza in pastures R.H. Posada et al.
138 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140
ª 2007 The Authors

Fontenla, S., Godoy, R., Rosso, P. and Havrylenko, M.
(1998) Root associations in Austrocedrus forests and sea-
sonal dynamics of arbuscular mycorrhizas. Mycorrhiza 8,
29–33.
Gehring, C.A. and Connell, J.H. (2006) Arbuscular mycorrhizal
fungi in the tree seedlings of two Australian rain forests:
occurrence, colonization, and relationships with plant per-
formance. Mycorrhiza 16, 89–98.
Heijne, B., van Dam, D., Heil, G.W. and Bobbink, R. (1996)
Acidification effects on vesicular-arbuscular mycorrhizal
(VAM) infection, growth and nutrient uptake of estab-
lished heathland herb species. Plant Soil 179, 197–206.
Herrera, R., Rodrıguez, A. and Furrazola, E. (1986) Metodo para
determinar la biomasa de micelio extramatrico vesiculo-
arbuscular. In Ciclo lectivo sobre el tema: Tecnicas de investi-
gacion en micorriza, pp. 18–28. Turrialba, Costa Rica.
IGAC (1999) Metodos analıticos de CALS, recomendaciones
establecidas para la reunion de representantes de laboratorios
efectuada en el IBAC. Bogota, Colombia: IGAC.
IGAC-INPA (1993) Aspectos ambientales para el ordenamiento
territorial del occidente del Departamento de Caqueta, Vol.
2. Bogota, Colombia: IGAC-INPA, Editorial Tropembos.
Koomenn, I., Grace, C. and Hayman, D.S. (1987) Effectiveness
of single and multiple mycorrhizal inocula on growth or
clover and strawberry plants at two soil pHs. Soil Biol Bio-
chem 16, 539–544.
Koske, R.E. (1987) Distribution of VAM fungi along a latitudi-
nal temperature gradient. Mycologia 79, 55–68.
Lodge, D.J. (1989) The influence of soil moisture and flooding
on formation of VA-endo-and ectomycorrhizae in Populus
and Salix. Plant Soil 117, 243–253.
Malagon, D., Pulido, C., Llinas, R.D. and Chamorro, C. (1995)
Suelos de Colombia. Bogota, Colombia: IGAC.
Mendoza, R., Goldmann, V., Rivas, J., Escudero, V., Pagani,
E., Collantes, M. and Marban, L. (2002) Poblaciones de
hongos micorrızicos arbusculares en relacion con propied-
ades del suelo y planta hospedante en pastizales de Tierra
del Fuego. Ecol Austr 12, 9–20.
Miller, S.P. (2000) Arbuscular mycorrhizal colonization of
semi-aquatic grasses along a wide hydrologic gradient.
New Phytol 145, 145–155.
Miller, S.P. and Bever, J.D. (1999) Distribution of arbuscular
mycorrhizal fungi in stands of the wetland grass Panicum
hemitomon along a wide hydrologic gradient. Oecologia
119, 586–592.
Oliveira, L.A., Moreira, F.W. and Moreira, F.M. (1997) Ocor-
rencias de microorganismos beneficos em ecossistemas
amazonicos. In Duas decadas de contribuicoes do INPA a
pesquisa agronomica no tropico umido ed. Noda, H., Souza,
L.A.G. and Fonseca, O.J.M. pp. 221–240 Manaus: Instituto
Nacional de Pesquisas da Amazonia.
Pabon, A. (2000) Presencia de estructuras similares a las que
componen el hongo de micorriza arbuscular en hojarasca de
Cedrela montana Turczaninov. BSc Thesis, Bogota: Pontifi-
cia Universidad Javeriana.
Pattinson, G.S., Sutton, B.G. and McGee, P.A. (2000) Leachate
from a Waste Disposal Centre reduces the initiation of
arbuscular mycorrhiza, and spread of hyphae in soil. Plant
Soil 227, 35–45.
Philips, J.M. and Hayman, D.S. (1970) Improved procedures
for clearing roots and staining parasitic and vesicular-
arbuscular mycorrhizal fungi for rapid assessment of
infection. Trans Br Mycol Soc 55, 158–161.
Porter, W.M., Robson, A.D. and Abbott, L.K. (1987a) Factors
controlling the distribution of vesicu1ar arbuscu1ar mycor-
rhizal fungi in re1ation to soil pH. J Appl Ecol 24, 663–
672.
Porter, W.M., Robson, A.D. and Abbott, L.K. (1987b) Field
survey of the distribution of vesicu1ar arbuscu1ar mycor-
rhizal fungi in relation to soil pH. J Appl Ecol 24, 659–662.
Posada, R.H. (2001) Presencia de estructuras de posibles hongos
de micorriza arbuscular en hojarasca de un bosque primario
y un sistema agricultural en los llanos orientales, 143 pp.
MSc Thesis, Bogota: Universidad de los Andes.
Read, J., Koucheki, H.K. and Hodgson, J. (1976) Vesicular-
arbuscular mycorrhiza in natural vegetation systems. New
Phytol 77, 641–653.
Rickerl, D.H., Sancho, F.O. and Ananth, S. (1994) Vesicular-
arbuscular endomycorrhizal colonization of wetland plants.
J Environ Qual 23, 913–916.
Robertson, G.P., Klingensmith, K.M., Klug, M.J., Paul, E.A.,
Crum, J.R. and Ellis, B.E. (1997) Soil Resources, microbial
activity, and primary production across and agricultural
ecosystem. Ecol Appl 7, 158–170.
Schack-Kirchner, H., Wilpert, K.V. and Hildebrand, E. (2000)
The spatial distribution of soil hyphae in structured
spruce-forest soils. Plant Soil 224, 195–205.
Sieverding, E. (1983) Manual de metodos para la investigacion
de la micorriza vesıculo-arbuscular en el laboratorio. Cali:
Centro Internacional de Agricultura Tropical (CIAT).
Simpson, D. and Daft, M.J. (1990) Interactions between water
stress and different mycorrhizal inocula on plant growth
and mycorrhizal development in maize and sorghum.
Plant Soil 121, 179–186.
Siqueira, J.O. and Saggin-Junior, O.J. (2001) Dependency
on arbuscular mycorrhizal fungi and responsiveness of
some Brazilian native woody species. Mycorrhiza 11,
245–255.
Siqueira, J., Rocha, J.W.F., Oliveira, E. and Colozzi-Filho, A.
(1990) The relationship between vesicular-arbuscu1ar
mycorrhiza and lime: associated effects on the growth and
nutrition of Brachiaria grass (Brachiaria decumbens). Biol
Fert Soils 10, 65–71.
Siqueira, J.O., Carbone, M.A., Curi, N., Silva, S.C. and Davide,
A.C. (1998) Micorrhizal colonization and mycotrophic
growth of native woody species as related to successional
groups in Southeastern Brazil. For Ecol Manage 107,
241–252.
Soedarjo, M. and Habte, M. (1995) Mycorrhizal and nonmy-
corrhizal host growth in response to changes in pH and P
R.H. Posada et al. Arbuscular mycorrhiza in pastures
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140 139

concentration in a manganiferous oxisol. Mycorrhiza 5,
337–345.
Stevens, K.J. and Peterson, R.L. (1996) The effect of a water
gradient on the vesicular-arbuscular mycorrhizal status of
Lythrum salicaria L. (purple loosestrife). Mycorrhiza 6, 99–
104.
Van der Heijden, M.G.A. and Sanders, I.R. (2002) Mycorrhizal
ecology: synthesis and perspectives. In Mycorrhizal Ecology
pp. 441–456. ed. Van der Heijden, M.G.A and Sanders,
I.R. Berlin, Heidelberg, New York: Springer
Velasquez, J.E. and Cuesta, P.A. (1990) Productividad animal
de Brachiaria decumbens (Stapf) bajo pastoreo continuo
con tres cargas en el piedemonte amazonico. Livestock
Research for Rural Development 2, 2. Available via
DIALOG. http://www.cipav.org.co.ezproxy.library.tufts.edu/
lrrd/lrrd2/3/cont23.htm (Cited June 2006).
Wang, G.M., Stribley, D.P., Tinker, P.B and Walker, C. (1993)
Effects of pH on arbuscular mycorrhiza I. Field observa-
tions on the long-term liming experiments at Rothamsted
and Wobum. New Phytol 124, 465–472.
Arbuscular mycorrhiza in pastures R.H. Posada et al.
140 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 132–140
ª 2007 The Authors

HOME / LIFE SCIENCES / MICROBIOLOGY & VIROLOGY / APPLIED MICROBIOLOGY /
CART | MY ACCOUNT | CONTACT US | HELP
Journal of Applied MicrobiologyIncluding Letters in Applied Microbiology & Annual Symposium
Published on behalf of the Society for Applied Microbiology
Edited by:A. Gilmour
Print ISSN: 1364-5072Online ISSN: 1365-2672Frequency: MonthlyCurrent Volume: 110 / 2011
Editorial Information
Chief EditorA. Gilmour, Agri-Food and Biosciences Institute, Northern Ireland, UKFax: +44 (0) 28 90255009email: [email protected]
Managing EditorK. Brister, Wiley-Blackwell, Oxford, UKemail: [email protected]
EditorsB. Austin, Institute of Aquaculture, University of Stirling, Stirling, UKM.M. Bagdasarian, Michigan State University, East Lansing, MI, USAL. Baillie, Welsh School of Pharmacy, Cardiff University, Cardiff, UKM.D. Barton, University of South Australia, Adelaide, AustraliaE. Bartowsky, The Australian Wine Research Institute, Glen Osmond, AustraliaA.M. Bojesen, Department of Veterinary Disease Biology, University of Copenhagen, DenmarkA. Bosch, Universitat de Barcelona, Barcelona, SpainP. Calik, Middle East Technical University, Ankara, TurkeyM.L. Chikindas, The State University of New Jersey, New Brunswick, NJ, USAP.J. Collier, University of Abertay, Dundee, UKK.L. Cook, USDA-ARS Animal Waste Management Research Unit, Bowling Green, KY, USAJ.E. Cooper, Queen's University of Belfast, Belfast, UKD.A. Cowan, University of the Western Cape, Bellville, Capetown, South AfricaS.P. Cummings, School of Applied Sciences, Northumbria University, Newcastle upon Tyne, UKS.M. Cutting, Royal Holloway, University of London, UKL.M.T. Dicks, University of Stellenbosch, Matieland, South AfricaM. Dow, BioSciences Institute, University College Cork, Cork, IrelandN. Fegan, Food Science Australia, Brisbane, AustraliaC. Gilbert, Laboratoire de Microbiologie et Génétique Moleculaire, Université Claud Bernard Lyon, Lyon, FranceJ. Glassey, University of Newcastle Upon Tyne, Newcastle upon Tyne, UKA.E. Glenn, USDA-ARS Toxicology & Mycotoxin Research Unit, Athens, GA, USAP.C. Gowland, Staffordshire University, Stoke-on-Trent, UKD.A. Grinstead, Diversey Lever Innovation Center, Cincinnati, OH, USAJ. Guard, USDA/ARS, Athens, GA, USAJ.H. Hill, Department of Plant Biology, Iowa State University, Ames, IA, USAC.R. Jackson, USDA-ARS Resistance Research Unit, Athens, GA, USAF. Jorgensen, Health Protection Agency, Food Water and Environmental Microbiology Network, Porton Down, Salisbury, UKT. Kuchta, Food Research Institute, Bratislava, SlovakiaG. LaPointe, Laval University, Quebec, CanadaA. Leaphart, Clemson University, Clemson, SC, USAN. Lima, MUM, Minho University, Braga, PortugalJ. Lisle, US Geological Survey, Center for Coastal & Watershed Studies, St Petersburg, FL, USAG.T. Macfarlane, MRC Microbiology and Gut Biology Group, Ninewells Hospital and Medical School, Dundee, UKS. Macfarlane, University of Dundee, Dundee, UKJ.-Y. Maillard, Welsh School of Pharmacy, Cardiff University, Cardiff, UKD.V. Mavrodi, Washington State University, Pullman, WA, USAB. Mayo, IPLA-CSIC, Villaviciosa, SpainA. McBain, University of Manchester, Manchester, UKJ.A. McGarvey, USDA-ARS Plant Mycotoxin Research Unit, Albany, CA, USAA. Mohagheghi, National Renewable Energy Laboratory, Golden, CO, USAF. Mozzi, CERELA-CONICET, Tucumán, ArgentinaK. Nickerson, Biological Sciences, University of Nebraska, Lincoln, NE, USAD.R. Noguera, University of Wisconsin, Madison, WI, USAC.E. Nwoguh, Health Protection Agency, CEPR, Porton Down, Salisbury, UKG-J.E. Nychas, Department of Food Science & Technology, Laboratory Food Microbiology & Biotechnology of Foods, Athens,GreeceG.K. Paterson, Department of Veterinary Medicine, University of Cambridge, Cambridge, UKC.A. Phillips, University of Northampton, UKW. Qin, Lakehead University, Thunder Bay, CanadaC. Rees, University of Nottingham, Sutton Bonington, UKS. Roller, London South Bank University, London, UKC.P. Saint, SA Water Centre for Water Management and Re-use, University of South Australia, Mawson Lakes, AustraliaR. Seviour, Biotechnology Research Centre, La Trobe University, Bendigo, AustraliaP. Silley, MB Consult Limited, Hampshire, UKT.J. Smith, Biomedical Research Centre, Sheffield Hallam University, Sheffield, UKM.B. Taylor, Department of Medical Virology, NHLS/University of Pretoria, Pretoria, South AfricaK.Thomas, University of Sunderland, Sunderland, UKV.P. Valdramidis, University of Malta, MaltaA. Venâncio, Dept de Engenharia Biológica, Universidade do Minho, PortugalK. Venema, TNO Nutrition and Food Research, The NetherlandsT.M. Wassenaar, Molecular Microbiology and Genomics Consultants, Tannenstrasse, Zotzenhein, GermanyJ. Wells, USDA-ARS, Meat Animal Research Center, NE, USAR. Zdor, Andrews University, Berrien Springs, MI, USAX.-H. Zhang, College of Marine Life Sciences, Ocean University of China, Qingdao, China
View content online
Journal of Applied Microbiology: Including Letters in Applied Microbiol... http://www.wiley.com/bw/editors.asp?ref=1364-5072&site=1
1 de 2 08/01/2012 06:18 p.m.




















