
Forest recovery in abandoned tropical pastures in Puerto Rico

Frugivorous Bats in Isolated Trees and Riparian Vegetation Associated withHuman-Made Pastures in a Fragmented Tropical Landscape
Jorge Galindo-Gonzaacutelez Vinicio J Sosa
The Southwestern Naturalist Vol 48 No 4 (Dec 2003) pp 579-589
Stable URL
httplinksjstororgsicisici=0038-49092820031229483A43C5793AFBIITA3E20CO3B2-C
The Southwestern Naturalist is currently published by Southwestern Association of Naturalists
Your use of the JSTOR archive indicates your acceptance of JSTORs Terms and Conditions of Use available athttpwwwjstororgabouttermshtml JSTORs Terms and Conditions of Use provides in part that unless you have obtainedprior permission you may not download an entire issue of a journal or multiple copies of articles and you may use content inthe JSTOR archive only for your personal non-commercial use
Please contact the publisher regarding any further use of this work Publisher contact information may be obtained athttpwwwjstororgjournalsswanhtml
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission
The JSTOR Archive is a trusted digital repository providing for long-term preservation and access to leading academicjournals and scholarly literature from around the world The Archive is supported by libraries scholarly societies publishersand foundations It is an initiative of JSTOR a not-for-profit organization with a mission to help the scholarly community takeadvantage of advances in technology For more information regarding JSTOR please contact supportjstororg
httpwwwjstororgThu Aug 9 192443 2007
FRUGIVOROUS BATS IN ISOLATED TREES AND RIPAFUAN VEGETATION ASSOCIATED WITH HUMAN-MADE PASTURES IN A
FRAGMENTED TROPICAL LANDSCAPE
Biotecnologia jEcologia Aplicnda Cniuersidad Veramuzann A P 250 Xalapa Vmcrur 91001 Mixiro lJGG)
Institute de Ecologin A C A P 63 Xalapa Verarrur 91000 Mixico (155) Correspondent jgal indo~umx
~ S T R ~ C T - C O ~ ~ ~ ~ S ~ O ~of tropical rain forest to pasture and cropland often leads to a loss of local species However information on the role of some isolated elements of the fragmented landscape on maintaining biodiversity is lacking This study carried out in human-made pastures in Veracruz Mexico demonstrated the year-round presence of frugivorous bats in isolated trees inside pastures and in riparian corridors of vegetation among pastures During a 1-year study (October 1995 through September 1996) we captured 652 bats belonging to 20 species repre- senting 56 of the total species recorded for the area Higher abundance and diversity of bats were recorded for riparian sites than for isolated trees Of the bats we captured 82 were fru- givorous 15 insectivorous 2 nectarivorous-frugivorous and 2 sanguivorous Bats visited iso- lated trees regardless of tree species or phenological state Abundance decreased with distance from the nearest forest fragment We discuss how riparian corridors and isolated trees in pastures might contribute to maintaining diversih of frugivorous bats and plants in a fragmented landscape
RESUMEN-La transformation de las selvas tropicales en pastizales 7 areas de cultivo frecuente- mente conduce a la perdida de especies locales Sin embargo falta informaci6n sobre el papel de algunos elementos aislados del paisaje fragmentado en el mantenimiento de la biodiversidad Este estudio registr6 la presencia de murciilagos frugivoros durante todo el ano bajo arboles aislados y en la vegetaci6n riparia remanente en pastizales antropogknicos en Veracruz Mexico Durante un aho de estudio (octubre 1995-septiembre 1996) capturamos 652 murciilagos de 20 especies que representan el 36 de las especies registradas en el irea Los sitios en la vegetation riparia registra- ron mayor abundancia 7 diversidad que 10s sitios bajo irboles aislados El 82 del total de murcik- lagos fueron frugivoros 15 insectivoros 2 nectarivoros-frugivorosy 2 sanguivoros Los murcie- lagos lisitaron 10s irboles aislados independientemente de la especie del irbol o de su estado fen016 gico La abundancia disminuyo a mayor distancia del fragment0 forestal mis cercano Discutimos c6mo 10s corredores riparios y irboles aislados en 10s pastizales pueden contribuir a1 mantenimiento de cierta diversidad de murciklagos 7 plantas en el paisaje fragmentado
Conversion of tropical rain forests to pas- number of functional groups and in species in tures and croplands often leads to a loss of lo- tropical ecosystems (Heithaus et al 1975 cal plant and animal species because many Fleming 1988) They affect biological com-species are sparsely distributed or intolerant to munities through predation pollination and open sites (Turner 1996) Human-made pas- seed dispersal (Vizquez-Yanes et al 1975 tures with little vegetative cover have been Fleming et al 1977 Fleming and Heithaus identified as a barrier to natural dispersal of 1981 Fleming 1982 Bonaccorso and Hum- species (Guevara et al 1986 Aide and Cave- phrey 1984 Thomas et al 1988 Fleming and lier 1994) Distance between forest fragments Williams 1990 Charles-Dominique 1991 Kal- also might affect species abundance (Estrada ko 1997) A small number of published studies et al 1993 Andsen 1994 Galindo-Gonzalez have specifically dealt with patterns of bat di- 1999) Bats like many groups of plants and an- versity within fragmented landscapes (de Jong imals reach their highest diversity both in 1995 Brosset et al 1996) including artificial
580 Thp Southwestm Naturalist vol 48 no 4
gaps (02 ha) produced by selective logging (Crome and Richards 1988) strips (30 X 150 m) cleared for logging (Gorchov et al 1993) 2 trees in a pasture (Janzen et al 1976) and an abandoned pasture (Nepstad et al 1996) Specifically one study conducted in the tropi- cal region of Los Tuxtlas Mexico reported on how bats differentially used habitats within modified landscapes in the continuous forest 91 of the total number of species (ie 35 species) were caught followed by 40 to 60 in plantations shaded by rain forest trees 43 to 51 in unshaded plantations and 34 in live fences (Estrada et al 1993) Although not cap- tured some bats (including the insectivorous Pteronotus) have been observed flying across pastures above net levels (Estrada et al 1993 Galindo-Gonzilez 1999)
Based on foraging behavior food habits and seed rain attributed to bats utilizing pastures Galindo-Gonzilez (1998) suggested that frugiv- orous bats are important in the regeneration of tropical rain forest in abandoned pastures In many tropical regions 2 kinds of habitats are created by the conversion of forest to pas- ture 1) strips of riparian vegetation are often left behind along watercourses to protect against erosion and 2) isolated trees are left standing to provide shade for cattle firewood for cooking or for aesthetic reasons These iso- lated trees often serve as perches for birds and bats (the presence of the latter is inferred from nocturnal seed rain) that bring viable seeds to pastures from nearby forest remnants or ripar- ian vegetation (Guevara and Laborde 1993 Galindo-GonzBlez et al 2000) On the other hand riparian vegetation might function as corridors for frugivorous bats and as a source of propagules for tropical forest species (Bon- accorso and Humphrey 1984 Malanson 1993 Naiman et al 1993 Knopf and Samson 1994 Rautenbach et al 1996 Beier and Noss 1998) These studies strongly suggest that fi-ugivorous bats travel across pastures and visit isolated trees while foraging commuting or looking for roosts or to decrease predation risk how- ever we need more information on how bats use isolated trees and riparian vegetation in fragmented habitats and their significance for forest regeneration and maintenance of bat di- versity
Visits to isolated trees in pastures might be a function not only of their quality as roosts
food or shelter but of their degree of isolation (ie distance from the nearest patch of for- est) Frugivorous bats are known to travel long distances when foraging or commuting from their day roosts to their feeding areas (Galin- do-Gonzklez 1998) Assuming that pastures are a less suitable habitat than forest remnants both in terms of food resources and shelter it is reasonable to predict that the probabilih of recording a bat at isolated trees decreases with distance from sources of bats
The general purpose of this study was to de- scribe the frugivorous bat community captured in ground-level mist-nets set under isolated trees and in riparian vegetation (remnants of the original vegetation canopy) associated with pastures in a fragmented tropical landscape Specifically we wanted to determine species richness and relative abundance in this com- ponent of the bat community We predicted 1) more captures at trees with seeds that are bat- dispersed rather than wind-dispersed 2) that captures at isolated Ficus trees in pastures would be highest when these trees bore ripe fruits and 3) that bat abundance would be in- versely proportional to distance to the nearest forest fragment We specifically emphasized fruit bats because of their potential impor- tance in seed dispersal and forest regeneration (Galindo-Gonzalez 1999 Galindo-Gonzale~ et al 2000)
METHODS-StudyA r e ~ F i e l d w o r k was conducted i n t h e northeastern portion o f t h e Sierra d e Los Tuxtlas Veracruz Mexico (1836N 9505W) t h e northernmost limit o f rain forest i n North America (Dirzo and Miranda 1991) Mean annual tempera- ture is 27degC Total annual rainfall is ca 4725 m m with a relatively dry period between March and May ( 1 117 2 117 m m m o n t h ) and a rainy period be-tween J u n e and February (486 i- 870 m m m o n t h Guevara e t al 1992) Cndisturbed vegetation and forest fragments consisted o f tropical rain forest with a closed canopy 30 to 35 m high Plant communit ies were dominated by Astrocaryum mexicanurn (Areca-ceae) i n t h e understory Pseudolmedia oxyphyllana (Moraceae) i n t h e m e d i u m stratum and A~ctandm anzbigens (Lauraceae) i n t h e canopy (Bongers e t al 1988) Land management practices have converted 65 t o 80 o f these forests t o cattle pastures (Dirzo and Garcia 1992)
A t present t h e study area is a heterogeneous land- scape Specifically it is 1 ) dominated by human- m a d e o p e n pastures for cattle grazing and crop- lands (56 of surface area) 2 ) interspersed with
581 December 2003 Galindo-Gonrilez and Sosa-Frugivorous
small forest remnants (36) and 3) interconnected by riparian vegetation and live fences (4) (Guevara et al 1998) Many isolated trees occur in pastures (04 to 119 treesha Guevara et al 1998) Trans- formation of the area into pastureland began in the 1960s and since that time cattle have been grazing on a year-round basis
The st~ldy site was located within a clear-cut basin (the average distance between forested ridges was 25 km) with forest fragments distributed along the surrounding ridge Plant communities within pas- tures were dominated by early successional species from the families Piperaceae Solanaceae and Me- lastomataceae and by isolated trees from the fami- lies h4oraceae and Cecropiaceae whose fruits are known to be eaten by bats (Galindo-GonzPlez 1999 Galindo-GonzPlez et al 2000) Riparian vegetation consisted o f remnant trees and shrubs of 112 species (these represented 71 of the species from the orig- inal forest) located on both sides of a stream whose banks were 2 to 5 m wide Riparian remnant trees were taller than 18 m Their closed canopy over the stream formed a tunnel-like flight corridor for birds and bats (for more detail on floristic composition and structure of vegetation in pastures see Guevara et al 1992)
Samplzng Strategy-We selected 8 nettlng sites rep- resenting 3 habitat types or elements of the frag- mented landscape to sample for bats and measure fruit availability 1) isolated trees producing bat-dis- persed seeds 2) isolated trees producing wind-dis- persed seeds (control) and 3) riparian vegetation Netting sites were placed under the canop of each of the following isolated trees (all trees 25 to 30 in h ~ g h )Izcus ~oponenszsFtcus coluhnae Cezba pentan dra and Cedrella odorata Vertebrates disperse the seeds of the 2 former species and the latter 2 are wind-dispersed Ceiba pentandra is a bat-pollinated tree flowering between December and March how- ever we did not record flowering of this tree during the study period Fzcus was considered particularly important because it is the most common tree ge- nus encountered among isolated trees in pastures (Guevara et al 1986) Isolated trees were 250 m from any forested fragment or riparian vegetation The remaining 4 sites were randomly selected along the main vegetated stream in the study area Netting sites varied in distance from a forest fragment Ri-parian sites were the nearest to forest (95 to 276 m) and isolated trees were the most distant (280 to 381 m) The distance from each isolated tree to the near- est forest fragment (a forested area gt1 ha) was nlea- sured on an aerial photograph (1991) scale l 75000 measurements of distance between 5 refer- ence points in situ were used to calibrate measure- ments Each month ~e recorded presence-absence of ripe ficus fruits for later correlation with bat abundance We then calculated the percentage of
bats in fragmented tropical landscape
the crown that was covered with fruits and estimated the proportion of ripe and unripe fruits (see meth- ods in Chapman et al 1992)
Bat CapturesBats were captured from October 1995 through September 1996 using mist-nets placed at ground level Mist-netting at ground level is known to be biased for fruit-eating bats and against high-flying insectivorous bats bats with low body mass and some species that tend to be cap- tured in the canopy stratum of tropical forests (Rau- tenbach eta] 1996 Kalko 1997 Bernard 2001 Kal- ko and Handley 2001) Two mist-nets (each 12 X
25 m) were placed in an L-shaped array under the canopy of isolated trees At each riparian site we used 1 net (6 X 25 nl) placed across the stream because each stream was lt5 m wide and because preliminary sampling showed that bat captures were more frequent in this habitat Netting sites were sam- pled once each month around the time of the new moon ( 2 3 days) to avoid lunar phobia (hlorrison 1978b) An a-erage of 41 hnight (SE 005) and 33 nightsmonth (SE + 02) were dedicated to sam- pling Nets were opened 30 min after sunset Total sampling effort in this study was 261320 net-m X
hours derived from 235 h of netting 1112 m of net during a total of 40 netting nights All captured bats were identified to species using field keys (hledellin et a] 1997 see Ramirez-Pulido et al 1996 for no- menclature) and fitted with unique color-coded plastic necklaces (Dennison Secur-A-Tie Framing- ham Massachusetts USA) Sex mass (to nearest 01 g) reproductive condition and time and site of cap- ture were recorded
Data AnalysisBat community structure was deter- mined according to family species and foraging- guild as well as by relative species abundance Spe- cies richness in each habitat type was compared through accumulation species f~mctions which pre- dict the maximum number of species (the asymp- tote) present in a certain area based on the cumu- lative number of species (Sober611 and Ilorente 1993) Several functions can be derived depending on the assumptions made about the probability of adding a new species after a collecting period UTe chose the exponential function [St = ab(l - bt) where S is species richness at time t and a h param-eters] which assumes that the probability of adding new species decreases linearly with the size of the list This function might be adequate when sampling a relatively small area or a well kuuwn group or both (our case) and eventually all species will be recorded It also allows one to estimate confidence intervals for S(t) which is the number of species collected at time tk (the time needed to lower the increase rate of the accumulated species list in k pro-portion Sober6n and Llorente 1993) We fitted the function by non-linear regression using the quasi-
582 Thr Southwestern iVaturaltst vol 48 no 4
Newton algorithm in the package STATISTICA (Statsoft 1991)
Due to our use of mist-nets of different sizes we calculated adjusted relative abundance (number of individuals per 1000 net-m X hour) by sampling ef- fort with the following formula adjusted relative abundance = 1000 [b (nh)] where b = total Ilum- ber of captured bats and nh = sampling effort as already defined Bat species diversity for each habitat type (ie riparian vegetation isolated Fzcus trees and isolated control trees) was estimated with the Shannon-Uiener index (H) using the total annual captures Diversity indices among habitat ty-pes were then compared using a I-way ANOVA test applied to ranks The Morisita-Horn index (CmH) was used to compare species similarity among habitats (Magur- ran 1988)
We conducted ANOVA and Tukey pairwise mul- tiple comparisons to check whether total relative abundance (ie total bat species captured during the year) differed among habitat types We also used ANOVA to determine whether the relative abun- dance of frugivorous bats differed between fruiting Ficus trees and non-fruiting Ceibu and Cedrella trees (ie controls) in the study area For this analysis we used total capture of frugivorous bat5 from April through August 1996 months when Ficus trees bore fruits Relative abundance was normalized b a square root transformation before conducting these analyses Linear regression analysis was conducted to determine the association between relative abun- dance of all bats and the percentage of canopy with ripe fruits at isolated Ficus trees in pastures Finally simple linear correlation analysis was conducted to determine the degree of association behveen dis- tance to the nearest forest fragment and bat abun- dance in riparian vegetation and around isolated trees in pastures
RESULTS-Species Richness and Community Structurewe captured 652 bats from 20 spe- cies in 4 families (Table 1) Most species were from the family Phyllostomidae (15 species) The families Mormoopidae Vespertilionidae and Molossidae were represented by 5 species (Table 1) As expected frugivorous bats were the most abundant (81 of the total captures) followed by insectivores (15) nectarivore- frugivores (2) and sanguivores (2) Total relative abundance varied greatly with species Five species accounted for 84 of the cap- tures These were the frugivores Sturnira liliurn (40) Artibeus jamaicensis (15) Carollia per-spinllata ( lo) and Dermanura tolteca (9) and the insectivore Pteronotus parnellii (10)
The cumulative number of species over all
habitat types tended to level off after 20 nights of netting The species accumulation function predicted a maximum of 23 species According to Moreno and Halffter (1999) 19 nights at least are enough to record 90 of frugivorous bats in a heterogeneous habitat Therefore our sampling provided a reasonably good es- timation of species richness at ground level The maximum numbers of bat species for each habitat predicted by the accumulation func- tion were 17 13 and 12 for the riparian the Acus trees and control trees respectively (Fig l ) and we captured 18 13 and 12 species respectively
Bat diversity varied among habitat types Abundance species richness and Shannon-Wiener index (H)were higher in riparian veg- etation than under isolated trees in pastures despite the lower sampling effort in the ripar- ian habitat (Table 2) Some species were pre- sent only in riparian vegetation whereas others used both habitats (Table 1) The adjusted rel- ative abundance of all bats differed among habitat types (ANOVA F = 2184 df = 2 P lt 0001 Fig 2) According to a posteriori multi- ple comparison tests relative abundance was significantly higher in the riparian habitat than at isolated Ficus trees (q = 833 P lt 005) or control trees (q = 784 P lt 005) relative abundance was not significantly different be- tween isolated Ficus trees and control trees ( q = 048 P gt 003) The Shannon-Wiener index was marginally different between the riparian and control trees (F = 581 P = 006 Table 2) Species similarity was highest between iso- lated trees (ie between Ficus and corltrol trees) and lowest between isolated trees and riparian habitats (Table 2) Bats also were cap- tured in different habitat types according to their feeding habits In particular insectivo- rous bats were more abundant in riparian sites (70) than in isolated tree sites (30) in con- trast frugivores were more abundant at isolat- ed trees (75) than at riparian sites (25)
Use of Isolated Trees and fiparinn Vegetation Habitats-Frugivorous bats were caught under the canopies of isolated trees irrespective of the seed dispersal mechanism (bat or wind-dis-persed) or phenological state of the tree Rela- tive abundance for frugivorous bats at control trees was slightly higher than at ficus trees dur- ing most of the year However no significant differences were found among mean relative
589 1)ecember 2003 Oalindo-Gonrilez and Sosa-Frugivorous bats in fragmented tropiral landscape
TABLE1-Bat species recorded in a fragmented landscape in Los Tuxtlas lkracruz Mexico October 1995 through September 1996 Habitat type I = under isolated tree R = riparian (number of individuals cap- tured)
Relative Habitat type
abundance n ( ) I K Feeding guild
Mormoopidae
Pteronotus dauyi 8 12 1 7 insectivore Pteronotus parnelliz 64 98 19 43 insectibore Mor~nocr megalophylla 16 25 2 14 insectivore
Phvllostomldae
Phyllostorninae Phgtllostomus dzscolor 1 02 0 1 frugivore
Carolliinae Carollza hreurcauda 1 02 0 1 frugivore Carollza perspzczllata 65 99 28 37 frugivore
Glossophaginae Glossophaga soricina 10 15 s 3 nectar-frugivore Choeronzscus godmani 1 02 0 I nectar-frugivore
Stenoderrnatlnae Sturnmi izlzum frugirore Di-oderma bzlobatuni frugivore Platyrrhznus hellm frugivore Vamnpyrodes caracczolz frugivore Chzrod~rma uzllosum frughore Artzbeu lamazcenzi frugivore Artzbvus lzturatus frugivore Dermanura toltrca frugivore Dernlanura pharotzs frugivorc
Desmodontinae Desmodus rotundus 10 15 4 6 sanguivore
Vcspertilionidae
My otis keaysz 11 17 0 11 insectivore
Molossidae
iMolossuc rufus I 02 1 0 insectivore
Total 652 391 261
abundance values for frugivorous bats at control parian sites and bats moved from 1 habitat vs Ficus trees during their fruiting seasons (AN- type to another (ie among riparian sites iso- OVA F = 035 P = 057 df = 1)Finally no lated Ficus trees and control trees) Most re- significant relationship was found between bat captures (20 individuals) were caught on sep- abundance and percentage of ripe Ficus fruits arate nights and at different sites and habitats in isolated trees ( r = 027 P = 066 n = 5) from where they were first captured Distance
We recaptured 22 individuals belonging to 3 among recapture sites ranged from 117 to 692 frugivorous species a total of 34 times (Table m (mean 2805 n~ 234 m SE) Recapture 3) Carollia perspiczllata was the most recaptured times range from a few hours in the same night species even though Sturnira lilium was more to 8 months after the first capture Recaptured abundant indicating that the former shows bats did not seem to have any harm caused by more fidelity to the studied pastures than the the necklaces latter We recorded more recaptures at the ri- Relative abundance of all bats decreased
584 The Southwestern Vuturalzst vol 48 no 4
20 1 A A A
0 Ficus
+ Control
+ R~par~an
A Total 1995 Months 1996
0 v 0 5 10 15 20 25 30 35 40 45
-0 -Riparian -v-Ficus trees -0-Control trees
Accumulated netting nightslmonth FIG2-Mean adjusted relative abundance of bats
FIG l-Accumulation curves (lines) for bat spe- in different habitat t7rpes of
ties adjusted by exponential function for each hab- landscape a -year at Tuxtlas Ve-itat and for total captures in pastures at Los Tuxtlas racruZ
Veracruz Mexico
with increasing distance from the nearest for-est edge (Fig 3 ) To eliminate potential bias produced by the presence of water (which could function as an attractor to bats) at ripar-ian sites we analyzed separately 1) the corre-lation between bat relative abundance and dis-tance to isolated trees ( r = -072 P gt 005) and 2) the relationship between bat relative abundance and distance to riparian sites ( r =
-095 P = 005) Most bats were captured in the riparian habitat located nearest to a forest fragment The fewest number of bats were cap-tured at isolated Ficus trees located the greatest distance from the forest edge
D~scus s~o~-Ourresults show that bats (mostly frugivorous species) flew around iso-
lated trees and in strips of riparian vegetation in pastures within a fragmented tropical land-scape while foraging commuting or looking for perches Bat abundance richness and di-versity were higher in riparian corridors than under the canopy of isolated trees Frugivorous bats were caught under the canopies of differ-ent isolated trees throughout the year irre-spective of the seed dispersal mechanism of the tree or the presence or absence of fruits As predicted bat abundance decreased with distance to the nearest forest fragment
In both the riparian and isolated tree sites frugivorous species from the family Phyllo-stomidae were most common Species in the subfamily Stenodermatinae were more numer-ous (9 out of 12 potentially present) than the subfamily Phyllostominae (1 out of 7 poten-
TABLE2-Bat species richness sampling effort (SE h = netting hours m = total meters of installed nets) adjusted relative abundance (ARA see text for units) mean Shannon-Wiener diversity index (IF) and pairwise comparisons of species similarity among habitats (Morisita-Horn index CmH) on different habitat types sampled in pastures in Los Tuxtlas Veracruz Mexico
BatsSpecies expected
Species cap- SE Mean Habitat type n observed St CI tured (h X m) ARA IF CmH2
- - ---
Fzcus (a) 2 13 113 88-137 150 36324 413 0776 097 (a-b) Control trees (b) 2 12 112 99-125 241 48654 495 0670 083 (b-c) Riparian (c) 3 18 156 141-172 261 8096 3224 0824 090 (c-a)
Total 652
At time tk when an increase of 5 (k = 005) in the species list is predicted according to the exponential species accumulation function Lower and upper 93 confidence limits shown
CmH = 1 (100 similarity)
December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

FRUGIVOROUS BATS IN ISOLATED TREES AND RIPAFUAN VEGETATION ASSOCIATED WITH HUMAN-MADE PASTURES IN A
FRAGMENTED TROPICAL LANDSCAPE
Biotecnologia jEcologia Aplicnda Cniuersidad Veramuzann A P 250 Xalapa Vmcrur 91001 Mixiro lJGG)
Institute de Ecologin A C A P 63 Xalapa Verarrur 91000 Mixico (155) Correspondent jgal indo~umx
~ S T R ~ C T - C O ~ ~ ~ ~ S ~ O ~of tropical rain forest to pasture and cropland often leads to a loss of local species However information on the role of some isolated elements of the fragmented landscape on maintaining biodiversity is lacking This study carried out in human-made pastures in Veracruz Mexico demonstrated the year-round presence of frugivorous bats in isolated trees inside pastures and in riparian corridors of vegetation among pastures During a 1-year study (October 1995 through September 1996) we captured 652 bats belonging to 20 species repre- senting 56 of the total species recorded for the area Higher abundance and diversity of bats were recorded for riparian sites than for isolated trees Of the bats we captured 82 were fru- givorous 15 insectivorous 2 nectarivorous-frugivorous and 2 sanguivorous Bats visited iso- lated trees regardless of tree species or phenological state Abundance decreased with distance from the nearest forest fragment We discuss how riparian corridors and isolated trees in pastures might contribute to maintaining diversih of frugivorous bats and plants in a fragmented landscape
RESUMEN-La transformation de las selvas tropicales en pastizales 7 areas de cultivo frecuente- mente conduce a la perdida de especies locales Sin embargo falta informaci6n sobre el papel de algunos elementos aislados del paisaje fragmentado en el mantenimiento de la biodiversidad Este estudio registr6 la presencia de murciilagos frugivoros durante todo el ano bajo arboles aislados y en la vegetaci6n riparia remanente en pastizales antropogknicos en Veracruz Mexico Durante un aho de estudio (octubre 1995-septiembre 1996) capturamos 652 murciilagos de 20 especies que representan el 36 de las especies registradas en el irea Los sitios en la vegetation riparia registra- ron mayor abundancia 7 diversidad que 10s sitios bajo irboles aislados El 82 del total de murcik- lagos fueron frugivoros 15 insectivoros 2 nectarivoros-frugivorosy 2 sanguivoros Los murcie- lagos lisitaron 10s irboles aislados independientemente de la especie del irbol o de su estado fen016 gico La abundancia disminuyo a mayor distancia del fragment0 forestal mis cercano Discutimos c6mo 10s corredores riparios y irboles aislados en 10s pastizales pueden contribuir a1 mantenimiento de cierta diversidad de murciklagos 7 plantas en el paisaje fragmentado
Conversion of tropical rain forests to pas- number of functional groups and in species in tures and croplands often leads to a loss of lo- tropical ecosystems (Heithaus et al 1975 cal plant and animal species because many Fleming 1988) They affect biological com-species are sparsely distributed or intolerant to munities through predation pollination and open sites (Turner 1996) Human-made pas- seed dispersal (Vizquez-Yanes et al 1975 tures with little vegetative cover have been Fleming et al 1977 Fleming and Heithaus identified as a barrier to natural dispersal of 1981 Fleming 1982 Bonaccorso and Hum- species (Guevara et al 1986 Aide and Cave- phrey 1984 Thomas et al 1988 Fleming and lier 1994) Distance between forest fragments Williams 1990 Charles-Dominique 1991 Kal- also might affect species abundance (Estrada ko 1997) A small number of published studies et al 1993 Andsen 1994 Galindo-Gonzalez have specifically dealt with patterns of bat di- 1999) Bats like many groups of plants and an- versity within fragmented landscapes (de Jong imals reach their highest diversity both in 1995 Brosset et al 1996) including artificial
580 Thp Southwestm Naturalist vol 48 no 4
gaps (02 ha) produced by selective logging (Crome and Richards 1988) strips (30 X 150 m) cleared for logging (Gorchov et al 1993) 2 trees in a pasture (Janzen et al 1976) and an abandoned pasture (Nepstad et al 1996) Specifically one study conducted in the tropi- cal region of Los Tuxtlas Mexico reported on how bats differentially used habitats within modified landscapes in the continuous forest 91 of the total number of species (ie 35 species) were caught followed by 40 to 60 in plantations shaded by rain forest trees 43 to 51 in unshaded plantations and 34 in live fences (Estrada et al 1993) Although not cap- tured some bats (including the insectivorous Pteronotus) have been observed flying across pastures above net levels (Estrada et al 1993 Galindo-Gonzilez 1999)
Based on foraging behavior food habits and seed rain attributed to bats utilizing pastures Galindo-Gonzilez (1998) suggested that frugiv- orous bats are important in the regeneration of tropical rain forest in abandoned pastures In many tropical regions 2 kinds of habitats are created by the conversion of forest to pas- ture 1) strips of riparian vegetation are often left behind along watercourses to protect against erosion and 2) isolated trees are left standing to provide shade for cattle firewood for cooking or for aesthetic reasons These iso- lated trees often serve as perches for birds and bats (the presence of the latter is inferred from nocturnal seed rain) that bring viable seeds to pastures from nearby forest remnants or ripar- ian vegetation (Guevara and Laborde 1993 Galindo-GonzBlez et al 2000) On the other hand riparian vegetation might function as corridors for frugivorous bats and as a source of propagules for tropical forest species (Bon- accorso and Humphrey 1984 Malanson 1993 Naiman et al 1993 Knopf and Samson 1994 Rautenbach et al 1996 Beier and Noss 1998) These studies strongly suggest that fi-ugivorous bats travel across pastures and visit isolated trees while foraging commuting or looking for roosts or to decrease predation risk how- ever we need more information on how bats use isolated trees and riparian vegetation in fragmented habitats and their significance for forest regeneration and maintenance of bat di- versity
Visits to isolated trees in pastures might be a function not only of their quality as roosts
food or shelter but of their degree of isolation (ie distance from the nearest patch of for- est) Frugivorous bats are known to travel long distances when foraging or commuting from their day roosts to their feeding areas (Galin- do-Gonzklez 1998) Assuming that pastures are a less suitable habitat than forest remnants both in terms of food resources and shelter it is reasonable to predict that the probabilih of recording a bat at isolated trees decreases with distance from sources of bats
The general purpose of this study was to de- scribe the frugivorous bat community captured in ground-level mist-nets set under isolated trees and in riparian vegetation (remnants of the original vegetation canopy) associated with pastures in a fragmented tropical landscape Specifically we wanted to determine species richness and relative abundance in this com- ponent of the bat community We predicted 1) more captures at trees with seeds that are bat- dispersed rather than wind-dispersed 2) that captures at isolated Ficus trees in pastures would be highest when these trees bore ripe fruits and 3) that bat abundance would be in- versely proportional to distance to the nearest forest fragment We specifically emphasized fruit bats because of their potential impor- tance in seed dispersal and forest regeneration (Galindo-Gonzalez 1999 Galindo-Gonzale~ et al 2000)
METHODS-StudyA r e ~ F i e l d w o r k was conducted i n t h e northeastern portion o f t h e Sierra d e Los Tuxtlas Veracruz Mexico (1836N 9505W) t h e northernmost limit o f rain forest i n North America (Dirzo and Miranda 1991) Mean annual tempera- ture is 27degC Total annual rainfall is ca 4725 m m with a relatively dry period between March and May ( 1 117 2 117 m m m o n t h ) and a rainy period be-tween J u n e and February (486 i- 870 m m m o n t h Guevara e t al 1992) Cndisturbed vegetation and forest fragments consisted o f tropical rain forest with a closed canopy 30 to 35 m high Plant communit ies were dominated by Astrocaryum mexicanurn (Areca-ceae) i n t h e understory Pseudolmedia oxyphyllana (Moraceae) i n t h e m e d i u m stratum and A~ctandm anzbigens (Lauraceae) i n t h e canopy (Bongers e t al 1988) Land management practices have converted 65 t o 80 o f these forests t o cattle pastures (Dirzo and Garcia 1992)
A t present t h e study area is a heterogeneous land- scape Specifically it is 1 ) dominated by human- m a d e o p e n pastures for cattle grazing and crop- lands (56 of surface area) 2 ) interspersed with
581 December 2003 Galindo-Gonrilez and Sosa-Frugivorous
small forest remnants (36) and 3) interconnected by riparian vegetation and live fences (4) (Guevara et al 1998) Many isolated trees occur in pastures (04 to 119 treesha Guevara et al 1998) Trans- formation of the area into pastureland began in the 1960s and since that time cattle have been grazing on a year-round basis
The st~ldy site was located within a clear-cut basin (the average distance between forested ridges was 25 km) with forest fragments distributed along the surrounding ridge Plant communities within pas- tures were dominated by early successional species from the families Piperaceae Solanaceae and Me- lastomataceae and by isolated trees from the fami- lies h4oraceae and Cecropiaceae whose fruits are known to be eaten by bats (Galindo-GonzPlez 1999 Galindo-GonzPlez et al 2000) Riparian vegetation consisted o f remnant trees and shrubs of 112 species (these represented 71 of the species from the orig- inal forest) located on both sides of a stream whose banks were 2 to 5 m wide Riparian remnant trees were taller than 18 m Their closed canopy over the stream formed a tunnel-like flight corridor for birds and bats (for more detail on floristic composition and structure of vegetation in pastures see Guevara et al 1992)
Samplzng Strategy-We selected 8 nettlng sites rep- resenting 3 habitat types or elements of the frag- mented landscape to sample for bats and measure fruit availability 1) isolated trees producing bat-dis- persed seeds 2) isolated trees producing wind-dis- persed seeds (control) and 3) riparian vegetation Netting sites were placed under the canop of each of the following isolated trees (all trees 25 to 30 in h ~ g h )Izcus ~oponenszsFtcus coluhnae Cezba pentan dra and Cedrella odorata Vertebrates disperse the seeds of the 2 former species and the latter 2 are wind-dispersed Ceiba pentandra is a bat-pollinated tree flowering between December and March how- ever we did not record flowering of this tree during the study period Fzcus was considered particularly important because it is the most common tree ge- nus encountered among isolated trees in pastures (Guevara et al 1986) Isolated trees were 250 m from any forested fragment or riparian vegetation The remaining 4 sites were randomly selected along the main vegetated stream in the study area Netting sites varied in distance from a forest fragment Ri-parian sites were the nearest to forest (95 to 276 m) and isolated trees were the most distant (280 to 381 m) The distance from each isolated tree to the near- est forest fragment (a forested area gt1 ha) was nlea- sured on an aerial photograph (1991) scale l 75000 measurements of distance between 5 refer- ence points in situ were used to calibrate measure- ments Each month ~e recorded presence-absence of ripe ficus fruits for later correlation with bat abundance We then calculated the percentage of
bats in fragmented tropical landscape
the crown that was covered with fruits and estimated the proportion of ripe and unripe fruits (see meth- ods in Chapman et al 1992)
Bat CapturesBats were captured from October 1995 through September 1996 using mist-nets placed at ground level Mist-netting at ground level is known to be biased for fruit-eating bats and against high-flying insectivorous bats bats with low body mass and some species that tend to be cap- tured in the canopy stratum of tropical forests (Rau- tenbach eta] 1996 Kalko 1997 Bernard 2001 Kal- ko and Handley 2001) Two mist-nets (each 12 X
25 m) were placed in an L-shaped array under the canopy of isolated trees At each riparian site we used 1 net (6 X 25 nl) placed across the stream because each stream was lt5 m wide and because preliminary sampling showed that bat captures were more frequent in this habitat Netting sites were sam- pled once each month around the time of the new moon ( 2 3 days) to avoid lunar phobia (hlorrison 1978b) An a-erage of 41 hnight (SE 005) and 33 nightsmonth (SE + 02) were dedicated to sam- pling Nets were opened 30 min after sunset Total sampling effort in this study was 261320 net-m X
hours derived from 235 h of netting 1112 m of net during a total of 40 netting nights All captured bats were identified to species using field keys (hledellin et a] 1997 see Ramirez-Pulido et al 1996 for no- menclature) and fitted with unique color-coded plastic necklaces (Dennison Secur-A-Tie Framing- ham Massachusetts USA) Sex mass (to nearest 01 g) reproductive condition and time and site of cap- ture were recorded
Data AnalysisBat community structure was deter- mined according to family species and foraging- guild as well as by relative species abundance Spe- cies richness in each habitat type was compared through accumulation species f~mctions which pre- dict the maximum number of species (the asymp- tote) present in a certain area based on the cumu- lative number of species (Sober611 and Ilorente 1993) Several functions can be derived depending on the assumptions made about the probability of adding a new species after a collecting period UTe chose the exponential function [St = ab(l - bt) where S is species richness at time t and a h param-eters] which assumes that the probability of adding new species decreases linearly with the size of the list This function might be adequate when sampling a relatively small area or a well kuuwn group or both (our case) and eventually all species will be recorded It also allows one to estimate confidence intervals for S(t) which is the number of species collected at time tk (the time needed to lower the increase rate of the accumulated species list in k pro-portion Sober6n and Llorente 1993) We fitted the function by non-linear regression using the quasi-
582 Thr Southwestern iVaturaltst vol 48 no 4
Newton algorithm in the package STATISTICA (Statsoft 1991)
Due to our use of mist-nets of different sizes we calculated adjusted relative abundance (number of individuals per 1000 net-m X hour) by sampling ef- fort with the following formula adjusted relative abundance = 1000 [b (nh)] where b = total Ilum- ber of captured bats and nh = sampling effort as already defined Bat species diversity for each habitat type (ie riparian vegetation isolated Fzcus trees and isolated control trees) was estimated with the Shannon-Uiener index (H) using the total annual captures Diversity indices among habitat ty-pes were then compared using a I-way ANOVA test applied to ranks The Morisita-Horn index (CmH) was used to compare species similarity among habitats (Magur- ran 1988)
We conducted ANOVA and Tukey pairwise mul- tiple comparisons to check whether total relative abundance (ie total bat species captured during the year) differed among habitat types We also used ANOVA to determine whether the relative abun- dance of frugivorous bats differed between fruiting Ficus trees and non-fruiting Ceibu and Cedrella trees (ie controls) in the study area For this analysis we used total capture of frugivorous bat5 from April through August 1996 months when Ficus trees bore fruits Relative abundance was normalized b a square root transformation before conducting these analyses Linear regression analysis was conducted to determine the association between relative abun- dance of all bats and the percentage of canopy with ripe fruits at isolated Ficus trees in pastures Finally simple linear correlation analysis was conducted to determine the degree of association behveen dis- tance to the nearest forest fragment and bat abun- dance in riparian vegetation and around isolated trees in pastures
RESULTS-Species Richness and Community Structurewe captured 652 bats from 20 spe- cies in 4 families (Table 1) Most species were from the family Phyllostomidae (15 species) The families Mormoopidae Vespertilionidae and Molossidae were represented by 5 species (Table 1) As expected frugivorous bats were the most abundant (81 of the total captures) followed by insectivores (15) nectarivore- frugivores (2) and sanguivores (2) Total relative abundance varied greatly with species Five species accounted for 84 of the cap- tures These were the frugivores Sturnira liliurn (40) Artibeus jamaicensis (15) Carollia per-spinllata ( lo) and Dermanura tolteca (9) and the insectivore Pteronotus parnellii (10)
The cumulative number of species over all
habitat types tended to level off after 20 nights of netting The species accumulation function predicted a maximum of 23 species According to Moreno and Halffter (1999) 19 nights at least are enough to record 90 of frugivorous bats in a heterogeneous habitat Therefore our sampling provided a reasonably good es- timation of species richness at ground level The maximum numbers of bat species for each habitat predicted by the accumulation func- tion were 17 13 and 12 for the riparian the Acus trees and control trees respectively (Fig l ) and we captured 18 13 and 12 species respectively
Bat diversity varied among habitat types Abundance species richness and Shannon-Wiener index (H)were higher in riparian veg- etation than under isolated trees in pastures despite the lower sampling effort in the ripar- ian habitat (Table 2) Some species were pre- sent only in riparian vegetation whereas others used both habitats (Table 1) The adjusted rel- ative abundance of all bats differed among habitat types (ANOVA F = 2184 df = 2 P lt 0001 Fig 2) According to a posteriori multi- ple comparison tests relative abundance was significantly higher in the riparian habitat than at isolated Ficus trees (q = 833 P lt 005) or control trees (q = 784 P lt 005) relative abundance was not significantly different be- tween isolated Ficus trees and control trees ( q = 048 P gt 003) The Shannon-Wiener index was marginally different between the riparian and control trees (F = 581 P = 006 Table 2) Species similarity was highest between iso- lated trees (ie between Ficus and corltrol trees) and lowest between isolated trees and riparian habitats (Table 2) Bats also were cap- tured in different habitat types according to their feeding habits In particular insectivo- rous bats were more abundant in riparian sites (70) than in isolated tree sites (30) in con- trast frugivores were more abundant at isolat- ed trees (75) than at riparian sites (25)
Use of Isolated Trees and fiparinn Vegetation Habitats-Frugivorous bats were caught under the canopies of isolated trees irrespective of the seed dispersal mechanism (bat or wind-dis-persed) or phenological state of the tree Rela- tive abundance for frugivorous bats at control trees was slightly higher than at ficus trees dur- ing most of the year However no significant differences were found among mean relative
589 1)ecember 2003 Oalindo-Gonrilez and Sosa-Frugivorous bats in fragmented tropiral landscape
TABLE1-Bat species recorded in a fragmented landscape in Los Tuxtlas lkracruz Mexico October 1995 through September 1996 Habitat type I = under isolated tree R = riparian (number of individuals cap- tured)
Relative Habitat type
abundance n ( ) I K Feeding guild
Mormoopidae
Pteronotus dauyi 8 12 1 7 insectivore Pteronotus parnelliz 64 98 19 43 insectibore Mor~nocr megalophylla 16 25 2 14 insectivore
Phvllostomldae
Phyllostorninae Phgtllostomus dzscolor 1 02 0 1 frugivore
Carolliinae Carollza hreurcauda 1 02 0 1 frugivore Carollza perspzczllata 65 99 28 37 frugivore
Glossophaginae Glossophaga soricina 10 15 s 3 nectar-frugivore Choeronzscus godmani 1 02 0 I nectar-frugivore
Stenoderrnatlnae Sturnmi izlzum frugirore Di-oderma bzlobatuni frugivore Platyrrhznus hellm frugivore Vamnpyrodes caracczolz frugivore Chzrod~rma uzllosum frughore Artzbeu lamazcenzi frugivore Artzbvus lzturatus frugivore Dermanura toltrca frugivore Dernlanura pharotzs frugivorc
Desmodontinae Desmodus rotundus 10 15 4 6 sanguivore
Vcspertilionidae
My otis keaysz 11 17 0 11 insectivore
Molossidae
iMolossuc rufus I 02 1 0 insectivore
Total 652 391 261
abundance values for frugivorous bats at control parian sites and bats moved from 1 habitat vs Ficus trees during their fruiting seasons (AN- type to another (ie among riparian sites iso- OVA F = 035 P = 057 df = 1)Finally no lated Ficus trees and control trees) Most re- significant relationship was found between bat captures (20 individuals) were caught on sep- abundance and percentage of ripe Ficus fruits arate nights and at different sites and habitats in isolated trees ( r = 027 P = 066 n = 5) from where they were first captured Distance
We recaptured 22 individuals belonging to 3 among recapture sites ranged from 117 to 692 frugivorous species a total of 34 times (Table m (mean 2805 n~ 234 m SE) Recapture 3) Carollia perspiczllata was the most recaptured times range from a few hours in the same night species even though Sturnira lilium was more to 8 months after the first capture Recaptured abundant indicating that the former shows bats did not seem to have any harm caused by more fidelity to the studied pastures than the the necklaces latter We recorded more recaptures at the ri- Relative abundance of all bats decreased
584 The Southwestern Vuturalzst vol 48 no 4
20 1 A A A
0 Ficus
+ Control
+ R~par~an
A Total 1995 Months 1996
0 v 0 5 10 15 20 25 30 35 40 45
-0 -Riparian -v-Ficus trees -0-Control trees
Accumulated netting nightslmonth FIG2-Mean adjusted relative abundance of bats
FIG l-Accumulation curves (lines) for bat spe- in different habitat t7rpes of
ties adjusted by exponential function for each hab- landscape a -year at Tuxtlas Ve-itat and for total captures in pastures at Los Tuxtlas racruZ
Veracruz Mexico
with increasing distance from the nearest for-est edge (Fig 3 ) To eliminate potential bias produced by the presence of water (which could function as an attractor to bats) at ripar-ian sites we analyzed separately 1) the corre-lation between bat relative abundance and dis-tance to isolated trees ( r = -072 P gt 005) and 2) the relationship between bat relative abundance and distance to riparian sites ( r =
-095 P = 005) Most bats were captured in the riparian habitat located nearest to a forest fragment The fewest number of bats were cap-tured at isolated Ficus trees located the greatest distance from the forest edge
D~scus s~o~-Ourresults show that bats (mostly frugivorous species) flew around iso-
lated trees and in strips of riparian vegetation in pastures within a fragmented tropical land-scape while foraging commuting or looking for perches Bat abundance richness and di-versity were higher in riparian corridors than under the canopy of isolated trees Frugivorous bats were caught under the canopies of differ-ent isolated trees throughout the year irre-spective of the seed dispersal mechanism of the tree or the presence or absence of fruits As predicted bat abundance decreased with distance to the nearest forest fragment
In both the riparian and isolated tree sites frugivorous species from the family Phyllo-stomidae were most common Species in the subfamily Stenodermatinae were more numer-ous (9 out of 12 potentially present) than the subfamily Phyllostominae (1 out of 7 poten-
TABLE2-Bat species richness sampling effort (SE h = netting hours m = total meters of installed nets) adjusted relative abundance (ARA see text for units) mean Shannon-Wiener diversity index (IF) and pairwise comparisons of species similarity among habitats (Morisita-Horn index CmH) on different habitat types sampled in pastures in Los Tuxtlas Veracruz Mexico
BatsSpecies expected
Species cap- SE Mean Habitat type n observed St CI tured (h X m) ARA IF CmH2
- - ---
Fzcus (a) 2 13 113 88-137 150 36324 413 0776 097 (a-b) Control trees (b) 2 12 112 99-125 241 48654 495 0670 083 (b-c) Riparian (c) 3 18 156 141-172 261 8096 3224 0824 090 (c-a)
Total 652
At time tk when an increase of 5 (k = 005) in the species list is predicted according to the exponential species accumulation function Lower and upper 93 confidence limits shown
CmH = 1 (100 similarity)
December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

580 Thp Southwestm Naturalist vol 48 no 4
gaps (02 ha) produced by selective logging (Crome and Richards 1988) strips (30 X 150 m) cleared for logging (Gorchov et al 1993) 2 trees in a pasture (Janzen et al 1976) and an abandoned pasture (Nepstad et al 1996) Specifically one study conducted in the tropi- cal region of Los Tuxtlas Mexico reported on how bats differentially used habitats within modified landscapes in the continuous forest 91 of the total number of species (ie 35 species) were caught followed by 40 to 60 in plantations shaded by rain forest trees 43 to 51 in unshaded plantations and 34 in live fences (Estrada et al 1993) Although not cap- tured some bats (including the insectivorous Pteronotus) have been observed flying across pastures above net levels (Estrada et al 1993 Galindo-Gonzilez 1999)
Based on foraging behavior food habits and seed rain attributed to bats utilizing pastures Galindo-Gonzilez (1998) suggested that frugiv- orous bats are important in the regeneration of tropical rain forest in abandoned pastures In many tropical regions 2 kinds of habitats are created by the conversion of forest to pas- ture 1) strips of riparian vegetation are often left behind along watercourses to protect against erosion and 2) isolated trees are left standing to provide shade for cattle firewood for cooking or for aesthetic reasons These iso- lated trees often serve as perches for birds and bats (the presence of the latter is inferred from nocturnal seed rain) that bring viable seeds to pastures from nearby forest remnants or ripar- ian vegetation (Guevara and Laborde 1993 Galindo-GonzBlez et al 2000) On the other hand riparian vegetation might function as corridors for frugivorous bats and as a source of propagules for tropical forest species (Bon- accorso and Humphrey 1984 Malanson 1993 Naiman et al 1993 Knopf and Samson 1994 Rautenbach et al 1996 Beier and Noss 1998) These studies strongly suggest that fi-ugivorous bats travel across pastures and visit isolated trees while foraging commuting or looking for roosts or to decrease predation risk how- ever we need more information on how bats use isolated trees and riparian vegetation in fragmented habitats and their significance for forest regeneration and maintenance of bat di- versity
Visits to isolated trees in pastures might be a function not only of their quality as roosts
food or shelter but of their degree of isolation (ie distance from the nearest patch of for- est) Frugivorous bats are known to travel long distances when foraging or commuting from their day roosts to their feeding areas (Galin- do-Gonzklez 1998) Assuming that pastures are a less suitable habitat than forest remnants both in terms of food resources and shelter it is reasonable to predict that the probabilih of recording a bat at isolated trees decreases with distance from sources of bats
The general purpose of this study was to de- scribe the frugivorous bat community captured in ground-level mist-nets set under isolated trees and in riparian vegetation (remnants of the original vegetation canopy) associated with pastures in a fragmented tropical landscape Specifically we wanted to determine species richness and relative abundance in this com- ponent of the bat community We predicted 1) more captures at trees with seeds that are bat- dispersed rather than wind-dispersed 2) that captures at isolated Ficus trees in pastures would be highest when these trees bore ripe fruits and 3) that bat abundance would be in- versely proportional to distance to the nearest forest fragment We specifically emphasized fruit bats because of their potential impor- tance in seed dispersal and forest regeneration (Galindo-Gonzalez 1999 Galindo-Gonzale~ et al 2000)
METHODS-StudyA r e ~ F i e l d w o r k was conducted i n t h e northeastern portion o f t h e Sierra d e Los Tuxtlas Veracruz Mexico (1836N 9505W) t h e northernmost limit o f rain forest i n North America (Dirzo and Miranda 1991) Mean annual tempera- ture is 27degC Total annual rainfall is ca 4725 m m with a relatively dry period between March and May ( 1 117 2 117 m m m o n t h ) and a rainy period be-tween J u n e and February (486 i- 870 m m m o n t h Guevara e t al 1992) Cndisturbed vegetation and forest fragments consisted o f tropical rain forest with a closed canopy 30 to 35 m high Plant communit ies were dominated by Astrocaryum mexicanurn (Areca-ceae) i n t h e understory Pseudolmedia oxyphyllana (Moraceae) i n t h e m e d i u m stratum and A~ctandm anzbigens (Lauraceae) i n t h e canopy (Bongers e t al 1988) Land management practices have converted 65 t o 80 o f these forests t o cattle pastures (Dirzo and Garcia 1992)
A t present t h e study area is a heterogeneous land- scape Specifically it is 1 ) dominated by human- m a d e o p e n pastures for cattle grazing and crop- lands (56 of surface area) 2 ) interspersed with
581 December 2003 Galindo-Gonrilez and Sosa-Frugivorous
small forest remnants (36) and 3) interconnected by riparian vegetation and live fences (4) (Guevara et al 1998) Many isolated trees occur in pastures (04 to 119 treesha Guevara et al 1998) Trans- formation of the area into pastureland began in the 1960s and since that time cattle have been grazing on a year-round basis
The st~ldy site was located within a clear-cut basin (the average distance between forested ridges was 25 km) with forest fragments distributed along the surrounding ridge Plant communities within pas- tures were dominated by early successional species from the families Piperaceae Solanaceae and Me- lastomataceae and by isolated trees from the fami- lies h4oraceae and Cecropiaceae whose fruits are known to be eaten by bats (Galindo-GonzPlez 1999 Galindo-GonzPlez et al 2000) Riparian vegetation consisted o f remnant trees and shrubs of 112 species (these represented 71 of the species from the orig- inal forest) located on both sides of a stream whose banks were 2 to 5 m wide Riparian remnant trees were taller than 18 m Their closed canopy over the stream formed a tunnel-like flight corridor for birds and bats (for more detail on floristic composition and structure of vegetation in pastures see Guevara et al 1992)
Samplzng Strategy-We selected 8 nettlng sites rep- resenting 3 habitat types or elements of the frag- mented landscape to sample for bats and measure fruit availability 1) isolated trees producing bat-dis- persed seeds 2) isolated trees producing wind-dis- persed seeds (control) and 3) riparian vegetation Netting sites were placed under the canop of each of the following isolated trees (all trees 25 to 30 in h ~ g h )Izcus ~oponenszsFtcus coluhnae Cezba pentan dra and Cedrella odorata Vertebrates disperse the seeds of the 2 former species and the latter 2 are wind-dispersed Ceiba pentandra is a bat-pollinated tree flowering between December and March how- ever we did not record flowering of this tree during the study period Fzcus was considered particularly important because it is the most common tree ge- nus encountered among isolated trees in pastures (Guevara et al 1986) Isolated trees were 250 m from any forested fragment or riparian vegetation The remaining 4 sites were randomly selected along the main vegetated stream in the study area Netting sites varied in distance from a forest fragment Ri-parian sites were the nearest to forest (95 to 276 m) and isolated trees were the most distant (280 to 381 m) The distance from each isolated tree to the near- est forest fragment (a forested area gt1 ha) was nlea- sured on an aerial photograph (1991) scale l 75000 measurements of distance between 5 refer- ence points in situ were used to calibrate measure- ments Each month ~e recorded presence-absence of ripe ficus fruits for later correlation with bat abundance We then calculated the percentage of
bats in fragmented tropical landscape
the crown that was covered with fruits and estimated the proportion of ripe and unripe fruits (see meth- ods in Chapman et al 1992)
Bat CapturesBats were captured from October 1995 through September 1996 using mist-nets placed at ground level Mist-netting at ground level is known to be biased for fruit-eating bats and against high-flying insectivorous bats bats with low body mass and some species that tend to be cap- tured in the canopy stratum of tropical forests (Rau- tenbach eta] 1996 Kalko 1997 Bernard 2001 Kal- ko and Handley 2001) Two mist-nets (each 12 X
25 m) were placed in an L-shaped array under the canopy of isolated trees At each riparian site we used 1 net (6 X 25 nl) placed across the stream because each stream was lt5 m wide and because preliminary sampling showed that bat captures were more frequent in this habitat Netting sites were sam- pled once each month around the time of the new moon ( 2 3 days) to avoid lunar phobia (hlorrison 1978b) An a-erage of 41 hnight (SE 005) and 33 nightsmonth (SE + 02) were dedicated to sam- pling Nets were opened 30 min after sunset Total sampling effort in this study was 261320 net-m X
hours derived from 235 h of netting 1112 m of net during a total of 40 netting nights All captured bats were identified to species using field keys (hledellin et a] 1997 see Ramirez-Pulido et al 1996 for no- menclature) and fitted with unique color-coded plastic necklaces (Dennison Secur-A-Tie Framing- ham Massachusetts USA) Sex mass (to nearest 01 g) reproductive condition and time and site of cap- ture were recorded
Data AnalysisBat community structure was deter- mined according to family species and foraging- guild as well as by relative species abundance Spe- cies richness in each habitat type was compared through accumulation species f~mctions which pre- dict the maximum number of species (the asymp- tote) present in a certain area based on the cumu- lative number of species (Sober611 and Ilorente 1993) Several functions can be derived depending on the assumptions made about the probability of adding a new species after a collecting period UTe chose the exponential function [St = ab(l - bt) where S is species richness at time t and a h param-eters] which assumes that the probability of adding new species decreases linearly with the size of the list This function might be adequate when sampling a relatively small area or a well kuuwn group or both (our case) and eventually all species will be recorded It also allows one to estimate confidence intervals for S(t) which is the number of species collected at time tk (the time needed to lower the increase rate of the accumulated species list in k pro-portion Sober6n and Llorente 1993) We fitted the function by non-linear regression using the quasi-
582 Thr Southwestern iVaturaltst vol 48 no 4
Newton algorithm in the package STATISTICA (Statsoft 1991)
Due to our use of mist-nets of different sizes we calculated adjusted relative abundance (number of individuals per 1000 net-m X hour) by sampling ef- fort with the following formula adjusted relative abundance = 1000 [b (nh)] where b = total Ilum- ber of captured bats and nh = sampling effort as already defined Bat species diversity for each habitat type (ie riparian vegetation isolated Fzcus trees and isolated control trees) was estimated with the Shannon-Uiener index (H) using the total annual captures Diversity indices among habitat ty-pes were then compared using a I-way ANOVA test applied to ranks The Morisita-Horn index (CmH) was used to compare species similarity among habitats (Magur- ran 1988)
We conducted ANOVA and Tukey pairwise mul- tiple comparisons to check whether total relative abundance (ie total bat species captured during the year) differed among habitat types We also used ANOVA to determine whether the relative abun- dance of frugivorous bats differed between fruiting Ficus trees and non-fruiting Ceibu and Cedrella trees (ie controls) in the study area For this analysis we used total capture of frugivorous bat5 from April through August 1996 months when Ficus trees bore fruits Relative abundance was normalized b a square root transformation before conducting these analyses Linear regression analysis was conducted to determine the association between relative abun- dance of all bats and the percentage of canopy with ripe fruits at isolated Ficus trees in pastures Finally simple linear correlation analysis was conducted to determine the degree of association behveen dis- tance to the nearest forest fragment and bat abun- dance in riparian vegetation and around isolated trees in pastures
RESULTS-Species Richness and Community Structurewe captured 652 bats from 20 spe- cies in 4 families (Table 1) Most species were from the family Phyllostomidae (15 species) The families Mormoopidae Vespertilionidae and Molossidae were represented by 5 species (Table 1) As expected frugivorous bats were the most abundant (81 of the total captures) followed by insectivores (15) nectarivore- frugivores (2) and sanguivores (2) Total relative abundance varied greatly with species Five species accounted for 84 of the cap- tures These were the frugivores Sturnira liliurn (40) Artibeus jamaicensis (15) Carollia per-spinllata ( lo) and Dermanura tolteca (9) and the insectivore Pteronotus parnellii (10)
The cumulative number of species over all
habitat types tended to level off after 20 nights of netting The species accumulation function predicted a maximum of 23 species According to Moreno and Halffter (1999) 19 nights at least are enough to record 90 of frugivorous bats in a heterogeneous habitat Therefore our sampling provided a reasonably good es- timation of species richness at ground level The maximum numbers of bat species for each habitat predicted by the accumulation func- tion were 17 13 and 12 for the riparian the Acus trees and control trees respectively (Fig l ) and we captured 18 13 and 12 species respectively
Bat diversity varied among habitat types Abundance species richness and Shannon-Wiener index (H)were higher in riparian veg- etation than under isolated trees in pastures despite the lower sampling effort in the ripar- ian habitat (Table 2) Some species were pre- sent only in riparian vegetation whereas others used both habitats (Table 1) The adjusted rel- ative abundance of all bats differed among habitat types (ANOVA F = 2184 df = 2 P lt 0001 Fig 2) According to a posteriori multi- ple comparison tests relative abundance was significantly higher in the riparian habitat than at isolated Ficus trees (q = 833 P lt 005) or control trees (q = 784 P lt 005) relative abundance was not significantly different be- tween isolated Ficus trees and control trees ( q = 048 P gt 003) The Shannon-Wiener index was marginally different between the riparian and control trees (F = 581 P = 006 Table 2) Species similarity was highest between iso- lated trees (ie between Ficus and corltrol trees) and lowest between isolated trees and riparian habitats (Table 2) Bats also were cap- tured in different habitat types according to their feeding habits In particular insectivo- rous bats were more abundant in riparian sites (70) than in isolated tree sites (30) in con- trast frugivores were more abundant at isolat- ed trees (75) than at riparian sites (25)
Use of Isolated Trees and fiparinn Vegetation Habitats-Frugivorous bats were caught under the canopies of isolated trees irrespective of the seed dispersal mechanism (bat or wind-dis-persed) or phenological state of the tree Rela- tive abundance for frugivorous bats at control trees was slightly higher than at ficus trees dur- ing most of the year However no significant differences were found among mean relative
589 1)ecember 2003 Oalindo-Gonrilez and Sosa-Frugivorous bats in fragmented tropiral landscape
TABLE1-Bat species recorded in a fragmented landscape in Los Tuxtlas lkracruz Mexico October 1995 through September 1996 Habitat type I = under isolated tree R = riparian (number of individuals cap- tured)
Relative Habitat type
abundance n ( ) I K Feeding guild
Mormoopidae
Pteronotus dauyi 8 12 1 7 insectivore Pteronotus parnelliz 64 98 19 43 insectibore Mor~nocr megalophylla 16 25 2 14 insectivore
Phvllostomldae
Phyllostorninae Phgtllostomus dzscolor 1 02 0 1 frugivore
Carolliinae Carollza hreurcauda 1 02 0 1 frugivore Carollza perspzczllata 65 99 28 37 frugivore
Glossophaginae Glossophaga soricina 10 15 s 3 nectar-frugivore Choeronzscus godmani 1 02 0 I nectar-frugivore
Stenoderrnatlnae Sturnmi izlzum frugirore Di-oderma bzlobatuni frugivore Platyrrhznus hellm frugivore Vamnpyrodes caracczolz frugivore Chzrod~rma uzllosum frughore Artzbeu lamazcenzi frugivore Artzbvus lzturatus frugivore Dermanura toltrca frugivore Dernlanura pharotzs frugivorc
Desmodontinae Desmodus rotundus 10 15 4 6 sanguivore
Vcspertilionidae
My otis keaysz 11 17 0 11 insectivore
Molossidae
iMolossuc rufus I 02 1 0 insectivore
Total 652 391 261
abundance values for frugivorous bats at control parian sites and bats moved from 1 habitat vs Ficus trees during their fruiting seasons (AN- type to another (ie among riparian sites iso- OVA F = 035 P = 057 df = 1)Finally no lated Ficus trees and control trees) Most re- significant relationship was found between bat captures (20 individuals) were caught on sep- abundance and percentage of ripe Ficus fruits arate nights and at different sites and habitats in isolated trees ( r = 027 P = 066 n = 5) from where they were first captured Distance
We recaptured 22 individuals belonging to 3 among recapture sites ranged from 117 to 692 frugivorous species a total of 34 times (Table m (mean 2805 n~ 234 m SE) Recapture 3) Carollia perspiczllata was the most recaptured times range from a few hours in the same night species even though Sturnira lilium was more to 8 months after the first capture Recaptured abundant indicating that the former shows bats did not seem to have any harm caused by more fidelity to the studied pastures than the the necklaces latter We recorded more recaptures at the ri- Relative abundance of all bats decreased
584 The Southwestern Vuturalzst vol 48 no 4
20 1 A A A
0 Ficus
+ Control
+ R~par~an
A Total 1995 Months 1996
0 v 0 5 10 15 20 25 30 35 40 45
-0 -Riparian -v-Ficus trees -0-Control trees
Accumulated netting nightslmonth FIG2-Mean adjusted relative abundance of bats
FIG l-Accumulation curves (lines) for bat spe- in different habitat t7rpes of
ties adjusted by exponential function for each hab- landscape a -year at Tuxtlas Ve-itat and for total captures in pastures at Los Tuxtlas racruZ
Veracruz Mexico
with increasing distance from the nearest for-est edge (Fig 3 ) To eliminate potential bias produced by the presence of water (which could function as an attractor to bats) at ripar-ian sites we analyzed separately 1) the corre-lation between bat relative abundance and dis-tance to isolated trees ( r = -072 P gt 005) and 2) the relationship between bat relative abundance and distance to riparian sites ( r =
-095 P = 005) Most bats were captured in the riparian habitat located nearest to a forest fragment The fewest number of bats were cap-tured at isolated Ficus trees located the greatest distance from the forest edge
D~scus s~o~-Ourresults show that bats (mostly frugivorous species) flew around iso-
lated trees and in strips of riparian vegetation in pastures within a fragmented tropical land-scape while foraging commuting or looking for perches Bat abundance richness and di-versity were higher in riparian corridors than under the canopy of isolated trees Frugivorous bats were caught under the canopies of differ-ent isolated trees throughout the year irre-spective of the seed dispersal mechanism of the tree or the presence or absence of fruits As predicted bat abundance decreased with distance to the nearest forest fragment
In both the riparian and isolated tree sites frugivorous species from the family Phyllo-stomidae were most common Species in the subfamily Stenodermatinae were more numer-ous (9 out of 12 potentially present) than the subfamily Phyllostominae (1 out of 7 poten-
TABLE2-Bat species richness sampling effort (SE h = netting hours m = total meters of installed nets) adjusted relative abundance (ARA see text for units) mean Shannon-Wiener diversity index (IF) and pairwise comparisons of species similarity among habitats (Morisita-Horn index CmH) on different habitat types sampled in pastures in Los Tuxtlas Veracruz Mexico
BatsSpecies expected
Species cap- SE Mean Habitat type n observed St CI tured (h X m) ARA IF CmH2
- - ---
Fzcus (a) 2 13 113 88-137 150 36324 413 0776 097 (a-b) Control trees (b) 2 12 112 99-125 241 48654 495 0670 083 (b-c) Riparian (c) 3 18 156 141-172 261 8096 3224 0824 090 (c-a)
Total 652
At time tk when an increase of 5 (k = 005) in the species list is predicted according to the exponential species accumulation function Lower and upper 93 confidence limits shown
CmH = 1 (100 similarity)
December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

581 December 2003 Galindo-Gonrilez and Sosa-Frugivorous
small forest remnants (36) and 3) interconnected by riparian vegetation and live fences (4) (Guevara et al 1998) Many isolated trees occur in pastures (04 to 119 treesha Guevara et al 1998) Trans- formation of the area into pastureland began in the 1960s and since that time cattle have been grazing on a year-round basis
The st~ldy site was located within a clear-cut basin (the average distance between forested ridges was 25 km) with forest fragments distributed along the surrounding ridge Plant communities within pas- tures were dominated by early successional species from the families Piperaceae Solanaceae and Me- lastomataceae and by isolated trees from the fami- lies h4oraceae and Cecropiaceae whose fruits are known to be eaten by bats (Galindo-GonzPlez 1999 Galindo-GonzPlez et al 2000) Riparian vegetation consisted o f remnant trees and shrubs of 112 species (these represented 71 of the species from the orig- inal forest) located on both sides of a stream whose banks were 2 to 5 m wide Riparian remnant trees were taller than 18 m Their closed canopy over the stream formed a tunnel-like flight corridor for birds and bats (for more detail on floristic composition and structure of vegetation in pastures see Guevara et al 1992)
Samplzng Strategy-We selected 8 nettlng sites rep- resenting 3 habitat types or elements of the frag- mented landscape to sample for bats and measure fruit availability 1) isolated trees producing bat-dis- persed seeds 2) isolated trees producing wind-dis- persed seeds (control) and 3) riparian vegetation Netting sites were placed under the canop of each of the following isolated trees (all trees 25 to 30 in h ~ g h )Izcus ~oponenszsFtcus coluhnae Cezba pentan dra and Cedrella odorata Vertebrates disperse the seeds of the 2 former species and the latter 2 are wind-dispersed Ceiba pentandra is a bat-pollinated tree flowering between December and March how- ever we did not record flowering of this tree during the study period Fzcus was considered particularly important because it is the most common tree ge- nus encountered among isolated trees in pastures (Guevara et al 1986) Isolated trees were 250 m from any forested fragment or riparian vegetation The remaining 4 sites were randomly selected along the main vegetated stream in the study area Netting sites varied in distance from a forest fragment Ri-parian sites were the nearest to forest (95 to 276 m) and isolated trees were the most distant (280 to 381 m) The distance from each isolated tree to the near- est forest fragment (a forested area gt1 ha) was nlea- sured on an aerial photograph (1991) scale l 75000 measurements of distance between 5 refer- ence points in situ were used to calibrate measure- ments Each month ~e recorded presence-absence of ripe ficus fruits for later correlation with bat abundance We then calculated the percentage of
bats in fragmented tropical landscape
the crown that was covered with fruits and estimated the proportion of ripe and unripe fruits (see meth- ods in Chapman et al 1992)
Bat CapturesBats were captured from October 1995 through September 1996 using mist-nets placed at ground level Mist-netting at ground level is known to be biased for fruit-eating bats and against high-flying insectivorous bats bats with low body mass and some species that tend to be cap- tured in the canopy stratum of tropical forests (Rau- tenbach eta] 1996 Kalko 1997 Bernard 2001 Kal- ko and Handley 2001) Two mist-nets (each 12 X
25 m) were placed in an L-shaped array under the canopy of isolated trees At each riparian site we used 1 net (6 X 25 nl) placed across the stream because each stream was lt5 m wide and because preliminary sampling showed that bat captures were more frequent in this habitat Netting sites were sam- pled once each month around the time of the new moon ( 2 3 days) to avoid lunar phobia (hlorrison 1978b) An a-erage of 41 hnight (SE 005) and 33 nightsmonth (SE + 02) were dedicated to sam- pling Nets were opened 30 min after sunset Total sampling effort in this study was 261320 net-m X
hours derived from 235 h of netting 1112 m of net during a total of 40 netting nights All captured bats were identified to species using field keys (hledellin et a] 1997 see Ramirez-Pulido et al 1996 for no- menclature) and fitted with unique color-coded plastic necklaces (Dennison Secur-A-Tie Framing- ham Massachusetts USA) Sex mass (to nearest 01 g) reproductive condition and time and site of cap- ture were recorded
Data AnalysisBat community structure was deter- mined according to family species and foraging- guild as well as by relative species abundance Spe- cies richness in each habitat type was compared through accumulation species f~mctions which pre- dict the maximum number of species (the asymp- tote) present in a certain area based on the cumu- lative number of species (Sober611 and Ilorente 1993) Several functions can be derived depending on the assumptions made about the probability of adding a new species after a collecting period UTe chose the exponential function [St = ab(l - bt) where S is species richness at time t and a h param-eters] which assumes that the probability of adding new species decreases linearly with the size of the list This function might be adequate when sampling a relatively small area or a well kuuwn group or both (our case) and eventually all species will be recorded It also allows one to estimate confidence intervals for S(t) which is the number of species collected at time tk (the time needed to lower the increase rate of the accumulated species list in k pro-portion Sober6n and Llorente 1993) We fitted the function by non-linear regression using the quasi-
582 Thr Southwestern iVaturaltst vol 48 no 4
Newton algorithm in the package STATISTICA (Statsoft 1991)
Due to our use of mist-nets of different sizes we calculated adjusted relative abundance (number of individuals per 1000 net-m X hour) by sampling ef- fort with the following formula adjusted relative abundance = 1000 [b (nh)] where b = total Ilum- ber of captured bats and nh = sampling effort as already defined Bat species diversity for each habitat type (ie riparian vegetation isolated Fzcus trees and isolated control trees) was estimated with the Shannon-Uiener index (H) using the total annual captures Diversity indices among habitat ty-pes were then compared using a I-way ANOVA test applied to ranks The Morisita-Horn index (CmH) was used to compare species similarity among habitats (Magur- ran 1988)
We conducted ANOVA and Tukey pairwise mul- tiple comparisons to check whether total relative abundance (ie total bat species captured during the year) differed among habitat types We also used ANOVA to determine whether the relative abun- dance of frugivorous bats differed between fruiting Ficus trees and non-fruiting Ceibu and Cedrella trees (ie controls) in the study area For this analysis we used total capture of frugivorous bat5 from April through August 1996 months when Ficus trees bore fruits Relative abundance was normalized b a square root transformation before conducting these analyses Linear regression analysis was conducted to determine the association between relative abun- dance of all bats and the percentage of canopy with ripe fruits at isolated Ficus trees in pastures Finally simple linear correlation analysis was conducted to determine the degree of association behveen dis- tance to the nearest forest fragment and bat abun- dance in riparian vegetation and around isolated trees in pastures
RESULTS-Species Richness and Community Structurewe captured 652 bats from 20 spe- cies in 4 families (Table 1) Most species were from the family Phyllostomidae (15 species) The families Mormoopidae Vespertilionidae and Molossidae were represented by 5 species (Table 1) As expected frugivorous bats were the most abundant (81 of the total captures) followed by insectivores (15) nectarivore- frugivores (2) and sanguivores (2) Total relative abundance varied greatly with species Five species accounted for 84 of the cap- tures These were the frugivores Sturnira liliurn (40) Artibeus jamaicensis (15) Carollia per-spinllata ( lo) and Dermanura tolteca (9) and the insectivore Pteronotus parnellii (10)
The cumulative number of species over all
habitat types tended to level off after 20 nights of netting The species accumulation function predicted a maximum of 23 species According to Moreno and Halffter (1999) 19 nights at least are enough to record 90 of frugivorous bats in a heterogeneous habitat Therefore our sampling provided a reasonably good es- timation of species richness at ground level The maximum numbers of bat species for each habitat predicted by the accumulation func- tion were 17 13 and 12 for the riparian the Acus trees and control trees respectively (Fig l ) and we captured 18 13 and 12 species respectively
Bat diversity varied among habitat types Abundance species richness and Shannon-Wiener index (H)were higher in riparian veg- etation than under isolated trees in pastures despite the lower sampling effort in the ripar- ian habitat (Table 2) Some species were pre- sent only in riparian vegetation whereas others used both habitats (Table 1) The adjusted rel- ative abundance of all bats differed among habitat types (ANOVA F = 2184 df = 2 P lt 0001 Fig 2) According to a posteriori multi- ple comparison tests relative abundance was significantly higher in the riparian habitat than at isolated Ficus trees (q = 833 P lt 005) or control trees (q = 784 P lt 005) relative abundance was not significantly different be- tween isolated Ficus trees and control trees ( q = 048 P gt 003) The Shannon-Wiener index was marginally different between the riparian and control trees (F = 581 P = 006 Table 2) Species similarity was highest between iso- lated trees (ie between Ficus and corltrol trees) and lowest between isolated trees and riparian habitats (Table 2) Bats also were cap- tured in different habitat types according to their feeding habits In particular insectivo- rous bats were more abundant in riparian sites (70) than in isolated tree sites (30) in con- trast frugivores were more abundant at isolat- ed trees (75) than at riparian sites (25)
Use of Isolated Trees and fiparinn Vegetation Habitats-Frugivorous bats were caught under the canopies of isolated trees irrespective of the seed dispersal mechanism (bat or wind-dis-persed) or phenological state of the tree Rela- tive abundance for frugivorous bats at control trees was slightly higher than at ficus trees dur- ing most of the year However no significant differences were found among mean relative
589 1)ecember 2003 Oalindo-Gonrilez and Sosa-Frugivorous bats in fragmented tropiral landscape
TABLE1-Bat species recorded in a fragmented landscape in Los Tuxtlas lkracruz Mexico October 1995 through September 1996 Habitat type I = under isolated tree R = riparian (number of individuals cap- tured)
Relative Habitat type
abundance n ( ) I K Feeding guild
Mormoopidae
Pteronotus dauyi 8 12 1 7 insectivore Pteronotus parnelliz 64 98 19 43 insectibore Mor~nocr megalophylla 16 25 2 14 insectivore
Phvllostomldae
Phyllostorninae Phgtllostomus dzscolor 1 02 0 1 frugivore
Carolliinae Carollza hreurcauda 1 02 0 1 frugivore Carollza perspzczllata 65 99 28 37 frugivore
Glossophaginae Glossophaga soricina 10 15 s 3 nectar-frugivore Choeronzscus godmani 1 02 0 I nectar-frugivore
Stenoderrnatlnae Sturnmi izlzum frugirore Di-oderma bzlobatuni frugivore Platyrrhznus hellm frugivore Vamnpyrodes caracczolz frugivore Chzrod~rma uzllosum frughore Artzbeu lamazcenzi frugivore Artzbvus lzturatus frugivore Dermanura toltrca frugivore Dernlanura pharotzs frugivorc
Desmodontinae Desmodus rotundus 10 15 4 6 sanguivore
Vcspertilionidae
My otis keaysz 11 17 0 11 insectivore
Molossidae
iMolossuc rufus I 02 1 0 insectivore
Total 652 391 261
abundance values for frugivorous bats at control parian sites and bats moved from 1 habitat vs Ficus trees during their fruiting seasons (AN- type to another (ie among riparian sites iso- OVA F = 035 P = 057 df = 1)Finally no lated Ficus trees and control trees) Most re- significant relationship was found between bat captures (20 individuals) were caught on sep- abundance and percentage of ripe Ficus fruits arate nights and at different sites and habitats in isolated trees ( r = 027 P = 066 n = 5) from where they were first captured Distance
We recaptured 22 individuals belonging to 3 among recapture sites ranged from 117 to 692 frugivorous species a total of 34 times (Table m (mean 2805 n~ 234 m SE) Recapture 3) Carollia perspiczllata was the most recaptured times range from a few hours in the same night species even though Sturnira lilium was more to 8 months after the first capture Recaptured abundant indicating that the former shows bats did not seem to have any harm caused by more fidelity to the studied pastures than the the necklaces latter We recorded more recaptures at the ri- Relative abundance of all bats decreased
584 The Southwestern Vuturalzst vol 48 no 4
20 1 A A A
0 Ficus
+ Control
+ R~par~an
A Total 1995 Months 1996
0 v 0 5 10 15 20 25 30 35 40 45
-0 -Riparian -v-Ficus trees -0-Control trees
Accumulated netting nightslmonth FIG2-Mean adjusted relative abundance of bats
FIG l-Accumulation curves (lines) for bat spe- in different habitat t7rpes of
ties adjusted by exponential function for each hab- landscape a -year at Tuxtlas Ve-itat and for total captures in pastures at Los Tuxtlas racruZ
Veracruz Mexico
with increasing distance from the nearest for-est edge (Fig 3 ) To eliminate potential bias produced by the presence of water (which could function as an attractor to bats) at ripar-ian sites we analyzed separately 1) the corre-lation between bat relative abundance and dis-tance to isolated trees ( r = -072 P gt 005) and 2) the relationship between bat relative abundance and distance to riparian sites ( r =
-095 P = 005) Most bats were captured in the riparian habitat located nearest to a forest fragment The fewest number of bats were cap-tured at isolated Ficus trees located the greatest distance from the forest edge
D~scus s~o~-Ourresults show that bats (mostly frugivorous species) flew around iso-
lated trees and in strips of riparian vegetation in pastures within a fragmented tropical land-scape while foraging commuting or looking for perches Bat abundance richness and di-versity were higher in riparian corridors than under the canopy of isolated trees Frugivorous bats were caught under the canopies of differ-ent isolated trees throughout the year irre-spective of the seed dispersal mechanism of the tree or the presence or absence of fruits As predicted bat abundance decreased with distance to the nearest forest fragment
In both the riparian and isolated tree sites frugivorous species from the family Phyllo-stomidae were most common Species in the subfamily Stenodermatinae were more numer-ous (9 out of 12 potentially present) than the subfamily Phyllostominae (1 out of 7 poten-
TABLE2-Bat species richness sampling effort (SE h = netting hours m = total meters of installed nets) adjusted relative abundance (ARA see text for units) mean Shannon-Wiener diversity index (IF) and pairwise comparisons of species similarity among habitats (Morisita-Horn index CmH) on different habitat types sampled in pastures in Los Tuxtlas Veracruz Mexico
BatsSpecies expected
Species cap- SE Mean Habitat type n observed St CI tured (h X m) ARA IF CmH2
- - ---
Fzcus (a) 2 13 113 88-137 150 36324 413 0776 097 (a-b) Control trees (b) 2 12 112 99-125 241 48654 495 0670 083 (b-c) Riparian (c) 3 18 156 141-172 261 8096 3224 0824 090 (c-a)
Total 652
At time tk when an increase of 5 (k = 005) in the species list is predicted according to the exponential species accumulation function Lower and upper 93 confidence limits shown
CmH = 1 (100 similarity)
December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

582 Thr Southwestern iVaturaltst vol 48 no 4
Newton algorithm in the package STATISTICA (Statsoft 1991)
Due to our use of mist-nets of different sizes we calculated adjusted relative abundance (number of individuals per 1000 net-m X hour) by sampling ef- fort with the following formula adjusted relative abundance = 1000 [b (nh)] where b = total Ilum- ber of captured bats and nh = sampling effort as already defined Bat species diversity for each habitat type (ie riparian vegetation isolated Fzcus trees and isolated control trees) was estimated with the Shannon-Uiener index (H) using the total annual captures Diversity indices among habitat ty-pes were then compared using a I-way ANOVA test applied to ranks The Morisita-Horn index (CmH) was used to compare species similarity among habitats (Magur- ran 1988)
We conducted ANOVA and Tukey pairwise mul- tiple comparisons to check whether total relative abundance (ie total bat species captured during the year) differed among habitat types We also used ANOVA to determine whether the relative abun- dance of frugivorous bats differed between fruiting Ficus trees and non-fruiting Ceibu and Cedrella trees (ie controls) in the study area For this analysis we used total capture of frugivorous bat5 from April through August 1996 months when Ficus trees bore fruits Relative abundance was normalized b a square root transformation before conducting these analyses Linear regression analysis was conducted to determine the association between relative abun- dance of all bats and the percentage of canopy with ripe fruits at isolated Ficus trees in pastures Finally simple linear correlation analysis was conducted to determine the degree of association behveen dis- tance to the nearest forest fragment and bat abun- dance in riparian vegetation and around isolated trees in pastures
RESULTS-Species Richness and Community Structurewe captured 652 bats from 20 spe- cies in 4 families (Table 1) Most species were from the family Phyllostomidae (15 species) The families Mormoopidae Vespertilionidae and Molossidae were represented by 5 species (Table 1) As expected frugivorous bats were the most abundant (81 of the total captures) followed by insectivores (15) nectarivore- frugivores (2) and sanguivores (2) Total relative abundance varied greatly with species Five species accounted for 84 of the cap- tures These were the frugivores Sturnira liliurn (40) Artibeus jamaicensis (15) Carollia per-spinllata ( lo) and Dermanura tolteca (9) and the insectivore Pteronotus parnellii (10)
The cumulative number of species over all
habitat types tended to level off after 20 nights of netting The species accumulation function predicted a maximum of 23 species According to Moreno and Halffter (1999) 19 nights at least are enough to record 90 of frugivorous bats in a heterogeneous habitat Therefore our sampling provided a reasonably good es- timation of species richness at ground level The maximum numbers of bat species for each habitat predicted by the accumulation func- tion were 17 13 and 12 for the riparian the Acus trees and control trees respectively (Fig l ) and we captured 18 13 and 12 species respectively
Bat diversity varied among habitat types Abundance species richness and Shannon-Wiener index (H)were higher in riparian veg- etation than under isolated trees in pastures despite the lower sampling effort in the ripar- ian habitat (Table 2) Some species were pre- sent only in riparian vegetation whereas others used both habitats (Table 1) The adjusted rel- ative abundance of all bats differed among habitat types (ANOVA F = 2184 df = 2 P lt 0001 Fig 2) According to a posteriori multi- ple comparison tests relative abundance was significantly higher in the riparian habitat than at isolated Ficus trees (q = 833 P lt 005) or control trees (q = 784 P lt 005) relative abundance was not significantly different be- tween isolated Ficus trees and control trees ( q = 048 P gt 003) The Shannon-Wiener index was marginally different between the riparian and control trees (F = 581 P = 006 Table 2) Species similarity was highest between iso- lated trees (ie between Ficus and corltrol trees) and lowest between isolated trees and riparian habitats (Table 2) Bats also were cap- tured in different habitat types according to their feeding habits In particular insectivo- rous bats were more abundant in riparian sites (70) than in isolated tree sites (30) in con- trast frugivores were more abundant at isolat- ed trees (75) than at riparian sites (25)
Use of Isolated Trees and fiparinn Vegetation Habitats-Frugivorous bats were caught under the canopies of isolated trees irrespective of the seed dispersal mechanism (bat or wind-dis-persed) or phenological state of the tree Rela- tive abundance for frugivorous bats at control trees was slightly higher than at ficus trees dur- ing most of the year However no significant differences were found among mean relative
589 1)ecember 2003 Oalindo-Gonrilez and Sosa-Frugivorous bats in fragmented tropiral landscape
TABLE1-Bat species recorded in a fragmented landscape in Los Tuxtlas lkracruz Mexico October 1995 through September 1996 Habitat type I = under isolated tree R = riparian (number of individuals cap- tured)
Relative Habitat type
abundance n ( ) I K Feeding guild
Mormoopidae
Pteronotus dauyi 8 12 1 7 insectivore Pteronotus parnelliz 64 98 19 43 insectibore Mor~nocr megalophylla 16 25 2 14 insectivore
Phvllostomldae
Phyllostorninae Phgtllostomus dzscolor 1 02 0 1 frugivore
Carolliinae Carollza hreurcauda 1 02 0 1 frugivore Carollza perspzczllata 65 99 28 37 frugivore
Glossophaginae Glossophaga soricina 10 15 s 3 nectar-frugivore Choeronzscus godmani 1 02 0 I nectar-frugivore
Stenoderrnatlnae Sturnmi izlzum frugirore Di-oderma bzlobatuni frugivore Platyrrhznus hellm frugivore Vamnpyrodes caracczolz frugivore Chzrod~rma uzllosum frughore Artzbeu lamazcenzi frugivore Artzbvus lzturatus frugivore Dermanura toltrca frugivore Dernlanura pharotzs frugivorc
Desmodontinae Desmodus rotundus 10 15 4 6 sanguivore
Vcspertilionidae
My otis keaysz 11 17 0 11 insectivore
Molossidae
iMolossuc rufus I 02 1 0 insectivore
Total 652 391 261
abundance values for frugivorous bats at control parian sites and bats moved from 1 habitat vs Ficus trees during their fruiting seasons (AN- type to another (ie among riparian sites iso- OVA F = 035 P = 057 df = 1)Finally no lated Ficus trees and control trees) Most re- significant relationship was found between bat captures (20 individuals) were caught on sep- abundance and percentage of ripe Ficus fruits arate nights and at different sites and habitats in isolated trees ( r = 027 P = 066 n = 5) from where they were first captured Distance
We recaptured 22 individuals belonging to 3 among recapture sites ranged from 117 to 692 frugivorous species a total of 34 times (Table m (mean 2805 n~ 234 m SE) Recapture 3) Carollia perspiczllata was the most recaptured times range from a few hours in the same night species even though Sturnira lilium was more to 8 months after the first capture Recaptured abundant indicating that the former shows bats did not seem to have any harm caused by more fidelity to the studied pastures than the the necklaces latter We recorded more recaptures at the ri- Relative abundance of all bats decreased
584 The Southwestern Vuturalzst vol 48 no 4
20 1 A A A
0 Ficus
+ Control
+ R~par~an
A Total 1995 Months 1996
0 v 0 5 10 15 20 25 30 35 40 45
-0 -Riparian -v-Ficus trees -0-Control trees
Accumulated netting nightslmonth FIG2-Mean adjusted relative abundance of bats
FIG l-Accumulation curves (lines) for bat spe- in different habitat t7rpes of
ties adjusted by exponential function for each hab- landscape a -year at Tuxtlas Ve-itat and for total captures in pastures at Los Tuxtlas racruZ
Veracruz Mexico
with increasing distance from the nearest for-est edge (Fig 3 ) To eliminate potential bias produced by the presence of water (which could function as an attractor to bats) at ripar-ian sites we analyzed separately 1) the corre-lation between bat relative abundance and dis-tance to isolated trees ( r = -072 P gt 005) and 2) the relationship between bat relative abundance and distance to riparian sites ( r =
-095 P = 005) Most bats were captured in the riparian habitat located nearest to a forest fragment The fewest number of bats were cap-tured at isolated Ficus trees located the greatest distance from the forest edge
D~scus s~o~-Ourresults show that bats (mostly frugivorous species) flew around iso-
lated trees and in strips of riparian vegetation in pastures within a fragmented tropical land-scape while foraging commuting or looking for perches Bat abundance richness and di-versity were higher in riparian corridors than under the canopy of isolated trees Frugivorous bats were caught under the canopies of differ-ent isolated trees throughout the year irre-spective of the seed dispersal mechanism of the tree or the presence or absence of fruits As predicted bat abundance decreased with distance to the nearest forest fragment
In both the riparian and isolated tree sites frugivorous species from the family Phyllo-stomidae were most common Species in the subfamily Stenodermatinae were more numer-ous (9 out of 12 potentially present) than the subfamily Phyllostominae (1 out of 7 poten-
TABLE2-Bat species richness sampling effort (SE h = netting hours m = total meters of installed nets) adjusted relative abundance (ARA see text for units) mean Shannon-Wiener diversity index (IF) and pairwise comparisons of species similarity among habitats (Morisita-Horn index CmH) on different habitat types sampled in pastures in Los Tuxtlas Veracruz Mexico
BatsSpecies expected
Species cap- SE Mean Habitat type n observed St CI tured (h X m) ARA IF CmH2
- - ---
Fzcus (a) 2 13 113 88-137 150 36324 413 0776 097 (a-b) Control trees (b) 2 12 112 99-125 241 48654 495 0670 083 (b-c) Riparian (c) 3 18 156 141-172 261 8096 3224 0824 090 (c-a)
Total 652
At time tk when an increase of 5 (k = 005) in the species list is predicted according to the exponential species accumulation function Lower and upper 93 confidence limits shown
CmH = 1 (100 similarity)
December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

589 1)ecember 2003 Oalindo-Gonrilez and Sosa-Frugivorous bats in fragmented tropiral landscape
TABLE1-Bat species recorded in a fragmented landscape in Los Tuxtlas lkracruz Mexico October 1995 through September 1996 Habitat type I = under isolated tree R = riparian (number of individuals cap- tured)
Relative Habitat type
abundance n ( ) I K Feeding guild
Mormoopidae
Pteronotus dauyi 8 12 1 7 insectivore Pteronotus parnelliz 64 98 19 43 insectibore Mor~nocr megalophylla 16 25 2 14 insectivore
Phvllostomldae
Phyllostorninae Phgtllostomus dzscolor 1 02 0 1 frugivore
Carolliinae Carollza hreurcauda 1 02 0 1 frugivore Carollza perspzczllata 65 99 28 37 frugivore
Glossophaginae Glossophaga soricina 10 15 s 3 nectar-frugivore Choeronzscus godmani 1 02 0 I nectar-frugivore
Stenoderrnatlnae Sturnmi izlzum frugirore Di-oderma bzlobatuni frugivore Platyrrhznus hellm frugivore Vamnpyrodes caracczolz frugivore Chzrod~rma uzllosum frughore Artzbeu lamazcenzi frugivore Artzbvus lzturatus frugivore Dermanura toltrca frugivore Dernlanura pharotzs frugivorc
Desmodontinae Desmodus rotundus 10 15 4 6 sanguivore
Vcspertilionidae
My otis keaysz 11 17 0 11 insectivore
Molossidae
iMolossuc rufus I 02 1 0 insectivore
Total 652 391 261
abundance values for frugivorous bats at control parian sites and bats moved from 1 habitat vs Ficus trees during their fruiting seasons (AN- type to another (ie among riparian sites iso- OVA F = 035 P = 057 df = 1)Finally no lated Ficus trees and control trees) Most re- significant relationship was found between bat captures (20 individuals) were caught on sep- abundance and percentage of ripe Ficus fruits arate nights and at different sites and habitats in isolated trees ( r = 027 P = 066 n = 5) from where they were first captured Distance
We recaptured 22 individuals belonging to 3 among recapture sites ranged from 117 to 692 frugivorous species a total of 34 times (Table m (mean 2805 n~ 234 m SE) Recapture 3) Carollia perspiczllata was the most recaptured times range from a few hours in the same night species even though Sturnira lilium was more to 8 months after the first capture Recaptured abundant indicating that the former shows bats did not seem to have any harm caused by more fidelity to the studied pastures than the the necklaces latter We recorded more recaptures at the ri- Relative abundance of all bats decreased
584 The Southwestern Vuturalzst vol 48 no 4
20 1 A A A
0 Ficus
+ Control
+ R~par~an
A Total 1995 Months 1996
0 v 0 5 10 15 20 25 30 35 40 45
-0 -Riparian -v-Ficus trees -0-Control trees
Accumulated netting nightslmonth FIG2-Mean adjusted relative abundance of bats
FIG l-Accumulation curves (lines) for bat spe- in different habitat t7rpes of
ties adjusted by exponential function for each hab- landscape a -year at Tuxtlas Ve-itat and for total captures in pastures at Los Tuxtlas racruZ
Veracruz Mexico
with increasing distance from the nearest for-est edge (Fig 3 ) To eliminate potential bias produced by the presence of water (which could function as an attractor to bats) at ripar-ian sites we analyzed separately 1) the corre-lation between bat relative abundance and dis-tance to isolated trees ( r = -072 P gt 005) and 2) the relationship between bat relative abundance and distance to riparian sites ( r =
-095 P = 005) Most bats were captured in the riparian habitat located nearest to a forest fragment The fewest number of bats were cap-tured at isolated Ficus trees located the greatest distance from the forest edge
D~scus s~o~-Ourresults show that bats (mostly frugivorous species) flew around iso-
lated trees and in strips of riparian vegetation in pastures within a fragmented tropical land-scape while foraging commuting or looking for perches Bat abundance richness and di-versity were higher in riparian corridors than under the canopy of isolated trees Frugivorous bats were caught under the canopies of differ-ent isolated trees throughout the year irre-spective of the seed dispersal mechanism of the tree or the presence or absence of fruits As predicted bat abundance decreased with distance to the nearest forest fragment
In both the riparian and isolated tree sites frugivorous species from the family Phyllo-stomidae were most common Species in the subfamily Stenodermatinae were more numer-ous (9 out of 12 potentially present) than the subfamily Phyllostominae (1 out of 7 poten-
TABLE2-Bat species richness sampling effort (SE h = netting hours m = total meters of installed nets) adjusted relative abundance (ARA see text for units) mean Shannon-Wiener diversity index (IF) and pairwise comparisons of species similarity among habitats (Morisita-Horn index CmH) on different habitat types sampled in pastures in Los Tuxtlas Veracruz Mexico
BatsSpecies expected
Species cap- SE Mean Habitat type n observed St CI tured (h X m) ARA IF CmH2
- - ---
Fzcus (a) 2 13 113 88-137 150 36324 413 0776 097 (a-b) Control trees (b) 2 12 112 99-125 241 48654 495 0670 083 (b-c) Riparian (c) 3 18 156 141-172 261 8096 3224 0824 090 (c-a)
Total 652
At time tk when an increase of 5 (k = 005) in the species list is predicted according to the exponential species accumulation function Lower and upper 93 confidence limits shown
CmH = 1 (100 similarity)
December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman
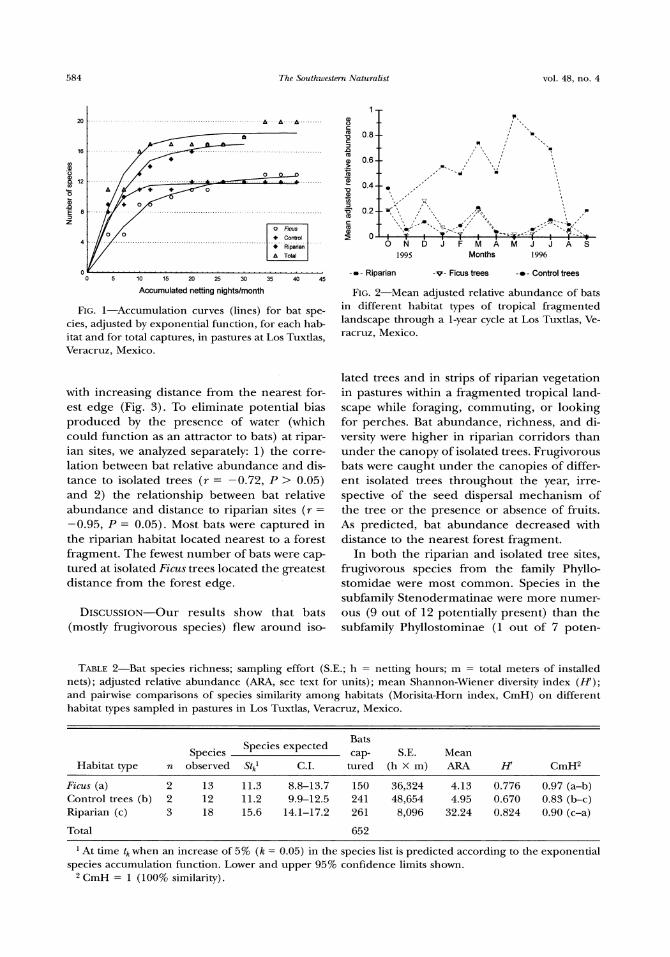
584 The Southwestern Vuturalzst vol 48 no 4
20 1 A A A
0 Ficus
+ Control
+ R~par~an
A Total 1995 Months 1996
0 v 0 5 10 15 20 25 30 35 40 45
-0 -Riparian -v-Ficus trees -0-Control trees
Accumulated netting nightslmonth FIG2-Mean adjusted relative abundance of bats
FIG l-Accumulation curves (lines) for bat spe- in different habitat t7rpes of
ties adjusted by exponential function for each hab- landscape a -year at Tuxtlas Ve-itat and for total captures in pastures at Los Tuxtlas racruZ
Veracruz Mexico
with increasing distance from the nearest for-est edge (Fig 3 ) To eliminate potential bias produced by the presence of water (which could function as an attractor to bats) at ripar-ian sites we analyzed separately 1) the corre-lation between bat relative abundance and dis-tance to isolated trees ( r = -072 P gt 005) and 2) the relationship between bat relative abundance and distance to riparian sites ( r =
-095 P = 005) Most bats were captured in the riparian habitat located nearest to a forest fragment The fewest number of bats were cap-tured at isolated Ficus trees located the greatest distance from the forest edge
D~scus s~o~-Ourresults show that bats (mostly frugivorous species) flew around iso-
lated trees and in strips of riparian vegetation in pastures within a fragmented tropical land-scape while foraging commuting or looking for perches Bat abundance richness and di-versity were higher in riparian corridors than under the canopy of isolated trees Frugivorous bats were caught under the canopies of differ-ent isolated trees throughout the year irre-spective of the seed dispersal mechanism of the tree or the presence or absence of fruits As predicted bat abundance decreased with distance to the nearest forest fragment
In both the riparian and isolated tree sites frugivorous species from the family Phyllo-stomidae were most common Species in the subfamily Stenodermatinae were more numer-ous (9 out of 12 potentially present) than the subfamily Phyllostominae (1 out of 7 poten-
TABLE2-Bat species richness sampling effort (SE h = netting hours m = total meters of installed nets) adjusted relative abundance (ARA see text for units) mean Shannon-Wiener diversity index (IF) and pairwise comparisons of species similarity among habitats (Morisita-Horn index CmH) on different habitat types sampled in pastures in Los Tuxtlas Veracruz Mexico
BatsSpecies expected
Species cap- SE Mean Habitat type n observed St CI tured (h X m) ARA IF CmH2
- - ---
Fzcus (a) 2 13 113 88-137 150 36324 413 0776 097 (a-b) Control trees (b) 2 12 112 99-125 241 48654 495 0670 083 (b-c) Riparian (c) 3 18 156 141-172 261 8096 3224 0824 090 (c-a)
Total 652
At time tk when an increase of 5 (k = 005) in the species list is predicted according to the exponential species accumulation function Lower and upper 93 confidence limits shown
CmH = 1 (100 similarity)
December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

December 2003 Galindo-Gonzalez and Sosa-Frugivorous bats in fragmented tropical landscape 585
TABLE3-Recaptured species in pastures of Los Tuxtlas Veracruz Mexico showing for each species the total captures number and percentage of recaptures from total captures of each species number of times some individuals were recaptured average distance (and range) between recapture sites (ADRS) and time between recaptures (TBR) Habitat type F T = Ficus tree CT = control tree R = riparian
Recaptures Times individuals Habitat type recaptured
Num- ADRS TBR Bat species n ber F T C T R 1 2 3 4 5 (m) (days)
Carollia perspicillata 65 23 55 8 2 13 12 6 3 1 1 284 (175-417) 0-244 Stumira lilium 263 7 27 2 3 2 6 1 - - - 348 (0-692) 1-153 Dermanura tolteca 58 4 74 1 0 3 4 - - - - 295 (0-417) 30-122
Total 34 11 5 18
tially present Navarro 1982) These trends were also found in disturbed habitats of the Yucatan Peninsula and of the Selva Lacandona in Mexico (Fenton et al 1992Medellin et al 2000) In addition similar results including the capture of the insectivore Pteronotus parnel-lii have been recorded for disturbed habitats in French Guiana (Brosset et al 1996)
Despite a capture bias related to netting at ground level we caught a few individuals of Artibeus lituratus Phyllostomus discolor and Chi-roderma uillosum species that are classified as highly clutteredgleaning frugivores of the canopy stratum (Bernard 2001 Kalko and Handley 2001) and Molossus rufus an aerial insectivorous species that usually forages above
I Riparian
I
I
Isolated tree ~ ~ ~ ~ ~ ~ ~ ~ ~~~~
95 143 262 276 281 286 333 381
Distance to closest forest patch (m)
FIG3-Effect of distance from monitoring sites to nearest forest fragment on adjusted relative abundance of bats in riparian and isolated tree habitats at Los Tuxtlas Veracruz Mexico Notice logarithmic scale of y-axis
586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

586 The Southwestern vaturalzst vol 48 no 4
canopy Fifty-six percent o f the bat species his- torically recorded for the tropical rain forest at Los Tuxtlas were captured in a single year in our study [ie 20 species caught and 37 spe- cies reported (Navarro 1982 Estrada et al 1993)l Thus we conclude that considerable species richness is still maintained at the pre- sent degree o f forest fragmentation and aver-age distance between habitat patches riparian vegetation and isolated trees Although dis-tances between forest patches ranged from 10 to 500 m average distance between riparian corridors and isolated trees was only 40 to 50 m (Guevara et al 1998) This distance is easily traversed by bats whose commuting and for- aging movements range from hundreds to thousands o f meters according to radio-track- ing data For instance the foraging distance during 1 night for Carollia perspicillata ranges from 64 to 132 k m (Heithaus and Fleming 1978 Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991) and ranges for Artibeus jamaicensis from 1 to 8 k m (Janzen et al 1976 Morrison 1978a 1978c 1980b) T h e average distance between feeding areas and the day roost ranges from 015 to 8 k m for A jamaicensis A Lituratus C perspicillata and kmpyrodes caracciolz (Morrison 1 9 8 0 ~ ) T h e traveling distances o f bats varied according to the time o f the year (dry and wet season) plant species density and distribution o f the fruit trees habitat fragmentation m o o n phase bat sex and bat behavior to avoid predators ( Janzen et al 1976 Heithaus and Fleming 1978 hlorrison 1978a 1978c 1980a 1980h Fleming and Heithaus 1981 Fleming 1988 Charles-Dominique 1991 Galindo-Gonziilez 1998)
Given the 23 bat species predicted at the study site by the accumulation function 38 (14 species) were already extinct at our studied fragmented landscape I f the forest and ripar- ian corridors are removed the distance be- tween forest fragments will increase and we will certainly lose additional species in this re- gion Bat abundance was inversely proportion- al to forest proximity However the presence o f a given species i n a patch might be a func- tion not only o f patch size and isolation bu t also o f the neighbouring habitat size and qual- ity (AndrGn 1994) Species richness in pasture habitats might depend o n nearby forests Iso- lated trees and shrubs and riparian vegetation
might help maintain the number o f bat species that remain in the fragmented landscape o f Los Tuxtlas but only i f contiguous forest rem- nants are preserved
Bat abundance and species richness at Los Tuxtlas were highest i n the riparian habitat a finding similar to that reported for riverine for- ests in southern Africa (Rautenbach et al 1996) Bats might use riparian habitat as a source o f water as a foraging area or as a shel- tered corridor to commute between forest remnants and isolated trees in pastures 41-though isolated trees and riparian vegetation might provide day roosts none were located in the Los Tuxtlas study sites T h e scarcity o f day roosts is likely to be a limiting factor in a land- scape where 63 to 80 o f the forest has been removed Landscapes with remnant forest patches edges riparian corridors live fences and isolated trees are common in many re-gions o f the Neotropics Populations o f bat species that are highly dependent o n forests might coexist within such fragmented land-scapes but only i f forest patches o f adequate size are retained (Andrkn 1994 Granjon et a] 1996 Krohne 1997)
Our hypothesis that frugivorous bats visit iso- lated trees in pastures only for ripe fruits was not supported W e found n o differences in captures around fig trees and control trees during the fruiting time o f figs Actually we recorded less captures around fig trees during this period W e think that the presence at the same time o f many other fruiting Ficus trees o f the same and different species in the pas- tures or in nearby forest patches and live fenc- es diluted the number o f bats flying toward or around our studied isolated fig trees O n the other hand based on morphological traits col- or and scent and fruiting asynchrony Fin~s col- ubnnne fruits are proposed to be dispersed mainly by birds and in a minor proportion by bats (Korine et al 2000) Our data suggest that frugivorous bats flew near isolated trees shrubs and riparian vegetation in pastures year-round possibly seeking ripe fruits com- muting from one place to another or hiding from predators Isolated nonfruiting trees in pastures might have provided bats with roosts and shelter or acted as stepping stones for trav- eling across fragmented landscapes to differ- ent forest remnants as has been suggested for birds ( G ~ ~ e v a r a et a] 1998)
587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

587 December 2003 Galindo-Gonzilez and Sosa-Frugivorous
Frugivorous bats deposit a comparable amount of seeds under isolated Ficus trees in pastures as do birds (Galindo-Gonzalez et al 2000) They also might be dispersing seeds from isolated trees to forest fragments as evi- denced by paternity analysis (Aldrich and Hamrick 1998)Although abundant visual ev- idence suggests that abandoned fields and pas- tures lacking isolated trees can be quickly re- generated with bat-dispersed plants in tropical landscapes we presently ignore the threshold distance that allows frugivorous bats to fly across pastures with no isolated trees The pres- ence of isolated trees and riparian corridors in pastures would promote bat flights across pas- tures This would consequently increase the probability of dispersing seeds from nearby remnants into the pastures thus enhancing the potential for forest regeneration We need more studies on distance thresholds limiting nonstop bat flights across landscape habitats on forest patch size quality and distribution and on tree architecture as requisites for bat presence in fragmented landscapes
In conclusion bats at Los Tuxtlas were pre- sent and diverse year-round near isolated trees and in strips of riparian vegetation in pastures that are relatively close to surrounding patches of native forest Bats were caught under the canopies of isolated trees even when they lacked ripe fruits Abundance of bats was in- versely proportional to distance from nearest forest fragment We suggest that riparian veg- etation and isolated trees might be crucial for assuring bat mobility among different human- made habitats of the Neotropical fragmented landscape as well as the successful dispersal of bat-visited plant species
U7e thank J Laborde 0 Mu~ioz 0 Barrera T Fleming and G Sanchez for their help and com-panionship in fieldwork We also thank V Rico-Gray R A Medellin H Arita S Guevara A Kurta D Bennack R Manson M B Fenton T Fleming and an anonymous reviewer for their helpful and valu- able comments on early drafts of this paper This research was partially funded by doctoral grants for J Galindo-Gonzalez from CONACyT and Banco de Mexico additional support for field work was pro- vided by CONACyT Number 4127P-N9607 Europe- an Union Number B7-811095957D4 and the Departments of Ecologia Vegetal (90216) and Eco- sistemas Templados (90219) from Instituto de Ecol- ogia AC U C Hanson donated the mist-nets A
bats in fragmented tropical landscape
M Gonzilez G C Pulido B and L G6mez B pro- vided unconditional support
AIDE T M WD J CIVELIER 1994 Barriers to low- land tropical forest restoration in the Sierra Ne- vada de Santa Marta Colombia Restoration Ecol- ogy 2219-229
ALDRICHP R f i ~ 1998 Reproduc- J L W~RICK tive dominance of pasture trees in a fragmented tropical forest mosaic Science 281103-105
A S D ~ N H 1994 Effects of habitat fragmentation on birds and mammals in landscapes with differ- ent proportions of suitable habitat a review Oi- kos 713355-366
BEIERP AYD R F NOSS 1998 Do habitat corridors provide connectivity Conservation Biology 12 1241-1252
BERNARDE 2001 Vertical stratification of bat com- munities in primary forest of Central Amazon Brazil Journal of Tropical Ecology 17115-126
Bo~acco~so 1984 Fruit F J ~ L U DS R HL~MPHREY bat niche dynamics their role in maintaining tropical forest diversity In Chadwick A C and S L Sutton editors Tropical rain forest Special Publication Leeds Philosophical and Literary So- ciety United Kingdom Pp 169-183
BONGERSF J J POPSW LYDJ MWLT DEI CASTILLO J CIR~IIS 1988 Structure and floristic compo- sition of the lowland rain forest of Los Tuxtlas Mexico Vegetatio 7455-80
BROSSET A P CEL~ES-DOMINIQVE JA COCKLE C o s s o ~ ampYD D Mxsso~1996 Bat communities and deforestation in French Guiana Canadian Journal of Zoology 741974-1982
CHAPMAS C A L J CW~LLYR W ~ G H A M K HYNTD GEBOLUD L G ~ D N E R 1992 Estima- tors of fruit abundance of tropical trees Biotro- pica 24527-531
C E L ~ E S - D ~ ~ ~ I S I Q L ~ E P 1991 Feeding strategy and activity budget of the frugivorous bat Carollia per-spicillata (Chiroptera Phyllostomidae) in French Guiana Journal of Tropical Ecology 7243-256
CROMEF H J r ~ l l ~G C RICMDS 1988 Bats and gaps microchiropteran community structure in a Queensland rain forest Ecology 691960-1969
DE JONGJ 1995 Habitat use and species richness of bats in patchy landscape Acta Theriologica 40 237-248
DIRZO R LUD M C GARC~A 1992 Rates of defor- estation in Los Tuxtlas a Neotropical area in southeast Mexico Conservation Biology 684-90
DIRZO R WD A MIRKYDA 1991 El limite boreal de la selva en el Continente Americano contraccion de la vegetacihn y solucihn de una controversia Interciencia 16240-247
E s ~ ~ 4 n 4 WD JRA R COATES-ESTRADA D MERITT
588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

588 The Southwesta Naturalzst vol 48 no 4
1993 Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas Mexico Ecography 16 309-318
FENTONM B L D ACH~RW D AUDET M B C HICKEYC MERRIMAN D M SYME M K OBRIST AYD B ADKINS 1992 Phyllostomid bats (Chirop- tera Pyllostomidae) as indicators of habitat dis- ruption in the Neotropics Biotropica 24440-446
FLEMINGT H 1982 Foraging strategies of plant-vis- iting bats In Kunz T H editor Ecology of bats Plenum Press New York Pp 287-325
FLEMINGT H 1988 The short-tailed fruit bat a study in plant-animal interactions University of Chicago Press Chicago Illinois
FLEMISGT H ampYD E R HEITHAUS 1981 Frugivo- roils bats seed shadows and the structure of the tropical forest Biotropica 13(Supplement)45-53
FLEMING AND W B SAMWRT H E R HEITHAUS 1977 An experimental analysis of the food loca- tion behavior of frugivorous bats Ecology 58 619-627
FLEMING 1990 Phenology T H AND C F WILLIAMS seed dispersal and recruitment in Cemopiapeltata (Moraceae) in Costa Rican tropical dry forest
Journal of Tropical Ecology 6163-178 GALINDO-GONZ~EZJ 1998 Dispersion de semillas
por murciitlagos su importancia en la conserva- ci6n y regeneracion del bosque tropical Acta Zoologica Mexicana (nueva serie) 73357-74
GALINDO-Goszk~zJ 1999 Los murcitlagos frugi- voros en la regeneracion de la vegetaci6n del pai- saje pastizal-selva de Los Tuxtlas Veracruz Un- published PhD thesis Iristituto de Ecologia 4C Xalapa Veracruz Mtxico
GALINDO-GOSZ~EZ ~ L U DV J SOSA J S GUEVARA 2000 Bat and bird generated seed rains at isolat- ed trees in grazing pastures their importance in restoring tropical rain forest in Mexico Conser- vation Biology 141693-1703
GORCHOVD L F COKVEIOC ~coRRA AND M J-A- RAMIILO1993 The role of seed dispersal in the natural regeneration of rain forest after strip-cut- ting in the Peruvian Amazon In Fleming T H and A Estrada editors Frugivorv and seed dis- persal ecological and evolutionary aspects Klu- wer Academic Publishers Dordrecht The Neth- erlands Pp 339-349
GRANJON J JUDM AND S RINGUET L J F COSSON 1996 Influence of tropical rainforest on mammal communities in French Guiana short-term ef- fects Acta Oecologica 17673-684
GUEVXRA 1993 Monitoring seed S AXD J LPLBORDE dispersal at isolated standing trees in pastures consequences for local species availability In Fleming T H and A Estrada editors Frugivory
and seed dispersal ecological and evolutionary aspects Kluwer Academic Publishers Dordrecht The Netherlands Pp 319-338
GL-Ex S J LBORDEAND G S~VCHEZ 1998 Are isolated remnant trees in pastures a fragmented canopy Selbyana 1934-43
GuEv~RS J MEEP MORENO-CMASOLAAN11 J L~BORDE1992 Floristic composition and struc- ture of vegetation under isolated trees in Neo- tropical pastures Journal of Vegetation Science 3655-664
G u E v ~ S S PURATA LVD E YAY DER M ~ E L 1986 The role of remnant trees in tropical sec- ondary succession Vegetatio 6674-84
HEITHAUSE R LYD T H FLEMING1978 Foraging movements of a frugivorous bat Carollia perspicil- latu (Phyllostomatidae) Ecological Monographs 48127-143
HEITHAUS -LUDE R T H FLEMING P A OPLER 1975 Foraging patterns and resource utilization in seven species of bats it1 a seasonal tropical for- est Ecology 56841-854
JANZEN ~~CKFORTH-JONESCD H G A MILLERJ M PONDK HOOPERAKD D P JWOS 1976 TWO Costa Rican bat-generated seed shadows of An-dira ircermis (Leguminosae) Ecology 57 1068- 1075
KALKO E K T 1997 Diversity in tropical bats In Urlich H editor Tropical biodiversity and sys- tematics Proceedings of the International Sy~n- posium on Biodiversity and Systematics in Trop- ical Ecosystems Bonn 1994 Alexander Koenig Research Institute and Zoological Museum Bonn Germany Pp 13-43
K~LKO JR 2001 Neo- E K v AND C 0 HANDI-EY tropical bats in the canopy diversity communi structure and implications for conservation Plant Ecology 153319-333
KNOPF F L a-D 1994 Scale perspec- F B SLISON tive on avian diversity in western riparian ecosps- tems Conservation Biology 8669-676
KORINEC E K EL-lrltO AN11 E A HERRE 2000 Fruit characteristics and factors affecting fruit re- moval in a Panamanian community of strangler figs Oecologia 123560-568
KROHNFD 1997 Dynamics of metapopulations of small mammals Journal of Mammalogy 781014- 1026
MAGCRRAYA E 1988 Ecological diversity and its measurement Princeton University Press Prince- ton New Jersey
MAIANSON 1993 Riparian landscapes Cam- G P bridge University Press Cambridge United King- dom
MEDELI~NR A H T ARITAAND 0S~YCHEZ1997 Identificacidn de 10s murcielagos de Mexico clave de campo Publicaciones Especiales NGme- ro 2 Asociacion Mexicana de Mastozoologia
589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman

589 December 2003 Galindo-Gonzalez and Sosa-Frugivorous
Bat diversity and abundance as indicators of dis- turbance in Neotropical rainforests Conserva- tion Biology 141666-1675
MORESOC E XVD G HALFFTER1999 Assessing the completeness of bat biodiversity inventories using species accumulation curves Journal of Applied Ecology 37l-11
MORRISOND Mi 1978a Foraging ecology and en- ergetics of the frugivorous bat Artibeus jamaicen- sis Ecology 59716-723
MORRISON 19786 Lunar phobia in a Neotrop- D UT ical fruit bat Artibeus jamaicensis (Chiroptera Phyllostomidae) Animal Behaviour 26852-855
MORRISOSD W 1978c Influence of habitat on the foraging distance of the fruit bat Artibeus jamai- censis Journal of Mammalogy 59622-624
MORRISOND W 1980a Foraging and day-roosting dynamics of canopy fruits bats in Panama Jour- nal of Mammalogy 6120-29
MORRISOND W 19806 Efficiency of food utilization by fruit bats Oecologia 45270-273
NXgtLLR J H DEMPS WD M POLLOCK 1993 The role of riparian corridors in maintaining re- gional biodiversity Ecological Applications 3 209-212
NAVRRO D 1982 Mamiferos de la Estaci6n de Biol- ogia Tropical Los Tuxtlas Veracruz Unpub- lished BSc thesis Facultad de Ciencias Univer- sidad Nacional Aut6noma de Mkxico
NEPSTAD S P DA C ~ O S O D C C UHLA CSSIO LNDSJ DA C ~ O S O 1996 A comparative study
bats in fragmented tropical landscape
of tree establishment in abandoned pasture and mature forest of eastern Amazonia Oikos 7625- 39
RAM~REz-PULIDO J ARROYO-J A CASTRO-CAMPILLO CABIULESm~ F A CERVANTES1996 Lista tax- onomica de 10s mamiferos terrestres de Mkxico Occasional Papers The Museum Texas Tech University 158 1-62
RAUTENBACH WDI L M B FENTOK M J MIHITING
1996 Bats in riverille forests and woodlands a latitudinal transect in southern Africa Canadian Journal of Zoology 74312-322
SOBERON 1993 The use of spe- J Ah-D J LIORENTE cies accumulation functions for the prediction of species richness Conservation Biology 7480-488
STATSOFT1991 CSS Statistics handbook volume 11 Statsoft Inc Tulsa Oklahoma
THOILIASD W D CLOCTIER M PROVENCHER AND
C HOUIE 1988 The shape of bird-and bat-gen- erated seed shadows around a tropical fruiting tree Biotropica 20347-348
TURNERM I 1996 Species loss in fragments of t rop ical rain forest a review of the evidence Journal of Applied Ecology 33200-209
~ z Q L ~ E z - ~ ~ N E s C A OROZCOG FRLVGOISAND L TREJO 1975 Observations on the seed dispersal by bats in a tropical humid region in ~eracruz Mexico Biotropica 773-76
Submitted 28 October 2001 Accepted 25 November 2002 Assoczate Editor was Loren K Ammerman
Copyright © 2022 FDOKUMEN




















