
Distribution of noncollagenous proteins in the matrix of adult human bone: Evidence of anatomic and...
11
JOURNALOFBONEANDMINERALRESEARCH Volume 8, Number 9,1993 Muy Ann Lkbcrt, lnc., Publlshen Distribution of Noncollagenous Proteins in the Matrix of Adult Human Bone: Evidence of Anatomic and Functional Heterogeneity RONALD T. INGRAM,' BART L. CLARKE,' LARRY W. FISHER,' and LORRAINE A. FITZPATRICK' ABSTRACT The microanatomic distribution of several noncollagenous proteins (NCPs) in bone matrix was examined by immunohistochemical analysis of glycol-methyl methacrylate-embedded normal adult human bone biopsies. Osteopontin and bone sialoprotein stained throughout the lamellae of both trnbecular and cortical bone. Ce- ment lines (cortical and trabeeular) and the mineralized matrix immediately adjacent to each Haversian canal were intensely stained. Osteocalcin was detected in cement lines; however, lamellar staining varied depending on the location within the individual unit of bone. In cortical bone, the inner concentric lamellae of osteons were often unstained but the outer Iamellae were heavily stained for osteocalcin. Osteonectin was not de- tected in cement lines and in most specimens revealed a pattern similar to that of osteocalcin with respect to the absence of immunostaining within the inner concentric Iamellae. Decorin was prominent in the peri- lacunar matrix, the canaliculi of osteocytes, and the matrix immediately adjacent to quiescent Haversian canals. Biglycan appeared evenly distributed throughout cortical and trabecular bone matrix. These results suggest that the incorporation of NCPs into matrix may vary depending on the stage of formation of indi- vidual bone units. The specific distribution and spatial relationship of these NCPs may be related to the function of each protein during bone resorption and formation. The distinct patterns of NCP localization in bone support the hypothesis that in addition to their structural and mineral-inducing properties, these pro- teins may influence the events associated with bone remodeling, such as recruitment, attachment, differenti- ation, and activity of bone cells. INTRODUCTION ONCOLLA~ENOUS PROTEINS (NCPs) comprise 10% of N the total bone protein. Several NCPs have been iden- tified, including osteopontin (OPN, SPP-I), bone sialo- protein (BSP),") osteocalcin (OC),"' osteonectin (ON),(3.4) biglycan (BGN, PG-11). and decorin (DCN, PG- I).('.*' Although it is known that NCPs are expressed by bone cells at specific stages of and that these proteins are incorporated into mineralized bone, (I-*) our understanding of the specific roles of these proteins during the formation and remodeling of bone is not well understood. Accumulating evidence suggests that in addi- tion to structural roles, such as collagen fibrillogenesis and regulation of matrix mineralization, (9-13) NCPs may partic- ipate in bone cell signaling and attachment. Several studies provide evidence that the composition of bone matrix may influence the type and extent of remodel- ing that occurs in this For example, differ- ences in the content of NCPs in cortical compared to tra- becular bone suggest functional heterogeneity in these two regions.(ao) Using an in vivo rat model, bone matrix parti- cles deficient in osteocalcin recruited fewer osteoclasts than normal bone. (I4) Furthermore, osteopontin immuno- activity was localized ultrastructurally to sites of osteoclast attachment in vivo. (I7) Compositional changes in bone 'Endocrine Research Unit, Department of Internal Medicine, Mayo Clinic and Mayo Foundation, Rochester, Minnesota. 'National Institute of Dental Research, National Jnstitutes of Health, Bethesda. Maryland. 1019
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Distribution of noncollagenous proteins in the matrix of adult human bone: Evidence of anatomic and...

JOURNALOFBONEANDMINERALRESEARCH Volume 8, Number 9,1993 M u y Ann Lkbcrt, lnc., Publlshen
Distribution of Noncollagenous Proteins in the Matrix of Adult Human Bone: Evidence of Anatomic and
Functional Heterogeneity
RONALD T. INGRAM,' BART L. CLARKE,' LARRY W. FISHER,' and LORRAINE A. FITZPATRICK'
ABSTRACT
The microanatomic distribution of several noncollagenous proteins (NCPs) in bone matrix was examined by immunohistochemical analysis of glycol-methyl methacrylate-embedded normal adult human bone biopsies. Osteopontin and bone sialoprotein stained throughout the lamellae of both trnbecular and cortical bone. Ce- ment lines (cortical and trabeeular) and the mineralized matrix immediately adjacent to each Haversian canal were intensely stained. Osteocalcin was detected in cement lines; however, lamellar staining varied depending on the location within the individual unit of bone. In cortical bone, the inner concentric lamellae of osteons were often unstained but the outer Iamellae were heavily stained for osteocalcin. Osteonectin was not de- tected in cement lines and in most specimens revealed a pattern similar to that of osteocalcin with respect to the absence of immunostaining within the inner concentric Iamellae. Decorin was prominent in the peri- lacunar matrix, the canaliculi of osteocytes, and the matrix immediately adjacent to quiescent Haversian canals. Biglycan appeared evenly distributed throughout cortical and trabecular bone matrix. These results suggest that the incorporation of NCPs into matrix may vary depending on the stage of formation of indi- vidual bone units. The specific distribution and spatial relationship of these NCPs may be related to the function of each protein during bone resorption and formation. The distinct patterns of NCP localization in bone support the hypothesis that in addition to their structural and mineral-inducing properties, these pro- teins may influence the events associated with bone remodeling, such as recruitment, attachment, differenti- ation, and activity of bone cells.
INTRODUCTION
ONCOLLA~ENOUS PROTEINS (NCPs) comprise 10% of N the total bone protein. Several NCPs have been iden- tified, including osteopontin (OPN, SPP-I), bone sialo- protein (BSP),") osteocalcin (OC),"' osteonectin (ON),(3.4) biglycan (BGN, PG-11). and decorin (DCN, PG- I).('.*' Although it is known that NCPs are expressed by bone cells at specific stages of and that these proteins are incorporated into mineralized bone, ( I - * )
our understanding of the specific roles of these proteins during the formation and remodeling of bone is not well understood. Accumulating evidence suggests that in addi-
tion to structural roles, such as collagen fibrillogenesis and regulation of matrix mineralization, (9-13) NCPs may partic- ipate in bone cell signaling and attachment.
Several studies provide evidence that the composition of bone matrix may influence the type and extent of remodel- ing that occurs in this For example, differ- ences in the content of NCPs in cortical compared to tra- becular bone suggest functional heterogeneity in these two regions.(ao) Using an in vivo rat model, bone matrix parti- cles deficient in osteocalcin recruited fewer osteoclasts than normal bone. ( I 4 ) Furthermore, osteopontin immuno- activity was localized ultrastructurally to sites of osteoclast attachment in vivo. (I7) Compositional changes in bone
'Endocrine Research Unit, Department of Internal Medicine, Mayo Clinic and Mayo Foundation, Rochester, Minnesota. 'National Institute of Dental Research, National Jnstitutes of Health, Bethesda. Maryland.
1019

1020 INCRAM ET AL.
matrix that are age related may lead to deficits in recruit- ment of osteoclasts and cause abnormalities of remodeling associated with aging."') In metabolic bone diseases, such as ostcogenesis imperfecta, deficiencies in one or more of the NCPs suggest that normal bone architecture and min- eralization are highly dependent on matrix proteins. These studies suggest that metabolic and pathologic pro- cesses that alter NCP production and incorporation in bone may compromise the architecture, remodeling, and structural integrity of this tissue.
With few exceptions, immunohistochemical studies of NCPs in bone have been limited to fetal or neonatal tis- sues. (5~a4-29 ) Although these studies have provided useful information regarding the developmental expression of NCPs by osteoblasts, the methods of tissue preparation often precluded detection of NCPs in bone matrix. The in- timate association of most NCPs with the mineral phase makes their retention difficult during fixation and demin- eralization. Although plastic-embedded bone enables sec- tioning of undemineralized specimens, this method often limits immunodetection because of masking of epitopes by mineral and embedding medium. Nevertheless, plastic-em- bedded bone specimens have been used successfully in ultrastructural immunohistochemical studies, possibly be- cause of their enhanced signal Some in- vestigators have noted improved immunostaining of pro- teins in bone matrix sections decalcified after embed-
Since the microanatomic distribution and relationship of various NCPs within adult bone are not well known, our understanding of how these proteins might influence cell- matrix interactions and, ultimately, bone remodeling is limited. In this study, we developed methodology for im- munohistochemical staining of NCPs in the extracellular matrix of glycol-methyl methacrylate-embedded, iliac crest bone biopsies. This method has been used successfully on sections from specimen blocks stored for up to 10 years. We hypothesize that distinct patterns of distribution for each NCP within individual units of bone provide further insight into the function of these proteins in adult bone.
ding. (14 .25 .31 )
MATERIALS AND METHODS Bone biopsies
Iliac crest bone biopsies were obtained under local anes- thesia from 16 male volunteers (ages 20-70) after informed consent (institutional review board approval No. 0478x90). Serum and urine chemical indices were normal (Table 1). Biopsies were fixed in 70% ethanol under vacuum and de- hydrated in ascending ethanols for 4 days at 4°C prior to embedding. In a separate experiment, additional biopsies were fmed in either buffered formalin (5%. 4"C, 3 days) or 70% ethanol prior to demineralization in 0.5 M EDTA (SOC, 10 days) and embedding in either glycol-methyl methacrylate (GMA) or paraffin.
Em bedding
Biopsies were embedded in modified glycol-methyl methacrylate using a temperature-controlled method
(Rainier Technical Products, Seattle, WA). Briefly, biop- sies were infiltrated for 3 days in a mixture of the follow- ing: 81 Yo (vol/vol) uninhibited methyl methacrylate, 8% (wt/vol) polyethylene glycol distearate (1540). 6.5% (vol/ vol) 2-hydroxyethyl methacrylate, 4% dibutylphthalate, and 0.65% benzoyl peroxide. Infiltrated biopsies were placed in fresh monomer containing accelerator and al- lowed to polymerize onto aluminum chucks at room tem- perature in the presence of nitrogen. Sections (5 pm) were cut using a Reichert Jung Autocut microtome equipped with a carbide steel knife and immunostained prior to mounting. Adjacent sections were stained with Goldner- Masson-trichrome stain. (33)
A n tibodies
Rabbit polyclonal antibodies LF-6,('.") LF-7,") LF-15, and LF-30(5) were generated against human noncollage- nous proteins or their sequences. Bovine bone-derived pro- teins were used to generate antisera LF-32 and LF- BONII,(16) and these antibodies cross-react to their respec- tive human bone-derived proteins. Bovine osteocalcin pro- tein, a generous gift of Dr. Paul Price, was further purified by high-performance liquid chromatography prior to use as an immunogen.
Immunohistology
Bone sections were rehydrated in 50% ethanol and decal- cified with acetic acid (1% for 10 minutes). After rinsing in Tris-buffered saline (TBS, 0.05 M Tris, 0.01 To bovine serum albumin, and 0.9% NaCI, pH 7.5). sections were blocked with 10% normal goat serum in TBS for 30 minutes. Sec- tions were stained using a Vectastain Elite ABC kit (Vector Laboratories, Burlingame, CA) according to the manufac- turer's recommendations with modifications. All steps were carried out at room temperature and were followed by a 30 minute wash. Sections were incubated with primary anti- body (1:200 in TBS containing 1.5% normal goat serum) for 1 h and washed in TBS containing 0.02% Triton X-100 ('ITBS). Primary antibody was detected by incubation with biotinylated secondary antibody (goat antirabbit) for 30 minutes and washed with TTBS. Endogenous peroxidase ac- tivity was inhibited with 1.5% H,O, and 0.1'70 sodium azide in 50% methanol for 15 minutes. Peroxidase-conjugated avidin-biotin complex (ABC) in TBS containing 0.5 M NaCl was allowed to react with secondary antibody for 30 min- utes. Antibody complexes were visualized after the addition of 0.01% diaminobenzidine and 0.01% H202 in Tris, pH 7.2, for 4 minutes. Sections were rinsed with tap water, de- hydrated with ascending alcohols, cleared with xylene, and mounted on glass slides with coverslips using Eukitt mount- ing medium (Calibrated Instruments, Inc., Hawthorne, NY). Control sections were stained using normal rabbit serum at the same dilution as primary antibody.
Sections stained for the two proteoglycans, biglycan and decorin, were preincubated with chondroitinase ABC (1.25 U/ml in 0.1 M Tris and 0.05 M calcium sulfate, pH 7.4) for 10 minutes at 37°C prior to blocking. Chondroitinase ABC enhanced specific staining, as described elsewhere. 0')
In a separate experiment, sections were treated as fol-

HETEROGENEITY OF NCP IN HUMAN BONE lo21
lows prior to immunostaining: (1) n o acetic acid pretreat- ment; (2) 1% gluteraldehyde fixation("); (3) 0.05% trypsin digestion (1-5 minutes, 37"C).(")
RESULTS
Table 1 summarizes the clinical parameters and serum values of calcium, phosphorus, and alkaline phosphatase for each normal subject. The observed immunostaining patterns for each NCP are summarized in Tiible 2 and ex- pressed diagrammatically in Fig. 1.
Antisera and nonimmune sera were tested on adjacent sections. Elimination of any one of the immunostaining steps resulted in negative immunostaining of the matrix (data not shown). Distinct, highly specific staining was ob- served using each of the antisera against matrix proteins (Fig. 2). and nonimmune rabbit sera at the :same concen- tration did not stain bone matrix (Fig. 3B).
The distribution of osteopontin and bone sialoprotein in the mineralized matrix of normal male bone is shown in Figs. 2A and B, respectively. Immunodetectable BSP and OPN demonstrated similar patterns of distribution. In- tense, highly specific staining was observed in the cement lines within the cortex (Fig. 2A and B) and trabeculae (Fig. 3D). In a similar manner, the bone matrix immediately ad- jacent to the Haversian canal was distinctly positive for both OPN and BSP. The remaining lamellae of osteons and trabecular bone were moderately immunostained for these phosphoproteins. In roughly 70% of the sections ex- amined, immunodectable BSP was evident in the perila- cunar matrix of osteocytes, and canaliculi were not stained greater than surrounding matrix.
The distribution of immunostaining for osteonectin is shown in Figs. 2C and 3E. Immunodetectable ON was not demonstrable in cement lines or osteocytes of cortical or
trabecular bone matrix. Within osteons in the cortex, posi- tive staining was most apparent in the outer lamellae; the inner lamellae were essentially devoid of stain. In the tra- beculae (Fig. 3E), regions of variable ON staining were noted in a pattern similar to the pattern of staining in cor- tical matrix (Fig. 2C).
Figure 2D illustrates the distribution of osteocalcin. Ce- ment lines in both cortical and trabecular bone stained in- tensely. Staining of the osteons varied within the same sec- tion of bone. Although the outer concentric lamellae al- ways stained positive, the inner lamellae of certain osteons were lightly stained or negative. The absence of immuno- staining for OC and ON in the inner lamellae is not thought to represent artifact since other antisera were able t o stain these regions.
Decorin stained heavily at specific sites within the bone matrix in a pattern unique to this NCP (Figs. 2E and 3C). Intense immunostaining was evident in the perilacunar matrix and canaliculi of osteocytes (Fig. 3C). In a pattern similar to that obtained using antisera against BSP and OPN, the matrix immediately adjacent to the Haversian canal stained heavily for DCN. However, the cement lines, most of the interstitial lamellae, and some osteons were devoid of immunostaining.
Figure 2F illustrates immunodetectable biglycan. In comparison t o other NCPs, light homogeneous staining was evident throughout the cortex and trabeculae. Cement lines and the matrix adjacent to each Haversian canal were generally equal in staining intensity to that of the remain- ing osteon.
Using developing bone, previous reports have described in detail the localization of NCPs to osteoblasts and osteo- cytes. ( y 1 9 3 4 4 . 3 6 . 3 7 ) The specificity of antisera for the various NCPs has been characterized previously.(y) Within the re- maining extracellular matrix, staining was highly dependent on the method of tissue preparation. In the present study, NCPs
TABLE 1. PARAMETERS FROM NORMAL MALE SUBIECTS~
1 2 3 4 5 6 7 8 9
10 1 1 12 13 14 15 16
Mean SEM
20 26 29 30 33 36 40 41 41 41 48 59 65 67 69 70 44.7
3.5
187.5 178.8 172.3 182.4 173.0 174.5 18O.O 177.0 183.0 167.6 171.2 177.8 182.0 182.3 185 .O 162.5 177.4
1.4
74.9 67.3 77.9 80.4 82.4 80.0 69.6 91.8 96.7 67.9 82.1 74.6 79.0 91.6 96.0 76.8 80.6 2.0
9.2 9.8 9.6 9.3 9.5 9.3 9.2 9.6 9.6 9.1 9.2 9.3 9.1 8.9 9.1 9.2 9.3 0.1
3.3 4.0 3.4 4.1 3.0 4.1 3.3 3.0 3.7 3.8 3.6 3.1 3.1 2.4 2.8 3.3 3.4 0.1
167 141 124 108 161 164 %
120 160 163 I88 176 145 142 225 173 153
6.8 ~~ ~~
aALP, alkaline phosphatase; SEM. standard error of the mean.
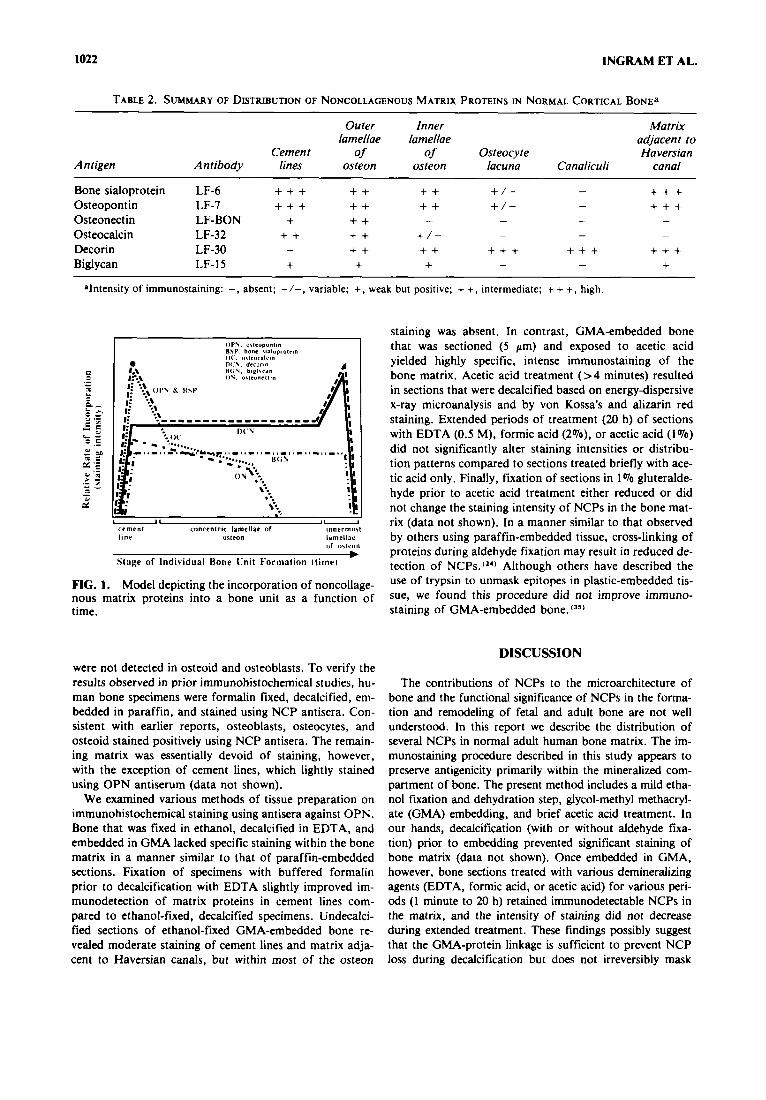
1022 INGRAM ET AL.
TABLE 2. SUMMARY OF DISTRIBUTION OF NONCOLLAGENOUS MATRIX PROTEINS IN NORMAL CORTICAL BONE^
Outer Inner Matrix lamellae lamellae adjacent to
Cemen t of of Osteocyte Haversian Antigen Antibody lines osteon osteon lacuna Canaliculi canal
+ + + + + +
Bone sialoprotein LF-6 + + + + + + + + / - - Osteopontin LF-7 + + + + + + + + / - - Osteonectin LF-BON + + + Osteocalcin LF-32 + + + + + / - Decorin LF-30 -
Biglycan LF-15 + + +
- - - -
- - -
+ + + + + + + + + + + + + + - -
alntcnsity of immunostaining: -, absent; -I-, variable; +, weak but positive; + +, intermediate; + + +, high.
O P Y . allroponlln BSP. bone rialuprolcin ( I ( , ,Itl.OCIICI.
M;\, bigl?can o\, Ollll ,nc'lln
I ) ( \, drcorin
I 4
-1 1- cement concentric lamellac or innermint l ine oileon lilmellic
of iitlrun
Stage of Individual Bone [!nit Formation ltirnel
FIG. 1. Model depicting the incorporation of noncollage- nous matrix proteins into a bone unit as a function of time.
were not detected in osteoid and osteoblasts. To verify the results observed in prior immunohistochemical studies, hu- man bone specimens were formalin fixed, decalcified, em- bedded in paraffin, and stained using NCP antisera. Con- sistent with earlier reports, osteoblasts, osteocytes, and osteoid stained positively using NCP antisera. The remain- ing matrix was essentially devoid of staining, however, with the exception of cement lines, which lightly stained using OPN antiserum (data not shown).
We examined various methods of tissue preparation on immunohistochemical staining using antisera against OPN. Bone that was fiied in ethanol, decalcified in EDTA, and embedded in GMA lacked specific staining within the bone matrix in a manner similar to that of paraffin-embedded sections. Fixation of specimens with buffered formalin prior to decalcification with EDTA slightly improved im- munodetection of matrix proteins in cement lines com- pared to ethanol-fined, decalcified specimens. Undecalci- fied sections of ethanol-fied GMA-embedded bone re- vealed moderate staining of cement lines and matrix adja- cent to Haversian canals, but within most of the osteon
staining was absent. In contrast, GMA-embedded bone that was sectioned (5 pm) and exposed to acetic acid yielded highly specific, intense immunostaining of the bone matrix. Acetic acid treatment (> 4 minutes) resulted in sections that were decalcified based on energy-dispersive x-ray microanalysis and by von Kossa's and alizarin red staining. Extended periods of treatment (20 h) of sections with EDTA (0.5 M), formic acid (2%). or acetic acid (1 070)
did not significantly alter staining intensities or distribu- tion patterns compared to sections treated briefly with ace- tic acid only. Finally, fiiation of sections in 1 Vo gluteralde- hyde prior to acetic acid treatment either reduced or did not change the staining intensity of NCPs in the bone mat- rix (data not shown). In a manner similar to that observed by others using paraffin-embedded tissue, cross-linking of proteins during aldehyde fixation may result in reduced de- tection of NCPS.(~'' Although others have described the use of trypsin to unmask epitopes in plastic-embedded tis- sue, we found this procedure did not improve immuno- staining of GMA-embedded bone. (35)
DISCUSSION
The contributions of NCPs to the microarchitecture of bone and the functional significance of NCPs in the forma- tion and remodeling of fetal and adult bone are not well understood. In this report we describe the distribution of several NCPs in normal adult human bone matrix. The im- munostaining procedure described in this study appears to preserve antigenicity primarily within the mineralized com- partment of bone. The present method includes a mild etha- nol f ia t ion and dehydration step, glycol-methyl methacryl- ate (GMA) embedding, and brief acetic acid treatment. In our hands, decalcification (with or without aldehyde f i a - tion) prior to embedding prevented significant staining of bone matrix (data not shown). Once embedded in GMA, however, bone sections treated with various demineralizing agents (EDTA, formic acid, or acetic acid) for various peri- ods (1 minute to 20 h) retained immunodetectable NCPs in the matrix, and the intensity of staining did not decrease during extended treatment. These findings possibly suggest that the GMA-protein linkage is sufficient to prevent NCP loss during decalcification but does not irreversibly mask


1024 INGRAM ET AL.
the immunodetection of these proteins. GMA may aid in the retention of NCPs in bone matrix during decalcifica- tion and may be used without prior aldehyde fixation. Our studies indicate that in normal adult human bone the in- corporation of each of the matrix proteins is distinct and highly organized and corresponds to their proposed func- tions.
Osteopontin and bone sialoprotein belong to a family of unique phosphorylated glycoproteins that contain highly conserved polyaspartic acid- and polyglutamic acid-rich se- quences that are common to mineral binding proteins.(13) In addition, these phosphoproteins contain the tripeptide sequence Arg-Gly-Asp (RGD) common to cell attachment type proteins that bind the cell surface receptors called in- tegrins. ( 3 9 . u ) ) Integrins are expressed in osteoblasts and osteoclasts and are requisite for attachment of these cells to b ~ n e . ' ~ . ' ~ ) In a similar manner, osteopontin is ex- pressed in osteoblasts and osteoclasts. (14.41.43) Attachment, differentiation, and activation of osteoclasts were en- hanced in the presence of osteopontin and bone sialopro- tein in v i t r ~ , ( ~ - ' " ) and osteopontin has been localized to sites of osteoclast attachment in developing rat bone in vivo. ( I 7 ) Recent evidence suggests a relationship between the concentration of osteopontin and bone sialoprotein at osteoclast attachment sites and the ability of these cells to resorb bone.(47) Overall, these studies support the sugges- tion that osteopontin and bone sialoprotein influence cell- matrix interactions and activity. However, the high affinity of these proteins for calcium may also regulate the deposi- tion of mineral in bone matrix.'") The latter hypothesis is supported, in part, by reports localizing these NCPs to areas of mineralization in rat and chicken Therefore, the complex physicochemical properties of osteopontin and bone sialoprotein may produce multiple actions in bone formation and remodeling.
In adult human bone, bone sialoprotein and osteopontin were detected throughout the concentric lamellae of each osteon. Intense staining of these phosphoproteins was ob- served within the cement lines and in the matrix adjacent to Haversian canals. As noted on serial specimens stained with Goldner-Masson-trichrome (Fig. 3A), von Kossa's, or ali- zarin red (not shown), these regions of increased phospho- protein staining of the inner osteon are fully mineralized and d o not contain osteoid. In light of previous reports and our immunohistochemical findings, the following theoreti- cal model is suggested for the possible role of osteopontin and bone sialoprotein in bone remodeling (Fig. 4) and is based, in part, on the model described by H t ~ f f e r " ' ~ . ~ ~ . ' ~ ) At the beginning of a remodeling cycle, the osteopontin- and bone sialoprotein-rich inner surface of the matrix lin- ing the Haversian canal promotes osteoclast attachment'l') and activation (stage l ) , leading to the resorption phase (stage 2). The osteoclasts expose an acellular surface of hy- droxyapatite. Osteopontin and bone sialoprotein, derived from the cellular milieu and/or circulation, may bind to these denuded mineral surfaces (via acid rich residues, stage 3), as indicated by the intense staining of cement lines immunohistochemically (Fig. 2A and B). During the reversal phase (stage 4), preosteoblasts and osteoblasts are recruited and attach (via RGD-integrin complexes) to these
osteopontin-and bone sialoprotein-rich surfaces. Preosteo- blasts differentiate into osteoblasts (stage S ) , and matrix is deposited (stage 6; see Fig. 4).
Osteopontin and bone sialoprotein appear to be incor- porated into matrix at lower concentrations than those ob- served in cement lines. During the formative phase, lower levels of these phosphoproteins within the majority of the osteon might be sufficient to maintain osteoblast attach- ment yet reduce the likelihood of premature osteoclast at- tachment and resorption of bone within units that are not completed. During osteoid formation, as osteoblasts be- come distant from the mineralization front, the ability of osteopontin and bone sialoprotein t o provide a linkage be- tween mineral and osteoblasts may be limited. Accord- ingly, the presence of these proteins in the lamellae of oste- ons may reflect their suggested roles in mineraliza- tion.(12.'n1 A dual role for osteopontin in both bone cell at- tachment and matrix mineralization was previously sug- gested by In contrast, at the completion of a bone unit (stage 7), osteopontin and bone sialoprotein in- corporation increases. The possibility exists that these pro- teins are secreted by osteoblasts at higher rates at the com- pletion of the formative phase. Alternatively, these phos- phoproteins accumulate in the innermost matrix of the os- teon over time as a result of their high affinity for mineral. Regardless of their origin, osteopontin and bone sialopro- tein incorporated into mineralized bone adjacent to com- pleted Haversian canals would provide attachment sites for osteoclasts at the beginning of the next remodeling cycle. Perhaps osteons that are less mature and have less phos- phoprotein accumulated on the mineralized surface may be less susceptible to bone-resorbing activity. Although specu- lative, this model provides a means by which resorption and formation are coupled to yield organized, complete units of bone.
The pattern of distribution of osteocalcin, a 6 kD glu- tamic acid-containing protein, was similar to that of OPN and BSP in that increased immunostaining was noted within the cement lines (Fig. 2D). This suggests that during the reversal phase, osteocalcin may be incorporated into de novo matrix along with osteopontin and bone sialoprotein. Alternatively, the high affinity of osteocalcin for calcium and hydroxyapatite may cause it to bind to mineral sur- faces that are exposed by osteoclastic a c t i ~ i t y . ( ~ . ~ * ~ ~ ) In contrast to the distribution of osteopontin and bone sialo- protein within the osteon, a marked reduction in immuno- staining of osteocalcin and osteonectin suggests that the rate of incorporation of these proteins into matrix may de- cline as inner lamellae are formed. This may be associated with the progressive decrease in the rate of matrix deposi- tion that occurs as the Haversian system As a re- sult of reduced osteoblast activity, the synthesis and secre- tion of osteonectin and osteocalcin may decrease as well. Alternatively, there may be a delay in the incorporation of these NCPs into bone matrix. Osteocalcin inhibits mineral- ization and promotes bone resorption in ~ i t r o . ( ~ ~ . ' ' ) Since osteocalcin is chemotactic for osteoclasts,(ls) a reduction in its incorporation into inner lamellae may allow comple- tion of each bone remodeling unit prior to initiation of the next resorptive phase. Local factors produced by osteo-

HETEROGENEITY OF NCP IN HUMAN BONE 1025
FIG. 3. Immunohistochemical distribution of NCPs in normal adult male iliac crest bone. Sections were treated and stained as described in Materials and Methods. (B) Cortical bone stained using normal rabbit serum (conirol). (C) Single Haversian system in cortical bone immunostained with decorin. Positive immunostaining was present in the matrix adjacent to the Haversian canal and in the lacuna and canaliculi of osteocytes. Inset a t x loo0 (bar equal to 25 pm). (D-F) Trabecular bone immunostained for osteopontin D), osteonectin (E), and decorin (F). Bar equal to 100 pm.
(A) Goldner-Masson-trichrome staining of cortical bone.

1026 INGRAM ET AL.
FIG. 4. Theoretical model of the role of phosphoproteins (osteopontin and bone sialoprotein) in bone remodeling. State of remodeling: (1) activation/attachment of osteoclasts to OPN- and BSP-rich surfaces; (2) resorption; (3) increased binding/incorporation of OPN and BSP to exposed mineral surfaces; (4) preosteoblast recruitment and attachment to ce- ment lines (reversal); ( 5 ) preosteoblast differentiation and osteoblast recruitment; (6) bone formation; and (7) increased incorporation of OPN and BSP. Abbreviations: HC, Haversian canal; LC, lining cell; POCL, preosteoclast; OCL, osteoclast; OPN, osteopontin; BSP, bone sialoprotein; POB, preosteoblast; OB, osteoblast; CAP, capillary; CL, cement line. [Modified from Huffer.(49)]
blasts, such as transforming growth factor B, may progres- sively regulate the synthesis and secretion of osteocalcin in an autocrine or paracrine manner during the formation of a bone unit.(55-5')
Osteonectin, a 38 kD glycoprotein. is one of the most abundant noncollagenous matrix proteins in fetal bone. (9 )
Osteonectin has a high affinity for both type I collagen and hydroxyapatite and is thought to promote mineraliza- tion by stabilizing hydroxyapatite and providing sites for nucleation of the mineral phase. ('.lo) Although the possi- bility exists that osteocalcin and osteonectin are extracted specifically from the inner aspect of the osteon during decalcification and staining, this effect appears to be highly selective since the remaining NCPs were retained in these same regions of osteons in adjacent sections. Fur- thermore, many osteons stained for osteocalcin through- out the concentric lamellae. The similar patterns of distri- bution between osteonectin and osteocalcin within the osteon are interesting considering their presumptive yet paradoxical roles in regulating mineralization.
Decorin and biglycan are chondroitin sulfate proteogly- cans derived from separate genes. ( I 8 ) Unlike other bone matrix proteins, these structural proteins are present in many nonskeletal connective tissue matrices. Decorin is thought to be involved in the regulation of collagen fibril- logenesis."') Osteons that are rich in decorin (Fig. 2E) may reflect continued organization of collagen fibers at these sites. Recent evidence suggests decorin and other proteo- glycans bind local growth factors. (59.w0) Therefore, decorin
may sequester growth factors to specific sites within units of bone. Interestingly, the matrix surrounding Haversian canals was rich in decorin and cement lines were com- pletely devoid of immunoreactivity.
Although the function of biglycan is at present un- known, recent studies suggest that this protein may be im- portant in tissues undergoing morphogenesis and differen- t i a t i~n . (~ ' .~ ' ) In the present study using normal adult bone, biglycan was diffusely distributed throughout cortical and trabecular matrix in a manner distinct from that of other matrix proteins. In developing human skeletal tissues, dec- orin and biglycan were distributed in separate regions of both cartilage and bone.(l') However, in contrast to our findings in adult bone, biglycan, but not decorin, was lo- calized to osteocytes and canaliculi in fetal bone (Table 2). These findings suggest that the specific roles of these pro- teoglycans may differ depending on the developmental state of the tissue.
Our histologic findings provide evidence of anatomic and functional heterogeneity within individual bone-re- modeling units and provide further indirect evidence that NCPs may influence cell-matrix interactions. The compo- sition of cement lines with respect to NCPs differs from the composition of the inner Haversian matrix (OPN, BSP. and OC versus OPN, BSP, and DCN, respectively; see Table 2). The observed distinct distribution of certain NCPs at these two initial cell binding surfaces supports the proposal that these proteins may be partially responsible for mediating the attachment and activation of appropri-

HETEROGENEITY OF NCP IN HUMAN BONE 1027
ate cells (i.e., osteoblast versus osteoclast) at the appropri- ate stage (i.e., formation versus resorption) in the bone-re- modeling cycle. The ability of certain NCPs t o bind and incorporate growth factors into bone matrix suggests that NCPs in conjunction with growth factors might influence bone unit remodeling by regulating the migration. differ- entiation, attachment, and activity of bone cells.('*~59~60)
Evidence suggests that during osteoblast differentiation, the expression of various matrix proteins follows a tempo- ral sequence.(6' These observations are consistent with im- munohistochemical and in situ hybridization studies of de- veloping bone in the rat and human.(',''.*') In light of these results, it appears that not only are NCPs expressed at dis- tinct stages of osteoblast maturation but that the produc- tion and incorporation of each of these proteins into mat- rix during normal bone remodeling may vary depending on the stage of formation of individual units of bone. Com- positional differences in NCPs within bone may reflect a regulatory mechanism whereby extracellular matrix influ- ences cell dynamics and consequently the remodeling and turnover of bone. These findings provide insight as to the function of NCPs and support the suggestion that these proteins play an integral role in both the architecture and remodeling of normal adult bone.
REFERENCES
I .
2.
3.
4.
5 .
6.
7.
8.
9.
10.
Fisher LW, Hawkins GR, Tuross N, Termine JD 1987 Puri- fication and partial characterization of small proteoglycans 1 and I I , bone sialoproteins 1 and 11, and osteoiiectin from the mineral compartment of developing human bone. J Biol Chem M2:9702-9708. Hauschka PV, Lian JB, Gallop PM 1975 Direct identifica- tion of the calcium binding amino acid, gamma-carboxyglu- tamate in mineralized tissue. Proc Natl Acaci Sci USA 72:
Termine JD, Kleinman HK, Whitson SW, Conn KM, Mc- Garvey ML, Martin GR 1981 Osteonectin, a bone-specific protein linking mineral to collagen. Cell M:W-IOS. Young MF, Day AA, Dominquez P, McQuillan CI, Fisher LW, Termine JD 1990 Structure and expression of osteonec- tin mRNA in human tissue. Connect Tiss Res 24:17-28. Fisher LW, Termine JD, Young MF 1989 Deduced protein sequence of bone small proteoglycan 1 (biglycan) shows ho- mology with proteoglycan I1 (decorin) and several noneon- nective tissue proteins in a variety of species. J Biol Chem
Stein GS, Lian JB, Owen TA 1990 Relaticinship of cell growth to the regulation of tissue-specific gene expression during osteoblast differentiation. FASEB J 4 3 1 11-3123. Weinreb M, Shinar D, Rodan GA 1990 Different pattern of alkaline phosphatase, osteopontin, and osteocalcin expres- sion in developing rat bone visualized by in situ hybridiza- tion. J Bone Miner Res 5:831-842. Robey PG, Bianco P, Termine JD 1992 The cellular biology and molecular biochemistry of bone formation. In: Coe FL, Favus MJ (eds.) Disorders of Bone and Mineral Metabolism. Raven Press, New York, pp. 241-263. Termine JD. Belcourt AB. Conn KM, Kleinnian HK 1981 Mineral and collagen binding proteins of fetal calf bone. J Biol Chem 256:10403-10408. Fisher LW. Termine JD 1985 Noncollagenous proteins influ-
3925-3929.
264:4571-4576.
encing the local mechanisms of calcification. Clin Orthop
11. Boskey A, Marsca M, Appel J 1989 The effects of noncollag- enous matrix proteins on hydroxyapatite formation and pro- liferation in a collagen gel system. Connect Tissue Res 21:
12. Glimcher MJ 1989 Mechanism of calcification: Role of colla- gen fibrils and collagen-hydroxyapatite formation and prolif- eration in a collagen gel system. Connect Tissue Res 21:171- 178.
13. Gorski JP 1992 Acidic phosphoproteins from bone matrix: A structural rationalization of their role in mineralization. Cal- cif Tissue Int 59:391-3%.
14. Lian JB, Tassinari M, Glowacki J 1984 Resorption of im- planted bone prepared from normal and warfarin-treated rats. J Clin Invest 73:1223-1226.
IS. Glowacki J, Lian JB 1987 lmpaired recruitment and differ- entiation of osteoclast progenitors by osteocalcin-deplete bone implants. Cell Differ 21:247-254.
16. Oldberg A, Franzen A, Heinegard D, Pierschbacher M, Ruoslahti E 1988 Identification of a bone sialoprotein recep- tor in osteosarcoma cells. J Biol Chem M3:19433-19436.
17. Reinholt FP, Hultenby K, Oldberg A, Heinegard D 1990 Osteopontin: A possible anchor of osteoclasts to bone. Proc Natl Acad Sci USA 87:4473-4475.
18. Rinklemen RD. Linkhart TA, Mohan S, Lau KHW, Baylink DJ, Bell NH 1991 Vitamin D deficiency causes a selective re- duction in deposition of transforming growth factor 6 in rat bone: Possible mechanism for impaired osteoinduction. Proc Natl Acad Sci USA 88:3657-3660.
19. Groessner-Schreiber B, Krukowski M, Lyons C, Osdoby P 1992 Osteoclast recruitment in response to human bone mat- rix is age related. Mech Ageing Dev 62:143-154.
20. Ninomiya JT, Tracy RP, Calore JD, Gendreau MA, Kelm RJ, Mann KG 1990 Heterogeneity of human bone. J Bone Miner Res 5933-938.
21. Vetter U, Fisher LW, Mintz JP, Tuross N, Termine JD, Robey PG 1991 Osteogenesis imperfecta: Changes in noncol- lagenous proteins in bone. J Bone Miner Res 6501-505.
22. Termine JD, Robey P Gehron. Fisher LW, Shimokawa H, Drum MA, Conn Kh4. Hawkins GR, Cruz JB. Thompson KG 1984 Osteonectin, bone proteoglycan and phosphoryn defects in a form of bovine osteogenesis imperfecta. Proc Natl Acad Sci USA 81:2213-2217.
23. Jundt G, Berghauser KH, Termine JD, Schulz A 1987 Osteo- nectin-a differentiation marker of bone cells. Cell Tissue Res 248:409-415.
24. Mark MP. Prince CW. Oosawa T, Gay S, Bronckers AL. Butler WT 1987 lmmunohistochemical demonstration of a 44-kD phosphoprotein in developing rat bones. J Histochem Cytochem 39707-715.
25. Bianco P. Silvestrini G, Termine JD, Bonucci E 1988 Immu- nohistochemical localization of osteonectin in developing hu- man and calf bone using monoclonal antibodies. Calcif Tis- sue lnt 43:155-161.
26. Bolander ME, Robey PG, Fisher LW. Conn KM. Prabhakar BS. Termine JD 1989 Monoclonal antibodies against osteo- nectin show conservation of epitopes across species. Calcif Tissue Int 45:74-80.
27. Bianco P. Fisher LW, Young MF, Termine JD, Robey PG 1990 Expression and localization of the two small proteogly- cans biglycan and decorin in developing human skeletal and non-skeletal tissues. J Histochem Cytochem 1: 1549-1563.
28. Bianco P, Fisher LW, Young MF, Termine JD, Robey PG 1991 Expression of bone sialoprotein (BSP) in developing hu- man tissues. Calcif Tissue Int 49421-426.
200:362-385.
171-178.

1028 INCRAM ET AL.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
Chen J , Zang Q, McCulloch CAG, Sodek J 1991 Immuno- histochemical localization of bone sialoprotein in foetal bone tissues: Comparisons with secreted phosphoprotein 1 (SPP- I , osteopontin) and SPARC (osteonectin). Histochem J 23:
McKee MD, Nanci A, Landis WJ, Gerstenfeld LC, Gotoh Y, Glimcher MJ 1989 Ultrastructural immunolocalization of a major phosphoprotein in embryonic chick bone. Connect Tissue Res 21:21-29. Hultenby K, Reinholt FP, Oldberg A, Heingard D 1991 Ultrastructual immunolocalization of osteopontin in meta- physeal and cortical bone. Matrix 11:206-213. Bonucci E, Reurink J 1978 The fine structure of decalcified cartilage and bone: A comparison between decalcification procedures performed before and after embedding. Calcif Tissue Res 25179-190. Goldners J 1937 A modification of the Masson trichrome technique for routine laboratory purposes. Am J Path 14:
McKee MD, Nanci A, Landis WJ, Gotoh Y , Gerstenfeld LC, Glimcher MJ 1991 Effects of fixation and demineralization on the retention of bone phosphoprotein and other matrix components as evaluated by biochemical analyses and quan- titative immunocytochemistry. J Bone Miner Res 6:937-945. VanGoor H, Harms G, Gerrits PO, Kroese FGM, Poppema S, Grond J 1988 lmmunohistochemical antigen demonstra- tion in plastic-embedded lymphoid tissue. J Histochem Cyto- chem 36:115-120. Bianco P, Hayashi Y, Silvestrini G. Termine JD, Bonucci E 1985 Osteonectin and GLA-protein in calf bone: Ultrastruc- tural immunohistochemical localization using the protein A-gold method. Calcif Tissue Int 37:684-686. Mark MP, Butler WT, Prince CW, Finkelman RD, Ruch JV 1988 Developmental expression of 44-Da bone phosphopro- tein (osteopontin) and bone gamma-carboxyglutamic acid (Gla)-containing protein (osteocalcin) in calcifying tissues of rat. Differentiation 37:123-136. Fisher LW, Lindner W, Young MF 1989 Synthetic peptide antisera: Their production and use in the cloning of matrix protein. Connect Tissue Res 21:43-50. Oldberg A, Franzen A, Heinegard D 1986 Cloning and se- quence analysis of rat bone sialoprotein (osteopontin) cDNA reveals in Arg-Gly-Asp cell-binding sequence. Proc Natl Acad Sci USA 83:8819-8823. Hynes RO 1987 Integrins: A family of cell surface receptors. Cell 48549-554. Horton MA, Davies J 1989 Perspectives: Adhesion receptors in bone. J Bone Miner Res 4:803-808. Franzen A, Heinegard D 1985 Proteoglycans and proteins of rat bone; purification and biosynthesis of major noncollage- nous macromolecules. In: Butler WT (ed.) The Chemistry and Biology of Mineralized Tissues. EBSCO Media, Birmingham,
Tezuka K, Sato T, Kamioka H, Nijweide PJ , Tanaka K, Matsuo T, Ohta M, Kurihara N, Hakeda Y, Kumegawa M 1992 Identification of osteopontin in isolated rabbit osteo- clasts. Biochem Biophys Res Commun 186:911-917. VanderPluijm G. Mouthaan H , Papapoulos S, Lowik C 1991 Inhibition of osteoclast precursor attachment and subsequent bone resorption by synthetic RGD-peptides. J Bone Miner Res 6:S146.
281-289.
237-243.
pp. 132-141.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
5 5 .
56.
57.
58.
59.
60.
Helfrich MH, Nesbitt SA, Dorey EL. Horton MA 1992 Rat osteoclasts adhere to a wide range of KGD (Arg-Gly-Asp) peptide-containing proteins, including the bone sialoproteins and fibronectin, via a 4, integrin. J Bone Miner Res 7:335- 343. Hruska KA. Rolnick F, Huskey M 1992 Occupancy o f the osteoclast 6,4, integrin by osteopontin stimlates a novel SRC associated phosphatidylinositol 3 kinase ( P I3 kinase) result- ing in phosphatidylinositol trisphosphate (PIP,) formation. J Bone Miner Res 7:S106. Blair HC, Lynch JA, Prince CW I992 Either osteopontin or bone sialoprotein are sufficient to reconstitute normal osteo- clastic degradation of protease stripped bone. J Bone Miner Res 75103. McKee MD, Nanci A, Landis WJ, Gotoh Y. Gerstenfeld 1.C. Glimcher MJ 1990 Developmental appearance and ultra- structural immunolocalization of a major 66 kDa phospho- protein in embryonic and post-natal chicken bone. Anat Rec
Huffer WE 1988 Biology of disease: Morphology and bio- chemistry of bone remodeling: Possible control by vitamin D. parathyroid hormone, and other substances. Lab Invest
Poser JW, Price PA 1979 A method for decarboxylation of gamma-carboxyglutamic acid in proteins: Properties of the decarboxylated gamma-carboxyglutamic acid from calf bone. J Biol Chem 254:431-436. Hauschka PV, Carr SA 1982 Calcium-dependent alpha-heli- cal structure of osteocalcin. Biochemistry 21:2538-2547. Delmas PD, Stenner D. Wahner HW. Mann KG, Riggs BL 1983 Increase in serum bone gamma-carboxyglutamic acid protein with aging in women. Implications for the mecha- nism of age-related bone loss. J Clin Invest 71:1316-1321. Amprino R, Engstrom A 1952 Studies on x-ray absorption and diffraction of bone tissue. Acta Anat (Basel) 1 5 1 . DeFranco DJ, Glowacki J , Cox KA, Lian JB 1991 Normal bone particles are preferentially resorbed in the presence of osteocalcin-deficient boine particles in v i v o . Calcif Tissue Int
Robey PG, Young MF, Flanders KC, Roche NS, Kondaiah P, Reddi AH, Termine J D , Sporn MB, Roberts AB 1987 Osteoblasts synthesize and respond to transforming growth factor-type 4 (TGFP) in vitro. J Cell Biol 105:457-463. Noda M 1989 Transcriptional regulation of osteocalcin pro- duction by transforming growth factor-p in rat osteoblast- like cells. Endocrinology 124:612-617. Oursler MJ, Cortese C, Keeling P, Anderson MA, Bonde SK. Riggs BI., Spelsberg TC 1991 Modulation of transform- ing growth factor-4 production in normal human osteoblast- like cells by 174-estradiol and parathyroid hormone. Endo- crinology 129: 1-8. McBride OW. Fisher 1.W, Young %IF 1990 Localization of PGI (biglycan, BGN) and PGII (decorin, I X N , PCi-40) genes on human chromosomes Xql3-qter and I2q, respec- tively. Genomics 6:219-225. Yamaguchi Y , Mann DM, Ruoslahti E 1990 Negative regula- tion of transforming growth factor+ by the proteoglycan decorin. Nature 346:281-284. Andres JL., Ronnstrand I., Cheifetz S, Massague J 1991 Pur - ification of the transforming growth factor+ (TGF-0) bind- ing proteoglycan betaglycan. J Biol Chem 266:23282-23287.
228~77-92.
S9:418-442.
49~43-50.

HETEROGENEITY OF NCP IN HUMAIS BONE
61. Fisher LW, Heegaard AM, Vetter U , Just W, Termine JD, Young MF 1991 Human biglycan gene: Puiative promoter, intron-exon junctions, and chromosomal localization. J Biol Chem 266: 14371-14377.
Address reprint requests to: L. A. Fitzpatrick, M. D.
Endocrine Research Unit 5-164 West Jclseph Building
Mayo Clinic and Mayo Foundalion Rochesrer, MN 55905
1029
Received in original form August 13, 1992; in revised form Janu- ary 5 , 1993; accepted February 6. 1993.




















