
Distribution, movements and diet of nocturnal fishes on temperate reefs
14
Distribution, movements and diet of nocturnal fishes on temperate reefs Danielle M. Annese a & Michael J. Kingsford b a School of Earth and Environmental Sciences, University of Wollongong, Wollongong, NSW 2522, Australia (e-mail: [email protected]) b School of Marine Biology & Aquaculture, James Cook University, Townsville, QLD 4811, Australia Received 6 February 2003 Accepted 12 April 2004 Key words: bullseyes, Pempherididae, diel patterns, tagging, site fidelity, plankton Synopsis We counted nocturnal fishes both day and night, and monitored the position of tagged individuals on temperate reefs in New South Wales, Australia. Pempheris affinis and P. multiradiata were the most abundant nocturnal planktivores on Sydney’s rocky reefs and showed great differences in diel migration behaviour. Both species were observed in deep shelter sites during the day (5–10 m), and most emerged into the water column at night. P. multiradiata was found to undergo extensive vertical and horizontal migrations. In contrast, P. affinis remained within daytime depth strata, with tagged individuals often moving less than 20 m at night. Tagged adult P. affinis returned to tagging sites for up to 7 weeks, indicating high site fidelity. Dietary analysis demonstrated that small and large pempherids differed in diet and the timing of foraging, suggesting a size-based transition from diurnal to nocturnal foraging. Stratified sampling of planktonic assemblages at different depths during the day and night showed spatial variation in the availability of prey items at different times of the day. Amphipods, the main prey of large fish, were only available during the night, and concentrated in shallow water, whereas decapod larvae, consumed mainly by small fish, were abundant day and night. Large P. affinis also fed on polychaetes, which were never found in the stomachs of P. multiradiata, suggesting that these species may have different prey require- ments, or that these polychaetes are only found in deep water where foraging P. affinis were abundant. We found no general model for the Pempheridae. The movements and behaviour of nocturnal fishes varied greatly by species, and this may be due to differences in body size, and/or physiological (e.g. visual ability) and ecological constraints. Introduction Historically, most research on reef fishes has fo- cussed on diurnal species, especially patterns of abundance, habitat preferences, patterns of recruitment and intra- and interspecific interac- tions (Sale 1991). Although nocturnal fishes have been counted on reefs (Lincoln Smith et al. 1991), they are invariably observed in daytime shelter sites and are often lumped into assemblage-based analyses (Sale 1978). Abundant nocturnal reef fishes include bullseyes (Pempherididae) and car- dinal fishes (Apogonidae). Although common inhabitants of coral and temperate reefs, little information is available on the behaviour and feeding ecology of these fishes. Casual observa- tions (Hobson 1974) and some quantitative data from tropical waters (Gladfelter 1979, Marnane & Bellwood 2002) suggest that there are great chan- ges in the distribution of pempherids and apogo- nids at night; some species are known to move distances of tens to hundreds of metres from daytime shelter. Gladfelter (1979) and Hobson (1972, 1973) have argued that nocturnal Environmental Biology of Fishes (2005) 72: 161–174 ȑ Springer 2005
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Distribution, movements and diet of nocturnal fishes on temperate reefs

Distribution, movements and diet of nocturnal fishes on temperate reefs
Danielle M. Annesea & Michael J. KingsfordbaSchool of Earth and Environmental Sciences, University of Wollongong, Wollongong, NSW 2522, Australia(e-mail: [email protected])bSchool of Marine Biology & Aquaculture, James Cook University, Townsville, QLD 4811, Australia
Received 6 February 2003 Accepted 12 April 2004
Key words: bullseyes, Pempherididae, diel patterns, tagging, site fidelity, plankton
Synopsis
We counted nocturnal fishes both day and night, and monitored the position of tagged individuals ontemperate reefs in New South Wales, Australia. Pempheris affinis and P. multiradiata were the mostabundant nocturnal planktivores on Sydney’s rocky reefs and showed great differences in diel migrationbehaviour. Both species were observed in deep shelter sites during the day (5–10 m), and most emerged intothe water column at night. P. multiradiata was found to undergo extensive vertical and horizontalmigrations. In contrast, P. affinis remained within daytime depth strata, with tagged individuals oftenmoving less than 20 m at night. Tagged adult P. affinis returned to tagging sites for up to 7 weeks,indicating high site fidelity. Dietary analysis demonstrated that small and large pempherids differed in dietand the timing of foraging, suggesting a size-based transition from diurnal to nocturnal foraging. Stratifiedsampling of planktonic assemblages at different depths during the day and night showed spatial variation inthe availability of prey items at different times of the day. Amphipods, the main prey of large fish, were onlyavailable during the night, and concentrated in shallow water, whereas decapod larvae, consumed mainlyby small fish, were abundant day and night. Large P. affinis also fed on polychaetes, which were neverfound in the stomachs of P. multiradiata, suggesting that these species may have different prey require-ments, or that these polychaetes are only found in deep water where foraging P. affinis were abundant. Wefound no general model for the Pempheridae. The movements and behaviour of nocturnal fishes variedgreatly by species, and this may be due to differences in body size, and/or physiological (e.g. visual ability)and ecological constraints.
Introduction
Historically, most research on reef fishes has fo-cussed on diurnal species, especially patterns ofabundance, habitat preferences, patterns ofrecruitment and intra- and interspecific interac-tions (Sale 1991). Although nocturnal fishes havebeen counted on reefs (Lincoln Smith et al. 1991),they are invariably observed in daytime sheltersites and are often lumped into assemblage-basedanalyses (Sale 1978). Abundant nocturnal reeffishes include bullseyes (Pempherididae) and car-
dinal fishes (Apogonidae). Although commoninhabitants of coral and temperate reefs, littleinformation is available on the behaviour andfeeding ecology of these fishes. Casual observa-tions (Hobson 1974) and some quantitative datafrom tropical waters (Gladfelter 1979, Marnane &Bellwood 2002) suggest that there are great chan-ges in the distribution of pempherids and apogo-nids at night; some species are known to movedistances of tens to hundreds of metres fromdaytime shelter. Gladfelter (1979) and Hobson(1972, 1973) have argued that nocturnal
Environmental Biology of Fishes (2005) 72: 161–174 � Springer 2005

movements relate to the abundance of prey andavailability of shelter. Therefore, descriptions ofdistribution patterns and the processes that influ-ence these should focus on several aspects includ-ing: movements within reefs and adjacent habitats(i.e. availability of shelter and habitats with anabundance of prey), dietary preferences and thedistribution of prey.Nocturnal species such as pempherids and
apogonids tend to be cryptic, sheltering in holesand crevices during the day (Greenfield & Johnson1990, Mooi 2000). This may be associated with ahigh degree of site specificity resulting in fisheseither returning to the same shelter site after for-aging (Apogonidae: Marnane & Bellwood 2002) ormaintaining a limited home-range during the day(Cheilodactylidae: Lowry & Suthers 1998). Evi-dence from some tropical pempherids indicatesthat movement patterns are often highly predict-able in space and time (Gladfelter 1979, Mooi2000). Although studies have used external tagsand radiotelemetry to track the movements ofdiurnal fishes (Matthews 1992, Lowry & Suthers1998), there are few data on the distribution andmovements of individual fish, or the long-termcontinuity of shelter sites. Furthermore, no datahas been collected at more than one site to deter-mine the generality of movements and compari-sons among taxa.Patterns of movement and distribution of fishes
commonly vary according to fish size, but arepoorly described for nocturnal planktivores. Al-though it is generally accepted that nocturnalspecies shelter during the day and emerge at night,ontogenetic differences in the patterns of move-ment and behaviour of fishes are common onrocky reefs (Jones 1988). Juveniles have been ob-served remaining closer to shelter sites duringmigration (Hobson 1973) and in doing so, they areless often found high in the water column (Platell& Potter 1999). Furthermore, juveniles of a speciesare sometimes diurnal, becoming nocturnal duringthe adult life history stage (Lockett & Suthers1998, Mooi 2000). Hence, size and species-basedcomparisons are crucial in descriptions of theecology of nocturnal fishes.The patchy distribution of fishes in rocky reef
environments can often be related to resourcepartitioning. This may include habitat preferenceor specificity (Greenfield & Johnson 1990), com-
bined with different preferred prey (Platell & Pot-ter 1999). Nocturnal fishes commonly consumedemersal zooplankters that are undergoing noc-turnal excursions from the substratum, holo-plankton and some small fishes (Russell 1983,Hobson 1974, Gladfelter 1979, MacDiarmid1981). Demersal zooplankters typically emergefrom the substratum shortly after dark and des-cend to the substratum before dawn, however, theexact timing may vary according to taxa, size, sexand season (Alldredge & King 1980, Jacoby &Greenwood 1988). The availability and quantity ofzooplanktonic organisms as a source of prey may,therefore, have a strong influence on distributionand behaviour. Furthermore, information on thefeeding behaviour of nocturnal fishes may increaseour knowledge of reef trophodynamics in that theymay consume food at great distances from theirshelter sites and defecate in daytime shelters. Ananalogous situation is found for diurnal plankti-vores (e.g. temperate Chromis species) that forageconsiderable distances from night shelter sites(Bray et al. 1981, Kingsford & MacDiarmid 1988).The objectives of this study were to describe the
distribution and movements of the most abundantnocturnal planktivores on Sydney’s rocky reefs,and explore the potential role of feeding and thedistribution of prey for influencing behaviour. Ourspecific aims were to: (1) describe depth relatedpatterns of abundance at spatial scales of hundredsof metres to kilometres, and from this infer pat-terns of movement; (2) determine the site fidelityand movements of pempherids at time scales ofdays to weeks by tagging individuals; (3) describethe diet of pempherids of different sizes; and (4)determine the availability of prey with time of dayand depth.
Materials and methods
Study sites
Most study sites were located in and around Bot-any Bay, New South Wales, Australia (Figure 1).Botany Bay (34� 01¢S, 151� 11¢ E) is a semi-land-locked shallow estuary with a surface area ofapproximately 42 km2 (Steffe & Pease 1988). Thedomination of the bay by ocean swell and windwaves, as well as the natural topography leads to
162

both exposed and relatively sheltered locationsnear the entrance and within the bay. Locationswere separated by hundreds of metres to kilome-tres. Sandstone reefs were sampled at Henry Head,Cape Banks, Bare Island (southern end), Suther-land Point, Inscription Point and North Bondi(north of Botany Bay) (Figure 1).
Fish abundance by depth and habitat
We hypothesised that the diel activities of noc-turnal fishes alter patterns of relative abundanceamong depths during the day and night. Depthand diel patterns of abundance were determinedusing depth stratified sampling at six locationsduring the day and night. Nocturnal fishes werecounted in five non-overlapping 5 · 20 m randombelt transects in each depth range: deep (10–14 m),mid (4–10 m) and shallow (<4 m). Optimal tran-sect length and number were determined by a pilotstudy where we tested 10, 15, 20, 25 and 30 mlengths (Annese 2000). Counts were made onSCUBA as the tape was unreeled to minimise
fright response. A 5 m transect width was main-tained by calibrating the eye of observers beforeeach replicate transect by running out the tape to2.5 m. Thorough searches were made of all cre-vices and ledges within the belt transects. Inaddition, the locations of fishes were recorded, i.e.whether in caves, associated with the substratumor in the water column. Fishes were defined asbeing in the water column if they were 20 cmabove the substratum. This procedure was re-peated again on the same day approximately 1–3 hafter sunset to allow time for twilight to nocturnalmigrations (Gladfelter 1979). We counted fishes atnight by torch light. They were easy to count atnight and were generally diver neutral (i.e. theywere not startled or frightened away by the light).We analyzed data with a mixed-model Analysis ofVariance (ANOVA). Depth (treatments: shallow,mid, deep) and time of day (treatments: day, night)were treated as fixed factors and location as ran-dom.We estimated the standard length of all pem-
pherids. Small fishes were those with a standard
Figure 1. Location of sampling sites around Botany Bay, Sydney: BI – Bare Island, CB – Cape Banks, SP – Sutherland Point, IP –
Inscription Point, HH – Henry Head, and North Bondi (above).
163

length less than 55 mm. Medium fishes were be-tween 55 and 100 mm, and large fishes were largerthan 100 mm. No individual from the three specieswas found to have a larger standard length than190 mm. Collections of fishes allowed verificationof the accuracy of these estimates. We also exam-ined temporal variation in diel patterns at KurnellPoint. A detailed map of Kurnell Point was com-pleted so that the positions of fishes could be re-corded. Thorough searches of the mapped area(50 · 80 m and depth range 2–10 m) wereconducted on nine occasions during the day, andthen repeated on the same night 2 h after sunset.This was achieved by counting fishes along lanesparallel to the shore that traversed the entiremapped area, and plotting fish locations. Weestimated the standard lengths of all fish, as well asthe habitats in which they were found.We hypothesised that diel and depth-related
patterns would remain consistent among times.Non-overlapping transect data were extractedfrom the map on nine occasions over a 7 weekperiod. We analysed data by a mixed modelANOVA with time of day and depth treated asfixed factors, and times as random.
Site fidelity
We observed nocturnal fishes (P. affinis andP. multiradiata) in caves and crevices during theday. We anesthetised fishes from four discrete cavesites (n ¼ 10 individuals from each site) with a 1:5mix of clove oil to ethanol and tagged them usingvisible implant fluorescent elastomer (NorthwestMarine Technology Inc., USA), injected sub-cutaneously to the caudal peduncle. We tagged allfishes on both sides of the body to enable detectionwhen viewed from either side. They were recogni-sable at a cave-specific level by position and colourof tags. Pempherids are relatively translucent sosuch tags were clearly visible by eye during theday, and with the aid of a UV light at night.We held tagged individuals in a recovery cage
for between 10 and 40 min. This allowed fishes torecover from the tagging procedure, therebyreducing the risk of predation once released. Wemonitored all fishes for a 10 min period followingrelease to ensure they survived the procedure andwere integrated back into the school.
We monitored tagging sites for returns of taggedindividuals the day after tagging, and then once aweek thereafter during the day and night to detectany differences in the location of tagged individu-als. We monitored Sites 1 and 2 for 7 weeks, andSites 3 and 4 for 6 weeks. On each occasion, wecompleted a search of concentric circles aroundeach site, commencing at a radius of 2 m andextending to 10 m in 2 m increments. Addition-ally, we completed a search of the entire mappedarea on each occasion to monitor any longer dis-tances covered by individuals. The location oftagged individuals relative to their cave of taggingwas plotted on a map of the study area, as well asrecorded in terms of distance.
Diet and availability of prey
We hypothesised that the diet of pempheridswould differ with fish size. We collected small andlarge P. affinis (n ¼ 22) and P. multiradiata(n ¼ 19) during the day and night from severalsites including Cape Banks, Henry Head, Suther-land Point, Bare Island and Inscription Point. Wecollected all fishes while SCUBA diving in depthsless than 14 m. We collected fishes with a standardlength smaller than 40 mm using hand nets, whilewe collected those greater than this using a mul-tipronged hand spear. We injected fishes with 10%formalin soon after collection to reduce deterio-ration of the prey prior to dissection. Final pres-ervation was in 5% formalin.We counted individual prey items and expressed
them as a numerical percentage. We also estimatedthe percentage representation of prey items in eachstomach using the grid method (Jones 1968). Wecalculated the percent occurrence of prey items asthe percentage of fishes that had consumed eachtype of prey. We measured size of prey itemsacross the greatest distance, excluding appendages.We hypothesised that the composition of
planktonic assemblages would vary between dayand night, and between shallow and deep water,and that these patterns would be consistent amongdays. We sampled zooplankton on three occasionsduring the day and night at Kurnell Point usingthree non-overlapping depth integrated replicatetows in shallow (from 2 m deep) and deep (from10 m deep) water. We examined two subsamplesfrom each plankton tow, and identified, counted
164

and measured all organisms. We analysed plank-ton data using a 3-factor mixed model ANOVA(factors depth and time of day were treated asfixed, and times as random). Analyses were onlydone for abundant taxa.
Results
Assemblage of nocturnal planktivores
We observed four species of nocturnal plankti-vores in fish counts at six locations (totaln ¼ 2497). Pempheris affinis and P. multiradiataranked first and second in total abundance(Table 1). Apogon limenus ranked third and rarespecies included P. compressa. Species richness andabundance of all nocturnal planktivores wasgreatest at mid- and deep-depth strata. There werediel changes in depth-related patterns of distribu-tion of some species. For example, P. multiradiatawere more abundant in shallow water at night(Table 1). Most nocturnal fishes were in crevicesduring the day (89%), and almost all fishes werefound out of crevices at night (92%).
Fish abundance by depth and habitat
Pempheris affinis was found at all locations duringthe day and night, and we observed 94% of the fishin mid- and deep strata, regardless of time of day(n ¼ 1644). We found a significant interactionbetween depth and time of day (Table 2) as rankabundance changed between depths both day andnight. The magnitude of day/night differences alsovaried with location resulting in a significantinteraction between depth and location (Table 2).
Diel patterns of distribution and abundancevaried for fish of different size classes. For example,we observed most small and medium fish (72%), indeep strata regardless of time of day. We foundgreat differences in diel patterns of abundanceamong depths for P. multiradiata (n ¼ 540). Dur-ing the day, they were only found at mid and deepstrata. In contrast, 63% of fish counted at nightwere found in shallow water. Differences in rankabundance between depths at day and night, aswell as differences in total abundance at differentlocations (Figure 2), resulted in a three-way inter-action between depth, time of day and location(Table 2). Patterns of distribution found at BareIsland and North Bondi suggested that P. multi-radiata had moved laterally into the locationsduring the night as few residents were found duringthe day (Figure 2). No small P. multiradiata werefound during sampling at the six locations.Although the vast majority of nocturnal fishes in
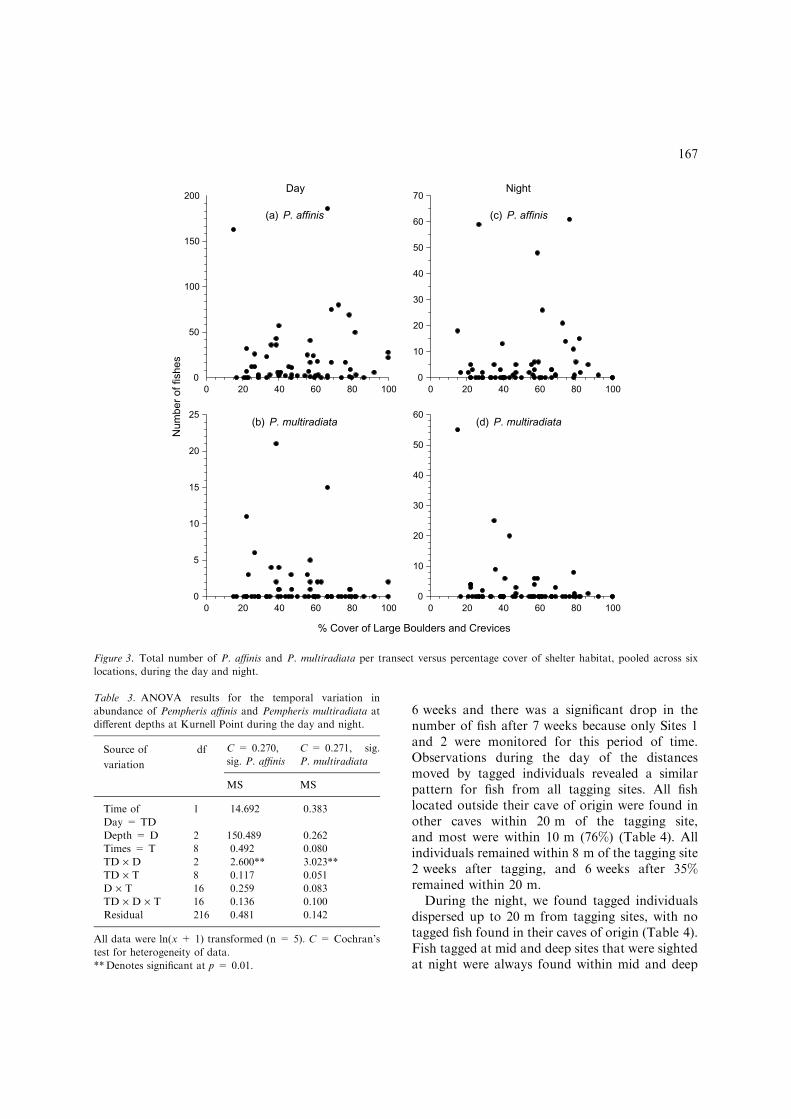
all parts of the study were observed in, or close to,shelter sites during the day, we found little evi-dence for a relationship between the abundance ofnocturnal fishes and the percentage cover of hab-itat with adequate shelter (Figure 3). Most specieswere found in aggregations and this resulted inhigh variance among replicate transects (25–100%of the mean).
Temporal variation in distribution and abundanceamong depths
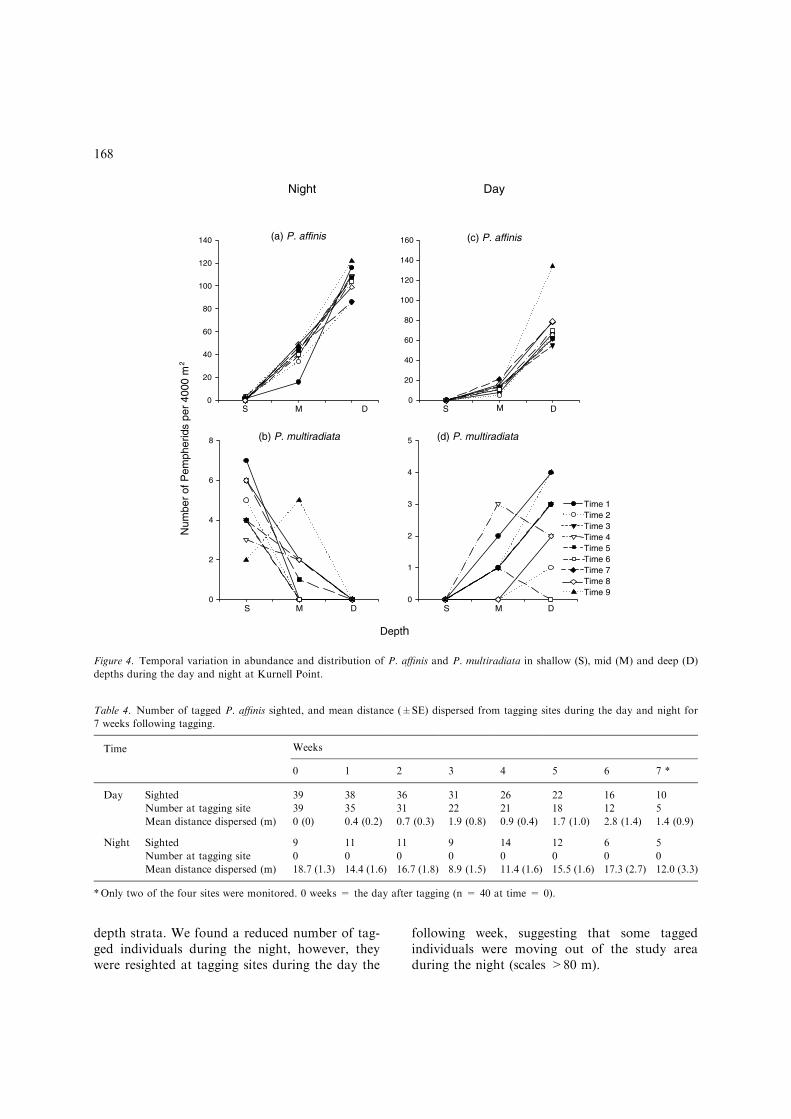
Temporal diel and depth related patterns of dis-tribution and abundance differed notably betweenP. affinis and P. multiradiata. We found a signifi-cant interaction between depth and time of day forboth species (Table 3). The abundance of P. affiniswas greatest in deep water during the day and
Table 1. Mean abundance (±SE) of nocturnal planktivorous fishes (per 25 m2) in shallow, mid and deep depths, at six locations,
during the day and night (n = 2497).
Species Day Night Total Abundance Rank
Shallow Mid Deep Shallow Mid Deep
Pempheris affinis 8.6 (6.4) 31.5 (24.1) 85.7 (22.6) 15.2 (12.1) 28.4 (6.2) 62.2 (17.3) 213.6 (88.7) 1
Pempheris multiradiata 0 5.1 (3.2) 4.5 (2.2) 57.4 (7.5) 10.0 (6.8) 18.2 (16.2) 95.2 (35.9) 2
Apogon limenus 2.1 (1.2) 3.8 (2.6) 10.0 (6.4) 0 5.5 (4.3) 9.0 (8.2) 30.4 (22.7) 3
Pempheris compressa 0 4.4 (3.0) 7.2 (6.4) 0 4.0 (1.8) 4.7 (3.5) 20.3 (14.7) 4
Total abundance 10.7 (7.6) 44.8 (32.9) 107.4 (37.6) 72.6 (19.6) 47.9 (19.1) 94.1 (45.2)
165

abundance was generally higher at mid depthsduring the night on all sampling occasions atKurnell Point (Figure 4a and c). The greatestabundance of P. multiradiata was also in deepwater during the day, but the opposite was true atnight, with the majority of fish being found inthe shallows on all but one occasion (Figure 4b andd).
Site fidelity of Pempheris affinis
Site fidelity was high for tagged P. affinis on a scaleof days, and at longer time scales (months). Wepooled results due to similar patterns at all taggingsites. All tagged fish except one were present in thecave of capture on the day following tagging(week ¼ 0, Table 4). Most of the tagged fish werefound at tagging sites after 2 weeks (78%). Thisnumber progressively decreased over the following5 weeks, however, 53% of tagged fish remained attagging sites after 1 month. We found 30% after
Table 2. ANOVA results for the total numbers of Pempheris
affinis and Pempheris multiradiata at different depths at six
locations during the day and night.
Source of variation df C = 0.268,
sig. P. affinis
C = 0.253,
sig. P. multi-
radiata
MS MS
Time of
day = TD
1 14.362 9.614
Location = L 5 1.904 2.664
Depth = D 2 39.949 0.319
TD · L 5 0.886 5.065
TD · D 2 6.811* 8.062
L · D 10 2.781* 3.025
TD · L · D 10 1.236 1.257**
Residual 144 1.226 0.491
All data were ln(x + 1) transformed (n = 5). C = Cochran’s
test for heterogeneity of data.
*Denotes significant at p < 0.05, ** Significant at p < 0.01.
0
5
10
15
20
25
30
35
40
Cape Banks Henry Head SutherlandPoint
InscriptionPoint
Bare Island North Bondi
0
5
10
15
20
25
Cape Banks Henry Head SutherlandPoint
Location
InscriptionPoint
Bare Island North Bondi
DayNight
Mea
n nu
mbe
r of P
emph
erid
s pe
r 100
m2
(a) P.affinis
(b) P. multiradiata
Figure 2. Mean abundance of pempherids during the day and night at six locations.
166
6 weeks and there was a significant drop in thenumber of fish after 7 weeks because only Sites 1and 2 were monitored for this period of time.Observations during the day of the distancesmoved by tagged individuals revealed a similarpattern for fish from all tagging sites. All fishlocated outside their cave of origin were found inother caves within 20 m of the tagging site,and most were within 10 m (76%) (Table 4). Allindividuals remained within 8 m of the tagging site2 weeks after tagging, and 6 weeks after 35%remained within 20 m.During the night, we found tagged individuals
dispersed up to 20 m from tagging sites, with notagged fish found in their caves of origin (Table 4).Fish tagged at mid and deep sites that were sightedat night were always found within mid and deep
(b) P. multiradiata
0 20 40 60 80 1000
5
10
15
20
25(d) P. multiradiata
0 20 40 60 80 1000
10
20
30
40
50
60
(a) P. affinis
0 20 40 60 80 100
Num
ber o
f fis
hes
0
50
100
150
200
(c) P. affinis
0 20 40 60 80 1000
10
20
30
40
50
60
70Day Night
% Cover of Large Boulders and Crevices
Figure 3. Total number of P. affinis and P. multiradiata per transect versus percentage cover of shelter habitat, pooled across six
locations, during the day and night.
Table 3. ANOVA results for the temporal variation in
abundance of Pempheris affinis and Pempheris multiradiata at
different depths at Kurnell Point during the day and night.
Source of
variation
df C = 0.270,
sig. P. affinis
C = 0.271, sig.
P. multiradiata
MS MS
Time of
Day = TD
1 14.692 0.383
Depth = D 2 150.489 0.262
Times = T 8 0.492 0.080
TD · D 2 2.600** 3.023**
TD · T 8 0.117 0.051
D · T 16 0.259 0.083
TD · D · T 16 0.136 0.100
Residual 216 0.481 0.142
All data were ln(x + 1) transformed (n = 5). C = Cochran’s
test for heterogeneity of data.
**Denotes significant at p = 0.01.
167
depth strata. We found a reduced number of tag-ged individuals during the night, however, theywere resighted at tagging sites during the day the
following week, suggesting that some taggedindividuals were moving out of the study areaduring the night (scales >80 m).
(a) P. affinis
0
20
40
60
80
100
120
140 (c) P. affinis
0
20
40
60
80
100
120
140
160
(b) P. multiradiata
Depth
Num
ber
ofP
emph
erid
spe
r40
00m
2
0
2
4
6
8 (d) P. multiradiata
0
1
2
3
4
5
Night Day
S M D S M D
S M DS M D
Time 1Time 2Time 3Time 4Time 5Time 6Time 7Time 8Time 9
Figure 4. Temporal variation in abundance and distribution of P. affinis and P. multiradiata in shallow (S), mid (M) and deep (D)
depths during the day and night at Kurnell Point.
Table 4. Number of tagged P. affinis sighted, and mean distance (±SE) dispersed from tagging sites during the day and night for
7 weeks following tagging.
Time Weeks
0 1 2 3 4 5 6 7 *
Day Sighted 39 38 36 31 26 22 16 10
Number at tagging site 39 35 31 22 21 18 12 5
Mean distance dispersed (m) 0 (0) 0.4 (0.2) 0.7 (0.3) 1.9 (0.8) 0.9 (0.4) 1.7 (1.0) 2.8 (1.4) 1.4 (0.9)
Night Sighted 9 11 11 9 14 12 6 5
Number at tagging site 0 0 0 0 0 0 0 0
Mean distance dispersed (m) 18.7 (1.3) 14.4 (1.6) 16.7 (1.8) 8.9 (1.5) 11.4 (1.6) 15.5 (1.6) 17.3 (2.7) 12.0 (3.3)
*Only two of the four sites were monitored. 0 weeks = the day after tagging (n = 40 at time = 0).
168

Diet
The main prey consumed by small and largepempherids were amphipods, decapods and os-tracods, and we found similar prey items amongcollection days. Relative proportions of these,however, differed between fish size classes withinspecies. Dietary evidence corroborated withobservations of juvenile pempherids feeding dur-ing the day. Juvenile pempherids collected duringthe day were found to have a greater variety andabundance of prey items in the stomach than thosecollected at night. The main prey items consumedby small pempherids of both species were mero-planktonic decapods (especially crab zoel stages),representing 47.5% of the stomach contents ofsmall P. affinis collected during the day and 49.3%in those collected at night (Table 5a). Similarly,they represented 32.6% of the stomach contents ofsmall P. multiradiata collected during the day and40.0% for those collected at night (Table 6a).Other prey items consumed in abundance by smallpempherids were calanoid copepods, ostracodsand amphipods.For both species, the proportion of demersal
amphipods consumed was greatest for large indi-
viduals, contributing to 33.9% representation inthe stomachs of large P. affinis collected at night(Table 5b) and 41.8% in P. multiradiata specimens(Table 6b). In addition, the variety and abundanceof prey items in the stomachs of large P. multi-radiata were significantly greater during the nightwhen fish were observed foraging outside of theirday time caves. In addition to demersal amphi-pods, these fish also consumed a variety of mero-planktonic organisms such as anomuran zoea,crab megalopa and the occasional holoplanktoniccopepod Paracalanus sp. (Table 6b). Although themajority of prey items consumed by both smalland large pempherids were <5 mm in size, therewas greater variation in the size of prey consumedby large pempherids of both species.
Distribution of prey
Analysis of plankton tows revealed a clear dis-tinction between demersal and meroplanktonicorganisms (Table 7), with some plankton presentin greater numbers during the day (copepod, Te-mora sp.) or night (amphipods). Others occurredin similar concentrations during both day and
Table 5. Stomach contents of small (n = 10) and large (n = 12) P. affinis collected during the day and night.
Prey item Day Night
Size (mm) mean
(range)
Mean numerical
abundance
(±SE)
Mean%
representation
(±SE)
Size (mm) mean
(range)
Mean numerical
abundance
(±SE)
Mean% repre-
sentation
(±SE)
(a) Small ðSL 29–55 mmÞParacalanus spp. 0.8 (0.2–1.2) 5.2 (2.2) 17.7 (10.5) – – –
Decapod fragment 1.2 (0.3–4.1) 21.0 (8.4) 47.5 (12.9) 1.2 (0.2–2.4) 5.0 (2.3) 49.3 (13.6)
Amphipod 1.3 (0.3–2.3) 3.0 (0.4) 11.5 (1.1) 1.3 (0.3–2.4) 1.2 (0.7) 26.7 (11.3)
Ostracod 1.0 (0.4–2.2) 3.2 (2.7) 16.7 (11.7) 1.2 (1.2–2.2) 0.2 (0.2) 2.0 (2.0)
Other 1.2 (0.3–3.5) 0.6 (0.2) 2 (1.9) 5.3 1.0 (1.0) 2 (2.0)
(b) Large ðSL 108–152 mmÞTemora sp. 1.6 (0.4–0.9) 3.4 (3.2) 9.5 (6.6) 0.8 (0.4–1.8) 1.7 (1.1) 1.9 (1.9)
Oncaea sp. 0.6 (0.4–1.0) 2.4 (1.3) 10.7 (6.9) 0.9 (0.5–1.2) 0.3 (0.3) <0.1
Decapod fragment 1.7 (0.9–2.8) 10.0 (7.6) 11.3 (7.6) 3.8 (0.4–6.8) 20.0 (6.5) 14.0 (6.1)
Amphipod 2.7 (0.7–3.5) 1.2 (1.0) <0.1 2.5 (0.5–4.8) 34.6 (10.6) 33.9 (6.2)
Ostracod 0.7 (0.4–1.3) 8.0 (3.6) 32.8 (14.0) 1.6 (1.2–1.9) 1.7 (0.7) 1.8 (1.1)
Isopod 1.0 (0.8–1.2) 0.8 (0.4) 2.9 (2.9) 1.9 (0.9–3.9) 6.0 (3.2) 6.4 (3.2)
Nemertia 4.8 (1.8–6.5) 0.8 (0.8) 0.3 (0.3) 8.7 (2.6–2.0) 9.7 (9.1) 9.6 (6.7)
Polychaete 2.5 (2.4–2.5) 0.6 (0.4) 27.9 (14.4) 16.0 (6.2–29.0) 2.0 (0.8) 14.8 (8.8)
Anomuran zoea – – – 4.2 (2.8–6.8) 4.7 (2.3) 9.0 (6.2)
Crab megalopa – – – 8.5 (3.4–13.2) 0.9 (0.4) 6.8 (3.0)
Other 0.8 (0.2–1.0) 0.7 (0.4) 1 (0.7) 2.8 (1.2–8.5) 0.6 (0.2) 1 (0.4)
169

night, for example, crab larvae (brachyuran zoeaand crab megalopa).Amphipods were often responsible for composi-
tional differences between day and night. Clear dielpatterns of distribution and abundance of amphi-pods between depths during the day and night wereevident across all three sampling times and thisresulted in a significant interaction between time ofday and depth (Table 8). Amphipods were demer-sal with only negligible numbers inhabiting thewater column during the day, and great increases in
concentration occurring at night. Highest concen-trations were found in shallow water.Conversely, meroplanktonic decapods were
found in the water column during the day and nightand among times, however, the abundance wasgreatest during the night. Although inhabiting bothdepths, decapods were collected in greatest con-centrations in deep water during the day, and theyhad a more variable distribution evident at night.
Table 6. Stomach contents of small (n = 9) and large (n = 10) P. multiradiata collected during the day and night.
Prey Item Day Night
Size (mm)
mean (range)
Mean numeri-
cal abundance
(±SE)
Mean%representation
(±SE)
Size (mm)
mean (range)
Mean numeri-
cal abundance
(±SE)
Mean%representation
(±SE)
ðaÞ Small ðSL 37–54 mmÞDecapod fragment 0.9 (0.2–2.3) 8.4 (2.6) 32.6 (9.1) 1.1 (0.4–1.2) 3.8 (2.5) 40.0 (17.2)
Amphipod 1.3 (0.4–2.5) 5.6 (1.9) 21.0 (8.0) 0.9 (0.2–1.6) 2.5 (1.6) 17.2 (10.3)
Ostracod 0.7 (0.2–1.4) 13.6 (5.9) 41.7 (12.2) 0.6 (0.4–0.9) 0.8 (0.5) 0.3 (0.2)
Gastropod fragment – – – 2.0 0.3 (0.3) 12.5 (12.5)
Other 0.8 (0.2–1.2) 0.8 (0.2) 1 (0.6) 0.9 (0.7–1.2) 0.4 (0.3) <0.1
ðbÞ Large ðSL 105–155 mmÞDecapod fragment 1.9 (1.4–2.8) 1.8 (1.6) 6.0 (6.0) 2.9 (0.9–6.8) 9.6 (4.0) 11.8 (5.2)
Amphipod 1.7 (0.9–2.8) 0.6 (0.6) <0.1 2.5 (0.7–4.7) 31.0 (6.4) 41.8 (10.2)
Ostracod – – – 1.4 (0.7–1.2) 1.2 (0.2) 4.5 (1.4)
Isopod – – – 1.8 (0.7–2.5) 3.6 (2.1) 5.5 (2.8)
Nemertia 6.4 (3.4–8.2) 2.6 (2.4) 34.0 (21.4) 9.8 (3.2–34.2) 7.0 (6.5) 10.0 (7.8)
Anomuran zoea – – – 3.1 (0.4–8.9) 6.4 (2.8) 15.6 (10.5)
Crab megalopa – – – 1.6 (1.6–2.8) 0.6 (0.4) 6.0 (3.9)
Other 0.1 0.2 (0.2) < 0.1 2.5 (1.1–4.2) 0.7 (0.3) 2 (0.8)
Table 7. Comparison of the mean number of main prey
plankters per m3 during the day and night at Kurnell Point
(data pooled for 3 days and nights; n = 18).
Taxa Number of individual plankters per m3
Day Night
Mean ±SE Mean ±SE
Ostracod 3.5 1.6 15.5 3.7
Copepods
Paracalanus sp. 1015.3 287.3 738.8 169.4
Temora sp. 558.8 90.2 278.5 83.7
Oncaea sp. 497.9 137.1 363.2 124.8
Brachyuran zoea 48.8 7.8 59.7 20.1
Anomuran zoea 10.3 3.7 23.5 5.6
Crab megalopa 2.9 1.3 3.2 1.2
Amphipods 3.2 1.4 183.7 58.4
Table 8. ANOVA for abundance of amphipods and decapods
at different depths during the day and night, on three occasions
at Kurnell Point.
Source of
variation
df C = 0.389, not
sig.
Amphipods
C = 0.399,
not sig.
Decapods
MS MS
Times = T 2 0.10 4.46
Time of
day = TD
1 102.23 12.49
Depth = D 1 13.66 5.07
T · TD 2 1.08 0.38
T · D 2 1.89 1.47
TD · D 1 9.57* 4.77*
T · TD · D 2 2.93 0.72
Residual 24 2.08 0.51
All data were ln(x + 1) transformed (n = 3). C = Cochran’s
test for heterogeneity of data.
*Denotes significant at p < 0.05.
170

Discussion
Diel distributions and movements
The most abundant nocturnal planktivores onSydney’s rocky reefs were two species of pem-pherids P. affinis and P. multiradiata. Apogonidswere found but were relatively rare compared totropical reefs (Marnane & Bellwood 2002). Al-though there are few comparative studies, pem-pherids are often the most abundant planktivoresfound on temperate reefs in the southern hemi-sphere (MacDiarmid 1981, Russell 1983, LincolnSmith 1989). Historical models of the movementsof pempherids have described similar patterns formost species (Mooi 2000). Qualitative assessmentsof the timing of migrations have been done (Fi-shelson et al. 1971, MacDiarmid 1981) with littleemphasis on their actual extent and nature(Gladfelter 1979). On the basis of this project, it isclear that there are striking differences in thefeeding ecology of pempherids, both betweenspecies and among size-classes within species.The diel distribution and abundance of large
P. affinis and P. multiradiata varied considerably.P. multiradiata consistently moved into the shal-lows during the night while P. affinis remainedwithin the same mid and deep depth strata, andmoved only short distances from daytime sites.Furthermore, differences in the abundance of P.multiradiata at study locations during the day andnight suggested this species underwent both hori-zontal migrations into and away from studylocations, as well as vertical migrations at night.Movement from deep to shallow depths duringforaging migrations has been documented amongother pempherid species on a scale of tens ofmetres (MacDiarmid 1981), as well as on a smallerscale of within 2 m in shallow waters (Gladfelter1979). Variation in the patterns of movement oftwo such closely related species, however, has notpreviously been demonstrated.It is widely acknowledged that migratory
behaviour leads to increased risk of predation(Gliwicz & Jachner 1992). Morphological andphysiological differences between the two speciesmay render P. affinis more vulnerable to preda-tion, and hence less extensive migrations. Vulner-ability to predation commonly varies withstandard length of fishes (Bray 1980). Pempheris
affinis generally grows to a smaller maximumstandard length than P. multiradiata (Edgar,1997). This smaller size is likely to increase the riskof predation during longer migrations. The col-ouration of the two species is slightly different.Pempheris multiradiata is bronze-coloured whileP. affinis has a silver colouration. This may lead toincreased reflection of ambient light, therebymaking them more noticeable in the water columnand hence more vulnerable to predation. Physio-logical considerations may include differences inswimming ability or visual perception of prey(Pankhurst 1982). Pempheris multiradiata mayhave superior swimming ability, enabling them tocover longer distances efficiently, and withoutusing excess energy. Further research on theseareas may help elucidate the reasons pempheridsundergo variable migrations.Differences in diel migration behaviour among
size-classes within species were also evident. Incontrast to large pempherids, small (0+ fish) werefound to have a close association with the substra-tum away from shelter during the day. This is acommon feature among rocky reef fishes wheresmall members of a species either undergo lessextensive movements (Hobson 1973) or migrate atdifferent times of the day (Mooi 2000). Small fishesmay undergo less extensive movements due to riskof predation (Gliwicz & Jachner 1992) or in order toreduce energy expenditure associated with extensivemovement away from shelter sites (Bray 1980).Another possibility may be that nocturnal vision ispoorly developed early in life (Pankhurst 1982).
Site fidelity
Although tagged individuals migrated away fromshelter sites during the night, most individualswere found returning to the same cave for 7 weeks,while other fish moved to nearby caves (<20 maway). A previous tagging study on P. oualensisfound some fish returning every morning to thesame cave for 3 months (Fishelson et al. 1971).This behaviour indicates a high degree of sitespecificity and predictability of the nocturnalmigrations of pempherid species. The number oftagged fishes sighted in this study did, however,decrease during the sampling period. Possiblereasons that tagged fishes were not sighted duringsampling may be movement outside the sample
171

area, tag loss, observer failure to see tags ormortality of tagged individuals. Movement be-yond the mapped area at night is a strong possi-bility given the sighting of tagged individualsduring the day in other caves up to 20 m away. Inaddition, tagged individuals not sighted at nightwere found later at the tagging site. Loss of VIFEtags is unlikely on the basis of past research whichhas found a retention rate of 91.3% when elasto-mer is injected into the caudal peduncle (Farooqi& Morgan 1996) as it was in this study. Addi-tionally, VIFE tags have not been found to in-crease the susceptibility of tagged fishes topredation (Malone et al. 1999), however, a highpredation rate is plausible given the high mortalityrates and low age maxima (1–2 years) of closelyrelated apogonids (Chrystal et al. 1985).
Diet and prey availability
Dietary data supported field observations thatjuvenile and adult pempherids have different forag-ing behaviour. Juveniles of both species feed duringthe day while adults feed at night. Both species,therefore, undergo an ontogenetic transition fromdiurnal to nocturnal foraging. Ontogenetic change ofthis type has also been recorded for the nocturnalplanktivore Pempheris adspersa (Mooi 2000). Dif-ferences in the timing of foraging were accompaniedby the consumption of different prey items. Juvenilesconsumed a range of plankton including copepodsand decapod larvae, while amphipods constitutedthe greatest percent representation in the stomachsof large fishes. Other demersal zooplankters suchostracods, were also common. Pempherids, there-fore, have important trophic links with benthicinvertebrate assemblages at night.An ontogenetic shift in the size of prey items was
also evident. There was an increase in the varianceof prey size with increasing standard length offishes. This is probably due to small and largefishes feeding at different times of the day, andmany large prey items being available only at night(Magnan & Fitzgerald 1984). There may also be arelaxation of morphological constraints (e.g. jawgape), with increasing fish size, resulting in greaterprey capture success (Norton 1995). The ability toforage on a wider range of prey items may enablegreater flexibility of feeding behaviour (Lowe et al.1996). Large fishes may therefore be able to exploit
a range of habitats to maximise prey intake(Gladfelter 1979, Sanchez-Velasco 1998) anddiversity of prey as well as minimising the timespent foraging (Grossman 1980). The movementsof large fishes must also increase the chances ofencountering patches of suitable prey above dif-ferent habitats and at different depths.The composition of planktonic assemblages was
found to differ between day and night. Manydemersal species such as amphipods, ostracods andmysids were therefore migrating vertically into thewater column at different times of the day. Thispattern has been demonstrated on a number ofoccasions in tropical waters (Alldredge & King1980, 1985, Hamner 1981). The timing of suchmigrations is often highly variable and can occursuddenly, resulting in a significant change inplanktonic assemblages within 1–2 h (Gallahar1997). Abundance of prey also varied with depthon the reefs in our study. Although spatial varia-tion in plankton is well documented, there are fewdata from reefs day and night at different depths.This variation could influence movements and diet.In conclusion, we have demonstrated greater
differences in behaviour between congeneric pem-pherids. Although both species showed a transitionfrom diurnal to nocturnal feeding behaviour, onespecies only moved a few metres from day timeresting sites, while the other moved 10–100 ofmetres at night. These movements need to be con-sidered in models of nutrient and energy transfer onreefs, as well as the management of reef resources.
Acknowledgements
Thanks to I. Carlson, M. Flood, S. Gartenstein andH. Patterson for help in the field. Thanks also to H.Patterson for commenting on the manuscript.
References
Alldredge, A.L. & J.M. King. 1980. Effects of moonlight in the
vertical migration patterns of demersal zooplankton. J. Exp.
Mar. Biol. Ecol. 44: 133–156.
Alldredge, A.L. & J.M. King. 1985. The distance demersal
zooplankton migrate above the benthos: implications for
predation. Mar. Biol. 84: 253–260.
Annese, D.M. 2000. Distribution and movements of nocturnal
rocky reef fishes. Bachelor of Science with Honours Thesis,
The University of Sydney, 84 pp.
172

Bray, R.N. 1980. Daily foraging migrations of the blacksmith
Chromis punctipinnis, a planktivorous kelp-bed damsel. Bull.
Mar. Sci. 30: 325–326.
Bray, R.N., A.C. Miller & G.G. Geesey. 1981. The fish con-
nection: a trophic link between planktonic and rocky reef
communities? Science 214: 204–205.
Chrystal, P.J., I.C. Potter, N.R. Loneragan & C.P.Holt. 1985.
Age structure, growth rates, movement patterns and feeding
in an estuarine population of the cardinalfish Apogon rue-
ppellii. Mar. Biol. 85: 185–197.
Edgar, G.J. 1997. Australian Marine Life: the plants and ani-
mals of temperate waters. Reed Books, Victoria. 544 pp.
Farooqi, M.A. & C.E. Morgan. 1996. Elastomer visible implant
(EVI) tag retention and the effect of tagging on the growth
and survival of barbel, Barbus barbus (L.). Fish. Manage.
Ecol. 3: 181–183.
Fishelson, L., D. Popper & N. Gunderman. 1971. Diurnal
cyclic behaviour of Pempheris oualensis Cuv. & Val. (Pemp-
heridae, Teleostei). J. Nat. History 5: 503–506.
Gallahar, N.K. 1997. The Ecology of Gammaridean Amphi-
pods on the Kelp, Ecklonia radiata. Ph.D. Thesis. The Uni-
versity of Sydney. 226 pp.
Gladfelter, W.B. 1979. Twilight migrations and foraging
behaviour of the Copper Sweeper Pempheris schomburgki
(Teleosti: Pempheridae). Mar. Biol. 50: 109–119.
Gliwicz, Z.M. & A. Jachner. 1992. Diel migration of juvenile
fish: a ghost of predation past or present? Archiv fur Hy-
drobiol. 124: 385–410.
Greenfield, D.W. & R.K. Johnson. 1990. Heterogeneity in
habitat choice in cardinalfish community structure. Copeia 4:
1107–1114.
Grossman, G.D. 1980. Ecological aspects of ontogenetic shifts
in prey size utilization in the bay goby (Pisces: Gobiidae).
Oecologia 47: 233–238.
Hamner, R.M. 1981. Day-night differences in the emergence of
demersal zooplankton from a sand substrate in a kelp forest.
Mar. Biol. 62: 275–280.
Hobson, E.S. 1972. Activity of Hawaiian reef fishes. Fish. Bull.
70: 715–740.
Hobson, E.S. 1973. Diel feeding migrations in tropical reef fishes.
Helgolander wissenschaften Meeresuntersuchen 24: 361–370.
Hobson, E.S. 1974. Feeding relationships of teleostean fishes on
coral reefs in Kona, Hawaii. Fish. Bull. 72: 915–1031.
Jacoby, C.A. & J.G. Greenwood. 1988. Spatial, temporal, and
behavioural patterns in emergence of zooplankton in the la-
goon of Heron Reef, Great Barrier Reef, Australia. Mar.
Biol. 97: 309–328.
Jones, G.P. 1988. Experimental evaluation of the effects of
habitat structure and competitive interactions on the juve-
niles of two coral reef fishes. J. Exp. Mar. Biol. Ecol. 123:
115–126.
Jones, R.S. 1968. A suggested method for quantifying gut
contents in herbivorous fishes. Micronesica 4: 369–371.
Kingsford, M.J. & A.B. MacDiarmid. 1988. Interrelations be-
tween planktivorous reef fish and zooplankton in temperate
waters. Mar. Ecol. Progr. Ser. 48: 103–117.
Lincoln Smith, M.P. 1989. Improving multispecies rocky reef
fish censuses by counting different groups of species using
different procedures. Environ. Biol. Fishes 26: 29–37.
Lincoln Smith, M.P., J.D. Bell & C.A. Hair. 1991. Spatial
variation in abundance of recently settled rocky reef fish in
southeastern Australia – Implications for detecting change.
Mar. Ecol. Progr. Series 77: 95–103.
Lockett, M.M. & I.M. Suthers. 1998. Ontogenetic diet shift and
feeding activity in the temperate reef fish Heilodactylus Fus-
cus. Proc. Linnean Soc. 120: 105–116.
Lowe, C.G., B.M. Wetherbee, G.L. Crow & A.L. Tester. 1996.
Ontogenetic dietary shifts and feeding behaviour of the tiger
shark, Galeocerdo cuvier, in Hawaiian waters. Environ. Biol.
Fishes 47: 203–211.
Lowry, M.B. & I.M. Suthers. 1998. Home range, activity and
distribution patterns of a temperate rocky-reef fish, Cheilo-
dactylus fuscus. Mar. Biol. 132: 569–578.
MacDiarmid, A.B. 1981. Factors influencing the distribution
and abundance of two temperate reef fish, Pempheris ad-
spersa and Scorpis violaceus. Master of Science Thesis. Uni-
versity of Auckland. 101 pp.
Magnan, P. & G.J. Fitzgerald. 1984. Ontogenetic changes in
diel activity, food habits and spatial distribution of juvenile
and adult creek chub, Semotilus atromaculatus. Environ.
Biol. Fishes 11: 301–307.
Malone, J.C., G.E. Forrester & M.A. Steele. 1999. Effects of
subcutaneous microtags on the growth, survival, and vul-
nerability to predation of small reef fishes. J. Exp. Mar. Biol.
Ecol. 237: 243–253.
Marnane, M.J. & D.R. Bellwood. 2002. Diet and nocturnal
foraging in cardinalfishes (Apogonidae) at One Tree Reef,
Great Barrier Reef, Australia. Mar. Ecol. Progr. Ser. 231:
261–268.
Matthews, K. 1992. A telemetric study of the home ranges and
homing routes of lingcod Ophiodon elongatus on shallow
rocky reefs off Vancouver Island, British Columbia. Fish.
Bull. 90: 784–790.
Mooi, R.D. 2000. Review of New Zealand bullseyes (Percifor-
mes: Pempheridae). New Zealand J. Mar. Freshwater Res.
34: 87–102.
Norton, S.F. 1995. A functional approach to ecomorphological
patterns of feeding in cottid fishes. Environ. Biol. Fishes 44:
61–78.
Pankhurst, N.W. 1982. Relation of visual changes to the onset
of sexual maturation in the European eel Anguilla anguilla
(L.). J. Fish Biol. 21: 127–140.
Platell, M.E. & I.C. Potter. 1999. Partitioning of habitat and
prey by abundant and similar-sized species of the Triglidae
and Pempherididae (Teleostei) in coastal waters. Estuarine
Coastal & Shelf Science 48: 235–252.
Russell, B.C. 1983. The food and feeding habits of rocky reef
fishes of northeastern New Zealand. New Zealand J. Mar.
Freshwater Res. 17: 121–145.
Sale, P.F. 1978. Reef fishes and other invertebrates: a compar-
ison of social structures. pp. 313–346. In: E.S. Reese & F.J.
Lichter (eds.) Contrasts in Behaviour: Adaptations in the
Aquatic and Terrestrial Environments, Wiley & Sons, New
York.
Sale, P.F. 1991. Reef fish communities: Open nonequilib-
rial systems. pp. 564–600. In: P.F. Sale (ed.) The
Ecology of Fishes on Coral Reefs, Academic Press, San
Diego.
173

Sanchez-Velasco, L. 1998. Diet composition and feeding habits
of fish larvae of two co-occurring species (Pisces: Calli-
onymidae and Bothidae) in the north-western Mediterra-
nean. J. Mar. Sci. 55: 299–308.
Steffe, A.S. & B.C. Pease. 1988. Diurnal survey of ichthyo-
plankton abundance, distribution and seasonality in Bot-
any Bay, New South Wales. Proc. Linnean Soc. 110: 1–
10.
174




















