
Distribution, colonization and diversity of arbuscular mycorrhizal fungi associated with central...
10
Distribution, colonization and diversity of arbuscular mycorrhizal fungi associated with central Himalayan rhododendrons Bhaskar Chaurasia a , Anita Pandey a, * , Lok Man S. Palni b a Environmental Physiology and Biotechnology, G B Pant Institute of Himalayan Environment and Development, Kosi-Katarmal, Almora-263 643, Uttaranchal, India b State Biotechnology Programme, Govt. of Uttaranchal, Biotech Bhawan, P.O. Haldi, Pantnagar-263 146, India Received 25 February 2004; received in revised form 16 July 2004; accepted 12 October 2004 Abstract The objective of the present study was to investigate arbuscular mycorrhizal status of five species of rhododendrons distributed in Kumaun region of the Indian Central Himalaya. Root and rhizosphere soil samples of all the five species of rhododendrons, namely, Rhododendron anthopogon, R. arboreum, R. campanulatum, R. barbatum and R. lepidotum were collected from temperate, sub-alpine to alpine location in altitudinal range from 1500 to 4500 m amsl. The arbuscular mycorrhizal colonization in root samples ranged from 28 to 42%; and maximum and minimum colonization was observed in R. arboreum and R. lepidotum, respectively. The highest number of intraradical vesicles (12.5 2.8 cm 1 root segment) was recorded in R. arboreum and the lowest (7.0 1.7 cm 1 root segment) in R. barbatum; vesicles were not observed in R. lepidotum. Spores were extracted from the rhizosphere soil by wet sieving to perform microscopic identification of the species. The maximum and minimum populations of spores were detected in the rhizosphere soil samples of R. anthopogon (52.0 1.5 spores 25 g 1 soil) and R. lepidotum (32.0 2.5 spore 25 g 1 soil), respectively. Spore populations were found to belong to five genera—Acaulospora, Glomus, Gigaspora, Sclerocystis and Scutellispora; genus Glomus was found to be dominant in the rhizosphere soil samples of all the rhododendron species. The most frequent and abundant species was G. fasciculatum, however, this species was not isolated from the rhizosphere soil of R. barbatum. Finger millet (Eleucine coracana) was used to cultivate the trap culture of arbuscular mycorrhizal fungi to confirm the species identity. Spores of Glomus pustulatum, not detected in the rhizosphere soil, were recovered from the trap culture. Contrary to this, genus Gigaspora, which was present in the rhizosphere soil, did not sporulate in the trap culture. Shannon and Wiener index of diversity and Simpson’s index of dominance indicated that the species richness, dominance and diversity of arbuscular mycorrhizal fungi decrease with increasing altitude. In two species of rhododendrons, namely R. campanulatum and R. anthopogon, dark septate mycelium was also observed. # 2004 Elsevier B.V. All rights reserved. Keywords: Arbuscular mycorrhizal colonization; Dark septate endophytes (DSE); Glomus; Vesicles; Species richness www.elsevier.com/locate/foreco Forest Ecology and Management 207 (2005) 315–324 * Corresponding author. Tel.: +91 5962 241041; fax: +91 5962 241150. E-mail addresses: [email protected], [email protected] (A. Pandey). 0378-1127/$ – see front matter # 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.foreco.2004.10.014
Transcript of Distribution, colonization and diversity of arbuscular mycorrhizal fungi associated with central...

www.elsevier.com/locate/foreco
Forest Ecology and Management 207 (2005) 315–324
Distribution, colonization and diversity of arbuscular mycorrhizal
fungi associated with central Himalayan rhododendrons
Bhaskar Chaurasiaa, Anita Pandeya,*, Lok Man S. Palnib
aEnvironmental Physiology and Biotechnology, G B Pant Institute of Himalayan Environment and Development,
Kosi-Katarmal, Almora-263 643, Uttaranchal, IndiabState Biotechnology Programme, Govt. of Uttaranchal, Biotech Bhawan, P.O. Haldi, Pantnagar-263 146, India
Received 25 February 2004; received in revised form 16 July 2004; accepted 12 October 2004
Abstract
The objective of the present study was to investigate arbuscular mycorrhizal status of five species of rhododendrons
distributed in Kumaun region of the Indian Central Himalaya. Root and rhizosphere soil samples of all the five species of
rhododendrons, namely, Rhododendron anthopogon, R. arboreum, R. campanulatum, R. barbatum and R. lepidotum were
collected from temperate, sub-alpine to alpine location in altitudinal range from 1500 to 4500 m amsl. The arbuscular
mycorrhizal colonization in root samples ranged from 28 to 42%; and maximum and minimum colonization was observed in R.
arboreum and R. lepidotum, respectively. The highest number of intraradical vesicles (12.5 � 2.8 cm�1 root segment) was
recorded in R. arboreum and the lowest (7.0 � 1.7 cm�1 root segment) in R. barbatum; vesicles were not observed in R.
lepidotum. Spores were extracted from the rhizosphere soil by wet sieving to perform microscopic identification of the species.
The maximum and minimum populations of spores were detected in the rhizosphere soil samples of R. anthopogon
(52.0 � 1.5 spores 25 g�1 soil) and R. lepidotum (32.0 � 2.5 spore 25 g�1 soil), respectively. Spore populations were found
to belong to five genera—Acaulospora, Glomus, Gigaspora, Sclerocystis and Scutellispora; genus Glomus was found to be
dominant in the rhizosphere soil samples of all the rhododendron species. The most frequent and abundant species was G.
fasciculatum, however, this species was not isolated from the rhizosphere soil of R. barbatum. Finger millet (Eleucine coracana)
was used to cultivate the trap culture of arbuscular mycorrhizal fungi to confirm the species identity. Spores of Glomus
pustulatum, not detected in the rhizosphere soil, were recovered from the trap culture. Contrary to this, genus Gigaspora, which
was present in the rhizosphere soil, did not sporulate in the trap culture. Shannon and Wiener index of diversity and Simpson’s
index of dominance indicated that the species richness, dominance and diversity of arbuscular mycorrhizal fungi decrease with
increasing altitude. In two species of rhododendrons, namely R. campanulatum and R. anthopogon, dark septate mycelium was
also observed.
# 2004 Elsevier B.V. All rights reserved.
Keywords: Arbuscular mycorrhizal colonization; Dark septate endophytes (DSE); Glomus; Vesicles; Species richness
* Corresponding author. Tel.: +91 5962 241041; fax: +91 5962 241150.
E-mail addresses: [email protected], [email protected] (A. Pandey).
0378-1127/$ – see front matter # 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.foreco.2004.10.014

B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324316
1. Introduction
The Indian Himalayan region consists of five
geographical zones i.e., Trans, North West, West,
Central and East Himalaya and covers an area of
approximately 419,873 km2.
Arbuscular mycorrhizal fungi are symbiotic asso-
ciations formed between the members of phylum
Glomeromycota and the roots of most terrestrial
flowering plants (Gianinazzi and Gianinazzi-Pearson,
1986). Arbuscular mycorrhizal fungi are the most
wide spread group (Schubler et al., 2001) found in the
tropics (Chaurasia, 2000), temperate (Vestberg, 1995)
and arctic regions (Dalpe and Aiken, 1998).
The mycorrhizal status from different ecological
regions i.e. aquatic and marshy habitats (Sengupta and
Chaudhuri, 1990), tropical plains (Ragupathy and
Mahadevan, 1993), tropical forests (Chaurasia, 2000),
sand dunes (Mohankumar et al., 1988), mangroves
(Mohankumar and Mahadevan, 1986) and the Western
Ghat region (Muthukumar and Udaiyan, 2000) of the
Indian sub-continent has been investigated.
Dark septate endophytes (DSE) are a miscella-
neous group of ascomycetous fungi that colonize roots
of many plants, both intracellularly and intercellularly
(Christie and Nicolson, 1983; O’Dell et al., 1993;
Ahlich and Sieber, 1996; Jumpponen, 2001). These
fungi were first described as Mycelium radicis
atrovirens (Smith and Read, 1997) and were found
to melanize hyphae and colonized the roots, occa-
sionally forming microsclerotia and sometimes a weak
hartig net in conifers (O’Dell et al., 1993; Ahlich and
Sieber, 1996).
The available information on mycorrhizal associa-
tions of rhododendrons indicates a number of
mycorrhizal partners (Largent et al., 1980); the main
associates appear to be ericoid in nature (Read and
Stribley, 1975; Duddrige and Read, 1982). However,
arbuscular mycorrhizal status of the rhododendron
species has received little attention (Cross, 1975;
Dighton and Coleman, 1992). Dighton and Coleman
(1992) reported the presence of intracellular arbus-
cules, vesicles and intraradical spores of arbuscular
mycorrhizal fungi in the roots of R. maximum. In the
present study the occurrence, colonization and the
diversity of arbuscular mycorrhizal fungi associated
with all the five species of rhododendrons found on an
altitudinal range from 1500 to 4500 m amsl in
Kumaun region of Indian Central Himalaya have
been investigated.
2. Materials and methods
2.1. Site description
The present study was conducted in the temperate,
sub-alpine and alpine zones of Pindari Glacier region
(33850–308100N to 798480–798520E) of Kumaun
Himalaya in the Indian Central Himalaya. A total
of five species of rhododendrons (family Ericaceae),
distributed from 1500 to 4500 m amsl (up to timber-
line), are found in this region. R. arboreum is most
widely distributed and starts appearing around 1500 m
elevation; it increases in importance with elevation
especially in oak dominated forests. Beyond 2800 m it
is replaced by R. barbatum in the birch forest.
Eventually, vegetation is represented by scrubs or
heaths (above 3400 m amsl), where R. campanulatum
and R. anthopogon are the sole representatives of
woody vegetation, and R. lepidotum forms sparse
bushy appearance in the alpine meadows. The general
description of the study sites is presented in Table 1.
2.2. Collection of soil and root samples
The rhizosphere soil and root samples were
collected from all the five species of rhododendrons,
preferably from the sites where the species were
found to grow in form of a pure stand or pure patch in
natural forest. The samples were taken after
removing the top litter layer (5–10 cm) and digging
out an appropriate amount of soil close to the roots of
the host plant from a depth of 10–15 cm. The samples
were collected in triplicate. Soil particles attached to
fine feeder roots were removed by generous shaking.
The soil samples brought to the laboratory were
stored at 4 8C to sustain the viability of arbuscular
mycorrhizal spores and to lower the activity of the
rhizosphere micro flora.
2.3. Assessment of arbuscular mycorrhizal
colonization
The root samples were rinsed thoroughly with
running tap water several times to remove the debris
B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324 317
Table 1
Distribution of rhododendrons found in Indian Central Himalaya and the study locations
Plant species/habit and height Locationa (m) Soil pH Other associates
R. anthopogon Don, shrub, 0.18–0.95 m Phurkia forest, 3000–4500 5.12 R. lepidotum, R. campanulatum,
Cotoneaster microphyllus, Pyrus foliolosa
R. arboreum Sm, tree, 7.62–12.20 m Khaljhuni forest, 1500–3000 5.32 Quercus semicarpifolia, Q. floribunda,
Lyonia ovalifolia, Symplocos chinensis,
Litsea umbrosa
R. barbatum G. Don, tree, 6.09–7.62 m Sundardunga forest, 2600–3200 4.74 Prunus cornuta, Acer accuminatum, Abies
pindrow, R. arboreum, Q. semicarpifolia
R. campanulatum Don, shrub, 3.04–4.57 m Phurkia forest, 2800–4000 4.60 Rosa sericea, R. anthopogon, Lonicera
angustifolia, Betula utlis, Prunus cornuta,
Pyrus foliolosa, Syringa emodi
R. lepidotum Wall ex G. Don, shrub, 0.30–0.61 m Phurkia forest, 3300–4500 4.62 R. anthopogon, Pyrus foliolosa
a All located in district Bageshwar, Kumaun region of Uttaranchal in the Indian Central Himalaya.
and adhering soil particles. Roots were cut into 1 cm.
segments for cleaning and staining. Root segments
were cleared in KOH (10%,w/v; 90 8C, 2 h) and
stained with acid fuschin (0.01%, w/v; Kormanik
et al., 1980).
The assessment of root colonization by arbuscular
mycorrhizal fungi was done by the slide method; root
segments were selected randomly from the stained
samples and observed for the presence or absence of
functional structures (mycelium, vesicles and arbus-
cules) of arbuscular mycorrhizal fungi. A minimum
of 100 root segments were used for this enumeration,
and the colonization by arbuscular mycorrhizal
fungi was calculated using the following formula:
%Colonization¼ total number of root segments colonizedtotal number of root segments studied �100
2.4. Extraction and identification of arbuscular
mycorrhizal spores from the rhizosphere soil
Extraction of arbuscular mycorrhizal spores was
carried out following the wet-sieving and decanting
method (Gerdemann and Nicolson, 1963). The soil–
water mixture (25:1000; w/v) was decanted through
stacked sieves (the coarse sieve on the top and the
finest sieve at the bottom). The sieve sizes ranged from
36 to 420 mm. Population of arbuscular mycorrhizal
spores was estimated by the method of Gour and
Adholeya (1994). Spores were separated into groups
according to their size and the general morphological
similarities recorded under a stereomicroscope
(Nikon). The slides were prepared using PVL
(polyvinyl alcohol + lactophenol) or PVLG (polyvinyl
alcohol + lactic acid + glycerol) for the identification
of arbuscular mycorrhizal species. Spores were
identified up to species level according to Schenck
and Perez (1990). Observations on the size, wall and
other spore structures were carefully recorded with an
optiphot-2 (Nikon) and the micrographs were taken
with fx35 dx (Nikon) camera.
2.5. Trap cultures
Since arbuscular mycorrhizal fungi do not grow
independently on routine culture media, greenhouse
pot cultures were established with the soil samples to
recover spores of arbuscular mycorrhizal fungal
species present in the soil, including those that may
not have sporulated at the time of sampling. A mixture
of soil and sand (3:1) (autoclaved three times at 121 8Cand 1.1 kg/cm2 for 1 h) was used for pot cultures
(30 cm height � 25 cm diameter) for providing a
suitable host, the seeds of a local finger millet (E.
coracana) previously surface sterilized with 0.01%
HgCl2 for 2 min and washed several 3–4 times with
sterilized distilled water, were sown in the pots (20
seeds per pot). Approximately 50 g of rhizosphere soil
was mixed with the seeds at the time of sowing. The
spores were extracted after 3–4 months, as described
earlier.
2.6. Data analysis
Shannon and Wiener index of diversity (Hutch-
enson, 1970), Simpson’s index of dominance (Simp-
son, 1949), similarity index, and species richness
(Franke-Synder et al., 2001) were calculated for the
B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324318
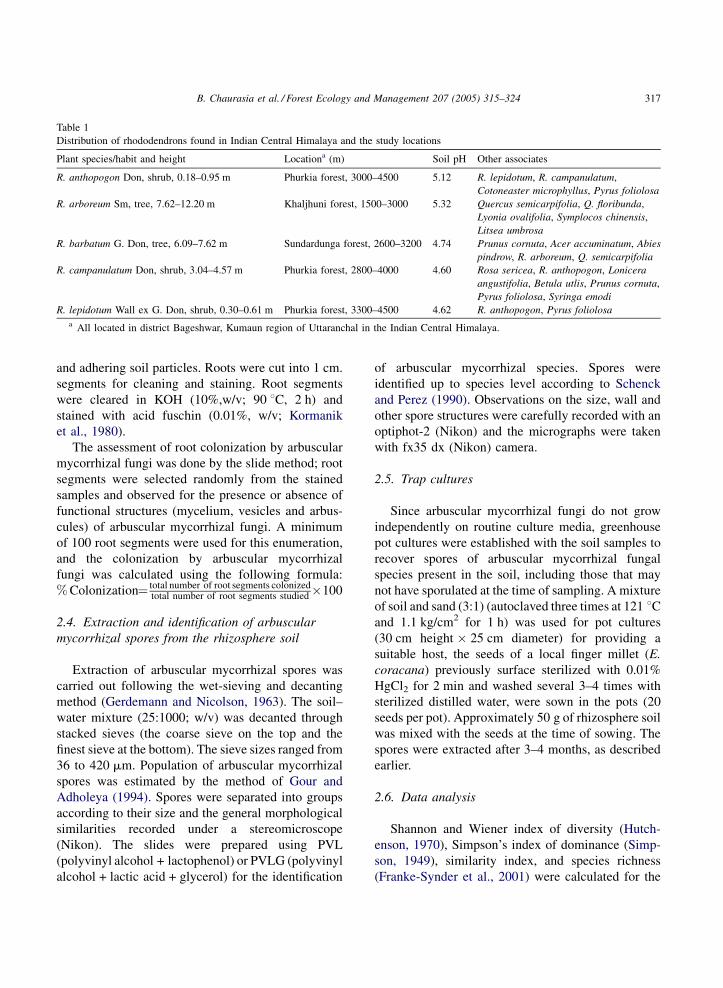
Fig. 1. Arbuscular mycorrhizal colonization in rhododendrons of Kumaun, Indian Central Himalaya. (A) Extraradical chlamydospore and
vesicles (bar = 100 mm). (B) Intraradical vesicles (bar = 100 mm). (C) Intraradical chlamydospore of Glomus sp. (bar = 50 mm). (D) Hyphal
coiling inside the cell (bar = 40 mm). (E) Constriction in the mycelium and intraradical vesicles in the root (bar = 20 mm). (F) Dark septate
mycelium (bar = 40 mm).
distribution and diversity of arbuscular mycorrhizal
fungi.
3. Results
Colonization by the arbuscular mycorrhizal fungi
was observed in the root samples of different species
of rhododendrons. Extraradical spore and vesicles
(Fig. 1A) as wall as intraradical vesicles (Fig. 1B) and
spore (Fig. 1C) were observed during the study.
Extraradical as well as intraradical spores and vesicles
varied from spherical to ellipsoidal in shape.
Colonization of arbuscular mycorrhizal fungi was
almost similar in all rhododendron species. Maximum
arbuscular mycorrhizal percent colonization was
observed in R. arboreum (42%) followed by R.
barbatum (40%), R. anthopogon (37%), R. campa-
nulatum (33%) and R. lepidotum (28%). Intraradical
vesicles were not observed in R. lepidotum samples.
Arbuscules were not detected in the root segments of
all the species, however, the hyphal coiling (Fig. 1D)
and constriction in hyphae (Fig. 1E) were observed.
The results indicated that there was no correlation
between the arbuscular mycorrhizal colonization and
spore population. The number of species of arbuscular
mycorrhizal fungi associated with the rhizosphere soil
samples of all the rhododendrons showed a limited
B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324 319
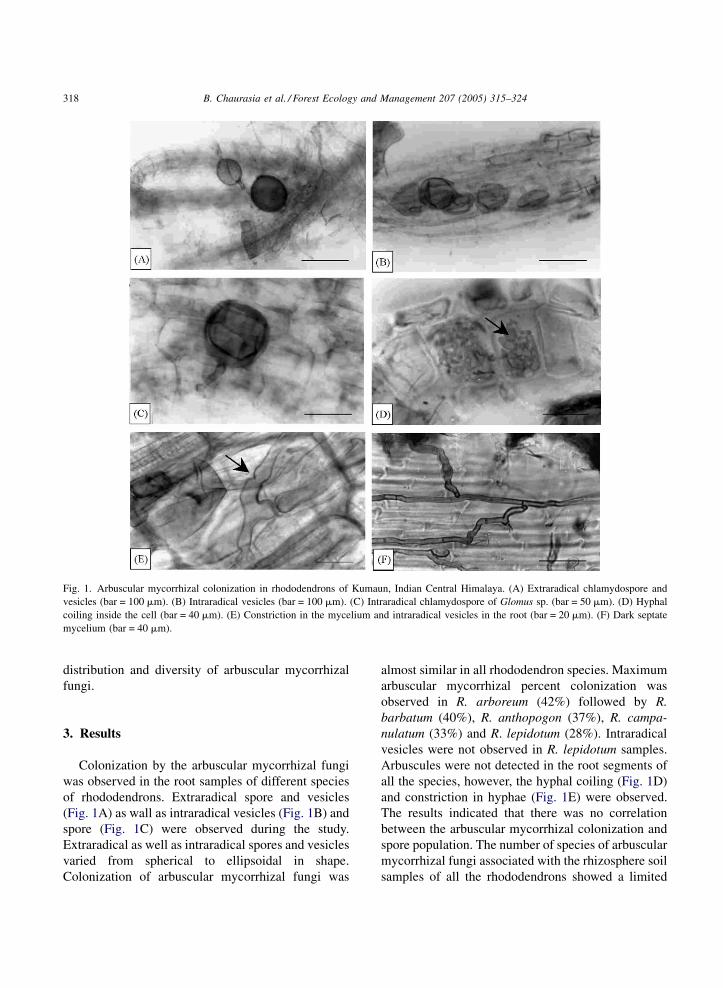
Fig. 2. Arbuscular mycorrhizal colonization (%) and number of arbuscular mycorrhizal species found in rhizosphere soil (RS) of rhododen-
drons.
correlation with arbuscular mycorrhizal colonization
(Fig. 2). The rhizosphere soil of R. arboreum was found
to harbor maximum number of arbuscular mycorrhizal
fungal species (12) followed by R. barbatum (9), R.
anthopogon (8), R. campanulatum (8) and R. lepidotum
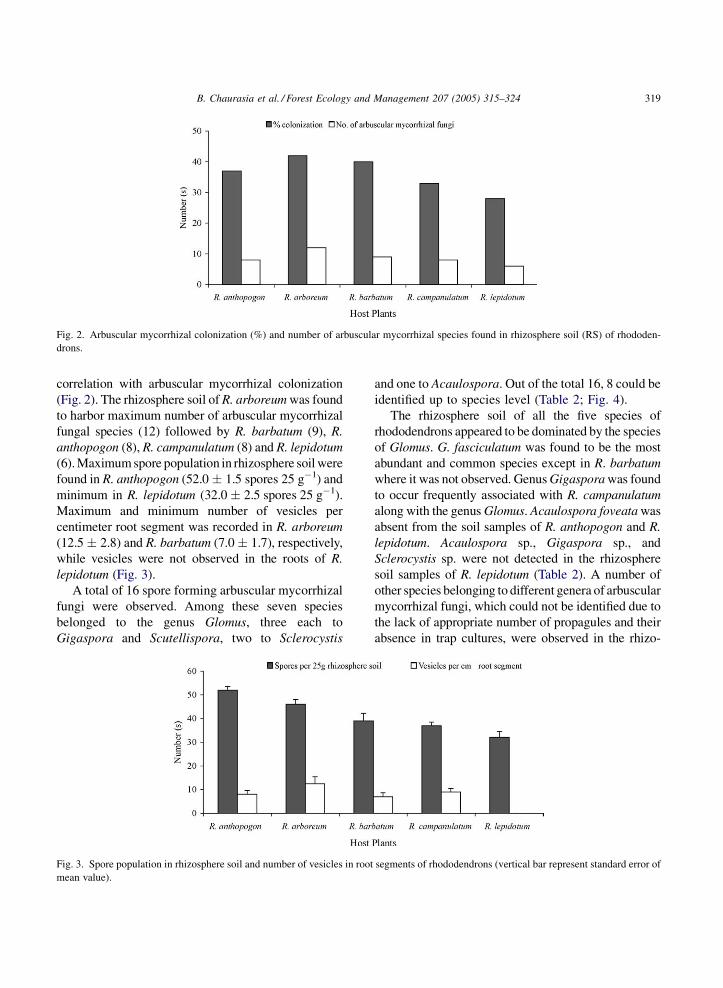
(6). Maximum spore population in rhizosphere soil were
found in R. anthopogon (52.0 � 1.5 spores 25 g�1) and
minimum in R. lepidotum (32.0 � 2.5 spores 25 g�1).
Maximum and minimum number of vesicles per
centimeter root segment was recorded in R. arboreum
(12.5 � 2.8) and R. barbatum (7.0 � 1.7), respectively,
while vesicles were not observed in the roots of R.
lepidotum (Fig. 3).
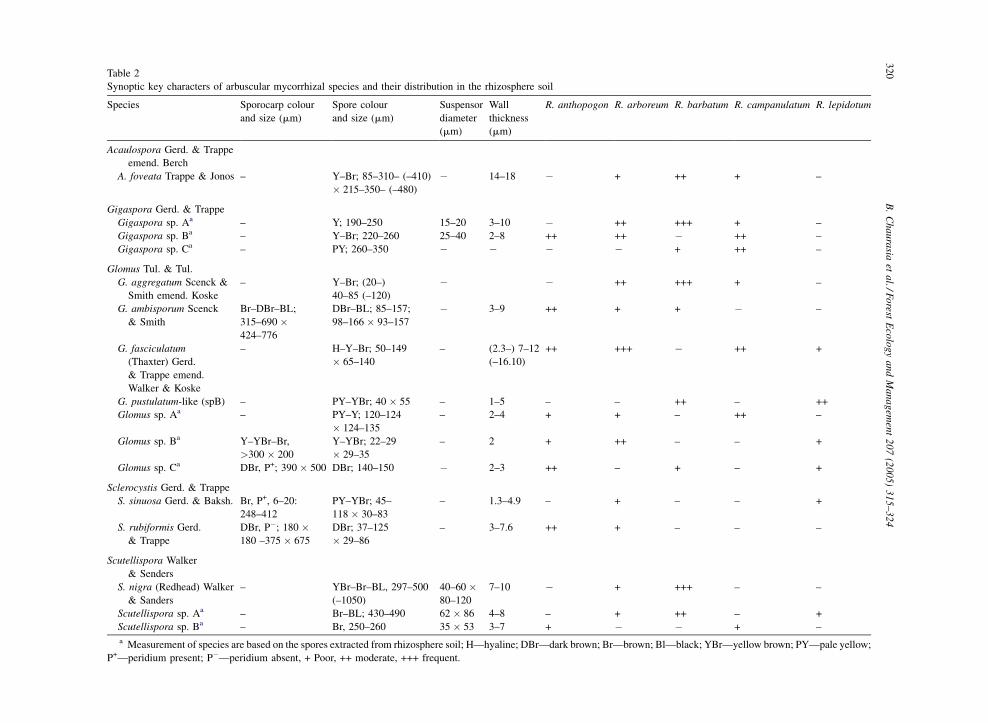
A total of 16 spore forming arbuscular mycorrhizal
fungi were observed. Among these seven species
belonged to the genus Glomus, three each to
Gigaspora and Scutellispora, two to Sclerocystis
Fig. 3. Spore population in rhizosphere soil and number of vesicles in root
mean value).
and one to Acaulospora. Out of the total 16, 8 could be
identified up to species level (Table 2; Fig. 4).
The rhizosphere soil of all the five species of
rhododendrons appeared to be dominated by the species
of Glomus. G. fasciculatum was found to be the most
abundant and common species except in R. barbatum
where it was not observed. Genus Gigaspora was found
to occur frequently associated with R. campanulatum
along with the genus Glomus. Acaulospora foveata was
absent from the soil samples of R. anthopogon and R.
lepidotum. Acaulospora sp., Gigaspora sp., and
Sclerocystis sp. were not detected in the rhizosphere
soil samples of R. lepidotum (Table 2). A number of
other species belonging to different genera of arbuscular
mycorrhizal fungi, which could not be identified due to
the lack of appropriate number of propagules and their
absence in trap cultures, were observed in the rhizo-
segments of rhododendrons (vertical bar represent standard error of
B.
Ch
au
rasia
eta
l./Fo
restE
colo
gy
an
dM
an
ag
emen
t2
07
(20
05
)3
15
–3
24
32
0Table 2
Synoptic key characters of arbuscular mycorrhizal species and their distribution in the rhizosphere soil
Species Sporocarp colour
and size (mm)
Spore colour
and size (mm)
Suspensor
diameter
(mm)
Wall
thickness
(mm)
R. anthopogon R. arboreum R. barbatum R. campanulatum R. lepidotum
Acaulospora Gerd. & Trappe
emend. Berch
A. foveata Trappe & Jonos – Y–Br; 85–310– (–410)
� 215–350– (–480)
� 14–18 � + ++ + –
Gigaspora Gerd. & Trappe
Gigaspora sp. Aa – Y; 190–250 15–20 3–10 � ++ +++ + –
Gigaspora sp. Ba – Y–Br; 220–260 25–40 2–8 ++ ++ � ++ –
Gigaspora sp. Ca – PY; 260–350 � � � � + ++ –
Glomus Tul. & Tul.
G. aggregatum Scenck &
Smith emend. Koske
– Y–Br; (20–)
40–85 (–120)
� � ++ +++ + –
G. ambisporum Scenck
& Smith
Br–DBr–BL;
315–690 �424–776
DBr–BL; 85–157;
98–166 � 93–157
� 3–9 ++ + + � –
G. fasciculatum
(Thaxter) Gerd.
& Trappe emend.
Walker & Koske
– H–Y–Br; 50–149
� 65–140
– (2.3–) 7–12
(–16.10)
++ +++ � ++ +
G. pustulatum-like (spB) – PY–YBr; 40 � 55 – 1–5 – – ++ – ++
Glomus sp. Aa – PY–Y; 120–124
� 124–135
– 2–4 + + – ++ –
Glomus sp. Ba Y–YBr–Br,
>300 � 200
Y–YBr; 22–29
� 29–35
– 2 + ++ – – +
Glomus sp. Ca DBr, P+; 390 � 500 DBr; 140–150 � 2–3 ++ – + – +
Sclerocystis Gerd. & Trappe
S. sinuosa Gerd. & Baksh. Br, P+, 6–20:
248–412
PY–YBr; 45–
118 � 30–83
– 1.3–4.9 – + – – +
S. rubiformis Gerd.
& Trappe
DBr, P�; 180 �180 –375 � 675
DBr; 37–125
� 29–86
– 3–7.6 ++ + – – –
Scutellispora Walker
& Senders
S. nigra (Redhead) Walker
& Sanders
– YBr–Br–BL, 297–500
(–1050)
40–60 �80–120
7–10 � + +++ – –
Scutellispora sp. Aa – Br–BL; 430–490 62 � 86 4–8 – + ++ – +
Scutellispora sp. Ba – Br, 250–260 35 � 53 3–7 + � � + –
a Measurement of species are based on the spores extracted from rhizosphere soil; H—hyaline; DBr—dark brown; Br—brown; Bl—black; YBr—yellow brown; PY—pale yellow;
P+—peridium present; P�—peridium absent, + Poor, ++ moderate, +++ frequent.
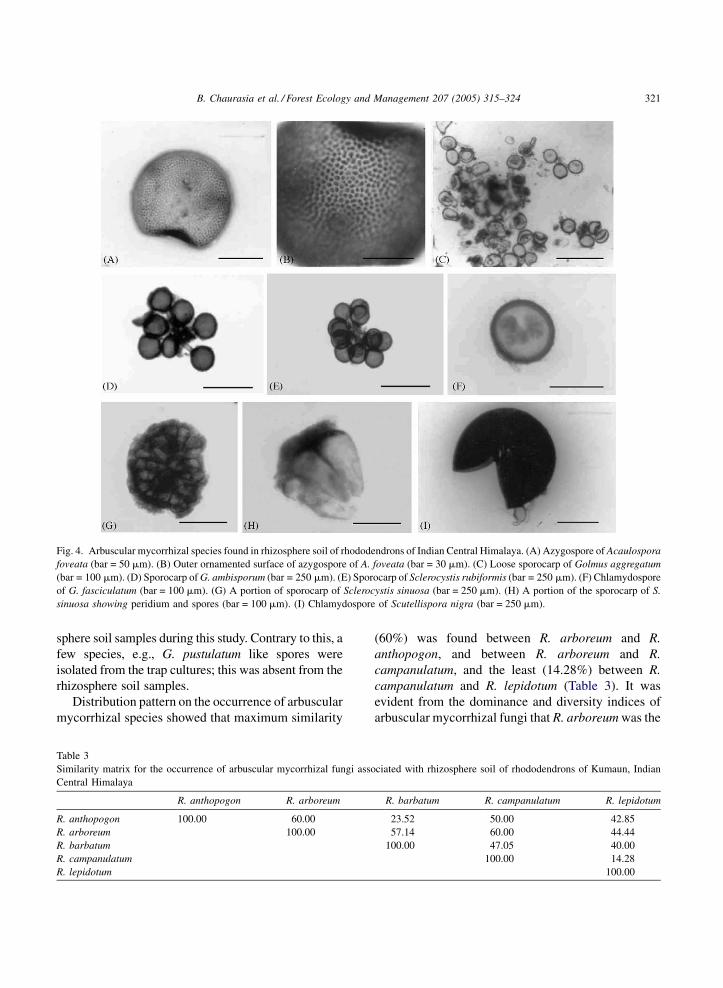
B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324 321
Fig. 4. Arbuscular mycorrhizal species found in rhizosphere soil of rhododendrons of Indian Central Himalaya. (A) Azygospore of Acaulospora
foveata (bar = 50 mm). (B) Outer ornamented surface of azygospore of A. foveata (bar = 30 mm). (C) Loose sporocarp of Golmus aggregatum
(bar = 100 mm). (D) Sporocarp of G. ambisporum (bar = 250 mm). (E) Sporocarp of Sclerocystis rubiformis (bar = 250 mm). (F) Chlamydospore
of G. fasciculatum (bar = 100 mm). (G) A portion of sporocarp of Sclerocystis sinuosa (bar = 250 mm). (H) A portion of the sporocarp of S.
sinuosa showing peridium and spores (bar = 100 mm). (I) Chlamydospore of Scutellispora nigra (bar = 250 mm).
sphere soil samples during this study. Contrary to this, a
few species, e.g., G. pustulatum like spores were
isolated from the trap cultures; this was absent from the
rhizosphere soil samples.
Distribution pattern on the occurrence of arbuscular
mycorrhizal species showed that maximum similarity
Table 3
Similarity matrix for the occurrence of arbuscular mycorrhizal fungi asso
Central Himalaya
R. anthopogon R. arboreum
R. anthopogon 100.00 60.00
R. arboreum 100.00
R. barbatum
R. campanulatum
R. lepidotum
(60%) was found between R. arboreum and R.
anthopogon, and between R. arboreum and R.
campanulatum, and the least (14.28%) between R.
campanulatum and R. lepidotum (Table 3). It was
evident from the dominance and diversity indices of
arbuscular mycorrhizal fungi that R. arboreum was the
ciated with rhizosphere soil of rhododendrons of Kumaun, Indian
R. barbatum R. campanulatum R. lepidotum
23.52 50.00 42.85
57.14 60.00 44.44
100.00 47.05 40.00
100.00 14.28
100.00

B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324322
Table 4
Dominance and diversity indices of arbuscular mycorrhizal fungi associated with rhizosphere soil of rhododendrons of Kumaun, Indian Central
Himalaya
Host plants Species richness (%) Shimpson’s index of dominance Shannon–Weiner index
R. anthopogon 50 (8 sp.) 0.1360 2.0315
R. arboreum 75 (12 sp.) 0.0987 2.3991
R. barbatum 56.25 (9 sp.) 0.1296 2.1099
R. campanulatum 50 (8 sp.) 0.1388 2.0228
R. lepidotum 37.5 (6 sp.) 0.1836 1.7478
species where arbuscular mycorrhizal species were
dominant with the highest diversity (2.3991) and the
least dominance (1.7478) and diversity of arbuscular
mycorrhizal fungi (Table 4) in respect to R. lepidotum.
Dark septate endophytes (DSE) were found to
occur frequently along with arbuscular mycorrhizal
mycelium in R. campanulatum and R. anthopogon
(Fig. 1F). Roots of both the hosts were found sparsely
colonized by hyphae of DSE without appressoria in
the root segments. Primordia for microsclerotia were
also recorded in some of the root segments of R.
campanulatum. These structures were, however, not
observed in other species of rhododendrons.
4. Discussion
Due to their non-specific nature, arbuscular
mycorrhizae represent widely spread symbiotic
associations both geographically and in terms of the
number of plant host species involved. There are a few
reports on the occurrence of arbuscular mycorrhizae
(Cross, 1975; Koske et al., 1990; Dighton and
Coleman, 1992; Blaszkowski, 1994) in plants belong-
ing to the order Ericales, where ericoid type of
mycorrhizae is commonly found in their roots
(Duddrige and Read, 1982).
Mycorrhizal flora of rhododendrons, e.g., R.
ponticum, R. simsii, and the occurrence of arbuscular
mycorrhizal fungi and their use as inoculum for
enhancing their growth have been reported from
Europe (Cross, 1975; Dighton and Coleman, 1992).
The present study would appear to be the first detailed
report on the mycorrhizal associations found in all the
five species of rhododendrons found in Kumaun
region of the Indian Central Himalaya, representing
temperate to alpine locations.
This study describes the distribution of arbuscular
mycorrhizal fungi in the rhizosphere soil of rhodo-
dendrons along with details of externally associated
functional structures (extra radical mycelium, vesicles
and chlamydospores) as well as internal associations
(intraradical hyphal coiling, intraradical vesicles and
chlamydospores), except the arbuscules. The presence
of arbuscules in roots is generally used to associate
plants with functional arbuscular mycorrhizae. While
the arbuscules were almost absent in the root samples,
hyphal coils were commonly seen. The amount of
internal colonization by arbuscular mycorrhizal fungi
might depend on differences in the root morphology.
Smith and Smith (1997) reviewed structural diversity
of arbuscular mycorrhizal fungi and recognized two
types of colonizing pattern, viz., Arum-type and Paris-
type. In the Arum-type, extensive intercellular hyphae
and arbuscules develop, while in the Paris-type these
structures are absent and hyphal coils occur com-
monly. In the present investigation the Paris-type
colonization of arbuscular mycorrhizal fungi was
found to be prominent in all the five species of
rhododendrons. These structures may have a role in bi-
directional transfer of nutrients in the absence of
arbuscules.
The frequent occurrence of intraradical vesicles
(ellipsoid) indicated that a large part of the arbuscular
mycorrhizal fungi belonged to order Glomerales
(Schubler et al., 2001). Approximately 50% of total
isolated species were identified as species of Glomus;
G. fasciculatum being the most dominant. Predomi-
nance of G. fasciculatum, under various climatic
conditions, tropical to high arctic has been reported by
several workers (Dalpe and Aiken, 1998; Ragupathy
and Mahadevan, 1993; Chaurasia, 2000). The depen-
dence of spore population on pH of the soil was also
reflected in the present study. The spore population of

B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324 323
the total arbuscular mycorrhizal fungi was lower in
alpine region where the soil pH was relatively lower.
Although genus Glomus has been found to
dominate in the rhizosphere soil, the species
composition of arbuscular mycorrhizal fungi varies
in different rhododendrons. The study also reflected a
trend indicating decrease in the richness and diversity
of arbuscular mycorrhizal fungi with the increasing
altitude. It coincides with decreased richness and
diversity in the vegetational flora along an increase in
the altitude. The diversity of several groups of
terrestrial organisms has been found to be inversely
related with altitude (Gentry, 1988; Stevens, 1992).
Results of the present study showed that higher
arbuscular mycorrhizal diversity supports higher
aboveground diversity, as also proposed by van der
Heijden et al. (1998). According to them the below-
ground diversity of vesicular arbuscular mycorrhizal
fungi is a major factor in the maintenance of plant
diversity and ecosystem functioning.
In two rhododendron species, namely R. campa-
nulatum and R. anthopogon, presence of dark septate
fungi was also observed. Functional role of DS fungi
in rhododendrons is a matter of investigation. As in
the present work, DS fungal structures (intra cellular
mycelium, microsclerotium primordial) were also
reported from antarctic and sub-arctic sites (Christie
and Nicolson, 1983; Treu et al., 1996). The
occurrence of DS fungi in high stress environments
such as the alpine and arctic habitats strongly
suggests a more mutualistic role of these fungi
(Jumpponen, 2001).
The present study confirms the wide occurrence of
arbuscular mycorrhizal fungi in members of Erica-
ceae. It also extends the range of host plants of
arbuscular mycorrhizae which may play essential
ecological role in highly stressed environments like
arctic and alpine habitats where plant communities
experience short growing period, low temperature,
low nutrient status and low decomposition rates.
Acknowledgements
The Council of Scientific and Industrial Research
(CSIR), and the Union Ministry of Forests and
Environment, Government of India, New Delhi, are
thanked for the financial support.
References
Ahlich, K., Sieber, T.N., 1996. The profusion of dark septate
endophytic fungi in non-ectomycorrhizal fine roots of forest
trees and shrubs. New Phytol. 132, 259–270.
Blaszkowski, J., 1994. Arbuscular fungi and mycorrhizae (Glo-
males) of the Hel Peninsula, Poland. Mycorrhiza 5, 71–88.
Chaurasia, B., 2000. Ecological study of tropical forest trees with
special reference to vesicular–arbuscular-mycorrhizal (VAM)
association. Ph.D. Thesis. Dr. H. S. Gour University Sagar,
M.P., India, 172 pp.
Christie, P., Nicolson, T.H., 1983. Are mycorrhizas absent from the
Antarctic? Trans. Brit. Mycol. Soc. 80, 557–560.
Cross, J.R., 1975. Biological flora of the British Isles: Rhododendron
ponticum L. J. Ecol. 63, 345–364.
Dalpe, Y., Aiken, S.G., 1998. Arbuscular mycorrhizal fungi asso-
ciated with Festuca species in the Canadian High Arctic. Can. J.
Bot. 76, 1930–1938.
Dighton, J., Coleman, D.C., 1992. Phosphorus relations of roots and
mycorrhizas of Rhododendron maximum L. in the southern
Appalachians, North Carolina. Mycorrhiza 1, 175–184.
Duddrige, J., Read, D.J., 1982. An ultrastructural analysis of the
development of mycorrhizas in Rhododendron ponticum. Can. J.
Bot. 60, 2345–2356.
Franke-Synder, M., Douds Jr., D.D., Galvez, l., Phillips, J.G.,
Wagoner, P., Drinkwater, L., Morton, J.B., 2001. Diversity of
communities of arbuscular mycorrhizal (AM) fungi present in
conventional verses low-input agricultural sites in eastern Penn-
sylvania, USA. Appl. Soil Ecol. 16, 35–48.
Gerdemann, J.W., Nicolson, T.H., 1963. Spores of mycorrhizal
Endogone species extracted from soil by wet sieving and
decanting. Trans. Brit. Mycol. Soc. 46, 235–244.
Gentry, A.H., 1988. Changes in plant community diversity and
floristic composition on environmental and geographical gradi-
ent. Ann. Misso. Bot. Gard. 75, 1–34.
Gianinazzi, S., Gianinazzi-Pearson, V., 1986. Progress and head-
aches in endomycorrhiza biotechnology. Symbiosis 2, 139–149.
Gour, A., Adholeya, A., 1994. Estimation of VAMF spores in soil: a
modified method. Mycorrhiza News 6 (1), 10–11.
Hutchenson, K., 1970. A test for comparing diversities based on the
Shannon formula. Theor. Biol. 29, 151–154.
Jumpponen, A., 2001. Dark septate endophytes—are they
mycorrhizal? Mycorrhiza 11, 207–211.
Kormanik, P.P., Bryan, W.C., Schultz, R.C., 1980. Procedure and
equipment for staining large number of plant roots for endo-
mycorrhizal assay. Can. J. Microbiol. 26, 536–538.
Koske, R.E., Gemma, J.N., Englander, L., 1990. Vesicular–arbus-
cular mycorrhizae in Hawaiian Ericales. Am. J. Bot. 77, 64–68.
Largent, D.L., Sugihara, N., Wishner, C., 1980. Occurrence of
mycorrhizae on ericaceous and pyrolaceous plants in Northern
California. Can. J. Bot. 58, 2275–2279.
Mohankumar, V., Mahadevan, A., 1986. Survey of vesicular–arbus-
cular mycorrhizae in mangrove vegetation. Curr. Sci. 55,
936.
Mohankumar, V., Ragupathy, S., Nirmala, C.V., Mahadevan, A.,
1988. Distribution of vesicular–arbuscular mycorrhizae (VAM)
in the sandy beach soil of Madras coast. Curr. Sci. 57, 367–368.

B. Chaurasia et al. / Forest Ecology and Management 207 (2005) 315–324324
Muthukumar, T., Udaiyan, K., 2000. Arbuscular mycorrhizas of
plants growing in the Western Ghat region, southern India.
Mycorrhiza 9, 297–313.
O’Dell, T.E., Massicotte, H.B., Trappe, J.M., 1993. Root coloniza-
tion of Lupinus latifolius Agardh. and Pinus contorta Dougl. by
Phialocephala fortinii Wang & Wilcox. New Phytol. 124, 93–
100.
Ragupathy, S., Mahadevan, A., 1993. Distribution of vesicular–
arbuscular mycorrhizae in plants and rhizosphere soils of
the tropical plains, Tamilnadu, India. Mycorrhiza 3, 123–
136.
Read, D.J., Stribley, D.P., 1975. Some mycological aspects of the
biology of mycorrhiza in the Ericaceae. In: Sanders, F.E.,
Mosse, B., Tinker, P.B. (Eds.), Endomycorrhizas. Academic
Press, London, pp. 105–117.
Schenck, N.C., Perez, Y., 1990. A manual for identification of
vesicular–arbuscular mycorrhizal fungi. INVAM University of
Florida, Gainesville, FL, 286 pp.
Schubler, A., Schwarzott, D., Walker, C., 2001. A new fungal
phylum, the Glomeromycota: phylogeny and evolution. Mycol.
Res. 105 (12), 1413–1421.
Sengupta, A., Chaudhuri, S., 1990. Vesicular–arbuscular mycorrhi-
zal (VAM) fungi in pioneer salt marsh plants of the Ganges river
delta in West Bengal (India). Plant Soil 122, 111–113.
Simpson, E.H., 1949. Measurement of diversity. Nature 163–168.
Smith, F.A., Smith, S.E., 1997. Structural diversity in vesicular–
arbuscular mycorrhizal symbioses. New Phytol. 137, 373–388.
Smith, S.E., Read, D.J., 1997. Mycorrhizal Symbiosis, 2nd ed.
Academic Press, San Diego, 605 pp.
Stevens, G.C., 1992. The elevational gradient in altitudinal range: an
extension of Papoport’s latitudinal rule of altitude. Am. Nat. 140,
893–911.
Treu, R., Laursen, G.A., Stephenson, S.L., Landolt, J.C., Densmore,
R., 1996. Mycorrhizae from Denali National Park and Preserve,
Alaska. Mycorrhiza 6, 21–29.
van der Heijden, M.G.A., Klironomos, J.M., Ursie, M., Moutoglis,
P., Streitwolf-Engel, R., Boller, T., Wiemken, A., Sanders, I.R.,
1998. Mycorrhizal fungal diversity determines plant biodiver-
sity, ecosystem variability and productivity. Nature 396 (5), 69–
72.
Vestberg, M., 1995. Occurrence of some Glomales in Finland.
Mycorrhiza 5, 329–336.




















