
Distribution and functional characterization of equilibrative nucleoside transporter-4, a novel...
31
ISSN: 1524-4571 Copyright © 2006 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online TX 72514 Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas, DOI: 10.1161/01.RES.0000238359.18495.42 published online Jul 27, 2006; Circ. Res. Baldwin E. Cass, James D. Young, Rudi Billeter-Clark, Mark R. Boyett and Stephen A. R. Scullion, Lijie Sun, James Tellez, Mabel W.L. Ritzel, William C. Claycomb, Carol Kay Barnes, Halina Dobrzynski, Sophie Foppolo, Paul R. Beal, Fouzia Ismat, Elspeth Transporter-4, a Novel Cardiac Adenosine Transporter Activated at Acidic pH Distribution and Functional Characterization of Equilibrative Nucleoside http://circres.ahajournals.org/cgi/content/full/01.RES.0000238359.18495.42/DC1 Data Supplement (unedited) at: http://circres.ahajournals.org located on the World Wide Web at: The online version of this article, along with updated information and services, is http://www.lww.com/reprints Reprints: Information about reprints can be found online at [email protected] 410-528-8550. E-mail: Fax: Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters http://circres.ahajournals.org/subscriptions/ Subscriptions: Information about subscribing to Circulation Research is online at by on May 18, 2011 circres.ahajournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Distribution and functional characterization of equilibrative nucleoside transporter-4, a novel...

ISSN: 1524-4571 Copyright © 2006 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online
TX 72514Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas,
DOI: 10.1161/01.RES.0000238359.18495.42 published online Jul 27, 2006; Circ. Res.
Baldwin E. Cass, James D. Young, Rudi Billeter-Clark, Mark R. Boyett and Stephen A.
R. Scullion, Lijie Sun, James Tellez, Mabel W.L. Ritzel, William C. Claycomb, Carol Kay Barnes, Halina Dobrzynski, Sophie Foppolo, Paul R. Beal, Fouzia Ismat, Elspeth
Transporter-4, a Novel Cardiac Adenosine Transporter Activated at Acidic pHDistribution and Functional Characterization of Equilibrative Nucleoside
http://circres.ahajournals.org/cgi/content/full/01.RES.0000238359.18495.42/DC1Data Supplement (unedited) at:
http://circres.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circres.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation Research is online at
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Distribution and Functional Characterization ofEquilibrative Nucleoside Transporter-4, a Novel Cardiac
Adenosine Transporter Activated at Acidic pHKay Barnes, Halina Dobrzynski,* Sophie Foppolo,* Paul R. Beal,* Fouzia Ismat, Elspeth R. Scullion,Lijie Sun, James Tellez, Mabel W.L. Ritzel, William C. Claycomb, Carol E. Cass, James D. Young,
Rudi Billeter-Clark, Mark R. Boyett, Stephen A. Baldwin
Abstract—Adenosine plays multiple roles in the efficient functioning of the heart by regulating coronary blood flow,cardiac pacemaking, and contractility. Previous studies have implicated the equilibrative nucleoside transporter familymember equilibrative nucleoside transporter-1 (ENT1) in the regulation of cardiac adenosine levels. We report here thata second member of this family, ENT4, is also abundant in the heart, in particular in the plasma membranes ofventricular myocytes and vascular endothelial cells but, unlike ENT1, is virtually absent from the sinoatrial andatrioventricular nodes. Originally described as a monoamine/organic cation transporter, we found that both human andmouse ENT4 exhibited a novel, pH-dependent adenosine transport activity optimal at acidic pH (apparent Km values0.78 and 0.13 mmol/L, respectively, at pH 5.5) and absent at pH 7.4. In contrast, serotonin transport by ENT4 wasrelatively insensitive to pH. ENT4-mediated nucleoside transport was adenosine selective, sodium independent and onlyweakly inhibited by the classical inhibitors of equilibrative nucleoside transport, dipyridamole, dilazep, and nitroben-zylthioinosine. We hypothesize that ENT4, in addition to playing roles in cardiac serotonin transport, contributes to theregulation of extracellular adenosine concentrations, in particular under the acidotic conditions associated with ischemia.(Circ Res. 2006;99:0-0.)
Key Words: nucleoside � adenosine � transport � ischemia � pH
The purine nucleoside adenosine is produced by the action ofboth endo- and ecto-nucleotidases on adenine nucleotides in
the heart and plays key roles in the regulation of coronary bloodflow and myocardial O2 supply–demand balance.1–4 For exam-ple, action of adenosine on A2A receptors on vascular smoothmuscle and endothelial cells causes coronary vasodilatation.1,5 Incontrast, the negative inotropic and dromotropic effects ofadenosine on the heart are mediated primarily by A1 receptors.2
Similarly, the negative chromotropic effect of adenosine in-volves action of A1 receptors in the sinoatrial (SA) node on theinwardly rectifying potassium channel current IK-Ado and thehyperpolarization-activated pacemaker current If.2,6 Endogenousadenosine, acting on mitochondrial KATP channels via A1 and A3
receptors, also makes a major contribution to the phenomenon ofischemic preconditioning.5,7
Extracellular adenosine concentrations in the heart are gov-erned both by action of ecto-5�-nucleotidase on adenine nucle-otides released from cells and by transporter-mediated flux ofadenosine across cell membranes.3,4 Although most adenosine
production occurs intracellularly, under normoxic conditions,metabolism maintains a low intracellular concentration and,therefore, the net flux of adenosine is into cardiomyocytes andendothelial cells. Under such conditions, administration of trans-port inhibitors increases extracellular concentrations of adeno-sine, leading to vasodilatation.8 However, increased adeninenucleotide breakdown and inhibition of adenosine kinase duringhypoxia reverses the concentration gradient across membranes,resulting in nucleoside efflux.9 The importance of nucleosidetransporters in regulating extracellular adenosine concentrationsin the heart is demonstrated not only by the vasodilatory effectsof transport inhibitors but also by their effects on guinea pig SAand atrioventricular (AV) nodes, for which dipyridamole, apotent inhibitor of equilibrative transporters, was found topotentiate the chronotropic and dromotropic effects of adeno-sine, respectively.10
In mammalian cells adenosine transport is mediated bymembers of the SLC29 family of equilibrative nucleosidetransporters (ENTs)11 and the SLC28 family of concentrative
Original received February 22, 2006; revision received July 18, 2006; accepted July 19, 2006.From the Institute of Membrane and Systems Biology (K.B., S.F., P.R.B., F.I., E.R.S., L.S., R.B.-C., S.A.B.), University of Leeds, UK; Division of
Cardiovascular and Endocrine Sciences (H.D., J.T., M.R.B.), University of Manchester, UK; Membrane Protein Research Group (M.W.L.R., C.E.C.,J.D.Y.) and Departments of Physiology (M.W.L.R., J.D.Y.) and Oncology (C.E.C.), University of Alberta, and Cross Cancer Institute (C.E.C.),Edmonton, Alberta, Canada; and Department of Biochemistry and Molecular Biology (W.C.C.), Louisiana State, University Health Sciences Center, NewOrleans. 8Current address for P.R.B.: Department of Biology, University of York, UK.
*These authors contributed equally to this study.Correspondence to Kay Barnes, Institute of Membrane and Systems Biology, University of Leeds, Leeds LS2 9JT, UK. E-mail [email protected]© 2006 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000238359.18495.42
1 by on May 18, 2011 circres.ahajournals.orgDownloaded from

nucleoside transporters (CNTs).12 Two members of the latter,CNT2 and CNT3, exhibit robust adenosine transport activity,but substantial levels of mRNA have been reported in theheart only for CNT2.12 Moreover, Na�-dependent uptake hasbeen reported to represent only a small fraction of totaladenosine uptake in normal rat cardiomyocytes.13 Instead, themajority of published studies have reported equilibrative,Na�-independent uptake of adenosine into cardiac cells,consistent with involvement of members of the ENT family.
Mammalian genomes encode 4 members of the ENTfamily that share the ability to transport adenosine and alikely 11-transmembrane (TM) helix topology.11 One mem-ber, ENT3, although abundant in the heart, has been shown tobe a lysosomal transporter and, therefore, is unlikely tocontribute directly to regulation of interstitial adenosineconcentrations.14 However, the best-characterized familymembers, ENT1 and -2, are cell surface proteins that couldregulate access of adenosine to its receptors.11 Moreover,their activity appears to be subject to physiological regulationby the receptors themselves.15 These broad-selectivity equili-brative transporters differ in their sensitivity to the nucleosideanalog nitrobenzylthioinosine (nitrobenzylmercaptopurine ri-bonucleoside [NBMPR]), which inhibits ENT1-mediatedtransport with a Ki value of �5 nmol/L, whereas ENT2-mediated transport is only weakly inhibited.11 Human ENT1(hENT1) is also more potently inhibited than human ENT2(hENT2) by coronary vasodilators such as dipyridamole,dilazep, and draflazine. The corresponding rodent transport-ers are much less sensitive to inhibition by such agents.11 Thetransport characteristics of pig and human coronary smoothmuscle cells,16 guinea pig cardiac endothelial cells,17 and ratcardiomyocytes18,19 suggest that adenosine flux is mediatedprimarily by ENT1, although low levels of NBMPR-insensitive transport in cardiomyocytes have recently beenattributed to ENT2.13 Consistent with these findings, we haveshown that ENT1 is abundantly expressed in atrial andventricular myocytes and in the SA node of the rat heart.20,21
The fourth member of the SLC29 family to be identified,ENT4, exhibits only low sequence identity (�20%) to theother family members, indicating an ancient divergence fromthese isoforms.22 Although in a preliminary study we reportedthat this protein was a low-affinity adenosine transporter,11 itwas subsequently found by Wang and coworkers to functionas a polyspecific organic cation transporter and was desig-nated plasma membrane monoamine transporter (PMAT).23,24
These authors reported that PMAT exhibited no significantinteraction with nucleosides but efficiently transported sero-tonin (5-hydroxytryptamine [5-HT]).24 Although most highlyexpressed in brain and skeletal muscle, Northern blottingrevealed significant expression in heart. Serotonin is pro-duced by cardiac myocytes25 and, by acting on 5-HT2B
receptors, regulates cardiac development as well as cardiacstructure and function in adults.26 Given the potential impor-tance of serotonin in the heart, and our preliminary findingthat ENT4 transported adenosine at acidic pH but not at pH7.4, the present study was undertaken to examine the func-tional properties of the human and rodent transporters andtheir tissue distributions in more detail. We confirm thatadenosine is efficiently transported by ENT4, but in a fashion
highly sensitive to extracellular pH. Although its activity hasnot been measured in situ, its cardiac abundance suggests acontribution to regulation of extracellular adenosine concen-trations, in particular during ischemia.
Materials and MethodsFor expression in Xenopus oocytes the coding regions of mouseENT4 (mENT4) and hENT cDNAs were subcloned into the vectorpGEM-HE.27 Production of ENT4 proteins in Xenopus oocytes,assays of nucleoside and serotonin uptake activity, and preparationof oocyte membranes were performed as previously described.14,20
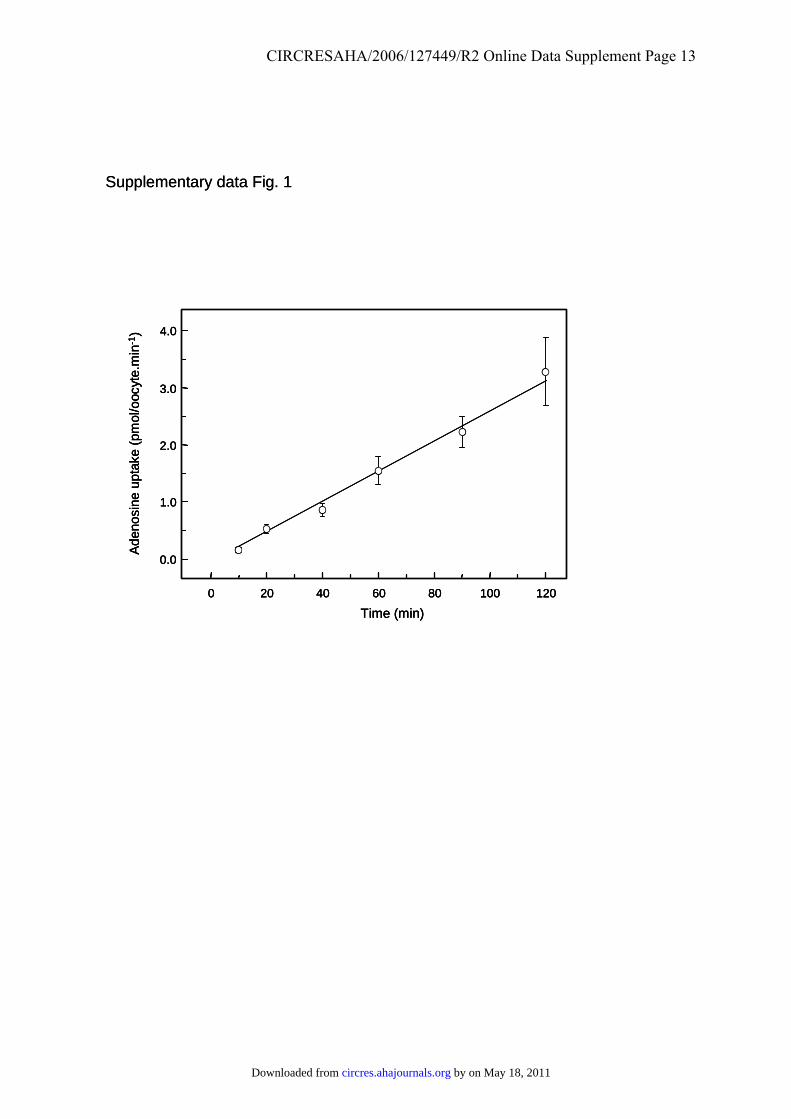
Uptake was typically measured for 30 minutes, during which periodrates were linear (Figure I in the online data supplement). HL-1cardiomyocytes were cultured as described elsewhere28 and seroto-nin uptake measured at 37°C on cells at 80% confluence. Adenosineuptake into rat ventricular myocytes was measured at 37°C. Humanumbilical vein endothelial cells (HUVECs) were cultured as de-scribed by Herbert et al.29
Tissue distribution of hENT4 transcripts was investigated byprobing a human multiple tissue expression (MTE mRNA array;Clontech) with an [�-32P]dATP-labeled antisense DNA probe.14 ForWestern blotting and immunocytochemistry, anti–synthetic peptidebodies, designated anti-hENT4301–319, were raised in rabbits byPepceuticals Ltd (Leicester, UK) and affinity purified by chroma-tography on a column of immobilized peptide.30 Rat, mouse, andhuman tissue lysates and membrane samples for blotting wereprepared as previously described21 and are detailed in the online datasupplement. Permission to use human tissue was granted by theLeeds (West) Research Ethics Committee. Deglycosylation withN-glycosidase F was performed according to the instructions of themanufacturer (Roche Applied Sciences). For quantification of ENT4,known amounts of a glutathione S-transferase (GST) fusion proteinbearing residues 252 to 347 of mENT4 were included as standards onblots.
The subcellular distribution of ENT4 in primary rat cardiomyo-cytes and cultured cells was investigated by immunofluorescencemicroscopy with anti-hENT4301–319, using an Olympus IX70 micro-scope equipped with a Delta Vision deconvolution system.14 A ZeissLSM510 META laser scanning confocal microscope was used forimmunofluorescence microscopy of tissue sections.21
An expanded Materials and Methods section is available in theonline data supplement at http://circres.ahajournals.org.
ResultsTissue Distribution and Glycosylation Stateof ENT4Northern blotting previously revealed abundant expression ofhENT4 (PMAT) mRNA in adult human brain and skeletalmuscle.24 In the present study, probing a multiple tissueexpression RNA array derived from 76 human tissues andcell types with a hENT4 oligonucleotide probe confirmedabundant expression of hENT4 transcripts in various brainregions and skeletal muscle (supplemental Figure II). Sub-stantial levels of hENT4 mRNA were also found in manyother adult and fetal tissues, with particularly high levels inregions of adult heart and intestine, as well as in pancreas,kidney, liver, bone marrow, and lymph node.
To investigate whether the abundance of hENT4 transcripts inheart and other tissues was paralleled by an abundance of theprotein itself, antibodies were raised against residues 301 to 319of hENT4. This region exhibits 63% and 68% identity to thecorresponding regions of mENT4 and rENT4 (GenBank acces-sion no. XP_221940) (Figure 1A). Anti-hENT4301–319 stained asingle major band of size �55kDa (predicted size 58.1 kDa) inmembranes prepared from Xenopus oocytes injected with
2 Circulation Research September 1, 2006
by on May 18, 2011 circres.ahajournals.orgDownloaded from
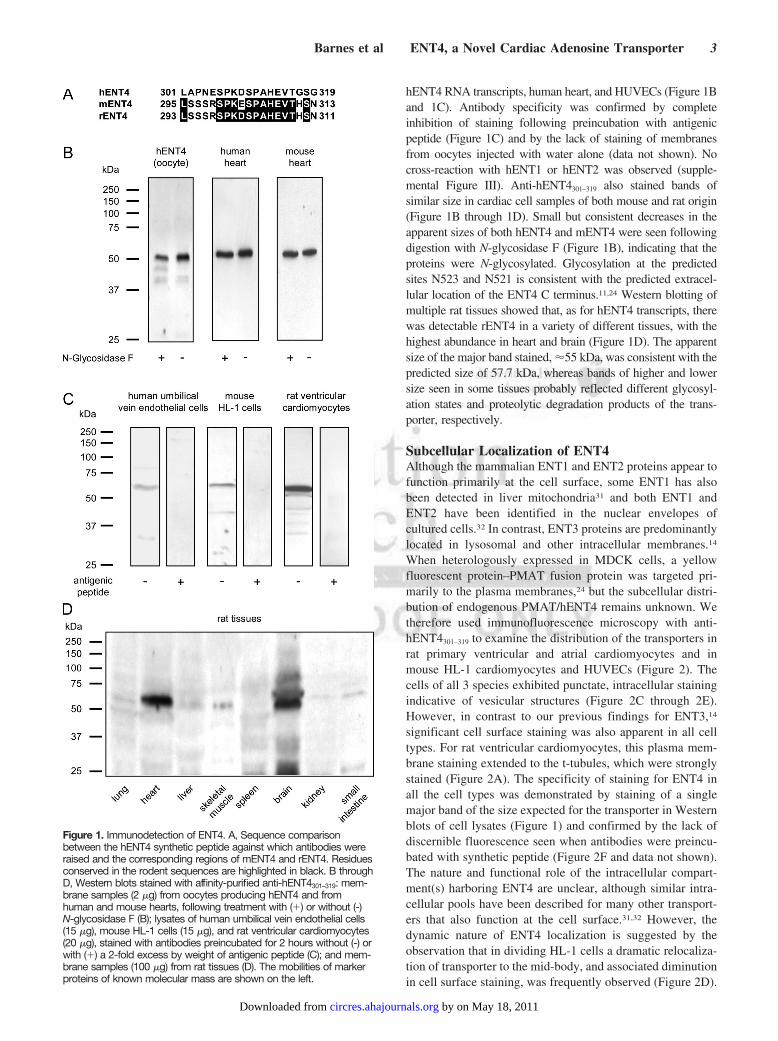
hENT4 RNA transcripts, human heart, and HUVECs (Figure 1Band 1C). Antibody specificity was confirmed by completeinhibition of staining following preincubation with antigenicpeptide (Figure 1C) and by the lack of staining of membranesfrom oocytes injected with water alone (data not shown). Nocross-reaction with hENT1 or hENT2 was observed (supple-mental Figure III). Anti-hENT4301–319 also stained bands ofsimilar size in cardiac cell samples of both mouse and rat origin(Figure 1B through 1D). Small but consistent decreases in theapparent sizes of both hENT4 and mENT4 were seen followingdigestion with N-glycosidase F (Figure 1B), indicating that theproteins were N-glycosylated. Glycosylation at the predictedsites N523 and N521 is consistent with the predicted extracel-lular location of the ENT4 C terminus.11,24 Western blotting ofmultiple rat tissues showed that, as for hENT4 transcripts, therewas detectable rENT4 in a variety of different tissues, with thehighest abundance in heart and brain (Figure 1D). The apparentsize of the major band stained, �55 kDa, was consistent with thepredicted size of 57.7 kDa, whereas bands of higher and lowersize seen in some tissues probably reflected different glycosyl-ation states and proteolytic degradation products of the trans-porter, respectively.
Subcellular Localization of ENT4Although the mammalian ENT1 and ENT2 proteins appear tofunction primarily at the cell surface, some ENT1 has alsobeen detected in liver mitochondria31 and both ENT1 andENT2 have been identified in the nuclear envelopes ofcultured cells.32 In contrast, ENT3 proteins are predominantlylocated in lysosomal and other intracellular membranes.14
When heterologously expressed in MDCK cells, a yellowfluorescent protein–PMAT fusion protein was targeted pri-marily to the plasma membranes,24 but the subcellular distri-bution of endogenous PMAT/hENT4 remains unknown. Wetherefore used immunofluorescence microscopy with anti-hENT4301–319 to examine the distribution of the transporters inrat primary ventricular and atrial cardiomyocytes and inmouse HL-1 cardiomyocytes and HUVECs (Figure 2). Thecells of all 3 species exhibited punctate, intracellular stainingindicative of vesicular structures (Figure 2C through 2E).However, in contrast to our previous findings for ENT3,14
significant cell surface staining was also apparent in all celltypes. For rat ventricular cardiomyocytes, this plasma mem-brane staining extended to the t-tubules, which were stronglystained (Figure 2A). The specificity of staining for ENT4 inall the cell types was demonstrated by staining of a singlemajor band of the size expected for the transporter in Westernblots of cell lysates (Figure 1) and confirmed by the lack ofdiscernible fluorescence seen when antibodies were preincu-bated with synthetic peptide (Figure 2F and data not shown).The nature and functional role of the intracellular compart-ment(s) harboring ENT4 are unclear, although similar intra-cellular pools have been described for many other transport-ers that also function at the cell surface.31,32 However, thedynamic nature of ENT4 localization is suggested by theobservation that in dividing HL-1 cells a dramatic relocaliza-tion of transporter to the mid-body, and associated diminutionin cell surface staining, was frequently observed (Figure 2D).
Figure 1. Immunodetection of ENT4. A, Sequence comparisonbetween the hENT4 synthetic peptide against which antibodies wereraised and the corresponding regions of mENT4 and rENT4. Residuesconserved in the rodent sequences are highlighted in black. B throughD, Western blots stained with affinity-purified anti-hENT4301–319: mem-brane samples (2 �g) from oocytes producing hENT4 and fromhuman and mouse hearts, following treatment with (�) or without (-)N-glycosidase F (B); lysates of human umbilical vein endothelial cells(15 �g), mouse HL-1 cells (15 �g), and rat ventricular cardiomyocytes(20 �g), stained with antibodies preincubated for 2 hours without (-) orwith (�) a 2-fold excess by weight of antigenic peptide (C); and mem-brane samples (100 �g) from rat tissues (D). The mobilities of markerproteins of known molecular mass are shown on the left.
Barnes et al ENT4, a Novel Cardiac Adenosine Transporter 3
by on May 18, 2011 circres.ahajournals.orgDownloaded from
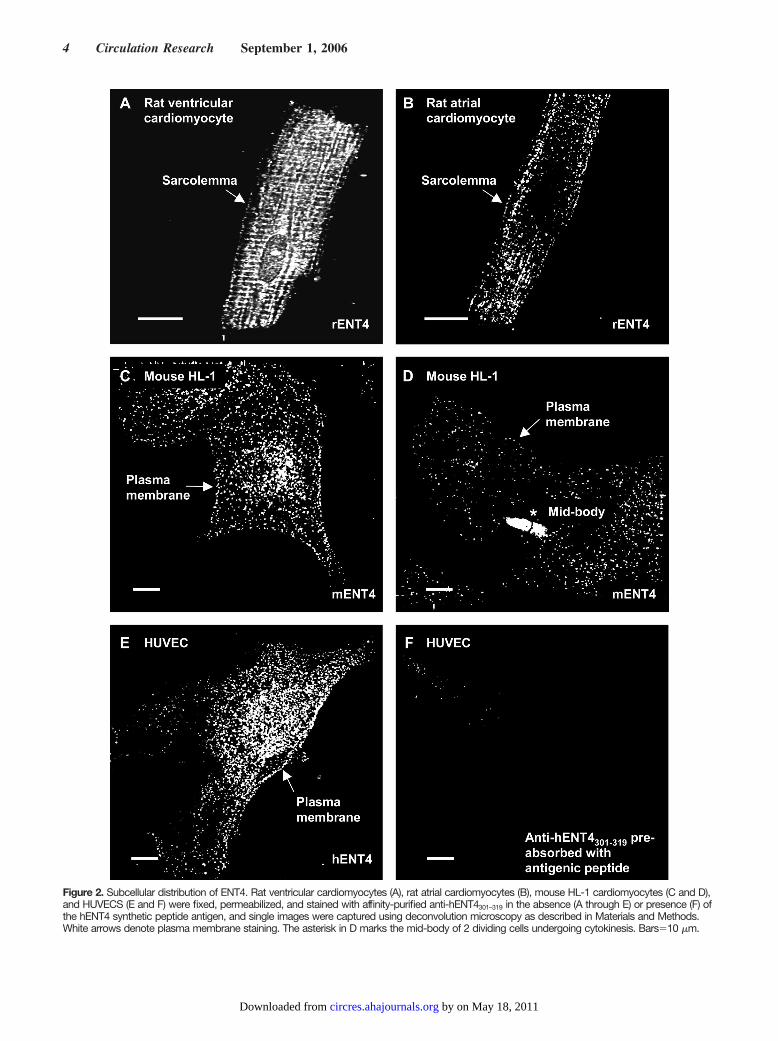
Figure 2. Subcellular distribution of ENT4. Rat ventricular cardiomyocytes (A), rat atrial cardiomyocytes (B), mouse HL-1 cardiomyocytes (C and D),and HUVECS (E and F) were fixed, permeabilized, and stained with affinity-purified anti-hENT4301–319 in the absence (A through E) or presence (F) ofthe hENT4 synthetic peptide antigen, and single images were captured using deconvolution microscopy as described in Materials and Methods.White arrows denote plasma membrane staining. The asterisk in D marks the mid-body of 2 dividing cells undergoing cytokinesis. Bars�10 �m.
4 Circulation Research September 1, 2006
by on May 18, 2011 circres.ahajournals.orgDownloaded from
Kinetic Properties of hENT4 and mENT4The localization of ENT4 in cardiomyocytes and culturedcells suggested that it functioned at least in part to transportpermeants across plasma membranes. When expressed inMDCK cells, hENT4/PMAT was reported to function as apolyspecific transporter of organic cations, rather than ofnucleosides.23,24 Our preliminary measurements of transportat pH 7.5 in Xenopus oocytes producing mENT4 and hENT4revealed mediated transport of adenosine, although at lowactivities.11 Given the presence of an intracellular pool ofENT4 (Figure 2) and our recent finding that the lysosomalnucleoside transporter ENT3 is optimally active at acid pH,14
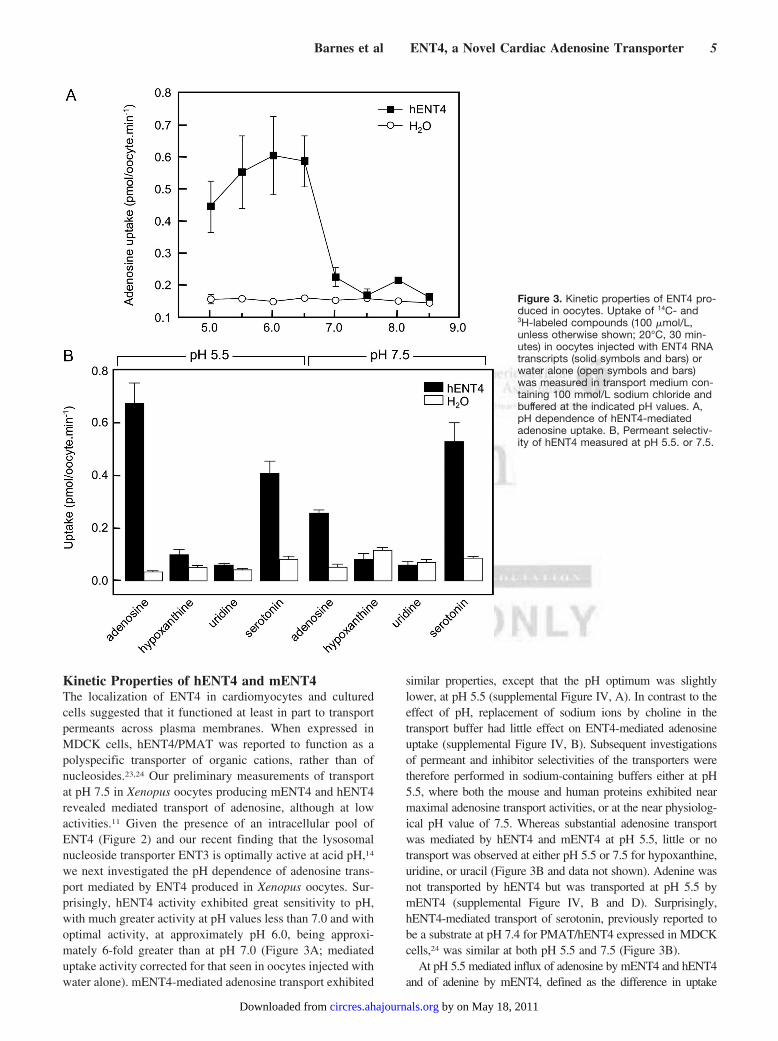
we next investigated the pH dependence of adenosine trans-port mediated by ENT4 produced in Xenopus oocytes. Sur-prisingly, hENT4 activity exhibited great sensitivity to pH,with much greater activity at pH values less than 7.0 and withoptimal activity, at approximately pH 6.0, being approxi-mately 6-fold greater than at pH 7.0 (Figure 3A; mediateduptake activity corrected for that seen in oocytes injected withwater alone). mENT4-mediated adenosine transport exhibited
similar properties, except that the pH optimum was slightlylower, at pH 5.5 (supplemental Figure IV, A). In contrast to theeffect of pH, replacement of sodium ions by choline in thetransport buffer had little effect on ENT4-mediated adenosineuptake (supplemental Figure IV, B). Subsequent investigationsof permeant and inhibitor selectivities of the transporters weretherefore performed in sodium-containing buffers either at pH5.5, where both the mouse and human proteins exhibited nearmaximal adenosine transport activities, or at the near physiolog-ical pH value of 7.5. Whereas substantial adenosine transportwas mediated by hENT4 and mENT4 at pH 5.5, little or notransport was observed at either pH 5.5 or 7.5 for hypoxanthine,uridine, or uracil (Figure 3B and data not shown). Adenine wasnot transported by hENT4 but was transported at pH 5.5 bymENT4 (supplemental Figure IV, B and D). Surprisingly,hENT4-mediated transport of serotonin, previously reported tobe a substrate at pH 7.4 for PMAT/hENT4 expressed in MDCKcells,24 was similar at both pH 5.5 and 7.5 (Figure 3B).
At pH 5.5 mediated influx of adenosine by mENT4 and hENT4and of adenine by mENT4, defined as the difference in uptake
Figure 3. Kinetic properties of ENT4 pro-duced in oocytes. Uptake of 14C- and3H-labeled compounds (100 �mol/L,unless otherwise shown; 20°C, 30 min-utes) in oocytes injected with ENT4 RNAtranscripts (solid symbols and bars) orwater alone (open symbols and bars)was measured in transport medium con-taining 100 mmol/L sodium chloride andbuffered at the indicated pH values. A,pH dependence of hENT4-mediatedadenosine uptake. B, Permeant selectiv-ity of hENT4 measured at pH 5.5. or 7.5.
Barnes et al ENT4, a Novel Cardiac Adenosine Transporter 5
by on May 18, 2011 circres.ahajournals.orgDownloaded from
between RNA-injected and water-injected oocytes, was saturableand conformed to simple Michaelis–Menten kinetics (supplementalFigures IV, C and D, and V, A). In the case of mENT4, the apparentKm values for adenosine and adenine transport were 130�30 �mol/L and 2600�500 �mol/L, respectively. To facilitate a
comparison of the ability of hENT4 to transport serotonin andadenosine, measurements were made using the same batch ofoocytes (supplemental Figure V, A and B). These yielded apparentKm values of 780�240 �mol/L and 1900�220 �mol/L for aden-osine and serotonin, respectively. The corresponding Vmax valueswere 7.7�0.7 and 8.9�0.4 pmol/oocyte � min�1, respectively. Itfollows that at acidic pH the transport efficiency (Vmax/Km) ofhENT4 for adenosine was approximately twice that for seroto-nin. The higher Km for serotonin than reported for PMATexpressed in MDCK cells (110�12 �mol/L24) may reflectdifferences both in the expression systems used and the pHvalues at which transport was measured.
Sensitivity of ENT4 to Transport InhibitorsMammalian ENT proteins differ in their sensitivities toinhibition by nucleoside analogs and coronary vasodilators.
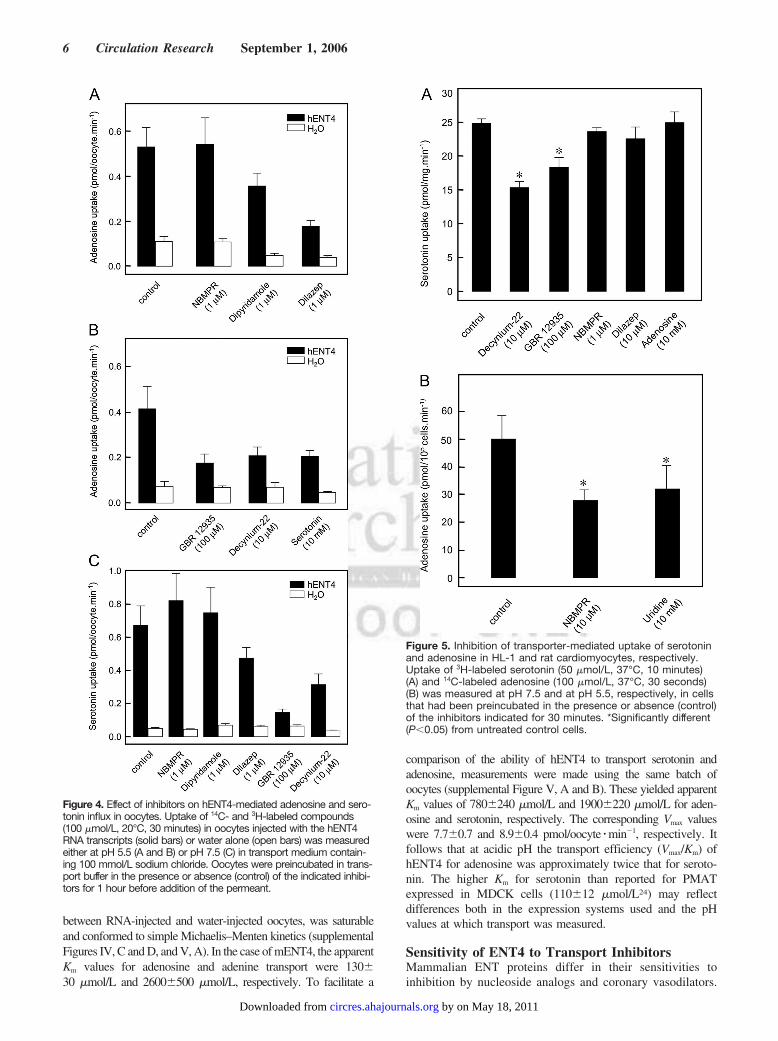
Figure 4. Effect of inhibitors on hENT4-mediated adenosine and sero-tonin influx in oocytes. Uptake of 14C- and 3H-labeled compounds(100 �mol/L, 20°C, 30 minutes) in oocytes injected with the hENT4RNA transcripts (solid bars) or water alone (open bars) was measuredeither at pH 5.5 (A and B) or pH 7.5 (C) in transport medium contain-ing 100 mmol/L sodium chloride. Oocytes were preincubated in trans-port buffer in the presence or absence (control) of the indicated inhibi-tors for 1 hour before addition of the permeant.
Figure 5. Inhibition of transporter-mediated uptake of serotoninand adenosine in HL-1 and rat cardiomyocytes, respectively.Uptake of 3H-labeled serotonin (50 �mol/L, 37°C, 10 minutes)(A) and 14C-labeled adenosine (100 �mol/L, 37°C, 30 seconds)(B) was measured at pH 7.5 and at pH 5.5, respectively, in cellsthat had been preincubated in the presence or absence (control)of the inhibitors indicated for 30 minutes. *Significantly different(P�0.05) from untreated control cells.
6 Circulation Research September 1, 2006
by on May 18, 2011 circres.ahajournals.orgDownloaded from
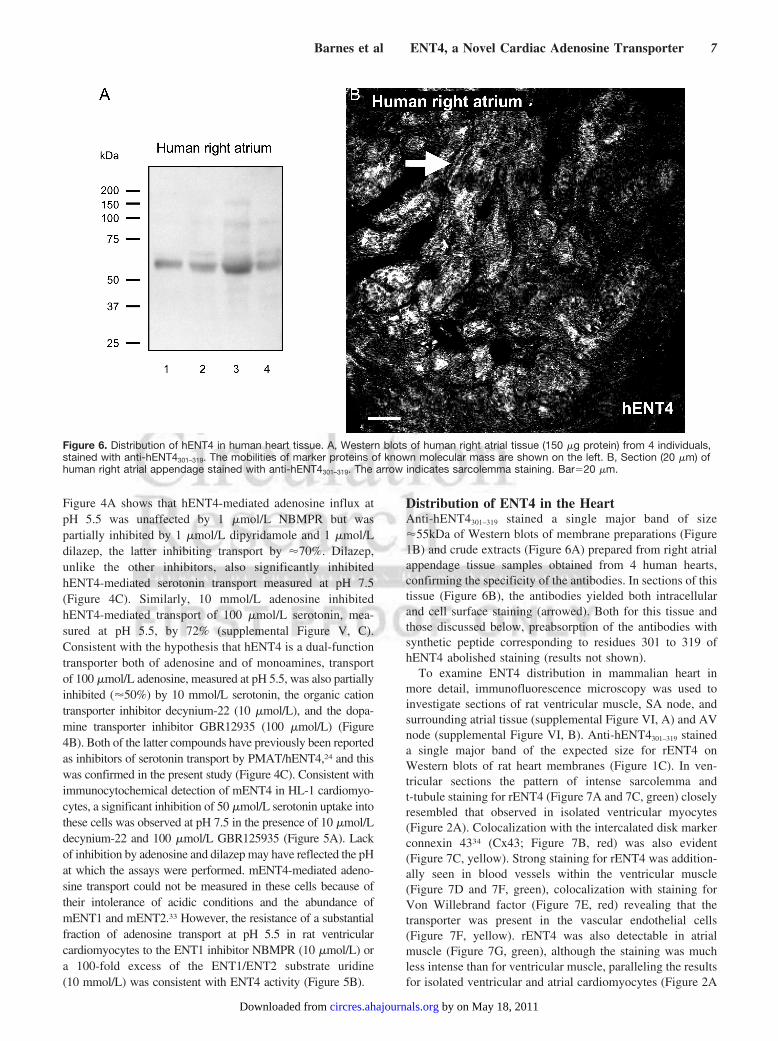
Figure 4A shows that hENT4-mediated adenosine influx atpH 5.5 was unaffected by 1 �mol/L NBMPR but waspartially inhibited by 1 �mol/L dipyridamole and 1 �mol/Ldilazep, the latter inhibiting transport by �70%. Dilazep,unlike the other inhibitors, also significantly inhibitedhENT4-mediated serotonin transport measured at pH 7.5(Figure 4C). Similarly, 10 mmol/L adenosine inhibitedhENT4-mediated transport of 100 �mol/L serotonin, mea-sured at pH 5.5, by 72% (supplemental Figure V, C).Consistent with the hypothesis that hENT4 is a dual-functiontransporter both of adenosine and of monoamines, transportof 100 �mol/L adenosine, measured at pH 5.5, was also partiallyinhibited (�50%) by 10 mmol/L serotonin, the organic cationtransporter inhibitor decynium-22 (10 �mol/L), and the dopa-mine transporter inhibitor GBR12935 (100 �mol/L) (Figure4B). Both of the latter compounds have previously been reportedas inhibitors of serotonin transport by PMAT/hENT4,24 and thiswas confirmed in the present study (Figure 4C). Consistent withimmunocytochemical detection of mENT4 in HL-1 cardiomyo-cytes, a significant inhibition of 50 �mol/L serotonin uptake intothese cells was observed at pH 7.5 in the presence of 10 �mol/Ldecynium-22 and 100 �mol/L GBR125935 (Figure 5A). Lackof inhibition by adenosine and dilazep may have reflected the pHat which the assays were performed. mENT4-mediated adeno-sine transport could not be measured in these cells because oftheir intolerance of acidic conditions and the abundance ofmENT1 and mENT2.33 However, the resistance of a substantialfraction of adenosine transport at pH 5.5 in rat ventricularcardiomyocytes to the ENT1 inhibitor NBMPR (10 �mol/L) ora 100-fold excess of the ENT1/ENT2 substrate uridine(10 mmol/L) was consistent with ENT4 activity (Figure 5B).
Distribution of ENT4 in the HeartAnti-hENT4301–319 stained a single major band of size�55kDa of Western blots of membrane preparations (Figure1B) and crude extracts (Figure 6A) prepared from right atrialappendage tissue samples obtained from 4 human hearts,confirming the specificity of the antibodies. In sections of thistissue (Figure 6B), the antibodies yielded both intracellularand cell surface staining (arrowed). Both for this tissue andthose discussed below, preabsorption of the antibodies withsynthetic peptide corresponding to residues 301 to 319 ofhENT4 abolished staining (results not shown).
To examine ENT4 distribution in mammalian heart inmore detail, immunofluorescence microscopy was used toinvestigate sections of rat ventricular muscle, SA node, andsurrounding atrial tissue (supplemental Figure VI, A) and AVnode (supplemental Figure VI, B). Anti-hENT4301–319 staineda single major band of the expected size for rENT4 onWestern blots of rat heart membranes (Figure 1C). In ven-tricular sections the pattern of intense sarcolemma andt-tubule staining for rENT4 (Figure 7A and 7C, green) closelyresembled that observed in isolated ventricular myocytes(Figure 2A). Colocalization with the intercalated disk markerconnexin 4334 (Cx43; Figure 7B, red) was also evident(Figure 7C, yellow). Strong staining for rENT4 was addition-ally seen in blood vessels within the ventricular muscle(Figure 7D and 7F, green), colocalization with staining forVon Willebrand factor (Figure 7E, red) revealing that thetransporter was present in the vascular endothelial cells(Figure 7F, yellow). rENT4 was also detectable in atrialmuscle (Figure 7G, green), although the staining was muchless intense than for ventricular muscle, paralleling the resultsfor isolated ventricular and atrial cardiomyocytes (Figure 2A
Figure 6. Distribution of hENT4 in human heart tissue. A, Western blots of human right atrial tissue (150 �g protein) from 4 individuals,stained with anti-hENT4301–319. The mobilities of marker proteins of known molecular mass are shown on the left. B, Section (20 �m) ofhuman right atrial appendage stained with anti-hENT4301–319. The arrow indicates sarcolemma staining. Bar�20 �m.
Barnes et al ENT4, a Novel Cardiac Adenosine Transporter 7
by on May 18, 2011 circres.ahajournals.orgDownloaded from
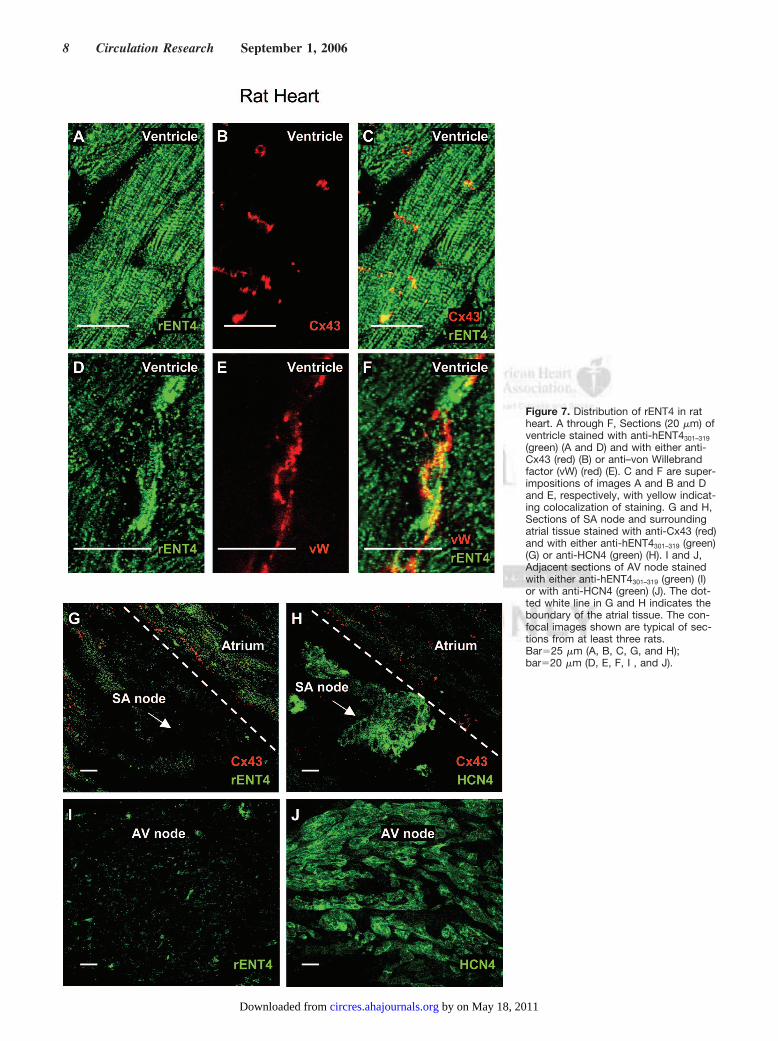
Figure 7. Distribution of rENT4 in ratheart. A through F, Sections (20 �m) ofventricle stained with anti-hENT4301–319
(green) (A and D) and with either anti-Cx43 (red) (B) or anti–von Willebrandfactor (vW) (red) (E). C and F are super-impositions of images A and B and Dand E, respectively, with yellow indicat-ing colocalization of staining. G and H,Sections of SA node and surroundingatrial tissue stained with anti-Cx43 (red)and with either anti-hENT4301–319 (green)(G) or anti-HCN4 (green) (H). I and J,Adjacent sections of AV node stainedwith either anti-hENT4301–319 (green) (I)or with anti-HCN4 (green) (J). The dot-ted white line in G and H indicates theboundary of the atrial tissue. The con-focal images shown are typical of sec-tions from at least three rats.Bar�25 �m (A, B, C, G, and H);bar�20 �m (D, E, F, I , and J).
8 Circulation Research September 1, 2006
by on May 18, 2011 circres.ahajournals.orgDownloaded from
and 2B) and probably reflecting the relative paucity oft-tubules in the latter. In contrast, little or no rENT4 stainingwas observed in cells of the SA and AV nodes (Figure 7G and7I, respectively, green). The latter were identified in closelyadjacent sections by their strong staining by antibodiesagainst the hyperpolarization-activated channel HCN435 (Fig-ure 7H and 7J, respectively, green) and their lack of stainingfor Cx4334 (Figure 7G and 7H, respectively, and data notshown).
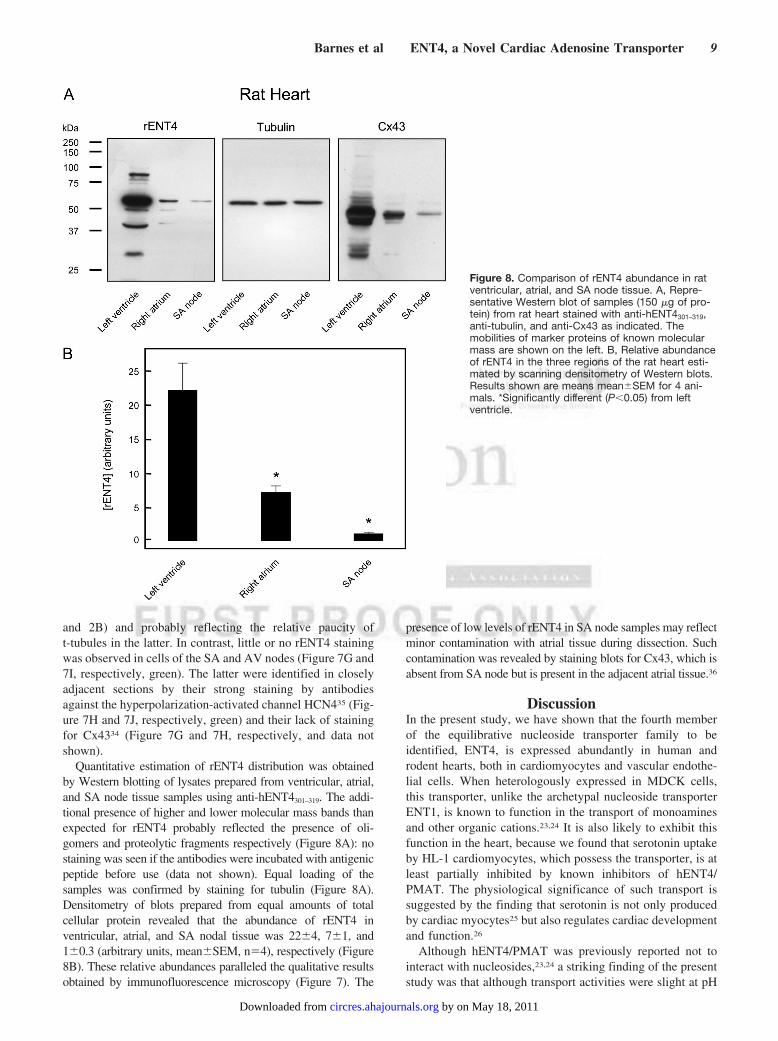
Quantitative estimation of rENT4 distribution was obtainedby Western blotting of lysates prepared from ventricular, atrial,and SA node tissue samples using anti-hENT4301–319. The addi-tional presence of higher and lower molecular mass bands thanexpected for rENT4 probably reflected the presence of oli-gomers and proteolytic fragments respectively (Figure 8A): nostaining was seen if the antibodies were incubated with antigenicpeptide before use (data not shown). Equal loading of thesamples was confirmed by staining for tubulin (Figure 8A).Densitometry of blots prepared from equal amounts of totalcellular protein revealed that the abundance of rENT4 inventricular, atrial, and SA nodal tissue was 22�4, 7�1, and1�0.3 (arbitrary units, mean�SEM, n�4), respectively (Figure8B). These relative abundances paralleled the qualitative resultsobtained by immunofluorescence microscopy (Figure 7). The
presence of low levels of rENT4 in SA node samples may reflectminor contamination with atrial tissue during dissection. Suchcontamination was revealed by staining blots for Cx43, which isabsent from SA node but is present in the adjacent atrial tissue.36
DiscussionIn the present study, we have shown that the fourth memberof the equilibrative nucleoside transporter family to beidentified, ENT4, is expressed abundantly in human androdent hearts, both in cardiomyocytes and vascular endothe-lial cells. When heterologously expressed in MDCK cells,this transporter, unlike the archetypal nucleoside transporterENT1, is known to function in the transport of monoaminesand other organic cations.23,24 It is also likely to exhibit thisfunction in the heart, because we found that serotonin uptakeby HL-1 cardiomyocytes, which possess the transporter, is atleast partially inhibited by known inhibitors of hENT4/PMAT. The physiological significance of such transport issuggested by the finding that serotonin is not only producedby cardiac myocytes25 but also regulates cardiac developmentand function.26
Although hENT4/PMAT was previously reported not tointeract with nucleosides,23,24 a striking finding of the presentstudy was that although transport activities were slight at pH
Figure 8. Comparison of rENT4 abundance in ratventricular, atrial, and SA node tissue. A, Repre-sentative Western blot of samples (150 �g of pro-tein) from rat heart stained with anti-hENT4301–319,anti-tubulin, and anti-Cx43 as indicated. Themobilities of marker proteins of known molecularmass are shown on the left. B, Relative abundanceof rENT4 in the three regions of the rat heart esti-mated by scanning densitometry of Western blots.Results shown are means mean�SEM for 4 ani-mals. *Significantly different (P�0.05) from leftventricle.
Barnes et al ENT4, a Novel Cardiac Adenosine Transporter 9
by on May 18, 2011 circres.ahajournals.orgDownloaded from

7.5, both hENT4 and mENT4 exhibited robust adenosinetransport activity at lower pH values, with hENT4 exhibitingmaximal activity in the pH range 5.5 to 6.5 (Figure 3A).Indeed, at pH 5.5, the transport efficiency (Vmax/Km) of hENT4for adenosine was greater than that for serotonin. Transportwas not dependent on sodium ions, was unaffected byNBMPR, and was only partially inhibited by dipyridamoleand dilazep at concentrations (1 �mol/L) that potently inhibitthe archetypal equilibrative nucleoside transporter hENT1.
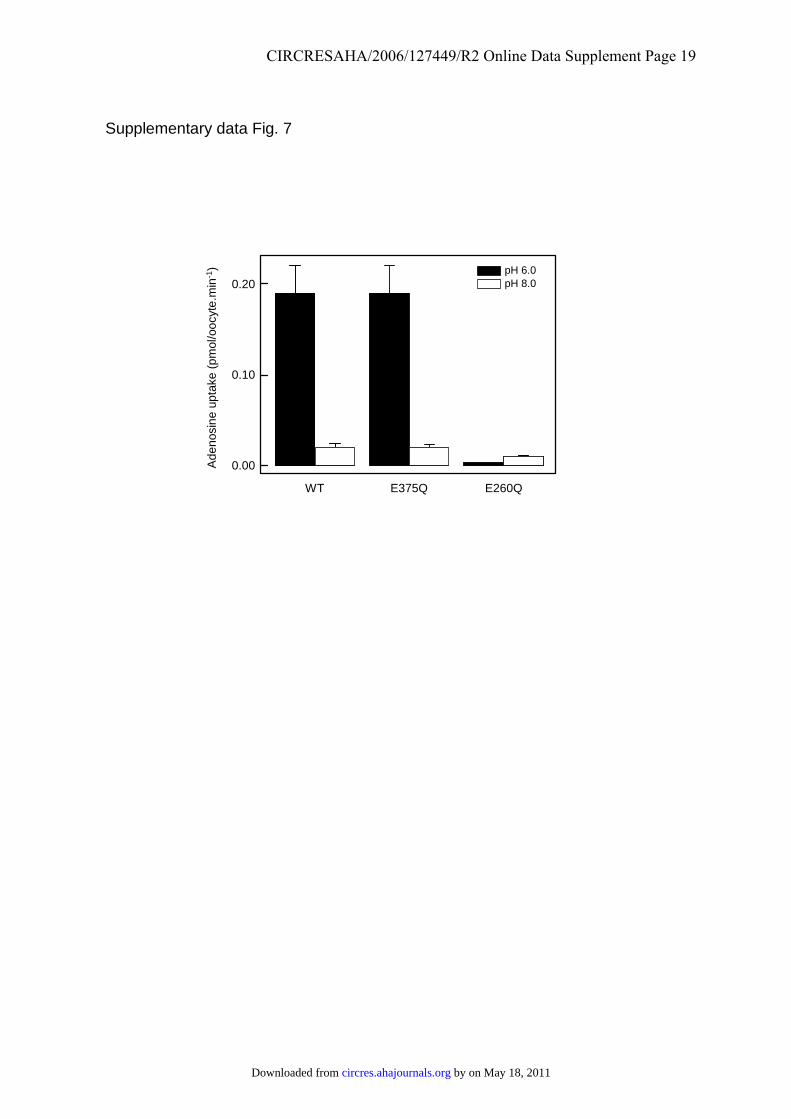
From a mechanistic point of view, it remains unclearwhether the pH dependence of transport reflects nucleoside/proton cotransport. However, it is noteworthy that serotonintransport, which has been reported to be sensitive to mem-brane potential but does not involve cotransport of ions,24 wasrelatively insensitive to pH. Nor is it clear which residue(s) inhENT4 is responsible for the pH sensitivity of adenosinetransport. In a preliminary attempt to identify such residues,we examined the effects of mutating the ionizable residuesE206 in putative TM5 and E375 in putative TM7 to glu-tamine: these positions are also occupied by glutamate inclose homologs of hENT4 but are occupied by unchargedresidues, typically glutamine or threonine, respectively, inother ENT family members, including hENT1 and hENT2.Although both mutants were expressed in oocytes at nearwild-type levels (data not shown), E206Q proved to beessentially inactive, whereas the activity and pH sensitivity ofE375Q resembled that of the wild-type protein (supplementalFigure VII). Although this finding highlights the importanceof E206 in transport, further studies will be necessary toidentify the residue(s) responsible for pH sensitivity.
In rat heart, rENT4 was found to be most abundant inventricular cardiomyocytes, which represent a major site ofadenosine production.8 Interestingly, it was absent from boththe SA and AV nodes, in contrast to rENT1, which we havepreviously shown is abundant in the SA node.21 It was alsoabundantly expressed in vascular endothelial cells, whichrepresent a major site for adenosine metabolism.8 Immuno-cytochemistry of isolated cardiomyocytes and cultured endo-thelial cells revealed that the protein was present at the cellsurface, where it could mediate adenosine flux and thusinfluence the extracellular concentration of this key purine.Myocardial ischemia and anaerobic metabolism followingcardiac arrest can lead to a rapid fall in interstitial fluid pH tovalues as low as pH 6.6.37 Thus, although the adenosinetransport activity of ENT4 is optimal at acidic pH andvirtually absent at pH 7.4, our findings on cardiomyocytesand ENT4 produced in oocytes suggest it is likely to play animportant role in ischemic conditions. Given the importanceof endogenous adenosine in the phenomenon of ischemicpreconditioning,5,7 and the observation that oral therapy withthe ENT1 inhibitor dipyridamole limits reperfusion injury inhumans,38 ENT4 represents a possible future therapeutictarget for cardiac disease.
AcknowledgmentsWe thank J.C. Ingram for technical support and Dr S. Ponnambalamfor provision of HUVECs.
Sources of FundingThis study was funded by the British Heart Foundation and Well-come Trust. C.E.C. is Canada Research Chair in Oncology; J.D.Y. isHeritage Scientist of the Alberta Heritage Foundation for MedicalResearch.
DisclosuresNone.
References1. Mubagwa K, Mullane K, Flameng W. Role of adenosine in the heart and
circulation. Cardiovasc Res. 1996;32:797–813.2. Shryock JC, Belardinelli L. Adenosine and adenosine receptors in the
cardiovascular system: biochemistry, physiology, and pharmacology.Am J Cardiol. 1997;79:2–10.
3. Deussen A. Metabolic flux rates of adenosine in the heart. NaunynSchmiedebergs Arch Pharmacol. 2000;362:351–363.
4. Koszalka P, Ozuyaman B, Huo Y, Zernecke A, Flogel U, Braun N,Buchheiser A, Decking UK, Smith ML, Sevigny J, Gear A, Weber AA,Molojavyi A, Ding Z, Weber C, Ley K, Zimmermann H, Godecke A,Schrader J. Targeted disruption of cd73/ecto-5�-nucleotidase alters throm-boregulation and augments vascular inflammatory response. Circ Res.2004;95:814–821.
5. Mubagwa K, Flameng W. Adenosine, adenosine receptors and myo-cardial protection: an updated overview. Cardiovasc Res. 2001;52:25–39.
6. Zaza A, Rocchetti M, DiFrancesco D. Modulation of the hyperpolariza-tion-activated current (If) by adenosine in rabbit sinoatrial myocytes.Circulation. 1996;94:734–741.
7. Riksen NP, Smits P, Rongen GA. Ischaemic preconditioning: frommolecular characterisation to clinical application–part I. Neth J Med.2004;62:353–363.
8. Deussen A, Stappert M, Schafer S, Kelm M. Quantification of extra-cellular and intracellular adenosine production: understanding the trans-membranous concentration gradient. Circulation. 1999;99:2041–2047.
9. Decking UK, Schlieper G, Kroll K, Schrader J. Hypoxia-induced inhi-bition of adenosine kinase potentiates cardiac adenosine release. Circ Res.1997;81:154–164.
10. Meester BJ, Shankley NP, Welsh NJ, Meijler FL, Black JW. Pharmaco-logical analysis of the activity of the adenosine uptake inhibitor, dipyri-damole, on the sinoatrial and atrioventricular nodes of the guinea-pig.Br J Pharmacol. 1998;124:729–741.
11. Baldwin SA, Beal PR, Yao SYM, King AE, Cass CE, Young JD. Theequilibrative nucleoside transporter family, SLC29. Pflugers Arch. 2004;447:735–743.
12. Gray JH, Owen RP, Giacomini KM. The concentrative nucleoside trans-porter family, SLC28. Pflugers Arch. 2004;447:728–734.
13. Podgorska M, Kocbuch K, Grden M, Szutowicz A, Pawelczyk T. Prev-alence of unidirectional Na�-dependent adenosine transport and alteredpotential for adenosine generation in diabetic cardiac myocytes. Basic ResCardiol. 2006;101:214–222.
14. Baldwin SA, Yao SYM, Hyde RJ, Ng AML, Foppolo S, Barnes K, RitzelMWL, Cass CE, Young JD. Functional characterization of novel humanand mouse equilibrative nucleoside transporters (hENT3 and mENT3)located in intracellular membranes. J Biol Chem.2005;280:15880–15887.
15. Pinto-Duarte A, Coelho JE, Cunha RA, Ribeiro JA, Sebastiao AM.Adenosine A2A receptors control the extracellular levels of adenosinethrough modulation of nucleoside transporters activity in the rat hip-pocampus. J Neurochem. 2005;93:595–604.
16. Dhalla AK, Dodam JR, Jones AW, Rubin LJ. Characterization of anNBTI-sensitive equilibrative nucleoside transporter in vascular smoothmuscle. J Mol Cell Cardiol. 2001;33:1143–1152.
17. Schwartz LM, Bukowski TR, Ploger JD, Bassingthwaighte JB. Endothe-lial adenosine transporter characterization in perfused guinea pig hearts.Am J Physiol. 2000;279:H1502–H1511.
18. Ford DA, Rovetto MJ. Rat cardiac myocyte adenosine transport andmetabolism. Am J Physiol. 1987;252:H54–H63.
19. Geisbuhler TP, Johnson DA, Rovetto MJ. Cardiac myocyte guanosinetransport and metabolism. Am J Physiol. 1987;253:C645–C651.
20. Sundaram M, Yao SYM, Ingram JC, Berry ZA, Abidi F, Cass CE,Baldwin SA, Young JD. Topology of a human equilibrative, nitroben-zylthioinosine (NBMPR)-sensitive nucleoside transporter (hENT1)
10 Circulation Research September 1, 2006
by on May 18, 2011 circres.ahajournals.orgDownloaded from

implicated in the cellular uptake of adenosine and anti-cancer drugs.J Biol Chem. 2001;276:45270–45275.
21. Musa H, Dobrzynski H, Berry Z, Abidi F, Cass CE, Young JD, BaldwinSA, Boyett MR. Immunocytochemical demonstration of the equilibrativenucleoside transporter rENT1 in rat sinoatrial node. J HistochemCytochem. 2002;50:305–309.
22. Acimovic Y, Coe IR. Molecular evolution of the equilibrative nucleosidetransporter family: identification of novel family members in prokaryotesand eukaryotes. Mol Biol Evol. 2002;19:2199–2210.
23. Engel K, Wang J. Interaction of organic cations with a newly identifiedplasma membrane monoamine transporter (PMAT). Mol Pharmacol.2005;68:1397–1407.
24. Engel K, Zhou M, Wang J. Identification and characterization of a novelmonoamine transporter in human brain. J Biol Chem. 2004;279:50042–50049.
25. Ikeda K, Tojo K, Otsubo C, Udagawa T, Kumazawa K, Ishikawa M,Tokudome G, Hosoya T, Tajima N, Claycomb WC, Nakao K, KawamuraM. 5-Hydroxytryptamine synthesis in HL-1 cells and neonatal rat cardio-cytes. Biochem Biophys Res Commun. 2005;328:522–525.
26. Nebigil CG, Maroteaux L. A novel role for serotonin in heart. TrendsCardiovasc Med. 2001;11:329–335.
27. Liman ER, Tytgat J, Hess P. Subunit stoichiometry of a mammalian K�
channel determined by construction of multimeric cDNAs. Neuron. 1992;9:861–871.
28. Claycomb WC, Lanson NA Jr, Stallworth BS, Egeland DB, Delcarpio JB,Bahinski A, Izzo NJ Jr. HL-1 cells: a cardiac muscle cell line thatcontracts and retains phenotypic characteristics of the adult cardiomyo-cyte. Proc Natl Acad Sci U S A. 1998;95:2979–2984.
29. Herbert SP, Ponnambalam S, Walker JH. Cytosolic phospholipaseA2-alpha mediates endothelial cell proliferation and is inactivated byassociation with the Golgi apparatus. Mol Biol Cell. 2005;16:3800–3809.
30. Davies A, Meeran K, Cairns MT, Baldwin SA. Peptide-specific anti-bodies as probes of the orientation of the glucose transporter in the humanerythrocyte membrane. J Biol Chem. 1987;262:9347–9352.
31. Lai Y, Tse CM, Unadkat JD. Mitochondrial expression of the humanequilibrative nucleoside transporter 1 (hENT1) results in enhanced mito-chondrial toxicity of antiviral drugs. J Biol Chem. 2004;279:4490–4497.
32. Mani RS, Hammond JR, Marjan JJ, Graham KA, Young JD, Baldwin SA,Cass CE. Demonstration of equilibrative nucleoside transporters (hENT1and hENT2) in nuclear envelopes of cultured human choriocarcinoma(BeWo) cells by functional reconstitution in proteoliposomes. J BiolChem. 1998;273:30818–30825.
33. Chaudary N, Shuralyova I, Liron T, Sweeney G, Coe IR. Transportcharacteristics of HL-1 cells: a new model for the study of adenosinephysiology in cardiomyocytes. Biochem Cell Biol. 2002;80:655–665.
34. Dobrzynski H, Rothery SM, Marples DD, Coppen SR, Takagishi Y,Honjo H, Tamkun MM, Henderson Z, Kodama I, Severs NJ, Boyett MR.Presence of the Kv1.5 K� channel in the sinoatrial node. J HistochemCytochem. 2000;48:769–780.
35. Stieber J, Herrmann S, Feil S, Loster J, Feil R, Biel M, Hofmann F,Ludwig A. The hyperpolarization-activated channel HCN4 is required forthe generation of pacemaker action potentials in the embryonic heart.Proc Natl Acad Sci U S A. 2003;100:15235–15240.
36. Dobrzynski H, Marples DD, Musa H, Yamanushi TT, Henderson Z,Takagishi Y, Honjo H, Kodama I, Boyett MR. Distribution of the mus-carinic K� channel proteins Kir3.1 and Kir3.4 in the ventricle, atrium, andsinoatrial node of heart. J Histochem Cytochem. 2001;49:1221–1234.
37. Carrier M, Trudelle S, Thai P, Pelletier L. Ischemic threshold during coldblood cardioplegic arrest: monitoring with tissue pH and pO2. J Car-diovasc Surg (Torino). 1998;39:593–597.
38. Riksen NP, Oyen WJ, Ramakers BP, Van den Broek PH, Engbersen R,Boerman OC, Smits P, Rongen GA. Oral therapy with dipyridamolelimits ischemia-reperfusion injury in humans. Clin Pharmacol Ther.2005;78:52–59.
Barnes et al ENT4, a Novel Cardiac Adenosine Transporter 11
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 1
Online Data Supplement Barnes et al.
Materials and Methods
Construction of ENT4 expression vectors
cDNAs encoding mouse (m) and human (h) ENT4 were obtained from the IMAGE
consortium as cloneID 5250967 (mouse mammary tumour) and cloneID 4816109 (human
hippocampus) respectively. DNA sequencing showed that the amino acid sequence encoded
by the mouse cDNA was identical to that reported in GenBank accession NP_666369. In
contrast, the nucleotide sequence of the human cDNA (GenBank accession BC047592)
differed from that reported for hENT4/PMAT (GenBank accession AY485959) and predicted
from the human genome sequence (GenBank accession BK000627) at the codons for residues
E79, K124 and T429. Since the differences rendered the protein inactive in transport (data
not shown), they were mutated to those reported for hENT4/PMAT (V, N and P respectively)
using the QuickChangeR Multi Site-Directed Mutagenesis method (Stratagene) and the
following primers:
E79V 5′ P-GATGCTGCTGGCTGGCGTGGGCTTCCTGCTGCC 3′
K124N 5′ P-CTGTCCTCCTGAACAACGTCCTGGTGGAGAGAC 3′
T429P 5′ P-GCGTGTGGTCTTCATCCCCCTCTTCATCCTGTG 3′
For efficient expression in Xenopus oocytes the following primers were used to introduce
XbaI sites (underlined) before the initiation codon (italicised) and after the stop codon (bold)
of the mENT4 and mutated hENT4 coding regions by PCR amplification:
mENT4 forward 5′ CGAATTCTCTAGAATGGGCTCTATCGGAAGC 3′
mENT4 reverse 5′ GCTCTAGACTCAGGGACCGACAGGGATGG 3′
hENT4 forward 5′ GAGTCTAGAATGGGCTCCGTGGG 3′
hENT4 reverse 5′ GCTTGCTCTAGACTCAGAGGCCTGCG 3′
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 2
Following insertion of the amplification products into the XbaI site of the Xenopus expression
vector pGEM-HE,1 constructs harbouring inserts with the desired orientation were identified
by restriction digestion and designated pGmENT4 and pGhENT4, respectively.
Mutants E206Q and E375Q of hENT4 were made in the pGhENT4 template using the
QuickChangeR Site-Directed Mutagenesis method (Stratagene) and the following primers:
E206Q forward 5′ GTGATGACCGGGCAGAGCACGGCGGGC 3′
E206Q reverse 5′ GCCCGCCGTGCTCTGCCCGGTCATCAC 3′
E375Q forward 5′ CCCCGGCCTCCAGTCTGAGATCCGC 3′
E375Q reverse 5′ GCGGATCTCAGACTGGAGGCCGGGG 3′
For quantification of ENT4, a glutathione S-transferase (GST) fusion protein encoding the
central cytoplasmic loop of mENT4 (residues 252 to 347) was produced by PCR
amplification of the cognate region of pGmENT4 using the following primers:
5′ CGAGGATCCGTGCGGCGCAGCCGTTTTGTG 3′
5′ CGAGAATTCTCAGACCCGCGCCACGACGTATC 3′
The primer-encoded BamHI and EcoRI restriction sites (underlined) were then used to insert
the amplification product into a version of the Escherichia coli expression vector pGEX-KT
modified to include a tobacco etch virus protease cleavage site.2 The coding regions of all
constructs were sequenced to ensure their fidelity.
Production and characterisation of hENT4 and mENT4 in Xenopus oocytes
For production of mENT4 and wild-type or mutant hENT4 in Xenopus oocytes, pGmENT4,
pGhENT4 or mutant derivatives were linearised with NheI and transcribed with T7
polymerase in the presence of m7GpppG cap using the mMESSAGE mMACHINE™
(Ambion) transcription system. Preparation of oocytes, injection with RNA transcripts (20 ng
per oocyte) and assays of the uptake of radiolabelled nucleosides and nucleobases (GE
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 3
Healthcare UK Limited, Chalfont St.Giles, Bucks, UK) were performed essentially as
previously described.3 Transport measurements were made in a solution containing 100
mmol/L NaCl (or 100 mmol/L choline chloride), 2 mmol/L KCl, 1 mmol/L CaCl2 and 1
mmol/L MgCl2, buffered with either 10 mmol/L 2-morpholinoethanesulfonic acid (MES; pH
5.0-6.0), 10 mmol/L N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid (HEPES; pH 6.0-
7.5) or 10 mmol/L Tris-HCl (pH 8.0-8.5). Unless otherwise indicated, the incubation time
was 30 min and the permeant concentration was 100 μmol/L. Uptake of serotonin (5-
hydroxy[3H]tryptamine, GE Healthcare UK Limited) was measured in the presence of
ascorbic acid (100 µmol/L) and paragyline (10 µmol/L) to prevent oxidation. Each
experiment was performed at least twice on different batches of oocytes. For estimates of Km
and Vmax values the data, corrected for non-mediated uptake seen in oocytes injected with
water rather than with RNA transcripts, were fitted to the Michaelis-Menten equation using
non-linear regression.
Measurement of transport activity in HL-1 cells
HL-1 cardiomyocytes were cultured in 6-well plates as previously described.4 Uptake of
serotonin (5-hydroxy[3H]tryptamine, 50 µmol/L, 10 min) was measured in triplicate at 37ºC
in the same transport buffer as used for oocytes, on cells at 80% confluence. Uptake was
terminated by washing cells three times with ice-cold phosphate-buffered saline (8 mmol/L
Na2HPO4, 1.6 mmol/L KH2PO4, 2.4 mmol/L KCl, 140 mmol/L NaCl, pH 7.4). The cells were
then solubilised in 1% (v/v) Triton X-100 and samples taken for quantification of
radioactivity by scintillation counting and protein content using the bicinchoninic acid assay
(Pierce). Results from representative experiments, repeated at least three times, are expressed
as means ± S.D. Statistical analysis was performed using the two-tailed Student’s t test.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 4
Measurement of transport activity in rat ventricular myocytes
Uptake of [14C]-labelled adenosine (100 µmol/L at 37ºC) into freshly isolated
cardiomyocytes (100 µl; 105 cells) in 1.5 ml tubes was measured at pH 5.5 for 30 sec (during
which period the rate of permeant uptake was linear) in the same choline chloride transport
buffer as used for oocytes in the absence or presence of either 10 µmol/L NBMPR or 10
mmol/L uridine. Prior to addition of the assay buffer, cells were incubated for 30 min at room
temperature in the presence of the inhibitors to allow binding to the transporter to reach
equilibrium. After stopping the assay by adding 200 µl of adenosine (10 mmol/L) in ice-cold
assay buffer, 400 µl cells were added to a 200 µl layer of oil (a 1:1 ratio of dinonyl phthalate,
Fisher Scientific, UK Limited, Bishop Meadow Road, Loughborough, Leicestershire LE11
5RG, UK and Dow-Corning 550 silicone fluid, BDH Laboratory Supplies, Poole, BH15 1TD,
UK) in a 1.5 ml tube and separated from the assay medium by centrifugation at 13,000 rpm in
a microcentrifuge. The assay medium was aspirated and the tube washed 5X with water prior
to aspiration of the oil and water. The cell pellet was dissolved in 25 µl 1 mol/L NaOH at
37ºC for 1 h for quantification of radioactivity by scintillation counting. Results from
representative experiments, repeated at least three times, are expressed as means ± S.D.
Statistical analysis was performed using the two-tailed Student’s t test.
Tissue distribution of hENT4 mRNA
A human multiple tissue expression (MTE™) mRNA array (Clontech) was incubated with an
[α-32P]dATP-labelled antisense DNA probe corresponding to residues 79-141 of hENT4,
produced using a Strip-EZ™ PCR kit (Ambion) and the following primers:
hENT4-F 5' GACAGTCACCAGCTGGAGGAGG 3'
hENT4-R 5' GAGGTAGCCTGCGGTGATCCTG 3'
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 5
Hybridization, washing and quantification were performed using the methods described in
detail previously.3 The signals for each tissue sample were normalized to that for human
DNA (500 ng), which was given a value of 1. Possible cross-hybridization between the
hENT4 probe and other ENT family members was tested on dot blots of dilutions (100 ng-
0.05 ng RNA) of hENT1, hENT2, hENT3 or hENT4 in vitro transcripts, using the same
procedures for hybridization and washing. The results showed that the probe was specific for
the hENT4 transcript under the highly stringent hybridization and washing conditions used in
the analysis (data not shown).
Detection and quantification of ENT4 by western blotting
Western blotting of samples from mouse, human and rat tissues and cultured cells was
employed to investigate the distribution of the corresponding ENT4 proteins. Permission to
use human tissue was granted by the Leeds (West) Research Ethics Committee. Human
umbilical vein endothelial cells (HUVECs) were cultured as described by Herbert et al..5
Lysates of isolated rat ventricular myocytes, HL-1 cells, HUVECS and rat heart tissue
samples were prepared in the presence of protease inhibitors (Sigma-Aldrich protease
inhibitor cocktail P8340 containing 4-(2-aminoethyl)benzenesulfonyl fluoride, pepstatinA, E-
64, bestatin, leupeptin, and aprotinin) as described previously.6 For preparation of
membranes, samples (1 g) of mouse, human and rat tissues were homogenised on ice using
an Ultra-Turrax® homogeniser (3 x 5 sec bursts) in 5 ml 0.25 mol/L sucrose, 1 mmol/L
ethylenediaminetetraacetic acid (EDTA), 10 mmol/L HEPES, pH 7.4 containing the same
cocktail of mammalian protease inhibitors. The homogenates were depleted of nuclei and
mitochondria by centrifugation at 3,000 x g for 10 min at 4°C. Membranes were then
prepared by centrifuging the supernatants at 100,000 x g for 45 min at 4°C. Xenopus oocyte
membranes were prepared as previously described.7 Where indicated in the text, samples of
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 6
oocyte, mouse or human membranes (10 µg) were deglycosylated by treatment with N-
Glysosidase F according to the manufacturer’s instructions (Roche Applied Sciences).
Antibodies against a synthetic peptide corresponding to residues 301-319 of hENT4 were
raised in rabbits by Pepceuticals Ltd. (Leicester, UK) and affinity-purified by
chromatography on a column of immobilised peptide as previously described.8 The resultant
antibodies, designated anti-hENT4301-319, were used at a concentration of 2 µg/ml for western
blotting. Blots were also stained with mouse monoclonal antibodies against a synthetic
peptide corresponding to residues 252-270 of rat cardiac connexin 43 (Cx43; Chemicon
International, Harrow, UK, clone 4E6.2, used at a dilution of 1:1000) and against α-tubulin
(Sigma-Aldrich, clone DM1A, used at a dilution of 1:2000). For western blotting, protein
samples were resolved by SDS/10% (w/v) polyacrylamide gel electrophoresis, electroblotted
onto nitrocellulose membranes, blocked by incubation with 5% (w/v) skimmed milk powder
and then incubated with primary antibodies at 4ºC overnight. Following incubation with
horseradish peroxidase conjugates of goat anti-rabbit or anti-mouse IgG as appropriate
(1/50,000; Jackson ImmunoResearch Laboratories Inc.), antigens were visualised using
Supersignal West Pico Chemiluminescent Substrate (Pierce). Where applicable blots were
stripped of antibodies (Restore™ Western Blot Stripping Buffer, Pierce) and reprobed for
relevant proteins. Staining intensities were quantified using a Bio-Rad Fluor-S gel
documentation system and Multi-analyst software. For quantitative estimation of rENT4
levels in different regions of the heart, known amounts of a purified GST fusion protein
bearing the central cytoplasmic loop of mENT4 (residues 252 to 347) were included in the
gels used for blotting. The amounts of heart proteins used were chosen such that their staining
intensities fell within the region where a linear relationship existed between signal intensity
and the amount of fusion protein standard present.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 7
Immunocytochemistry
Freshly isolated rat atrial and ventricular cardiomyocytes adhering to coverslips, and
HUVECs and HL-1 cells cultured on coverslips, were fixed for 20 min in 4% formaldehyde
in phosphate-buffered saline (PBS; 0.01 M phosphate buffer, 0.0027 M potassium chloride
and 0.137 M sodium chloride, pH 7.4, Sigma-Aldrich), washed in PBS (three times 10 min),
blocked in 10% normal goat serum (NGS; GE Healthcare UK Limited) for 1 h at room
temperature and incubated overnight at 4°C with 20 μg/mL anti-hENT4301-319 in PBS
containing 1% NGS and 0.1% saponin. To confirm the specificity of the hENT4 staining
parallel samples were stained with anti-hENT4301-319 that had been pre-incubated with a 2-
fold excess by weight of synthetic peptide. After subsequent washing in PBS, cells were
incubated in a goat anti-rabbit IgG fluorescein isothiocyanate (FITC) conjugate (1/50)
(Sigma-Aldrich), washed again in PBS and mounted in Vectashield® medium (Vector
Laboratories, Ltd., Peterborough, UK). For imaging, twenty optical sections were captured at
0.2 µm intervals with a Delta Vision® system (Applied Precision, Issaquah WA) comprising
an Olympus IX70 microscope linked to a CCD camera, and processed using the
manufacturer’s deconvolution software.
To prepare heart tissue sections, rats of either sex weighing 0.2 to 0.3 kg were sacrificed
and the tissue dissected, cryosectioned and mounted on poly-L-lysine-coated slides (BDH;
Poole, Dorset) as described previously.6 Sections (20 µm) through the rat sinoatrial (SA)
node and surrounding atrial tissue, and from the atrioventricular (AV) node and ventricular
tissue were prepared from at least four rats. Human heart sections (samples of right atrial
appendage from at least three human hearts) were fixed, stained and mounted as described
above, except that sections were treated with 0.1% Triton X-100 prior to the blocking step to
enhance permeabilization of plasma membranes. For double-labelling experiments, sections
were incubated simultaneously with either rabbit anti-hENT4301-319 or rabbit anti-
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 8
Hyperpolarization-Activated Cyclic Nucleotide-Gated Potassium Channel 4 (HCN4;
Chemicon International, used at a dilution of 1:20) plus mouse monoclonal antibodies against
Cx43 (Chemicon International, used at a dilution of 1:50) or human von Willebrand factor
(vW; Chemicon International, used at a dilution of 1:50). Following incubation and washing
as described above, sections were stained with a mixture of goat anti-rabbit IgG FITC
conjugate (1:50) and goat anti-mouse IgG tetramethylrhodamine isothiocyanate (TRITC)
conjugate (1:100) and then washed and mounted as described above. Images were captured
on a LSM510 META laser scanning confocal microscope (Carl Zeiss, Herts, UK). Optical
sections (1 µm) were collected using the appropriate pinhole setting.
Figure Legends
Supplementary Figure 1. Time course of ENT4-mediated adenosine transport. Uptake of
14C-labelled adenosine (100 µmol/L) was measured in oocytes injected with mENT4 RNA
transcripts or water alone for the indicated periods. Mediated uptake values were obtained by
correction for uptake by oocytes injected with water alone, which typically represented ≤4%
that of oocytes expressing mENT4, and are shown are means ± SEM (n = 12).
Supplementary Figure 2. Tissue distribution of hENT4. (A) and (B), a commercial human
multiple tissue expression RNA array probed with a 32P-labelled antisense probe
corresponding to hENT4 amino acid residues 79-141. The numbered samples are: 1,
leukemia (undifferentiated HL-60); 2, HeLa S3; 3, leukemia (K-562); 4, leukemia (MOLT-
4); 5, Burkitt’s lymphoma (Raji); 6, Burkitt’s lymphoma (Daudi); 7, colorectal
adenocarcinoma (SW480); 8, lung carcinoma (A549); 9, whole brain ; 10, cerebral cortex;
11, frontal lobe; 12, parietal lobe; 13, occipital lobe; 14 temporal lobe; 15, paracentral gyrus
of cerebral cortex; 16, pons; 17, cerebellum (left); 18, cerebellum (right); 19, corpus
callosum; 20, amygdala; 21, caudate nucleus; 22, hippocampus; 23, medulla oblongata; 24,
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 9
putamen; 25, substantia nigra; 26, accumbens nucleus; 27, thalamus; 28, pituitary gland; 29,
spinal cord; 30, esophagus; 31 stomach; 32, duodenum; 33, jejunum; 34, ileum; 35, ilocecum;
36, appendix; 37, colon (ascending); 38, colon (transverse); 39, colon (descending); 40,
rectum; 41, heart; 42, aorta; 43, atrium (left); 44, atrium (right); 45, ventricle (left); 46,
ventricle (right); 47, interventricular septum; 48, apex of the heart; 49, kidney; 50, skeletal
muscle; 51, spleen; 52, thymus; 53, peripheral blood leukocyte; 54, lymph node; 55, bone
marrow; 56, trachea; 57, lung; 58, placenta; 59, bladder; 60, uterus; 61, prostate; 62, testis;
63, ovary; 64, liver; 65, pancreas; 66, adrenal gland; 67, thyroid gland; 68, salivary gland; 69,
mammary gland; 70, fetal brain; 71 fetal heart; 72, fetal kidney; 73, fetal liver; 74, fetal
spleen; 75, fetal thymus; 76, fetal lung; 77, human DNA (100 ng); 78 human DNA (500 ng).
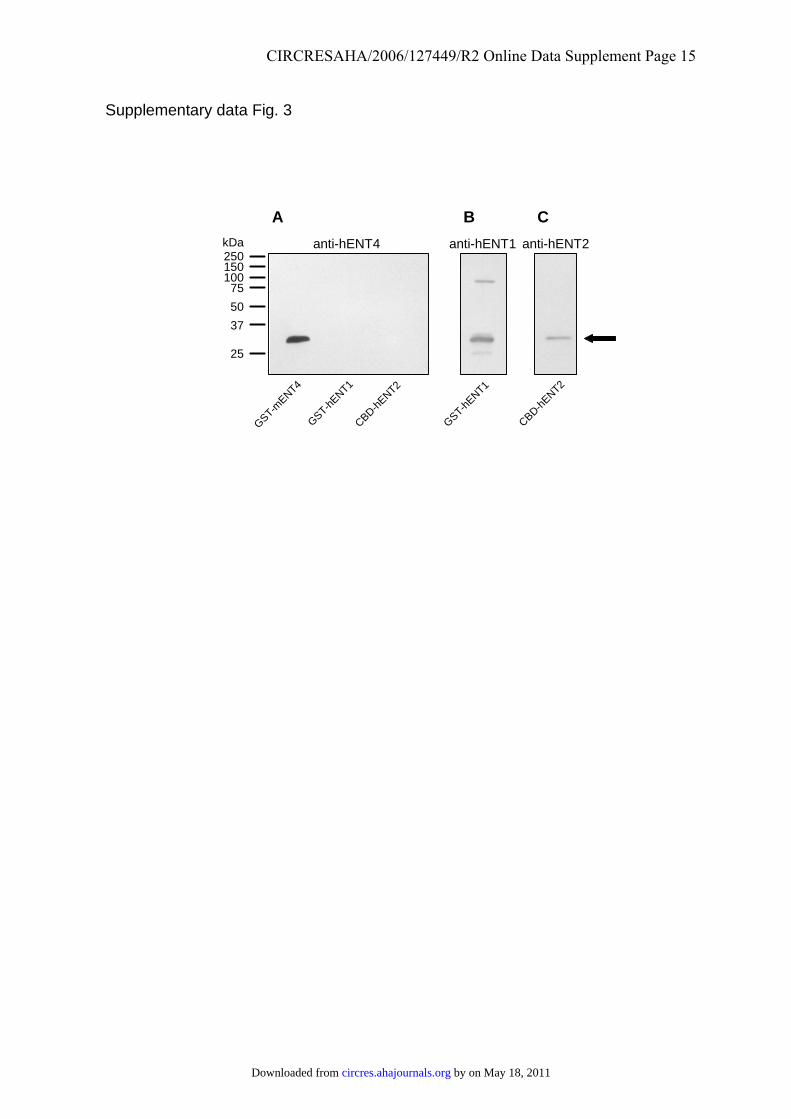
Supplementary Figure 3. Specificity of anti-hENT4 antibodies. (A) Samples (1 µg) of
glutathione S-transferase (GST) fusion proteins bearing the central cytoplasmic loops of
mENT4 (residues 252-350) or of hENT1 (residues 226-291) and of a cellulose binding
domain (CBD) fusion protein bearing the central cytoplasmic loop of hENT2 (residues 214-
292) were separated on a SDS/10% (w/v) polyacrylamide gel. After transfer to nitrocellulose,
the blot was probed with antibodies to hENT4301-319. For (B) and (C) the antibodies were
stripped from blot (A) and the portions containing GST-hENT1 and CBD-hENT2 were re-
probed with antibodies to hENT1 (B)6 and hENT2 (C)9, respectively, as indicated. The
apparent size of the bands stained by the antibodies, ~ 30 kDa, was consistent with the
predicted molecular mass of the fusion proteins (arrow). The mobilities of proteins of known
molecular mass are shown on the left.
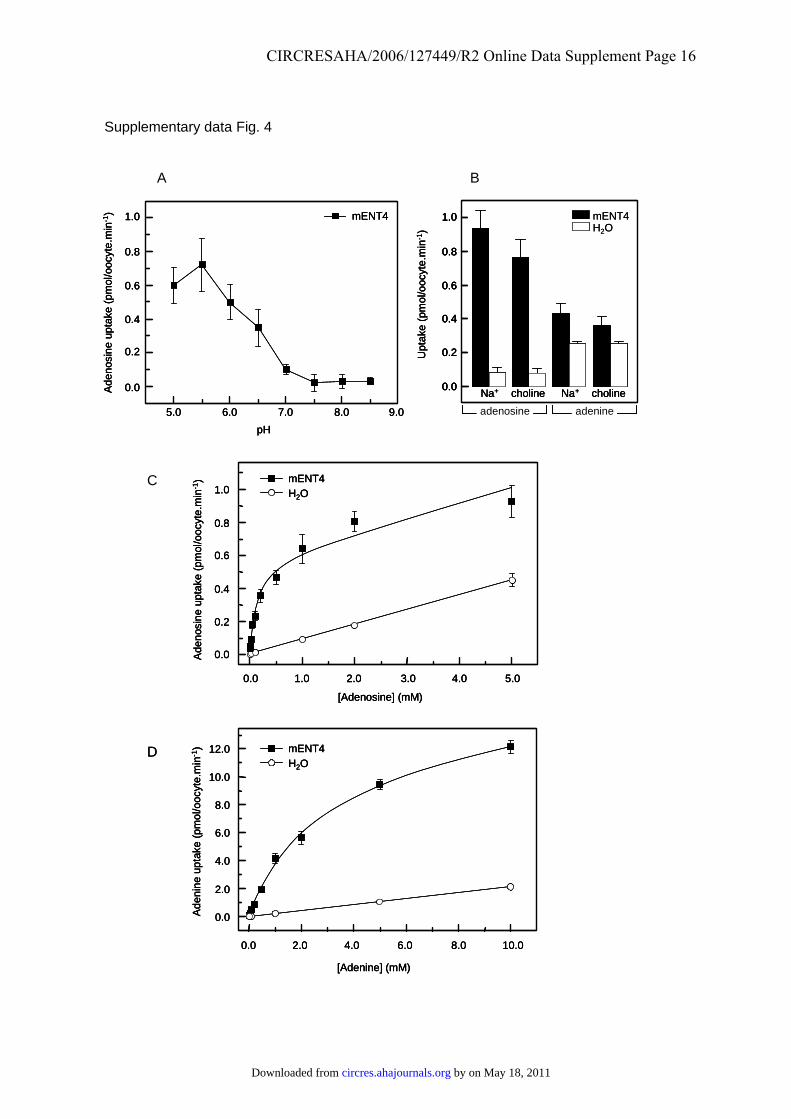
Supplementary Figure 4. Cation- and concentration-dependence of mENT4-mediated
transport. Uptake of 14C-labelled adenosine (A, B, C) and adenine (B, D) was measured at a
permeant concentration of 100 µmol/L (A, B) or at the indicated concentrations (C, D) at
20ºC for 30 min in oocytes injected with mENT4 RNA transcripts (solid symbols and bars)
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 10
or water alone (open symbols and bars). Data shown in (A) were corrected for endogenous
transport activity by subtraction of uptake measured in oocytes injected with water only.
Results shown are means ± SEM (n = 12). Uptake was measured in transport medium
containing 100 mM sodium chloride, except for the indicated samples in Panel (B) where
sodium chloride was replaced by 100 mM choline chloride to generate sodium-free medium,
and buffered at the indicated pH values (A) or at pH 5.5 (B, C, D). (A) pH-dependence of
mENT4-mediated adenosine uptake; (B) sodium-dependence of mENT4-mediated adenosine
uptake; (C) concentration-dependence of mENT4-mediated adenosine uptake; and (D)
concentration-dependence of mENT4-mediated adenine uptake.
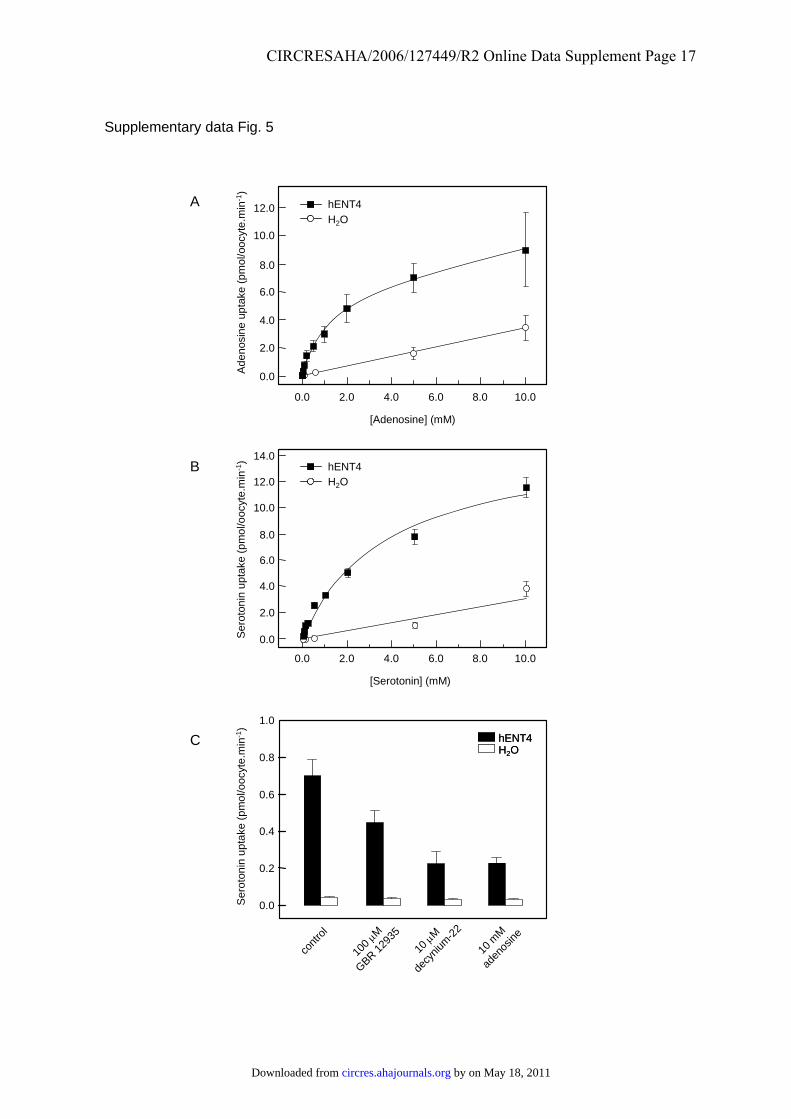
Supplementary Figure 5. Concentration-dependence and inhibition of hENT4-mediated
transport. Uptake of 14C-labelled adenosine (A) or 3H-labelled serotonin (B, C) was measured
at the indicated permeant concentrations (A, B) or at 100 μmol/L (C) at 20ºC for 30 min in
oocytes injected with hENT4 RNA transcripts (solid symbols and bars) or water alone (open
symbols and bars). Uptake was measured in transport medium containing 100 mM sodium
chloride and buffered at pH 5.5. (A) and (B), concentration-dependence of the uptake of
adenosine and serotonin, respectively. Apparent Km and Vmax values, determined by non-
linear regression analysis of permeant influx in RNA-injected oocytes minus that measured in
water-injected oocytes, are given in the Results section. (C) inhibition of serotonin uptake:
oocytes were pre-incubated in transport buffer in the presence or absence (control) of the
inhibitors indicated for 1 h prior to addition of the permeant.
Supplementary Figure 6. Location of nodal tissue in the heart. Cryosections (20 µm) of the
SA node (A) and AV node (B) regions of the rat heart were cut and stained with Masson’s
trichrome as previously described.10 The SA node is marked by a red line in (A) and the AV
node by a dotted red line in (B).
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 11
Supplementary Figure 7. Effect of pH on adenosine uptake mediated by hENT4 and its
mutants produced in oocytes. Uptake of 14C-labelled adenosine (100 μmol/L, 20°C, 30 min)
in oocytes injected with the indicated mRNA transcripts was measured at pH 6.00 (solid bars)
or pH 8.00 (open bars) in transport medium containing 100 mmol/L sodium chloride. Data
were corrected for endogenous transport by subtraction of uptake measured in oocytes
injected with water only. Results shown are means ± SEM (n = 12).
References
1. Liman ER, Tytgat J, Hess P. Subunit stoichiometry of a mammalian K+ channel
determined by construction of multimeric cDNAs. Neuron. 1992;9:861-871.
2. Hakes DJ, Dixon JE. New vectors for high level expression of recombinant proteins in
bacteria. Anal Biochem. 1992;202:293-298.
3. Baldwin SA, Yao SYM, Hyde RJ, Ng AML, Foppolo S, Barnes K, Ritzel MWL, Cass
CE, Young JD. Functional characterization of novel human and mouse equilibrative
nucleoside transporters (hENT3 and mENT3) located in intracellular membranes. J Biol
Chem. 2005;280:15880-15887.
4. Claycomb WC, Lanson NA, Jr., Stallworth BS, Egeland DB, Delcarpio JB, Bahinski A,
Izzo NJ, Jr. HL-1 cells: a cardiac muscle cell line that contracts and retains phenotypic
characteristics of the adult cardiomyocyte. Proc Natl Acad Sci USA. 1998;95:2979-
2984.
5. Herbert SP, Ponnambalam S, Walker JH. Cytosolic phospholipase A2-alpha mediates
endothelial cell proliferation and is inactivated by association with the Golgi apparatus.
Mol Biol Cell. 2005;16:3800-3809.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 12
6. Musa H, Dobrzynski H, Berry Z, Abidi F, Cass CE, Young JD, Baldwin SA, Boyett
MR. Immunocytochemical demonstration of the equilibrative nucleoside transporter
rENT1 in rat sinoatrial node. J Histochem Cytochem. 2002;50:305-309.
7. Sundaram M, Yao SYM, Ingram JC, Berry ZA, Abidi F, Cass CE, Baldwin SA, Young
JD. Topology of a human equilibrative, nitrobenzylthioinosine (NBMPR)-sensitive
nucleoside transporter (hENT1) implicated in the cellular uptake of adenosine and anti-
cancer drugs. J Biol Chem. 2001;276:45270-45275.
8. Davies A, Meeran K, Cairns MT, Baldwin SA. Peptide-specific antibodies as probes of
the orientation of the glucose transporter in the human erythrocyte membrane. J Biol
Chem. 1987;262:9347-9352.
9. Redzic ZB, Biringer J, Barnes K, Baldwin SA, Al Sarraf H, Nicola PA, Young JD, Cass
CE, Barrand MA, Hladky SB. Polarized distribution of nucleoside transporters in rat
brain endothelial and choroid plexus epithelial cells. J Neurochem. 2005;94:1420-1426.
10. Dobrzynski H, Nikolski VP, Sambelashvili AT, Greener ID, Yamamoto M, Boyett MR,
Efimov IR. Site of origin and molecular substrate of atrioventricular junctional rhythm
in the rabbit heart. Circ Res. 2003;93:1102-1110.
by on May 18, 2011 circres.ahajournals.orgDownloaded from
CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 13
0 20 40 60 80 100 120
0.0
1.0
2.0
3.0
4.0
Time (min)
Aden
osin
e up
take
(pm
ol/o
ocyt
e.m
in-1
)
Supplementary data Fig. 1
0 20 40 60 80 100 120
0.0
1.0
2.0
3.0
4.0
Time (min)
Aden
osin
e up
take
(pm
ol/o
ocyt
e.m
in-1
)
0 20 40 60 80 100 120
0.0
1.0
2.0
3.0
4.0
Time (min)
Aden
osin
e up
take
(pm
ol/o
ocyt
e.m
in-1
)
Supplementary data Fig. 1
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 14
A
RN
A d
ot b
lot s
igna
l int
ensi
ty(a
rbitr
ary
units
)
0
10
20
30
40
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 392 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
cell lines brain intestine
RN
A d
ot b
lot s
igna
l int
ensi
ty(a
rbitr
ary
units
)
10
20
30
40
041 43 45 47 49 51 53 55 57 59 61 63 65 67 69 71 73 75 77
42 44 46 48 50 52 54 56 58 60 62 64 66 68 70 72 74 76 78
B
heart other tissues fetal tissues DNA
Supplementary data Fig. 2
A
0
10
20
30
40
sity
ten
gnal
inR
NA
dot
blo
t si
(arb
itrar
y un
its)
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 392 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
cell lines brain intestine
RN
A d
ot b
lot s
igna
l int
ensi
ty(a
rbitr
ary
units
)
10
20
30
40
041 43 45 47 49 51 53 55 57 59 61 63 65 67 69 71 73 75 77
42 44 46 48 50 52 54 56 58 60 62 64 66 68 70 72 74 76 78
B
heart other tissues fetal tissues DNA
Supplementary data Fig. 2
by on May 18, 2011 circres.ahajournals.orgDownloaded from
CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 15
GST-mENT4
GST-hENT1
CBD-hENT2
GST-hENT1
CBD-hENT2
anti-hENT4 anti-hENT1 anti-hENT2
25
375075
100150250kDa
A B C
Supplementary data Fig. 3
by on May 18, 2011 circres.ahajournals.orgDownloaded from
CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 16
Supplementary data Fig. 4
[Adenosine] (mM)
0.0 1.0 2.0 3.0 4.0 5.0
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
0.0
1.0
0.2
0.4
0.6
0.8
mENT4H2O
[Adenosine] (mM)
0.0 1.0 2.0 3.0 4.0 5.0
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
0.0
1.0
0.2
0.4
0.6
0.8
[Adenosine] (mM)
0.0 1.0 2.0 3.0 4.0 5.0
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
0.0
1.0
0.2
0.4
0.6
0.8
mENT4H2OmENT4H2O
C
A
0.0 2.0 4.0 6.0 8.0 10.0
[Adenine] (mM)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
Ade
nine
upt
ake
(pm
ol/o
ocyt
e.m
in-1
) mENT4H2O
D
0.0 2.0 4.0 6.0 8.0 10.0
[Adenine] (mM)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
Ade
nine
upt
ake
(pm
ol/o
ocyt
e.m
in-1
)
0.0 2.0 4.0 6.0 8.0 10.0
[Adenine] (mM)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
Ade
nine
upt
ake
(pm
ol/o
ocyt
e.m
in-1
) mENT4H2OmENT4H2O
D
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
pH5.0 6.0 7.0 8.0 9.0
0.0
0.2
0.4
0.6
0.8
1.0 mENT4
Upt
ake
(pm
ol/o
ocyt
e.m
in-1
)
0.0
0.2
0.4
0.6
1.0
0.8
Na+ choline Na+ cholineadenosine adenine
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
pH5.0 6.0 7.0 8.0 9.0
0.0
0.2
0.4
0.6
0.8
1.0 mENT4
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
pH5.0 6.0 7.0 8.0 9.0
0.0
0.2
0.4
0.6
0.8
1.0 mENT4
Upt
ake
(pm
ol/o
ocyt
e.m
in-1
)
0.0
0.2
0.4
0.6
1.0
0.8
Na+ choline Na+ cholineadenosine adenine
Upt
ake
(pm
ol/o
ocyt
e.m
in-1
)
0.0
0.2
0.4
0.6
1.0
0.8
0.0
0.2
0.4
0.6
1.0
0.8
Na+ choline Na+ cholineadenosine adenine
B
mENT4H2OmENT4H2O
by on May 18, 2011 circres.ahajournals.orgDownloaded from
CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 17
A
B
C
hENT4H2O
0.0
2.0
4.0
6.0
8.0
10.0
12.0
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
0.0 2.0 4.0 6.0 8.0 10.0
[Adenosine] (mM)
Sero
toni
n up
take
(pm
ol/o
ocyt
e.m
in-1
)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
0.0 2.0 4.0 6.0 8.0 10.0
[Serotonin] (mM)
hENT4H2O
0.0
0.2
0.4
0.6
1.0
0.8
Ser
oton
in u
ptak
e (p
mol
/ooc
yte.
min
-1)
hENT4H2OhENT4H2O
contr
ol
100 μ
M
GBR 1293
5
10 m
M
aden
osine
10 μM
decy
nium-22
Supplementary data Fig. 5
by on May 18, 2011 circres.ahajournals.orgDownloaded from

CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 18
Supplementary data Fig. 6
by on May 18, 2011 circres.ahajournals.orgDownloaded from
CIRCRESAHA/2006/127449/R2 Online Data Supplement Page 19
Ade
nosi
ne u
ptak
e (p
mol
/ooc
yte.
min
-1)
WT E375Q E260Q
0.00
0.10
0.20pH 6.0pH 8.0
Supplementary data Fig. 7
by on May 18, 2011 circres.ahajournals.orgDownloaded from




















