
The small heat shock proteins αB-crystallin and Hsp27 suppress SOD1 aggregation in vitro
Upload
independentCategory
view
3download
0
Distinct heat-shock element arrangements that mediate the heatshock, but not the late-embryogenesis induction of smallheat-shock proteins, correlate with promoter activationin root-knot nematode feeding cells
Marta Barcala Æ Alejandra Garcıa Æ Pilar Cubas Æ Concepcion Almoguera ÆJuan Jordano Æ Carmen Fenoll Æ Carolina Escobar
Received: 19 July 2007 / Accepted: 2 November 2007 / Published online: 28 November 2007
� Springer Science+Business Media B.V. 2007
Abstract Genes coding small heat-shock proteins
(sHSPs) show distinct behaviours with respect to environ-
mental and developmental signals. Their transcriptional
regulation depends on particular combinations of heat
stress cis-elements (heat-shock elements; HSEs) but many
aspects regarding their regulation remain unclear. Cyst and
root-knot nematodes induce, in the roots of infected plants,
the differentiation of special feeding cells with high met-
abolic activity (syncytia and giant cells, respectively), a
process accompanied by extensive gene expression chan-
ges. The Hahsp17.7G4 (G4) promoter was active in giant
cells and its HSE arrangements were crucial for this acti-
vation. In the present work, we provide further basis to
associate giant cell expression with the heat-shock
response of this gene class, by analysing additional pro-
moters. The Hahsp17.6G1 (G1) promoter, not induced by
heat shock, was silent in giant cells, while Hahsp18.6G2
(G2), which responds to heat shock, was specifically
induced in giant cells. In addition, a mutated Hahsp17.7G4
promoter version (G4MutP) with a strong heat-shock
induction was also induced in giant cells. The responses of
the different promoters correlated with distinct HSE con-
figurations, which might have implications on differential
trans-activation. Furthermore, the shortest giant cell and
heat-shock-inducible sHSP promoter version analysed in
tobacco (-83pb Hahsp17.7G4) fully maintained its
expression profile in Arabidopsis. Cyst nematodes did not
induce the Hahsp17.7G4 promoter, revealing additional
specificity in the nematode response. These findings, toge-
ther with the fact that the class I sHSP products of
endogenous genes accumulated specifically in tobacco giant
cells, support the idea that these nematode-induced giant
cells represent a transcriptional state very similar to that
produced by heat shock regarding this class of genes. The
high metabolic rate of giant cells may result in unfolded
proteins requiring class I sHSPs as chaperones, which might,
somehow, mimic heat-shock and/or other stress responses.
Keywords Heat-shock cis-element � Heat-shock factor �Late embryogenesis � Nematode � Giant cell �Promoter regulation
Introduction
Heat-shock proteins (HSPs) are ubiquitous proteins with a
great abundance and diversity in plants. They have been
divided into six different classes on the basis of their
subcellular localisation, immunological cross-reactivity
and sequence similarity (Waters et al. 1996; Scharf et al.
2001). Their role during heat shock has been often studied
(Sun et al. 2002; Yu et al. 2005); small heat-shock proteins
(sHSPs) form aggregates with other sHSPs and different
protein substrates, through which they act as molecular
chaperones (Lee et al. 1997; Heckathorn et al. 1998; Low
et al. 2000). sHSPs are not only induced after heat shock but
M. Barcala � A. Garcıa � C. Fenoll � C. Escobar (&)
Facultad de Ciencias del Medio Ambiente, Universidad de
Castilla-La Mancha, Campus de la Real Fabrica de Armas,
45071 Toledo, Spain
e-mail: [email protected]
P. Cubas
Centro Nacional de Biotecnologıa, Departamento de Genetica
Molecular de Plantas, Campus Universidad Autonoma de
Madrid, 28049 Cantoblanco, Madrid, Spain
C. Almoguera � J. Jordano
Instituto de Recursos Naturales y Agrobiologıa, Consejo
Superior de Investigaciones Cientıficas, Apartado 1052,
41080 Seville, Spain
123
Plant Mol Biol (2008) 66:151–164
DOI 10.1007/s11103-007-9259-3

also by other abiotic stimuli, such as cold, water deficit,
heavy metals, ozone, UV radiation and c-irradiation
(Almoguera et al. 1993; Banzet et al. 1998; Hong and
Vierling 2001; Sun et al. 2002). Furthermore, sHSP accu-
mulation is developmentally regulated, notably during seed
and pollen development (Coca et al. 1994; Wehmeyer and
Vierling 2000; Volkov et al. 2005) and fruit maturation
(Fray et al. 1990; Dure 1993; Low et al. 2000). The
expression of sHSP genes in these situations is regulated
mainly by cis-elements, described as heat-shock elements
(HSEs) present in their promoters. The HSEs are arranged in
clusters of three or more inverted repeats of the sequence 50-nGAAn-30, to which, heat-shock factors (HSFs) bind to
control transcription (Carranco et al. 1997; Nover et al.
2001). The role of these elements in sHSP genes expression
in response to different stimuli is beginning to be understood
and different combinations and/or particular sequences in
the HSE cores are crucial for their regulation under different
circumstances, such as embryogenesis or heat shock (Coca
et al. 1996; Almoguera et al. 1998, 2002b). In addition,
plant HSFs belong to gene families with high complexity
and more than 21 different HSFs have been described so far
(Nover et al. 1996, 2001). Their functional specificity has
been demonstrated only in a few cases. These include par-
ticular HSFs involved in transcriptional activation during
embryogenesis (Almoguera et al. 2002b), in apoptotic
responses (Yamanouchi et al. 2002), in early oxidative
stress responses to H2O2 (Davletova et al. 2005) or in the
heat-stress response (Mishra et al. 2002).
We found that the promoter of Hahsp17.7G4 (G4 in this
work), a sunflower gene that codes an sHSP expressed during
seed development and in response to water stress or heat
shock (Coca et al. 1996; Almoguera et al. 1998), is also
active in giant cells (GCs) (Escobar et al. 2003). So far, this
is the only sHSP promoter reported to respond to biotic
interactions, in particular to nematode infection (Meloido-
gyne spp.). A short version of the G4 gene promoter
(-83G4) includes only one HSE array (HSE I), which is
crucial for the G4 activation after root-knot nematode
infection in tobacco (Escobar et al. 2003). This minimal
promoter version is also up-regulated after heat shock and, to
a lesser extent, during late embryogenesis (Coca et al. 1996).
Cyst and root-knot nematodes are obligate sedentary
endoparasites which need plants for their reproduction. Upon
infection, they induce the transformation of a selected set of
root cells into a feeding site that contains a few highly spe-
cialised cells (syncytia for cyst nematodes and GCs for root-
knot nematodes) on which they feed. Despite accomplishing
similar functions, the ontogeny of both nematode-induced
cell types is very different. GCs are the result of a re-
differentiation process, linked to dramatic changes in gene
expression (Gheysen and Fenoll 2002; Ramsay et al. 2004;
Bar-Or et al. 2005; Jammes et al. 2005; Wieczorek et al.
2006). GC differentiation transforms a normal root xylem-
associated cell into a large, multinucleate cell with enlarged
nucleoli, whose dense cytoplasm is rich in mitochondria,
plastids, ribosomes, rough and smooth endoplasmic reticulum
and crystalline, electron-dense inclusions (possibly proteina-
ceous) frequently associated to spiny vesicles (Paulson and
Webster 1970; Bleve-Zacheo and Melillo 1997). These
characteristics are typical of a cell with very active protein
synthesis, possibly needed to attend the nematode demand as
its only nutrient source during the weeks needed for the
nematode to complete its life cycle (Grundler and Bockenhoff
1997). In this cellular context, a transcriptional state that
favours chaperone (such as sHSP) accumulation might par-
tially mimic the response to heat shock.
To obtain a deeper insight into the HSE-mediated reg-
ulation of sHSPs promoters after nematode infection in
comparison to that of the heat-shock and embryogenesis
responses, we have studied the activity in GCs of addi-
tional sHSP gene promoters, Hahsp17.6G2 (G2) and
Hahsp18.6G1 (G1), closely related to G4, that differ in
their HSE array structures and in their regulation during
late embryogenesis and heat shock (Coca et al. 1996;
Carranco et al. 1997, 1999; Almoguera et al. 2002b).
Our results show that the G2 promoter was activated after
nematode infection in tobacco transgenic plants. In contrast,
G1 was not induced after nematode infection. We have also
found that a mutated version of the G4 promoter (G4MutP),
which shows a greatly reduced late embryogenesis response
(Almoguera et al. 2002b), was active in GCs. Taken together,
our results indicate that transcriptional activation in response
to root-knot nematode infection rely on mechanisms different
from those operating during late embryogenesis but is similar
to those of the heat-shock response and that distinct HSE
array structures may be related to this behaviour. We have
also found that the G4 promoter is specific for root-knot
nematodes, since it is not induced in syncytia formed by the
cyst nematode Globodera tabacum. We have consistently
shown that the class I sHSPs encoded by endogenous genes
accumulate differentially in tobacco GCs, parallelling the
nematode-induction of the heterologous class I sHSP pro-
moters. Therefore, we suggest for these proteins a putative
role during GC formation and/or maintenance, perhaps sim-
ilar to their role during the heat-shock response.
Materials and methods
Infections with nematodes
Meloidogyne incognita infections
Transgenic tobacco seeds from T0 plants containing
different sHSP promoter constructs (Hahsp17.7G4,
152 Plant Mol Biol (2008) 66:151–164
123

Hahsp17.6G1 and Hahsp18.6G2) were provided by
Dr. J. Jordano (Coca et al. 1996; Carranco et al. 1999;
Almoguera et al. 2002b). The seeds were surface-sterilised
and selected in MS medium (Murashige and Skoog 1962)
supplemented with 50 lg/ll of kanamycin. For nematode
infection, resistant tobacco and Arabidopsis homozygous
plants were transferred to Gamborg B5 plates (as in
Escobar et al. 2003) and grown at 24–25�C, 70% humidity
in long day photoperiods. We tested at least five indepen-
dent lines per construct for the analyses in tobacco and four
lines in Arabidopsis.
The M. incognita MIK population (Escobar et al. 2003)
was maintained in an in vitro culture on cucumber roots.
The plates were kept in darkness at 28�C and 60%
humidity. Mature egg masses were collected from the
cucumber roots and kept in sterile tap water in the previous
conditions. After 3–4 days, freshly hatched J2 were used to
inoculate either tobacco or Arabidopsis roots. An average
of 10–12 J2s were added per root tip. Gall formation was
carefully followed by monitoring the infection every 24 h
under a Leica MZ6 microscope. Each newly formed gall
was single-labelled. Thus, days post-infection (dpi) were
measured with an error of less than ±24 h. A minimum of
30–80 galls were scored for each independent line tested.
Globodera tabacum infections
Tobacco plants were selected in MS medium (Murashige
and Skoog 1962) supplemented with kanamycin (50 lg/ll)
for 1 week. After this period, the seedlings were transferred
either to 50-ml conical tubes filled with 16 ml of sand
under sterile conditions and kept in a growth chamber at
24�C in long day periods or to Petri dishes with Gamborg
B5 medium, as previously described (Escobar et al. 2003).
Plants growing in conical tubes were watered twice a week
alternately with either water or liquid MS medium sup-
plemented with vitamins. The tubes were covered with
Parafilm1 to limit water loss. The cysts were sterilised as
in Heungens et al. (1996) with minor modifications. Plants
in tubes were infected with 150 Globodera tabacum J2 and
those grown in Petri dishes with 250 J2 per plate. On
average, 35 cysts were tested in each line.
Arabidopsis transformation and characterisation
Arabidopsis thaliana Col-0 plants were Agrobacterium
tumefaciens-infiltrated with the construct -83G4::GUS, as
described in Coca et al. (1996) using the floral dip method
(Clough and Bent 1998). Transformants were selected by
consecutive cycles of kanamycin selection and self-prog-
eny production. Only the lines with T1 and T2 progeny
showing a classical segregation rate for one locus were
considered as homozygous lines carrying a single copy
insertion. Seeds from T3 plants were used for nematode
infection and plant characterisation. To achieve embryo-
genesis histochemical analysis, every newly opened flower
was labelled in order to set its age. For this analysis, the
plants were grown in soil at 23–24�C and 70% humidity
and with a long day photoperiod.
Histochemical GUS assay
Histochemical assays of GUS activity were performed
mainly as described by Escobar et al. (2003). To avoid
temperature variations that could promote sHSP induction,
histochemical analyses were performed under the same
growth conditions in darkness. For heat-shock experiments,
the plants were placed at 42�C for 3 h and were allowed to
recover at growth conditions for another 3 h before car-
rying out the histochemical analysis. For histological GUS
localisation, positive galls were fixed and embedded in
Araldyte (TAAB, Aldermaston, UK) as in Escobar et al.
(2003). Micrographs from semi-thin sections (6–7-lm
thick) were taken with either Nikon 90i or Leica DM IRB
stereomicroscopes.
Western analysis
Protein extracts were obtained from hand-dissected tobacco
galls or uninfected or heat-shocked roots. The total protein
was extracted in 19 SDS buffer containing 62.5 mM Tris
HCl pH 6.8, SDS 1% (w/v), 2-mercaptoethanol 2.5% (v/v)
and a protease inhibitor cocktail (according to the manu-
facturer’s instructions; Sigma-Aldrich, St. Louis, Missouri).
Protein extracts were concentrated using Microcon YM-10
centrifugal filters (Millipore, Bedford, Massachusetts) and
quantified by Bradford assay. Samples were separated by
sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE) in a 12.5% polyacrylamide gel and transferred
to a PVDF membrane (Hybond-P, Amersham Biosciences
Technologies, UK) with a semi-dry transfer cell (Bio-Rad,
Hercules, California). PVDF membrane background binding
was blocked with 5% dry milk in Tris-buffered saline with
Tween (TBST) and incubated with either anti-class I sHSP
antibodies (Almoguera et al. 1993) or the respective pre-
immune sera, diluted 1:1000 in 5% dry milk. Anti-rabbit
IgG-AP-conjugated antibodies (Sigma-Aldrich, St. Louis,
Missouri) diluted 1:5000 were used as secondary antibodies,
followed by colorimetric detection with a nitrotetrazolium
blue/5-bromo-4-chloro-3-indolyl phosphate solution (NBT/
BCIP; Sigma-Aldrich, St. Louis, Missouri and Roche
Diagnostics GmbH, Mannheim, Germany).
Plant Mol Biol (2008) 66:151–164 153
123

Immunolocalisation assay
Tobacco galls and uninfected control roots were sectioned
and incubated in freshly made fixative (4% (w/v) formal-
dehyde, 0.1 M PIPES, 0.25% (v/v) glutaraldehyde, 1 mM
DTT, 0.1% (v/v) Tween-20, 0.1% (v/v) Triton X-100)
twice for 10 min under vacuum. The fixative was replaced
with fresh solution and the samples were kept at 4�C
overnight. Then, they were dehydrated in ethanol (10%,
30%, 60%, 70%, 85%, 90% and 100% in water) at 4�C. At
the 90% and 100% ethanol steps, 0.1% (w/v) eosin Y was
added to stain the tissues. A complete dehydration was
carried out by incubating the samples in a 3:1, 1:1 and 1:3
series of ethanol (Histoclear; National Diagnostics,
Atlanta, Georgia). Finally, the samples were embedded in
Paramat extra Pastillated Gurr (BDH Lab Supplies, Poole,
UK).
Paraffin blocks were sectioned with a Leica RM 2125
rotary microtome. The sections (10-lm thick) were
mounted on poly-lysine-coated slides (Polysine, Menzel-
Glaser, KG, Germany). After de-waxing and rehydration,
the sections were incubated in primary antibody (1:200, at
4�C overnight), followed by the secondary antibody pre-
viously described for Western blot (1:200 for 45 min at
room temperature), both diluted in TBST.
Statistical analysis of data
The value distributions of the percentages of GUS-stained
galls were compared between plants carrying different
promoter::GUS fusions by the non-parametric test of U
Mann-Whitney and using the Statistical Package for the
Social Sciences (SPSS Inc., Chicago, Illinois). Pairwise
comparisons between all possible groups were performed
to analyse the differences of the effect of the chimaeric
gene on GUS expression. The level of significance was set
to P \ 0.05.
Results
Three differentially regulated sHSP gene promoters
show differential nematode response in tobacco
To gain insight into the regulation of sHSP genes through
their HSEs in GCs, we compared the promoter structure
and response of G4 with those of two sHSP genes, G1 and
G2, that code for very similar proteins but differ in their
expression patterns. We also analysed a mutant version of
the G4 promoter called the ‘‘perfect HSE mutant’’ G4MutP,
as it has a higher number of ‘‘perfect’’ core HSE repeats
(exactly matching the consensus sequence (50-nGAAn-30).
These changes in promoter structure are due to three point
mutations at positions -86 (TCAA changed by agAA),
-108 (TTT changed by TTc) and -116 (GGAA changed
by aGAA; Fig. 1A), which improves HSF binding and
maintains the heat-shock activation but reduces the late-
embryogenesis response (Almoguera et al. 2002b).
A detailed analysis of the promoter structure reveals
that, similarly to the G4 promoter (Escobar et al. 2003), the
G2 promoter has two HSE arrays, one proximal and one
distal (HSE I and HSE II, respectively; Fig. 1A). In con-
trast, G1 carries only one HSE array and in a distal position
(Fig. 1A). In addition, the G4 promoter contains three HSE
head–tail motifs (nGAAn; nTTCn), which include two
active head modules, one inactive head module and three
active tail modules in HSE I (ThTHTH; Fig. 1A). G2
carries fewer HSE head–tail motif combinations (THTHt)
in HSE I (Fig. 1A). In contrast, G1 contains a higher
number of HSE motifs as compared to G2. However, G1
HSE motifs present a distinct configuration (THHHtH;
Fig. 1A), as three adjacent active head modules are local-
ised in between an active and an inactive tail module
(Fig. 1A). Moreover, in G1, the distance between the HSE
and the TATA box is much longer than that of HSE I in G2
or G4 (49 bp as compared to 12 bp in G2 and 21 bp in G4;
Fig. 1A). Additionally, several putative CAAT boxes were
identified immediately upstream and in between the HSE
arrays of G2 and G4, at positions -177 and -72 bp for G4
and -141, -127 and -84 bp for G2. In contrast, a single
CAAT box was identified in the G1 promoter downstream
of the HSE array at position -67 pb.
Transgenic tobacco plants carrying translational fusions
of G2 and G1 promoters to the b-glucuronidase (GUS)
reporter gene (G2::GUS and G1::GUS) were infected with
Meloidogyne incognita and analysed 8–15 dpi, a stage
when GCs have a very active metabolism and during which
a high activity of the G4 promoter was previously reported
(Escobar et al. 2003).
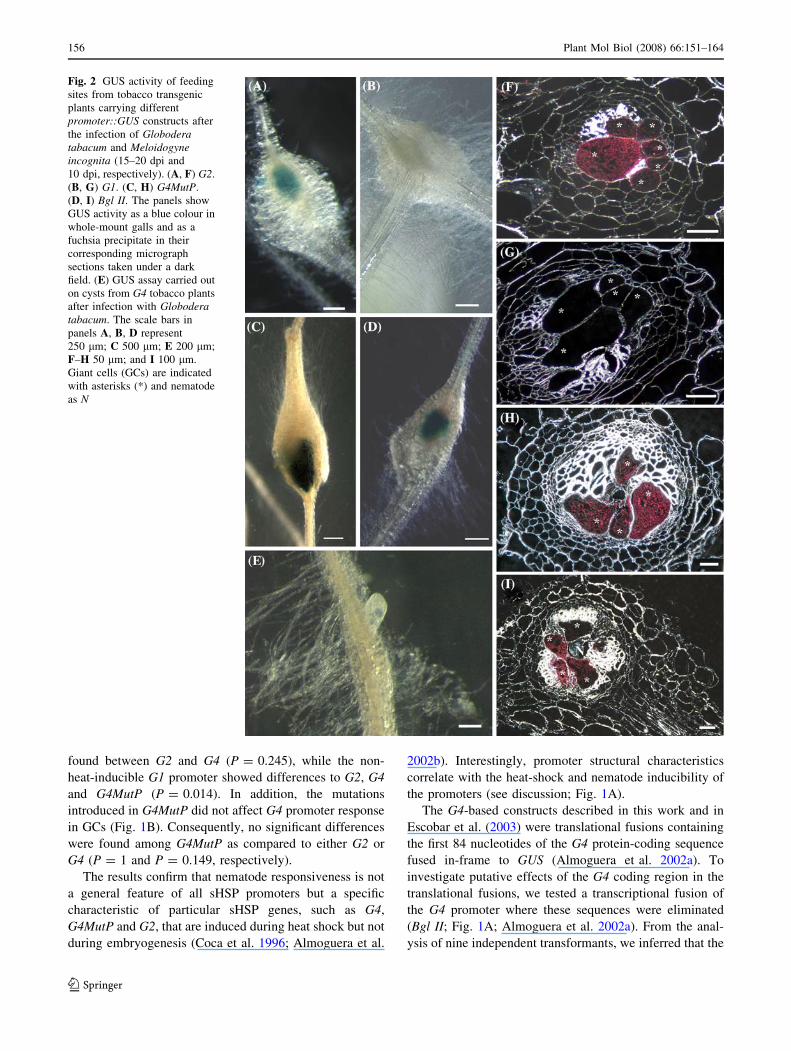
Plants carrying the G2 promoter showed a blue signal
centred in galls (Fig. 2A), which corresponded to a blue
precipitate strongly concentrated in GCs, as shown in semi-
thin sections (Fig. 2F). A faint patchy expression was
occasionally detected along the root and at the root tip
(data not shown) in all of the tested lines. In contrast, G1
nematode inducibility was undetectable, as shown in galls
and semi-thin sections of galls (Fig. 2B, G). The mutated
version of the G4 promoter, G4MutP, containing point
mutations in the HSE arrays that convert inactive to active
modules (Fig. 1A; Nover et al. 2001), was active in galls,
with the signal centred in GCs (Fig. 2C, H).
Analysis of the frequency of blue gall occurrence con-
firmed that the G2 promoter responds to nematode
infection similarly to G4 and that G1 is not responsive to
this stimulus (Fig. 1B). No significant differences were
154 Plant Mol Biol (2008) 66:151–164
123

Fig. 1 Small heat-shock protein (sHSP) promoter structures and
frequency of blue galls. (A) Structure of the different sHSP promoters
analysed based on Scharf et al. (2001). Core heat-shock elements
(HSEs) are referred as active head (nGAAn, nGAnn or nGnAn) or tail
(nTTCn, nnTCn or nTnCn) modules and are represented as white
boxes ( , ). Inactive head or tail modules lack G or C residues
and/or the other two core nucleotides (AA or TT) and are indicated
with grey boxes. HSE arrays are named I and II and the TATA box is
represented with a striped box. Numbers under promoter diagrams
indicate the position of the initial and last nucleotide of either the HSEarrays or the TATA box to the transcriptional starting point. The
numbers between slashes represent the distance to adjacent promoter
elements. (B) The histograms represent the frequency of blue galls
(10 dpi) of the different tobacco transformants. The data are means of
at least five independent lines per construct. The data from four
independent experiments per line were pooled. The table shows
activation of the G4, G2, G1 and G4MutP promoters after nematode
infection, heat shock and during late embryogenesis
Plant Mol Biol (2008) 66:151–164 155
123
found between G2 and G4 (P = 0.245), while the non-
heat-inducible G1 promoter showed differences to G2, G4
and G4MutP (P = 0.014). In addition, the mutations
introduced in G4MutP did not affect G4 promoter response
in GCs (Fig. 1B). Consequently, no significant differences
were found among G4MutP as compared to either G2 or
G4 (P = 1 and P = 0.149, respectively).
The results confirm that nematode responsiveness is not
a general feature of all sHSP promoters but a specific
characteristic of particular sHSP genes, such as G4,
G4MutP and G2, that are induced during heat shock but not
during embryogenesis (Coca et al. 1996; Almoguera et al.
2002b). Interestingly, promoter structural characteristics
correlate with the heat-shock and nematode inducibility of
the promoters (see discussion; Fig. 1A).
The G4-based constructs described in this work and in
Escobar et al. (2003) were translational fusions containing
the first 84 nucleotides of the G4 protein-coding sequence
fused in-frame to GUS (Almoguera et al. 2002a). To
investigate putative effects of the G4 coding region in the
translational fusions, we tested a transcriptional fusion of
the G4 promoter where these sequences were eliminated
(Bgl II; Fig. 1A; Almoguera et al. 2002a). From the anal-
ysis of nine independent transformants, we inferred that the
Fig. 2 GUS activity of feeding
sites from tobacco transgenic
plants carrying different
promoter::GUS constructs after
the infection of Globoderatabacum and Meloidogyneincognita (15–20 dpi and
10 dpi, respectively). (A, F) G2.
(B, G) G1. (C, H) G4MutP.
(D, I) Bgl II. The panels show
GUS activity as a blue colour in
whole-mount galls and as a
fuchsia precipitate in their
corresponding micrograph
sections taken under a dark
field. (E) GUS assay carried out
on cysts from G4 tobacco plants
after infection with Globoderatabacum. The scale bars in
panels A, B, D represent
250 lm; C 500 lm; E 200 lm;
F–H 50 lm; and I 100 lm.
Giant cells (GCs) are indicated
with asterisks (*) and nematode
as N
156 Plant Mol Biol (2008) 66:151–164
123

28 extra amino acids in G4 as compared to Bgl II were not
affecting the GUS expression in GCs (Fig. 1B, 2D, I).
Hence, the translational fusions used were clear indicators
of the promoter activity.
The G4 promoter is activated by root-knot
but not by cyst nematodes
It has been previously reported that some plant genes and/or
promoters are differentially regulated in the feeding cells
induced by cyst and root-knot nematodes (Vercauteren et al.
1998; Gheysen and Fenoll 2002; Mitchum et al. 2004).
Therefore, we decided to analyse the Meloidogyne spp.-
inducible G4 promoter upon infection with the cyst nema-
tode Globodera tabacum. We selected those transgenic lines
that showed a higher induction upon M. incognita infection;
one line contained the G4::GUS construct and two lines
had the minimal GC-responsive HSE promoter elements
(-83G4; Escobar et al. 2003). Syncytia at 10–20 dpi did not
show any GUS activity, either in in vitro-grown plants or in
hydroponic sand cultures, for either promoter construct
(Fig. 2E). Therefore, the G4 promoter responds to root-knot
but not to cyst nematodes.
Class I low-molecular-weight heat-shock proteins
accumulate in giant cells
In order to investigate whether G4 and G2 promoter acti-
vation in the transgenic lines is parallelled by the tobacco
endogenous genes and leads to the accumulation of sHSPs
in GCs, Western-blot assays were carried out in protein
extracts from tobacco galls and uninfected roots. Using an
anti-sHSP antibody that specifically recognises class I
sHSPs (Almoguera et al. 1993), a very faint band of the
expected size (about 18 kDa) could be detected in gall
extracts (Fig. 3A) but not in control uninfected root tissue
(Fig. 3A). Protein extracts from tobacco roots subjected to
heat-shock treatment and sunflower embryos were also
loaded as positive controls (Fig. 3A). To further confirm
that sHSPs were localised in gall cells, we performed im-
munolocalisation assays in gall sections and compared
them to control root sections using the same detection
system. A clear purple precipitate could be observed,
which was highly restricted to GCs (Fig. 3B), while the
surrounding tissues (Fig. 3B) control uninfected root sec-
tions (not shown), as well as gall sections incubated with
preimmune antisera did not show any signal (Fig. 3C). This
result indicates that the endogenous sHSP genes are highly
expressed at the protein level in GCs, thus, opening the
possibility of a biologically meaningful role for sHSPs in
GCs development and/or physiology.
The -83bp G4 promoter is also induced in giant cells
and during heat shock in transgenic Arabidopsis
Helianthus annum is a recalcitrant plant for transformation
(Lewi et al. 2006). Therefore, in order to assess whether
the nematode activation of sHSPs is conserved in plant
species other than tobacco, we transformed Arabidopsis. A
comparable response in Arabidopsis could simplify further
analysis by exploiting molecular genetics tools available in
Fig. 3 Abundance of class I sHSP in galls. (A) Western blot carried
out with anti-sHSP class I antibodies. Bands of approximately 18 kDa
appear in lanes loaded with total protein from sunflower embryos
(lane E, 4 lg) and heat-stressed tobacco roots (lane HR, 135 lg) used
as positive controls. A faint band can be detected in tobacco gall
protein extracts (lane G, 135 lg), whereas no signal was detected in
uninfected control root protein extracts (lane CR, 135 lg). (B, C)
Immunolocalisation of class I sHSPs in tobacco galls 10 dpi. Sections
were incubated with either anti-class I sHSP antibody (B) or pre-
immune serum (C). A purple precipitate centred in the GCs (*) is
detected in B. The scale bars represent 50 lm
Plant Mol Biol (2008) 66:151–164 157
123

Arabidopsis and this might help in finding the putative role
of the class I sHSPs and HSFs in GCs. Arabidopsis plants
carrying the minimal nematode-inducible promoter version
-83G4 fused to GUS were characterised after nematode
infection and also during plant development and heat
shock, since no information on the behaviour of the
Helianthus annus G4 promoter in Arabidopsis was avail-
able (Fig. 1A).
Arabidopsis transgenic plants carrying the -83G4::GUS
construct were analysed 7 days after nematode infection,
revealing a clear GUS signal inside galls (Fig. 4A),
although the percentage of positive galls for all tested lines
was reduced (30–50%) as compared to tobacco (Escobar
et al. 2003). Therefore, the nematode inducibility of the
sunflower promoter in tobacco plants was maintained in
Arabidopsis. Consistent with the activity observed in
tobacco (Escobar et al. 2003), semi-thin sections of the
positive galls showed a GUS signal located in GCs
(Fig. 4B), with a faint and diffuse signal in the surrounding
tissues, which might be due to diffusion of the GUS pre-
cipitate. Occasionally, patchy GUS expression in scattered
single cells was observed within the root in both infected
and uninfected plants (Fig. 4C).
During germination, an intense GUS expression signal
was found in cotyledons of seedlings at growth stages 0.5–
0.7 (as defined by Boyes et al. 2001; Fig. 5A). This coty-
ledon pattern disappeared completely by the 1.02–1.04
growth stages and GUS expression became restricted to the
shoot apical meristem (SAM; Fig. 5B). GUS activity could
not be detected any longer during development in any
vegetative tissues (Fig. 5C), apart from some occasional
activity in lateral root initiation points (data not shown). No
signal was detected in developing floral organs (Fig. 5D),
except for a very restricted blue signal at the base of the
siliques, 3–5 days post-anthesis (dpa) that decreased during
silique maturation (Fig. 5D, E). A faint signal was also
detected at the funiculus 1–5 dpa (Fig. 5F). However, no
GUS activity was detected during seed maturation either in
developing embryos or in mature seeds (Fig. 5F–I). In
tobacco, a low level of GUS activity was reported
in -83G4 developing seeds, albeit activity was not
completely abolished as in Arabidopsis (Coca et al. 1996).
We cannot rule out a transcriptional activation in Arabid-
opsis embryos below the detection level of our
histochemical assays or a different post-transcriptional
regulation of the -83G4 gene product in Arabidopsis as
compared to tobacco seeds, as proposed for similar chi-
maeric genes (Prandl et al. 1995).
After heat shock, all tissues of seedlings during germi-
nation responded to the treatment (data not shown;
Fig. 5J). Similarly, adult plants showed a strong blue
staining in the roots, in the leaf vascular tissue and espe-
cially intensely in the stem (Fig. 5K, L). During fruit
maturation, green siliques subjected to heat shock were
completely blue stained (Fig. 5M). As silique maturation
progressed, GUS signal was restricted to the funiculus, the
septum and the base of the silique (Fig. 5N). However, no
GUS signal was observed after heat shock in mature seeds
(Fig. 5N). In accordance with our observations in Arabid-
opsis, adult -83G4 tobacco plants showed a similar
response to heat shock, which was especially intense in
stems (Coca et al. 1996), suggesting a remarkable conser-
vation of the promoter regulation in two heterologous
systems. In this respect, the promoter activity of G1, G2
and G4 during heat-shock in tobacco was in agreement
with their mRNA accumulation in sunflower. In contrast,
the accumulation of G4 mRNAs in sunflower after water
stress did not correspond to its promoter activity in tobacco
(Coca et al. 1996; Carranco et al. 1999; Almoguera et al.
2002a).
Discussion
Many aspects regarding the regulation of sHSPs through
their HSEs remain unknown (Scharf et al. 2001). One
example is the sunflower G4 promoter, expressed in
tobacco GCs induced by root-knot nematodes, whose
expression is mediated by a specific array of HSEs
(Escobar et al. 2003). GC expression of various G4 pro-
moter versions indicated a parallel with their heat-shock-
mediated expression and also with some aspects of their
Fig. 4 Histochemical GUS assay in Arabidopsis plants carrying a -
83G4::GUS fusion, 7 dpi. (A) Gall after M. incognita infection
showing GUS activity as a blue precipitate. (B) Semi-thin section of a
gall. GUS signal, as a fuchsia precipitate, is mainly concentrated in
the GCs. (C) Scattered blue signals in roots. GCs are indicated with
asterisks (*) and nematode as N
158 Plant Mol Biol (2008) 66:151–164
123

late-embryogenesis regulation (Escobar et al. 2003). This
raises the issue of what could be the signals inside GCs that
induce the sHSP promoters and whether they are related to
their regulation during heat-stress and/or late embryogen-
esis. Therefore, we examined in further detail the
transcriptional responses of several sHSP gene promoters
in response to nematode infection. These promoters were
selected because they showed differences in their promoter
structures and in their regulation under heat shock or late
embryogenesis, which may correlate to their responses
after nematode infection.
The regulation of three sHSP promoters in giant cells
differs from their regulation during late embryogenesis
and is associated to different structural features
In a previous work, HSEs were identified as necessary for
the transcriptional activation of G4 in GCs, probably
through HSFs (Escobar et al. 2003) and partial differences
in its regulation after heat shock, embryogenesis and
nematode infection were described.
In the present work, we have studied the nematode-
inducibility of two additional sHSP promoters and one new
version of the G4 promoter. Results were compared to
previous observations of the G4 promoter. The mutant
version of the G4 promoter (G4MutP) showed the same
activation pattern as G4 after nematode infection (Fig. 1, 2)
but a very low activation during late embryogenesis
(Almoguera et al. 2002b). This indicates a clear difference
between GCs and late embryogenesis regarding the G4
promoter activation and suggests that imperfect repeats at
positions -86 and -108 are not crucial for the binding of
putative HSFs and/or other transcriptional regulators that
participate in nematode induction, whereas they were
essential for late embryogenesis activation (Fig 1;
Almoguera et al. 2002b).
The promoters of two additional sHSP genes (G1 and
G2) were tested for nematode-inducibility. These promot-
ers and that of G4 differ in their HSE array structure
(Fig. 1). The G1 promoter was inactive in GCs, as well as
during heat-shock (Figs. 1 and 2; Carranco et al. 1999),
despite its efficient activation in late embryogenesis
(Carranco et al. 1999), probably via HaHSFA9 or its
Fig. 5 Histochemical GUS assay of Arabidopsis transgenic plants
carrying a -83G4::GUS fusion. (A) During germination, GUS
activity is restricted to the cotyledons from stage 0.5 to 0.7 (Boyes
et al. 2001). (B) Stage 1.02 to 1.04 seedlings with GUS expression in
SAM. (C) No GUS staining in adult plants. (D) Flowers and siliques
0–5 dpa. (E) Blue staining at the base of the silique, 7–10 dpa. (F)
Silique 4–5 dpa with faint GUS staining at the funiculus (arrow-heads). (G, H) Siliques 15 and 18 dpa, respectively. (I) Embryos
isolated at 11, 15 and 18 dpa with no GUS staining. After heat-shock
treatment, b-glucuronidase is induced in: (J) Vegetative tissues of
seedlings; (K, L) Adult plants with GUS staining in leaf vascular
tissue and strong signal in stems; (M, N) GUS activity diminishes
through development of heat-stressed siliques from 5 to 9 dpa,
respectively. Scales bars in panel I represents 100 lm; A, B, D–F, J,
L and N 0.5 mm; and C, G, H, K and M 1 mm
Plant Mol Biol (2008) 66:151–164 159
123

orthologous tobacco HSF (Almoguera et al. 2002b). This
demonstrates that the presence of functional HSEs in an
sHSP promoter is not sufficient to confer GCs expression.
In contrast, the G2 promoter responded to nematodes
(Figs. 1 and 2), as well as to heat shock, but its transcripts
did not accumulate in late embryogenesis (Coca et al.
1996; Carranco et al. 1997). These results strongly suggest
that the HSFs involved in the nematode response in
tobacco are able to activate transcription from perfect
(G4MutP) or imperfect (G2, G4) HSEs but only from
promoter-proximal HSE locations, which are shared by the
G2 and G4 promoters but not by G1 (Fig. 1A). Further-
more, HSE II in G4 is dispensable for nematode activation
in tobacco and Arabidopsis (Escobar et al. 2003 and this
work). The heat-shock induction of sunflower sHSP pro-
moters in tobacco also depends on the presence of proximal
HSE, whereas distal HSE contributes to the developmental
regulation (Carranco et al. 1999). These also support a
higher similarity of the sHSP regulation during heat shock
and in GCs as compared to late embryogenesis.
Additionally, the G1 HSE array does not present the
alternate head-tail-head pattern observed in HSE I of G2
and G4, which could constitute a poor combination for
HSF primer binding (see results and Fig. 1). This fact
could explain, at least in part, their different regulation in
GCs and suggests that HSFs and other transcription factors
that participate in G1 promoter activation during late
embryogenesis differ from those involved in G2 and G4
expression in GCs and heat shock. In this respect, the
combination of active and inactive modules, as well as
their position, seem to be important for the proper inter-
action of HSFs and other factors in sHSP promoters (Scharf
et al. 2001; Bharti 2003), as described for the synergistic
effect between tomato HSFA1 and B1, as well as their
cooperation with other activators, such as HAC1 and
ASF1/2 (Bharti et al. 2004). Moreover, the HSE array in
G1 showed a low binding affinity to certain HSFs, such as
hHSF-1 (Carranco et al. 1997). In addition, in G1 the
distance between the HSE and the TATA box is much
longer than that of HSE I in G2 or G4 (Fig. 1). This dif-
ference may be relevant for the activation of G2, G4 and
-83G4 in GCs and not G1, as some HSFs interact directly
with elements of the basal transcription machinery (Reindl
and Schoffl 1998). Additionally, several putative CAAT
boxes were identified immediately upstream and in
between the HSE arrays of G2 and G4, whereas a single
CAAT box was identified in the G1 promoter downstream
of the HSE array. CAAT sequences placed in between and/
or immediately upstream to the HSEs have been described
to act cooperatively with HSE to increase promoter activity
in Gmhsp17.3-B (Rieping and Schoffl 1992). Therefore,
these CAAT boxes might participate in the activation of G2
and G4 after nematode infection but not in G1. All of these
structural differences of G2 and G4 as compared to G1
might be crucial for their differential activation in GCs.
Although the HSE I present in the -83G4 is crucial for
nematode activation in tobacco (Escobar et al. 2003), the
lines tested were translational fusions and included, besides
the promoter region, an additional 163 bp sequence
downstream from the transcription initiation site. A search
for putative cis-elements in the 246 pb that constitute the
minimal nematode-inducible sequence using three different
databases identified several homologies to known regula-
tory sequences from other plant genes (Escobar et al.
2003). Therefore, we cannot exclude the participation of
some of these putative cis-elements in the activation of
-83G4 in Arabidopsis GCs. However, in this work, we
found that a transcriptional fusion of the G4 promoter
where these 163 bp were eliminated (Bgl II; Almoguera
et al. 2002a) maintains its inducibility in tobacco GCs.
Thus, the possibility that the putative cis-elements identi-
fied could play a role in sHSP regulation in GCs is low.
Additionally, no relevant putative cis-elements in the
sequences between HSE I and the TATA box could be
identified in -83G4 (Fig. 1).
The sunflower -83G4 minimal promoter maintains its
regulation in Arabidopsis during development, heat
shock and nematode infection
We also demonstrate that both the nematode and the heat-
shock responses of the minimal promoter version -83G4
are conserved between tobacco and Arabidopsis. This short
promoter version, which does not contain the distal HSE
array (HSE II; Fig. 1), showed induction after heat shock in
vegetative tissues and after nematode infection in tobacco
(Coca et al. 1996; Almoguera et al. 1998; Escobar et al.
2003) and in Arabidopsis (Figs. 4 and 5). With respect to
late embryogenesis expression, the -83G4 promoter was
silent in Arabidopsis (this work) and had a low activity in
tobacco (Escobar et al. 2003). Therefore, the putative HSFs
and, possibly, other transcriptional regulators that must
interact with the G4 HSE I (Fig. 1) in GCs and during heat
shock seem to be functionally conserved in different plant
species.
Particular HSFs might be involved in sHSP gene
expression in giant cells
The data presented in this paper strongly suggest that the
regulation of sHSP promoters through HSEs in GCs
depends on HSFs, combinations of HSFs (Scharf et al.
1998; Rojas et al. 2002; Bharti et al. 2004) or combina-
tions of HSFs with other interacting proteins (Bharti et al.
160 Plant Mol Biol (2008) 66:151–164
123

2004; Diaz-Martin et al. 2005), which are different
from those involved in their regulation during late-
embryogenesis.
Plant HSFs show a high complexity and functional
specificity has been demonstrated in only a few cases
(reviewed by Baniwal et al. 2004). It is possible that still
uncharacterised HSFs might be involved in the nematode
response. The intrinsic characteristics of these HSFs
regarding binding to HSEs would match the observed HSE/
promoter structural requirements for nematode response in
tobacco and Arabidopsis.
Confirmation of the involvement of particular HSFs in
G4 and G2 promoter regulation after nematode infection
via loss-of-function mutants might be precluded by func-
tional redundancy amongst some members of the gene
family and lines carrying loss-of-function alleles for sev-
eral HSF genes might be necessary (Morimoto 1998; Miller
and Mittler 2006), since a master regulator of the heat-
shock response as in tomato (Mishra et al. 2002) has not
yet been identified in Arabidopsis (Reindl et al. 1997;
Wunderlich et al. 2003; Lohmann et al. 2004). GCs do not
seem to show a typical heat-shock stress cellular response.
Thus, it is possible that HSFs similar to HSFA3 from
tomato that could represent a developmentally regulated
HSF with expression only in rapidly dividing cells (Nover
et al. 2001) may be the crucial activators of the sHSP
promoters in GCs, whose differentiation involves repeated
endomitosis cycles (revised in Gheysen and Fenoll 2002).
Additionally, HSFs identified as important for the stress
response in Arabidopsis, such as AtHSF1 and AtHSF3,
might also participate in the activation of sHSPs promoters
in GCs (Wunderlich et al. 2003; Lohmann et al. 2004),
particularly AtHSF1, which is also activated during the cell
cycle (Reindl et al. 1997). Future work based on differen-
tially expressed HSFs in GCs identified in a transcriptome
analysis of micro-dissected Arabidopsis GCs (Barcala et al.;
unpublished) will help to distinguish amongst these multiple
possibilities and to assign functional roles of particular HSFs
in GC-specific expressions of sHSP genes.
A function for class I sHSPs in giant cells?
Our results show that class I sHSPs are present in galls and
are particularly abundant in tobacco GCs as compared to
the surrounding root tissues (Fig. 3). This is in complete
accordance with GUS expression data from several heter-
ologous promoters whose genes code for class I sHSPs
(Escobar et al. 2003 and this paper). sHSPs belong to a
large family of abundant proteins that play different roles
throughout plant life. They are not only induced by envi-
ronmental cues, such as extreme heat, heavy metals, water
stress etc. (Vierling 1991; Almoguera et al. 1993) but are
also developmentally programmed to be expressed during
embryogenesis. In prokaryotic cells, some sHSPs are
associated with inclusion bodies formed during high-level
protein expression (Laskowska et al. 1996). In GCs, crys-
talline inclusions have been detected in their cytosol
(Paulson and Webster 1970), perhaps related to the high
protein turnover that rapid GC growth predicts. The high
metabolic rate of these cells inferred from their ultra-
structure (Bird 1961; Bleve-Zacheo and Melillo 1997;
Gheysen and Fenoll 2002) might need chaperones to pre-
vent the aggregation of abundantly synthesised proteins.
Some sHSPs are particularly effective chaperones in pre-
venting protein aggregation, acting by an ATP-independent
mechanism through conformational changes both depen-
dent on and independent of heat (Sun et al. 2002; Haslbeck
et al. 2005). In addition, sHSPs are important for the pro-
tection of multiple cellular proteins and a wide variety of
cellular activities, such as transcription, cell signalling and
the assembly/disassembly of cytoskeletal components
(Basha et al. 2004), all of which are processes described to
be altered in GCs (Gheysen and Fenoll 2002; de Almeida
Engler et al. 2004). Nematode feeding habits should also
be taken into account, as they feed through disposable
‘‘feeding tubes,’’ thought to be formed by secretions
extruded from their stylets. These tubes are poorly
described crystalline structures found in the cytoplasm of
GCs and syncytia (Hussey and Mims 1991), which have to
be assembled and attached to the nematode stylet in each
feeding cycle and that might recruit plant chaperons as
well. Finally, the parasites might pre-digest proteins by
injecting proteases inside the feeding cells (Vanholme
et al. 2004; Shingles et al. 2007), which might also signal
chaperones overproduction.
Our finding that sHSP accumulate in GCs, together with
the great similarity between GCs and heat-shock responses
regarding sHSP promoter regulation, suggests that the
metabolic state of the GCs might somehow mimic a heat-
shock response or even a cellular state with conditions
similar to those elicited by osmotic, water or heat stress
when abundant unfolded proteins present in the cell need
chaperone activities. Hence, sHSP production in GCs
might be aimed at protecting protein activity during self-
generated stress conditions. GCs are nutrient sinks in which
soluble assimilates are continuously ingested by the nem-
atode and replenished by the plant (Hussey and Mims
1991). It is possible that cyclic changes in low-molecular-
weight metabolites result in cyclic alterations of the
osmotic balance in the GC. In this respect, several genes
involved in drought tolerance, including genes from car-
bohydrate metabolism, have been described as targets of
HSFs in Arabidopsis (Busch et al. 2005) and a recent work
by Swindell et al. (2007) suggests that sHSPs could con-
tribute to multiple stress tolerance in different plant
Plant Mol Biol (2008) 66:151–164 161
123

species. Moreover, HSFs and HSPs represent an interaction
point at the cross-talk of several stress response pathways.
At the moment, the signals that activate HSFs and other
possible transcription factors in GCs that lead to sHSP
activation are completely unknown. Future research,
including gain- and loss-of-function experiments by
exploiting molecular genetics tools available in Arabid-
opsis, will help in finding both the putative role of the class
I sHSPs in GCs and the transcriptional circuits that lead to
their accumulation.
Nematode-specificity in the transcriptional activation
of the G4 promoter
Syncytia and GCs show differential regulation of a set of
genes that share a common regulation during late-
embryogenesis and/or under different stress conditions,
such as drought (de Meutter et al. 2005). We had described
the induction of the G4 promoter in GCs induced by root-
knot nematodes (Escobar et al. 2003). Our results in
tobacco plants infected with the cyst nematode Globodera
tabacum now establish that G4 is a GC-specific promoter,
at least in tobacco, as we could not detect any GUS activity
in the syncytia of both in vitro and sand-grown plants
(Fig. 2). Furthermore, the minimal promoter version that
was active in tobacco GCs, -83G4, showed also a null
response after cyst nematode infection.
Expression studies of genes with common regulation
during late-embryogenesis, drought stress and nematode
infection (Van der Eycken et al. 1996; Escobar et al. 1999,
2003; de Meutter et al. 2005 and this study) have revealed
their complex regulation in nematode-feeding sites. Some
genes are syncytia-specific (ABI3 and CDeT-27-45; de
Meutter et al. 2005), whereas others are specific for GCs
(Hahsp17.7G4; Escobar et al. 2003). Other genes, such as
LEMMI9, are induced by both nematodes, although the
response to cyst nematodes is weaker (Van der Eycken
et al. 1996, 1999; de Meutter et al. 2005). Our results
support the view that complex sets of transcription factors
are differentially active in syncytia and GCs. Detailed
analysis of each particular promoter and its interacting
transcription factors in different physiological situations
will help in understanding the profound re-programming of
gene expression that takes place in nematode-feeding sites
and which leads to the widely described root morphologi-
cal changes elicited by plant parasitic nematodes.
Acknowledgements We thank Dr. Mugniery for kindly providing
the Globodera tabacum population and Ana Belen Yuste for her
technical help in the analysis of the Arabidopsis transformants. This
work was supported by grants from the Fundacion Ramon Areces and
the Ministerio de Educacion (AGL-2004-08103-C02-02) to CE, from
a collaboration project from the National Science Foundation of the
USA to Elizabeth Vierling and from the European Commission
(QLK5-1999-01501) and the Junta de Comunidades de Castilla-La
Mancha (JCCM, GC-02-011) to CF. MB was a recipient of an FPU
fellowship from the Ministerio de Educacion and AG from the ‘‘Junta
de Comunidades de Castilla-La Mancha.’’
References
Almoguera C, Coca MA, Jordano J (1993) Tissue-specific expression
of sunflower heat shock proteins in response to water stress.
Plant J 4:947–958
Almoguera C, Prieto-Dapena P, Jordano J (1998) Dual regulation of a
heat shock promoter during embryogenesis: stage-dependent role
of heat shock elements. Plant J 13:437–446
Almoguera C, Rojas A, Jordano J (2002a) Reversible heat-induced
inactivation of chimeric beta-glucuronidase in transgenic plants.
Plant Physiol 129:333–341
Almoguera C, Rojas A, Diaz-Martin J, Prieto-Dapena P, Carranco R,
Jordano J (2002b) A seed-specific heat-shock transcription factor
involved in developmental regulation during embryogenesis in
sunflower. J Biol Chem 277:43866–43872
Baniwal SK, Bharti K, Chan KY, Fauth M, Ganguli A, Kotak S,
Mishra SK, Nover L, Port M, Scharf K-D, Tripp J, Weber C,
Zielinski D, von Koskull-Doring P (2004) Heat stress response in
plants: a complex game with chaperones and more than twenty
heat stress transcription factors. J Biosci 29:471–487
Banzet N, Richaud C, Deveaux Y, Kazmaier M, Gagnon J,
Triantaphylides C (1998) Accumulation of small heat shock
proteins, including mitochondrial HSP22, induced by oxidative
stress and adaptive response in tomato cells. Plant J 13:
519–527
Bar-Or C, Kapulnik Y, Koltai H (2005) A broad characterization of
the transcriptional profile of the compatible tomato response to
the plant parasitic root knot nematode Meloidogyne javanica.
Eur J Plant Pathol 111:181–192
Basha E, Lee GJ, Breci LA, Hausrath AC, Buan NR, Giese KC,
Vierling E (2004) The identity of proteins associated with a
small heat shock protein during heat stress in vivo indicates that
these chaperones protect a wide range of cellular functions.
J Biol Chem 279:7566–7575
Bharti K (2003) Tomato heat stress transcription factor HsfB1
represents a novel type of general transcription coactivator with
a histone-like motif interacting with HAC1/CBP. Johann Wolf-
gang Goethe-Universitat, Frankfurt am Main, 126
Bharti K, von Koskull-Doring P, Bharti S, Kumar P, Tintschl-
Korbitzer A, Treuter E, Nover L (2004) Tomato heat stress
transcription factor HsfB1 represents a novel type of general
transcription coactivator with a histone-like motif interacting
with the plant CREB binding protein ortholog HAC1. Plant Cell
16:1521–1535
Bird AF (1961) The ultrastructure and histochemistry of a nematode-
induced giant cell. J Biophys Biochem Cytol 11:701–715
Bleve-Zacheo T, Melillo MT (1997) The biology of giant cells. In:
Fenoll C, Grundler FMW, Ohl SA (eds) Cellular and molecular
aspects of plant–nematode interactions. Kluwer Academic
Publishers, Dordrecht, the Netherlands, pp 65–79
Boyes DC, Zayed AM, Ascenzi R, McCaskill AJ, Hoffman NE, Davis
KR, Gorlach J (2001) Growth stage-based phenotypic analysis of
Arabidopsis: a model for high throughput functional genomics in
plants. Plant Cell 13:1499–1510
Busch W, Wunderlich M, Schoffl F (2005) Identification of novel heat
shock factor-dependent genes and biochemical pathways in
Arabidopsis thaliana. Plant J 41:1–14
162 Plant Mol Biol (2008) 66:151–164
123

Carranco R, Almoguera C, Jordano J (1997) A plant small heat shock
protein gene expressed during zygotic embryogenesis but
noninducible by heat stress. J Biol Chem 272:27470–27475
Carranco R, Almoguera C, Jordano J (1999) An imperfect heat shock
element and different upstream sequences are required for the
seed-specific expression of a small heat shock protein gene. Plant
Physiol 121:723–730
Clough SJ, Bent AF (1998) Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thali-ana. Plant J 16:735–743
Coca MA, Almoguera C, Jordano J (1994) Expression of sunflower
low-molecular-weight heat-shock proteins during embryogenesis
and persistence after germination: localization and possible
functional implications. Plant Mol Biol 25:479–492
Coca MA, Almoguera C, Thomas TL, Jordano J (1996) Differential
regulation of small heat-shock genes in plants: analysis of a
water-stress-inducible and developmentally activated sunflower
promoter. Plant Mol Biol 31:863–876
Davletova S, Rizhsky L, Liang H, Shengqiang Z, Oliver DJ, Coutu J,
Shulaev V, Schlauch K, Mittler R (2005) Cytosolic ascorbate
peroxidase 1 is a central component of the reactive oxygen gene
network of Arabidopsis. Plant Cell 17:268–281
de Almeida Engler J, Van Poucke K, Karimi M, De Groodt R,
Gheysen G, Engler G (2004) Dynamic cytoskeleton rearrange-
ments in giant cells and syncytia of nematode-infected roots.
Plant J 38:12–26
de Meutter J, Robertson L, Parcy F, Mena M, Fenoll C, Gheysen G
(2005) Differential activation of ABI3 and LEA genes upon plant
parasitic nematode infection. Mol Plant Pathol 6:321–325
Diaz-Martin J, Almoguera C, Prieto-Dapena P, Espinosa JM, Jordano
J (2005) Functional interaction between two transcription factors
involved in the developmental regulation of a small heat stressprotein gene promoter. Plant Physiol 139:1483–1494
Dure L 3rd (1993) The LEA proteins of higher plants. In: Verma DPS
(ed) Control of plant gene expression. CRC Press, Boca Raton,
Florida, pp 325–335
Escobar C, De Meutter J, Aristizabal FA, Sanz-Alferez S, del Campo
FF, Barthels N, Van der Eycken W, Seurinck J, van Montagu M,
Gheysen G, Fenoll C (1999) Isolation of the LEMMI9 gene and
promoter analysis during a compatible plant-nematode interac-
tion. Mol Plant Microbe Interact 12:440–449
Escobar C, Barcala M, Portillo M, Almoguera C, Jordano J, Fenoll C
(2003) Induction of the Hahsp17.7G4 promoter by root-knot
nematodes: involvement of heat-shock elements in promoter
activity in giant cells. Mol Plant Microbe Interact 16:1062–1068
Fray RG, Lycett GW, Grierson D (1990) Nucleotide sequence of a
heat-shock and ripening-related cDNA from tomato. Nucleic
Acids Res 18:7148
Gheysen G, Fenoll C (2002) Gene expression in nematode feeding
sites. Annu Rev Phytopathol 40:191–219
Grundler FMW, Bockenhoff A (1997) Physiology of nematode
feeding and feeding sites. In: Fenoll C, Grundler FMW, Ohl SA
(eds) Cellular and molecular aspects of plant-nematode interac-
tions. Kluwer Academic Publishers, Dordrecht, the Netherlands,
pp 107–119
Haslbeck M, Franzmann T, Weinfurtner D, Buchner J (2005) Some
like it hot: the structure and function of small heat-shock
proteins. Nat Struct Mol Biol 12:842–846
Heckathorn SA, Downs CA, Sharkey TD, Coleman JS (1998) The
small, methionine-rich chloroplast heat-shock protein protects
photosystem II electron transport during heat stress. Plant
Physiol 116:439–444
Heungens K, Mugniery D, Van Montagu M, Gheysen G, Niebel A
(1996) A method to obtain disinfected Globodera infective
juveniles directly from cysts. Fundam Appl Nematol 19:91–93
Hong S-W, Vierling E (2001) Hsp101 is necessary for heat tolerance
but dispensable for development and germination in the absence
of stress. Plant J 27:25–35
Hussey RS, Mims CW (1991) Ultrastructure of feeding tubes formed
in giant-cells induced in plants by the root-knot nematode
Meloidogyne incognita. Protoplasma 162:99–107
Jammes F, Lecomte P, de Almeida-Engler J, Bitton F, Martin-
Magniette ML, Renou JP, Abad P, Favery B (2005) Genome-
wide expression profiling of the host response to root-knot
nematode infection in Arabidopsis. Plant J 44:447–458
Laskowska E, Wawrzynow A, Taylor A (1996) IbpA and IbpB, the
new heat-shock proteins, bind to endogenous Escherichia coliproteins aggregated intracellularly by heat shock. Biochimie
78:117–122
Lee GJ, Roseman AM, Saibil HR, Vierling E (1997) A small heat
shock protein stably binds heat-denatured model substrates and
can maintain a substrate in a folding-competent state. EMBO J
16:659–671
Lewi DM, Hopp HE, Escandon AS (2006) Sunflower (Helianthusannuus L.). Methods Mol Biol 343:291–297
Lohmann C, Eggers-Schumacher G, Wunderlich M, Schoffl F (2004)
Two different heat shock transcription factors regulate immedi-
ate early expression of stress genes in Arabidopsis. Mol Genet
Genomics 271:11–21
Low D, Brandle K, Nover L, Forreiter C (2000) Cytosolic heat-stress
proteins Hsp17.7 class I and Hsp17.3 class II of tomato act as
molecular chaperones in vivo. Planta 211:575–582
Miller G, Mittler R (2006) Could heat shock transcription factors
function as hydrogen peroxide sensors in plants? Ann Bot (Lond)
98:279–288
Mishra SK, Tripp J, Winkelhaus S, Tschiersch B, Theres K, Nover L,
Scharf K-D (2002) In the complex family of heat stress
transcription factors, HsfA1 has a unique role as master regulator
of thermotolerance in tomato. Genes Dev 16:1555–1567
Mitchum MG, Sukno S, Wang X, Shani Z, Tsabary G, Shoseyov O,
Davis EL (2004) The promoter of the Arabidopsis thaliana Cel1endo-1,4-beta glucanase gene is differentially expressed in plant
feeding cells induced by root-knot and cyst nematodes. Mol
Plant Pathol 5:175–181
Morimoto RI (1998) Regulation of the heat shock transcriptional
response: cross talk between a family of heat shock factors,
molecular chaperones, and negative regulators. Genes Dev
12:3788–3796
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue culture. Physiol Plant 15:473–497
Nover L, Scharf K-D, Gagliardi D, Vergne P, Czarnecka-Verner E,
Gurley WB (1996) The Hsf world: classification and properties
of plant heat stress transcription factors. Cell Stress Chaperones
1:215–223
Nover L, Bharti K, Doring P, Mishra SK, Ganguli A, Scharf K-D
(2001) Arabidopsis and the heat stress transcription factor world:
how many heat stress transcription factors do we need? Cell
Stress Chaperones 6:177–189
Paulson RE, Webster JM (1970) Giant cell formation in tomato roots
caused by Meloidogyne incognita and Meloidogyne hapla(Nematoda) infection. A light and electron microscope study.
Can J Bot 48:271–276
Prandl R, Kloske E, Schoffl F (1995) Developmental regulation and
tissue-specific differences of heat shock gene expression in
transgenic tobacco and Arabidopsis plants. Plant Mol Biol
28:73–82
Ramsay K, Wang Z, Jones MGK (2004) Using laser capture
microdissection to study gene expression in early stages of
giant cells induced by root-knot nematodes. Mol Plant Pathol
5:587–592
Plant Mol Biol (2008) 66:151–164 163
123

Rieping M, Schoffl F (1992) Synergistic effect of upstream
sequences, CCAAT box elements, and HSE sequences for
enhanced expression of chimaeric heat shock genes in transgenic
tobacco. Mol Gen Genet 231:226–232
Reindl A, Schoffl F (1998) Interaction between the Arabidopsisthaliana heat shock transcription factor HSF1 and the TATA
binding protein TBP. FEBS Lett 436:318–322
Reindl A, Schoffl F, Schell J, Koncz C, Bako L (1997) Phosphor-
ylation by a cyclin-dependent kinase modulates DNA binding of
the Arabidopsis heat-shock transcription factor HSF1 in vitro.
Plant Physiol 115:93–100
Rojas A, Almoguera C, Carranco R, Scharf K-D, Jordano J (2002)
Selective activation of the developmentally regulated Hahsp17.6 G1 promoter by heat stress transcription factors. Plant
Physiol 129:1207–1215
Scharf K-D, Heider H, Hohfeld I, Lyck R, Schmidt E, Nover L (1998)
The tomato Hsf system: HsfA2 needs interaction with HsfA1 for
efficient nuclear import and may be localized in cytoplasmic heat
stress granules. Mol Cell Biol 18:2240–2251
Scharf K-D, Siddique M, Vierling E (2001) The expanding family of
Arabidopsis thaliana small heat stress proteins and a new family
of proteins containing alpha-crystallin domains (Acd proteins).
Cell Stress Chaperones 6:225–237
Shingles J, Lilley CJ, Atkinson HJ, Urwin PE (2007) Meloidogyneincognita: molecular and biochemical characterisation of a
cathepsin L cysteine proteinase and the effect on parasitism
following RNAi. Exp Parasitol 115:114–120
Sun W, Van Montagu M, Verbruggen N (2002) Small heat shock
proteins and stress tolerance in plants. Biochim Biophys Acta
1577:1–9
Swindell WR, Huebner M, Weber AP (2007) Transcriptional profiling
of Arabidopsis heat shock proteins and transcription factors
reveals extensive overlap between heat and non-heat stress
response pathways. BMC Genomics 8:125
Van der Eycken W, de Almeida Engler J, Inze D, Van Montagu M,
Gheysen G (1996) A molecular study of root-knot nematode-
induced feeding sites. Plant J 9:45–54
Vanholme B, De Meutter J, Tytgat T, Van Montagu M, Coomans A,
Gheysen G (2004) Secretions of plant-parasitic nematodes: a
molecular update. Gene 332:13–27
Vercauteren I, Goeleven E, Barthels N, Van Montagu M, Gheysen G
(1998) The rha1 gene, encoding a small GTP-binding protein, is
induced in nematode infection sites. Arch Physiol Biochem
106:158
Vierling E (1991) The roles of heat shock proteins in plants. Annu
Rev Plant Physiol Plant Mol Biol 42:579–620
Volkov RA, Panchuk II, Schoffl F (2005) Small heat shock proteins
are differentially regulated during pollen development and
following heat stress in tobacco. Plant Mol Biol 57:487–502
Waters ER, Lee GJ, Vierling E (1996) Evolution, structure and
function of the small heat shock proteins in plants. J Exp Bot
47:325–338
Wehmeyer N, Vierling E (2000) The expression of small heat shock
proteins in seeds responds to discrete developmental signals and
suggests a general protective role in desiccation tolerance. Plant
Physiol 122:1099–1108
Wieczorek K, Golecki B, Gerdes L, Heinen P, Szakasits D, Durachko
DM, Cosgrove DJ, Kreil DP, Puzio PS, Bohlmann H, Grundler
FMW (2006) Expansins are involved in the formation of
nematode-induced syncytia in roots of Arabidopsis thaliana.
Plant J 48:98–112
Wunderlich M, Werr W, Schoffl F (2003) Generation of dominant-
negative effects on the heat shock response in Arabidopsisthaliana by transgenic expression of a chimaeric HSF1 protein
fusion construct. Plant J 35:442–451
Yamanouchi U, Yano M, Lin H, Ashikari M, Yamada K (2002) A rice
spotted leaf gene, Spl7, encodes a heat stress transcription factor
protein. Proc Natl Acad Sci USA 99:7530–7535
Yu JH, Kim KP, Park SM, Hong CB (2005) Biochemical analysis of a
cytosolic small heat shock protein, NtHSP18.3, from Nicotianatabacum. Mol Cells 19:328–333
164 Plant Mol Biol (2008) 66:151–164
123
Copyright © 2022 FDOKUMEN




















