
Disrupted-in-schizophrenia 1 regulates transport of ITPR1 mRNA for synaptic plasticity
12
698 VOLUME 18 | NUMBER 5 | MAY 2015 NATURE NEUROSCIENCE ARTICLES Schizophrenia is a devastating psychiatric disorder affecting 1% of the population worldwide. The gene disrupted-in-schizophrenia 1 (DISC1) is a promising susceptibility factor for schizophrenia. The DISC1 locus was originally identified at the breakpoint of a bal- anced (1;11) (q42;q14) chromosome translocation that was found to cosegregate with schizophrenia, bipolar disorder and recurrent major depression in a large Scottish family 1,2 . Further analysis indicated that inheritance of the translocation was causal and increased the risk of these psychiatric disorders by 50-fold 1 . To understand the molecular function of DISC1, several groups have identified DISC1-interacting proteins, including nuclear dis- tribution gene E homolog-like 1 (NDEL1), growth factor receptor- bound protein 2 (GRB2), glycogen synthase kinase 3-β (GSK3β), lissencephaly-1 (LIS1), kinesin family member 5B (KIF5B), fas- ciculation and elongation protein-ζ 1 (FEZ1), girdin, kalirin and phosphodiesterase 4B 3,4 . We previously reported that DISC1 is required for the transport of some key regulators of axon outgrowth, including the NDEL1–LIS1–14-3-3ε complex and GRB2 through the kinesin-1 motor complex 5,6 . These results indicate that DISC1 functions as an adaptor molecule to link cargo molecules to kinesin-1 during neurodevelopment. These efforts have provided a basis for examining the processes and pathways affected by DISC1. Studies carried out in vitro and in intact cells have shown that DISC1 is involved in neurogenesis, neuronal migration, the formation of axons and dendrites and synapse formation through inter- actions with various partners 7 . We previously generated a Disc1- knockout (Disc1 −/− ) mouse and developed a set of antibodies to the N-terminal and C-terminal regions of DISC1 (DISC1n-Ab and DISC1c-Ab, respectively) 8 . Disc1 −/− mice revealed the follow- ing problems in earlier DISC1 studies: (1) the presence of DISC1 isoforms, (2) the poor specificity of commercial antibodies to DISC1 and (3) the use of improper mouse strains as pointed out previously 9 . DISC1n-Ab and DISC1c-Ab commonly detected a protein band of 100 kDa in a wild-type brain lysate but not in the Disc1 −/− lysate. Neither antibody detected any of the DISC1 isoforms postulated in previous studies 10,11 , suggesting that the isoforms are not expressed in the brain or that the expression levels are extremely low. When we examined the specificity of the commercially available antibodies to DISC1 used in previous studies 12–14 , we found that these antibodies did not detect endogenous DISC1, presumably because of its low abundance 8 . Gogos and others identified a 25-bp deletion in exon 6 of Disc1 in inbred 129 mice 15,16 . They claim that the micro- deletion in Disc1 causes a frame shift and interferes with the production of endogenous DISC1. We confirmed that the 129 and ICR outbred mouse strains 8 , which were employed in the earlier DISC1 studies 14,17–19 , carried the same deletion and that these mouse strains did not express DISC1. In the present study, it was reasonable to use the Disc1 −/− mouse to evaluate the physiological function of DISC1. 1 Department of Cell Pharmacology, Graduate School of Medicine, Nagoya University, Nagoya, Aichi, Japan. 2 JST, CREST, Tokyo, Japan. 3 Department of Physiology, Graduate School of Medicine, Nagoya University, Nagoya, Aichi, Japan. 4 Department of Biochemistry and Cellular Biology, National Institute of Neuroscience, Kodaira, Tokyo, Japan. 5 Department of Anatomy and Cell Biology, Graduate School of Medicine, Nagoya University, Nagoya, Aichi, Japan. 6 Department of Developmental and Regenerative Biology, Graduate School of Medical Science, Nagoya City University, Nagoya, Aichi, Japan. 7 Department of Neural Regeneration and Cell Communication, Mie University Graduate School of Medicine, Tsu, Mie, Japan. 8 Department of Neuronal Cell Biology, Okazaki Institute for Integrative Bioscience and National Institute for Basic Biology, National Institutes of Natural Sciences, Okazaki, Aichi, Japan. 9 Department of Physiology, Keio University School of Medicine, Shinjuku, Tokyo, Japan. 10 Brain Science Institute, RIKEN, Wako, Saitama, Japan. Correspondence should be addressed to K. Kaibuchi ([email protected]). Received 19 December 2014; accepted 22 February 2015; published online 30 March 2015; doi:10.1038/nn.3984 Disrupted-in-schizophrenia 1 regulates transport of ITPR1 mRNA for synaptic plasticity Daisuke Tsuboi 1,2 , Keisuke Kuroda 1,2 , Motoki Tanaka 3 , Takashi Namba 1,2 , Yukihiko Iizuka 1,2 , Shinichiro Taya 1,2,4 , Tomoyasu Shinoda 1,5 , Takao Hikita 1,6 , Shinsuke Muraoka 1 , Michiro Iizuka 2,7 , Ai Nimura 1 , Akira Mizoguchi 7 , Nobuyuki Shiina 8 , Masahiro Sokabe 3 , Hideyuki Okano 9 , Katsuhiko Mikoshiba 10 & Kozo Kaibuchi 1,2 Disrupted-in-schizophrenia 1 (DISC1) is a susceptibility gene for major psychiatric disorders, including schizophrenia. DISC1 has been implicated in neurodevelopment in relation to scaffolding signal complexes. Here we used proteomic analysis to screen for DISC1 interactors and identified several RNA-binding proteins, such as hematopoietic zinc finger (HZF), that act as components of RNA-transporting granules. HZF participates in the mRNA localization of inositol-1,4,5-trisphosphate receptor type 1 (ITPR1), which plays a key role in synaptic plasticity. DISC1 colocalizes with HZF and ITPR1 mRNA in hippocampal dendrites and directly associates with neuronal mRNAs, including ITPR1 mRNA. The binding potential of DISC1 for ITPR1 mRNA is facilitated by HZF. Studies of Disc1-knockout mice have revealed that DISC1 regulates the dendritic transport of Itpr1 mRNA by directly interacting with its mRNA. The DISC1-mediated mRNA regulation is involved in synaptic plasticity. We show that DISC1 binds ITPR1 mRNA with HZF, thereby regulating its dendritic transport for synaptic plasticity. npg © 2015 Nature America, Inc. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Disrupted-in-schizophrenia 1 regulates transport of ITPR1 mRNA for synaptic plasticity

698 VOLUME 18 | NUMBER 5 | MAY 2015 nature neurOSCIenCe
a r t I C l e S
Schizophrenia is a devastating psychiatric disorder affecting 1% of the population worldwide. The gene disrupted-in-schizophrenia 1 (DISC1) is a promising susceptibility factor for schizophrenia. The DISC1 locus was originally identified at the breakpoint of a bal-anced (1;11) (q42;q14) chromosome translocation that was found to cosegregate with schizophrenia, bipolar disorder and recurrent major depression in a large Scottish family1,2. Further analysis indicated that inheritance of the translocation was causal and increased the risk of these psychiatric disorders by 50-fold1.
To understand the molecular function of DISC1, several groups have identified DISC1-interacting proteins, including nuclear dis-tribution gene E homolog-like 1 (NDEL1), growth factor receptor-bound protein 2 (GRB2), glycogen synthase kinase 3-β (GSK3β), lissencephaly-1 (LIS1), kinesin family member 5B (KIF5B), fas-ciculation and elongation protein-ζ 1 (FEZ1), girdin, kalirin and phosphodiesterase 4B3,4. We previously reported that DISC1 is required for the transport of some key regulators of axon outgrowth, including the NDEL1–LIS1–14-3-3ε complex and GRB2 through the kinesin-1 motor complex5,6. These results indicate that DISC1 functions as an adaptor molecule to link cargo molecules to kinesin-1 during neurodevelopment. These efforts have provided a basis for examining the processes and pathways affected by DISC1.
Studies carried out in vitro and in intact cells have shown that DISC1 is involved in neurogenesis, neuronal migration, the formation
of axons and dendrites and synapse formation through inter-actions with various partners7. We previously generated a Disc1- knockout (Disc1−/−) mouse and developed a set of antibodies to the N-terminal and C-terminal regions of DISC1 (DISC1n-Ab and DISC1c-Ab, respectively)8. Disc1−/− mice revealed the follow-ing problems in earlier DISC1 studies: (1) the presence of DISC1 isoforms, (2) the poor specificity of commercial antibodies to DISC1 and (3) the use of improper mouse strains as pointed out previously9. DISC1n-Ab and DISC1c-Ab commonly detected a protein band of 100 kDa in a wild-type brain lysate but not in the Disc1−/− lysate. Neither antibody detected any of the DISC1 isoforms postulated in previous studies10,11, suggesting that the isoforms are not expressed in the brain or that the expression levels are extremely low. When we examined the specificity of the commercially available antibodies to DISC1 used in previous studies12–14, we found that these antibodies did not detect endogenous DISC1, presumably because of its low abundance8. Gogos and others identified a 25-bp deletion in exon 6 of Disc1 in inbred 129 mice15,16. They claim that the micro-deletion in Disc1 causes a frame shift and interferes with the production of endogenous DISC1. We confirmed that the 129 and ICR outbred mouse strains8, which were employed in the earlier DISC1 studies14,17–19, carried the same deletion and that these mouse strains did not express DISC1. In the present study, it was reasonable to use the Disc1−/− mouse to evaluate the physiological function of DISC1.
1Department of Cell Pharmacology, Graduate School of Medicine, Nagoya University, Nagoya, Aichi, Japan. 2JST, CREST, Tokyo, Japan. 3Department of Physiology, Graduate School of Medicine, Nagoya University, Nagoya, Aichi, Japan. 4Department of Biochemistry and Cellular Biology, National Institute of Neuroscience, Kodaira, Tokyo, Japan. 5Department of Anatomy and Cell Biology, Graduate School of Medicine, Nagoya University, Nagoya, Aichi, Japan. 6Department of Developmental and Regenerative Biology, Graduate School of Medical Science, Nagoya City University, Nagoya, Aichi, Japan. 7Department of Neural Regeneration and Cell Communication, Mie University Graduate School of Medicine, Tsu, Mie, Japan. 8Department of Neuronal Cell Biology, Okazaki Institute for Integrative Bioscience and National Institute for Basic Biology, National Institutes of Natural Sciences, Okazaki, Aichi, Japan. 9Department of Physiology, Keio University School of Medicine, Shinjuku, Tokyo, Japan. 10Brain Science Institute, RIKEN, Wako, Saitama, Japan. Correspondence should be addressed to K. Kaibuchi ([email protected]).
Received 19 December 2014; accepted 22 February 2015; published online 30 March 2015; doi:10.1038/nn.3984
Disrupted-in-schizophrenia 1 regulates transport of ITPR1 mRNA for synaptic plasticityDaisuke Tsuboi1,2, Keisuke Kuroda1,2, Motoki Tanaka3, Takashi Namba1,2, Yukihiko Iizuka1,2, Shinichiro Taya1,2,4, Tomoyasu Shinoda1,5, Takao Hikita1,6, Shinsuke Muraoka1, Michiro Iizuka2,7, Ai Nimura1, Akira Mizoguchi7, Nobuyuki Shiina8, Masahiro Sokabe3, Hideyuki Okano9, Katsuhiko Mikoshiba10 & Kozo Kaibuchi1,2
Disrupted-in-schizophrenia 1 (DISC1) is a susceptibility gene for major psychiatric disorders, including schizophrenia. DISC1 has been implicated in neurodevelopment in relation to scaffolding signal complexes. Here we used proteomic analysis to screen for DISC1 interactors and identified several RNA-binding proteins, such as hematopoietic zinc finger (HZF), that act as components of RNA-transporting granules. HZF participates in the mRNA localization of inositol-1,4,5-trisphosphate receptor type 1 (ITPR1), which plays a key role in synaptic plasticity. DISC1 colocalizes with HZF and ITPR1 mRNA in hippocampal dendrites and directly associates with neuronal mRNAs, including ITPR1 mRNA. The binding potential of DISC1 for ITPR1 mRNA is facilitated by HZF. Studies of Disc1-knockout mice have revealed that DISC1 regulates the dendritic transport of Itpr1 mRNA by directly interacting with its mRNA. The DISC1-mediated mRNA regulation is involved in synaptic plasticity. We show that DISC1 binds ITPR1 mRNA with HZF, thereby regulating its dendritic transport for synaptic plasticity.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature neurOSCIenCe VOLUME 18 | NUMBER 5 | MAY 2015 699
a r t I C l e S
The Disc1−/− mouse displays no gross abnormalities in the cytoar-chitecture of the brain, but it does show altered synaptic plasticity and abnormal emotional behaviors8, suggesting that DISC1 regulates synaptic plasticity and cognitive functions.
In the present study, we used proteomic analysis to identify RNA-binding proteins, including HZF, as DISC1 interactors. HZF is a component of RNA granules, along with inositol-1,4,5-trisphosphate receptor type 1 (ITPR1, also known as IP3R1) mRNA, which acts as a key regulator for synaptic plasticity. DISC1 colocalized with HZF and Itpr1 mRNA in hippocampal dendrites. Furthermore, DISC1 directly associated with several mRNAs, including that of Itpr1. The impair-ment of DISC1 function prohibited the dendritic transport of Itpr1 mRNA and caused altered synaptic plasticity. This study describes a novel role for DISC1 and provides an intriguing clue regarding the molecular mechanism underlying synaptic plasticity.
RESULTSProteomic analysis of DISC1-interacting proteinsTo identify previously unrecognized DISC1-interacting molecules in mature neurons, we used adult rat brain lysates for DISC1 affinity chromatography. The brain cytosol fraction was loaded onto affinity columns coated with maltose-binding protein (MBP) or MBP-hDISC1. Several protein bands were detected in the eluate of the MBP-hDISC1 affinity column but not in that of the MBP-only column (the negative control) (Fig. 1a). We identified KIF1B, KIF5A, heterogeneous nuclear ribonucleoprotein U (HnRNP U), synapto-tagmin-binding cytoplasmic RNA-interacting protein (SYNCRIP), dynein cytoplasmic 1 intermediate chain 1 (DYNC1I1), HZF, LIS1, purine-rich single-stranded DNA-binding protein alpha (PURα), RACK1 and GRB2 as DISC1 interactors by mass spectrometry (Fig. 1a and Supplementary Table 1). HnRNP U, SYNCRIP, HZF, PURα and RACK1 are RNA-binding proteins20,21. SYNCRIP and HZF are thought to be components of RNA granules, RNA-protein macrocomplexes that are sites of mRNA transport, storage and trans-lation regulation in dendrites22,23.
DISC1 as a component of RNA-transporting granulesIt has been reported that SYNCRIP and HZF participate in the den-dritic localization of ITPR1 mRNA20,24. Because these observations hint at the possible involvement of DISC1 in mRNA transport, we focused on SYNCRIP and HZF. We examined the interaction of DISC1 with SYNCRIP and HZF in RNA granules. RNase A treatment has been used to verify direct interactions between RNA-binding proteins21. Rat brain lysates with or without RNase A treatment were loaded onto an affinity column (Fig. 1b). The binding of DISC1 to HnRNP U, PURα and RACK1 decreased after RNA digestion, whereas the interactions of DISC1 with KIF5A, SYNCRIP and HZF appeared to be unaffected (Fig. 1b and Supplementary Fig. 1a). To examine in vivo associations of DISC1 with SYNCRIP and HZF, we carried out an immunoprecipitation assay with the validated DISC1 antibody (DISC1c-Ab)8. When DISC1 was immunoprecipi-tated from brain lysates, KIF5A, SYNCRIP and HZF coprecipitated (Fig. 1c). In addition, KIF5A immunoprecipitated with DISC1, SYNCRIP and HZF in brain lysate (Fig. 1d). These results indicate that DISC1 closely associates with KIF5A, HZF and SYNCRIP under physiological conditions.
Because neurons are used as a model system for the analysis of RNA granules25,26, we determined the localization of DISC1 in hippo-campal neurons using DISC1c-Ab8. As described previously, DISC1 was distributed in punctate structures in microtubule-associated protein 2 (MAP2)-positive dendrites of pyramidal neurons in the
hippocampal CA1 region, as well as in cultured hippocampal neurons (Supplementary Fig. 2)8. To analyze the subcellular distribution of DISC1 in detail, we used immunoelectron microscopy to examine dendrites of hippocampal CA1 pyramidal cells. DISC1 immuno-reactivity was detected in the bases of dendritic spines and along microtubules (Supplementary Fig. 2j,k). DISC1 puncta appeared as large (>1 µm in diameter) and small (<1 µm in diameter) granules in dendrites (Fig. 1e–g and Supplementary Fig. 2b–i). The larger granules were uniformly distributed in somatodendrites, whereas the smaller granules occurred in dendrites. Both types of DISC1 gran-ule colocalized with SYTO14-visualized RNA and rat Staufen-GFP, a reliable marker for RNA granules (Fig. 1e and Supplementary Fig. 2c)25,26. The larger granules primarily colocalized with argonaute 2 (AGO2) (Supplementary Fig. 2i)23. The smaller granules partly colocalized with HZF, SYNCRIP and fragile X mental retardation protein (FMRP), but not with eukaryotic translation initiation factor 4G (eIF4G) (Fig. 1f and Supplementary Fig. 2d–f). To quantify the relative colocalization of DISC1 with several endogenous components of RNA granules, we calculated the Pearson’s correlation coefficient (Supplementary Fig. 1b). The correlation coefficients for DISC1 with HZF, SYNCRIP, FMRP and AGO2 were higher than that with eIF4G, an unrelated component of RNA granules (Supplementary Fig. 1b). These results suggest that DISC1 is a component of neuronal RNA granules and localizes to a subpopulation of the RNA granules.
It has been shown that HZF interacts with the 3′ untranslated region (UTR) of ITPR1 mRNA. The 3′ UTR of ITPR1 mRNA is responsible for both its dendritic localization and its binding to HZF in hippo-campal neurons20,24. To visualize the localization pattern conferred by the 3′ UTR of ITPR1 mRNA, we used a modified MS2-GFP tagging system20,27,28. Hippocampal neurons were transfected with MS2bs–ITPR1 3′ UTR and MS2-nls-Venus. ITPR1 3′ UTR RNA visualized using Venus (Venus–ITPR1 3′ UTR) colocalized with DISC1 in the dendrites (Supplementary Fig. 2g). To further examine whether endogenous ITPR1 mRNA colocalized with DISC1, we carried out combined immunostaining and fluorescent in situ hybridization (FISH) analysis. DISC1 partly colocalized with the FISH signals corre-sponding to ITPR1 mRNA (Fig. 1g). These results suggest that ITPR1 mRNA forms a complex with DISC1 in hippocampal dendrites.
RNA granules are transported via the kinesin-1 motor21, which functions as a key component of RNA-transporting granules to localize various messenger ribonucleoprotein complexes in dendrites29. Kinesin-1 is a tetramer consisting of three kinesin heavy chains (KIF5A, KIF5B and KIF5C) and two kinesin light chains (KLC1 and KLC2). Because DISC1 directly interacts with KIF5A and links NDEL1 and GRB2 to kinesin-1, thereby promoting their transport into axons5,6, we hypothesized that DISC1 regulates the localization of dendritic mRNAs via a kinesin-1–mediated transport mechanism. To visualize DISC1 transport in real time, we monitored live neu-rons expressing human DISC1-GFP fusion protein (hDISC1-GFP). Granules containing hDISC1-GFP were observed in both axons and dendrites. The colocalization of DISC1 and Venus–ITPR1 3′ UTR appeared to be most prominent in dendrites (Supplementary Fig. 2g). There were two different size classes of hDISC1-GFP granules, small and large (Fig. 2), similar to what was observed in the fixed cells for the DISC1 puncta. The smaller granules colocalized with HZF (Supplementary Fig. 2h) and often moved bidirectionally in the dendrites of cultured neurons, whereas the larger granules localized mainly in somatodendrites and appeared to be immobile. These obser-vations raised the possibility that the smaller granules function as RNA-transporting granules for the dendritic localization of ITPR1 mRNA. Thus, we focused on the kinetics of the smaller hDISC1-GFP
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

700 VOLUME 18 | NUMBER 5 | MAY 2015 nature neurOSCIenCe
a r t I C l e S
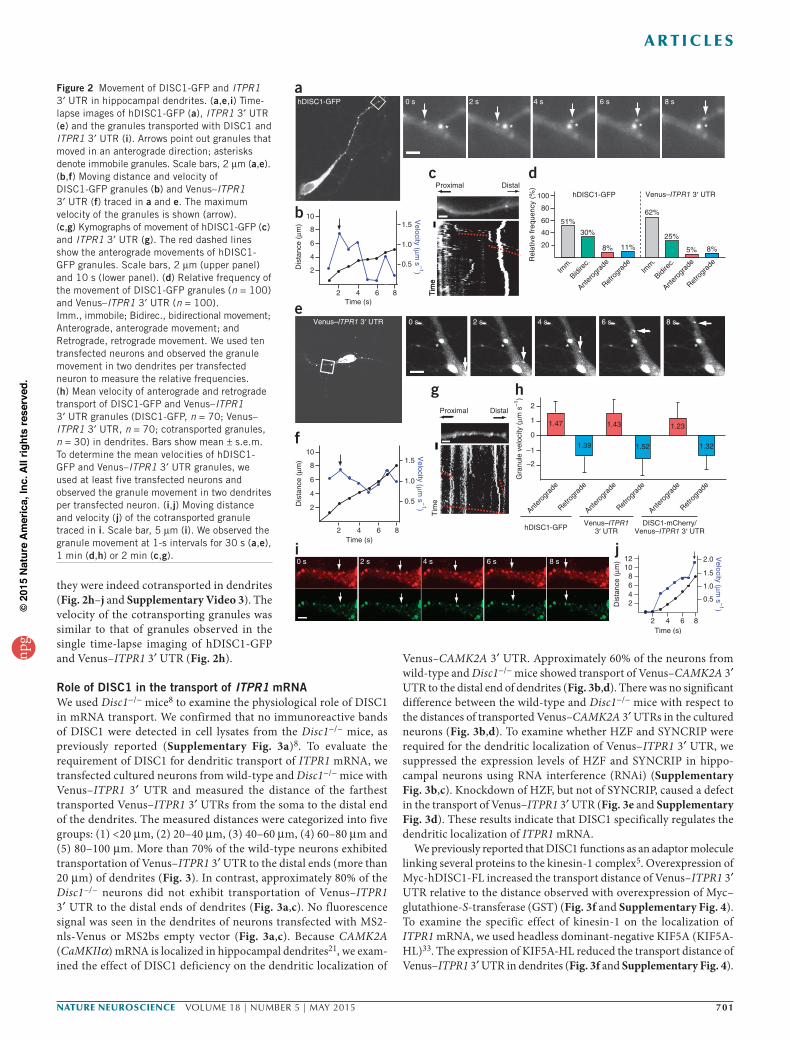
granules in dendrites (Fig. 2a–d). We categorized the movement patterns of hDISC1-GFP granules as follows: (1) immobile, (2) bidi-rectional and (3) anterograde or retrograde. Imaging of these granules revealed that approximately half of the observed granules were mobile, and the remainder were immobile (Fig. 2d). The mobile granules showed both bi- and unidirectional movement (Fig. 2a–c and Supplementary Video 1). Bidirectionally mobile granules were observed more frequently than anterograde- and retrograde-only granules (Fig. 2d). Single-granule tracking studies revealed that hDISC1-GFP granules displayed variable mobility (Fig. 2b). The mean transport rates for anterograde and retrograde hDISC1-GFP granules were 1.47 ± 0.8 µm s−1 and 1.39 ± 0.9 µm s−1, respectively (Fig. 2h). As it has been reported that the velocities of kinesin and dynein motors and their cargoes are around 1 µm s−1 (refs. 29–32), the kinetic study with hDISC1-GFP raises the possibility that DISC1 may attach to functional RNA-transporting granules through kinesin-mediated
and/or dynein-mediated linkage. We next examined the dynamics of Venus–ITPR1 3′ UTR in dendrites using time-lapse imaging. When we examined the motion pattern of the Venus–ITPR1 3′ UTR, we noted bidirectionally mobile granules more frequently than we did unidirectional granules (Fig. 2d), although some Venus–ITPR1 3′ UTR granules were observed to move unidirectionally (Fig. 2d–g and Supplementary Video 2). The mean velocities of Venus–ITPR1 3′ UTR granules moving in the anterograde and retrograde directions were 1.43 ± 0.9 µm s−1 and 1.52 ± 1.2 µm s−1, respectively (Fig. 2h), values similar to those determined for hDISC1-GFP granules. The bidirectional movement of hDISC1-GFP and Venus–ITPR1 3′ UTR granules was consistent with the kinesin/dynein-mediated tug-of-war movement model29. To examine whether DISC1 was cotransported with ITPR1 3′ UTR to the distal end of dendrites, we used dual-color time-lapse imaging with hDISC1-mCherry and Venus–ITPR1 3′ UTR. DISC1-mCherry colocalized with Venus–ITPR1 3′ UTR, and
Merge
DISC1SYTO14
*
HZF
Merge
DISC1
*
Antisense probe (ITPR1)
**
Sense probe (ITPR1)
DISC1
Merge
150
100
75
50
37
(kDa)
a
e f g
b c d
*
*
**
*
KIF1B
KIF5A,HnRNP-U
MBP
MBP-h
DISC1
*
*
SYNCRIPDYNC1I1
LIS1
RACK1
GRB2
PURα, HZF
(kDa)
37
50
150
RNase A
MBP
MBP-h
DISC1
MBP
MBP-h
DISC1
10075
37
KIF5A
SYNCRIP
PURα
RACK1
HnRNP-U
HZF
Elution
+ – + – + – + –
Input
DISC1
KIF5A
SYNCRIP
HZF
GAPDH
150Con
trol Ig
G
DISC1c
-Ab
Inpu
t
75
50
37
100
(kDa)Con
trol Ig
G
Anti-K
IF5
Inpu
t
DISC1
KIF5A
HZF
100
(kDa)
SYNCRIP75
50
150
Figure 1 Identification of RNA-binding proteins as novel DISC1 interactors. (a) Representative silver staining of the affinity-column eluates. Proteins that bound to affinity columns coated with MBP or MBP-hDISC1 were subjected to SDS-PAGE. The arrows and asterisks indicate DISC1 interactors and full-length MBP-DISC1 and its degradation products, respectively. (b) Immunoblot analysis of the affinity-column eluates. Brain lysates treated with or without RNase A were used in DISC1 affinity chromatography. Aliquots of cytosolic extracts (1% input) and eluates (10%) were subjected to SDS-PAGE and then immunoblotted with the indicated antibodies. (c,d) Immunoprecipitation assay with DISC1c-Ab (c) and anti-KIF5 (d) from mouse brain lysate. The precipitated proteins and inputs were immunoblotted with the indicated antibodies. Aliquots of original samples (1% input) and eluates (10%) were subjected to SDS-PAGE. GAPDH was used as a negative control. (e–g) Coimmunostainings of DISC1 with SYTO14 (e), HZF (f) and ITPR1 mRNA (g) in cultured rat hippocampal neurons at DIV 9. The Cy5-labeled DISC1 image in e is in pseudocolor (red). Scale bars in main images, 20 µm. Higher-magnification images of the boxed areas are shown in the lower right panels, in which asterisks and arrowheads indicate larger and smaller granules in dendrites, respectively. Scale bars in the magnified images represent 5 µm. Full-length blots are shown in Supplementary Figure 14.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.
nature neurOSCIenCe VOLUME 18 | NUMBER 5 | MAY 2015 701
a r t I C l e S
they were indeed cotransported in dendrites (Fig. 2h–j and Supplementary Video 3). The velocity of the cotransporting granules was similar to that of granules observed in the single time-lapse imaging of hDISC1-GFP and Venus–ITPR1 3′ UTR (Fig. 2h).
Role of DISC1 in the transport of ITPR1 mRNAWe used Disc1−/− mice8 to examine the physiological role of DISC1 in mRNA transport. We confirmed that no immunoreactive bands of DISC1 were detected in cell lysates from the Disc1−/− mice, as previously reported (Supplementary Fig. 3a)8. To evaluate the requirement of DISC1 for dendritic transport of ITPR1 mRNA, we transfected cultured neurons from wild-type and Disc1−/− mice with Venus–ITPR1 3′ UTR and measured the distance of the farthest transported Venus–ITPR1 3′ UTRs from the soma to the distal end of the dendrites. The measured distances were categorized into five groups: (1) <20 µm, (2) 20–40 µm, (3) 40–60 µm, (4) 60–80 µm and (5) 80–100 µm. More than 70% of the wild-type neurons exhibited transportation of Venus–ITPR1 3′ UTR to the distal ends (more than 20 µm) of dendrites (Fig. 3). In contrast, approximately 80% of the Disc1−/− neurons did not exhibit transportation of Venus–ITPR1 3′ UTR to the distal ends of dendrites (Fig. 3a,c). No fluorescence signal was seen in the dendrites of neurons transfected with MS2-nls-Venus or MS2bs empty vector (Fig. 3a,c). Because CAMK2A (CaMKIIα) mRNA is localized in hippocampal dendrites21, we exam-ined the effect of DISC1 deficiency on the dendritic localization of
Venus–CAMK2A 3′ UTR. Approximately 60% of the neurons from wild-type and Disc1−/− mice showed transport of Venus–CAMK2A 3′ UTR to the distal end of dendrites (Fig. 3b,d). There was no significant difference between the wild-type and Disc1−/− mice with respect to the distances of transported Venus–CAMK2A 3′ UTRs in the cultured neurons (Fig. 3b,d). To examine whether HZF and SYNCRIP were required for the dendritic localization of Venus–ITPR1 3′ UTR, we suppressed the expression levels of HZF and SYNCRIP in hippo-campal neurons using RNA interference (RNAi) (Supplementary Fig. 3b,c). Knockdown of HZF, but not of SYNCRIP, caused a defect in the transport of Venus–ITPR1 3′ UTR (Fig. 3e and Supplementary Fig. 3d). These results indicate that DISC1 specifically regulates the dendritic localization of ITPR1 mRNA.
We previously reported that DISC1 functions as an adaptor molecule linking several proteins to the kinesin-1 complex5. Overexpression of Myc-hDISC1-FL increased the transport distance of Venus–ITPR1 3′ UTR relative to the distance observed with overexpression of Myc–glutathione-S-transferase (GST) (Fig. 3f and Supplementary Fig. 4). To examine the specific effect of kinesin-1 on the localization of ITPR1 mRNA, we used headless dominant-negative KIF5A (KIF5A-HL)33. The expression of KIF5A-HL reduced the transport distance of Venus–ITPR1 3′ UTR in dendrites (Fig. 3f and Supplementary Fig. 4).
Venus–ITPR1 3′ UTR 0 s 2 s 4 s 6 s 8 s
Tim
e
Venus–ITPR13′ UTR
DISC1-mCherry/Venus–ITPR1 3′ UTR
Time (s)
* * * **
hDISC1-GFP
* * * * *
2468
10
1.0
0.5
1.5
2.012
Dis
tanc
e (µ
m)
2 6 84
Velocity (µm
s–1)
Proximal Distal
–1
–2
1
2
0
1.47
1.39
Anter
ogra
de
Gra
nule
vel
ocity
(µm
s–1
)
1.43
1.52
hDISC1-GFP
1.32
2
4
6
8
10
2 6
1.0
0.5
1.5
84
Dis
tanc
e (µ
m)
Velocity (µ m
s–1)
80
100
40
60
20
51%
30%
11%8%
Rel
ativ
e fr
eque
ncy
(%)
62%
25%
8%5%
hDISC1-GFP
Tim
e
Proximal
a
b
e
f
g
i j
h
c dDistal
2
4
6
8
10
2 6
1.0
0.5
1.5
84D
ista
nce
(µm
)
Velocity (µm
s–1)
Retro
grad
e
Anter
ogra
de
Retro
grad
e
Anter
ogra
de
Retro
grad
e
0 s 2 s 4 s 6 s 8 s
Venus–ITPR1 3′ UTR
Tim
e
Time (s)
Time (s)
0 s 2 s 4 s 6 s 8 s
Imm
.
Bidire
c.
Anter
ogra
de
Retro
grad
eIm
m.
Bidire
c.
Anter
ogra
de
Retro
grad
e
1.23
Figure 2 Movement of DISC1-GFP and ITPR1 3′ UTR in hippocampal dendrites. (a,e,i) Time-lapse images of hDISC1-GFP (a), ITPR1 3′ UTR (e) and the granules transported with DISC1 and ITPR1 3′ UTR (i). Arrows point out granules that moved in an anterograde direction; asterisks denote immobile granules. Scale bars, 2 µm (a,e). (b,f) Moving distance and velocity of DISC1-GFP granules (b) and Venus–ITPR1 3′ UTR (f) traced in a and e. The maximum velocity of the granules is shown (arrow). (c,g) Kymographs of movement of hDISC1-GFP (c) and ITPR1 3′ UTR (g). The red dashed lines show the anterograde movements of hDISC1-GFP granules. Scale bars, 2 µm (upper panel) and 10 s (lower panel). (d) Relative frequency of the movement of DISC1-GFP granules (n = 100) and Venus–ITPR1 3′ UTR (n = 100). Imm., immobile; Bidirec., bidirectional movement; Anterograde, anterograde movement; and Retrograde, retrograde movement. We used ten transfected neurons and observed the granule movement in two dendrites per transfected neuron to measure the relative frequencies. (h) Mean velocity of anterograde and retrograde transport of DISC1-GFP and Venus–ITPR1 3′ UTR granules (DISC1-GFP, n = 70; Venus–ITPR1 3′ UTR, n = 70; cotransported granules, n = 30) in dendrites. Bars show mean ± s.e.m. To determine the mean velocities of hDISC1-GFP and Venus–ITPR1 3′ UTR granules, we used at least five transfected neurons and observed the granule movement in two dendrites per transfected neuron. (i,j) Moving distance and velocity (j) of the cotransported granule traced in i. Scale bar, 5 µm (i). We observed the granule movement at 1-s intervals for 30 s (a,e), 1 min (d,h) or 2 min (c,g).
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

702 VOLUME 18 | NUMBER 5 | MAY 2015 nature neurOSCIenCe
a r t I C l e S
Taken together, these results support the idea that DISC1 regulates the transport of ITPR1 mRNA with kinesin-1.
To explore how DISC1 regulates the transport of ITPR1 mRNA, we examined the ability of DISC1 to bind ITPR1 3′ UTR RNA in UV cross-linking assays; these assays were performed because HZF has been shown to interact with the ITPR1 3′ UTR24. Recombinant GST or GST-hDISC1 protein was mixed with biotin-labeled ITPR1 3′ UTR RNA. The protein-RNA complex was then cross-linked by UV irradiation and detected by immunoblot analysis with streptavidin–horseradish peroxidase (HRP). GST-hDISC1, unlike GST and GST-GRB2, bound to the 3′ UTR of ITPR1 mRNA in vitro (Fig. 4a). The binding of GST-hDISC1 to the ITPR1 3′ UTR RNA could be competed away with unlabeled ITPR1 3′ UTR RNA (Fig. 4a). GST-hDISC1 also bound to a gfp reporter mRNA containing ITPR1 3′ UTR, but not to a gfp-coding region alone (Fig. 4a). In addition, we performed a gel-shift assay to confirm the interaction between DISC1 and ITPR1 3′ UTR RNA. GST-hDISC1-FL, but not GST or GST-hDISC1-CH, was gel-shifted with ITPR1 3′ UTR (Supplementary Fig. 5a). The dissociation constant of DISC1 for ITPR1 3′ UTR was 160 nM (Supplementary Fig. 5b,c). To identify the DISC1-binding region in ITPR1 mRNA, we repeated the RNA-binding assay with various fragments of ITPR1 3′ UTR RNA. DISC1 bound to the F4 fragment (bases 310–462) of ITPR1 3′ UTR (Fig. 4b,c). To determine the RNA-binding region in DISC1, we con-ducted UV cross-linking assays with various DISC1 fragments. ITPR1 3′ UTR RNA interacted with MBP-hDISC1-FL (amino acids (aa) 1–832), DISC1-NH (aa 1–361) and DISC1-N1 (aa 1–120) (Fig. 4d,e), indicating that the N1 region contained a site responsible for binding to ITPR1 3′ UTR. DISC1-N1 has protein motifs that contain positively charged residues. It has been reported that this cluster of the charged residues has multiple functions, including nuclear localization and
binding to both proteins and nucleic acids34. To examine whether the clusters of positively charged amino acids are required for binding to RNA, we prepared DISC1-∆ARM and -∆HRM mutants that lacked the arginine-rich motif (ARM) and the histidine-rich motif (HRM), respectively. DISC1-∆HRM bound to ITPR1 3′ UTR, whereas DISC1-∆ARM and DISC1-N1–∆ARM did not (Fig. 4d,e). These results indicate that the ARM in DISC1 is responsible for binding to RNA.
A few hundred mRNAs, including ARC and CAMK2A, are trans-ported in RNA granules to the dendritic spine35. To identify neuronal mRNAs that associate with DISC1, we used a modified RNA immunoprecipitation assay and quantitative PCR (RIP-qPCR). Briefly, DISC1-mRNA complexes were isolated from mouse brain extracts by immunoprecipitation with DISC1c-Ab. The copre-cipitated mRNAs were subjected to qPCR with the gene-specific primers. Among the 40 genes tested in the qPCR assay, mRNAs encoding the neuronal Rho guanine nucleotide exchange fac-tor kalirin or the subunits of voltage-gated ion channels such as KCNC1, KCNC4, CACNA1C and CACNA2D1 were twofold enriched in the DISC1 immunoprecipitate compared with control immunoglobulin G immunoprecipitate (Supplementary Table 2 and Supplementary Fig. 6a). To validate the direct interactions between DISC1 and the identified mRNAs, we carried out in vitro RNA binding assays. GST-hDISC1-N1 bound to the biotin-labeled 3′ UTR RNA from CACNA1C, CACNA2D1, ITPR1, KALRN, KCNC1 and KCNC4, whereas GST alone did not bind (Supplementary Fig. 6b). GST-hDISC1-N1 did not bind to the 3′ UTR of SCN2A or the CACNA2D1 coding sequence. Finally, to confirm the in vivo asso-ciation of DISC1 with the identified mRNA, we conducted RNA immunoprecipitation assays of hippocampal tissue lysates (Fig. 4f). DISC1c-Ab immunoprecipitated with CACNA1C, CACNA2D1, ITPR1,
ITPR1 3′ UTR
Disc1–/–Wild type
ITPR1 3′ UTR No UTR
Wild type
Merge Merge Merge
Ven
usV
enus
/MA
P2
Wild type
CAMK2A 3′ UTR CAMK2A 3′ UTR
Ven
usV
enus
/MA
P2
MergeMerge
Disc1–/–
20
40
60
80
100R
elat
ive
freq
uenc
y (%
)
Wild
type
Wild
type
Disc1–/
–
Venus–ITPR13′ UTR
NoUTR
: 0–20 µm : 20–40 µm : 40–60 µm : 60–80 µm : 80–100 µm
20
40
60
80
100
Rel
ativ
e fr
eque
ncy
(%)
Wild
type
Disc1–/
–
Venus–CAMK2A3′ UTR
Scram
ble si
RNA
HZF siRNA
SYNCRIP si
RNA
Rel
ativ
e fr
eque
ncy
(%)
20
40
60
80
100
Venus–ITPR13′ UTR
Myc
-GST
Myc
-hDIS
C1-FL
Myc
-KIF
5A-H
L
20
40
60
80
100
Rel
ativ
e fr
eque
ncy
(%)
Venus–ITPR13′ UTR
a
c d e f
bFigure 3 Inhibition of dendritic localization of ITPR1 3′ UTR by the loss of DISC1. (a,b) Effect of DISC1 depletion on the dendritic localization of ITPR1 3′ UTR (a) and CAMK2A 3′ UTR (b). Cultured neurons from wild-type and Disc1−/− mice were transfected with the indicated plasmids at DIV 12. The transfected neurons were fixed at DIV 14 and immunostained with antibodies to Venus (green) and MAP2 (red). Blue bars indicate the distance between the soma and the farthest transported granules containing the ITPR1 3′ UTR. Magnified images of these granules are shown in the outlined areas. Scale bars, 15 µm. (c–f) Relative frequency of the transport distance of ITPR1 3′ UTR–containing granules (c,e,f) or CAMK2A 3′ UTR–containing granules (d) in the DNA-transfected neurons. The transport distances of ITPR1 3′ UTR and CAMK2A 3′ UTR granules were categorized into five groups. The percentage of granules transported each distance is shown in stacked bar charts. The data in c–f are from three independent experiments for each treatment, with at least 25 neurons per experiment (n = 75 for each transfectant). Significant differences between transport distances were determined by one-way analysis of variance (ANOVA) with Bonferroni post hoc test (wild type versus Disc1−/− or no UTR in c, P < 0.01, F(2, 222) = 88.85; wild type versus Disc1−/− in d, no significant results, F(1, 148) = 0.3490; scramble short interfering RNA (siRNA) versus HZF siRNA in e, P < 0.01, F(2, 222) = 33.27; Myc-GST versus Myc-hDISC1-FL or Myc-KIF5A-HL in f, P < 0.01, F(2, 222) = 62.07). Bars show mean ± s.e.m.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature neurOSCIenCe VOLUME 18 | NUMBER 5 | MAY 2015 703
a r t I C l e S
KALRN, KCNC1 and KCNC4 mRNAs, but not with phosphoglycerate mutase (PGAM1) (Fig. 4f). These results indicate that DISC1 binds to several mRNAs encoding synaptic regulators.
Regulatory mechanism of DISC1-mediated mRNA transportWe addressed the mechanism underlying mRNA transport by DISC1. First, to clarify the requirement of the ARM in the DISC1 protein for dendritic transport of ITPR1 3′ UTR, we performed a rescue experi-ment in Disc1−/− neurons using the DISC1-∆ARM mutant, which is defective in RNA binding (Fig. 4e). In contrast to that of DISC1-FL, the expression of DISC1-∆ARM did not rescue the transport defect of ITPR1 3′ UTR in the Disc1−/− neurons (Fig. 5a,b). We confirmed that DISC1-∆ARM interacted with several DISC1 interactors such as HZF, KIF5A, GSK3β, LIS1 and PDE4B (Supplementary Fig. 7a). These results indicate that the arginine-rich motif in DISC1 is a region responsible for the dendritic transport of ITPR1 3′ UTR. To investigate the significance of the interaction between DISC1 and KIF5A for the transport of ITPR1 3′ UTR, we next performed a rescue experiment in Disc1−/− neurons using DISC1-∆KBR, which is defective in KIF5 binding but not in binding to HZF and ITPR1 3′ UTR (Supplementary Fig. 7a,b). In contrast to that of DISC1-FL, expression of DISC1-∆KBR did not rescue the transport defect of ITPR1 3′ UTR (Supplementary Fig. 8a,b). Although we cannot exclude the possibility that the
DISC1-∆KBR mutant lost its ability to bind to other DISC1 partners, the rescue experiment suggests that an interaction between DISC1 and KIF5 is involved in the dendritic transport of ITPR1 3′ UTR.
We next examined the functional significance of HZF in DISC1-mediated mRNA transport. Histidine-tagged HZF bound directly to MBP-hDISC1 in vitro (Fig. 6a). Because DISC1 bound to the F4 fragment of ITPR1 3′ UTR that is distinct from the HZF-bind-ing region (Fig. 4b,c)24, we evaluated whether HZF influences the RNA-binding potential of DISC1 in an RNA pulldown assay. Myc-hDISC1-FL coprecipitated moderately when biotin–ITPR1 3′ UTR RNA was precipitated with streptavidin beads in COS7 cells expressing Myc-hDISC1-FL (Fig. 6b). Interestingly, the amounts of copre-cipitated DISC1 increased in an HZF-dependent manner (Fig. 6b). DISC1 and HZF did not precipitate without biotin–ITPR1 3′ UTR in this assay (Fig. 6b). These results suggest that the binding affinity of DISC1 for ITPR1 mRNA is increased by HZF. To examine whether the interaction between DISC1 and HZF is required for ITPR1 mRNA transport, we determined the binding region between DISC1 and HZF. HZF interacted with DISC1-FL, DISC1-NH, DISC1-N1 and DISC1-N1-∆ARM but not with DISC1-CH and DISC1-N23 in COS7 cells (Fig. 6c and Supplementary Fig. 9a). We confirmed that DISC1-N1-∆ARM had binding activity for HZF but not for RNA, KIF5A, GSK3β or LIS1 (Fig. 4e and Supplementary Fig. 7a). The DISC1-N1-∆ARM
Figure 4 Interaction of DISC1 with ITPR1 3′ UTR RNA. (a) In vitro binding between DISC1 and ITPR1 3′ UTR RNA. Recombinant GST, GST-GRB2 or GST-hDISC1 protein was incubated with biotin-labeled ITPR1 3′ UTR, GFP, GFP–ITPR1 3′ UTR, or ITPR1 3′ UTR and excess (ex.) unlabeled 3′ UTR. After UV cross-linking and digestion of the unbound RNA, RNA-protein complexes were detected by immunoblotting with streptavidin-HRP. The asterisks indicate the degradation products of GST-hDISC1 recombinant protein. (b) An illustration of the various ITPR1 3′ UTR subfragments. (c) Binding of ITPR1 3′ UTR and its subfragments to DISC1. GST or GST-hDISC1 protein was incubated with the ITPR1 3′ UTR fragments shown in b and UV cross-linked. The cross-linked proteins were then subjected to SDS-PAGE followed by immunoblotting with streptavidin-HRP. (d) An illustration of the deletion fragments of DISC1. The purple and pink boxes represent globular and coiled-coil domains, respectively. (e) Binding of DISC1 and its subfragments to ITPR1 3′ UTR. The full-length DISC1 and the DISC1 mutants shown in d were cross-linked with ITPR1 3′ UTR and subjected to SDS-PAGE followed by immunoblotting with streptavidin-HRP. (f) Coprecipitation of ITPR1 mRNA with DISC1. Mouse hippocampal extracts were subjected to immunoprecipitation (IP) with DISC1c-Ab or control rabbit IgG. The coprecipitated RNAs were purified from the immunoprecipitates and were subjected to RT-PCR for ITPR1, KALRN, CACNA2D1, CACNA1C, KCNC1, KCNC4 and PGAM1 mRNAs. RT, reverse transcriptase. Full-length blots are shown in Supplementary Figure 14.
e
MBP
MBP-h
DISC1-
FL
FL, ∆HRM
NH
N1
MBP-h
DISC1-
NH
MBP-h
DISC1-
CH
MBP-h
DISC1-
N1
MBP-h
DISC1-
N1-∆A
RM
MBP-h
DISC1-
N23
MBP-h
DISC1-
∆ARM
MBP-h
DISC1-
∆HRM
(Streptavidin-H
RP
)
150
100
75
50
37
(kDa)
d
FL
NH
CH
N1
N23
∆ARM
1 832
1 361
362 832
1 120
121 361
8321
hDISC1
8321
35–42
77–84∆HRM
1 120N1-∆ARM
35–42
f
c
GST-hDISC1GST
15010075
5037
25
GST-hDISC1
GST
(kDa)
ITPR1 3′ UTR-
a
ITPR1 3′ UTRUnlabeled 3′ UTR
gfpgfp 3′ UTR
GST-hDISC1GST–Protein
GST-hDISC1
GST
(kDa)
10075
150
37
25
50
GST-GRB2
5 ex. 20 ex. 100 ex.
GST-hDISC1
**
+–––
+–––
––––
+–––
+
––
+
––
+
––
+–––
––+–
–––+
b
UGA PolyA signalAAUAAA
FL
F1
F2
F3
F4
+462+1
1 462
1 200
201 462
201 309
310 462
ITPR1 3′ UTR
FL F1 F2 F3 F4 FL F1 F2 F3 F4
IP:
KCNC1mRNA
DISC1100(kDa)
Inpu
t
PGAM1mRNA
RT
IP: Control IgG DISC1c-Ab
CACNA1CmRNA
KCNC4mRNA
ITPR1mRNAKALRNmRNACACNA2D1mRNA
– + – +
DISC1c
-Ab
Contro
l IgG
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

704 VOLUME 18 | NUMBER 5 | MAY 2015 nature neurOSCIenCe
a r t I C l e S
fragment can be used as a dominant-negative fragment that affects the interaction between DISC1 and HZF, although we cannot exclude the effect of DISC1-N1-∆ARM on the interaction of endogenous DISC1
with other DISC1 interactors such as PDE4B. DISC1-N1-∆ARM inhibited the interaction between GFP-hDISC1-FL and His-HZF in a dose-dependent manner (Supplementary Fig. 10a). The expression
Figure 5 Requirement of interaction between DISC1 and RNA for dendritic transport of ITPR1 3′ UTR. (a) Rescue of ITPR1 3′ UTR transport defect in Disc1−/− neurons by wild-type and mutant DISC1. Cultured neurons from wild-type and Disc1−/− mice were transfected with the indicated plasmids at DIV 12. The transfected neurons were fixed at DIV 14 and immunostained with antibodies to Venus (green) and MAP2 (red). The blue bars indicate the distance between the soma and the farthest transported granules containing ITPR1 3′ UTR. Magnified images of these granules are shown in the outlined areas. Scale bars, 15 µm. (b) Relative frequency of the transport distance of ITPR1 3′ UTR–containing granules. The transport distances of ITPR1 3′ UTR granules were categorized into five groups. The data in b are from three independent experiments for each treatment, with at least 25 neurons per experiment (n = 75). Significant differences between transport distances were determined by one-way ANOVA with Bonferroni post hoc test (Disc1−/−–Myc-GST versus Disc1−/−–Myc-hDISC1-FL, P < 0.01, F(2, 222) = 60.11; Disc1−/−–Myc-GST versus Disc1−/−–Myc-hDISC1-∆ARM, not significant). Bars show mean ± s.e.m.
a50
(kDa)
His-HZF
MBP-hDISC1-FL
MBP
150
100
50
His-HZF
MBPMBP-hDISC1
+––
–++
hDISC1
FL
NH
CH
HZFbinding
1 832 aa
832
832
361
362
+
c
N1
361
120
121N23
361
N1-∆ARM120
+
–
+
+
–
b
Myc-hDISC1-FL
FLAG-HZF
Myc-hDISC1-FL
FLAG-HZF
Pulldow
nInput
SA beads onlySA beads +
biotin–ITPR1 3′ UTR(kDa)
100
100
50
50
– –FLAG-HZF
Myc-hDISC1-FL(1 µg)
Myc-hDISC1-FL(1 µg)
1 μg 5 µg 1 µg 5 µg5 µg 5 µg
DISC1binding
1
HZF
HZF (11-351)
∆Zn1
∆Zn2,3
∆Zn3
Zn1 Zn2 Zn3
+
+
–
–
386 aa
351
351
11
11
11
164
144
240
e
Myc-HZF
Rel
ativ
e fr
eque
ncy
(%)
20
40
60
80
100f Venus–ITPR1 3′ UTR
: 0–20 µm: 20–40 µm: 40–60 µm: 60–80 µm: 80–100 µm
d
20
40
60
80
100
Myc-hDISC1
Rel
ativ
e fr
eque
ncy
(%)
Venus–ITPR1 3′ UTR
: 0–20 µm: 20–40 µm: 40–60 µm: 60–80 µm: 80–100 µm
Myc
-GST N1
CH
N1-∆ARM
Myc
-GST
∆Zn3
∆Zn2,3FL
Figure 6 Requirement of interaction between DISC1 and HZF for dendritic transport of ITPR1 3′ UTR. (a) Direct binding between DISC1 and HZF. An in vitro binding assay was performed using purified recombinant proteins. The bound proteins were analyzed using immunoblotting with anti-His and anti-MBP antibodies. (b) RNA pulldown assay. Streptavidin (SA) beads alone or SA beads preincubated with biotinylated ITPR1 3′ UTR were mixed with cell lysates expressing the indicated proteins. DISC1 and HZF bound to SA beads was detected by immunoblot analysis. (c) Identification of the HZF-binding region in DISC1. The red bars indicate the DISC1 regions responsible for binding to HZF. Primary structure of DISC1: blue box, globular domain; pink boxes, coiled-coil motifs. (d) Relative frequency of the transport distance of ITPR1 3′ UTR–containing granules in DNA-transfected neurons. The data in d are from three independent experiments for each treatment, with at least 25 neurons per experiment (n = 75). Significant differences between transport distances were determined by one-way ANOVA with Bonferroni post hoc test (Myc-GST versus hDISC1-N1 and hDISC1-N1-∆ARM, P < 0.01, F(3, 296) = 16.88). Bars show mean ± s.e.m. (e) Determination of the DISC1-binding region in HZF. The red bars indicate the HZF regions responsible for binding to DISC1. Primary structure of HZF: purple boxes, zinc finger motifs. (f) Relative frequency of the transport distance of ITPR1 3′ UTR–containing granules in DNA-transfected neurons. The data in f are from three independent experiments for each treatment, with at least 25 neurons per experiment (n = 75). Significant differences between transport distances were determined by one-way ANOVA with Bonferroni post hoc test (Myc-GST versus Myc-HZF-∆Zn3, P < 0.01, F(3, 296) = 16.28). Bars show mean ± s.e.m. Full-length blots are shown in Supplementary Figures 14 and 15.
aITPR1 3′ UTRMyc-hDISC1-∆ARM
ITPR1 3′ UTRMyc-hDISC1-FL
ITPR1 3′ UTRMyc-GST
Disc1–/–
Ven
usV
enus
/MA
P2
b
20
40
60
80
Rel
ativ
e fr
eque
ncy
(%)
100
Venus–ITPR1 3′ UTR
: 0–20 µm: 20–40 µm: 40–60 µm: 60–80 µm: 80–100 µm
Disc1–/
–
Myc
-GST
Disc1–/
–
Myc
-hDIS
C1-FL
Disc1–/
–
Myc
-hDIS
C1-∆ARM
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature neurOSCIenCe VOLUME 18 | NUMBER 5 | MAY 2015 705
a r t I C l e S
of the DISC1-N1-∆ARM mutant decreased the transport of ITPR1 3′ UTR in dendrites (Fig. 6d and Supplementary Fig. 10b). HZF (aa 11–351) and HZF-∆Zn3 (aa 11–240) bound DISC1, whereas the HZF-∆Zn2,3 (aa 11–144) and HZF-∆Zn1 (aa 164–351) fragments did not (Fig. 6e and Supplementary Fig. 9b). Because HZF interacts with ITPR1 3′ UTR via the region containing the third Zn finger motif24 (Zn3), HZF-∆Zn3 can act as a dominant-negative mutant to interrupt the binding between DISC1 and HZF without affecting RNA bind-ing. In fact, we were able to demonstrate that HZF-∆Zn3 inhibited the association of HZF with DISC1 (Supplementary Fig. 11a). The expression of HZF-∆Zn3 inhibited the dendritic localization of ITPR1 3′ UTR, whereas the expression of full-length HZF or HZF-∆Zn2,3 did not (Fig. 6f and Supplementary Fig. 11b). These expression stud-ies with dominant-negative fragments such as DISC1-N1-∆ARM and HZF-∆Zn3 suggest that the interaction between HZF and DISC1 is involved in the dendritic transport of ITPR1 mRNA.
Physiological function of DISC1 in synaptic plasticityTo examine the effect of the loss of DISC1 on the dendritic trans-port of ITPR1 3′ UTR under physiological conditions, we examined the localization of ITPR1 3′ UTR in hippocampal slices of wild-type and Disc1−/− mice (Supplementary Fig. 12a,b). In CA1 pyramidal neurons, DISC1 deficiency inhibited the transport of ITPR1 3′ UTR (Supplementary Fig. 12a,b), indicating that DISC1 regulates the transport of ITPR1 mRNA in brain tissue.
Because the maintenance phase of long-term potentiation (LTP) is thought to require the transport and translation of neuronal mRNAs in dendrites35,36, we examined the maintenance of LTP evoked by ten
trains of high-frequency stimulation (HFS; 100 pulses at 100 Hz) in Disc1−/− mice. In hippocampal slices from wild-type mice, the ten trains of HFS resulted in an enhanced slope of the field excitatory postsyn-aptic potential (fEPSP) that persisted for at least 3 h (Supplementary Fig. 12c,d). However, in hippocampal slices from the Disc1−/− mice, the same protocol resulted in the induction of LTP, and the fEPSP enhance-ment did not persist (Supplementary Fig. 12c,d). These results show that DISC1 has a key role in the late phase of LTP.
To examine the physiological relevance of DISC1-mediated mRNA regulation for synaptic plasticity, we conducted an electrophysiological study with the DISC1-inhibitory peptide DISC1Ntide. DISC1Ntide is a 33-aa peptide that contains a cell-permeable peptide, antennapedia homeodomain37, and an arginine-rich motif of DISC1 (Fig. 7a). To examine the cell permeabilities of DISC1Ntide and the control peptide, which contained antennapedia homeodomain alone, we bath-applied DISC1Ntide and the control peptide to dissociated hippocampal neu-rons at 14 d in vitro (DIV 14). The neurons were immunostained with an anti-antennapedia antibody 10 min after application of the pep-tides. Both of the peptides penetrated most of the neurons (Fig. 7a). We next examined the interactions of DISC1Ntide with the neuronal mRNAs that bound to DISC1. Hippocampal slices were treated with control peptide or DISC1Ntide. The cell lysates were subjected to an immunoprecipitation assay with anti-antennapedia antibody. We con-firmed that DISC1Ntide, but not the control peptides, coprecipitated with the mRNAs encoding ITPR1, KALIRIN and CACNA2D1, but not that for PGAM1 (Fig. 7b). To validate the inhibitory effect of DISC1Ntide in the interaction between DISC1 and RNA, we carried out UV cross-linking assays with DISC1Ntide (Fig. 7c). GST-hDISC1-FL
3′ UTRDISC1Ntide
+– 3 p 10 p 50 p
hDISC1-FL
GST-hDISC1-FL (1 p)
Control
*
(kDa)
10075
150
37
25
50
c
+ + +
– 50 p– – –
+–
a bHuman DISC1 (854 aa)
Control peptide:
Antennapediahomeodomain
DISC1Ntide:
32 48
Control peptide DISC1NtideITPR1 mRNA
KALRN mRNA
CACNA2D1 mRNA
PGAM1 mRNA
Inpu
t
IP:
Contro
l
DISC1N
tide
DISC1N
tide
Anti-antennapedia
RT – +– +Con
trol
d
Nor
mal
ized
fEP
SP
slop
e (%
)
–50
100
150
0 10 20 30 40Time (min)
Peptide
(N)-RQIKIWFQNRRMKWKK-(C)
ACFRRRRLARRPGYMRS
(N)-RQIKIWFQNRRMKWKK- ACFRRRRLARRPGYMRS-(C)
f
Time after HFS (min)
30 180
fEP
SP
slo
pe(%
bas
elin
e)
100
200
150
*
Control peptideDISC1Ntide
Peptide
200
100
Nor
mal
ized
fEP
SP
slop
e (%
)
–50 0 30 60 90 120 150 180
32010 Time (ms)Sw32010 Time (ms)Sw32010 Time (ms)Sw
V_i
npu
(mV
)
0
0
32010 Time (ms)Sw
V_i
npu
(mV
) 0
5 ms
0.5 mVControlpeptide
DISC1Ntide
12
12
Control peptideDISC1Ntide
e
Time (min)
1
2
Control peptideDISC1Ntide
Figure 7 Involvement of DISC1-mediated mRNA regulation in synaptic plasticity. (a) Diagram and permeabilization of control peptide and DISC1Ntide. The two lower panels show the permeabilization of the peptides at 10 min after bath application. The treated neurons (DIV 9) were stained with anti-antennapedia (red). Scale bars, 20 µm. (b) In vivo associations of DISC1Ntide with neuronal mRNAs. The indicated mRNAs (except for PGAM1) were coprecipitated with anti-antennapedia from the hippocampal lysates treated with DISC1Ntide (see Online Methods). IP, immunoprecipitate; RT, reverse transcriptase. (c) Dose-dependent inhibition of the interaction between DISC1 and ITPR1 3′ UTR by DISC1Ntide. Competition analysis was performed using excess DISC1Ntide or control peptide. p, picomoles. Asterisk marks a degradation product of the GST-hDISC1 protein. (d) The time course of the effects of control peptide and DISC1Ntide on basal synaptic transmission in the dentate gyrus (3-week-old C57BL/6J mice). A 30-min bath application of control peptide or DISC1Ntide (10 nM) had no significant effect on the baseline of the fEPSP slope in Mann-Whitney U-tests (control peptide: 101.8 ± 6.0% of baseline, nonsignificant P value, U-test, n = 7; DISC1Ntide: 107.7 ± 11.3% of baseline, nonsignificant P value, n = 7). The average fEPSP slopes in control-treated and DISC1Ntide-treated slices were −0.61 ± 0.12 mV ms−1 and −0.65 ± 0.13 mV ms−1, respectively. (e) Time course of the effects of control peptide and DISC1Ntide on the late phase of LTP in the dentate gyrus. The upper traces show the typical recordings of fEPSPs before (1) and after (2) tetanic stimulation. The slices treated with control peptide were subjected to ten trains of tetanic stimulation (HFS: 100 Hz and 100 pulses at 60-s intervals, indicated by arrows), which produced long-lasting potentiation persisting for at least 3 h after the HFS (140.2 ± 18.8% of baseline 180 min after HFS, n = 7). In contrast, the potentiation in the slices treated with DISC1Ntide returned to near baseline level by 3 h (100.4 ± 11.8% of baseline 180 min after HFS, n = 7). (f) LTP amplitude was averaged at 30 min and 180 min after HFS and is expressed as a percentage of the baseline value. Statistical analysis was done via Mann-Whitney U-test (LTP amplitude: *P < 0.05, control versus DISC1Ntide at 30 min, n = 7; control versus DISC1Ntide at 180 min, n = 7). Data are expressed as mean ± s.d. in d–f. Full-length blots are shown in Supplementary Figure 15.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

706 VOLUME 18 | NUMBER 5 | MAY 2015 nature neurOSCIenCe
a r t I C l e S
bound to biotin-labeled ITPR1 3′ UTR RNA in the absence of DISC1Ntide. DISC1Ntide blocked the interaction between DISC1-FL and ITPR1 3′ UTR in a dose-dependent manner, whereas the control peptide did not block this interaction (Fig. 7c). These results support the conclusion that DISC1Ntide is a useful tool for inhibiting the interaction between DISC1 and RNA. We prepared acute hippocampal slices from wild-type mice (C57BL/6J, 3 weeks in age) and examined LTP after treatment using either a control peptide or DISC1Ntide. Applications of control peptides and DISC1Ntide (10 nM) had no effect on the baseline of the fEPSP during a 30-min baseline record-ing period (Fig. 7d). In the slices treated with control peptide, ten trains of HFS resulted in enhancement of the fEPSP slope, and this enhancement persisted for at least 3 h (Fig. 7e,f). In contrast, LTP induction was observed in the slice treated with DISC1Ntide, and LTP maintenance was not detected (Fig. 7e,f). Although we cannot exclude an effect of DISC1Ntide on the interaction of endogenous DISC1 with other DISC1 interactors, these results suggest that the interaction between DISC1 and RNA is involved in the late phase of LTP.
DISCUSSIONWe here report the discovery of a role for DISC1 as an RNA-binding protein for mRNA transport. Impaired function of DISC1 or HZF inhibited the dendritic transport of ITPR1 mRNA. DISC1 interacted with RNA molecules via an ARM in its N terminus. The ARM in DISC1 was responsible for the transport of ITPR1 mRNA and seemed to contain a nuclear localization signal (NLS). It has been reported that the RNA-binding protein FMRP requires an NLS for mRNA export from the nucleus38; thus DISC1 may have a function similar to that of FMRP. DISC1 and HZF bound to distinct regions of ITPR1 mRNA, and the binding of DISC1 was stabilized by HZF. Moreover, the interaction of DISC1 with HZF was required for the transport of ITPR1 mRNA. These results suggest that DISC1 recognizes HZF and ITPR1 mRNA simultaneously to provide the combinatorial avidity necessary for transport of the mRNA (Supplementary Fig. 13a). Given that kinesin-1 interacts with DISC1 and regulates its trans-port along microtubules5, and that expression of a dominant- negative mutant of kinesin heavy chain inhibits the transport of ITPR1 mRNA, it can be assumed that a molecular complex composed of DISC1, HZF and ITPR1 mRNA would be transported to the distal end of dendrites in a kinesin-1–dependent manner. In this study, we found that the overexpression of DISC1 facilitated the dendritic transport of ITPR1 mRNA, whereas DISC1 depletion had a negative effect on ITPR1 mRNA transport. We have shown that DISC1 functions as a cargo adaptor, linking the bound mRNAs to the kinesin-1 complex for mRNA transport. Molecular motors such as kinesin associate with their cargo through adaptor proteins that regulate the attach-ment and unloading of the cargo39. The overexpression of DISC1 would increase the attachment rate between ITPR1 3′ UTR RNA and kinesin-1 to promote the transport of its 3′ UTR.
What is the physiological meaning of DISC1-mediated mRNA regulation? We previously showed altered synaptic plasticity and an LTP defect in Disc1−/− mice8. Here we found that DISC1 associated with neuronal mRNAs encoding intracellular calcium release channel ITPR1; the neuronal RhoGEF kalirin; and subunits of voltage-gated ion channels such as KCNC1, KCNC4, CACNA1C and CACNA2D1. IP3 and its receptor are essential for synaptic plasticity and motor learning in the cerebellum and hippocampus40, and HZF is required for the translation of ITPR1 in response to BDNF stimulation in hippocampal neurons24. Mice lacking HZF have severe impairments in motor coordination and cerebellum-dependent motor learning41.
The voltage-activated potassium channels KCNC1 and KCNC4 belong to the Kv3 channel family and are important for controlling spike fre-quency at synapses42. It has been reported that FMRP directly binds to KCNC1 mRNA43. DISC1 and FMRP may cooperatively regulate the localization and translation of Kcnc1 mRNA for synaptic plasticity. CACNA1C and CACNA2D1 encode α1 and α2/δ subunits of L-type voltage-gated calcium channels. A study with Cacna1c-knockout mice revealed an essential role in LTP and spatial learning in the hippo-campus44. Kalirin is a known binding partner of DISC1, and these two proteins cooperatively regulate synaptic morphology and neuronal plasticity45. The results of our electrophysiological analyses with the DISC1-inhibitory peptide DISC1Ntide support the proposal that the DISC1-RNA interaction is involved in the maintenance of LTP. The LTP dysregulation observed with DISC1Ntide treatment phenocopied the LTP deficit in Disc1−/− mice. Considering the RIP-qPCR results in the screen for DISC1-interacting mRNAs and the findings from the electrophysiological studies, we propose that DISC1 regulates synaptic plasticity via the regulation of synaptic-molecule mRNAs.
Gene silencing is widely used to evaluate the physiological role of DISC1 (ref. 7). In the present study, we confirmed that transient Disc1 knockdown by RNAi inhibited the dendritic transport of ITPR1 mRNA. Some groups have reported that Disc1 knockdown alters the frequency and amplitude of miniature excitatory postsynaptic currents in cortical neurons45,46. Given that downregulation of synaptic effi-ciency can contribute to altered synaptic plasticity, the results of Disc1 knockdown studies appear to be consistent with those of the Disc1-knockout study. In spite of DISC1’s function in synaptic plasticity, the Disc1−/− mouse displays no gross abnormalities in the brain’s cytoar-chitecture8, whereas Disc1 knockdown caused severe neurodevelop-mental abnormalities, including neurogenesis and cell migration7. This phenotypic discrepancy between Disc1−/− and Disc1-knockdown mice may be explained by off-target effects of short hairpin RNAs (shRNAs), as pointed out previously9. Some shRNAs are known to have off-target effects and nonspecific cytotoxicity47,48. Recently, Baek et al. reported that shRNAs against doublecortin cause cortical cell-migration defects via an off-target effect48. To examine whether Disc1 RNAi has an off-target effect on cortical migration, we focused on the specificity of Disc1 knockdown using a shRNA vector (shRNA-D1) that has been widely used in DISC1 studies12,14,49,50. Because the target sequence of shRNA-D1 is located in exon 2 of the Disc1 gene, shRNA-D1 did not affect Disc1 mRNA in our Disc1−/− mice lacking exons 2 and 3. When the shRNA-D1 vector was transfected into wild-type and Disc1−/− mice, both types of mice showed a severe migration defect in cortical neurons (Supplementary Fig. 13b). This result clearly dem-onstrates that shRNA-D1 has off-target effects. In addition, other Disc1 RNAi tools that confer a defective phenotype in the 129 and outbred ICR strain backgrounds require verification of the knockdown specifi-city. Some groups have reported that Disc1 knockdown in the brains of 129 and outbred ICR mice leads to extensive disruption of cortical migration and adult neurogenesis14,17–19. One of the Disc1 shRNAs used in these studies is shRNA-D1. We confirmed that shRNA-D1 was able to suppress endogenous DISC1 expression using our DISC1 antibodies. The other Disc1 shRNAs have not been verified for their knockdown specificity toward endogenous DISC1, or for their off-target effects13,17,45. Taken together, these results indicate that DISC1 studies using Disc1 shRNAs should be interpreted with caution.
METHODSMethods and any associated references are available in the online version of the paper.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature neurOSCIenCe VOLUME 18 | NUMBER 5 | MAY 2015 707
a r t I C l e S
Note: Any Supplementary Information and Source Data files are available in the online version of the paper.
AcknowledgmenTSWe gratefully acknowledge T. Takumi (Hiroshima University, Higashihiroshima, Japan) for providing rat Staufen cDNA, S. Okabe (University of Tokyo, Tokyo, Japan) for providing the pAct expression vector, H. Song (Johns Hopkins University school of Medicine, Baltimore, Maryland) for providing the Disc1 shRNA vector and K. Kosik (University of California, Santa Barbara, Santa Barbara, California) for providing the RSV-MS2-CAMK2A 3′ UTR construct. We thank T. Fujiwara (Kobe University) and A. Sawa (Johns Hopkins University) for helpful discussions. We also thank Y. Funahashi, Y. Fujino, H. Yano and T. Watanabe for providing materials; K. Kobayash and S. Suzuki for technical assistance; and T. Ishii for secretarial assistance. This work was supported in part by a Grant-in-Aid for the Strategic Research Program for Brain Science (SRPBS; Theme G) (26-J-JG38), a Grant-in-Aid for Scientific Research (S) (20227006), an Academic Frontier Project from the Ministry of Education, Culture, Sports, Science and Technology of Japan (MEXT) and a Grant-in-Aid for Creative Scientific Research from the Japan Society for the Promotion of Science, Japan Science and Technology Agency, CREST (26-J-Jc08).
AUTHoR conTRIBUTIonSD.T. carried out most of the cell biology experiments; K. Kuroda and S.T. prepared the gene-knockout mouse and antibodies to DISC1; T.N. carried out in vivo experiments with in utero electroporation; Y.I., T.S. and T.H. carried out biochemical experiments; M.I. and A.M. carried out immunoelectron microscopy analysis; Y.I., S.M. and A.N. carried out affinity chromatography; M.T. and M.S. carried out an electrophysiological study; N.S., H.O. and K.M. made materials and analyzed results; and K. Kaibuchi guided the research and wrote the paper.
comPeTIng FInAncIAl InTeReSTSThe authors declare no competing financial interests.
Reprints and permissions information is available online at http://www.nature.com/reprints/index.html.
1. Blackwood, D.H. et al. Schizophrenia and affective disorders—cosegregation with a translocation at chromosome 1q42 that directly disrupts brain-expressed genes: clinical and P300 findings in a family. Am. J. Hum. Genet. 69, 428–433 (2001).
2. Millar, J.K. et al. Disruption of two novel genes by a translocation co-segregating with schizophrenia. Hum. Mol. Genet. 9, 1415–1423 (2000).
3. Brandon, N.J. & Sawa, A. Linking neurodevelopmental and synaptic theories of mental illness through DISC1. Nat. Rev. Neurosci. 12, 707–722 (2011).
4. Chubb, J.E., Bradshaw, N.J., Soares, D.C., Porteous, D.J. & Millar, J.K. The DISC locus in psychiatric illness. Mol. Psychiatry 13, 36–64 (2008).
5. Taya, S. et al. DISC1 regulates the transport of the NUDEL/LIS1/14-3-3ε complex through kinesin-1. J. Neurosci. 27, 15–26 (2007).
6. Shinoda, T. et al. DISC1 regulates neurotrophin-induced axon elongation via interaction with Grb2. J. Neurosci. 27, 4–14 (2007).
7. Brandon, N.J. et al. Understanding the role of DISC1 in psychiatric disease and during normal development. J. Neurosci. 29, 12768–12775 (2009).
8. Kuroda, K. et al. Behavioral alterations associated with targeted disruption of exons 2 and 3 of the Disc1 gene in the mouse. Hum. Mol. Genet. 20, 4666–4683 (2011).
9. Kvajo, M., McKellar, H. & Gogos, J.A. Avoiding mouse traps in schizophrenia genetics: lessons and promises from current and emerging mouse models. Neuroscience 211, 136–164 (2012).
10. Nakata, K. et al. DISC1 splice variants are upregulated in schizophrenia and associated with risk polymorphisms. Proc. Natl. Acad. Sci. USA 106, 15873–15878 (2009).
11. Ishizuka, K. et al. Evidence that many of the DISC1 isoforms in C57BL/6J mice are also expressed in 129S6/SvEv mice. Mol. Psychiatry 12, 897–899 (2007).
12. Duan, X. et al. Disrupted-In-Schizophrenia 1 regulates integration of newly generated neurons in the adult brain. Cell 130, 1146–1158 (2007).
13. Mao, Y. et al. Disrupted in schizophrenia 1 regulates neuronal progenitor proliferation via modulation of GSK3beta/beta-catenin signaling. Cell 136, 1017–1031 (2009).
14. Kim, J.Y. et al. DISC1 regulates new neuron development in the adult brain via modulation of AKT-mTOR signaling through KIAA1212. Neuron 63, 761–773 (2009).
15. Koike, H., Arguello, P.A., Kvajo, M., Karayiorgou, M. & Gogos, J.A. Disc1 is mutated in the 129S6/SvEv strain and modulates working memory in mice. Proc. Natl. Acad. Sci. USA 103, 3693–3697 (2006).
16. Clapcote, S.J. & Roder, J.C. Deletion polymorphism of Disc1 is common to all 129 mouse substrains: implications for gene-targeting studies of brain function. Genetics 173, 2407–2410 (2006).
17. Kamiya, A. et al. A schizophrenia-associated mutation of DISC1 perturbs cerebral cortex development. Nat. Cell Biol. 7, 1167–1178 (2005).
18. Kubo, K. et al. Migration defects by DISC1 knockdown in C57BL/6, 129X1/SvJ, and ICR strains via in utero gene transfer and virus-mediated RNAi. Biochem. Biophys. Res. Commun. 400, 631–637 (2010).
19. Niwa, M. et al. Knockdown of DISC1 by in utero gene transfer disturbs postnatal dopaminergic maturation in the frontal cortex and leads to adult behavioral deficits. Neuron 65, 480–489 (2010).
20. Bannai, H. et al. An RNA-interacting protein, SYNCRIP (heterogeneous nuclear ribonuclear protein Q1/NSAP1) is a component of mRNA granule transported with inositol 1,4,5-trisphosphate receptor type 1 mRNA in neuronal dendrites. J. Biol. Chem. 279, 53427–53434 (2004).
21. Kanai, Y., Dohmae, N. & Hirokawa, N. Kinesin transports RNA: isolation and characterization of an RNA-transporting granule. Neuron 43, 513–525 (2004).
22. Kiebler, M.A. & Bassell, G.J. Neuronal RNA granules: movers and makers. Neuron 51, 685–690 (2006).
23. Anderson, P. & Kedersha, N. RNA granules. J. Cell Biol. 172, 803–808 (2006).24. Iijima, T. et al. Hzf protein regulates dendritic localization and BDNF-induced
translation of type 1 inositol 1,4,5-trisphosphate receptor mRNA. Proc. Natl. Acad. Sci. USA 102, 17190–17195 (2005).
25. Knowles, R.B. et al. Translocation of RNA granules in living neurons. J. Neurosci. 16, 7812–7820 (1996).
26. Köhrmann, M. et al. Microtubule-dependent recruitment of Staufen-green fluorescent protein into large RNA-containing granules and subsequent dendritic transport in living hippocampal neurons. Mol. Biol. Cell 10, 2945–2953 (1999).
27. Bertrand, E. et al. Localization of ASH1 mRNA particles in living yeast. Mol. Cell 2, 437–445 (1998).
28. Rook, M.S., Lu, M. & Kosik, K.S. CaMKIIα 3′ untranslated region-directed mRNA translocation in living neurons: visualization by GFP linkage. J. Neurosci. 20, 6385–6393 (2000).
29. Hirokawa, N. & Takemura, R. Molecular motors and mechanisms of directional transport in neurons. Nat. Rev. Neurosci. 6, 201–214 (2005).
30. Presley, J.F. et al. ER-to-Golgi transport visualized in living cells. Nature 389, 81–85 (1997).
31. King, S.J. & Schroer, T.A. Dynactin increases the processivity of the cytoplasmic dynein motor. Nat. Cell Biol. 2, 20–24 (2000).
32. Dictenberg, J.B., Swanger, S.A., Antar, L.N., Singer, R.H. & Bassell, G.J. A direct role for FMRP in activity-dependent dendritic mRNA transport links filopodial-spine morphogenesis to fragile X syndrome. Dev. Cell 14, 926–939 (2008).
33. Nakata, T. & Hirokawa, N. Microtubules provide directional cues for polarized axonal transport through interaction with kinesin motor head. J. Cell Biol. 162, 1045–1055 (2003).
34. Bayer, T.S., Booth, L.N., Knudsen, S.M. & Ellington, A.D. Arginine-rich motifs present multiple interfaces for specific binding by RNA. RNA 11, 1848–1857 (2005).
35. Bramham, C.R. & Wells, D.G. Dendritic mRNA: transport, translation and function. Nat. Rev. Neurosci. 8, 776–789 (2007).
36. Miller, S. et al. Disruption of dendritic translation of CaMKIIα impairs stabilization of synaptic plasticity and memory consolidation. Neuron 36, 507–519 (2002).
37. Jones, S.W. et al. Characterisation of cell-penetrating peptide-mediated peptide delivery. Br. J. Pharmacol. 145, 1093–1102 (2005).
38. Eberhart, D.E., Malter, H.E., Feng, Y. & Warren, S.T. The fragile X mental retardation protein is a ribonucleoprotein containing both nuclear localization and nuclear export signals. Hum. Mol. Genet. 5, 1083–1091 (1996).
39. Hirokawa, N., Niwa, S. & Tanaka, Y. Molecular motors in neurons: transport mechanisms and roles in brain function, development, and disease. Neuron 68, 610–638 (2010).
40. Bardo, S., Cavazzini, M.G. & Emptage, N. The role of the endoplasmic reticulum Ca2+ store in the plasticity of central neurons. Trends Pharmacol. Sci. 27, 78–84 (2006).
41. Iijima, T. et al. Impaired motor functions in mice lacking the RNA-binding protein Hzf. Neurosci. Res. 58, 183–189 (2007).
42. Rudy, B. & McBain, C.J. Kv3 channels: voltage-gated K+ channels designed for high-frequency repetitive firing. Trends Neurosci. 24, 517–526 (2001).
43. Strumbos, J.G., Brown, M.R., Kronengold, J., Polley, D.B. & Kaczmarek, L.K. Fragile X mental retardation protein is required for rapid experience-dependent regulation of the potassium channel Kv3.1b. J. Neurosci. 30, 10263–10271 (2010).
44. Moosmang, S. et al. Role of hippocampal Cav1.2 Ca2+ channels in NMDA receptor-independent synaptic plasticity and spatial memory. J. Neurosci. 25, 9883–9892 (2005).
45. Hayashi-Takagi, A. et al. Disrupted-in-Schizophrenia 1 (DISC1) regulates spines of the glutamate synapse via Rac1. Nat. Neurosci. 13, 327–332 (2010).
46. Maher, B.J. & LoTurco, J.J. Disrupted-in-schizophrenia (DISC1) functions presynaptically at glutamatergic synapses. PLoS ONE 7, e34053 (2012).
47. McBride, J.L. et al. Artificial miRNAs mitigate shRNA-mediated toxicity in the brain: implications for the therapeutic development of RNAi. Proc. Natl. Acad. Sci. USA 105, 5868–5873 (2008).
48. Baek, S.T. et al. Off-target effect of doublecortin family shRNA on neuronal migration associated with endogenous microRNA dysregulation. Neuron 82, 1255–1262 (2014).
49. Kang, E. et al. Interaction between FEZ1 and DISC1 in regulation of neuronal development and risk for schizophrenia. Neuron 72, 559–571 (2011).
50. Zhou, M. et al. mTOR inhibition ameliorates cognitive and affective deficits caused by Disc1 knockdown in adult-born dentate granule neurons. Neuron 77, 647–654 (2013).
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature neurOSCIenCe doi:10.1038/nn.3984
ONLINE METHODSPlasmid constructs and biological materials. Full-length human DISC1 cDNA (aa 1–832) was inserted into pGEX-4T (Pharmacia Biotech, Piscataway, NJ), pMAL-c2 (New England Biolabs, Ipswich, MA), pEFBos-GST, pCAGGS-Myc, pEGFP-N3 (Clontech Laboratories, Palo Alto, CA), p∆CMV51 and pßact (a gift from S. Okabe, Tokyo University) vectors. The DISC1-∆ARM (in-frame deletion spanning aa 35–42), DISC1-∆HRM (in-frame deletion spanning aa 77–84), DISC1-∆KBR (in-frame deletion spaning aa 120–400), DISC1-NH (aa 1–361), DISC1-CH (aa 362–832), DISC1-N1 (aa 1–120), DISC1-N23 (aa 121–361) and DISC1-N1-∆ARM fragments were inserted into pEFBos-GST, pMAL-c2, pCAGGS-Myc, pEGFP-N3, pGEX-4T1 and pßact. Unless otherwise stated, human full-length DISC1 and its subfragments were used in this study. Full-length HZF cDNA was inserted into pFLAG, pRSET (Invitrogen, Carlsbad, CA) and pCAGGS-Myc. The HZF (aa 11–351), HZF-∆Zn1 (aa 164–351), HZF-∆Zn2,3 (aa 11–145) and HZF-∆Zn3 (aa 11–240) cDNA fragments were inserted into pCAGGS-Myc. A fragment of KIF5A–headless (HL) (aa 402–1,027) was also inserted into pCAGGS-Myc. MS2-GFP constructs for CaM-kinase IIα were kindly provided by K. Kosik (UCSB, Santa Barbara, CA). In addition, ITPR1 3′ UTR–FL (bases 1–462), −F1 (bases 1–200), −F2 (bases 201–462), −F3 (bases 201–309) and −F4 (bases 310–462) were cloned into the pTNT vector (Promega, Madison, WI) for synthesis of in vitro–transcribed RNAs. Rabbit polyclonal anti-bodies to mouse DISC1 (DISC1c-Ab; dilution, 1:5,000) and HZF (dilution, 1: 1,000) were used as previously described by our group24. An HZF monoclonal antibody (dilution, 1:200; clone 132-2) was kindly provided by M. Sugimoto (National Center for Geriatrics and Gerontology, Aichi, Japan). Antibodies to Venus-GFP (dilution, 1:500; catalog no. 598, MBL, Nagoya, Japan), β-tubulin (dilution, 1:500; clone AA2, Sigma, St. Louis, MO), KIF5 (dilution, 1:500; clone H2, Merck Millipore, Billerica, MA), MAP2 (dilution, 1:500; M3696, Sigma), HnRNP-U (dilution, 1:200; clone 3G6, Sigma), SYNCRIP (dilution, 1:200; clone 18E4, Sigma), RACK1 (dilution, 1:500; 610177, BD Transduction Laboratories, Lexington, KY), GRB2 (dilution, 1:500; clone 81, BD Transduction Laboratories), PURα (dilution, 1:1,000; purified by our laboratory), GAPDH (dilution, 1:500; clone 71.1, Sigma), ZBP1 (dilution, 1:200; catalog no. RN001M, MBL), AGO2 (dilution, 1:200; clone 11A9, Sigma), eIF4G (dilution, 1:200; clone 2A9, Sigma) and FMRP (dilution, 1:200; clone 3E11, Abnova, Taipei, Taiwan) and Kusabira Orange (dilution, 1:500; catalog no. M104, MBL) were used in immunoblot and immunostaining assays. For the UV cross-linking assay, we used the 3′ UTR of ITPR1 cDNA from a previous study20. The 3′ UTRs of CACNA1C, CACNA2D1, KCNC1, KCNC4, KALRN and SCN2A mRNAs and the CACNA2D1 cDNA were amplified from a mouse cDNA library with the following prim-ers: CACNA1C, forward (Fwd), 5′-TGCCTCACTGTTCTCGTGAC-3′, reverse (Rev), 5′-CGTAAAGCTTTTCACACTCCAAA-3′; CACNA2D1, Fwd, 5′-CCTTCTAGAACCCAGATC-3′, Rev, 5′-ATTATTTCATCAGTTTAAAC-3′; KCNC1, Fwd, 5′-TGGTGGAGGAATGAGAAAGC-3′, Rev, 5′-CCTGGA CTGCAGGGACTCTA-3′; KCNC4, Fwd, 5′-AACTATGCCCACGCTGAAGT-3′, Rev, 5′-GGTACTCGAGCACAGCATGA-3′; KALRN, Fwd, 5′-CAGGA CTCCAATCACCCAAC-3′, Rev, 5′-GCGTGTGAGACACATGGAAG-3′; SCN2A, Fwd, 5′-CGACTCCCTCAGGAGGTCTA-3′, Rev, 5′-GCTTGATTGT AATGTGGCTTACC-3′; and CACNA2D1 cDNA, Fwd, 5′- CCTTGT CACACTGGCAAAAA-3′, Rev, 5′-TGTCTGCTGCCAGATACCAG-3′. Gene-specific primers for quantitative PCR are shown in Supplementary Table 2.
For the electrophysiological study with the DISC1-inhibitory peptide (DISC1Ntide), DISC1Ntide and control peptide (98% purity) were obtained from the peptide-synthetic service at MBL (Nagoya, Aichi, Japan).
Affinity chromatography and mass spectrometry analysis. Affinity chromatog-raphy was performed as described5. Briefly, rat brain cytosol or COS7 cell lysate was loaded onto amylose resin (New England Biolabs) coated with MBP alone or MBP-hDISC1. The columns were washed with buffer A (20 mM Tris-HCl, pH 8.0, 1 mM dithiothreitol (DTT), 1 mM EDTA, and protease inhibitors). The proteins that bound to the affinity columns were eluted with buffer A containing 500 mM NaCl. When necessary, rat brain cytosol was treated with RNase A (0.1 mg/ml) for 6 h at 4 °C before being applied to the column. The eluates were resolved by SDS-PAGE followed by silver staining (Thermo Scientific, Rockford, IL). For in-gel digestion, the excised gels were destained and dried according to the manufacturer’s protocol (Thermo Scientific). The dried gel pieces were digested at 37 °C overnight with 25 mM ammonium bicarbonate containing 0.05 µg
of trypsin. The tryptic peptides were analyzed in an LTQ Orbitrap mass spec-trometer (Thermo Scientific). Peak-list generation and database searches against the rodent proteome from the National Center for Biotechnology Information database were performed with Mascot (version 2.302, Matrix Science, Boston, MA). Carbamidomethylation of cysteine and oxidation of methionine were set as variable modifications. Up to one missed trypsin cleavage was allowed. The mass tolerance for peptide ions and fragment ions was set at 10 ppm and 0.8 Da, respectively. The significance threshold of the Mascot search was P < 0.05.
Biochemical analysis. GST pulldown and immunoprecipitation assays were per-formed as previously described5. Briefly, COS7 cells were transfected in various combinations with lipofectamine and cultured for 48 h. The cells or rodent brains were lysed with ice-cold lysis buffer (20 mM Tris-HCl, pH 8.0, 1 mM EDTA, 100 mM NaCl, 1 mM DTT, 1% (w/v) NP-40, and protease inhibitors). The lysates were mixed with DISC1 recombinant proteins or the indicated antibodies (Fig. 1c,d) and incubated for 1 h at 4 °C. The eluate was subjected to SDS-PAGE followed by immunoblotting with the primary antibodies to the indicated proteins (Fig. 1c,d and Supplementary Figs. 7, 9, 10a and 11a). In vitro binding assays were performed as described5. Briefly, MBP-hDISC1 fusion proteins were mixed with affinity beads coated with histidine (His)-tagged HZF protein in buffer A (see above). The beads were then washed with buffer A, and the bound proteins were eluted with buffer A containing 300 mM imidazole. The eluates were subjected to SDS-PAGE followed by immunoblotting. For the RIP-qPCR assay, P9 mouse brain tissues were lysed with the modified IMAC buffer (20 mM HEPES, pH 7.4, 140 mM potassium acetate, 1 mM magnesium acetate, 1 mM EDTA, 0.5% NP-40). The cell lysate was immunoprecipitated with DISC1c-Ab. Coprecipitated RNAs were puri-fied from the DISC1 immunoprecipitants by QIAZOL (Qiagen, Valencia, CA) and subjected to quantitative PCR with gene-specific primers. The mRNA enrich-ment was determined by normalizing the DISC1 signal against the control rabbit IgG signal in three independent experiments. For the immunoprecipitation assay with peptides, mouse hippocampal slices that were treated with control peptide or DISC1Ntide were lysed with the modified IMAC buffer. The cell lysates were sub-jected to immunoprecipitation with anti-antennapedia homeodomain (Abcam). The coprecipitated RNAs were purified from the immunoprecipitates and were subjected to RT-PCR for ITPR1, KALRN, CACNA2D1 and PGAM1 mRNAs. For the UV cross-linking assay, plasmid templates were digested with the appropriate restriction enzymes and transcribed (Riboprobe in Vitro Transcription Kit, Promega) with biotin-UTP (Roche, Laval, Canada). The RNA transcripts were purified on G-50 Sephadex columns (Roche). The biotin-labeled RNAs were incubated at room temperature with 1 pmol of human DISC1 recombinant pro-tein in 10 µl of a HEPES-based buffer (10 mM HEPES, pH 7.3, 150 mM KCl, 1 mM MgCl2, 1 mM DTT, 5% glycerol, and 2 µg tRNA) for 20 min. The samples were UV-irradiated on ice for 5 min, and unbound RNAs were then degraded by incubation with 1 mg/ml RNase A or T1. The UV cross-linked RNA-protein com-plexes were separated using 10% SDS-PAGE followed by immunoblot analysis with streptavidin-HRP (Promega). A gel retardation assay with ITPR1 3′ UTR RNA was carried out as previously described52,53.
culture of mouse hippocampal neurons and transfection. All of the ani-mal studies were conducted in accordance with the Animal Care and Use Committee guidelines of Nagoya University in Japan. The hippocampal neu-rons were prepared from E16 embryos of wild-type (C57BL/6J) and Disc1−/− mice as previously described5. Briefly, neurons were seeded onto a poly-d-lysine-coated 35-mm glass-based dish and cultured in neurobasal medium (Invitrogen) supplemented with B-27 (Invitrogen) and 1 mM l-glutamine. In some RNAi knockdown experiments, siRNAs targeting the following sequences were used to downregulate SYNCRIP and HZF expression and were used as previously described21,54: 5′-GUGCUGUUUGUACGCAACC-3′ for SYNCRIP and 5′-AGCCAUGUCCAUCCAACCC-3′ for HZF. The scrambled sequence 5′-CAGTCGCGTTTGCGACTGG-3′, which has no homology to any mamma-lian mRNA, was used as a negative control. For the studies with in utero electro-poration, we used shRNA vectors (pUEG-C1 and pUEG-D1) kindly provided by S. Hongjun (Johns Hopkins University School of Medicine). To visualize the 3′ UTR of ITPR1 mRNA, we co-transfected cultured neurons with expression vectors encoding the 3′ UTR of mouse Itpr1 mRNA fused with 12 copies of the MS2 binding sequence (Itpr1 3′ UTR–MS2bs) and Venus fused with the MS2-nls protein (MS2-nls–Venus), as previously reported20. The bacteriophage MS2 coat
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature neurOSCIenCedoi:10.1038/nn.3984
protein binds with high affinity to specific RNA-binding sequences (MS2bs), multiple repeats of which were introduced into a reporter transcript. MS2 was fused to Venus, a yellow fluorescent protein, with a nuclear localization signal (nls) to force its accumulation in the nucleus. When necessary, siRNAs and plas-mids were transfected into the cultured neurons with MS2-nls–Venus and Itpr1 3′ UTR–MS2bs vectors. The dissociated neurons were transfected using a CalPhos transfection kit (Clontech), Lipofectamine 2000 (Invitrogen) and a Neon transfec-tion system (Invitrogen). For the transfection of DNA into organotypic slices from mice aged 7 postnatal days, a Helios gene gun system (Bio-Rad, Richmond, CA) was used according to the manufacturer’s instructions. In utero electroporation was performed as previously described, with some modifications55.
Immunofluorescence analysis. We immunostained cultured hippocampal neu-rons as previously described56. Briefly, hippocampal neurons were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) for 10 min and treated with PBS containing 0.05% Triton X-100 for 10 min. The fixed neurons were incubated overnight at 4 °C with primary antibodies, washed, and incubated for 1 h with sec-ondary antibodies. Fluorescent images were taken with a laser-scanning confocal microscope (LSM PASCAL, Carl Zeiss, Oberkochen, Germany). For the visu-alization of RNA molecules, living neurons were incubated with 50 nM SYTO14 dye (S7576, Life Technologies, Carlsbad, CA). The stained neurons were fixed at 30 min after dye application. All of the images taken using the confocal microscope were digitally smoothed to reduce noise levels using NIH ImageJ software (NIH, Bethesda, MD). For the combined immunostaining and in situ hybridization assay, the neurons were fixed with 3% paraformaldehyde for 10 min at 4 °C, treated with 0.2% Triton X-100 and 2 mM ribonucleoside vanadyl complex (NEB, Ipswich, MA) in PBS for 10 min, and washed three times with PBS. The neurons were blocked with 1% bovine serum albumin, 1% normal donkey serum in PBS and incubated with DISC1c-Ab at 4 °C for 1 h. After the antibody had been washed off, the neurons were incubated with secondary antibody at 4 °C for 30 min. After washes with 2× saline–sodium citrate (SSC) buffer, the stained neurons were hybridized at 50 °C overnight with hybridization buffer (100 µg/ml yeast tRNA, 25% formamide, 10% dextran sulfate, Denhardt’s solution in 2× SSC) containing 200 ng/ml of the RNA probe for ITPR1. After washes with SSC buffer, we amplified the probe signals (Green) using the TSA-biotin system (Perkin-Elmer, Boston, MA).
Immunoelectron microscopy. One-week-postnatal mouse brains and cultured hippocampal neurons at DIV 14 were fixed with 2% paraformaldehyde in 100 mM phosphate buffer (pH 7.4) and incubated at room temperature for 5 h with the primary antibody to DISC1 described above. This step was followed by incuba-tion with secondary anti-rabbit coupled to 1.4-nm gold particles (Nanoprobes Inc., Yaphank, NY). The sample-bound gold particles were silver-enhanced using the HQ Silver kit (Nanoprobes) at room temperature for 8 min. The samples were post-fixed with 0.5% osmium oxide in a buffer (pH 7.3) containing 100 mM cacodylate. The samples were dehydrated in propylene oxide and embedded in epoxy resin. Ultrathin sections were cut from this sample, stained with uranyl acetate and lead citrate, and observed with an electron microscope (JEM-1011 EX, JEOL, Akishima, Tokyo, Japan).
electrophysiology. Electrophysiological analyses were conducted on male mice aged 3 weeks. Mice were decapitated while under deep anesthesia with chloral hydrate (400 mg/kg). Brains were quickly removed, and transverse 350-µm-thick slices were cut from the hippocampus using a vibratome in ice-cold modified artificial cerebrospinal fluid (ACSF) containing 206 mM sucrose, 5 mM KCl, 8 mM MgCl2, 1.25 mM KH2PO4, 1 mM CaCl2, 26 mM NaHCO3 and 10 mM d-glucose. ACSF was gassed with 95% O2, 5% CO2, and the pH was adjusted to 7.4. Slices were maintained for at least 1 h at room temperature (26–28 °C) in an incubation chamber containing gassed standard ACSF containing 128 mM NaCl, 5 mM KCl, 1.3 mM MgSO4, 1.25 mM KH2PO4, 2.41 mM CaCl2, 26 mM NaHCO3 and 10 mM d-glucose. A single hippocampal slice was transferred to and maintained for 1 h in the recording chamber, where it was superfused con-tinuously with gassed low-Mg2+ ACSF (0.7 mM MgSO4, instead of 1.3 mM as in standard ACSF) at a rate of 2–2.5 mL/min at 30 °C. A stimulating electrode (monopolar stimulation) was positioned in the molecular layer 30–50 µm from the granule cell layer in the dentate gyrus to activate the medial perforant path. Constant-current pulses (50 µs) were supplied by a stimulator (SEN-3301, Nihon Kohden, Tokyo, Japan) every 60 s. The intensity of the test stimuli was adjusted to
evoke approximately 50% to 60% of the maximum response. Using the conven-tional extracellular recording technique, fEPSPs were recorded from the middle molecular layer in the dentate gyrus with a glass pipette filled with 2 M NaCl (2–3 MΩ). During the recordings in experiments using peptides, a total volume of 25 mL of gassed low-Mg2+ ACSF with or without peptides was recirculated. The late phase of long-term potentiation (L-LTP) was induced by tetanic stimulation (10 trains, 1 s each, 100 Hz within the train, 60-s intertrain interval, 100-µs pulse width) under low-Mg2+ conditions to reliably induce L-LTP57. Signals were amplified and filtered at 5 kHz with an amplifier (Axopatch 200B, Axon Instruments). Data acquisition and analysis were performed using pCLAMP 9.0 software (Axon Instruments).
Imaging analysis. Time-lapse imaging analyses were performed as described previously56. Briefly, hippocampal neurons from E16 wild-type (C57BL/6J) and Disc1−/− mouse embryos were grown on glass-bottomed dishes (Matsunami, Osaka, Japan) and transfected with the indicated plasmids (Fig. 2). The time-lapse images of the cells were acquired with an inverted fluorescence microscope (IX81, Olympus, Tokyo, Japan) or with LSM PASCAL. For the measurement of the veloci-ties of DISC1-GFP and Venus–ITPR1 3′ UTR granules, we measured the peak velocities in unidirectionally moving granules during a given observation period (at 1-s intervals for 1 min) and defined the mean of the peak values as the velocity. To measure the transport distance of ITPR1 3′ UTR RNA, we performed line-scan imaging of the fluorescent signal in dendrites with NIH ImageJ software (http://rsb.info.nih.gov/ij/). The full width at half-maximum (FWHM) was measured for fluorescent peaks in the line-scan image. We calculated the area enclosed by peak boundaries and an FWHM line as the FWHM region. We defined the peaks that were located within 5 µm distal and proximal to a strong peak as background peaks and calculated the mean FWHM region of the background peaks. A positive fluorescent signal was defined as a granule with an FWHM value more than three times greater than the mean FWHM of the background peaks.
Statistical analysis. Parametric (n > 10) and post hoc tests or non-parametric (n ≤ 10, distribution-free tests) statistical tests were performed, depending on the number of data sets in the study. In the parametric tests, we used ANOVA with a Bonferroni post hoc test to determine significance, depending on the number of treatments. ANOVA was used to test for mean movement distances of transported reporter RNAs. Data distribution was assumed to be normal, but this was not formally tested. For the qRT-PCR, ANOVA with Dunnett’s test was used to determine significance (tested mRNAs versus control PGAM1 mRNA; see Supplementary Table 2). We used the Mann-Whitney U-test to determine statistical significance in tests of the knockdown efficiency of RNAi against HZF or SYNCRIP. All of the data shown are means with s.e.m. No statistical methods were used to predetermine sample sizes, but our sample sizes were similar to those reported in previous studies on mRNA trafficking21 and L-LTP58. The data distribution was assumed to be normal, but this was not formally tested. Data collection and analysis were not performed with blinding to the conditions of the experiments. Data were collected and processed randomly.
A Supplementary methods checklist is available.
51. Watanabe, N. & Mitchison, T.J. Single-molecule speckle analysis of actin filament turnover in lamellipodia. Science 295, 1083–1086 (2002).
52. Yeap, B.B. et al. Novel binding of HuR and poly(C)-binding protein to a conserved UC-rich motif within the 3′-untranslated region of the androgen receptor messenger RNA. J. Biol. Chem. 277, 27183–27192 (2002).
53. Li, Y., Jiang, Z., Chen, H. & Ma, W.J. A modified quantitative EMSA and its application in the study of RNA–protein interactions. J. Biochem. Biophys. Methods 60, 85–96 (2004).
54. Sugimoto, M., Gromley, A. & Sherr, C.J. Hzf, a p53-responsive gene, regulates maintenance of the G2 phase checkpoint induced by DNA damage. Mol. Cell. Biol. 26, 502–512 (2006).
55. Funahashi, Y. et al. ERK2-mediated phosphorylation of Par3 regulates neuronal polarization. J. Neurosci. 33, 13270–13285 (2013).
56. Arimura, N. et al. Anterograde transport of TrkB in axons is mediated by direct interaction with Slp1 and Rab27. Dev. Cell 16, 675–686 (2009).
57. Nguyen, P.V. & Kandel, E.R. A macromolecular synthesis-dependent late phase of long-term potentiation requiring cAMP in the medial perforant pathway of rat hippocampal slices. J. Neurosci. 16, 3189–3198 (1996).
58. Huang, Y.Y. et al. Genetic evidence for a protein-kinase-A-mediated presynaptic component in NMDA-receptor-dependent forms of long-term synaptic potentiation. Proc. Natl. Acad. Sci. USA 102, 9365–9370 (2005).
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.




















