
Differentiation plasticity of stromal vascular fraction cells ...
115
Differentiation plasticity of stromal vascular fraction cells towards adipose tissue and endothelial structures Von der Medizinischen Fakultät der Rheinisch-Westfälischen Technischen Hochschule Aachen zur Erlangung des akademischen Grades einer Doktorin der Medizin genehmigte Dissertation vorgelegt von Melanie Esther Vicky Wosnitza aus Düsseldorf Berichter: Herr Universitätsprofessor Dr.med. Dr.univ.med. Prof. h.c.mult. Norbert Pallua Herr Universitätsprofessor Dr.med. Hans Merk Tag der mündlichen Prüfung: 21. April 2009 Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek online verfüg- bar.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Differentiation plasticity of stromal vascular fraction cells ...

Differentiation plasticity of stromal vascular fraction cells towards adipose tissue
and endothelial structures
Von der Medizinischen Fakultät
der Rheinisch-Westfälischen Technischen Hochschule Aachen
zur Erlangung des akademischen Grades
einer Doktorin der Medizin
genehmigte Dissertation
vorgelegt von
Melanie Esther Vicky Wosnitza
aus
Düsseldorf
Berichter: Herr Universitätsprofessor
Dr.med. Dr.univ.med. Prof. h.c.mult. Norbert Pallua
Herr Universitätsprofessor
Dr.med. Hans Merk
Tag der mündlichen Prüfung: 21. April 2009
Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek online verfüg-
bar.

2
Differentiation plasticity of stromal vascular fraction cells towards
adipose tissue and endothelial structures
Der Medizinischen Fakultät der Rheinisch-Westfälischen Technischen Hochschule
Aachen vorgelegte Dissertation zur Erlangung des akademischen Grades einer Dok-
torin der Medizin
von
Melanie Esther Vicky Wosnitza
aus Düsseldorf

3
CONTENTS
ABBREVIATIONS ............................................................................................. 6
GERMAN SUMMARY ....................................................................................... 8
SUMMARY ...................................................................................................... 11
INTRODUCTION ............................................................................................. 12
Adipose Tissue ....................................................................................................................... 12 Source and location ............................................................................................................ 12
Cellular components ........................................................................................................... 12
Harvesting of adipose tissue derived cells ......................................................................... 13
Miscellaneous populations due to isolation and culture methods ....................................... 14
Adipose derived adult stem cells ............................................................................................ 16 Adipose tissue as source of mesenchymal stem cells ....................................................... 16
Adipose derived adult stem cells with a phenotype of bone marrow stem cells ................. 18
Relevant CD markers and their expression pattern ............................................................ 23
Differentiation capacity towards multiple lineages .............................................................. 23
Mesodermal differentiation ................................................................................................. 27 Adipogenesis ................................................................................................................................ 27 Chondrogenesis ............................................................................................................................ 29 Osteogenesis ................................................................................................................................ 30 Skeletal myogenesis ..................................................................................................................... 31 Cardiomyogenesis ........................................................................................................................ 31 Immune cells ................................................................................................................................. 32
Ectodermal differentiation ................................................................................................... 33 Neurogenesis ................................................................................................................................ 33 Epithelial differentiation ................................................................................................................. 33
Endodermal differentiation ................................................................................................. 34 Hepatogenic and intestinal differentiation ..................................................................................... 34
Vasculogenesis and angiogenesis ......................................................................................... 34 Angiogenesis in adipose tissue .......................................................................................... 35

4
Adipogenesis and angiogenesis ......................................................................................... 35
STUDIES ......................................................................................................... 41
MATERIAL AND METHODS .......................................................................... 44
Technical equipment .............................................................................................................. 44
Reagents ................................................................................................................................ 45
Culture media ......................................................................................................................... 47
Buffers and solutions .............................................................................................................. 49
Isolation of SVF cells .............................................................................................................. 50 CD31 Dynabead separation of SVF cells ........................................................................... 52
CD31 MicroBead separation of SVF cells .......................................................................... 54
Culturing of SVF cells ......................................................................................................... 56
Cell splitting ........................................................................................................................ 57
Fibronectin and MatrigelTM coating ..................................................................................... 58
Human dermal microvascular endothelial cell (HDMEC) isolation ......................................... 58
Human dermal fibroblast isolation .......................................................................................... 59
Human umbilical vein endothelial cell isolation ...................................................................... 59
Flow cytometry analysis ......................................................................................................... 60
Immunostaining with von-Willebrand-factor ........................................................................... 61
Determination of glycerol-3-phosphate dehydrogenase (GPDH) activity ............................... 61
Oil Red O staining .................................................................................................................. 63
Cell counting ........................................................................................................................... 63
Statistical evaluation ............................................................................................................... 63
RESULTS ........................................................................................................ 64
Comparison of excised fat tissue versus liposuction material ................................................ 64
Characterisation of surface markers and morphological changes in SVF cells in culture ...... 67
In vitro differentiation of unpurified SVF cells under endothelial medium conditions ............. 68
Splitting and purity analysis of CD31- and CD31+ SVF cells .................................................. 70

5
Differentiation potential of CD31- and CD31+ SVF cells towards EC lineage ......................... 72
In vitro differentiation of SVF cells under adipogenic medium conditions .............................. 76
DISCUSSION .................................................................................................. 79
Tissue engineering with adipose derived adult stem cells ..................................................... 80 Present problems and hurdles............................................................................................ 80
How to manage insufficient vascularisation ........................................................................ 81
Characterization of the SVF ................................................................................................... 81
ADAS cell differentiation towards endothelial cells ................................................................ 84 Therapeutic angiogenesis .................................................................................................. 85
Endothelial cell differentiation to adipocytes ....................................................................... 88
Arteriosclerosis and adipose tissue ........................................................................................ 90 The impact of progenitor cells in atherosclerosis ............................................................... 94
CONCLUSION ................................................................................................. 95
REFERENCES ................................................................................................ 96
PUBLICATIONS ............................................................................................ 110
Full papers ............................................................................................................................ 110
Published abstracts .............................................................................................................. 110
Scientific presentations ........................................................................................................ 110
ACKNOWLEDGEMENT................................................................................ 112
ERKLÄRUNG § 5 ABS. 1 ZUR DATENAUFBEWARUNG .......................... 113
CURRICULUM VITAE ................................................................................... 114

6
ABBREVIATIONS
1,25 (OH)2D3 1alpha,25-dihydroxyvitamin D3 IgG Immunoglobulin
acLDL Acetylated low-density lipoprotein IL-3 Interleukin-3
ACLP Aortic carboxypeptidase-like protein IL-6 Interleukin-6
ADAS cell Adipose-derived adult stem cell IMDM Isocove’s Modified Dulbecco’s Medium
Ang Angiopoietin KCl Kalium chloride
aP2 Adipocyte fatty acid binding protein KDR Kinase insert domain receptor
APC Allophycocyanin KHCO3 Potassiumhydrogencarbonate Kit
AT Adipose tissue LDL Low-density lipoprotein
ATP Adenosintriphosphate LPL Lipoprotein lipase
ATRA All-trans retinoic acid Mac-1 Macrophage antigen-1/CD11b
BAT Brown adipose tissue MAP2ab Microtubule associated protein 2 a/ b
bFGF Basic fibroblast growth factor MCP-1 Monocyte chemotactic protein-1
BMP-2 Bone morphogenetic protein-2 M-CSF Macrophage colony-stimulating factor
BMSC Bone marrow stem cell MMP Matrix metalloproteinases
BSA Bovine serum albumin MOMA-2 Anti-macrophage monoclonal antibody-2
CaCl2 Calcium chloride MS Metabolic syndrome
CD Cluster of differentiation MSC Mesenchymal stem cell
C/EBP CCAAT/enhancer binding protein Myf Myogenic factor
C/EBP-α CCAAT/enhancer binding protein alfa MyoD1 Myogenic differentiation 1
CNS Central nervous system NaCl Natrium chloride
CO2 Carbone dioxide NAD Atrial natriuretic peptide
CYP Cytochrome P450 NaHCO3 Sodium bicarbonate
DAPI 4’,6’-Diamidin-2’-phenylindol-dihydrochlorid Neu N Neuronal nuclear protein
DiI 1,1'-dioctadecyl-3,3,3',3’-tetramethylindocarbocyanine
perchlorate
NH4Cl Ammonium chloride
DMEM Dulbecco’s Modified Eagle Medium NMDA N-methyl-D-aspartate
DMSO Dimethylsulfoxid NF Neurofilament
EC Endothelial cell NO Nitric oxide
ECGM Endothelial Cell Growth Medium PA Plasminogen activator
ECM Extracellular matrix PAI-1 Plasminogen activator inhibitor 1
ED2 Anti perivascular monocytes/macrophages antibody PBS Phosphate buffered saline
EDTA Ethylene diamine tetraacetic acid PD Population doublings
EGF Epithelial growth factor PDGF Platelet-derived growth factor
EGM-2 (MV) Endothelial Growth Medium 2 (Microvascular) PE Phycoerythrin
eNOS Endothelial nitric oxide synthetase PECAM-1 Platelet endothelial cell adhesion molecule 1
EPC Endothelial progenitor cell PFA Paraformaldehyde
F12 Ham’s F12 PFF Pulsating fluid flow
FA Fatty acid PLGA Poly(lactic-co-glycolic acid)
FACS Fluorescence activated cell sorting PPARγ Pperoxisome proliferator-activated receptor gamma
FCS Fetal calf serum PRELP Proline/arginie-rich end leucine-rich repeat protein
FGF Fibroblast growth factor PTH Parathyroid hormone
FITC Fluorescein-Isothiocyanat RBM Reconstituted basement membrane
Flk-1 Fetal liver kinase 1 SCC Sideward scatter
Flt-1 Fms like tyrosine kinase 1 SCD1 Stearoyl-CoA desaturase
FSC Forward scatter SCF Stem cell factor

7
GABA Gamma-amino butyric acid SCGF-β Stem cell growth factor beta
GFAP Glial fibrillary acidic protein SD Standard deviation
Glut4 Glucose transporter 4 SM Smooth muscle
GPDH Glycerol-3-phosphate dehydrogenase SMC Smooth muscle cell
Gy Gray SSC Sideward scatter
HDL High density lipoprotein SVF Stromal vascular fraction
HDMEC Human dermal microvascular endothelial cell TE Tissue engineering
HEPES N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid TGF-β Transforming growth factor beta
hATSC Human adipose tissue stem cell TIMI-3 Thrombolysis in Myocardial Infarction 3
hMADS cell Human multipotent adipose-derived stem cell TIMP Tissue inhibitor matrix metalloproteinases
hTERT Human telomerase TNF-α Tumor necrosis factor alfa
HMG-CoA 3-hydroxy-3-methylglutaryl coenzyme A VCAM-1 Vascular cell adhesion molecule 1
HNF4α Hepatocyte nuclear factor 4 alfa VE-cadherin Vascular endothelial cadherin
hS Human autologous serum VEGF Vascular endothelial growth factor
HSC Hematopoietic stem cell VEGF-R1 Vascular endothelial growth factor receptor 1
HUVEC Human umbilical vein endothelial cell VEGF-R2 Vascular endothelial growth factor receptor 2
IBMX Isobutylmethylxantine vWF Von-Willebrand factor
ICAM-1 Intercellular adhesion molecule 1 WAT White adipose tissue
IGF-1 Insulin-like growth factor 1

8
GERMAN SUMMARY In den letzten Jahrzehnten hat sich gezeigt, dass Fettgewebe weitaus mehr darstellt als ein
simples Energiespeicherorgan. Es besteht neben reifen Fettzellen, sog. Adipozyten, aus
Fettvorläuferzellen, auch Präadipozyten genannt. Präadipozyten gehören zu den mesen-
chymalen Stammzellen des Körpers und konnten das erste Mal vor mehr als dreißig Jahren
aus humanem menschlichem Fettgewebe isoliert werden. Seither sind die Vorläuferzellen im
Fettgewebe mehr und mehr in das Interesse der Forschung gerückt. Es ließ sich zeigen,
dass Präadipozyten keine homogene Zellpopulation darstellen, wie anfangs geglaubt, son-
dern ganz im Gegenteil ein heterogener Mix aus verschiedenen Zelltypen sind. Die Hetero-
genität ist eine Konsequenz der Methode zur Gewinnung von Fettvorläuferzellen, nämlich
Fettgewebsexzision oder Liposuktion. Sowohl das Herausschneiden als auch das Absaugen
von Fett beinhaltet die Gewinnung eines ganzen Gemisches von stromalen Zellen, nicht nur
selektiv von Fettvorläuferzellen. Die isolierten Zellen sind neben den klassischen Präadipo-
zyten auch Endothelzellen, endotheliale glatte Muskelzellen, Fibroblasten, Makrophagen und
Blutzellen; daher spricht man auch allumfassend von der stromalen vaskulären Fraktion
(SVF). Es hat sich gezeigt, dass sich SVF Zellen nicht nur zu reifen Adipozyten differenzie-
ren lassen, sondern auch in viele andere mesenchymale sowie nicht-mesenchymale Zellty-
pen. Die SVF des Fettgewebes gehört zu den Zellpopulationen, die sehr schwer zu charakte-
risieren sind. Daher ist die Terminologie auch nicht einheitlich. Sehr häufig finden sich neben
dem Begriff adipose SVF Bezeichnungen wie human adipose tissue-derived stem cells oder
adipose lineage cells. Ihre außerordentliche Differenzierungskapazität und ihre Oberflä-
chenmerkmale unterstreichen die Ähnlichkeit von SVF Zellen zu Knochenmarkstammzellen.
Aufgrund ihrer breiten Differenzierungskapazität in meso-, ekto- und endodermale Zelltypen
können die fettgeweblichen SVF Zellen als multipotente Stammzellen betrachtet werden.
Neueste Forschungsergebnisse legen nahe, dass Fettgewebszellen und Endothelzellen ei-
nen gemeinsamen Zellvorläufer haben könnten. Dies wird durch die enge Beziehung zwi-
schen Angiogenese und der Entstehung von Fettgewebe unterstrichen. Angiogenese und
Adipogenese beeinflussen sich gegenseitig; so verbessert die Interaktion zwischen Endo-
thelzellen und Adipozyten das Wachstum beider Zelltypen und bewirkt gleichzeitig eine Re-
duktion der Apoptoserate. In Hinblick auf eine gemeinsame Vorläuferzelle bleibt die For-
schung allerdings noch entscheidende Erklärungen schuldig, insbesondere der Verbin-
dungsweg zwischen Fettgewebe und Endothel, sowie das Differenzierungspotential der Zel-
len in die eine oder andere Richtung über eine gemeinsame Vorläuferzelle konnten noch
nicht demonstriert werden. Diese Themen waren daher ein zentraler Fokus dieser Arbeit.

9
SVF Zellen wurden aus dem Fettgewebe isoliert und ihre Oberflächeneigenschaften, das CD
(cluster of differentiation) Profil, untersucht. Für die Isolation von SVF Zellen kommen zwei
Methoden in Frage, zum einen die Fettgewebsexzision mit anschließender Präparation der
gewonnenen Fettgewebsläppchen oder die Liposuktion, bei der die anschließende Gewebs-
präparation entfällt. Um beide Methoden hinsichtlich ihrer Unterschiede in der Zellgewinnung
zu vergleichen, wurden die Oberflächeneigenschaften von SVF Zellen aus exzidiertem Ge-
webe und aus liposuktioniertem Gewebe mittels Durchflusszytometrie bestimmt. Hierbei
wurde ein besonderes Augenmerk auf endotheliale Marker sowie Stammzellmarker gelegt,
um eventuelle Kontaminationen mit anderen Zelltypen aufzudecken. Des Weiteren wurde die
Expression der CD Marker während einer Zellkulturphase untersucht, um Veränderungen
der Zelloberflächeneigenschaften im Kultivierungsverlauf darzustellen. Dies ist insbesondere
von Bedeutung, wenn SVF Zellen zur Zellvermehrung über mehrere Passagen kultiviert wer-
den und erst dann für Differenzierungsexperimente verwendet werden sollen. Das zelluläre
CD Profil wurde neben der Durchflusszytometrie mit Hilfe von Fluoreszenzfärbungen be-
stimmt. Als endotheliale Marker wurden hauptsächlich CD31, vWF und CD105 verwendet,
als Stammzellmarker CD34, KDR und S100. Weiterhin wurde die fettgewebliche SVF mit
Hilfe von magnetischen Beads (Dynabeads und MicroBeads) in eine CD31+ und eine CD31-
Fraktion aufgeteilt und die Differenzierungskapazität der jeweiligen CD31-Gruppe in Rich-
tung Fettgewebe sowie Endothel parallel untersucht. Die Differenzierung zu Endothel wurde
mit Hilfe verschiedener Endothelzellmedien und der Kultur auf Extrazellularmatrix (Matri-
gelTM) durchgeführt. Hier wurde das Augenmerk insbesondere auf die Ausbildung kapillärer
Strukturen auf MatrigelTM gelegt, zudem wurden die zellulären Oberflächeneigenschaften mit
Hilfe der Durchflusszytometrie bestimmt. Die Fettzelldifferenzierung der CD31- und CD31+
SVF Zellen erfolgte mittels Kultivierung in einem Fettzelldifferenzierungsmedium über insge-
samt 21 Tage. Die Differenzierung wurde mit dem Glycerol-3-Phosphat Dehydrogenase
(GPDH) Enzymassay sowie durch Öl-Rot Färbung und Zellzählung validiert.
Frisch isolierte SVF Zellen aus exzidiertem Fettgewebe zeigen signifikante Unterschiede in
ihrer Oberflächenbeschaffenheit im Vergleich zu Zellen aus Liposuktionsgewebe. SVF Zellen
aus exzidiertem Fettgewebe sind in 5-10% positiv für CD31, die Zellen aus Liposuktionsge-
webe zeigen hingegen einen deutlich höheren Anteil an CD31 positiven Zellen von bis zu
40%. Ähnliches gilt für KDR. Beide Zellpopulationen haben einen vergleichbaren, deutlich
positiven Nachweis von CD34. Im Verlauf kam es bei beiden Fraktionen nach 14 Tagen in
der Zellkultur zu einem Abfall der Stammzellmarker (CD34, KDR, S100). Um das endothelia-
le Differenzierungspotential von CD31- SVF Zellen zu evaluieren, wurden die Zellen auf Mat-
rigelTM Extrazellularmatrix in Endothelzelldifferenzierungsmedium kultiviert. Hier zeigten die
CD31- Zellen eine ausgeprägte Tendenz, dreidimensionale, tubuläre Strukturen im Sinne

10
eines kapillären Netzwerkes auszubilden. Die Ergebnisse wurden durch die Durchflusszyto-
metrie bestätigt. Frisch isolierte SVF Zellen ohne weitere Selektierung differenzieren in vitro
zu ca. 90% zu reifen Adipozyten. Hier wurden CD31- und CD31+ SVF Zellen im Fettzelldiffe-
renzierungsmedium kultiviert und die Ausbeute der reifen Fettzellen bestimmt. Die CD31-
Zellfraktion differenziert zu fast 100% zu Adipozyten. Interessanterweise sind auch die
CD31+ SVF Zellen in der Lage eine Fettzelldifferenzierung zu vollziehen, hier zeigten ca.
65% der Zellen Lipidvakuolen. Die Bestimmung der GPDH Aktivität ergab ebenfalls einen
Wert von 64% ± 11%, verglichen mit der GPDH Aktivität der CD31- Zellen. Während der Dif-
ferenzierung zu reifen Adipozyten trugen die CD31+ SVF Zellen weiterhin die CD31-
Dynabeads auf der Zelloberfläche.
Die frisch isolierten SVF Zellen des Fettgewebes zeigen unabhängig von der Isolationsme-
thode eine starke Expression von Stammzellmarkern. Die SVF enthält jedoch auch eine Po-
pulation von CD31+ Zellen, insbesondere in Liposuktionsgewebe. Dies weist auf eine hohe
Kontaminationsrate mit Endothelzellen in liposuktioniertem Fettgewebe im Vergleich zu exzi-
diertem Gewebe hin. Insgesamt fallen die Stammzellmarker während der Kultivierung deut-
lich ab, was wiederum in Übereinstimmung mit der Erfahrung ist, dass Präadipozyten ihre
Differenzierungskapazität nach mehreren Passagen verlieren.
Die Ergebnisse der endothelialen und adipogenen Differenzierung belegen, dass die fettge-
webliche SVF einen CD31- Zelltypen enthält, der zu Adipozyten sowie Endothelzellen diffe-
renziert werden kann. Zudem gibt es CD31+ Zellen, die, obwohl sie einen endothelialen Phä-
notyp besitzen, in reife Fettzellen differenziert werden können. Diese Resultate demonstrie-
ren die Fähigkeit der SVF Zellen sich in beide Richtungen, und zwar Adipozyt sowie Endo-
thelzelle, differenzieren zu können. Allgemein gesprochen zeigen die Ergebnisse die Plastizi-
tät ausgereifter, mesenchymaler Zellen aus dem Fettgewebe, eine Differenzierung zwischen
Endothel und Fettgewebe sowie auch umgekehrt vollziehen zu können.
Des Weiteren könnten diese Resultate von Relevanz für die Pathogenese der Arteriosklero-
se sein. Hier scheinen Transdifferenzierungsprozesse endothelialer Vorläuferzellen eine Rol-
le zu spielen.

11
SUMMARY Recent research findings postulate that adipocytes and endothelial cells (ECs) may share a
common progenitor. However, the interlinking pathways between adipose tissue and endo-
thelium, and the differentiation potential of cells to convert from one tissue into the other via
progenitor cells, have not been elucidated so far and are therefore the focus of this thesis.
Stromal vascular fraction (SVF) cells were isolated from liposuction aspirates or excised adi-
pose tissue and separated into CD31+ and CD31- populations by magnet-assisted cell sort-
ing. Differentiation to fat tissue was induced in both CD31 fractions. Differentiation was as-
sayed enzymatically and by cell counting. Maturation to endothelium was performed with
vascular endothelial growth factor, insulin like growth factor-1 plus 2% FCS and confirmed by
flow cytometry and tube formation assays on Matrigel™. The results presented here show
that the SVF contains a CD31-, S100+ cell type which can differentiate into adipocytes and
EC. The SVF also comprises CD31+ cells that, although they have an endothelial phenotype,
can be converted into mature adipocytes. These findings demonstrate the potency of SVF
cells to perform both adipogenic and endothelial differentiation. Further, they reveal the plas-
ticity of mature cells of mesenchymal origin to undergo conversion from endothelium to adi-
pose tissue and vice versa.

INTRODUCTION
ADIPOSE TISSUE
SOURCE AND LOCATION
In the last decades the belief was corroborated that adipose tissue is much more than a sim-
ple fat storage. Adipose tissue plays a central role in energy homeostasis and was found to
be a highly active metabolic organ335. Many secretory factors produced by fat cells influence
the endocrine regulation of the body, affecting growth, metabolism and behaviour. Fat tissue
is a highly specialized connective tissue located in different places of the body and displaying
various functions. In humans, white adipose tissue composes as much as 20% of the body
weight in men and 25% of the body weight in women. A major portion is the subcutaneous
fat tissue located directly underneath the dermis. Furthermore, adipose tissue is found in
intraabdominal locations, as omental fat pads and surrounding several organs of the human
body. Dependent on the location there are two different types of fat: white and brown adipose
tissue. Both cushion and insulate the body, but each of them has further specialized func-
tions as well. White adipose tissue (WAT) is mainly found in the subcutaneous regions, but
also in the Omentum Majus, lying intraabdominally between the abdominal muscles and the
visceral organs. WAT basically functions as energy source for the body, whereas brown adi-
pose tissue (BAT) displays the heat source, primarily at young age. BAT is surrounding
nearly all organs in the abdomen as well as in the chest. During the natural aging process
brown fat tissue is gradually replaced by white adipose tissue.
CELLULAR COMPONENTS
The primary cellular component of adipose tissue is a collection of so called adipocytes, lipid
filled fat cells. Focussing on the two different types of fat tissue, morphological and ultrastruc-
tural properties of brown and white adipocytes reflect the different functions in thermoregula-
tion and energy balance. White adipocytes contain only a single large fat vacuole, whereas
brown fat cells consist of several smaller vacuoles and a much higher number of mitochon-
dria76. Brown and white adipose tissue are both innervated by the sympathetic nervous sys-
tem, which controls metabolic and lipolytic activity as well as the vascularisation in adipo-
cytes18,19.
White mature adipocytes consist of about 90% lipids303 and each of them stands in close
contact with at least one capillary, providing a vascular network that allows continued growth
of the organ304. A further component located in the adipose organ is the stromal vascular
fraction (SVF), stromal cells which are mainly located around the blood vessels. This adipose

13
SVF contains a large population of preadipocytes, first identified by Poznanski et al. in 1971
following the enzymatic digestion of adipose tissue282,317. Preadipocytes were recently identi-
fied to have major properties of adult mesenchymal stem cells. These precursor cells are
present troughout adult life, and differentiation into new mature adipocytes can still be
achieved in old animals211,269.
HARVESTING OF ADIPOSE TISSUE DERIVED CELLS
A number of factors have to be taken into consideration when isolating preadipocytes or SVF
cells from adipose tissue. Overall cell yield is determined by donor age, obesity, the anatomi-
cal site of harvest, the methods of tissue harvest, storage and preadipocyte isolation and the
in vitro culture environment (for review158). Besides the isolation method of preadipocyte that
defines the cell numbers harvested, the donor side exhibits a major factor for the adipogenic
capacity of preadipocytes. In general human abdominal subcutaneous preadipocytes seem
to have a greater potential for adipogenesis as omental or intraabdominal preadipo-
cytes150,265,384, however, other authors state that there is no difference in adipogenic potential
between both locations352,399. In subcutaneous locations abdominal preadipocytes have a
greater potential to convert into mature adipocytes than femoral preadipocytes146. Addition-
ally proliferation capacity in subcutaneous preadipocytes is superior to those of omental cells
since subcutaneous cells proliferate faster under in vitro conditions399. Additionally the donor
age plays an important role in proliferation rates of harvested cells where a negative correla-
tion can be observed between age and doubling capacity33,399.
To release the broad spectrum of different cell types harboured in adipose tissue, minced fat
lobules from excised tissue or plain lipoaspirate are used. The material is washed after har-
vesting to remove possible blood contamination. The extracellular matrix (ECM) holding the
adipocytes in place is digested with collagenase solution, followed by centrifugation to divide
mature fat cells from stromal cells. The cells located in the stromal fraction mainly appear in
a fibroblast-like morphology after adherence to cell culture dishes, macroscopically indistin-
guishable from factual fibroblasts. At the beginning of fat tissue research these cells were
referred to as preadipocytes or adipoblasts5,52,130, as they were found to give rise to fat vacu-
ole-containing cells. Both terms refer to fat cell precursors, but adipoblasts were regarded to
be the juvenile adipose stem cell, contrariwise preadipocytes as the cell type in between adi-
poblast and adipocyte5. However this could not be asserted. Instead, the term preadipocyte
was established over the last three decades and is still frequently used in the literature for
precursor cells derived from fat tissue65,79,92,118,140,158,160,200,350,407.
Preadipocytes are mainly located in the surrounding of small blood vessels and between
mature fat cells in the stromal vascular fraction of fat tissue260,288,290 and can be recruited a
lifelong6. Nevertheless, harvested adipose stromal cells display a heterogeneous cell popula-

14
tion directly after isolation and are not as homogenous as initially believed. Regarding the
isolation method of adipose stromal cells it is likely that not solely preadipocytes are digested
from the ECM but also cells located in the blood vessels and the vessel walls. As a result the
adipose SVF not exclusively comprises preadipocytes but also microvascular endothelial
cells, endothelial smooth muscle cells, fibroblasts, macrophages, and blood cells (Figure
1)427,434.
SVFSVF
preadipocytes
endothelialcells
smooth musclecells
fibroblasts
macrophages
bloodcells
Major progress was made in the last few years concerning the nomenclature of the adipose
stromal cells due to newly discovered cell features and cell characteristics158,314,433. At the
beginning of fat tissue research the differentiation process from preadipocyte to adipocyte
was believed to be an irreversible process, with mature fat cells as an end-stage product. In
vitro studies, however, have demonstrated that the phenomenon of differentiation seems to
be reversible. It has been proven in cell culture experiments that dedifferentiation of adipo-
cytes back into preadipocytes is possible281,334,430. In addition, “preadipocytes” have been
found to be able to differentiate not only into fat cells but also into many other mesenchymal
and non-mesenchymal cell lineages55,382,433. Further research revealed that adipose stromal
cells have surface characteristics comparable to bone-marrow stem cells (compare Table
1)133. Taking their remarkable differentiation potential433, these cells can be regarded as adult
mesenchymal stem cells capable of multipotency372,388,433.
MISCELLANEOUS POPULATIONS DUE TO ISOLATION AND CULTURE METHODS
Although intensive research has been performed on fat tissue progenitor cells, the adipose
SVF remains a cell population very hard to characterize. Consequently, in recent literature
Figure 1. Adipose stromal
vascular fraction (SVF)
Freshly isolated SVF cells
from fat tissue display a het-
erogeneous cell population
comprising contrasting cell
types, such as preadipocytes,
endothelial cells, smooth mus-
cle cells, fibroblasts, blood cells
and macrophages.

15
many different terms refer to this cell mixture isolated from adipose tissue by collagenase
digestion. Most often these cells are called preadipocytes, but adipose tissue-derived stem
cells113,260, adipose-derived adult stem cells23, adipose tissue mesenchymal stem cells244,382,
multi-lineage cells from adipose tissue98, adipose tissue-derived stromal cells109,133, or adi-
pose lineage cells314 have been used to describe this population as well. Since different do-
nor sites such as abdominal, gluteal, mammary or intraabdominal fat tissue are applied for
SVF harvesting, nonuniform populations of preadipocytes are often used for cell surface
characterisation and differentiation assays. Consequently, literature results vary widely re-
garding the surface protein expression and the differentiation plasticity of adipose stromal
cells.
Two miscellaneous procedures are described for primary isolation of SVF cells, liposuction
and plain excision during procedures, such as abdominoplasty or breast reduction. It remains
controversial whether liposuction aspirate or excised adipose tissue displays the superior
source for preadipocyte harvesting. Some studies state that liposuction aspirates represent a
better source compared to excised fat405, others claim the contrary176. After excision of adi-
pose tissue (AT), fat lobules need to be isolated by removing capillaries and connective tis-
sue, a possible contamination source for endothelial cells and fibroblasts. Liposuction is per-
formed by manually applying negative pressure using a 10-cc-syringe with a blunt tip
cannula80. Due to the harvesting procedure the isolation by liposuction damages not only AT
but also the surrounding structures including capillaries and fibrous tissue. Therefore minced
fat tissue from lipoaspirates contains vessels and connective tissue besides the fat compart-
ment.
The adipose SVF harbours not only preadipocytes, but also other mature cell types, e.g. en-
dothelial cells (CD31+/CD34+), macrophages (CD14+/CD31+), and blood cells (see Figure 1).
Certain cell fractions can be isolated with magnet-assisted cell sorting due to the expression
of surface proteins including stem cell markers like CD34260 or kinase insert domain receptor
(KDR)60,252, macrophage markers as CD14350, and fibroblast markers. This cell sorting can
assist in creating a more pure and homogenous population of SVF cells by using a negative
selection method to gain the non-labelled cells, the classical preadipocytes.
Rodriguez et al.331 established a method to isolate two distinguishable cell populations from
the adipose SVF based on adhesion characteristics. Cells either displayed a fast- or a slow-
adherence one. Both populations exhibited similar characteristics concerning doubling time
(36h) and differentiation potential towards adipocytes and osteoblasts for up to 60-80 pas-
sages. After this growth period slow-adherent SVF cells ceased to proliferate and also to
differentiate. In contrary fast-adherent SVF cells exhibited a slow proliferation rate over time
(72h) and expressed significant levels of telomerase activity, whereas slow-adherent SVF
cells did not. Fast-adherent SVF cells responded mainly to basic fibroblast growth factor

16
(bFGF) with expansion, but not to epithelial growth factor (EGF) or platelet-derived growth
factor (PDGF) and could be cultured for more than 200 passages. This fast-adherent cell
type from the adipose SVF was termed human multipotent adipose-derived stem cells
(hMADS cells)332.
ADIPOSE DERIVED ADULT STEM CELLS
ADIPOSE TISSUE AS SOURCE OF MESENCHYMAL STEM CELLS
Stem cells are defined as being qualified to distribute renewal cells for a particular mature
cell type29. Three miscellaneous aspects delineate this special cell fraction and underline
their uniqueness and difference from all other cell populations harboured in the human body.
To begin with, stem cells are capable to replicate themselves leading to self-renewal and an
extensive proliferation capacity. Second, stem cells display an immature and unspecialized
cell type, which indicates that they do not have any tissue specificity and therefore lack spe-
cialized, tissue-specific functions. Thirdly, stem cells are able to differentiate into at least one
specialized cell type. How many different cell phenotypes a stem cell can convert into, known
as stem cell plasticity or potency, can be outlined with different stem cell-defining terms.
Stem cells may be categorized as either totipotent, pluripotent, or multipotent, whereby the
stem cell is able to form all, most, or a small number of miscellaneous cells of the body. Fur-
ther, stem cells capable of producing the blood cells of an organism are defined as hemato-
poietic stem cells.
Several sources of stem cells were established in the past to harvest multipotent stem cells
from lineages of the three different germinal sheets (Figure 2). According to the embryonal
sheet and tissue location, stem cells can be classified as mesodermal, ectodermal or endo-
dermal412. Stem cells exist in humans throughout adult life in special stem cell niches418. Fur-
ther stem cells can be harvested as embryonic stem cells from the inner cell mass of the pre-
implantation blastocyst. Embryonic stem cells were isolated from either human, non-human
primates or mice from 3-5 days old embryos43,403. Still capable of pluripotency, these cells
have the capacity to undergo nearly the full range of differentiation pathways, including all
three germinal lines264,318,343,359,416. Differentiation phenotypes include adipocytes, endothelial
cells, chondrocytes, muscle cells, neuronal cells as well as blood cells and several others
(Figure 2)43,165,236,403. However, major concerns regarding ethical topics in application of em-
bryonic stem cells in research, restrict a wider range of promising clinical usage and trials. By
applying adult stem cells instead, most of these ethical problems can be avoided.

17
Epiblast-like pluripotent SC
Endodermal SCMesodermal SCEctodermal SC
LiverSC
PancreaticSC
IntestinalSC
MesenchymalSC
Hemangioblast
Vascular SCHematopoieticSC
SkinSC
NeuralSC
Epithelial cellMelanocyteHair follicle
NeuronsAstrocyte
Oligo-dendrocyte
LymphoidProgenitor
cell
MyeloidProgenitor
cell
EndothelialProgenitor
cell
EndothelialCell
Pericyte
PreadipocyteFibroblastOsteoblast
ChondroblastMyoblast
Cardiomyoblast
AdipocyteFibrocyte
Smooth muscle cellOsteocyte
ChondrocyteMyocyte
Cardiomyocyte
Hepatocyte
Islet cell
Intestinalcell
MonocyteMakrophageGranulocyteErythrocyte
PlateletDendritic
cell
T-lymphocyteB-lymphocyte
NK cell
Adult stem cells, often termed mesenchymal or somatic stem cells, are mature, undifferenti-
ated cells which retained their unique capacity to differentiate into a wide spectrum of differ-
ent tissue types. They display a reserve of fibroblast-like cells that lie secluded in mature
tissue yet can home to and repair damaged tissue when necessary219. Therefore these cells
display an interesting source for multiple clinical applications135,258,259. Since most mesen-
chymal stem cells (MSCs) can be easily isolated, cultured and expanded, application for a
cell-based therapy is highly attractive, e.g. the treatment of children with osteogenesis imper-
fecta168, patients with Alzheimer’s37,375,376, Parkinson’s191,394, and Crohn´s disease122,
Duchenne muscular dystrophy332, burns321,357, cerebral ischemia193, spinal cord injuries196,
hematopoietic recovery214, heart disease131,170,373, osteoarthritis105,327 and others.
Mesenchymal stem cells preserve their ablility to differentiate into mature specialized cell
phenotypes28,30,38,135,164,294,312,403 and perform self-renewal combined with maintaining their
Figure 2. Postnatal stem and progenitor cells
Differentiation cascade of stem and progenitor cells from mesoderm, ectoderm and endoderm.

18
specific stem cell plasticity and phenotype for several generations218. Adult stem cells com-
prise bone marrow stromal cells (BMSCs), hematopoietic stem cells (HSCs), endothelial pro-
genitor cells, as well as neural, dermal, hepatic and adipose stem cells, among others.
Located in distinct stem cell niches in divergent tissues and organs, adult stem cells can be
found all over the human body. One of the best known niches exists in bone marrow, har-
bouring HSCs as well as MSCs185. Additiononally multiple other sources for MSCs exist in
the human body, including trabecular bone285, skeletal muscle428, dermis428, synovium344,
deciduous teeth395, lung, liver, intestine, pancreas, heart35, central nervous system (CNS)
tissue, human umbilical cord blood106,209, human placenta178, perivascular cells346 as well as
adipose tissue209.
Adipose tissue comprises a stromal vascular fraction comparable to the stromal fraction of
bone marrow. SVF cells from adipose tissue have been identified as mesenchymal cells,
being directly derived from mesenchyme, just like bone marrow433. Stromal cells from fat tis-
sue include adult mesenchymal stem cells, often referred to as preadipocytes, and in addi-
tion various other cell types as mentioned earlier98,113,434. Under stem cell supporting culture
conditions, the SVF turns into a more homogeneous population, termed human adipose-
derived adult stem (ADAS) cells. ADAS cells have nearly equal morphology and characteris-
tics as BMSCs. Most notably they demonstrate extensive expansion potential, plasticity to
undergo multilineage differentiation and surface protein phenotype. ADAS cells are therefore
an alternative source for mesenchymal stem cells96,113,133,186,233,304,326,332. Additionally an ideal
source of stem cells should be abundant, accessible and readily isolated with minimal risk to
the patient. ADAS cells seem to outfit precisely this assortment of specifications.
ADIPOSE DERIVED ADULT STEM CELLS WITH A PHENOTYPE OF BONE MARROW
STEM CELLS
The best known origin for mesenchymal stem cells is the stroma of bone marrow which con-
tains adult stem cells, the BMSCs. ADAS cells have been identified as mesenchymal cells as
they are located in the stroma of adipose tissue, which is derived from mesenchyme much
like bone marrow. Located between mature adipocytes both stem cell populations (ADAS
cells and BMSCs) are harboured in a highly vascularised tissue with close contact to blood
circulation.
Not surprisingly ADAS cells exhibit a differentiation repertoire similar to that described for
bone marrow stem cells. The cells are multipotent adult stem cells and have the ability for
multilineage differentiation429,433,434 comprising the mesenchymal and non-mesenchymal
branches. When cultured in vitro, ADAS cells as well as BMSCs show a fibroblast-like mor-
phology after adhering to the culture flask. Both cell types can be easily cultured and ex-
panded in vitro. Further similarities between ADAS cells and BMSCs have been described in

19
the literature focussing on growth kinetics and expansion capacity, cell senescence, gene
transduction efficiency, gene transcription and protein surface expression.
Noted exceptions comprise the presence of CD49d as well as the low expression of CD133,
the lack of telomerase activity and absence of STRO-1 and CD62L on ADAS cells (Table 1).
CD49d, STRO-1 and CD62L are thought to be molecules involved in cell homing to bone
marrow or adipose tissue compartments. STRO-1 is expressed by stromal elements in hu-
man bone marrow and valuable for identification and functional characterisation of human
bone marrow stromal cell precursors134. Both cell types are stromal populations isolated from
fatty compartments based on their ability to adhere to plastic.
It remains questionable if CD106, CD117 and ABCG2 are expressed on ADAS cells due to
diverse reports in the literature (Table 1).
In contrary to BMSCs lack ADAS cells the natural activity of telomerase but by transduction
with a retrovirus containing the gene for the catalytic subunit of human telomerase (hTERT)
stable clones can be produced and isolated188. The tranduced clones (hATSC-TERTs) have
high telomerase activity, which could be maintained for more than 100 population doublings
(PD). hATSC-TERTs had a normal karyotype and replicated constantly, whereas control
cells underwent senescence-associated proliferation arrest after 36-40 PD. Supplementary
hATSC-TERTs revealed an increased bone-forming capacity188. These results further sup-
port a similarity between BMSCs and hATSCs as studies with BMSCs demonstrated that
telomerase expression not only extends the proliferative life-span but also maintains the os-
teogenic differentiation potential of these stem cells.
The following table summarizes the surface expression patterns of CD markers for ADAS
cells and BMSCs.
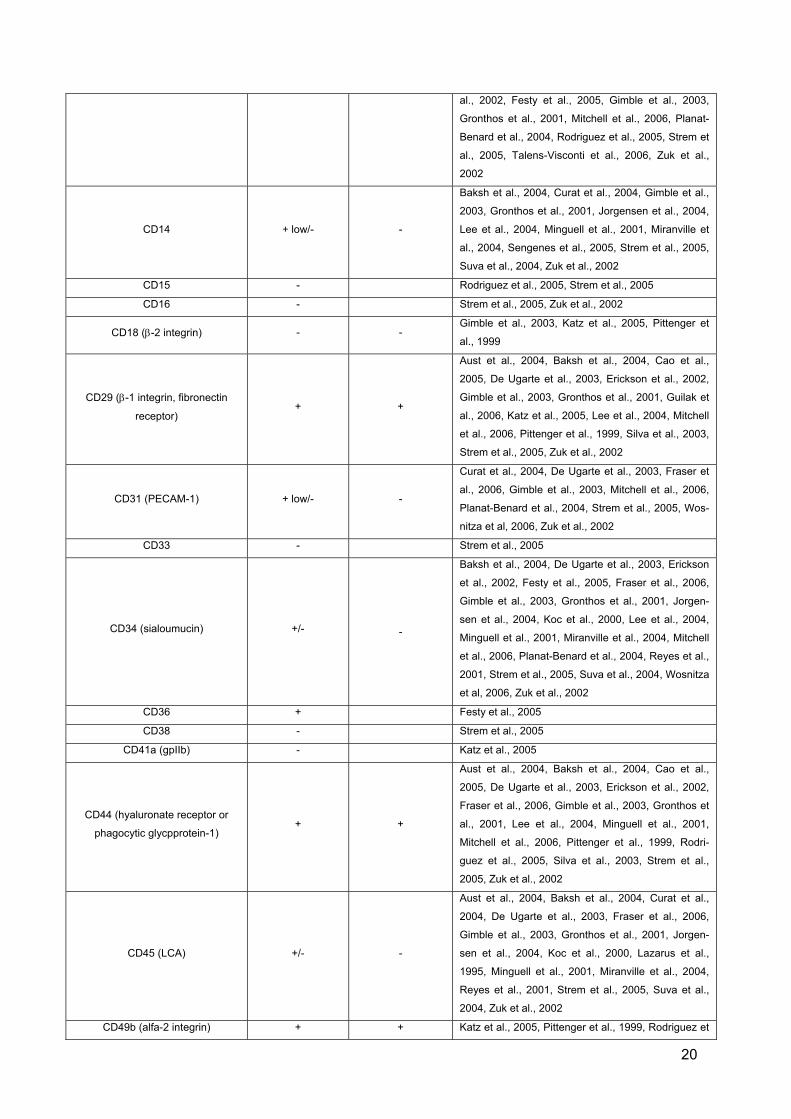
Table 1. Characterization of adipose tissue stem cells compared to bone marrow stem cells
Cluster of Differentiation Surface Expression
Adipose Tissue Stem Cells
BoneMarrow
Stem Cells Reference
CD3 - Strem et al. 2005
CD4 (MHC class II co-receptor; T4) - - Katz et al., 2005, Pittenger et al., 1999
CD8 (MHC class I co-receptor; T8) - Katz et al., 2005
CD9 (tetraspan) + +
De Ugarte et al., 2003, Erickson et al., 2002, Gimble
et al., 2003, Gronthos et al., 2001, Guilak et al.,
2006, Strem et al., 2005, Zuk et al., 2002
CD10 (CALLA) + +
Aust et al., 2004, De Ugarte et al., 2003, Erickson et
al., 2002, Festy et al., 2005, Gimble et al., 2003,
Gronthos et al., 2001, Strem et al., 2005, Zuk et al,
2002,
CD11 (alfa-integrin) - +/-
Aust et al., 2004, Barry et al., 2004, Gimble et al.,
2003, Gronthos et al., 2001, Katz et al., 2005, Pit-
tenger et al., 1999, Suva et al., 2004
CD13 (aminopeptidase) + + Aust et al., 2004, De Ugarte et al., 2003, Erickson et
20
al., 2002, Festy et al., 2005, Gimble et al., 2003,
Gronthos et al., 2001, Mitchell et al., 2006, Planat-
Benard et al., 2004, Rodriguez et al., 2005, Strem et
al., 2005, Talens-Visconti et al., 2006, Zuk et al.,
2002
CD14 + low/- -
Baksh et al., 2004, Curat et al., 2004, Gimble et al.,
2003, Gronthos et al., 2001, Jorgensen et al., 2004,
Lee et al., 2004, Minguell et al., 2001, Miranville et
al., 2004, Sengenes et al., 2005, Strem et al., 2005,
Suva et al., 2004, Zuk et al., 2002
CD15 - Rodriguez et al., 2005, Strem et al., 2005
CD16 - Strem et al., 2005, Zuk et al., 2002
CD18 (β-2 integrin) - - Gimble et al., 2003, Katz et al., 2005, Pittenger et
al., 1999
CD29 (β-1 integrin, fibronectin
receptor) + +
Aust et al., 2004, Baksh et al., 2004, Cao et al.,
2005, De Ugarte et al., 2003, Erickson et al., 2002,
Gimble et al., 2003, Gronthos et al., 2001, Guilak et
al., 2006, Katz et al., 2005, Lee et al., 2004, Mitchell
et al., 2006, Pittenger et al., 1999, Silva et al., 2003,
Strem et al., 2005, Zuk et al., 2002
CD31 (PECAM-1) + low/- -
Curat et al., 2004, De Ugarte et al., 2003, Fraser et
al., 2006, Gimble et al., 2003, Mitchell et al., 2006,
Planat-Benard et al., 2004, Strem et al., 2005, Wos-
nitza et al, 2006, Zuk et al., 2002
CD33 - Strem et al., 2005
CD34 (sialoumucin) +/- -
Baksh et al., 2004, De Ugarte et al., 2003, Erickson
et al., 2002, Festy et al., 2005, Fraser et al., 2006,
Gimble et al., 2003, Gronthos et al., 2001, Jorgen-
sen et al., 2004, Koc et al., 2000, Lee et al., 2004,
Minguell et al., 2001, Miranville et al., 2004, Mitchell
et al., 2006, Planat-Benard et al., 2004, Reyes et al.,
2001, Strem et al., 2005, Suva et al., 2004, Wosnitza
et al, 2006, Zuk et al., 2002
CD36 + Festy et al., 2005
CD38 - Strem et al., 2005
CD41a (gpIIb) - Katz et al., 2005
CD44 (hyaluronate receptor or
phagocytic glycpprotein-1) + +
Aust et al., 2004, Baksh et al., 2004, Cao et al.,
2005, De Ugarte et al., 2003, Erickson et al., 2002,
Fraser et al., 2006, Gimble et al., 2003, Gronthos et
al., 2001, Lee et al., 2004, Minguell et al., 2001,
Mitchell et al., 2006, Pittenger et al., 1999, Rodri-
guez et al., 2005, Silva et al., 2003, Strem et al.,
2005, Zuk et al., 2002
CD45 (LCA) +/- -
Aust et al., 2004, Baksh et al., 2004, Curat et al.,
2004, De Ugarte et al., 2003, Fraser et al., 2006,
Gimble et al., 2003, Gronthos et al., 2001, Jorgen-
sen et al., 2004, Koc et al., 2000, Lazarus et al.,
1995, Minguell et al., 2001, Miranville et al., 2004,
Reyes et al., 2001, Strem et al., 2005, Suva et al.,
2004, Zuk et al., 2002
CD49b (alfa-2 integrin) + + Katz et al., 2005, Pittenger et al., 1999, Rodriguez et

21
al., 2005
CD49d (alfa-4 integrin) + -
De Ugarte et al., 2003, Erickson et al., 2002, Fraser
et al., 2006, Gimble et al., 2003, Gronthos et al.,
2001, Katz et al., 2005, Pittenger et al., 1999, Silva
et al., 2003, Strem et al., 2005, Zuk et al., 2002
CD49e (alfa-5 integrin) + +
Aust et al., 2004, De Ugarte et al., 2003, Erickson et
al., 2002, Gronthos et al., 2001, Katz et al., 2005,
Pittenger et al., 1999, Silva et al., 2003, Strem et al.,
2005, Zuk et al., 2002
CD49f (alfa-6 integrin) - Katz et al., 2005
CD50 (ICAM-3) - - Gimble et al., 2003, Minguell et al., 2001
CD51/61 (vitronectin receptor) +/- +/- Katz et al., 2005, Pittenger et al., 1999, Silva et al.,
2003, Wosnitza et al., 2006
CD54 (ICAM-1) + +
De Ugarte et al., 2003, Erickson et al., 2002, Gimble
et al., 2003, Gronthos et al., 2001, Strem et al.,
2005, Zuk et al., 2002
CD55 (DAF) + +
De Ugarte et al., 2003, Erickson et al., 2002, Festy
et al., 2005, Gimble et al., 2003, Gronthos et al.,
2001, Strem et al., 2005, Zuk et al., 2002
CD56 (NCAM) - Gimble et al., 2003, Gronthos et al., 2001, Strem et
al., 2005, Zuk et al., 2002
CD59 (complement protein) + +
Aust et al., 2004, De Ugarte et al., 2003, Erickson et
al., 2002, Festy et al., 2005, Gimble et al., 2003,
Gronthos et al., 2001, Strem et al., 2005, Zuk et al.,
2002
CD62E (E-selectin) - - Gimble et al., 2003, Gronthos et al., 2001, Pittenger
et al., 1999, Zuk et al., 2002
CD62L (L-selectin) - + Katz et al., 2005, Pittenger et al., 1999
CD62P (P-selectin) - - Katz et al., 2005, Pittenger et al., 1999, Strem et al.,
2005
CD63 + Mitchell et al., 2006
CD65 + Festy et al., 2005
CD71 (transferrin receptor) + + Baksh et al., 2004, Gimble et al., 2003, Zuk et al.,
2002
CD73 (5´nucleotidase) + Gimble et al., 2003, Mitchell et al., 2006
CD90 (Thy-1) + +
Aust et al., 2004, Baksh et al., 2004, De Ugarte et
al., 2003, Gimble et al., 2003, Jorgensen et al.,
2004, Katz et al., 2005, Lee et al., 2004, Mitchell et
al., 2006, Pittenger et al., 1999, Rodriguez et al.,
2005, Silva et al., 2003, Strem et al., 2005, Suva et
al., 2004, Talens-Visconti et al., 2006, Zuk et al.,
2002
CD104 - Gimble et al., 2003, Strem et al., 2005, Zuk et al.,
2002
CD105 (endoglin) + +
Baksh et al., 2004, Cao et al., 2005, De Ugarte et
al., 2003, Erickson et al., 2002, Fraser et al., 2006,
Gimble et al., 2003, Gronthos et al., 2001, Guilak et
al., 2006, Jorgensen et al., 2004, Knippenberg et al.,
2005, Lee et al., 2004, Majumdar et al., 1998, Rodri-
guez et al., 2005, Strem et al., 2005, Talens-Visconti
et al., 2006, Zuk et al., 2002

22
CD106 (VCAM-1) +/- +
Baksh et al., 2004, De Ugarte et al., 2003, Erickson
et al., 2002, Fraser et al., 2006, Gimble et al., 2003,
Gronthos et al., 2001, Gronthos et al., 2003, Jorgen-
sen et al., 2004, Katz et al., 2005, Majumdar et al.,
1998, Pittenger et al., 1999, Strem et al., 2005, Zuk
et al., 2002
CD117 (c-Kit) +/- +
De Ugarte et al., 2003, Katz et al., 2005, Lee et al.,
2004, Minguell et al., 2001, Rodriguez et al., 2005,
Strem et al., 2005, Zuk et al., 2002
CD133/AC133 + low/- +
Gronthos et al., 2001, Gronthos et al., 2003, Katz et
al., 2005, Rodriguez et al., 2005, (Prentice et al.,
2004)
CD138 (syndecan-1) + Katz et al., 2005
CD140a (PDGFR-alfa) + + Katz et al., 2005, Pittenger et al., 1999
CD144 (VE-cadherin) + low/- Mitchell et al., 2006, Strem et al., 2005
CD146 (Muc18) + +
De Ugarte et al., 2003, Erickson et al., 2002, Gimble
et al., 2003, Gronthos et al., 2001, Strem et al.,
2005, Zuk et al., 2002
CD166 (ALCAM) + +
Aust et al., 2004, De Ugarte et al., 2003, Erickson et
al., 2002, Gimble et al., 2003, Gronthos et al., 2001,
Guilak et al., 2006, Knippenberg et al., 2005, Mit-
chell et al., 2006, Strem et al., 2005, Zuk et al., 2002
CD243 (MDR-1) - Katz et al., 2005
ABCG2 +/- + Katz et al., 2005, Mitchell et al., 2006
aldehyde dehydrogenase + Mitchell et al., 2006
alfa smooth muscle actin + + Gimble et al., 2003, Minguell et al., 2001
Collagen I + + Gimble et al., 2003, Minguell et al., 2001
Collagen II + + Gimble et al., 2003, Minguell et al., 2001
Glycophorin A - Rodriguez et al., 2005
HLA-ABC + +
Aust et al., 2004, Baksh et al., 2004, Barry et al.,
2004, Cao et al., 2005, Gimble et al., 2003, Katz et
al., 2005, Planat-Benard et al., 2004, Silva et al.,
2003, Suva et al., 2004
HLA-DR - -
Aust et al., 2004, Barry et al., 2004, Gimble et al.,
2003, Gronthos et al., 2001, Katz et al., 2005, Lee et
al., 2004, Rodriguez et al., 2005, Silva et al., 2003
KDR/Flk1 (VEGF-R2) + Cao et al., 2005, Mitchell et al., 2006, Wosnitza et
al., 2006
Osteonectin + + Gimble et al., 2003, Katz et al., 2005
Osteopontin + + Gimble et al., 2003, Katz et al., 2005, Minguell et al.,
2001
S100 + Atanassova et al., 2001, Kato et al., 1988, Wosnitza
et al., 2006
SH3 + + De Ugarte et al., 2003, Koc et al., 2000, Zuk et al.,
2002
STRO-1 +/- +
Baksh et al., 2004, De Ugarte et al., 2003, Fraser et
al., 2006, Gimble et al., 2003, Gronthos et al., 2001,
Gronthos et al., 2003, Majumdar et al., 1998, Rodri-
guez et al., 2005, Strem et al., 2005, Zuk et al., 2002
telomerase - + Katz et al., 2005

23
Vimentin + + Gimble et al., 2003, Minguell et al., 2001
vWF low + Mitchell et al., 2006, Wosnitza et al., 2006
RELEVANT CD MARKERS AND THEIR EXPRESSION PATTERN
In this thesis, the major aim was to differentiate between cells of endothelial versus adipo-
genic origin, and also to determine the level of maturation of the screened cells. Therefore,
the following markers were selected to be applied for flow cytometry analyses. CD31, also
known as platelet endothelial cell adhesion molecule 1 (PECAM-1), is commonly used for
labeling endothelial cells but is also expressed by monocytes, platelets, lymphocytes, and
granulocytes. CD51/61 labels various types of endothelial cells while vWF is expressed by
endothelial cells and platelets. CD105, or endoglin, is expressed by endothelial cells, acti-
vated monocytes and tissue macrophages, hematopoietic progenitor cells, as well as stromal
cells of certain tissues including bone marrow. Expression of CD105 is increased on acti-
vated endothelium in tissues undergoing angiogenesis and is a component of the TGF-beta
receptor system in human umbilical vein endothelial cells (HUVECs). CD34 is found on he-
matopoietic stem cells, vascular endothelial cells, embryonic fibroblasts and adult nervous
tissue. CD34 is absent on adult hematopoietic cells. VEGF signalling is performed through
KDR or VEGF receptor 2, a receptor tyrosine kinase. KDR is expressed during vasculogene-
sis and hematopoiesis and can be found on circulating endothelial progenitor cells, mature
vascular endothelial cells, and HUVECs. Acetylated-LDL (acLDL) can be applied to differen-
tially label endothelial progenitor cells, vascular endothelial cells, and macrophages. S100
labels nervous tissue, skeletal and cardiac muscle, malignant melanoma cells and some
other cell types. Additionally, it is well accepted as an early marker of adipogenesis21,199.
DIFFERENTIATION CAPACITY TOWARDS MULTIPLE LINEAGES
ADAS cells have been proven to be capable of multilineage differentiation429,433,434. As stem
cells from adipose tissue have their origin in the mesoderm, it is no longer surprising that
these cells can undergo different mesenchymal lineage conversions. Interestingly besides
the classic lineage differentiation, ADAS cells are likewise capable of undergoing transdiffer-
entiation towards the non-mesenchymal branches. By differentiating into neural cells, epithe-
lium, cardiomyocytes, skeletal muscle cells and hepatocytes, these cells show a wide plastic-
ity towards the ectodermal and endodermal lineage (Figure 3 and 4).
Clonal expansion of ADAS cells has proven that at least one part of the plasticity is situated
in a fraction of multipotent cells332,433. Despite pluri- and multipontent populations, fat tissue
harbours above all bi- and unipotent stem cells, solely capable of undergoing only two or one
differentiation pathways261,332.

24
To investigate multipotency, Rodriguez et al. created a single clone from fast-adherent ADAS
cells and proved that two out of 12 clones were able to undergo multilineage differentia-
tion331. The remaining ten clones had bipotent capacity. In contrary to these findings Kang et
al. found that a high percentage of single clones derived from primate ADAS cells were able
to retain adipogenic (82%), osteogenic (64%) and neurogenic (79%) differentiation potential.
The majority of clones (17 out of 20) showed even pluripotency along several mesodermal
and neuronal lineages195. These findings indicate that a high percentage of ADAS cells have
multipotential as well as pluripotential capacity in vitro due to self-renewal and creation of
daughter cells with the potency to convert into the major mesodermal and ectodermal line-
ages. Guilak et al. found that at least 81% out of 45 human ADAS cell clones showed unipo-
tency by differentiating into adipocytes, chondrocytes, osteocytes or neuronal cells138. Fur-
ther 52% of the ADAS cell clones converted into two or more lineages. The major number of
clones expressed phenotypes of osteoblasts (48%), chondrocytes (43%), and neuron-like
cells (52%) than of adipocytes (12%) which could be due to the repeated doublings leading
to the loss of adipogenic differentiation capacity amoung ADAS cells. These findings also
support the hypothesis that ADAS cells are capable of undergoing multipotent differentiation
pathways of the mesodermal and ectodermal lineages and do not solely consist of a mixture
of different unipotent stem cells. However, further investigation on incidence and percentage
between multipotent, bipotent and unipotent adipose mesenchymal stem cells remains es-
sential.

25
adipocytesendothelial cells
chondrocytes
osteocytes
myocyteshepatocytes
cardiomyocytes
neuronal cells
macrophages
ADAS
epithelium
Figure 3. Multiple differentiation pathways of human SVF cells
Human SVF cells convert into multiple cell lineages under appropriate culture conditions, including fat tissue, endothelial cells,
chondrocytes, osteocytes, myocytes, epithelial cells, hepatocytes, cardiomyocytes, neuronal cells and macrophages.

26
proliferationdifferentiation
chondrocyte
endothelial cell
osteocyte
myocyte
other tissues
ADAS
Figure 4. Scheme of SVF cell differentiation
SVF cells are capable to perform self-renewal and additionally to differentiate into numerous different cell types. The
lineage-committed cell progresses through several stages of maturation prior to the onset of terminal differentiation,
which is marked by the cessation of proliferation and shift towards increased expression of tissue-specific markers.

27
MESODERMAL DIFFERENTIATION
ADIPOGENESIS
Adipose-tissue derived adult mesenchymal stem cells naturally differentiate into mature adi-
pocytes. In adipogenic induction medium, ADAS cells develop intracellular lipid vacuoles
which coalesce and give rise to a single, cytoplasma-filling vacuole. Besides this definite
marker of adipogenesis349,414, several fat cell specific genes are expressed, comprising glyc-
erol-3-phosphate dehydrogenase (GPDH)216, lipoprotein lipase127,272, peroxisome proliferator-
activated receptor γ (PPARγ)366, leptin246, adipocyte fatty acid binding protein (aP2)11,
CCAAT/enhancer binding protein (C/EBP) and glucose transporter 4 (Glut4) (Figure 5)149.
GPDH is a key enzyme in adipose progenitor cell differentiation335 and has been widely ap-
plied for decades to measure the degree of adipocyte conversion298,368. The occurrence of
the genes given above in ADAS derived fat cells personates a very analogical pattern to that
noted in adipogenesis in MSCs82,308,312.
Adipocyte conversion of ADAS cells is not exclusively mined by miscellaneous growth factors
and the absence of serum (which acts as inhibitor during differentiation), but also by extracel-
lular matrix proteins, like fibronectin and collagen among others369.
ADAS proliferation and differentiation are two intimately connected processes, in adipogene-
sis as well as differentiation into other cell types (compare also Figure 4). Long-term prolif-
eration in fetal calf serum (FCS)-supplemented medium notedly and irreversibly decrease
ADAS cell potency to undergo fat differentiation149,158,160. Contrary to FCS, human autologous
serum (hS) enables long-term proliferation as well as subsequent differentiation, even after
prolonged culture and doubling rates160. Some studies support a composite of FCS with hS,
due to the fact that ADAS cells cultured in hS alone were found to proliferate weekly123, did
not attach to the culture flask75, or were not viable148. Hence the inhibitory effect of prolonged
proliferation periods in FCS-supplemented medium is most likely due to serum factors in the
FCS, varying between sundry charges and unpredictable to change or control. According to
this, a serum-free culture medium would be highly desirable136,202,387.
Optimal differentiation results are achieved when adipogenic induction medium is added at
maximal cell confluence. Also, an age-related decrease in proliferation and differentiation
capacity in ADAS cells has been observed7,147,198.
During acquisition of the adipocyte phenotype chronological changes in the expression of
several genes appear, including early, intermediate and late mRNA and protein markers as
well as triglyceride accumulation (see Figure 5)57,68,247,271,286.

28
Cell confluence and cell-cell contact both improve adipocyte conversion but are not manda-
tory for fat differentiation298. Growth arrest, in contrary, appears to be inevitable for differen-
tiation of ADAS cells into mature adipocytes.
Adipocyte specific genes are mainly transactivated by two transcription factors, PPAR-γ and
CCAAT/enhancer binding protein α (C/EBP-α)72,93,223,366. Both of them are also involved in
growth arrest required for adipocyte conversion. Furthermore C/EBP-α acts as an antimitotic
agent and leads to growth inhibition390,396. PPAR-γ is capable of inducing growth arrest in
fibroblasts and other cells, leading to cell cycle withdrawal9. PPAR-γ and C/EBP-α seem to
act cooperatively to bring about growth arrest that precedes differentiation9.
After withdrawal from the cell cycle in the early stage of adipocyte development, premature
signs of differentiation include the expression of lipoprotein lipase (LPL) mRNA4,83,132,247,272.
LPL is secreted in high amounts by mature fat cells and plays a central role in controlling lipid
accumulation89,129. LPL expression is independent of the addition of adipocyte differentiation
substrates and emerges spontaneously at cell confluence12,13. Therefore LPL occurrence
may display growth arrest rather than an early marker of adipogenesis and is not adipocyte
specific90,115,386. PPAR-γ and C/EBP expression is significantly upregulated during early adi-
pocyte differentiation, although low levels of these factors are also expressed in undifferenti-
ated ADAS cells. Both are subsequently involved in terminal differentiation by transactivation
of adipocyte specific genes and maximal levels of expression are attained in mature fat
cells54,71,72,420. During conversion into adipocytes, ADAS cells change their morphology from a
fibroblast-like shape to a spherical appearance. This process of lipid accumulation also in-
cludes major changes in cytoskeletal components, like actin and tubulin among others88,367,
as well as the level and types of extracellular matrix elements276 221,277. These changes in
cell shape reflect a distinct process in differentiation and are not results of lipid vacuole ac-
cumulation.
Two major events in terminal adipogenesis are de novo lipogenesis and acquisition of insulin
sensitivity. Lipogenic enzyme levels, including mRNA and protein expression, and activity of
triglycerol metabolism, increase 10- to 100-fold during final differentiation into adipocytes.
Maturation comprises mainly glycerol-3-phosphate dehydrogenase, glycerol-3-phosphate
acyltransferase, glyceraldehydes-3-phosphate dehydrogenase, fatty acid (FA) synthase, ace-
tyl-CoA carboxylase, stearoyl-CoA desaturase (SCD1), adenosintriphosphate (ATP) citrate
lyase and malic enzyme306,368,411. Insulin receptor numbers, insulin sensivity and glucose
transporter levels rise as well121,224,324,337. Furthermore, total number of adrenergic receptors
is upregulated during adipogenic differentiation, specially an increase in the β2- and the β3-
subtypes110,111,137,222. Leptin as well as aP2, an adipocyte-specific fatty acid binding protein,

29
are likewise increased during terminal adipogenesis11,40,73,246,368. PPAR-γ and C/EBP-α are
incorporated in the coordinate activation of several of these genes, comprising leptin, aP2,
Glut4 and SCD1 inter alia166,245,256,392,393.
growth arrestpost-confluent mitoses
clonal expansionearly late differentiation
Pref-1C/EBPβ
PPARγ
C/EBPα
adipocytegenes
lipogenicenzymes
FA bindingproteins
secretedfactors
ECM alteration cytosceletal remodelling
Although mature adipocytes were thought to remain in terminal differentiation without pro-
gressing towards dedifferentiation nor re-entering mitosis410,415, several studies support the
capacity of mature fat cells to undergo progressive dedifferentiation back to the primary
ADAS cell type378. Dedifferentiation followed by proliferation15,378 and subsequent differentia-
tion back into adipocytes355,377,379 as well as other lineages like endothelial cells have been
reported (Figure 4)314. A true reversion to an ADAS bi- or even multipotential cell phenotype
could play a role in the regulation of adipocyte and endothelial cell numbers and conse-
quently adipose tissue mass.
CHONDROGENESIS
Under chondrogenic medium conditions ADAS cells establish cellular nodules that produce
cartilage-related extracellular matrix molecules: collagen type II, VI, sulfate proteoglycans,
e.g. chondroitin 4-sulfate and keratan sulfate, proteoglycan aggrecan and proline/ arginine-
rich end leucine-rich repeat protein (PRELP) in vitro61,103,107,175,243,291,312,414,417,433,434. Optimal
conditions for differentiation are high density micro mass cultures supplemented with trans-
forming growth factor beta (TGF-β) and insulin, combined with dexamethasone, ascorbate-2-
phosphate, sodium pyruvate, or combined with transferrin, to name the most common possi-
bilities24,107,175,243.
Figure 5. Adipocyte
differentiation
Chronological changes in
the cellular processes and
transcriptional cascade
during fat cell matura-
tion.

30
ADAS cells seeded on scaffolds and subsequently implanted produce cartilage matrix in
vivo. Chondrogenic differentiation is influenced by different cell-biomaterial interactions, de-
pendent on the material applied. The biomaterial not only influences the chondrogenic differ-
entiation but also the mechanical properties of the construct25. ADAS cells seeded onto algi-
nate carriers followed by implantation into immunodeficient mice posses prolonged synthesis
of cartilage matrix molecules over 12 weeks107. In vivo, engineered cartilage contains ECM
molecules such as aggrecan, collagen II and collagen VI107,243. Besides the chondrogenic
growth factors and the ECM environment, oxygen tension also plays an important role in
chondrogenic differentiation. As cartilage is an avascular tissue, ADAS cells possible un-
dergo hypoxia during cartilage formation in vivo. ADAS cells differentiate in both, normoxic
as well as hypoxic environment, but hypoxia strongly reduces in vitro chondrogenesis248.
It remains controversial whether ADAS cells or MSCs are superior for cartilage engineer-
ing98,179,279,417. However, ADAS cells seem to have a greater chondrogenic potential than
MSCs when applied for engineering of cartilage in vitro as well as in a rabbit osteochondral
defect model98,279.
OSTEOGENESIS
ADAS cell differentiation towards the osteogenic cell lineage is well established for in vitro
as well as for in vivo animal tissue engineering mod-
els102,103,141,145,163,174,194,213,230,248,291,388,421,433,434. A clinical obervation is in part responsible for
the discovery of the osteogenic differentiation capacity of ADAS cells. A rare disorder named
“progressive osseus heteroplasia” together with the capacity of MSCs to convert into the os-
teogenic lineage led to the asssumption that ADAS cells are likewise able to differentiate into
osteocytes36,38,53,134,142,164,179,287,297. Patients with “progressive osseous heteroplasia” develop
calcified nodules representing normal bone which is placed at ectopic soft-tissue locations
including subcutaneous fat depots22,197,257,356,401.
Osteogenic induction of ADAS cells can be achieved by similar culture conditions as used in
MSCs, including supplementation with ascorbic acid together with 1-alpha,25-
dihydroxyvitamin D3 (1,25(OH)2D3), the hormonal metabolite of vitamin D, or
dexamethasone194,213,291. Under osteogenic differentiation medium ADAS cells are capable of
expressing diverse genes and proteins found in the osteoblast’s phenotype: type I collagen,
alkaline phosphatase, osteocalcin, osteonectin, osteopontin, parathyroid hormone (PTH)
receptor, bone morphogenetic protein-2 (BMP-2), BMP-4, BMP receptors I and II, bone sialo
protein and RunX-1141,213,291,433. Furthermore, ADAS cells exhibit a bone-like response to fluid
shear stress in vitro, achieved with mechanical loading by pulsating fluid flow (PFF). The re-
sult is increased nitric oxide (NO) production and cyclooxygenase-2 expression213. This might
indicate a bone-cell specific function during bone remodelling in vivo. ADAS cells are also

31
competent to generate mineralized matrix in vitro in both long term 2D or 3D osteogenic cul-
tures.
Human ADAS cells are able to generate bone tissue in immunodeficient rodent ectopic bone
models when seeded onto several different carrier scaffolds103,145,163,231. Furthermore these
cells have proven to regenerate bone defects in in vivo animal models81,85,104,and even in
humans when applied in a clinical trial for treatment of sizeable bilateral calvarial defects235.
Due to the restricted amount of bone marrow stem cells available in the patient with calvarial
defects, ADAS cells were applied to the defect in a single procedure together with autolo-
gous iliac crest bone and fibrin glue combined with resorbable sheets. CT-scans three
months after the reconstruction showed new ossification as well as nearly complete calvarial
continuity235.
ADAS cells have a capacity for bone formation that is comparable to bone marrow MSCs.
They maintain their osteogenic potential with aging when conferred to juvenile cells354.
SKELETAL MYOGENESIS
Cultured under appropriate myogenic medium conditions, ADAS cells undergo conversion
into a skeletal muscle cell phenotype in vitro263,332,433. Addition of hydrocortisone, dexa-
methasone and 5-azacytidine or coculture with primary skeletal myocytes induces myocyte-
related gene expression. Upregulated are key regulatory master factors like myogenic factor
5 (myf5), myf6, myogenic differentiation 1 (MyoD1) and myogenin232,263,433. ADAS cells also
respond with muscle cell differentiation in a myogenic environment, as they can be incorpo-
rated into muscle fibers of mdx mice after ischemia restoring dystrophin expression signifi-
cantly101.
Evidence exists that ADAS cells are a promising tool for restoration of muscle function in vivo
as well. ADAS cells attached to injectable poly(lactic-co-glycolic acid) (PLGA) spheres in-
jected subcutaneously into the necks of nude mice after culture in myogenic medium, gener-
ated new muscle tissue in vivo210. When transplanted into damaged regions of rabbit skeletal
muscle, ADAS cells increased fiber cross section area and muscle weight, resulting in sig-
nificantly raised contractile force, compared to the injured control group27. Furthermore, there
is striking indication that ADAS cells could be useful for the treatment of Duchenne muscular
dystrophy. When injected into the tibial and gastrocnemius muscle in dystrophin-deficient
mdx mice, an animal model for Duchenne muscular dystrophy, these cells induced a long-
term engraftment and a high expression of human dystrophin332.
CARDIOMYOGENESIS
The use of stem cells to regenerate damaged heart tissue is advocated as the new treatment
for heart failure secondary to heart disease or severe myocardial infarction. There are prom-

32
ising results of in vitro studies achieving myocardial differentiation in ADAS cells112,124,313,320.
When cultured in semisolid methylcellulose medium under low serum conditions and sup-
plemented with 2-mercaptoethanol, insulin, transferrin, interleukin-3 (IL-3), IL-6 and stem cell
factor (SCF), cohesive cell groups form and display branching fibers, sharing tight connec-
tions and beating at a single rate. These cells express cardiomyocyte-specific markers in-
cluding myosin-enhancing factor 2C, sacromeric α-actinin, ventricular and atrial myosin light
chain MLC-2v and MLC-2a, β-myosin heavy chain, atrial natriuretic peptide (NAD) and con-
nexin43 as well as connexin40313. In a murine in vivo model of acute myocardial injury, donor
ADAS cells were injected into the intraventricular chamber instantly after application of cryo-
injury to the myocardium373. Donor ADAS cells were identified exclusively in infarct areas of
animals treated with ADAS cells, but not in animals without myocardial damage or in the con-
trol group without cell treatment. Areas containing donor cells were positive for Heavy Chain,
Nkx2.5 and Troponin I373. So far this is one of the first study demonstrating that ADAS cells
have the potency both to undergo cardiomyocytic conversion and to engraft infracted myo-
cardium in vivo373. These are promising results for future application of ADAS cells in myo-
cardial infarction and other cardiovascular diseases, that are one of the leading causes for
morbidity and mortality in western countries.
IMMUNE CELLS
Preadipocytes as well as adipocytes secrete numerous inflammatory cytokines, such as tu-
mor necrosis factor α (TNF-α), and can be activated by lipopolysaccharide binding which
results in immunomodulatory molecule secretion56,169,207,208,242. Furthermore, PPARγ and aP2,
initially believed to be highly specific for adipocytes, were also found in macrophages309. In-
terestingly, preadipocytes were discovered to be genomically closer related to macrophages
than adipocytes69. Recent studies have demonstrated that ADAS cells display common func-
tional features of monocytes and macrophages by revealing specific phagocytosis activity
including antimicrobial activity via an oxygen-dependent mechanism342. These cells likewise
express CD80, CD86, CD45, anti perivascular monocyte/ macrophage antibody (ED2), anti-
macrophage monoclonal antibody-2 (MOMA-2), F4/80 and macrophage antigen-1 (Mac-1)/
CD11b69,84. In vivo transdifferentiation after grafting ADAS cells into the peritoneal cavity al-
lowed a transdifferentiation rate into macrophages of about 70% of cells69.

33
ECTODERMAL DIFFERENTIATION
NEUROGENESIS
The ability to undergo in vitro and in vivo neuronal lineage differentiation is another property
of ADAS cells that may have impact on CNS tissue regeneration113,195,196,385. With treatment
of different supplement combinations, ADAS cells achieve neurosphere formation when cul-
tured at high density195. Subsequent culture of the spheroid bodies on laminin leads to final
neural differentiation195. Differentiated cells protrude extensive cell processes with a neuronal
morphology and express neural markers, including nestin, neuronal nuclear protein (Neu N),
neurofilament (NF) 160, microtubule associated protein 2 a/b (MAP2ab) and the mature as-
trocyte marker glial fibrillary acidic protein (GFAP)195. While nestin and NF160 belong to the
early markers of the neuronal lineage, MAP2ab, Neu N as well as GFAP are thought to
match with terminal conversion of ADAS cells in neurogenesis195. Several protocols exist in
the literature to induce neurogenic conversion and achieve expression of neuronal-specific
markers20,117,192,339,341,419,423,433. Furthermore, positive staining for functional neuronal tissue,
comprising the neurotransmitter gamma-amino butyric acid (GABA), both subunits of the
glutamate N-methyl-D-aspartate (NMDA) receptor and α-1 voltage gated calcium channels,
was detected in neural differentiated ADAS cells341. Still it remains to be investigated if such
neurons are electrically active and functional with all essential characteristics of mature CNS
neurons364.
So far there are a few promising in vitro studies using ADAS cells for treatment of cerebral
ischemia, spinal cord injury as well as other neurological disorders193,196,340. ADAS cells mi-
grate into injured cerebral lesions after ischemic stroke and improve functional deficits and
motor recovery193. When applied in a rat model of spinal cord injury, ADAS cells migrate into
the damaged regions and significantly improve motor function after 4-5 weeks196.
Although neurogenesis is known to occur in adult life in limited regions of the brain119,385,
adult neuronal stem cells remain a population unattainable for clinical applications3,119. There-
fore ADAS cells display a highly promising source for cerebral injury treatment in the future.
EPITHELIAL DIFFERENTIATION
ADAS cells are known to accelerate epidermal regeneration when seeded together with
keratinocytes in a cutaneous reconstructive model. Furthermore, ADAS cells inhibit keratino-
cyte apoptosis and induced rete ridge- and epidermal ridge-like structure formation14. Fur-
thermore ADAS cells improve dermal regeneration in full thickness skin defects and reduce
wound contraction and subsequent scarring99. Treatment of human ADAS cells with all-trans
retinoic acid (ATRA) initiates cytokeratin 18 expression and the formation of keratin fibers

34
indicating epithelial differentiation. As demonstrated by Brzoska et al, more than 80% of cells
converted to an epithelial lineage cell type after incubation with ATRA55.
ENDODERMAL DIFFERENTIATION
HEPATOGENIC AND INTESTINAL DIFFERENTIATION
Although primary hepatocytes, liver stem cells and bone marrow stem cells represent the
main sources for liver regeneration, ADAS cells have recently been transdifferentiated into
hepatocyte-like cells351. Under treatment with a cytokine mixture and dimethylsulfoxid
(DMSO), ADAS cell conversion into the hepatic lineage was confirmed by the expression of
albumin, α-fetoprotein, as well as functional characteristics comprising low-density lipoprotein
(LDL) uptake and urea production. After transplantation into a hepatic injury mouse model,
ADAS cells differentiated into hepatocytes in vivo351. Other groups applied more complex
differentiation protocols to induce hepatic lineage differentiation in both ADAS cells and
MSCs, again finding upregulation of several liver-markers including albumine, cytochrome
P450 (CYP) isoenzymes, α-fetoprotein, C/EBPβ and hepatocyte nuclear factor 4 alfa
(HNF4α), which confirmed cellular conversion into hepatocytes381-383.
Autologous stem cell transplantation of ADAS cells has also been used for the treatment of
Crohn’s fistula in a phase I clinical trial122. Although a phase I trial does not allow final con-
clusions about effectiveness, the results achieved appear highly promising. After inoculation
of fistulas with autologous ADAS cells, six fistulas out of eight showed coverage with epithe-
lium at the external opening. A healing success was obtained in 75% and no adverse effects
were observed during the follow-up period of in average 22 months122.
VASCULOGENESIS AND ANGIOGENESIS
De novo blood vessel development in the embryo comprehends two different mechanisms,
vasculogenesis and angiogenesis. Vasculogenesis enfolds the primary vascular plexus
formed out of the mesoderm by differentiation of angioblasts followed by the generation of
primary blood vessels329. The fibroblast growth factor (FGF) family is crucial to conduct the
mesoderm in forming of angioblasts as well as hemangioblasts. Vascular endothelial growth
factor (VEGF) and its receptor tyrosine kinase VEGF-Receptor 2 (VEGF-R2/KDR/fetal liver
kinase 1 (flk-1)) represent a signalling system which is highly important for the differentiation
of endothelial cells and the development of the vascular system. Specified cell adhesion
molecules such as vascular endothelial cadherin (VE-cadherin) and platelet endothelial cell
adhesion molecule 1 (PECAM-1/CD31), as well as mechanical forces and vascular regres-
sion with remodelling, participate in the process of endothelial cell conversion, apoptosis, and

35
succeeding angiogenesis. Branching of several capillaries and pruning of others during vas-
culogenesis leads to de novo blood vessel formation and subsequently to a mature vascular
network.
Following the primary vascular plexus formation, endothelial cell differentiation and prolifera-
tion cause sprouting and splitting of blood vessels with subsequent development of new cap-
illary structures, an incident called angiogenesis. Angiogenesis consists of two different
types, sprouting angiogenesis from pre-existing capillaries and non-sprouting angiogenesis
or intussusception. Both require VEGF as a trigger for endothelial cell proliferation and mi-
gration62,63,328. Angiogenesis is a highly complex mechanism and displays an invasive cellular
process which is dependent on several growth factors, proteolytic enzymes, extracellular
matrix proteins and adhesion receptors46,143,144,180.
Primarily, it was adopted that vasculogenesis is a process constricted to embryonal devel-
opment, but recent studies proved that postnatal vasculogenesis takes place in the adult or-
ganism with the help of endothelial progenitor cells (EPCs)353. EPCs have a similar profile
when compared to an embryonic angioblast and these stem cells respond to angiogenic
growth factors with proliferation and migration forming in situ blood vessels via endothelial
differentiation. They incorporate into foci of neovascularization, thereby demonstrating that
postnatal neovascularization consists of both, angiogenesis as well as vasculogenesis16.
ANGIOGENESIS IN ADIPOSE TISSUE
The development and maturation of adipose tissue vascularity are critical to the function of
adipose tissue87,315. In fact adipose tissue begins to develop in well vascularized regions316
where adipocyte development is characterized by the appearance of a number of fat cell
clusters, termed “primitive organs”. Primitive fat organs are vascular structures in presump-
tive adipose tissue with few or no fat cells and increase throughout fetal development in
number and size. Fetal fat cell development is locally and temporarily dependent on capillary
formation. Arteriolar development clearly precedes adipocyte differentiation in fat tissue and
can be used to distinguish adipose tissue depots in the fetus87. Vascular development and
endothelial cell proliferation rate seem to be fat depot-dependent as endothelial cells from
several different rat fat depots have different replication rates after multiple passages226,227.
ADIPOGENESIS AND ANGIOGENESIS
Endothelial cells and preadipocytes share mutual developmental and autocrine as well as
paracrine relationships. Capillary formation in fat depots emerges synchronistically within the
early stages of adipogenesis, characterized by appearance of perivascular preadipo-
cytes152,184. Another evidence is that both endothelial cells and preadipocytes express αvβ3
integrin and secrete plasminogen activator inhibitor-1 (PAI-1), adjusting endothelial cell and

36
preadipocyte migration in vitro86. αvβ3 integrin is critical for vasculogenesis and angiogenesis
during development and mediates EC attachment, spreading and migration239,299. It binds
vitronectin, fibronectin and fibrin. PAI-1 regulates smooth muscle and endothelial cell motility
and migration through the interaction with vitronectin and its receptor (CD51/61) as well.
Preadipocyte-derived PAI-1 production ensures organization and coordination of adipogene-
sis and angiogenesis at local levels. Therefore preadipocytes are able to migrate together
with sprouting capillary endothelial cells in the formation of the primitive fat organ86.
Sprouting of new blood vessels is dependent on basement membrane protein degradation422.
The breakdown and degradation of connective tissue and basement membrane proteins be-
long to the critical phase of angiogenesis. Two major proteolytic enzymes are crucial for ex-
tracellular protein proteolysis, the matrix metalloproteinases (MMP) and the plasminogen
activator (PA)-plasmin system239,299,310. Inhibition of MMP activity leads to reduced or blocked
angiogenesis, indicating the implication of extracellular proteolysis in basement membrane
degradation, cell migration and ECM invasion, capillary lumen formation, regulation of cyto-
kine activity, and release of membrane bound VEGF and TNFα310,311.
MMP-2 and MMP-9 as well as their tissue inhibitors (TIMP) are expressed by ADAS cells
and adipose tissue in a depot-dependent manner51,70,100,120,251. MMP and TIMP significantly
influence adipose tissue development by adipocyte differentiation but may do so in coopera-
tion or independent of angiogenesis51,70,88,240.
Preadipocytes secrete PAI-1 in a depot-dependent manner32. PAI-1 inhibits both plasmino-
gen activators, t-PA and u-PA, and influences the migration of endothelial cells, smooth
muscle cells, and preadipocytes themselves86,311. Additionally, hypoxia induces PAI-1 ex-
pression in endothelial cells311. Elevated PAI-1 levels in the circulation are involved in the
development of the metabolic syndrome with insulin resistance and increased risk for cardio-
vascular disease8. Glitazones (insulin sensitizer) reduce human fat PAI-1 secretion leading to
improved endothelial function in chronic cardiovascular disease states300.
Both VEGF and FGF act as mitogenic agents in endothelial cells and preadipocytes by pro-
moting cell proliferation, migration and pericellular proteolysis (Figure 6). Their interaction on
angiogenesis is closely linked together41,389,422. VEGF acts through two tyrosine kinase recep-
tors, VEGFR-1 (fms like tyrosine kinase 1 (flt-1)) and VEGFR-2 (flk-1/KDR), which are mainly
expressed by endothelial cells, endothelial and hematopoietc progenitor cells and mono-
cytes. VEGF and its receptors both influence EC proliferation, migration, tubulogenesis, sur-
vival, vascular permeability, MMP proteinases, and the PA system. However, it seems to be
primarily VEGFR-2 which is crucial for angiogenesis. Besides, this receptor can be upregu-
lated in ECs by bFGF supplementation, indicating the synergism between these two growth
factors389. FGF may play a role in angiogenesis during tissue repair299 and enhances adipo-

37
cyte differentiation in vivo203. VEGF gene expression is influenced by a number of factors,
comprising hypoxia, insulin, growth factors and several different cytokines239.
VEGF is considered to be the major factor of the angiogenic activity of media conditioned by
adipocytes and adipose tissue432. Fat tissue expression and secretion of VEGF is induced by
hypoxia as well as insulin255,432. Vice versa insulin may be involved in increased adipose tis-
sue VEGF expression during rebound weight gain after diet restriction in rabbits268. Angio-
poietin (Ang)-1, Ang-2, and Tie 1 and Tie 2 receptors are regarded VEGF’s most fundamen-
tal partners (compare Figure 6)299,422. Ang-1 stabilizes blood vessel walls, whereas Ang-2
promotes destabilization. Hypoxia, VEGF, FGF, and cytokines induce Ang-2 expression in
microvascular ECs249,292. Several other studies have demonstrated the possibility that leptin
induces angiogenesis or influences angiogenic factors50,59,128,187,220,289,319,358,374,431. Both adi-
pose tissue and endothelial cells express the receptor for leptin binding49,50 and leptin is pro-
duced and secreted by adipocytes. Adipose tissue leptin and VEGF expression are similarly
regulated by hypoxia and insulin10,422. Therefore, leptin may potentiate VEGF-mediated an-
giogenesis by indirectly influencing VEGF secretion and uptake by endothelial cells302.
Several other growth factors are involved in angiogenesis as well, e.g. PDGF-β, TGF-β, TNF-
α and FGF, promoting vessel wall stability, establishing the integrity of capillary walls during
sprouting, and promoting tube formation in ECs (Figure 6)299,322.

38
Ang-1FGF
TGF-βPDGF
VEGF
EC
SMC
ECM
Strong evidence suggests adipogenesis to be influenced by angiogenesis and vice
versa338,358,360. Adipocyte endothelial cell interaction is thus presumed to be involved in the
development and maintenance of fat tissue. Also, adipose tissue itself is an origin of several
different vasoactive factors116. A close evolutionary relationship between preadipocytes and
ECs can be assumed since preadipocytes have been demonstrated to influence EC prolif-
eration, reduce apoptosis325, and increase mature EC viability and migration274. Impact on
proliferation and subsequent enhancement of tube formation, mainly meditated by secretion
of VEGF, TGF-β, bFGF and HGF by preadipocytes48,77,274,275,325. The expression of
leptin50,358, Ang 1370, Ang 278,139 and PAI-1345,365 by adipose tissue also has impact on angio-
genesis, as mentioned above, and the activity of these factors is interrelated. Leptin for in-
stance induces the expression of Ang 278 and enhances VEGF-mediated angiogenesis59.
Furthermore, preadipocytes are able to increase the amount of VEGF under hypoxia325 and
during fat pad development280. However, not exclusively adipocyte precursors are capable of
Figure 6. Regulation of blood vessel morphogenesis
during neovascularisation
New vessels arise from pre-existing capillaries and post-
capillary venoles. Pericytes start to migrate and blood
vessels dilate. Afterwards the basal membrane and the
extracellular matrix is degraded. Now endothelial migrate
into the perivascular space along the angiogenic stimulus
mediated through VEGF and its receptors, VEGF recep-
tor 1 and 2. Endothelial cells begin to proliferate and form
capillary-like structures including a lumen. TGF-β, FGF
and PDGF function as angiogenic growth factors as well.
FGF and VEGF function as mitogenic agents in ECs by
promoting cell proliferation, migration and pericellular
proteolysis. PDGF possesses a structural affinity with
VEGF. Ang-1 navigates the interaction between endothe-
lial cells and smooth muscle cells as well as pericytes by
stabilising the freshly formed blood vessels and acting as
an anti-permeability factor.

39
enhancing angiogenesis. Mature adipocytes also produce significant amounts of VEGF66.
MMP-9 which is upregulated in differentiated adipocytes51,204 increases the availability of ma-
trix-bound VEGF39. These findings indicate that mature fat cells, which require a very in-
tensely branched vascular network with direct contact to ECs, mediate endothelial cell ex-
pansion, migration and tube formation by VEGF secretion to optimize their oxygen supply
(Figure 7). This is supported by the fact that fat cells have a significant lower tolerance for
hypoxic conditions than preadipocytes and require higher concentration of oxygen406.
hypoxia
leptin
VEGF
TGF-β
FGF
PAI-1
TIMP
MMP-2MMP-9
proteolysis(MMP& PA)
angiogenicsprouting
cytokines Ang 1 ↑
Reciprocally, arteriolar differentiation precedes adipocyte development during fetal fat pat
formation87,151,154. Endothelial cell ECM maturation precedes differentiation of adipocyte
ECM87,153,156 and endothelial cells release ECM components that enhance preadipocyte dif-
ferentiation402. Remodelling of ECM components is important for both adipogenesis241 and
angiogenesis64. Preadipocyte spreading and migration during adipogenesis may be en-
hanced or even induced by blood vessel differentiation155. As VEGF expression can be gen-
erated by cold exposure in BAT18,19 and increased adipogenesis is promoted by enhanced
VEGF expression during rebound weight gain after diet restriction268, angiogenesis also
seems to play a crucial role in postnatal adipogenesis. Furthermore extracellular proteolysis
may be able to adjust fat tissue angiogenesis as well as fat cell maturation and develop-
ment167,267. Fat tissue proliferation is dependent on angiogenesis and can be regulated
through adipose tissue vascularization338. Anti-angiogenic agents leading to weight loss in
mice, as demonstrated by obesity models338, and are further proof of the close relationship
between endothelium and fat tissue. Several in vitro studies abet these findings, as ECs
Figure 7. Adipose angio-
genesis
Tissue hypoxia enhances
preadipocyte and adipocyte
secretion of proangiogenic
factors. VEGF, FGF, TGF-β
together with other cyto-
kines increase capillary
formation through EC
sprouting and Ang-1 ex-
pression. TIMP, MMP,
PAI-1 further support
vessel growth by tissue
proteolysis and tube forma-
tion.

40
promote adipose tissue derived precursor cell proliferation in conditioned medium177. Direct
adhesion of ECs promotes the active development of immature preadipocytes together with
increased growth of mature adipocytes15. These findings support the view that endothelium
closely regulates adipose tissue growth and homeostasis.
In conclusion, modulating the development of the vascular network in adipose tissue in either
an anti-angiogenic or pro-angiogenic manner may provide a strategy to solve problems with
obesity and cardiovascular complications on the one hand67, on the other hand may help
finding solutions for soft tissue engineering approaches to reconstruct defects after trauma or
tumor removal.

41
STUDIES
In this thesis, the differentiation plasticity and capacity of stromal vascular fraction cells to-
wards adipogenic and endothelial lineage was analyzed. The study was divided into the fol-
lowing six tasks:
1. Comparison of SVF cells from excised fat tissue versus liposuction material 2. Analysis of surface protein expression in SVF cells during the culturing process 3. Differentiation of unpurified SVF cells under endothelial medium conditions 4. Splitting of SVF cells in CD31- and CD31+ SVF cells and characterisation of purified frac-
tions 5. Differentiation potential of CD31- and CD31+ SVF cells towards the endothelial lineage 6. Differentiation potential of CD31- and CD31+ SVF cells towards the adipogenic lineage
1. Comparison of SVF cells from excised fat tissue versus liposuction material Two distinct isolation methods for gaining cells from the adipose SVF are established in the
literature, one from excised fat tissue and one from liposuction aspirates. Due to the harvest-
ing procedure liposuction material does not exclusively contain AT but also blood vessels
and fibrous tissue. Isolation of SVF cells from excised AT, in contrast, involves the manual
removal of capillaries and fibrous tissue. In order to prescind the differences in heterogenity
between SVF cells isolated from liposuctioned versus excised AT, freshly harvested SVF
cells from both isolation methods were analysed with flow cytometry. To quantify the amount
of endothelial cells, hematopoietic stem cells, and other cells contained in the two cell frac-
tions, antibodies against CD31, CD51/61, CD34 and KDR were applied.
2. Analysis of surface protein expression in SVF cells during the culturing process Not only the harvesting method but also the culture conditions influence the SVF cell charac-
teristics and their heterogenity. Long-term culturing of SVF cells generates high cell numbers
but changes major preadipocyte characteristics, i.e. protein surface expression and differen-
tiation capacity.
To analyse the changes in surface protein expression during culture of SVF cells in standard
medium, flow cytometry analyses were performed with stem cell markers for CD34, KDR and
S100 after 7 and 14 days.

42
3. Differentiation of unpurified SVF cells under endothelial medium conditions Usually, the harvested SVF is contaminated with ECs. When measuring preadipocyte trans-
differentiation potential to ECs, ECs embodied in the isolated SVF can mimic a successful
differentiation result even though ECs were already present in the applied cell fraction before
differentiation induction. To determine the growth potential of contaminating endothelial cells
after isolation, various growth media were tested including different EC supporting media
(EC-1, -2, -3). These culture media included standard EC media frequently used in the litera-
ture.
The first culture medium (EC-1) applied was an endothelial lineage differentiation medium for
angiogenic progenitor cells and contained Endothelial Growth Medium-2 Microvascular
(EGM-2MV) with 10 ng/ml VEGF and 10-6 mol/l rosiglitazone. The second medium (EC-2), an
endothelial differentiation medium, was Endothelial Cell Growth Medium (ECGM) supple-
mented with 50 ng/ml VEGF, 20 ng/ml IGF-1, and 2% FCS. The third culture medium (EC-3)
tested was a hematopoietic and endothelial progenitor cell medium for peripheral blood pro-
genitor cells containing IMDM, 50 ng/ml VEGF, 100 ng/ml SCGF-ß, 30% FCS, 1% BSA, and
10-4 mol/l ß-mercaptoethanol.
4. Splitting of SVF cells in CD31- and CD31+ SVF cells and characterisation of purified frac-
tions In order to seperately investigate the differentiation potential of both CD31- and CD31+ SVF
cells towards the adipogenic and endothelial lineage, splitting with paramagnetic beads was
performed (see studies 5. and 6.). To confirm the purity of both separated cell fractions, flow
cytometry analyses for CD105, vWF, CD34 and S100 were carried out in addition to vWF
immunofluorescence staining and histological evaluation. HUVECs were used as positive
control, fibroblasts as negative control.
5. Differentiation potential of CD31- and CD31+ SVF cells towards the endothelial lineage To investigate the differentiation potential of CD31- and CD31+ SVF cells towards the endo-
thelial lineage two endothelial differentiation media (EC-2 and EC-3) were applied to both
CD31 fractions separately. Endothelial differentiation was determined by (i) Matrigel assays
and (ii) flow cytometry analyses. 6. Differentiation potential of CD31- and CD31+ SVF cells towards the adipogenic lineage Both cell fractions, CD31- and CD31+ SVF cells, were tested for their differentiation potential
towards adipose tissue. After magnetic-assisted separation SVF cells were cultured in adi-
pogenic differentiation medium. Fat cell differentiation was measured (i) on a molecular level

43
by obtaining the GPDH activity and (ii) by cell counting under light microscopy and Oil Red O
Staining.

44
MATERIAL AND METHODS
TECHNICAL EQUIPMENT
♦ CO2 incubator, 37°C with 5% CO2
and > 95% humidity (Heraeus)
♦ Laminar flow hood (Flow Laborato-
ries GmbH)
♦ Laborwaage/ Laboratory scale (Sar-
torius)
♦ Laborfeinwaage/ Laboratory preci-
sion scale (Sartorius basic)
♦ Centrifuge Minifuge (T-Heraeus)
♦ Laboratory centrifuge (SIGMA)
♦ Centrifuge Biofuge A (Heraeus)
♦ Phasecontrast microscope, Telaval
31 (Zeiss)
♦ Photo device (Nikon D100)
♦ Fluorescence microscope Axioscope
(Leica)
♦ Programme DISCUS (Leica)
♦ Coverplates (Shandon)
♦ Counting cell chamber (Neubauer
improved)
♦ Water shaking bath (GFL
1092/1083)
♦ Shaking device Reax 2000
(Heidolph)
♦ Autoclave (Melag Autoklav 23)
♦ Conical tubes, 10 ml, 50 ml konisch
(Costar)
♦ Pipetting device Pipetboy
(Tecnomara)
♦ Culture flask T-25 (25 cm2/ 5 ml)
(Greiner)
♦ Culture flask T-75 (75 cm2/ 25 ml)
(Greiner)
♦ Culture dish/ plate, 6-well, 12-well,
24-well, 96-well (Falcon)
♦ Petri dish Ø 150 mm (Falcon)
♦ Glass vials 50 ml, 100 ml, 250 ml,
500 ml (Schott Duran)
♦ Pipettes, 1 ml, 2 ml, 5 ml, 10 ml, 25
ml (Greiner)
♦ Eppendorf cups 1.5 ml (Eppendorf)
♦ Disposable syringe Discardit II 5 ml,
10 ml, 20 ml (Becton Dickinson)
♦ Elisa-Reader with HP-Printer (Dynex
Technologies)
♦ Programme Revelation (Dynex
Technologies)
♦ Multi-channel pipette (Eppendorf)
♦ Repeater Pipette and combitips (Ep-
pendorf)
♦ Glass cylinder, Borosilikatglas
(Braun)
♦ Teflon flask (Braun)
♦ Syringe driven filter unit Millex GP
0.22 µm (Millipore)
♦ 70 μm nylon mesh (Verseidag
Techfab GmbH)
♦ 250 µm nylon mesh (Verseidag
Techfab GmbH)
♦ Sucking device Vacusafe (IBS In-
tegra Biosciences)
♦ Pasteurpipettes, 230 mm (Brand)
♦ Tweezers (Aesculap)
♦ Scalpels (Feather)
♦ Scissors (Aesculap)
♦ Clamps (Aesculap)

45
♦ Needles Microlance (Becton Dickin-
son)
♦ Glass slides (Engelbrecht)
♦ Quadriperms (Heraeus)
♦ Magnetic rack Dynal MPC-1 (Dynal
Biotech GmbH)
♦ MiniMACS separation unit (Miltenyi
Biotec GmbH)
♦ MACS Multi Stand (Miltenyi Biotec
GmbH)
♦ MACS columns type MS (Miltenyi
Biotec GmbH)
♦ FACS Calibur apparatus (Beckton
Dickinson)
♦ FACS tubes (Sarstedt)
♦ Programme Cell Quest Pro (Beckton
Dickinson)
♦ Parafilm (Pechiney Plastic Packing)
REAGENTS
♦ Dulbecco´s Modified Eagle Medium
(DMEM)/Ham’s F12 (F12) 1:1 (PAA)
♦ Dulbecco´s Modified Eagle Medium
(DMEM) High Glucose (PAA)
♦ Isocove´s Modified Dulbecco´s Me-
dium (IMDM) with GlutaMAX (Invi-
trogen)
♦ Endothelial Cell Growth Medium with
supplement mix (PromoCell)
♦ Endothelial Growth Medium-2 Mi-
crovascular (EGM-2 MV) with sup-
plement mix (Cambrex Bio Science)
♦ Penicillin (PAA Laboratories)
♦ Streptomycin (PAA Laboratories)
♦ Fetal calf serum (FCS) (Biochrom)
♦ Bovine serum albumin (BSA) (PAA)
♦ BSA for immunhistology (Sigma)
♦ Basic fibroblast growth factor (bFGF)
(Peprotech)
♦ Vascular endothelial growth factor
(VEGF) (Peprotech)
♦ Insulin-like growth factor 1 (IGF-1)
(Peprotech)
♦ Stem cell growth factor beta (SCGF-
β) (Peprotech)
♦ Dexamethasone (Sigma)
♦ Insulin (Roche)
♦ Transferrin (Sigma)
♦ Triiodo-L-thyronine (Sigma)
♦ Isobutylmethylxanthine (IBMX)
(Sigma)
♦ Pioglitazone (Takeda Pharmaceuti-
cals)
♦ Rosiglitazone (Cayman Chemical)
♦ Phosphat buffered saline (PBS2-)
(Biochrom)
♦ Aqua ad injectabile (Delta Select)
♦ Aqua bidest (Pharmacy RWTH
Aachen)
♦ Trypsin 0.02%, 0.05% EDTA in PBS
(PAA Laboratories)
♦ Collagenase CLS Typ I (Biochrom)
♦ Dispase Typ II (Roche)
♦ Fibronectin (Biochrom)
♦ Matrigel™, growth factor reduced
(BD Biosciences)

46
♦ Cell recovery solution (BD Biosci-
ences)
♦ Oil Red O (Merck)
♦ Glycerol-3-phosphat-dehydrogenase
(GPDH) activity assay kit (TaKaRa)
♦ HEPES (Sigma Aldrich Chemie)
♦ NaCl (Merck)
♦ CaCl2 (Merck)
♦ HCl (Merck)
♦ Glucose (Merck)
♦ NH4Cl (Merck)
♦ NaHCO3 (Merck)
♦ KHCO3 (Merck)
♦ β-mercaptoethanol (Fluka)
♦ Paraformaldehyde (PFA) (MERCK)
♦ Acetone (Pharmacy RWTH Aachen)
♦ 4’,6’-Diamidin-2’-phenylindol-
dihydrochlorid (DAPI) (Boehringer
Mannheim)
♦ DAPI working solution (10µl DAPI
stock solution plus 10ml methanol)
♦ Dimethylsulfonoxid (DMSO) (Sigma)
♦ Anti-CD31 paramagnetic Dynabeads
(Dynal Biotech GmbH)
♦ Anti-CD31 MicroBead Kit (Miltenyi
Biotec GmbH)
♦ Mouse anti-human S100 antibody
(DAKO GmbH)
♦ Rabbit anti-human von-Willebrand-
Factor (vWF) antibody (DAKO
GmbH)
♦ Secondary swine anti-rabbit FITC-
antibody, IgG (DAKO GmbH)
♦ Mouse anti-human CD31 antibody
(Dianova GmbH)
♦ Mouse anti-human CD34 antibody
(Dianova GmbH)
♦ Mouse anti-human CD51/61 anti-
body (Dianova GmbH)
♦ Mouse anti-human KDR antibody
(Dianova GmbH)
♦ Secondary goat anti-mouse FITC-
antibody, IgG (Dianova GmbH)
♦ Anti-human CD105 FITC-conjugated
antibody (ImmunoTools GmbH)
♦ Isotype control IgG1 and IgG2 anti-
bodies (ImmunoTools GmbH)
♦ CD31 APC-conjugated antibody
(Miltenyi Biotec GmbH)
♦ DiI acLDL cell tracker (Cell Systems)
♦ Mowiol 4-88 (Calbiochem)

CULTURE MEDIA
Attachment medium/Fibroblast medium
♦ DMEM/Ham’s F12
♦ 10% FCS
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
Table 2.
Endothelial differentiation media
EC-1 EC-2 EC-3
♦ Endothelial Growth
Medium-2 Microvascu-
lar (EGM-2MV)
♦ 10 ng/ml VEGF
♦ 10-6 mol/l rosiglitazone
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
♦ Endothelial Cell
Growth Medium
(ECGM)
♦ 50 ng/ml VEGF
♦ 20 ng/ml IGF-1
♦ 2% FCS
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
♦ IMDM
♦ 50 ng/ml VEGF
♦ 100 ng/ml SCGF-β
♦ 30% FCS
♦ 1% BSA
♦ 10-4 mol/l β-
mercaptoethanol
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
HDMEC medium
♦ EGM-2MV
♦ 15% FCS
♦ 100µg/ml Heparin
♦ 7µg/ml IBMX
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
HUVEC medium
♦ EGM-2
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin

48
Preadipocyte proliferation medium
♦ DMEM/Ham’s F12
♦ 10% FCS
♦ 100 µl bFGF
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
Table 3.
Adipogenic differentiation media
Adipocyte differentiation medium 1 (d1-5) Adipocyte differentiation medium 2 (≥ d6)
♦ DMEM/Ham’s F12
♦ 100 nM dexamethasone
♦ 10 µg/ml transferrin
♦ 66 nM insulin
♦ 1 nM triiodo-L-thyronine
♦ 0.5 mM IBMX
♦ 0.1 µg/ml pioglitazone
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
♦ DMEM/Ham’s F12
♦ 100 nM dexamethasone
♦ 10 µg/ml transferrin
♦ 66 nM insulin
♦ 1 nM triiodo-L-thyronine
♦ 100 U/ml penicillin
♦ 100 µg/ml streptomycin
Table 4.
Seading of SVF cells
Seeding
density
20.000/cm2
(cell num-
bers)
Seeding
density
30.000/cm2
(cell num-
bers)
Cells at
confluency
(cell num-
bers)
Trypsin/
EDTA (ml)
Growth me-
dium (ml)
6-well 181.400 272.100 1.2 x 106 1 3
T-25 0.5 x 106 0.75 x 106 2.8 x 106 2 5
T-75 1.5 x 106 2.25 x 106 8.4 x 106 5 10-15

49
BUFFERS AND SOLUTIONS
Collagenase buffer
♦ 100 mM HEPES
♦ 120 mM NaCl
♦ 50 mM KCl
♦ 1 mM CaCl2
♦ 50 mM glucose
♦ → pH 7.4
Collagenase solution
♦ 0.494 U/ml collagenase CLS Typ I
♦ 1.5 % BSA
♦ Collagenase buffer
♦ → 1 ml collagenase buffer per 1 g tissue
Erythrocyte lysis buffer
1. Stock solution: For 100 ml Aqua bidest:
♦ 8.02 g NH4Cl
♦ 0.84 g NaHCO3 or KHCO3
♦ 0.37 g EDTA (Di-Na-salt)
♦ 100 ml of this solution plus 900 ml Aqua bidest
♦ → pH 7.4
2. Working solution:
♦ Stock soltution diluted 1:10 with Auqa bidest
MACS buffer
♦ 2 mM EDTA
♦ 0.1% BSA
♦ PBS2-
♦ → degassed by ultrasound
FACS buffer
♦ 2 mM EDTA
♦ 0.2% FCS
♦ PBS2-

50
ISOLATION OF SVF CELLS
SVF cells were isolated from freshly excised human subcutaneous abdominal or mammary
fat tissue at the Department of Plastic Surgery and Hand Surgery - Burn Center from patients
who underwent elective operations (e.g. abdominoplasty, breast reduction, liposuction). In
regards to the amount of human preadipocytes gained from fat tissue, the harvesting method
plays a central role. There are two different surgical isolation methods for fat tissue. Firstly
harvesting of adipose material can be performed by simple excision of adipose tissue during
elective operations such as abdominoplasty, breast reduction and others. The second
method is liposuction in which numerous diminutive fat pieces are isolated. This displays the
most atraumatic version for the patient (Figure 8). Cell yields from liposuction vary enor-
mously in the literature, according to the harvesting procedure, from 1.6 x105 to approxi-
mately 2 x106 cells per gram of lipoaspirated human fat405,434.
Harvesting of excised fat tissue was arranged by isolating the single fat lobules which were
prepared by careful removal of capillaries and connective tissue. After mincing into small
pieces of 2-6 mm in diameter with scissors, adipose tissue was washed with 0.9% NaCl and
digested. Harvesting of adipose tissue by liposuction was performed according to Sydney
Coleman. With the Coleman method, manually applied negative pressure using a 10-cc-
syringe with a blunt tip cannula minces the fat lobules and sucks the small pieces into the
syringe (Figure 8)80. To guarantee cell viability no additional solutions were added. Harvest-
ing sides were predominantly abdomen and breast. Isolation methods were adapted to the
methods of Rodbell330 and Björntorp44,45 with minor modifications. All procedures were per-
formed under sterile conditions under the laminar flow hood. Adipose tissue was either used
directly after harvesting or stored for up to 24 hours at 4°C in DMEM High Glucose. Adipose
Figure 8. Different isolation
methods for fat tissue
Process of adipose tissue harvest
from tissue excision, e.g. abdomin-
oplasty (A.) or lipoasiprates (B.).
A.
B.

51
tissue from aspiration was digested by collagenase treatment after washing with 0.9% NaCl
to remove possible blood contamination.
Digestion of excized and liposuctioned fat tissue was performed with 0.2% collagenase CLS
type I (0.494 U/ml) and 1.5% BSA dissolved in collagenase buffer for 60 minutes at 37°C
under constant shaking. The gained suspension was filtered through a 250 µm filter mesh
and centrifuged at 700g for 7 minutes at room temperature. SVF cells were separated from
mature fat cells by discharging the fat layer on top, containing the lipid-filled adipocytes. SVF
cells located in the cell pellet on the bottom of the tube were resuspended in different culture
media dependent on their further use (Figure 9). For separation of CD31- and CD31+ cells,
the SVF cell pellet was resuspended in EGM-2 MV culture medium in order to enhance EC
numbers and maintain EC characteristics of CD31+ cells in the SVF. For plain flow cytometry
analyses and endothelial differentiation assays of SVF cells, cells were seeded in
DMEM/F12 (1:1) with 10% FCS. Seeding was performed on fibronectin-coated culture flasks
at a density of 30,000 cells/cm2. Cells were washed twice with 0.9% NaCl after 24 and 48
hours to remove contaminating blood cells.

52
adipose SVFin culture
fat tissue fat lobules
digestion
filtration
centrifugation
separation
resuspention
seeding
SVF pellet
CD31 DYNABEAD SEPARATION OF SVF CELLS
To split CD31- from CD31+ cells two widely accepted methods for cell separation were ap-
plied, cell splitting with Dynabeads and with Microbeads. For endothelial cell separation the
most frequently used surface protein is CD31 as it is expressed on all endothelial cells but
also on platelets. To minimize platelet contamination cells were washed thoroughly with NaCl
after harvesting and again with PBS2- before trypsinizing for Dynabead/Microbead separa-
tion. Cells were grown to subconfluence for three to five days, trypsinized after washing, re-
Figure 9. SVF cell isolation from
excised tissue
Excised adipose tissue is minced with
scissors into single fat lobules and digested
with a collagenase solution. The resulting
suspension is then filtered through a nylon
mesh to remove undigested components,
specially fibrous tissue and small vessels.
Subsequent centrifugation deposits the
stromal vascular fraction (SVF) as a pellet
at the bottom of the tube from the floating
lipid layer (mature adipocytes) on top. The
supernatant is removed by pipetting and
followed by resuspension of the cell pellet
which allows seeding of the SVF cells in a
culture dish.

53
suspended in 10 ml cold PBS2-/0.1% BSA with 2 mM EDTA (MACS buffer) and washed by
pipetting the cell suspension carefully. Cells were centrifuged at 400g for 5 minutes, resus-
pended in 8 ml cold MACS buffer and passed trough a 70 µm nylon mesh into a 15 ml tube.
The mesh ensured that only single cells passed through by excluding cell clusters thereby
maximizing the purity and percentage of CD31+ cells. Afterwards cells were counted with a
Neubauer chamber to calculate cell numbers. To ensure optimal binding of paramagnetic
beads cell concentration should not exceed 108 cells/ml sample volume. Before use Dyna-
beads were prepared by washing with cold MACS buffer. Dynabeads were suspended in 8
ml cold MACS buffer in a 15 ml tube and pipetted thoroughly. Next the tube was placed into
the magnetic rack for two minutes until the solution became clear and all beads had moved
to the magnet. The see-through supernatant was discharged, the tube removed from the
rack and the beads were resuspended anew in 8 ml cold MACS buffer by pipetting. This
washing step was performed three times and Dynabeads were resuspended in the original
volume again. To separate CD31+ cells from CD31- cells by anti-CD31 paramagnetic beads,
cells were incubated with 200 µl washed CD31 Dynabeads for endothelial cells (25 µl
beads/ml cell suspension) at 4°C for 30 minutes under constant rotation, allowing for binding
of endothelial cells to the beads. Afterwards the bead-cell suspension was pipetted a few
times to release cell clusters which may have formed during the incubation period. The tube
containing the Dynabead-labeled cells was placed in the magnetic rack (Dynal MPC-1) for 2
minutes until all beads were attached to the magnet (Figure 10). The supernatant with CD31-
cells was transferred to a fresh tube by pipetting, centrifuged at 400g for 5 minutes, washed
once with MACS buffer, and cells were subsequently seeded on fresh culture flasks. For iso-
lating CD31+ cells, three washing steps as described before were performed with Dynabead-
labelled cells. After the falcon tube was removed from the magnetic rack, CD31+ SVF cells
with Dynabeads attached to the surface were resuspended in fresh DMEM/F12.

54
For attachment to culture flasks both cell types were cultivated in DMEM/F12 supplemented
with 10% FCS in fibronectin-coated flasks over night at a density of 80.000 cells/cm2. Seed-
ing at high density was performed in order to keep proliferation times as low as possible until
complete confluence was achieved, necessary for further differentiation experiments towards
the adipogenic and endothelial lineage.
CD31 MICROBEAD SEPARATION OF SVF CELLS
Cells were grown to subconfluence, trypsinized after washing twice with PBS2-, resuspended
in 10 ml MACS buffer and passed trough a 70 µm nylon mesh to remove cell clusters. Cells
were resuspended in 60 µl cold MACS buffer to a maximum concentration of 1x 107 cells/
60 µl buffer. 20 µl FcR Blocking reagent was added to minimise unspecific binding of Micro-
Beads to FcR domains. The gained suspension was vortexed thoroughly and 20 µl of CD31
MicroBeads were added. Cells were incubated for 15 minutes at 4 °C to allow binding of
beads to the cell surface. During the incubation period MACS columns were prepared by
placing two columns in the magnetic fields of each MiniMACS separator. The columns were
rinsed with 1 ml MACS buffer three times. After the incubation time non-attaching beads
were removed from the suspension cells by washing once with 5 ml medium and subsequent
centrifugation at 400g for 5 minutes. The supernatant was discharged, cells were resus-
pended in 500 µl MACS buffer (up to 108 cells) and transferred onto prepared MACS col-
B.
C.
A. Figure 10. Splitting of SVF cells with
paramagnetic CD31 Dynabeads
Dynabeads are uniform, superparamagnetic
polystyrene beads with a diameter of about
4.5 µm. The beads are coated with a mouse
IgG1 monoclonal antibody specific for the
CD31 cell surface antigen PECAM-1. Re-
quired material for Dynabead separation
includes the Dynal magnet, CD31 Dyna-
beads and a conical tube (A.). Washed Dyna-
beads are mixed with the cell sample and the
beads ligate to the CD31+ target cells (red)
after incubation (B.). After binding the posi-
tive cells are separated from the negative cells
by a magnet, whereas the CD31+ cells attach
to the magnet and the CD31- cells remain in
the cell suspension (C.).
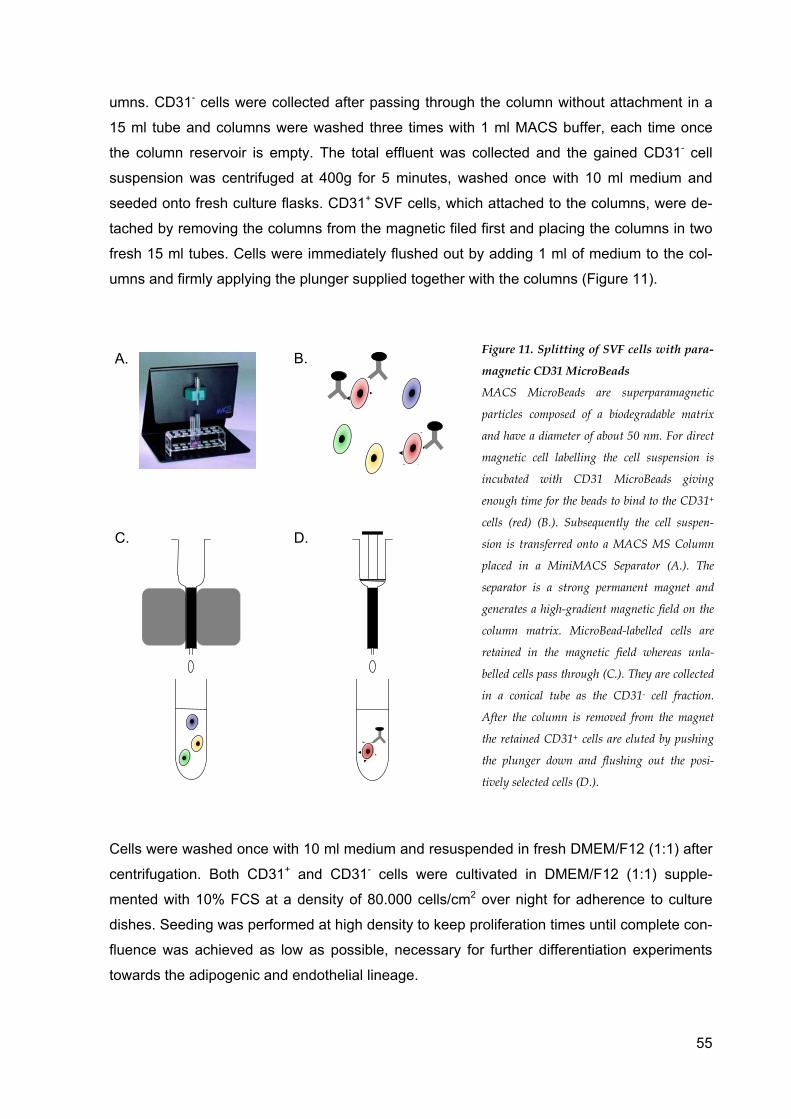
55
umns. CD31- cells were collected after passing through the column without attachment in a
15 ml tube and columns were washed three times with 1 ml MACS buffer, each time once
the column reservoir is empty. The total effluent was collected and the gained CD31- cell
suspension was centrifuged at 400g for 5 minutes, washed once with 10 ml medium and
seeded onto fresh culture flasks. CD31+ SVF cells, which attached to the columns, were de-
tached by removing the columns from the magnetic filed first and placing the columns in two
fresh 15 ml tubes. Cells were immediately flushed out by adding 1 ml of medium to the col-
umns and firmly applying the plunger supplied together with the columns (Figure 11).
Cells were washed once with 10 ml medium and resuspended in fresh DMEM/F12 (1:1) after
centrifugation. Both CD31+ and CD31- cells were cultivated in DMEM/F12 (1:1) supple-
mented with 10% FCS at a density of 80.000 cells/cm2 over night for adherence to culture
dishes. Seeding was performed at high density to keep proliferation times until complete con-
fluence was achieved as low as possible, necessary for further differentiation experiments
towards the adipogenic and endothelial lineage.
B.
C. D.
A.Figure 11. Splitting of SVF cells with para-
magnetic CD31 MicroBeads
MACS MicroBeads are superparamagnetic
particles composed of a biodegradable matrix
and have a diameter of about 50 nm. For direct
magnetic cell labelling the cell suspension is
incubated with CD31 MicroBeads giving
enough time for the beads to bind to the CD31+
cells (red) (B.). Subsequently the cell suspen-
sion is transferred onto a MACS MS Column
placed in a MiniMACS Separator (A.). The
separator is a strong permanent magnet and
generates a high-gradient magnetic field on the
column matrix. MicroBead-labelled cells are
retained in the magnetic field whereas unla-
belled cells pass through (C.). They are collected
in a conical tube as the CD31- cell fraction.
After the column is removed from the magnet
the retained CD31+ cells are eluted by pushing
the plunger down and flushing out the posi-
tively selected cells (D.).

56
CULTURING OF SVF CELLS
Culture media were changed for both CD31- and CD31+ to either endothelial or adipogenic
differentiation medium 12 hours after Dynabead or MicroBead separation, respectively (see
Figure 12).
To evaluate differentiation plasticity towards endothelium, three types of differentiation media
were tested (Table 2). The first culture medium (EC-1) used was an endothelial differentiat-
zion medium for angiogenic progenitor cells according to Wang et al.409 and contained Endo-
thelial Growth Medium-2 Microvascular (EGM-2MV) with 10 ng/ml VEGF and 10-6 mol/l
rosiglitazone. The second medium (EC-2), an endothelial differentiation medium, was Endo-
thelial Cell Growth Medium (ECGM) supplemented with 50 ng/ml VEGF, 20 ng/ml IGF-1, and
2% FCS, according to Miranville et al.260 with minor modifications. The third culture medium
(EC-3) tested was a hematopoietic and endothelial progenitor cell medium for peripheral
blood progenitor cells according to Gehling et al. 125, containing IMDM, 50 ng/ml VEGF, 100
ng/ml SCGF-ß, 30% FCS, 1% BSA, and 10-4 mol/l ß-mercaptoethanol.
To perform adipogenic differentiation, cells were cultured in DMEM/F12 (1:1) supplemented
with bFGF (10 ng/ml) and 10% FCS (referred to as proliferation medium) to obtain cell con-
fluency again. Lipid droplet development was then promoted for 21 days by changing me-
dium to DMEM/F12 (1:1) without serum addition, supplemented with 66 nM insulin, 100 nM
dexamethasone, 0.5 mM IBMX, 0.1 µg/ml pioglitazone, 1 nM triiodo-L-thyronine, and 10
µg/ml human transferrin (adipocyte differentiation medium 1, Table 3). After 5 days of incuba-
tion, medium was used as before but without IBMX and pioglitazone for 16 more days (adi-
pocyte differentiation medium 2, Table 3). Culture media were changed every three days.

57
10% 90%CD31 Dynabead/ Microbead
splitting
SVF culture in EC medium
CD31- SVFCD31+ SVF
adipose differentiation► GPDH assay
capillary structures► Matrigel
Extracellular Matrix
CD31- SVFCD31+ SVF
CELL SPLITTING
Medium was removed from the culture flasks and cells were washed twice with PBS2- to
eliminate residual medium. According to the size of the culture flask trypsin/EDTA was added
and cells were incubated at 37°C for 2-3 minutes. Serum-containing medium was supple-
mented to the trypsin/EDTA containing the detached cells in order to stop the enzymatic
process. Cells were centrifuged at 1350 rpm for 5 minutes, resuspended in 5 to 10 ml fresh
culture medium and counted with a Neubauer cell counting chamber under the phasecon-
trast microscope. Cell numbers were calculated as follows: Counted cells / 4 x 104 = cell
numbers / ml. Subsequently cells were seeded at the desired density and medium was
changed the following day to remove non-vital cells.
Figure 12. Experimental design of
Dynabead/Microbead splitting of
SVF cell culture and separate
differentiation of subpopulations
towards adipogenic and andothe-
lial lineage
SVF cells were cultured in EC me-
dium to enhance EC numbers prior to
cell separation. After splitting with
paramagnetic beads CD31- and
CD31+ SVF cells were both cultured
in endothelial and adipogenic differ-
entiation medium. Endothelial differ-
entiation was measured via capillary
structure formation on Matrigel
ECM and flow cytometry analysis.
Adipogenic conversion was deter-
mined by GPDH activity and Oil Red
O staining.

58
FIBRONECTIN AND MATRIGELTM COATING Fibronectin (1 mg) was diluted to a concentration of 0.02 mg/ml by adding 50 ml aqua ad
injectabile (fibronectin working solution). The bottom of the culture flask was covered with
fibronectin working solution and air-dried under sterile conditions.
MatrigelTM Basement Membrane Extracellular Matrix is made out of Engelbreth-Holm-Swarm
(EHS) mouse tumor. Major components of Matrigel include laminin, collagen IV, heparan
sulfate proteoglycanes, entactin and nidogen212. Further it contains TGF-β, FGF, tissue
plasminogen activator254 and other growth factors which naturally exist in the EHS tumor.
Matrigel is known to support and provide several factors necessary for the study of angio-
genesis283,284. For testing the angiogenic potential of SVF cells in vivo the thin gel method
was used. Culture flasks are coated with Matrigel in a thickness of about 0.5 mm and cells
are plated on top. Matrigel was diluted 1:2 with serum-free medium (EGM2-MV without sup-
plement mix), placed in aliquots and stored at -20°C. To handle Matrigel exclusively pre-
cooled material should be used as Matrigel gels at temperatures over 5-10°C. One day prior
to use Matrigel was thawn on ice at 4°C overnight and pipettes and culture plates were
placed in the freezer at -20°C for precooling. Next day a bucket was filled with ice and Ma-
trigel, pipettes and culture plates were placed on ice to keep them cold. All experiments with
Matrigel coating were perfomed with 6-well plates. About 500 µl of Matrigel were added per
well of a 6-well plate (50µl/cm2) and culture plate was placed for one hour at 37°C to allow
Matrigel to harden. Afterwards cells were seeded at a density of 30,000/cm2 (see Table 4) in
EC medium to examine EC differentiation and capillary formation capacity.
HUMAN DERMAL MICROVASCULAR ENDOTHELIAL CELL (HDMEC) ISOLATION
HDMECs were isolated from freshly excised human skin at the Department of Plastic Sur-
gery and Hand Surgery - Burn Center from patients who underwent elective operations (e.g.
abdominoplasty, breast reduction, flap surgery). The unprocessed skin was placed on a ster-
ile Petri dish with the epidermal, smooth side facing upward, and was cut into small pieces of
5 x 5 mm2 with a scalpel and digested overnight with dispase II at 4°C. Afterwards the skin-
dispase mixture was incubated under constant shaking at 37°C for 30 minutes. Dispase II
enables the separation and removal of the epidermis from the dermis. With sterile tweezers
skin pieces were transferred in a fresh tube with 20 ml cold PBS2- to stop the dispase diges-
tion. After transferring onto a petri dish the epidermis was separated from the dermis by care-
fully peeling back the epidermal layer using sterile tweezers. The dermis was minced with
scissors into small pieces in a 50 ml tube. The dermal layer was then digested in prewarmed
0.2% collagenase CLS type I with 1.5% BSA in collagenase buffer (sterile filtered) for 90
minutes at 37°C under constant shaking. Cell suspension was filtered through a 250 µm

59
mesh to remove contaminating fibrous tissue and afterwards culture medium was added.
After centrifugation at 400 g for 10 minutes the cell pellet was resuspended in HDMEC me-
dium and cells were seeded on fibronectine-coated culture dishes. Cells were washed twice
after 24 hours with 0.9% NaCl to remove possible blood contamination and cultured to sub-
confluency. At subconfluency cells were washed twice with PBS2- and incubated with Tryp-
sin/EDTA for two to three minutes and stopped with medium containing 10% FCS. Detached
cells were counted with a Neubauer counting chamber and separated with CD31-Dynabeads
and CD31-Microbeads as described earlier. Afterwards the CD31 positive HDMECs were
seeded on fibronectin-coated chamber slides and 6-well plates in HDMEC medium. They
were used as positive control for endothelial differentiation assays on MatrigelTM Basement
Membrane Extracellular Matrix as well as flow cytometry analysis.
HUMAN DERMAL FIBROBLAST ISOLATION
Fibroblasts were isolated following the same procedure as with HDMECs from human skin.
After separation with CD31 Dynabeads/Microbeads, the CD31 negative fibroblasts were
seeded on chamber slides, 6-well plates, and T75 culture flasks in fibroblast medium as
negative control.
HUMAN UMBILICAL VEIN ENDOTHELIAL CELL ISOLATION
HUVECs were isolated from human umbilical veins that had been received as a gift from the
Department of Pathology. The injured ends of each vein were removed and a feeding needle
was carefully inserted into the vein and fixed with a clamp. Afterwards the vein was rinsed
with 20 ml HEPES/6 mM EDTA via a syringe to remove possible blood contamination. An-
other feeding needle was inserted at the other end of the vein, fixed and the flow through
was checked with another 20 ml of HEPES/6 mM EDTA. A second syringe was filled with 5
ml trypsin/EDTA (37 °C warm) to replace the first one. Trypsin/EDTA was injected into the
umbilical vein and the cord was inserted in U-shape into a 500 ml vial filled with 37°C pre-
warmed PBS2-. After incubation for 20 minutes at 37°C in the water bath, the cord was
placed on a sterile operation cloth and massaged for five minutes to release HUVECs from
the vessel wall. 8 ml HEPES/6 mM EDTA was filled into another syringe and the syringe was
connected to the feeding needle. The cord vein was flushed thoroughly and the eluted fluid
was collected in a 50 ml tube containing 2 ml FCS to stop the digestion. After centrifugation
at 400g for 5 minutes at room temperature cells were resuspended in HUVEC medium and
subsequently seeded on fibronectin-coated T75 culture flasks. They were used as positive
control for endothelial differentiation assays on MatrigelTM Extracellular Matrix as well as flow
cytometry analysis.

60
FLOW CYTOMETRY ANALYSIS
To perform flow cytometry analyses, freshly isolated cells were incubated for 15 minutes with
erythrocyte lysis buffer at 4°C. For staining with DiI, cells were washed with PBS2-, and incu-
bated with DiI-ac-LDL (acLDL) (10 µg/ml) for one hour at 37°C. Afterwards cells were
washed twice with plain medium, once with PBS2-, and fixed with 4% paraformaldehyde
(PFA). Next, cells were centrifuged at 400g for 10 minutes and resuspended in PBS2- con-
taining 0.2% FCS and 2 mM EDTA. For incubation with primary anti-human CD31, CD34,
KDR, CD51/61, vWF, S100, and CD105-FITC, cells were also fixed with 4% PFA, resus-
pended in PBS containing 0.2% FCS and 2 mM EDTA, and kept at 4°C for 30 minutes.
Washing and incubation with secondary FITC-conjugated antibodies was performed for 30
minutes at 4°C with all samples except for the already FITC-labelled CD105. For additional
staining with CD31 APC-conjugated antibodies to generate double-labelling, cells were incu-
bated with CD31-APC for 10 minutes at 4 °C, washed and resuspended in PBS containing
0.2% FCS and 2 mM EDTA. Labelled cells were analyzed by flow cytometry using a FACS
Calibur apparatus (Beckton Dickinson) and CellQuest Pro analysis (Figure 14).
FL 3
SSC
FSC
FL 1
FL 2
FL 4 APC
PE
FITC
488nm
633nm
90° light scattergranularity
2°-16° light scattersize
ArgonLaser
Side scatter(SSC)
Forward scatter(FSC)
Light source
A.
B.
Figure 13. Flow cytometry
analysis
Flow cytometry analysis allows
simultaneous measuring of relative
cell size (FSC), cell granularity and
internal complexity (SSC) and
fluorescence intensity (A.). The
major components comprise a fluid
system, an optical system with
laser, reflector and filter device.
According to the fluorescence
staining applied wavelengths differ
between 450 nm and 700 nm. FITC
requires a laser wavelength of about
488 nm in the green range, con-
trariwise APC has its optimum at
633 nm in the red spectrum (B.).

61
Isotype controls for IgG1 and IgG2a were used as recommended by the supplier to exclude
non-specific binding of antibodies and were always negative. Each labelling with FITC-CD
markers was compared to control cells exclusively stained with the secondary FITC-
conjugated fluorescence antibody. The percentage of positively stained cells was determined
after correction for the percentage of cells reacting with the secondary FITC antibody. Stain-
ing of CD31+ as well as CD31- SVF cells was performed on the day of adipogenic and endo-
thelial differentiation induction.
IMMUNOSTAINING WITH VON-WILLEBRAND-FACTOR
For immunostaining with vWF cells were seeded on glass slides in chamber slides culture
dishes. Preceeding the staining procedure culture medium was removed from chamber
slides containing adherent cells and glass slides were washed twice with PBS2-. Cells were
fixed in cold acetone for 20 minutes at 4°C, acetone was removed again and glass slides
were dried at room temperature for a few minutes until completely parched. Glass slides
were placed into PBS2- and positioned into coverplates. Coverplates were rinsed once with
fresh PBS2- and each incubated with 100 µl primary anti-vWF antibodies for one hour at room
temperature. Antibodies against vWF were diluted 1:200 with PBS2-/1% BSA prior to use.
After washing three times with PBS2-, slides were stained with 100 µl FITC-conjugated sec-
ondary antibodies each for one hour at 37°C. FITC antibodies were diluted 1:30 with PBS/1%
BSA before use as well. After washing three times with PBS2-, staining of the cell nucleus
was performed by applying 100 µl DAPI solution for each coverplate and incubating for 15
minutes at room temperature. After washing three times with PBS2- glass slides were re-
moved from coverplates, coated with two drops of Mowiol 4-88 and covered with small glass
slides. Mowiol was hardened for a few hours at 4°C in the dark. Detection of vWF expression
was performed by fluorescence microscopy. vWF staining of CD31+ and CD31- SVF cells
was performed on the same day as adipogenic and endothelial differentiation induction to
ensure purity of cells used for differentiation assays.
DETERMINATION OF GLYCEROL-3-PHOSPHATE DEHYDROGENASE (GPDH) ACTIV-ITY
To evaluate preadipocyte differentiation on a molecular level, the activity of GPDH was
measured with a kit system from Takara. GPDH is a representative molecular key marker of
adipogenic conversion335, which has been used for decades to measure preadipocyte differ-
entiation298,368.

62
NADH/H+ NAD+
GPDH
cytosol
Dihydroxyaceton-phosphat
Alfa-Glycero-phosphat
GPOXFADFADH2
IMM
Qox Qred
To perform GPDH assays, culture medium was removed and cells were washed twice with
PBS2-. Cell lysis was induced by adding 1 ml “Enzyme Extraction Buffer” per T25 culture
flask. Culture flasks were incubated for three to five minutes at 37°C until cell lysis was com-
pleted. Afterwards the lysate was transferred to a 15 ml tube and diluted to two-fold by add-
ing 1 ml “Dilution Buffer”. To gain the desired four-fold dilution, 250 µl of the cell suspension
were added to another 250 µl “Dilution Buffer” in a fresh 1.5 ml Eppendorf cup. The samples
are stable for one hour on ice if necessary. 25 µl of each four-fold diluted sample was trans-
ferred into each 96-well. GPDH activity was determined by measuring absorbance at 340 nm
with an ELISA reader. The 96-well plate was inserted into the reader and 100 µl “Substrate
Solution” was pipetted to each sample via multi-channel pipette. As the reaction starts imme-
diately after mixing the cell suspension with “Substrate Solution”, it was substantial to add the
“Substrate Solution” quickly to each 96-well. This was achieved by the application of a multi-
channel pipette. Next, the ELISA reader was started at once to measure the GPDH activity.
The enzyme activity was measured ten times during the redox reaction. GPDH activity was
calculated with the following formula supplied by Takara:
GPDH activity (units / ml) = ΔOD340 x 2.06 x dilution ratio of the sample
coenzyme for the reversible conversion between both substrates and can be measured by the GPDH assay through the secondary
decrease of NADH. GPDH activity is calculated by measuring the reduction of NADH via declining absorbence at 340nm.
Dihydroxyacetone phosphate + NADH + H+ → Glycerol-3-phosphate + NAD+
Figure 14. Enzymatic reactions in glycerol-3-
phosphate dehydrogenase (GPDH) assay
Sequential binding of the fatty acids activated as
acyl-CoAs to glycerol-3-phosphate is a major step in
lipid biosythesis. In the liver glycerol-3-phosphate is
directly synthesised from glycerol with the help of
the glycerol kinase. Due to the absence of the glycerol
kinase in adipose tissue und muscle, glycerol-3-
phosphate is produced by the reduction of dihy-
droxyacetone phosphate in combination with
NADH. As dihydroxyacetone phosphate is an inter-
mediate product in the glycolytic pathway, lipid
synthesis in adipose tissue and muscle cannot pro-
ceed without glucose. The enzyme which catalyses
the conversion between dihydroxyacetone phosphate
and glycerol-3-phosphate is the glycerol-3-phosphate
dehydrogenase (GPDH). GPDH requires NAD as a

63
OIL RED O STAINING
Oil Red O staining is an established staining method for intracellular lipid vacuoles. This
method stains lipid droplets with a red colour so that even small droplets can be identified in
the cytoplasma and the percentage of lipid-containing cells can be validated. Besides lipid
droplets intracellular stress vacuoles are stained slightly red as well when incubated with Oil
Red O. The staining intensity of stress vacuoles is minor to that of lipids and these vacuoles
are extremely small in diameter so that they can be distinguished from lipid droplets in light
microscopy examination.
Oil Red O dye (0.5 g) was diluted in 100 ml of 99% isopropanol. This stock solution was
mixed 6:4 with aqua bidest and stored for 24h at room temperature. Afterwards the solution
was filtered to remove small dye particles. Monolayer cultures were washed twice with PBS2-
and fixed with 10% cold PFA and incubated for five hours at 4°C. After removing the PFA Oil
Red O working solution was added to culture flasks and samples were kept at room tempera-
ture for at least two hours. After washing twice with PBS2- the stained cells were kept in 10%
PFA and examined by light microscopy to count lipid-filled adipocytes.
CELL COUNTING
To dertermine (i) the percentage of differentiated to undifferentiated cells, (ii) to detect those
cells attached to Dynabeads as well as (iii) cells stained for vWF under the fluorescence mi-
croscopy, cell counting was performed under the phasecontrast or fluorescence microscope
with 100x magnification.
Cells were counted in six randomly chosen fields (six fields per culture flask). Differentiated
and undifferentiated cells were counted to determine the percentage of both cell types and
compare the differentiation level between different wells. The same was done with cells car-
rying Dynabeads on their surface and vWF positive cells appearing green under fluores-
cence light.
STATISTICAL EVALUATION
Data of expression of surface markers and GPDH activity were expressed as mean value ±
standard deviation. The significance of differences between CD31+ and CD31- cell differen-
tiation was evaluated using the t-test. Differences with p<0.05 were considered significant
and p<0.01 highly significant.

64
RESULTS
COMPARISON OF EXCISED FAT TISSUE VERSUS LIPOSUCTION MATERIAL
The surface expression of freshly isolated SVF cells from liposuction and excision was char-
acterized by flow cytometry without performing any previous purification. SVF cells harvested
from excised fat tissue show strong labelling for CD34 and a less intensive staining for KDR
(Figure 15 A., Table 5). These cells also comprise a small population of 5-10% of CD31 and
CD51/61 positive cells. SVF cells derived from liposuction aspirate (Figure 15 B.) are positive
for CD34, similar to cells from excised tissue (compare Table 5). CD31 and CD51/61 fluores-
cent labeling was stronger in liposuctioned SVF cells with up to 40% CD31 positive cells
(p=0.05 compared to CD31 expression in excised tissue). KDR was much more positive in
lipoaspirate than in excised fat (p<0.01).
Following standard culture conditions for two weeks in DMEM/F12 containing 10% FCS, cells
from lipoaspirate remained positive for CD34 in the same percentage as freshly isolated cells
while CD34 expression in excised fat decreased. Endothelial cell marker CD31 decreased in
liposuction and excised material by about 50%, CD51/61 shows a similar tendency (Table 5).
During cell cultivation, KDR expression or selection, respectively, increased in excised tissue
cells but decreased in liposuction material.
Fibroblasts, HDMECs and HUVECs were used as controls. Fibroblasts were negative for
CD31, vWF, CD51/61, KDR, S100, acLDL, positive for CD105, and slightly positive for
CD34. HDMECs showed positive results for CD31, vWF, acLDL, CD34, and KDR, as well as
slightly positive staining for CD51/61. HUVECs were positive for CD31, acLDL, CD34, KDR,
and negative for CD51/61 (Figure 21). Isotype controls for IgG1 and IgG2a were always
negative (Figure 15).

65
CD51
A. CD31 CD34
KDR
CD31
CD51
CD34
KDR
B.
FITC control IgG1 IgG2a
FITC control IgG1 IgG2a
Figure 15. Characterisation of SVF cells from excised versus liposuctioned adipose tissue
SVF cells were isolated from excised (A.) and liposuctioned (B.) adipose tissue by collagenase digestion and immedi-
ately analyzed by flow cytometry without any previous purification or culturing. Cells were screened for the expres-
sion of CD31, CD34, CD51/61, and KDR. IgG1 and IgG2a were used as isotype controls. Also, staining with secon-
dary FITC-conjugated fluorescence antibodies (FITC control) only was monitored.

66
Table 5. Expression of surface markers on adipose tissue derived SVF cells
SVF cells were isolated from excised adipose tissue or liposcution material, respectively, as described in Figure 15, and cul-
tured in DMEM/F12 plus 10% FCS for 14 days, followed by flow cytometry analysis. Given is the expression of the surface
markers CD31, CD34, CD51/61, and KDR in percentage. Fibroblasts were used as negative, HDMECs and HUVECs as
positive control. Data are derived from three individual experiments.
excision
day 0
excision
day 14
liposuction
day 0
liposuction
day 14
CD31+ 10% ± 2% 6% ± 3% 24% ± 15% 12% ± 5%
CD34+ 57% ± 3% 36% ± 10% 57% ± 20% 57% ± 4%
CD51/61+ 4% ± 1% 5% ± 2% 11% ± 8% 7% ± 3%
KDR+ 14% ± 5 52% ± 8 43% ± 23% 23% ± 7%

67
CHARACTERISATION OF SURFACE MARKERS AND MORPHOLOGICAL CHANGES IN SVF CELLS IN CULTURE
The developement and changes in surface marker expression under standard culture condi-
tions were measured after one and two weeks (Figure 16). Preadipocytes loose their differ-
entiation capacity when cultured in standard medium for several passages. Here we focused
additionally on stem cell markers (CD34, KDR and S100) to evaluate the differentiation po-
tential after long term culture (compare also Table 5).
Unpurified SVF cells from liposuctioned and excised fat tissue were cultured in DMEM/F12
plus 10% FCS for either one or two weeks and flow cytometry was performed. After one
week CD34, KDR and S100 were still fully expressed by SVF cells (Figure 16 A.), after two
weeks all three stem cells markers decreased (Figure 16 B., Table 5).
A.
B.
CD34 KDR
CD34 KDR
S100
S100
Figure 16. Flow cytometry analysis of preadipocytes during culture
SVF cells without prior purification were cultured in DMEM/HAM´s F12 with 10% FCS. Changes in surface
protein expression during proliferation were measured by flow cytometry for CD34, KDR and S100 on day 7 (A.)
and day 14 (B.).

68
IN VITRO DIFFERENTIATION OF UNPURIFIED SVF CELLS UNDER ENDOTHELIAL ME-DIUM CONDITIONS
Freshly isolated preadipocytes responded significantly different to the cultivation in the vari-
ous EC media (Figure 17). SVF cells were tested for their EC development on plain plastic
culture dishes and were stained with vWF and DAPI. In medium 1 (EC-1) only minor prolif-
eration and no formation of capillary structures was detected in vWF positive cells (Figure 17
A.-D.). Medium 2 (EC-2), in contrast, induced substantial proliferation and enhanced tubular
network formation of vWF positive cells (Figure 17 E.-H.). Further positive stained cells
showed capillary structures with widespread network connections (Figure 17 F.). Medium 3
(EC-3) also allowed significant cell proliferation but no network formation (Figure 17 I.-M.).
Standard proliferation medium for preadipocytes showed equal results in proliferation and
tubular structure formation as medium EC-1 (data not shown).

69
EC medium 1 EC medium 2 EC medium 3
A.
B.
C.
D.
E.
F.
G.
H.
I.
K.
L.
M.
Figure 17. Preadipocytes in different EC media without prior purification
Adipose progenitor cells were screened for EC development on plastic culture dishes by immunofluorescence staining with
vWF and DAPI. All pictures are vWF images except for D., H., M. (vWF-DAPI merged), A.-D. Medium 1, E.-H. Medium
2, I.-M. Medium 3. Magnification in A., E., I. is x20, in B., F., K. is x50, in C., G., L., D., H. and M. is x100.

70
SPLITTING AND PURITY ANALYSIS OF CD31- AND CD31+ SVF CELLS
To further characterize the unpurified SVF population, von-Willebrand factor immunofluores-
cence staining was performed 12h after cell isolation and culture in DMEM/F12, 10% FCS.
Freshly harvested SVF cells from excised fat tissue contain a small amount of approximately
10% of vWF positive (Figure 18 A.) and CD31 positive cells, as revealed by microscopical
counting and flow cytometry analyses (compare Table 5). Cultured under endothelial differ-
entiation conditions (EC-2) in the presence of VEGF and IGF-1, these SVF cells develop
tubular structures on plastic without Matrigel™. This was demonstrated by vWF-
immunofluorescence staining after culturing for two weeks (Figure 18 B.). Following separa-
tion into CD31- and CD31+ cells by Dynabeads or MicroBeads, respectively, CD31- SVF cells
show no expression of vWF or CD31 (Figure 18 C., E.), neither after one nor after two weeks
culturing in proliferation medium. Furthermore, CD31- cells do not reveal endothelial network
formation on plastic culture dishes (Figure 18 C.). Analyzing expression of S100, an early
marker of adipogenesis, these cells are found almost 100% positive in flow cytometry (Figure
18 D.-E.). CD31+ cells, in contrast, demonstrate strong vWF expression (97% ± 2% stained
cells) and CD105 staining after one week. The Dynabead isolation procedure demonstrates
to be effective since 95% ± 5% of the cells carry Dynabeads, confirming that they are posi-
tive for CD31 (Figure 18 F.-I., Figure 18 E.). Cells isolated with CD31 MicroBeads show
strong staining for CD31 in flow cytometry (Figure 18 L., N., P.). Furthermore CD31+ Mi-
croBead-isolated cells reveal the same surface expression characteristics for CD105 and
vWF as after isolation with Dynabeads. During proliferation, magnetic beads initially remain
bound to the cells and resist washing steps, however, cell proliferation reduces bead num-
bers continuously.

71
S100
E.
CD105
vWF
CD31 Dynabead/ MicroBead splitting
CD31- SVF
freshlyisolatedSVF cells(excisedfat tissue)
CD31+ SVF
cultured in EC mediumwithoutCD31 splitting
A. B.
C.
F.
90% 10%
D.
G.
H.
I.
CD31+ SVF
CD31+ SVF
CD31+ SVF
L.
M.
S100
CD
31
CD105
CD
31C
D31
vWF
CD34
K.
CD
31N.
CD34
P.
O.
Figure 18. Scheme of the application of CD31 Dynabead separation for SVF cells
Freshly isolated SVF cells from excised tissue were analyzed by fluorescence microscopy after isolation and transferred to
tissue culture dishes (A.). Cells were then either cultured in EC medium (EC-2) without Dynabead treatment and analyzed
again after two weeks (B.) or separated by CD31 Dynabeads or MicroBeads into a CD31- (C.-E.) and a CD31+ fraction (F.-
P.). To confirm the purity of the isolation procedure, flow cytometry was performed with CD31- cells double-stained for

72
CD31 and S100 expression (D. and E.) and with CD31+ cells double-labelled for CD31 in combination with CD105, vWF,
or CD34 (K.-P.). For fluorescence microscopy, cells were stained with vWF and DAPI. All pictures except for F. (vWF alone)
and I. (bright field and vWF, merged) are vWF-DAPI merged images. F.-I., K., M. and O.: Dynabead cell sorting, L., N. and
P.: MicroBead cell sorting. Arrows in H. and I. indicate Dynabeads attached to cell surfaces. Magnification in A., B., C., F.,
G. and I. is x200, in H. x400.
DIFFERENTIATION POTENTIAL OF CD31- AND CD31+ SVF CELLS TOWARDS EC LINEAGE
To evaluate endothelial differentiation potential of CD31- SVF cells morphologically, this cell
fraction was seeded onto Growth Factor Reduced Matrigel™ in EC-2 or EC-3 culture medium,
respectively, and tube formation was assayed. Cells form three-dimensional tube-like struc-
tures in both media (Figure 19 A.-D.), while EC-2 medium also allows formation of extensive
intercellular tube networks (Figure 19 C.-D.). CD31+ SVF cells showed equal results com-
pared to CD31- SVF cells when seeded on Matrigel™ ECM (data not shown). HUVECs, which
were used as a positive control, also formed networks but were not able to grow three-
dimensionally (Figure 19 E.-F., compare to Figure 19 C.).
Next, endothelial differentiation of CD31- cells grown in EC-2 or EC-3, respectively, was ana-
lyzed by flow cytometry to determine the quantitative expression of the surface markers
CD34, KDR, and acLDL (compare Table 6). In EC-3 medium CD31- SVF cells express CD34
by up to 46% and KDR up to 90%, demonstrating the capacity of this medium to enhance
stem cell characteristics of SVF cells (Figure 20 A.). In EC-2 medium CD34 and KDR ex-
pression is significantly lower (p=0.003 for CD34, p=0.006 for KDR) (Table 6, Figure 20 B.).
In both media, cells were positively stained for acLDL confirming differentiation towards an
endothelial phenotype (Figure 20).
HDMECs and HUVECs were used as positive (Figure 21), fibroblasts as negative control.

73
A. B.
C. D.
E. F.
►
►
►
►
200µm
200µm
200µm
200µm
200µm
200µm
►
►
►
Figure 19. Morphological analysis of endothelial differentiation of CD31- SVF cells and HUVECs
CD31- cells were isolated from the SVF by Dynabeads. Endothelial differentiation was performed in a stem cell medium (EC-
3, A.-B.) and an endothelial differentiation medium (EC-2, C.-D.) on Matrigel™. As control human umbilical vein endothe-
lial cells (HUVECs) were cultured in EGM-2 medium on Matrigel™ (E.-F.). Endothelial differentiation was analyzed by
light microscopy. Shown are bright field micrographs at a magnification of x100. ►, capillary tubules; , three-dimensional
networks.

74
A.
B.
CD34 KDR
acLDL
CD34 KDR
acLDL
Figure 20. Flow cytometry analysis of CD31- cells differentiation under optimized medium conditions
CD31- cells were isolated from the SVF and endothelial differentiation was induced by applying a stem cell medium (EC-3,
A.) or an endothelial differentiation medium (EC-2, B.), respectively. Quantitative expression of the surface markers CD34,
KDR, and acLDL was determined in flow cytometry.

75
culture medium EC-2 culture medium EC-3
CD34+ 2% ± 1% 37% ± 9%
KDR+ 19% ± 15% 78% ± 12%
acLDL 70% ± 5% 77% ± 7%
Table 6. Expression of surface markers on CD31- SVF cells under endothelial medium conditions
To evaluate differentiation of CD31- cells towards endothelium, CD31- cells were isolated from the SVF by Dynabeads and
grown in EC-2 or EC-3, respectively, followed by quantitative analysis of the expression of the surface markers CD34, KDR,
and acLDL (compare Figure 20). Data are from three individual experiments.
CD34 KDR
CD31 CD51/61
acLDL
Figure 21. Flow cytometry analysis of HUVECs
HUVECs were used as control in flow cytometry analysis for the presence of endothelial cell markers as CD31, CD51/61,
CD34, KDR and acLDL uptake.

76
IN VITRO DIFFERENTIATION OF SVF CELLS UNDER ADIPOGENIC MEDIUM CONDI-TIONS
Unseparated SVF cells from excised tissue differentiate under adipogenic culture conditions
to almost 90% into adipocytes showing large lipid vacuoles after 21 days in adipogenic me-
dium (data published elsewhere). Regarding the separated SVF cell fractions, the CD31-
cells could differentiate into mature adipocytes by nearly 100%, as demonstrated by light
microscopy and Oil Red O staining in Figure 22 A.-D.. Interestingly, purified CD31+ mature
endothelial cells from the SVF also convert into adipocytes after 21 days under adipogenic
culture conditions. Undergoing differentiation, cells are viable and healthy with no signs of
apoptosis present. From microscopical cell counting, approximately 65% of CD31+ cells pre-
sented accumulation of lipid vacuoles equivalent to an adipocyte phenotype. As can be seen
from Figure 22 E.-G., CD31+ cells have CD31 Dynabeads attached to their cell membranes
which confirms their CD31 surface expression. When comparing GDPH activity of both CD31
subsets, CD31+ cells showed a GPDH activity of 64% ± 11% compared to GPDH levels of
CD31- cells set 100% (Figure 23). Lipid droplets appeared approximately 2-3 days later dur-
ing differentiation in CD31+ than in CD31- cells. Applying flow cytometry, the proliferating
CD31+ cell fraction was screened for the presence of the surface marker CD34 before induc-
ing adipogenic differentiation. As shown in Figure 18 O.-P., approximately 45% of the cells
express CD34.
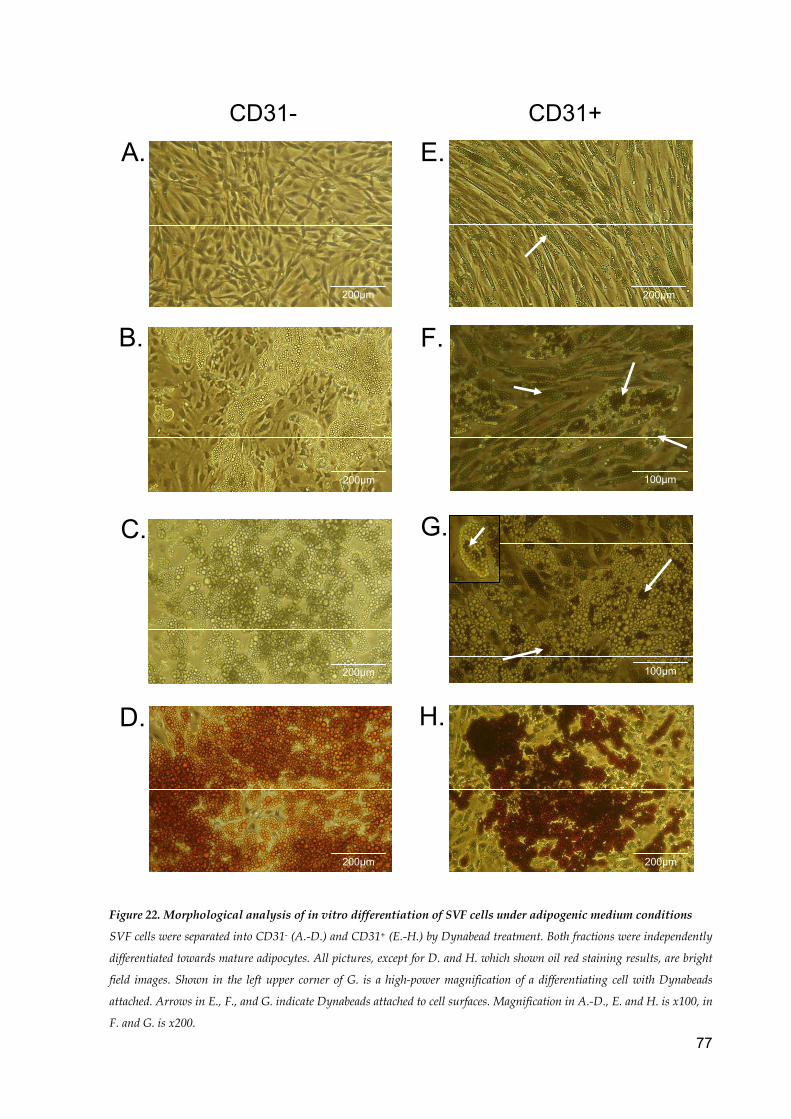
77
CD31- CD31+
A.
B.
C.
D.
E.
F.
G.
H.
200µm
200µm
200µm
200µm
200µm
100µm
100µm
200µm Figure 22. Morphological analysis of in vitro differentiation of SVF cells under adipogenic medium conditions
SVF cells were separated into CD31- (A.-D.) and CD31+ (E.-H.) by Dynabead treatment. Both fractions were independently
differentiated towards mature adipocytes. All pictures, except for D. and H. which shown oil red staining results, are bright
field images. Shown in the left upper corner of G. is a high-power magnification of a differentiating cell with Dynabeads
attached. Arrows in E., F., and G. indicate Dynabeads attached to cell surfaces. Magnification in A.-D., E. and H. is x100, in
F. and G. is x200.

78
0
20
40
60
80
100
120
CD31- Diff CD31+ Diff
Ext
ento
f diff
eren
tiatio
nre
lativ
e to
CD
31- c
ells
[%]
*Figure 23. Quantitative analysis of in vitro
differentiation of SVF cells in adipogenic
medium
SVF cells were separated into CD31- and
CD31+ cells by Dynabeads, independently
cultured in proliferation medium, and differen-
tiated at confluence. Differentiation of the
CD31+ fraction as determined by GPDH activ-
ity was measured after 3 weeks relative to
CD31- cells. Results are from three individual
experiments. *, p=0.01 compared to standard
differentiation of CD31- cells.

79
DISCUSSION
The stromal vascular fraction (SVF) of adipose tissue is a heterogenous cell population and
therefore very hard to characterize. Although there are similar findings about the surface pro-
tein expression of SVF cells, much controversy remains. The cell fractions that have been
used for surface analyses and differentiation assays cannot be compared since some re-
search groups apply liposuction aspirates60,252 133 or excised fat tissue260,314; others use more
purified, cell-sorted fractions260,350, while still others have identified cell surface structures
from cells not before culturing for several passages60. This leads to cell fractions with differ-
ent surface characteristics and different proliferation and differentiation behaviours. In this
thesis SVF cells from lipoaspirate and excised fat were screened for the presence of CD34,
KDR, CD31 and CD51/61. The rational for this was threefold: to (i) illustrate the differences
between SVF cells isolated from excised versus liposuction material, (ii) to determine the
relative amount of endothelial and hematopoietic progenitor cells in the freshly isolated SVF,
and (iii) to be aware of potential cellular contaminations, especially endothelial cells, which
might cause a false-positive differentiation result. Additionally, cells were cultured for two
weeks in standard medium to promote proliferation and to evalutate possible changes of sur-
face protein expression under long-term culture conditions.
After isolation of SVF cells the cell pellet is naturally contaminated with endothelial cells.
When measuring the differentiation potential of SVF cells into endothelial cells, already in-
corporated ECs might mimic a successful transdifferentiation of SVF cells. Therefore, it is not
possible to evaluate whether endothelial cells were transdifferentiated from SVF cells or if
they were already present in the applied SVF cell fraction before starting the experiment. To
determine the growth potential of contaminating ECs after isolation of SVF cells, different EC
growth media and a standard proliferation medium for preadipocytes were tested.
To analyze the differentiation plasticity of SVF cells towards endothelial structures and adi-
pose tissue, exclusively purified cell fractions were used to exclude cell contamination and
allow a solid analysis. Therefore, the cell pellet was splitted into a CD31+ and a CD31- cell
fraction using paramagnetic beads. After confirming purity of both cell fractions, CD31+ and
CD31- SVF cells were cultured in EC media or adipogenic differentiation medium. For endo-
thelial differentiation only EC-2 and EC-3 medium were applied as these two media showed
best potential for EC proliferation and differentiation according to Study 3.

80
TISSUE ENGINEERING WITH ADIPOSE DERIVED ADULT STEM CELLS
Soft tissue defects remain a challenge and often difficult problem in plastic and reconstruc-
tive surgery. There is high clinical need for adequate reconstruction of extended soft tissue
defects as found after deep burns, trauma, or tumor resections.
The application of autologous fat tissue to repair soft tissue defects seems most reasonable
in its approach as excess amounts of adipose tissue are found all over the human body. Adi-
pose tissue can be easily obtained through liposuction or elective operations and has been
used for transplantation or in soft tissue tissue engineering projects. Although there is re-
markable progress in this research field, serious problems remain. Transplantation of autolo-
gous adipose tissue to a defect site often results in 40-60% of graft volume loss over time.
One reason for significant tissue resorption is that large fat grafts never acquire sufficient
vascularization. Inadequate tissue vascularization restrains the supply of oxygen and nutri-
ents to the graft followed by consecutive tissue destruction406 and long-term tissue resorpti-
on304,303. Therefore the use of mature fat tissue for transplantation has not been consistently
successful in patients58,95,108,201,304,305,362.
More promising in the context of soft tissue reconstruction is the use of adipose tissue de-
rived progenitor cells seeded on biomaterials. By harvesting viable cells from patients healthy
autologous preadipocytes can be expanded through cell culture replication. Cells may then
be seeded onto scaffolds which need to support cell growth, proliferation and differentiation
of the progenitor cell fraction into mature fat cells. The scaffold construct is normally ab-
sorbed over time due to its biodegradable material and ideally, exclusively mature, well-
vascularized healthy adipose tissue remains.
Preadipocytes are deemed more advantageous as cell source for adipose tissue engineering
than mature adipocytes because they can be easily obtained, cultured, and expanded. Due
to the ability to generate large cell numbers and finally differentiate into various cell types
other than adipocytes97,157,162,433,434, preadipocytes are thought to have high potential in the
future development of soft tissue engineering.
PRESENT PROBLEMS AND HURDLES
Though several strategies using polymeric materials seeded with preadipocytes have been
successful to induce adipose tissue formation in vivo, one often quoted shortcoming with
these strategies is delayed growth in volume of the tissue constructs. This has been attrib-
uted mainly to slow vascularisation of the tissue leading to tissue resorption, oil cysts and
fibrosis253,307,363. Oxygen and nutrients supply are the limiting factors for the survival of trans-
planted cells. Especially adipose tissue engineering fails due to inadequate blood supply as
mature fat cells face cell death if they do not get in contact with surrounding capillaries by

81
inosculation within the first four days363,408. In conclusion, sufficient vascularisation and ap-
propriate cellular differentiation are two key problems in soft tissue engineering.
A further problem in organ and tissue transplants which will come into place once cell differ-
entiation and vascularisation have been addressed is the immunological rejection of the do-
nated tissue by the patient in non-autologous methods or cell culture using FCS. Strategies
in tissue engineering ascertain possible solutions to these problems225, including the applica-
tion of preadipocytes instead of adipocytes due to better proliferation capacity34,304 and hy-
poxia tolerance in precursor cells406, the exertion of autologous serum during in vitro cell cul-
ture160,161 and improvement of applied biomaterials159 among others.
HOW TO MANAGE INSUFFICIENT VASCULARISATION
Current methods to induce neovascularisation, the formation of a new vascular network,
within a tissue-engineered construct reveal limitations. Methods for adipose tissue angio-
genesis include co-culture of adipose cells with ECs in tubular scaffolds to induce vessel
formation. Although growth factors help in the formation of capillary structures, the generated
vessels are not long lasting304,303. The creation of vasculature prior to implantation allows the
tissue to receive nutrients faster because it does not have to form its vasculature. However,
the tissue may still not receive the required nutrition47. Other vascularisation methods com-
prise ingrowing of host vessels into the tissue construct leading to revascularisation of the
implanted material, but again insufficient nutrient supply remains a problem for the center of
the scaffold304. The relationship between ADAS cells and ECs as well as the role of certain
growth factors are not sufficiently understood to proceed towards a viable graft of relevant
size. Additionally, the potential of ADAS cells to differentiate into adipocytes and ECs upon
availability of the proper stimuli could be the key for confecting a vascularised fat graft with
sustained and increased volume. Therefore, this thesis studied the differentiation potential of
cells to convert from adipose tissue to endothelium and vice versa via progenitor cells.
CHARACTERIZATION OF THE SVF
Taking the broad mixture and combinations of cell surface CD markers, it is logical that the
SVF is not a unique population, but rather a pool of various progenitor cells including mature
ECs, too.
ADAS cells exhibit differentiation plasticity similar to that of bone marrow stem cells, as men-
tioned in the introduction. The cells are multipotent adult stem cells and have the ability for
multilineage differentiation429,433,434. Further similarities between ADAS cells and BMSCs
have been described in the literature focussing on growth kinetics and expansion capacity,
cell senescence, and protein surface expression (compare Table 1).

82
As shown by flow cytometry, freshly isolated SVF cells from excised as well as liposuctioned
adipose tissue comprise a significant population of progenitor cells as shown by strong ex-
pression of CD34 and KDR (Figure 15). However, the SVF also contains a population of
CD31, CD51/61, and vWF positive cells, demonstrating a contamination with mature ECs.
CD31 and CD51/61 expression was stronger in liposuction SVF cells, with up to 40% CD31
positive cells, emphazising the higher amount of contaminating ECs in lipoaspirates than in
excised fat (5-10% ECs, Table 5). The vWF expression of 10% for excised tissue as deter-
mined by microscopical cell counting (Figure 18 A.) completely matches the findings on
CD31 and CD51/61 expression in flow cytometry (compare Table 5).
After cell culture in DMEM/F12 with 10% FCS for two weeks, cells from lipoaspirate remain
positive for CD34 to nearly the same percentage as freshly isolated cells while cells from
excised tissue show a significant reduction (p<0.05) (Table 5). The missing reduction in
CD34 after cell adherence in liposuction material is surprising and the cause of this still is
under investigation. Endothelial surface characteristics, such as CD31 and CD51/61, how-
ever, decrease during culturing (Table 5) in all conditions, which might be due to selection
effects from culture medium as well as lower proliferation capacity of CD31+ SVF cells. CD31
expression in Figure 15 A. appears rather homogenous compared to Figure 15 B., however,
this is due to the two different preparation methods. In Figure 15 A., excised fat lobules were
carefully isolated to only gain preadipocytes. Since this is macroscopically not possible, there
is a very small but homogenous fraction of endothelial cells displaying CD31 characteristics.
The presence of these two cell fractions causes a double-peaked flow cytometry curve. In
Figure 15 B., liposuction material was used, which contains a much higher amount of endo-
thelials cells due to its origin and the method of yield which generates the two-peak appear-
ance of the flow cytometry.
KDR expression in liposuction material is initially 43% ± 23% and decreases to 23% ± 7%,
perhaps due to culturing conditions. This is in accordance with the reduction of endothelial
markers. In excised adipose tissue, however, the findings of KDR expression remain contro-
versial since KDR expression immediately after isolation is surprisingly low and its increase
due to cultivation is not in agreement with liposuction results (Table 5).
In general stem cell markers (CD34, KDR and S100) on SVF cells decrease during long-term
culture in standard medium as demonstrated in Figure 16. This is in agreement with the find-
ing that preadipocytes loose their differentiation capacity after multiple passages.
To analyze CD31+ and CD31- differentiation characteristics and surface markers independ-
ently, unpurified SVF cells from excised adipose tissue were split by CD31 Dynabeads. Al-
ternatively, CD31 MicroBeads were applied for the cell separation if double-labeling was per-
formed.

83
Whether or not Dynabead attachment to the cell surface influences the differentiation poten-
tial of preadipocytes has not been analyzed so far. In order to minimize any impact of the
Dynabeads, concentration of beads was kept low to allow proper attachment of cells to cul-
ture dishes which is otherwise impaired. As Dynabeads might bind non-specifically to adher-
ent and phagocytic cells, all steps with Dynabeads were carried out at 4°C. At this tempera-
ture, cell activity is low and non-specific binding reduced to a minimum. To further minimize
non-specific binding, the bead-cell suspension was pipetted a few times after incubation
(prior to separation on the magnet) to loosen any non-target cells adhering to the beads. All
these procedures reduced non-specific binding of Dynabeads which was finally evaluated by
staining Dynabead-carrying cells for vWF to ensure the endothelial phenotype. Cell counting
with fluorescence and light microscopy revealed that 95 ± 5% of the cells carried Dynabeads
(Figure 18 F.-I.). Of this fraction, 97 ± 2% were positive for vWF. This is supported by flow
cytometry after MicroBead cell sorting (Figure 18 M.-N.). These results reveal the beneficial
effects of the above shown technical modifications in Dynabead usage and show that 92 ±
4% of CD31+ cells are vWF positive. Together with the high expression of CD105 in CD31+
cells (Figure 18 K.-L.), this confirms the mature endothelial cell phenotype and the unique-
ness of the CD31+ fraction.
Freshly harvested CD31- SVF cells, in contrast to CD31+ cells, show no expression of vWF
and CD31 as ECs would do, neither after one nor after two weeks of culture in proliferation
medium (data not shown). Furthermore, CD31- cells do not present endothelial network for-
mation on plastic culture dishes (Figure 18 C.). Flow cytometry analyses reveal that this cell
fraction is 100% positive for S100 (Figure 18 D.-E.). Even though S100 labels nervous tis-
sue, skeletal and cardiac muscle, malignant melanoma cells and some other cell types, it is
well accepted as an early marker of adipogenesis21. It can therefore be assumed that the
CD31- fraction consists of adipocytes in a precursor state, the original preadipocytes. Thus,
separation of SVF cells by CD31 Dynabeads generates one cell fraction with preadipocyte
characteristics (CD31-) and one with endothelial properties (CD31+). SVF cells have recently
been shown to dedifferentiate from mature fat cells into a progenitor cell type and to differen-
tiate from there into ECs314. This has high implications for angiogenesis-related and cardio-
vascular topics.
In conclusion the surface protein expression of SVF cells demonstrates characteristics of
hematopoietic stem cells, early adipogenesis, ECs and endothelial progenitor cells (see Fig-
ure 24).

84
S100
CD34
KDR CD31
CD51/61
CD133
CD144
CD105
earlyadipogenesis
endothelial cell
hematopoietic stem cell
SVF
ADAS CELL DIFFERENTIATION TOWARDS ENDOTHELIAL CELLS
SVF cells have the capacity to differentiate into multiple cell phenotypes including all three
germ sheet lineages as mentioned earlier2,55,69,84,107,291,314,433,434. In addition to the remarkable
differentiation capacity of SVF cells, these cells have recently been differentiated into an en-
dothelial phenotype when cultured under EC conditions60,260,314. The first study demonstrating
the capacity of SVF cells to differentiate into capillary-like structures was already accom-
plished in 1999 by Zangani et al.429. When cultured on reconstituted basement membrane
(RBM) from Engelbreth-Holm-Swarm mouse sarcoma, mammary stromal cells rapidly turned
into capillary structures containing an internal lumen. The network formation was intensified
by VEGF supplementation and inhibited by the anti-angiogenic agent suramin. Differentiated
mammary stromal cells expressed KDR as well as ZO-1, a tight-junction-associated protein.
Conversion into mature adipocytes was still possible when cultured under adipogenic condi-
tions. More recent studies used other ECM components than RBM, such as methylcellulose
or Matrigel Extracellular Matrix as well as the presence of VEGF in combination with miscel-
laneous other growth factors for the differentiation of ADAS cells into ECs. When cultured in
the presence of ECM proteins and VEGF, ADAS cells develop branched tubular structures
and express various EC markers, such as CD31, CD34, vWF, CD144, and eNOS60,260,314.
The differentiation of mesenchymal stem cells into mature ECs has great impact on clinical
applications as well as tissue engineering research, as diffusion can supply cells just over a
distance of about 150 µm with nutrition, which is insufficient in ischemic disease and also in
larger three-dimensional fat tissue constructs in tissue engineering26. New capillaries from
the surrounding tissue invade after a delay of about five days and are not able to reach the
Figure 24. Cluster of
Differentiation com-
parable to bone
marrow stem cells
Freshly isolated SVF
cells have nearly the
same surface character-
istics as stem cells
from bone marrow and
display mesenchymal
stem cells.

85
centre of the construct. This leads to cell necrosis especially in highly vascularized tissues
like adipose tissue31.
THERAPEUTIC ANGIOGENESIS
In 2004, Planard-Bernard et al. demonstrated a common origin for adipose tissue and endo-
thelium314. They postulated that adipocytes and ECs have a common progenitor; however,
the interlinking pathways between adipose tissue and endothelium and the differentiation
potential from one tissue into the other via progenitor cells have not been analyzed. So far it
has only been shown that SVF cells are able to perform metaplasia from mature fat cells into
endothelial cells314. This thesis focused on this concept and aimed to further analyze the
interlinking pathways between adipose tissue and endothelium.
ADAS cells show a remarkable potential to form capillary-like structures in vitro. Further,
ADAS cells resemble EC characteristics in in vivo models as well. Angiogenic potential of fat
tissue has been known and clinically applied for treating wounds and ischemic organs for
more than 400 years (first mentioned in 1585)360,432. Applied for therapeutic angiogenesis,
ADAS cells have a wide range of possible clinical applications in neovascularization266,380,
including peripheral vascular disease, ischemic cardiomyopathy and myocardial infarction262,
ischemic stroke193, diabetic retinopathy, acute tubular necrosis91,189 and transplant related
ischemia. ADAS cells have first demonstrated enhanced angiogenesis and improved vascu-
larization in hindlimb ischemia models in mice60,260,266,274,275,314,325. Two potential but nonex-
clusive mechanisms could be involved in the pro-angiogenic effect of ADAS cells. Injected
cells could directly contribute to incorporation into the vessel wall60,260,314, or cells release
angiogenic growth factors which improve angiogenesis48,274,314. The interaction of both is
most likely the cause for blood flow enhancement, increased capillary density, and improved
angiogenesis induced by ADAS cells.
To study the angiogenic potential of adipose tissue derived cells, Cao et al. used CD34-
CD31- Flk-1+ cells60 that displayed features of EPCs. Miranville et al. applied CD34+ CD31-
ADAS cells260. Most other studies utilised unpurified adipose mesenchymal stem
cells274,314,325. Regardless of the applied subpopulation, all ADAS cell fractions improved an-
giogenesis, vascularization and blood flow, although CD34+ cells seem to be superior to
CD34- cells in increasing capillary density in hindlimb ischemia260. CD34- cells induce CD34
expression in ischemic tissue and vessels60 supporting the fact that CD34 is a crucial marker
for neo- or revascularization. Administration of ADAS cells can potentially affect revasculari-
zation to a degree similar to the application of BMSCs260.
A recent study has introduced a new method to approach decreased cardiac function after
myocardial infarction with transplantation of monolayered ADAS cells onto the scarred myo-
cardium four weeks after ischemic injury262. After application of ADAS cells with cell sheet

86
technology, the engrafted sheet gradually grew shaping a thick layer of tissue and compris-
ing freshly emerging blood vessels and a couple of cardiomyocytes. Additionally, angiogene-
sis was initiated in a paracrine manner and cardiac function improved subsequently in rats
with myocardial infarction262.
Another hint for the close relationship between adipose tissue and blood vessels is given by
preadipocytes that transdifferentiate into smooth muscle cells (SMCs)1. By overexpression of
aortic carboxypeptidase-like protein (ACLP) in adipogenic precursor cells a smooth muscle-
like cell phentotype can be induced in SVF cells and clonal preadipocyte cell lines1. ACLP is
expressed in ECM components of collagen-rich tissues229 and is up-regulated during vascu-
lar SMC differentiation228. Under smooth muscle (SM) differentiation environment, with β-
mercapto-ethanol and ascorbic acid, preadipocytes express anti SM22a, SM myosin heavy
chain, caldesmon, SM α-actin and develop stress fibers1.
To further verify the theory on mesenchymal differentiation plasticity, we performed morpho-
logical studies on CD31- SVF cells seeded onto growth factor reduced Matrigel™ in different
culture media, EC-2 or EC-3, respectively. EC-2 and EC-3 were chosen according to Study 3
as these two media showed the best results in inducing endothelial cell morphology in SVF
cells. The first medium (EC-2) is a culture medium to induce differentiation towards endothe-
lial lineage260. The second culture medium (EC-3) used has been applied as a hematopoietic
and endothelial progenitor cell medium for peripheral blood progenitors125. CD31- cells, which
were initially 100% positive S100 (compare Figure 18 D.-E.), were able to form three-
dimensional tube like structures in both media (Figure 19 A.-D.). EC-2 medium additionally
allowed the formation of extensive intercellular tube networks (Figure 19 C.-D.). HUVECs
used as a positive control also formed networks but were not able to form three-dimensional
capillary-like structures (Figure 19 E.-F., compare to Figure 19 C.). Analyzing endothelial
differentiation of CD31- cells grown in EC-2 or EC-3 by flow cytometry, we found that in EC-3
medium CD31- SVF cells express CD34 up to around 40% and KDR of 80% (Figure 20 A.,
Table 6). EC-3 medium therefore seems to support endothelial precursor characteristics in
the CD31- cells. In EC-2 medium, in contrast, CD34 and KDR expression was significantly
lower (Figure 20 B., Table 6) demonstrating the capacity of this medium to enhance endothe-
lial differentiation to mature ECs and diminish endothelial precursor cell characteristics of
SVF cells. In both media cells were positively stained for acLDL confirming differentiation
towards an endothelial phenotype (Figure 20, Table 6). AcLDL labels endothelial progenitor
cells, vascular endothelial cells, and macrophages.
Adipose tissue contains a subtype of mesenchymal stem cells that owes the ability to differ-
entiate into mature ECs in vitro, migrate into peripheral blood vessels, enhance angiogenesis

87
and revascularization by differentiating into ECs in vivo. Cells with the same abilities derived
from bone marrow as well as from blood circulation have been described since 199717 and
were termed “endothelial progenitor cells” for their EC potential. EPCs from bone marrow are
positive for CD34, VEGFR-2/KDR, CD133 and have the ability to take up
acLDL17,74,125,171,181,301,333,397,398,424. During mobilization and subsequent blood circulation they
lose CD133 and start expressing CD31, vascular endothelial cadherin, and vWF. Although
most subpopulations of ADAS cells have unequal surface protein patterns, various unique
markers were detected on them that resemble EPC characteristics. But not only the surface
characteristics reveal similarities between EPCs und ADAS cells. ADAS cells seem to par-
ticipate in endothelial repair, neovascularisation and angiogenesis in almost the same man-
ner as EPCs. Indeed, ADAS cells and EPCs seem to be involved in the repair of damaged
endothelium60,260,314. However, little is known about the signals associated in directing cells to
the centre of injured endothelium. Both ADAS cells and EPCs are capable of transdifferentia-
tion into smooth muscle cells1,114 underlining the ability of vessel wall integration. EPCs may
participate in remodelling of ischemic cardiac tissue and regeneration of ischemic myocar-
dium by modulation of angiogenesis and myogenesis190,205,215,348 in an almost comparable
manner as ADAS cells262,373.
EPCEC
smooth muscle cell
damaged EC
ADASEC
smooth muscle cell?
damaged EC
Figure 25. Vessel wall repair
through EPCs and ADAS cells
Similar to endothelial progenitor
cells ADAS cells may take part
in the repair of injured vessel
walls. Some of the ADAS cells
may integrate into the endothe-
lial monolayer after transdiffer-
entiation into endothelial and
even smooth muscle cells.

88
Some research groups aimed to identify a cell population that exhibits EPC characteristics in
adipose tissue. A recent study has isolated KDR positive cells from human lipoaspirates,
enhanced KDR expression through incubation in a hematopoietic stem cell medium and sub-
sequently differentiated the KDR+ subpopulation towards the endothelial lineage252. The
KDR+ subpopulation of LPA cells, pronounced as adipose EPCs, differentiated into ECs of
about 95%, being positive for VE-cadherin, vWF, LDL-lectin and formed tubular structures.
Although the KDR+ cells had striking characteristics of EPCs, an initial phenotyping and
characterization of the EPC fraction (KDR+ cells) was not performed. Flow cytometry for this
cell fraction was only performed for KDR alone, CD34, CD133 and acLDL uptake were not
obtained. Therefore it remains questionable whether the KDR+ subpopulation contained adi-
pose derived EPCs.
In another study, KDR+ ADAS cells were isolated and induced to perform endothelial lineage
conversion by plain culturing in EC medium. Differentiated ECs expressed CD31, CD34, VE-
cadherin and eNOS and were capable of tubular structure formation on Matrigel extracellular
matrix60. Even though the results so far display tremendous insights into the specific differen-
tiation potential of various cells, further investigations are needed to reveal the true potential
of the different mesenchymal stem cell fractions from adipose tissue to undergo EPC differ-
entiation.
ENDOTHELIAL CELL DIFFERENTIATION TO ADIPOCYTES
CD31+ and CD31- were separately cultivated under adipogenic medium conditions to analyze
their ability to convert into adipocytes. As expected, CD31- cells fully differentiated into ma-
ture adipocytes (Figure 22 A.-D.). Interestingly, purified SVF CD31+ cells also differentiated
into mature adipocytes after 21 days under adipogenic culture conditions. As can be seen
from Figure 22 E.-G., CD31+ preadipocytes and maturing adipocytes have CD31 Dynabeads
attached to their cell membranes confirming their CD31 surface expression. Further they
were positive for CD105 and vWF, additionally confirming their origin from endothelial line-
age (Figure 18 F.-N., Figure 26). While CD31+ and CD31- cells present a different morphol-
ogy during proliferation (compare Figure 22 A., 22 E.), they look identical in differentiated
state as adipocytes (Figure 22 D., 22 H.). Numbers from microscopic cell counting well
match with the more precise findings from GPDH assays in which CD31+ cells show a GPDH
activity of 64% ± 11% compared to CD31- cells set 100% (Figure 23). Thus, these findings
present for the first time a CD31+ progenitor cell type that can either convert into EC or adi-
pocyte, depending on the culture conditions (Figure 26). The CD31+ cell fraction is also
CD34+, since CD34 was expressed by about 45% of cells in flow cytometry (Figure 18 O.-P.).
The fact that CD34 and GPDH percentages do not completely match is due to the incongru-
ency between cell number (represented by CD34-level) and cell volume (GPDH-level).

89
Preadipocytes
Adipose Tissue EC
Endothelial diff.
Adipogenic diff.
MatrigelTM
Adipogenic diff.
MatrigelTM
Endothelial diff.
CD31- S100+
CD31+ CD105+ vWF+
Figure 26. Morphological analysis of in vitro differentiation of CD31- and CD31+ adipose progenitor cells under
endothelial and adipogenic differentiation conditions
Adipose progenitor cells were separated by Dynabeads for ECs in a CD31- and CD31+ cell fraction, both fractions were inde-
pendently differentiated towards endothelial structures on Matrigel as well as towards mature adipocytes.
In conclusion, the results presented here that adipose tissue is a source of mesenchymal
progenitor cells with broad differentiation plasticity. It is not exclusively the CD31- S100+ cell
population in adipose tissue, the classical preadipocytes, which can undergo fat differentia-
tion, but also the CD31+ CD105+ vWF+ cell fraction, phenotypically adipose tissue derived
ECs (compare Figure 26). In turn, preadipocytes even though predetermined to naturally
differentiate to adipocytes, have the potential to mature to endothelial network structures.
Conclusively, the adipose SVF appears to be a pool of multipotent cells with high plasticity
and differentiation capacity towards endothelium as well as adipose tissue. This might give
rise to innovative strategies in further tissue engineering applications where the adipose SVF
is applied for both, engineering of fat tissue and vascular structures.

90
ARTERIOSCLEROSIS AND ADIPOSE TISSUE
Arteriosclerosis affects large and medium-sized muscular and elastic arteries. Pathological
conditions, such as endothelial dysfunction, vascular inflammation, accumulation of lipids,
cell death and calcification of the intima lead to an atherosclerotic plaque of the vessel wall
(Figure 27). By protruding into the internal artery, plaques minimize the luminal diameter
available for blood flow, which results in abnormalities of blood flow, vascular remodelling,
acute and chronic obstruction and subsequent decreased oxygen supply.
In the past atherosclerosis was seen as a bland lipid storage disorder, however, substantial
advances in basic and experimental science have highlighted the role of inflammation as well
as cellular and molecular components supporting plaque evolvement.
Blood leukocytes, mediator cells of inflammation and host defence, play a major role in local-
ising the earliest lesions of arteriosclerosis and initiate the process of chronic plaque forma-
tion. Healthy endothelium does physiologically not allow leukocyte binding (Figure 27 A.).
However, after endothelial injury at sites of shear stress in the circulation and atherogenic
diet, inclusion of lipoproteins in the artery wall with accumulation of lipids in the subendothe-
lial space takes place (Figure 27 B.). Augmented expression of selective surface adhesion
molecules on arterial ECs promotes enhanced leukocyte recruitment237,273. When vascular
endothelium becomes inflamed, increased expression of adhesion molecules as vascular cell
adhesion molecule 1 (VCAM-1) and intercellular adhesion molecule 1 (ICAM-1) lead to bind-
ing of leukocytes to the activated ECs and integrins mediate firmer attachment (Figure 27
C.). Further blood flow disturbance promotes production of proteoglycans in the vessel wall,
that can bind lipoprotein particles, easing their oxidative modification and therefore advantag-
ing an inflammatory response234. Proinflammatory cytokines expressed within the artheroma
provide a chemotactic stimulus to the adherent leukocyte and promote penetration into the
intima. Infiltration of immune cells together with proteoglycan changes result in fatty streak
accumulation. Certain inflammatory mediators (MCP-1, M-CSF) support expression of
macrophage scavenger receptors, promoting the adhesion of circulating monocytes entering
the intima. Monocytes convert into macrophages when taking up the modified lipoproteins
located in the intima. These lipid-laden macrophages together with T-lymphocytes form a
lipid core that is isolated from the blood flow by a fibrous cap generated by migrated
SMCs371. Activated leukocytes and intrinsic arterial cells release mediators regulating this
process, comprising growth factors and cytokines such as TGF-β, PDGF BB, TNF-α and -β
and interferon-γ42,361. Fibrogenic growth factors stimulate SMC replication and the production
of a dense ECM leading to a more advanced atherosclerotic lesion (Figure 27 D.)336. But
inflammatory mediators can further evoke the expression of collagenase by foam cells in the
intimal plaque which comprehends thinning of the fibrous cap, leaving it weak and suscepti-
ble to rupture. Final evolvement of the end-stage plaque implies also further accumulation of

91
inflammatory cells, specially macrophages and T-lymphocytes, apoptosis and necrosis of the
central part of the lesion and calcification, resulting in evolvement of a necrotic core (Figure
27 E.)371. Cross-talk between macrophages and T-lymphocytes heightens the expression of
the potent pro-coagulant tissue factor238. When thinning of the fibrous cap and blood flow
disturbance lead to plaque rupture, the tissue factor triggers fatal thrombus formation that
causes arterial occlusion and a subsequent acute ischemic event of the target organ.

92
A.
B.
C.
D.
E.
endothelial cell
smooth muscle cell,contractile
normal ECM
hypertrophic ECM/fibrous cap
LDL/ lipoproteins
lymphocyte
monocyte/blood macrophage
foam cell
lipid core with necrosis
smooth muscle cell,synthetic
Figure 27. Stages of atheroscle-
rosis
A. Healthy endothelium. Blood
leukocytes adhere poorly to normal
endothelium. B. Inclusion of lipo-
proteins in the artery wall after
biochemical injury and subsequent
lipid accumulation. C. Leukocyte
recruitment to the nascent athero-
sclerotic lesion. When the endothe-
lial monolayer becomes inflamed, it
expresses adhesion molecules that
bind leukocytes. Proinflammatory
cytokines expressed within
atheroma provide a chemotactic
stimulus to the adherent leuko-
cytes, directing their migration into
the intima. Inflammatory mediators
can augment expression of macro-
phage scavenger receptors leading
to uptake of modified lipoprotein
particles and formation of lipid-
laden macrophages, foam cells. D.
Lymphocytes join macrophages in
the intima during lesion evolution.
Leukocytes, together with resident
vascular wall cells, secrete cyto-
kines and growth factors that can
promote the migration and prolif-
eration of smooth muscle cells. E.
Inflammatory factors can inhibit
collagen synthesis and evoke the
expression of collagenase by foam
cells. These alterations thin the
fibrous cap, leaving it fragile and
susceptible to rupture.

93
Numerous studies suggest that adipose tissue may have a strong impact on most obesity-
related cardiovascular complications, since an interrelation between adipocyte secreted pro-
teins and insulin resistance, hypofibrinolysis, inflammation, thrombosis and artherosclerosis
becomes more and more evident. Adipose tissue controls not only energy metabolism, but
further displays a secretory tissue for adipocytokines, which modify vascular response, for
proinflammatory cytokines, and for PAI-1, abetting fibrin accumulation. Most of these factors
produced by AT illustrate a major clue in the evolvement of the metabolic syndrome in obe-
sity. The metabolic syndrome is defined as cumulative cooccurrence of visceral obesity,
dyslipoproteinaemia with increased LDL and triglycerides but decreased high density lipopro-
tein (HDL) and essential hypertension. The beginning of metabolic syndrome is dominated
by an insulin resistance of the insulin-dependent tissues and impaired glucose tolerance,
requiring an increased insulin level for cellular glucose utilisation. This hyperinsulinaemia
heightens the feeling of hunger, resulting in obesity and progression of atherosclerosis323.
Recently, even more constituents of the metabolic syndrome have been revealed, compris-
ing subclinical inflammation, hyperuricaemia and microalbuminuria.
Among the adipocytokines, leptin as well as adiponectin play a role in the development of
cardiovascular risk factors. Leptin is a glycoprotein produced by mature adipocytes and acts
as a central part in controlling body fat storage. As adipose mass increases, its plasma level
increases as well, resulting in constricted appetite and enhanced energy expenditure. But the
mechanism of leptin does not work as efficient as it should since obese subjects remain
obese despite raised leptin concentration, possibly due to leptin resistance comparable to
insulin resistance in obesity.
Adiponectin is also secreted by adipocytes and correlates positively with insulin sensitivity
levels. This supports a negative association with body weight as plasma levels decrease with
the degree of insulin resistance413. Adiponectin acts as an anti-inflammatory and anti-
atherosclerosis adipokine and accumulates in an injured vessel wall by specifically binding to
collagen type I, III and IV293. Further adiponectin plays a role as a scaffold of newly formed
collagen in myocardial remodelling after ischaemic infarction182 and reduces the expression
of adhesion molecules on endothelial including VCAM-1 and ICAM-1296, which allow initial
leukocyte recruitment in atherosclerosis237,273. It inhibits TNF production by proliferation inhi-
bition of myelomonocytic lineages and mature macrophage functions425, reduces oxidized
LDL-induced EC proliferation270, and suppresses macrophage conversion into foam cells295.
Lower risk of myocardial infarction comes therefore together with high levels of circulating
adiponectin, whereas subjects with atherosclerosis seem to have low adiponectin concentra-
tions217,278. In contrary, resistin produced by adipose tissue induces the expression of endo-
thelial adhesion molecules VCAM-1 and ICAM-1206,404.

94
THE IMPACT OF PROGENITOR CELLS IN ATHEROSCLEROSIS
Besides the classical hypothesis of the atherosclerotic process recent research findings indi-
cate the involvement of stem and progenitor cells from the blood circulation in the develop-
ment of arteriosclerotic lesions. Progenitor cells were found to take part not only in endothe-
lial repair but also in smooth muscle cell accumulation in the vessel wall. In animal models
the injection of EPCs or BMSCs resulted in an aggravation of atherosclerosis and unstable
plaque evolvement126,426. Atherosclerosis turns out to be a pathological process initiated by
endothelial cell death, for instance due to trauma, which is followed by EC replacement
through progenitor cell aggregation. LDL can easily accumulate in the intima during the dif-
ferentiation of progenitor cells, a process which takes several days to weeks. Further blood
mononuclear cells adhere to freshly differentiated ECs and migrate into the subendothelial
space. Meanwhile, blood and resident progenitor cells harboured in the adventitia migrate
into the intima, followed by proliferation and differentiation into SMCs. According to that pro-
genitor cells seem to be accountable for the formation of atherosclerotic lesions as a major
cell source by giving rise to undue SMC accumulation172,173,347. In conclusion, stem cells
seem not simply to participate in vascular repair, but further in atherosclerotic lesion forma-
tion via pathological remodelling of vessel walls183,250.
Additionally Daub et al. proved that human CD34+ progenitor cells differentiate into foam
cells and ECs through the regulation of platelets. A changed balance of platelet-mediated
differentiation of CD34+ progenitor cells into foam cells and ECs may thus play a critical role
in atherogenesis as well94.
Taking these research findings and the differentiation plasticity of adipose SVF cells demon-
strated in this thesis, it might be possible that not only blood progenitor cells, such as EPCs,
take part in atherosclerosis, but also progenitor cells from adipose tissue (compare Figure
25).
The broad differentiation plasticity of CD31+ SVF cells could be explained by the common
mesenchymal origin of endothelium and adipose tissue and might have further implications
on the pathogenesis of atherosclerosis. Previous literature findings link the development and
advancement of atherosclerotic lesions to endothelial cells which are thought to be involved
in LDL binding and metabolism391,400, thereby displaying adipocyte characteristics. The
pathogenic processes in atherosclerosis might be not exclusively performed by endothelial or
hematopoietic progenitor cells, but also in part by CD34+ adipose SVF cells as well as CD31+
cells showing adipocyte-related metabolic activities.

95
CONCLUSION
This thesis demonstrates that the term “preadipocyte” can be misleading if purification by cell
sorting or equivalent methods have not been applied. Depending on the method of yield the
amount of endothelial cells in the SVF varies between 10% ± 2% in excised fat and 24% ±
15% in lipoaspirate. The SVF is therefore not a unique cell fraction but a mixture of various
cells with varying surface protein expressions, one of which is a CD31-, S100+ cell type which
has the capacity to differentiate into adipocyte as well as endothelial precursor cell and ma-
ture EC, depending on the involved differentiation-inducing factors. In addition, the SVF fur-
ther contains a CD31+, CD34+, S100- cell fraction with an endothelial phenotype which can
be converted into adipose tissue under adequate differentiation conditions. These results
encourage new approaches in adipose tissue engineering, since the use of purified multipo-
tent SVF cells with adipogenic and angiogenic potential instead of applying the unpurified
SVF cell pool might allow better vascularisation and tissue growth results. Furthermore, the
findings of this thesis might have relevant impact on the pathogenesis of atherosclerosis
since the development and advancement of atherosclerotic lesions seems to involve not only
progenitor cells, but also endothelial cells displaying both, endothelial and adipogenic proper-
ties.
Preadipocytes Adipose EC
Adipocytes Capillaries
Figure 28. Plasticity of hu-
man stromal vascular cells to
perform adipogenic and endo-
thelial differentiation
Besides their natural branch of
differentiation to fat cells and
tubular structures, SVF preadi-
pocytes and endothelial cells can
also differentiate into other
phenotypes. Preadipocytes
(CD31-, S100+) can form tubular
structures on Matrigel, and
endothelial cells (CD 31+,
CD34+, S100-) can aquire a fat
cell phenotype expressing fat-
specific enzymes.

96
REFERENCES
1. Abderrahim-Ferkoune A, Bezy O, Astri-Roques S, Elabd C, Ailhaud G, Amri EZ. Transdifferentiation of preadi-pose cells into smooth muscle-like cells: role of aortic carboxypeptidase-like protein. Exp Cell Res. 2004;293:219-28.
2. Abderrahim-Ferkoune A, Bezy O, Chiellini C, Maffei M, Grimaldi P, Bonino F, Moustaid-Moussa N, Pasqualini F, Mantovani A, Ailhaud G, Amri EZ. Characterization of the long pentraxin PTX3 as a TNFalpha-induced se-creted protein of adipose cells. J Lipid Res. 2003;44:994-1000.
3. Aboody KS, Brown A, Rainov NG, Bower KA, Liu S, Yang W, Small JE, Herrlinger U, Ourednik V, Black PM, Breakefield XO, Snyder EY. Neural stem cells display extensive tropism for pathology in adult brain: evidence from intracranial gliomas. Proc Natl Acad Sci U S A. 2000;97:12846-51.
4. Ailhaud G. Early adipocyte differentiation. Biochem Soc Trans. 1996;24:400-2. 5. Ailhaud G, Amri E, Czerucka D, Forest C, Gaillard D, Grimaldi P, Negrel R, Vannier C. [Lipoprotein lipase and
adipocyte differentiation]. Reprod Nutr Dev. 1985;25:153-8. 6. Ailhaud G, Grimaldi P, Négrel R. Cellular and molecular aspects of adipose tissue development. Ann Rev
Nutr. 1992;12:207-233. 7. Akanbi KA, Brodie AE, Suryawan A, Hu CY. Effect of age on the differentiation of porcine adipose stromal-
vascular cells in culture. J Anim Sci. 1994;72:2828-35. 8. Alessi MC, Juhan-Vague I. PAI-1 and the metabolic syndrome: links, causes, and consequences. Arterioscler
Thromb Vasc Biol. 2006;26:2200-7. 9. Altiok S, Xu M, Spiegelman BM. PPARgamma induces cell cycle withdrawal: inhibition of E2F/DP DNA-
binding activity via down-regulation of PP2A. Genes Dev. 1997;11:1987-98. 10. Ambrosini G, Nath AK, Sierra-Honigmann MR, Flores-Riveros J. Transcriptional activation of the human leptin
gene in response to hypoxia. Involvement of hypoxia-inducible factor 1. J Biol Chem. 2002;277:34601-9. 11. Amri EZ, Ailhaud G, Grimaldi P. Regulation of adipose cell differentiation. II. Kinetics of induction of the aP2
gene by fatty acids and modulation by dexamethasone. J Lipid Res. 1991;32:1457-63. 12. Amri EZ, Dani C, Doglio A, Etienne J, Grimaldi P, Ailhaud G. Adipose cell differentiation: evidence for a two-
step process in the polyamine-dependent Ob1754 clonal line. Biochem J. 1986;238:115-22. 13. Amri EZ, Dani C, Doglio A, Grimaldi P, Ailhaud G. Coupling of growth arrest and expression of early markers
during adipose conversion of preadipocyte cell lines. Biochem Biophys Res Commun. 1986;137:903-10. 14. Aoki S, Toda S, Ando T, Sugihara H. Bone marrow stromal cells, preadipocytes, and dermal fibroblasts pro-
mote epidermal regeneration in their distinctive fashions. Mol Biol Cell. 2004;15:4647-57. 15. Aoki S, Toda S, Sakemi T, Sugihara H. Coculture of endothelial cells and mature adipocytes actively pro-
motes immature preadipocyte development in vitro. Cell Struct Funct. 2003;28:55-60. 16. Asahara T, Kawamoto A. Endothelial progenitor cells for postnatal vasculogenesis. Am J Physiol Cell
Physiol. 2004;287:C572-9. 17. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner JM.
Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964-7. 18. Asano A, Kimura K, Saito M. Cold-induced mRNA expression of angiogenic factors in rat brown adipose
tissue. J Vet Med Sci. 1999;61:403-9. 19. Asano A, Morimatsu M, Nikami H, Yoshida T, Saito M. Adrenergic activation of vascular endothelial growth
factor mRNA expression in rat brown adipose tissue: implication in cold-induced angiogenesis. Biochem J. 1997;328 ( Pt 1):179-83.
20. Ashjian PH, Elbarbary AS, Edmonds B, DeUgarte D, Zhu M, Zuk PA, Lorenz HP, Benhaim P, Hedrick MH. In vitro differentiation of human processed lipoaspirate cells into early neural progenitors. Plast Reconstr Surg. 2003;111:1922-31.
21. Atanassova P. Immunohistochemical expression of S-100 protein in human embryonal fat cells. Cells Tissues Organs. 2001;169:355-60.
22. Athanasou NA, Benson MK, Brenton BP, Smith R. Progressive osseous heteroplasia: a case report. Bone. 1994;15:471-5.
23. Aust L, Devlin B, Foster SJ, Halvorsen YD, Hicok KC, du Laney T, Sen A, Willingmyre GD, Gimble JM. Yield of human adipose-derived adult stem cells from liposuction aspirates. Cytotherapy. 2004;6:7-14.
24. Awad HA, Halvorsen YD, Gimble JM, Guilak F. Effects of transforming growth factor beta1 and dexa-methasone on the growth and chondrogenic differentiation of adipose-derived stromal cells. Tissue Eng. 2003;9:1301-12.
25. Awad HA, Wickham MQ, Leddy HA, Gimble JM, Guilak F. Chondrogenic differentiation of adipose-derived adult stem cells in agarose, alginate, and gelatin scaffolds. Biomaterials. 2004;25:3211-22.
26. Awwad HK, el Naggar M, Mocktar N, Barsoum M. Intercapillary distance measurement as an indicator of hy-poxia in carcinoma of the cervix uteri. Int J Radiat Oncol Biol Phys. 1986;12:1329-33.
27. Bacou F, el Andalousi RB, Daussin PA, Micallef JP, Levin JM, Chammas M, Casteilla L, Reyne Y, Nougues J. Transplantation of adipose tissue-derived stromal cells increases mass and functional capacity of damaged skeletal muscle. Cell Transplant. 2004;13:103-11.
28. Baksh D, Song L, Tuan RS. Adult mesenchymal stem cells: characterization, differentiation, and application in cell and gene therapy. J Cell Mol Med. 2004;8:301-16.
29. Ballas CB, Zielske SP, Gerson SL. Adult bone marrow stem cells for cell and gene therapies: implications for greater use. J Cell Biochem Suppl. 2002;38:20-8.
30. Barry FP, Murphy JM. Mesenchymal stem cells: clinical applications and biological characterization. Int J Biochem Cell Biol. 2004;36:568-84.
31. Bartynski J, Marion M, Wang T. Histopathologic evaluation of adipose autografts in a rabbit ear model. Oto-laryngol Head Neck Surg. 1990;102:314-21.
32. Bastelica D, Morange P, Berthet B, Borghi H, Lacroix O, Grino M, Juhan-Vague I, Alessi MC. Stromal cells are the main plasminogen activator inhibitor-1-producing cells in human fat: evidence of differences between visceral and subcutaneous deposits. Arterioscler Thromb Vasc Biol. 2002;22:173-8.

97
33. Baum D, Beck RQ, Hammer LD, Brasel JA, Greenwood MR. Adipose tissue thymidine kinase activity in man. Pediatr Res. 1986;20:118-21.
34. Beahm EK, Walton RL, Patrick CW, Jr. Progress in adipose tissue construct development. Clin Plast Surg. 2003;30:547-58, viii.
35. Benarroch IS, Brodsky M, Rubinstein A, Viggiano C, Salama EA. Treatment of blood hyperviscosity with calcium Dobesilate in patients with diabetic retinopathy. Ophtalmis Res. 1985;17:131-138.
36. Bennett JH, Joyner CJ, Triffitt JT, Owen ME. Adipocytic cells cultured from marrow have osteogenic poten-tial. J Cell Sci. 1991;99 ( Pt 1):131-9.
37. Benvenuti S, Saccardi R, Luciani P, Urbani S, Deledda C, Cellai I, Francini F, Squecco R, Rosati F, Danza G, Gelmini S, Greeve I, Rossi M, Maggi R, Serio M, Peri A. Neuronal differentiation of human mesenchymal stem cells: changes in the expression of the Alzheimer's disease-related gene seladin-1. Exp Cell Res. 2006;312:2592-604.
38. Beresford JN, Bennett JH, Devlin C, Leboy PS, Owen ME. Evidence for an inverse relationship between the differentiation of adipocytic and osteogenic cells in rat marrow stromal cell cultures. J Cell Sci. 1992;102 ( Pt 2):341-51.
39. Bergers G, Brekken R, McMahon G, Vu TH, Itoh T, Tamaki K, Tanzawa K, Thorpe P, Itohara S, Werb Z, Hanahan D. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat Cell Biol. 2000;2:737-44.
40. Bernlohr D, Angus C, Lane M, Bolanowski M, Kelly Tj. Expression of specific mRNAs during adipose differen-tiation: identification of an mRNA encoding a homologue of myelin P2 protein. Proc Natl Acad Sci U S A. 1984;81:5468-72.
41. Bikfalvi A, Klein S, Pintucci G, Rifkin DB. Biological roles of fibroblast growth factor-2. Endocr Rev. 1997;18:26-45.
42. Binder CJ, Chang MK, Shaw PX, Miller YI, Hartvigsen K, Dewan A, Witztum JL. Innate and acquired immunity in atherogenesis. Nat Med. 2002;8:1218-26.
43. Bishop AE, Buttery LD, Polak JM. Embryonic stem cells. J Pathol. 2002;197:424-9. 44. Bjorntorp P, Karlsson M, Pertoft H, Pettersson P, Sjostrom L, Smith U. Isolation and characterization of cells
from rat adipose tissue developing into adipocytes. J Lipid Res. 1978;19:316-24. 45. Björntorp P, Karlsson M, Petterson P, Sypniewska G. Differentiation and function of rat adipocyte precursor
cells in primary culture. J Lipid Res. 1980;21:714-723. 46. Blood CH, Zetter BR. Tumor interactions with the vasculature: angiogenesis and tumor metastasis. Biochim
Biophys Acta. 1990;1032:89-118. 47. Borges J, Mueller MC, Padron NT, Tegtmeier F, Lang EM, Stark GB. Engineered adipose tissue supplied by
functional microvessels. Tissue Eng. 2003;9:1263-70. 48. Borges J, Torio-Padron N, Momeni A, Mueller MC, Tegtmeier FT, Stark BG. Adipose precursor cells (preadi-
pocytes) induce formation of new vessels in fibrin glue on the newly developed cylinder chorioallantoic membrane model (CAM). Minim Invasive Ther Allied Technol. 2006;15:246-52.
49. Bornstein SR, Abu-Asab M, Glasow A, Path G, Hauner H, Tsokos M, Chrousos GP, Scherbaum WA. Immuno-histochemical and ultrastructural localization of leptin and leptin receptor in human white adipose tissue and differentiating human adipose cells in primary culture. Diabetes. 2000;49:532-8.
50. Bouloumie A, Drexler HC, Lafontan M, Busse R. Leptin, the product of Ob gene, promotes angiogenesis. Circ Res. 1998;83:1059-66.
51. Bouloumie A, Sengenes C, Portolan G, Galitzky J, Lafontan M. Adipocyte produces matrix metalloproteinases 2 and 9: involvement in adipose differentiation. Diabetes. 2001;50:2080-6.
52. Bourgeois F, Goldstein AL, Johnson PR. Lipogenesis in primary cultures of adipoblasts derived from geneti-cally obese Zucker rats. Metabolism. 1983;32:673-80.
53. Bruder S, Jaiswal N. The osteogenic potential of human mesenchymal stem cells is not diminished after one billion-fold expansion in vitro. Trans Orthop Res Soc. 1996;21:580.
54. Brun RP, Kim JB, Hu E, Altiok S, Spiegelman BM. Adipocyte differentiation: a transcriptional regulatory cas-cade. Curr Opin Cell Biol. 1996;8:826-32.
55. Brzoska M, Geiger H, Gauer S, Baer P. Epithelial differentiation of human adipose tissue-derived adult stem cells. Biochem Biophys Res Commun. 2005;330:142-50.
56. Bullo M, Garcia-Lorda P, Megias I, Salas-Salvado J. Systemic inflammation, adipose tissue tumor necrosis factor, and leptin expression. Obes Res. 2003;11:525-31.
57. Burton GR, Nagarajan R, Peterson CA, McGehee RE, Jr. Microarray analysis of differentiation-specific gene expression during 3T3-L1 adipogenesis. Gene. 2004;329:167-85.
58. Büttemeyer R, Hohle R, Meyer-Gattermann W, Bruck JC. Late complications following autologous and ho-mologous fat transplantation for breast augmentation. Eur J Plast Surg. 1994;17:23-27.
59. Cao R, Brakenhielm E, Wahlestedt C, Thyberg J, Cao Y. Leptin induces vascular permeability and synergisti-cally stimulates angiogenesis with FGF-2 and VEGF. Proc Natl Acad Sci U S A. 2001;98:6390-5.
60. Cao Y, Sun Z, Liao L, Meng Y, Han Q, Zhao RC. Human adipose tissue-derived stem cells differentiate into endothelial cells in vitro and improve postnatal neovascularization in vivo. Biochem Biophys Res Commun. 2005;332:370-9.
61. Caplan AI. Mesenchymal stem cells. J Orthop Res. 1991;9:641-50. 62. Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nat Med. 2000;6:389-95. 63. Carmeliet P, Collen D. Molecular basis of angiogenesis. Role of VEGF and VE-cadherin. Ann N Y Acad Sci.
2000;902:249-62; discussion 262-4. 64. Carmeliet P, Jain RK. Angiogenesis in cancer and other diseases. Nature. 2000;407:249-57. 65. Carraro R, Li ZH, Johnson JE, Jr., Gregerman RI. Islets of preadipocytes highly committed to differentiation in
cultures of adherent rat adipocytes. Light- and electron-microscopic observations. Cell Tissue Res. 1991;264:243-51.
66. Castellot Jj, Karnovsky M, Spiegelman B. Potent stimulation of vascular endothelial cell growth by differenti-ated 3T3 adipocytes. Proc Natl Acad Sci U S A. 1980;77:6007-6011.
67. Castellot JJ, Karnovsky MJ, Spiegelman BM. Differentiation-dependent stimulation of neovascularization and endothelial cell chemotaxis by 3T3 adipocytes. Proc Natl Acad Sci U S A. 1982;79:5597-5601.

98
68. Chapman AB, Knight DM, Dieckmann BS, Ringold GM. Analysis of gene expression during differentiation of adipogenic cells in culture and hormonal control of the developmental program. J Biol Chem. 1984;259:15548-55.
69. Charriere G, Cousin B, Arnaud E, Andre M, Bacou F, Penicaud L, Casteilla L. Preadipocyte conversion to macrophage. Evidence of plasticity. J Biol Chem. 2003;278:9850-5.
70. Chavey C, Mari B, Monthouel MN, Bonnafous S, Anglard P, Van Obberghen E, Tartare-Deckert S. Matrix metal-loproteinases are differentially expressed in adipose tissue during obesity and modulate adipocyte differen-tiation. J Biol Chem. 2003;278:11888-96.
71. Chawla A, Lazar MA. Peroxisome proliferator and retinoid signaling pathways co-regulate preadipocyte phe-notype and survival. Proc Natl Acad Sci U S A. 1994;91:1786-90.
72. Chawla A, Schwarz E, Dimaculangan D, Lazar M. Peroxisome proliferator-activated receptor (PPAR) gamma: adipose- predominant expression and induction early in adipocyte differentiation. Endocrinology. 1994;135:798-800.
73. Chen XL, Hausman DB, Dean RG, Hausman GJ. Hormonal regulation of leptin mRNA expression and preadi-pocyte recruitment and differentiation in porcine primary cultures of S-V cells. Obes Res. 1998;6:164-72.
74. Choi K, Kennedy M, Kazarov A, Papadimitriou JC, Keller G. A common precursor for hematopoietic and endo-thelial cells. Development. 1998;125:725-32.
75. Choi YC, Morris GM, Lee FS, Sokoloff L. The effect of serum on monolayer cell culture of mammalian articular chondrocytes. Connect Tissue Res. 1980;7:105-12.
76. Cinti S, Morroni M. Brown adipocyte precursor cells: a morphological study. Ital J Anat Embryol. 1995;100 Suppl 1:75-81.
77. Claffey KP, Wilkison WO, Spiegelman BM. Vascular endothelial growth factor. Regulation by cell differentia-tion and activated second messenger pathways. J Biol Chem. 1992;267:16317-16322.
78. Cohen B, Barkan D, Levy Y, Goldberg I, Fridman E, Kopolovic J, Rubinstein M. Leptin induces angiopoietin-2 expression in adipose tissues. J Biol Chem. 2001;276:7697-700.
79. Coleman R, Bell R. Selective changes in enzymes of the sn-glycerol 3-phosphate and dihydroxyacetone-phosphate pathways of triacylglycerol biosynthesis during differentiation of 3T3-L1 preadipocytes. J Biol Chem. 1980;25:255(16) 7681-7687.
80. Coleman S. Facial recontouring with lipostructure. Clin Plast Surg. 1997;??:347-367. 81. Conejero JA, Lee JA, Parrett BM, Terry M, Wear-Maggitti K, Grant RT, Breitbart AS. Repair of palatal bone
defects using osteogenically differentiated fat-derived stem cells. Plast Reconstr Surg. 2006;117:857-63. 82. Conget PA, Minguell JJ. Phenotypical and functional properties of human bone marrow mesenchymal pro-
genitor cells. J Cell Physiol. 1999;181:67-73. 83. Cornelius P, MacDougald OA, Lane MD. Regulation of adipocyte development. Annu Rev Nutr. 1994;14:99-
129. 84. Cousin B, Munoz O, Andre M, Fontanilles AM, Dani C, Cousin JL, Laharrague P, Casteilla L, Penicaud L. A
role for preadipocytes as macrophage-like cells. Faseb J. 1999;13:305-12. 85. Cowan CM, Shi YY, Aalami OO, Chou YF, Mari C, Thomas R, Quarto N, Contag CH, Wu B, Longaker MT. Adi-
pose-derived adult stromal cells heal critical-size mouse calvarial defects. Nat Biotechnol. 2004;22:560-7. 86. Crandall DL, Busler DE, McHendry-Rinde B, Groeling TM, Kral JG. Autocrine regulation of human preadipo-
cyte migration by plasminogen activator inhibitor-1. J Clin Endocrinol Metab. 2000;85:2609-14. 87. Crandall DL, Hausman GJ, Kral JG. A review of the microcirculation of adipose tissue: anatomic, metabolic,
and angiogenic perspectives. Microcirculation. 1997;4:211-32. 88. Croissandeau G, Chretien M, Mbikay M. Involvement of matrix metalloproteinases in the adipose conversion
of 3T3-L1 preadipocytes. Biochem J. 2002;364:739-46. 89. Cryer A. Tissue lipoprotein lipase activity and its action in lipoprotein metabolism. Int J Biochem.
1981;13:525-41. 90. Cryer A, Wusteman FS, Casey JJ. Glycosaminoglycan: cell interactions; their role in lipoprotein lipase secre-
tion from isolated cardiac muscle cells. Cell Biochem Funct. 1984;2:53-6. 91. da Silva LB, Palma PV, Cury PM, Bueno V. Evaluation of stem cell administration in a model of kidney ische-
mia-reperfusion injury. Int Immunopharmacol. 2007;7:1609-16. 92. Dardick I, Poznanski W, Waheed I, Setterfield G. Ultrastructural observations on differentiating human
preadipocytes cultured in vitro. Tissue Cell. 1976;8:561-571. 93. Darlington GJ, Ross SE, MacDougald OA. The role of C/EBP genes in adipocyte differentiation. J Biol Chem.
1998;273:30057-60. 94. Daub K, Langer H, Seizer P, Stellos K, May AE, Goyal P, Bigalke B, Schonberger T, Geisler T, Siegel-Axel D,
Oostendorp RA, Lindemann S, Gawaz M. Platelets induce differentiation of human CD34+ progenitor cells into foam cells and endothelial cells. Faseb J. 2006;20:2559-61.
95. Davis RE, Guida RA, Cook TA. Autologous free dermal fat graft. Reconstruction of facial contour defects. Arch Otolaryngol Head Neck Surg. 1995;121:95-100.
96. De Ugarte DA, Alfonso Z, Zuk PA, Elbarbary A, Zhu M, Ashjian P, Benhaim P, Hedrick MH, Fraser JK. Differen-tial expression of stem cell mobilization-associated molecules on multi-lineage cells from adipose tissue and bone marrow. Immunol Lett. 2003;89:267-70.
97. De Ugarte DA, Ashjian PH, Elbarbary A, Hedrick MH. Future of fat as raw material for tissue regeneration. Ann Plast Surg. 2003;50:215-9.
98. De Ugarte DA, Morizono K, Elbarbary A, Alfonso Z, Zuk PA, Zhu M, Dragoo JL, Ashjian P, Thomas B, Benhaim P, Chen I, Fraser J, Hedrick MH. Comparison of multi-lineage cells from human adipose tissue and bone mar-row. Cells Tissues Organs. 2003;174:101-9.
99. de Vries HJ, Middelkoop E, van Heemstra-Hoen M, Wildevuur CH, Westerhof W. Stromal cells from subcuta-neous adipose tissue seeded in a native collagen/elastin dermal substitute reduce wound contraction in full thickness skin defects. Lab Invest. 1995;73:532-40.
100. Demeulemeester D, Collen D, Lijnen HR. Effect of matrix metalloproteinase inhibition on adipose tissue de-velopment. Biochem Biophys Res Commun. 2005;329:105-10.
101. Di Rocco G, Iachininoto MG, Tritarelli A, Straino S, Zacheo A, Germani A, Crea F, Capogrossi MC. Myogenic potential of adipose-tissue-derived cells. J Cell Sci. 2006;119:2945-52.

99
102. Dragoo JL, Choi JY, Lieberman JR, Huang J, Zuk PA, Zhang J, Hedrick MH, Benhaim P. Bone induction by BMP-2 transduced stem cells derived from human fat. J Orthop Res. 2003;21:622-9.
103. Dragoo JL, Samimi B, Zhu M, Hame SL, Thomas BJ, Lieberman JR, Hedrick MH, Benhaim P. Tissue-engineered cartilage and bone using stem cells from human infrapatellar fat pads. J Bone Joint Surg Br. 2003;85:740-7.
104. Dudas JR, Marra KG, Cooper GM, Penascino VM, Mooney MP, Jiang S, Rubin JP, Losee JE. The osteogenic potential of adipose-derived stem cells for the repair of rabbit calvarial defects. Ann Plast Surg. 2006;56:543-8.
105. English A, Jones EA, Corscadden D, Henshaw K, Chapman T, Emery P, McGonagle D. A comparative as-sessment of cartilage and joint fat pad as a potential source of cells for autologous therapy development in knee osteoarthritis. Rheumatology (Oxford). 2007;46:1676-83.
106. Erices A, Conget P, Minguell JJ. Mesenchymal progenitor cells in human umbilical cord blood. Br J Haematol. 2000;109:235-42.
107. Erickson GR, Gimble JM, Franklin DM, Rice HE, Awad H, Guilak F. Chondrogenic potential of adipose tissue-derived stromal cells in vitro and in vivo. Biochem Biophys Res Commun. 2002;290:763-9.
108. Ersek R. Transplantation of purified autologous fat: a 3-year follow-up is disappointing. Plast Reconstr Surg. 1991;87:219-227.
109. Festy F, Hoareau L, Bes-Houtmann S, Pequin AM, Gonthier MP, Munstun A, Hoarau JJ, Cesari M, Roche R. Surface protein expression between human adipose tissue-derived stromal cells and mature adipocytes. His-tochem Cell Biol. 2005;124:113-21.
110. Feve B, Emorine LJ, Briend-Sutren MM, Lasnier F, Strosberg AD, Pairault J. Differential regulation of beta 1- and beta 2-adrenergic receptor protein and mRNA levels by glucocorticoids during 3T3-F442A adipose differ-entiation. J Biol Chem. 1990;265:16343-9.
111. Feve B, Emorine LJ, Lasnier F, Blin N, Baude B, Nahmias C, Strosberg AD, Pairault J. Atypical beta-adrenergic receptor in 3T3-F442A adipocytes. Pharmacological and molecular relationship with the human beta 3-adrenergic receptor. J Biol Chem. 1991;266:20329-36.
112. Fraser JK, Schreiber R, Strem B, Zhu M, Alfonso Z, Wulur I, Hedrick MH. Plasticity of human adipose stem cells toward endothelial cells and cardiomyocytes. Nat Clin Pract Cardiovasc Med. 2006;3 Suppl 1:S33-7.
113. Fraser JK, Wulur I, Alfonso Z, Hedrick MH. Fat tissue: an underappreciated source of stem cells for biotech-nology. Trends Biotechnol. 2006;24:150-4.
114. Frid MG, Kale VA, Stenmark KR. Mature vascular endothelium can give rise to smooth muscle cells via endo-thelial-mesenchymal transdifferentiation: in vitro analysis. Circ Res. 2002;90:1189-96.
115. Friedman G, Chajek-Shaul T, Stein O, Stein Y. Modulation of lipoprotein lipase activity in cultured rat mesen-chymal heart cells and preadipocytes by dibutyryl cyclic AMP, cholera toxin and 3-isobutyl-1-methylxanthine. Biochim Biophys Acta. 1983;762:106-17.
116. Fruhbeck G. The adipose tissue as a source of vasoactive factors. Curr Med Chem Cardiovasc Hematol Agents. 2004;2:197-208.
117. Fujimura J, Ogawa R, Mizuno H, Fukunaga Y, Suzuki H. Neural differentiation of adipose-derived stem cells isolated from GFP transgenic mice. Biochem Biophys Res Commun. 2005;333:116-21.
118. Gaben-Cogneville AM, Aron Y, Idriss G, Jahchan T, Pello JY, Swierczewski E. Differentiation under the control of insulin of rat preadipocytes in primary culture. Isolation of homogeneous cellular fractions by gradient centrifugation. Biochim Biophys Acta. 1983;762:437-444.
119. Gage FH. Mammalian neural stem cells. Science. 2000;287:1433-8. 120. Galis ZS, Khatri JJ. Matrix metalloproteinases in vascular remodeling and atherogenesis: the good, the bad,
and the ugly. Circ Res. 2002;90:251-62. 121. Garcia der Herreros A, Birnbaum M. The acquisition of increased insulin-responsive hexose transport in 3T3-
L1 adipocytes correlates with expression of a novel transporter gene. J Biol Chem. 1989;264:19994-19999. 122. Garcia-Olmo D, Garcia-Arranz M, Herreros D, Pascual I, Peiro C, Rodriguez-Montes JA. A phase I clinical trial
of the treatment of Crohn's fistula by adipose mesenchymal stem cell transplantation. Dis Colon Rectum. 2005;48:1416-23.
123. Gargett CE, Bucak K, Rogers PA. Isolation, characterization and long-term culture of human myometrial microvascular endothelial cells. Hum Reprod. 2000;15:293-301.
124. Gaustad KG, Boquest AC, Anderson BE, Gerdes AM, Collas P. Differentiation of human adipose tissue stem cells using extracts of rat cardiomyocytes. Biochem Biophys Res Commun. 2004;314:420-7.
125. Gehling UM, Ergun S, Schumacher U, Wagener C, Pantel K, Otte M, Schuch G, Schafhausen P, Mende T, Kilic N, Kluge K, Schafer B, Hossfeld DK, Fiedler W. In vitro differentiation of endothelial cells from AC133-positive progenitor cells. Blood. 2000;95:3106-12.
126. George J, Afek A, Abashidze A, Shmilovich H, Deutsch V, Kopolovich J, Miller H, Keren G. Transfer of endo-thelial progenitor and bone marrow cells influences atherosclerotic plaque size and composition in apolipo-protein E knockout mice. Arterioscler Thromb Vasc Biol. 2005;25:2636-41.
127. Glick J, Adelman S. Established cell lines from rat adipose tissue that secrete lipoprotein lipase. In-Vitro. 1983;19:421-428.
128. Goetze S, Bungenstock A, Czupalla C, Eilers F, Stawowy P, Kintscher U, Spencer-Hansch C, Graf K, Nurnberg B, Law RE, Fleck E, Grafe M. Leptin induces endothelial cell migration through Akt, which is inhibited by PPARgamma-ligands. Hypertension. 2002;40:748-54.
129. Goldberg IJ. Lipoprotein lipase and lipolysis: central roles in lipoprotein metabolism and atherogenesis. J Lipid Res. 1996;37:693-707.
130. Goldstein AL, Johnson PR. Primary culture of adipoblasts from obese and lean Zucker rat adipose tissue. Metabolism. 1982;31:601-7.
131. Grauss RW, Winter EM, van Tuyn J, Pijnappels DA, Steijn RV, Hogers B, van der Geest RJ, de Vries AA, Steendijk P, van der Laarse A, Gittenberger-de Groot AC, Schalij MJ, Atsma DE. Mesenchymal stem cells from ischemic heart disease patients improve left ventricular function after acute myocardial infarction. Am J Physiol Heart Circ Physiol. 2007;293:H2438-47.
132. Gregoire FM, Johnson PR, Greenwood MR. Comparison of the adipoconversion of preadipocytes derived from lean and obese Zucker rats in serum-free cultures. Int J Obes Relat Metab Disord. 1995;19:664-70.

100
133. Gronthos S, Franklin DM, Leddy HA, Robey PG, Storms RW, Gimble JM. Surface protein characterization of human adipose tissue-derived stromal cells. J Cell Physiol. 2001;189:54-63.
134. Gronthos S, Graves SE, Ohta S, Simmons PJ. The STRO-1+ fraction of adult human bone marrow contains the osteogenic precursors. Blood. 1994;84:4164-73.
135. Gronthos S, Zannettino AC, Hay SJ, Shi S, Graves SE, Kortesidis A, Simmons PJ. Molecular and cellular characterisation of highly purified stromal stem cells derived from human bone marrow. J Cell Sci. 2003;116:1827-35.
136. Gstraunthaler G. Alternatives to the use of fetal bovine serum: serum-free cell culture. Altex. 2003;20:275-81. 137. Guest SJ, Hadcock JR, Watkins DC, Malbon CC. Beta 1- and beta 2-adrenergic receptor expression in differ-
entiating 3T3-L1 cells. Independent regulation at the level of mRNA. J Biol Chem. 1990;265:5370-5. 138. Guilak F, Lott KE, Awad HA, Cao Q, Hicok KC, Fermor B, Gimble JM. Clonal analysis of the differentiation
potential of human adipose-derived adult stem cells. J Cell Physiol. 2006;206:229-37. 139. Guo X, Liao K. Analysis of gene expression profile during 3T3-L1 preadipocyte differentiation. Gene.
2000;251:45-53. 140. Halvorsen YC, Pingerelli P, Petre MC, Saladin R, Briggs MR. Human primary preadipocytes and differentiated
adipocytes. Strategies. 1998;11:58-60. 141. Halvorsen YD, Franklin D, Bond AL, Hitt DC, Auchter C, Boskey AL, Paschalis EP, Wilkison WO, Gimble JM.
Extracellular matrix mineralization and osteoblast gene expression by human adipose tissue-derived stromal cells. Tissue Eng. 2001;7:729-41.
142. Hanada K, Dennis JE, Caplan AI. Stimulatory effects of basic fibroblast growth factor and bone morphoge-netic protein-2 on osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. J Bone Miner Res. 1997;12:1606-1614.
143. Hanahan D. Signaling vascular morphogenesis and maintenance. Science. 1997;277:48-50. 144. Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 1996;86:353-64. 145. Hattori H, Sato M, Masuoka K, Ishihara M, Kikuchi T, Matsui T, Takase B, Ishizuka T, Kikuchi M, Fujikawa K.
Osteogenic potential of human adipose tissue-derived stromal cells as an alternative stem cell source. Cells Tissues Organs. 2004;178:2-12.
146. Hauner H, Entenmann G. Regional variation of adipose differentiation in cultured stromal-vascular cells from the abdominal and femoral adipose tissue of obese women. Int J Obes. 1991;15:121-126.
147. Hauner H, Entenmann G, Wabitsch M, Gaillard D, Ailhaud G, Negrel R, Pfeiffer EF. Promoting effect of gluco-corticoids on the differentiation of human adipocyte precursor cells cultured in a chemically defined medium. J Clin Invest. 1989;84:1663-1670.
148. Hauner H, Loffler G. Adipogenic factors in human serum promote the adipose conversion of 3T3-L1 fibro-blasts. Int J Obes. 1986;10:323-30.
149. Hauner H, Röhrig K, Spelleken M, Liu LS, Eckel J. Development of insulin-responsive glucose uptake and GLUT4 expression in differentiating human adipocyte precursor cells. Int J Obes Relat Metab Disord. 1998;22:448-453.
150. Hauner H, Wabitsch M, Pfeiffer EF. Differentiation of adipocyte precursor cells from obese and nonobese adult women and from different adipose tissue sites. Horm Metab Res Suppl. 1988;19:35-9.
151. Hausman GJ. Identification of adipose tissue primordia in perirenal tissues of pig fetuses: utility of phos-phatase histochemistry. Acta Anat (Basel). 1987;128:236-42.
152. Hausman GJ, Richardson LR. Histochemical and ultrastructural analysis of developing adipocytes in the fetal pig. Acta Anat Basel. 1982;114:228-247.
153. Hausman GJ, Thomas GB. The development of the inner layer of backfat in fetal and young pigs. J Anim Sci. 1984;58:1550-60.
154. Hausman GJ, Thomas GB. Differentiation of blood vessels in the adipose tissue of lean and obese fetal pigs, studied by differential enzyme histochemistry. Acta Anat (Basel). 1985;123:137-44.
155. Hausman GJ, Wright JT, Richardson RL. The influence of extracellular matrix substrata on preadipocyte development in serum-free cultures of stromal-vascular cells. J Anim Sci. 1996;74:2117-2128.
156. Hausman GJ, Wright JT, Thomas GB. Vascular and cellular development in fetal adipose tissue: lectin bind-ing studies and immunhistochemistry for laminin and type IV collagen. Microvasc Res. 1991;41:111-125.
157. Hemmrich K, Meersch M, Wiesemann U, Salber J, Klee D, Gries T, Pallua N. Polyesteramide-derived nonwov-ens as innovative degradable matrices support preadipocyte adhesion, proliferation and differentiation. Tis-sue Eng. 2006;12(12):3557-65.
158. Hemmrich K, Thomas GPL, Wosnitza M, von Heimburg D, Morrison WA, Pallua N. The isolation and expan-sion of primary preadipocytes. Int J Adipose Tissue. 2007;1(1):24-33.
159. Hemmrich K, von Heimburg D. Biomaterials for adipose tissue engineering. Exp Rev Med Dev. 2006;in press. 160. Hemmrich K, von Heimburg D, Cierpka K, Haydarlioglu S, Pallua N. Optimization of the differentiation of hu-
man preadipocytes in vitro. Differentiation. 2005;73:28-35. 161. Hemmrich K, von Heimburg D, Haydarlioglu S, Pallua N. Die Kultivierung humaner Präadipozyten im autolo-
gen Serum bietet neue Perspektiven für die Rekonstruktion von Weichgewebsdefekten. Plastische Chirurgie. 2003;3:Suppl. 1.
162. Hemmrich K, von Heimburg D, Rendchen R, Di Bartolo C, Milella E, Pallua N. Implantation of preadipocyte-loaded hyaluronic acid-based scaffolds into nude mice to evaluate potential for soft tissue engineering. Bio-materials. 2005.
163. Hicok KC, Du Laney TV, Zhou YS, Halvorsen YD, Hitt DC, Cooper LF, Gimble JM. Human adipose-derived adult stem cells produce osteoid in vivo. Tissue Eng. 2004;10:371-80.
164. Hicok KC, Thomas T, Gori F, Rickard DJ, Spelsberg TC, Riggs BL. Development and characterization of con-ditionally immortalized osteoblast precursor cell lines from human bone marrow stroma. J Bone Miner Res. 1998;13:205-17.
165. Hirashima M, Kataoka H, Nishikawa S, Matsuyoshi N. Maturation of embryonic stem cells into endothelial cells in an in vitro model of vasculogenesis. Blood. 1999;93:1253-63.
166. Hollenberg AN, Susulic VS, Madura JP, Zhang B, Moller DE, Tontonoz P, Sarraf P, Spiegelman BM, Lowell BB. Functional antagonism between CCAAT/Enhancer binding protein-alpha and peroxisome proliferator-activated receptor-gamma on the leptin promoter. J Biol Chem. 1997;272:5283-90.

101
167. Hoover-Plow J, Ellis J, Yuen L. In vivo plasminogen deficiency reduces fat accumulation. Thromb Haemost. 2002;87:1011-9.
168. Horwitz EM, Gordon PL, Koo WK, Marx JC, Neel MD, McNall RY, Muul L, Hofmann T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc Natl Acad Sci U S A. 2002;99:8932-7.
169. Hotamisligil GS, Arner P, Caro JF, Atkinson RL, Spiegelman BM. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J Clin Invest. 1995;95:2409-15.
170. Hou M, Yang KM, Zhang H, Zhu WQ, Duan FJ, Wang H, Song YH, Wei YJ, Hu SS. Transplantation of mesen-chymal stem cells from human bone marrow improves damaged heart function in rats. Int J Cardiol. 2007;115:220-8.
171. Hristov M, Weber C. Endothelial progenitor cells: characterization, pathophysiology, and possible clinical relevance. J Cell Mol Med. 2004;8:498-508.
172. Hristov M, Weber C. Ambivalence of progenitor cells in vascular repair and plaque stability. Curr Opin Lipidol. 2008;19:491-7.
173. Hristov M, Zernecke A, Schober A, Weber C. Adult progenitor cells in vascular remodeling during atheroscle-rosis. Biol Chem. 2008;389:837-44.
174. Huang JI, Beanes SR, Zhu M, Lorenz HP, Hedrick MH, Benhaim P. Rat extramedullary adipose tissue as a source of osteochondrogenic progenitor cells. Plast Reconstr Surg. 2002;109:1033-41; discussion 1042-3.
175. Huang JI, Zuk PA, Jones NF, Zhu M, Lorenz HP, Hedrick MH, Benhaim P. Chondrogenic potential of multipo-tential cells from human adipose tissue. Plast Reconstr Surg. 2004;113:585-94.
176. Huss FR, Kratz G. Adipose tissue processed for lipoinjection shows increased cellular survival in vitro when tissue engineering principles are applied. Scand J Plast Reconstr Hand Surg. 2002;36:166-71.
177. Hutley LJ, Herington AC, Shurety W, Cheung C, Vesey DA, Cameron DP, Prins JB. Human adipose tissue endothelial cells promote preadipocyte proliferation. Am J Physiol Endocrinol Metab. 2001;281:E1037-44.
178. Igura K, Zhang X, Takahashi K, Mitsuru A, Yamaguchi S, Takashi TA. Isolation and characterization of mesen-chymal progenitor cells from chorionic villi of human placenta. Cytotherapy. 2004;6:543-53.
179. Im GI, Shin YW, Lee KB. Do adipose tissue-derived mesenchymal stem cells have the same osteogenic and chondrogenic potential as bone marrow-derived cells? Osteoarthritis Cartilage. 2005;13:845-53.
180. Ingber DE, Folkman J. How does extracellular matrix control capillary morphogenesis? Cell. 1989;58:803-5. 181. Ingram DA, Caplice NM, Yoder MC. Unresolved questions, changing definitions, and novel paradigms for
defining endothelial progenitor cells. Blood. 2005;106:1525-31. 182. Ishikawa Y, Akasaka Y, Ishii T, Yoda-Murakami M, Choi-Miura NH, Tomita M, Ito K, Zhang L, Akishima Y, Ishi-
hara M, Muramatsu M, Taniyama M. Changes in the distribution pattern of gelatin-binding protein of 28 kDa (adiponectin) in myocardial remodelling after ischaemic injury. Histopathology. 2003;42:43-52.
183. Iwata H, Sata M. Potential contribution of bone marrow-derived precursors to vascular repair and lesion for-mation: lessons from animal models of vascular diseases. Front Biosci. 2007;12:4157-67.
184. Iyama K, Ohzono K, Usuku G. Electron microscopical studies on the genesis of white adipocytes: differentia-tion of immature pericytes into adipocytes in transplanted preadipose tissue. Virchows Arch B Cell Pathol Incl Mol Pathol. 1979;31:143-155.
185. Janowska-Wieczorek A, Majka M, Ratajczak J, Ratajczak MZ. Autocrine/paracrine mechanisms in human hematopoiesis. Stem Cells. 2001;19:99-107.
186. Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du J, Aldrich S, Lisberg A, Low WC, Largaespada DA, Verfaillie CM. Pluripotency of mesen-chymal stem cells derived from adult marrow. Nature. 2002;418:41-9.
187. Jin X, Fukuda N, Su J, Takagi H, Lai Y, Lin Z, Kanmatsuse K, Wang ZW, Unger RH. Effects of leptin on endo-thelial function with OB-Rb gene transfer in Zucker fatty rats. Atherosclerosis. 2003;169:225-33.
188. Jun ES, Lee TH, Cho HH, Suh SY, Jung JS. Expression of telomerase extends longevity and enhances differ-entiation in human adipose tissue-derived stromal cells. Cell Physiol Biochem. 2004;14:261-8.
189. Kale S, Karihaloo A, Clark PR, Kashgarian M, Krause DS, Cantley LG. Bone marrow stem cells contribute to repair of the ischemically injured renal tubule. J Clin Invest. 2003;112:42-9.
190. Kamihata H, Matsubara H, Nishiue T, Fujiyama S, Tsutsumi Y, Ozono R, Masaki H, Mori Y, Iba O, Tateishi E, Kosaki A, Shintani S, Murohara T, Imaizumi T, Iwasaka T. Implantation of bone marrow mononuclear cells into ischemic myocardium enhances collateral perfusion and regional function via side supply of angioblasts, an-giogenic ligands, and cytokines. Circulation. 2001;104:1046-52.
191. Kan I, Ben-Zur T, Barhum Y, Levy YS, Burstein A, Charlow T, Bulvik S, Melamed E, Offen D. Dopaminergic differentiation of human mesenchymal stem cells--utilization of bioassay for tyrosine hydroxylase expres-sion. Neurosci Lett. 2007;419:28-33.
192. Kang SK, Jun ES, Bae YC, Jung JS. Interactions between human adipose stromal cells and mouse neural stem cells in vitro. Brain Res Dev Brain Res. 2003;145:141-9.
193. Kang SK, Lee DH, Bae YC, Kim HK, Baik SY, Jung JS. Improvement of neurological deficits by intracerebral transplantation of human adipose tissue-derived stromal cells after cerebral ischemia in rats. Exp Neurol. 2003;183:355-66.
194. Kang SK, Putnam L, Dufour J, Ylostalo J, Jung JS, Bunnell BA. Expression of telomerase extends the life-span and enhances osteogenic differentiation of adipose tissue-derived stromal cells. Stem Cells. 2004;22:1356-72.
195. Kang SK, Putnam LA, Ylostalo J, Popescu IR, Dufour J, Belousov A, Bunnell BA. Neurogenesis of Rhesus adipose stromal cells. J Cell Sci. 2004;117:4289-99.
196. Kang SK, Shin MJ, Jung JS, Kim YG, Kim CH. Autologous adipose tissue-derived stromal cells for treatment of spinal cord injury. Stem Cells Dev. 2006;15:583-94.
197. Kaplan FS, Hahn GV, Zasloff MA. Heterotopic Ossification: Two Rare Forms and What They Can Teach Us. J Am Acad Orthop Surg. 1994;2:288-296.
198. Karagiannides I, Tchkonia T, Dobson DE, Steppan CM, Cummins P, Chan G, Salvatori K, Hadzopoulou-Cladaras M, Kirkland JL. Altered expression of C/EBP family members results in decreased adipogenesis with aging. Am J Physiol Regul Integr Comp Physiol. 2001;280:R1772-80.
199. Kato K, Suzuki F, Ogasawara N. Induction of S100 protein in 3T3-L1 cells during differentiation to adipocytes and its liberating by lipolytic hormones. Eur J Biochem. 1988;177:461-6.

102
200. Katoh J, Ishida Y, Taniguchi H. Isolation of preadipocytes from rat adipose tissue in vitro. Folia Histochem Cytobiol. 1994;32:235-238.
201. Katz AJ, Llull R, Hedrick MH, Futrell JW. Emerging approaches to the tissue engineering of fat. Clin Plast Surg. 1999;26:587-603.
202. Kawada T, Aoki N, Kawai R, Sugimoto E. Proliferation of 3T3-L1 preadipocytes in a completely defined serum-free medium. Cell Biol Int Rep. 1990;14:567-74.
203. Kawaguchi N, Toriyama K, Nicodemou-Lena E, Inou K, Torii S, Kitagawa Y. De novo adipogenesis in mice at the site of injection of basement membrane and basic fibroblast growth factor. Proc Natl Acad Sci USA. 1998;95:1062-1066.
204. Kawaguchi N, Xu X, Tajima R, Kronqvist P, Sundberg C, Loechel F, Albrechtsen R, Wewer UM. ADAM 12 protease induces adipogenesis in transgenic mice. Am J Pathol. 2002;160:1895-903.
205. Kawamoto A, Gwon HC, Iwaguro H, Yamaguchi JI, Uchida S, Masuda H, Silver M, Ma H, Kearney M, Isner JM, Asahara T. Therapeutic potential of ex vivo expanded endothelial progenitor cells for myocardial ischemia. Circulation. 2001;103:634-7.
206. Kawanami D, Maemura K, Takeda N, Harada T, Nojiri T, Imai Y, Manabe I, Utsunomiya K, Nagai R. Direct recip-rocal effects of resistin and adiponectin on vascular endothelial cells: a new insight into adipocytokine-endothelial cell interactions. Biochem Biophys Res Commun. 2004;314:415-9.
207. Kern PA, Ranganathan S, Li C, Wood L, Ranganathan G. Adipose tissue tumor necrosis factor and inter-leukin-6 expression in human obesity and insulin resistance. Am J Physiol Endocrinol Metab. 2001;280:E745-51.
208. Kern PA, Saghizadeh M, Ong JM, Bosch RJ, Deem R, Simsolo RB. The expression of tumor necrosis factor in human adipose tissue. Regulation by obesity, weight loss, and relationship to lipoprotein lipase. J Clin In-vest. 1995;95:2111-9.
209. Kern S, Eichler H, Stoeve J, Kluter H, Bieback K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells. 2006;24:1294-301.
210. Kim M, Choi YS, Yang SH, Hong HN, Cho SW, Cha SM, Pak JH, Kim CW, Kwon SW, Park CJ. Muscle regenera-tion by adipose tissue-derived adult stem cells attached to injectable PLGA spheres. Biochem Biophys Res Commun. 2006;348:386-92.
211. Kirkland JL, Hollenberg CH, Gillon WS. Age, anatomic site, and the replication and differentiation of adipo-cyte precursors. Am J Physiol. 1990;258:206-210.
212. Kleinman HK, McGarvey ML, Liotta LA, Robey PG, Tryggvason K, Martin GR. Isolation and characterization of type IV procollagen, laminin, and heparan sulfate proteoglycan from the EHS sarcoma. Biochemistry. 1982;21:6188-93.
213. Knippenberg M, Helder MN, Doulabi BZ, Semeins CM, Wuisman PI, Klein-Nulend J. Adipose tissue-derived mesenchymal stem cells acquire bone cell-like responsiveness to fluid shear stress on osteogenic stimula-tion. Tissue Eng. 2005;11:1780-8.
214. Koc ON, Gerson SL, Cooper BW, Dyhouse SM, Haynesworth SE, Caplan AI, Lazarus HM. Rapid hematopoietic recovery after coinfusion of autologous-blood stem cells and culture-expanded marrow mesenchymal stem cells in advanced breast cancer patients receiving high-dose chemotherapy. J Clin Oncol. 2000;18:307-16.
215. Kocher AA, Schuster MD, Szabolcs MJ, Takuma S, Burkhoff D, Wang J, Homma S, Edwards NM, Itescu S. Neovascularization of ischemic myocardium by human bone-marrow-derived angioblasts prevents cardio-myocyte apoptosis, reduces remodeling and improves cardiac function. Nat Med. 2001;7:430-6.
216. Koekemoer TC, Litthauer D, Oelofsen W. Isolation and characterization of adipose tissue glycerol-3-phosphate dehydrogenase. Int J Biochem Cell Biol. 1995;27:625-32.
217. Kojima S, Funahashi T, Sakamoto T, Miyamoto S, Soejima H, Hokamaki J, Kajiwara I, Sugiyama S, Yoshimura M, Fujimoto K, Miyao Y, Suefuji H, Kitagawa A, Ouchi N, Kihara S, Matsuzawa Y, Ogawa H. The variation of plasma concentrations of a novel, adipocyte derived protein, adiponectin, in patients with acute myocardial infarction. Heart. 2003;89:667.
218. Kopen GC, Prockop DJ, Phinney DG. Marrow stromal cells migrate throughout forebrain and cerebellum, and they differentiate into astrocytes after injection into neonatal mouse brains. Proc Natl Acad Sci U S A. 1999;96:10711-6.
219. Korbling M, Estrov Z. Adult stem cells for tissue repair - a new therapeutic concept? N Engl J Med. 2003;349:570-82.
220. Kougias P, Chai H, Lin PH, Yao Q, Lumsden AB, Chen C. Effects of adipocyte-derived cytokines on endothe-lial functions: implication of vascular disease. J Surg Res. 2005;126:121-9.
221. Kubo Y, Kaidzu S, Nakajima I, Takenouchi K, Nakamura F. Organization of extracellular matrix components during differentiation of adipocytes in long-term culture. In Vitro Cell Dev Biol Anim. 2000;36:38-44.
222. Lai E, Rosen OM, Rubin CS. Dexamethasone regulates the beta-adrenergic receptor subtype expressed by 3T3 L1 preadipocytes and adipocytes. J Biol Chem. 1982;257:6691-6.
223. Lane MD, Lin FT, MacDougald OA, Vasseur-Cognet M. Control of adipocyte differentiation by CCAAT/enhancer binding protein alpha (C/EBP alpha). Int J Obes Relat Metab Disord. 1996;20 Suppl 3:S91-6.
224. Lane-Md, Reed-Bc, Clements-Pr. Insulin receptor synthesis and turnover in differentiating 3T3-L1 preadipo-cytes. Prog-Clin-Biol-Res. 1981;Pt:A 523-42.
225. Langer R. Tissue engineering: a new field and its challenges. Pharm Res. 1997;14:840-1. 226. Lau DC, Schillabeer G, Li ZH, Wong KL, Varzaneh FE, Tough SC. Paracrine interactions in adipose tissue
development and growth. Int J Obes Relat Metab Disord. 1996;20 Suppl 3:S16-25. 227. Lau DC, Shillabeer G, Wong KL, Tough SC, Russell JC. Influence of paracrine factors on preadipocyte replica-
tion and differentiation. Int J Obes. 1990;14:193-201. 228. Layne MD, Endege WO, Jain MK, Yet SF, Hsieh CM, Chin MT, Perrella MA, Blanar MA, Haber E, Lee ME. Aortic
carboxypeptidase-like protein, a novel protein with discoidin and carboxypeptidase-like domains, is up-regulated during vascular smooth muscle cell differentiation. J Biol Chem. 1998;273:15654-60.
229. Layne MD, Yet SF, Maemura K, Hsieh CM, Bernfield M, Perrella MA, Lee ME. Impaired abdominal wall devel-opment and deficient wound healing in mice lacking aortic carboxypeptidase-like protein. Mol Cell Biol. 2001;21:5256-61.
230. Lecoeur L, Ouhayoun JP. In vitro induction of osteogenic differentiation from non-osteogenic mesenchymal cells. Biomaterials. 1997;18:989-93.

103
231. Lee JA, Parrett BM, Conejero JA, Laser J, Chen J, Kogon AJ, Nanda D, Grant RT, Breitbart AS. Biological alchemy: engineering bone and fat from fat-derived stem cells. Ann Plast Surg. 2003;50:610-7.
232. Lee JH, Kemp DM. Human adipose-derived stem cells display myogenic potential and perturbed function in hypoxic conditions. Biochem Biophys Res Commun. 2006;341:882-8.
233. Lee RH, Kim B, Choi I, Kim H, Choi HS, Suh K, Bae YC, Jung JS. Characterization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell Physiol Biochem. 2004;14:311-24.
234. Lee RT, Yamamoto C, Feng Y, Potter-Perigo S, Briggs WH, Landschulz KT, Turi TG, Thompson JF, Libby P, Wight TN. Mechanical strain induces specific changes in the synthesis and organization of proteoglycans by vascular smooth muscle cells. J Biol Chem. 2001;276:13847-51.
235. Lendeckel S, Jodicke A, Christophis P, Heidinger K, Wolff J, Fraser JK, Hedrick MH, Berthold L, Howaldt HP. Autologous stem cells (adipose) and fibrin glue used to treat widespread traumatic calvarial defects: case re-port. J Craniomaxillofac Surg. 2004;32:370-3.
236. Levenberg S, Golub JS, Amit M, Itskovitz-Eldor J, Langer R. Endothelial cells derived from human embryonic stem cells. Proc Natl Acad Sci U S A. 2002;99:4391-6.
237. Li H, Cybulsky MI, Gimbrone MA, Jr., Libby P. An atherogenic diet rapidly induces VCAM-1, a cytokine-regulatable mononuclear leukocyte adhesion molecule, in rabbit aortic endothelium. Arterioscler Thromb. 1993;13:197-204.
238. Libby P, Simon DI. Inflammation and thrombosis: the clot thickens. Circulation. 2001;103:1718-20. 239. Liekens S, De Clercq E, Neyts J. Angiogenesis: regulators and clinical applications. Biochem Pharmacol.
2001;61:253-70. 240. Lijnen HR, Maquoi E, Hansen LB, Van Hoef B, Frederix L, Collen D. Matrix metalloproteinase inhibition im-
pairs adipose tissue development in mice. Arterioscler Thromb Vasc Biol. 2002;22:374-9. 241. Lilla J, Stickens D, Werb Z. Metalloproteases and adipogenesis: a weighty subject. Am J Pathol.
2002;160:1551-4. 242. Lin Y, Lee H, Berg AH, Lisanti MP, Shapiro L, Scherer PE. The lipopolysaccharide-activated toll-like receptor
(TLR)-4 induces synthesis of the closely related receptor TLR-2 in adipocytes. J Biol Chem. 2000;275:24255-63.
243. Lin Y, Luo E, Chen X, Liu L, Qiao J, Yan Z, Li Z, Tang W, Zheng X, Tian W. Molecular and cellular characteriza-tion during chondrogenic differentiation of adipose tissue-derived stromal cells in vitro and cartilage forma-tion in vivo. J Cell Mol Med. 2005;9:929-39.
244. Liu TM, Martina M, Hutmacher DW, Hui JH, Lee EH, Lim B. Identification of common pathways mediating differentiation of bone marrow- and adipose tissue-derived human mesenchymal stem cells into three mes-enchymal lineages. Stem Cells. 2007;25:750-60.
245. Long SD, Pekala PH. Regulation of GLUT4 gene expression by arachidonic acid. Evidence for multiple path-ways, one of which requires oxidation to prostaglandin E2. J Biol Chem. 1996;271:1138-44.
246. MacDougald OA, Hwang CS, Fan H, Lane MD. Regulated expression of the obese gene product (leptin) in white adipose tissue and 3T3-L1 adipocytes. Proc Natl Acad Sci U S A. 1995;92:9034-7.
247. MacDougald OA, Lane MD. Transcriptional regulation of gene expression during adipocyte differentiation. Annu Rev Biochem. 1995;64:345-73.
248. Malladi P, Xu Y, Chiou M, Giaccia AJ, Longaker MT. Effect of reduced oxygen tension on chondrogenesis and osteogenesis in adipose-derived mesenchymal cells. Am J Physiol Cell Physiol. 2006;290:C1139-46.
249. Mandriota SJ, Pepper MS. Regulation of angiopoietin-2 mRNA levels in bovine microvascular endothelial cells by cytokines and hypoxia. Circ Res. 1998;83:852-9.
250. Mannarino E, Pirro M. Endothelial injury and repair: a novel theory for atherosclerosis. Angiology. 2008;59:69S-72S.
251. Maquoi E, Munaut C, Colige A, Collen D, Lijnen HR. Modulation of adipose tissue expression of murine matrix metalloproteinases and their tissue inhibitors with obesity. Diabetes. 2002;51:1093-101.
252. Martinez-Estrada OM, Munoz-Santos Y, Julve J, Reina M, Vilaro S. Human adipose tissue as a source of Flk-1+ cells: new method of differentiation and expansion. Cardiovasc Res. 2005;65:328-33.
253. Masuda T, Furue M, Matsuda T. Novel strategy for soft tissue augmentation based on transplantation of frag-mented omentum and preadipocytes. Tissue Eng. 2004;10:1672-83.
254. McGuire PG, Seeds NW. The interaction of plasminogen activator with a reconstituted basement membrane matrix and extracellular macromolecules produced by cultured epithelial cells. J Cell Biochem. 1989;40:215-27.
255. Mick GJ, Wang X, McCormick K. White adipocyte vascular endothelial growth factor: regulation by insulin. Endocrinology. 2002;143:948-53.
256. Miller CW, Ntambi JM. Peroxisome proliferators induce mouse liver stearoyl-CoA desaturase 1 gene expres-sion. Proc Natl Acad Sci U S A. 1996;93:9443-8.
257. Miller ES, Esterly NB, Fairley JA. Progressive osseous heteroplasia. Arch Dermatol. 1996;132:787-91. 258. Minguell JJ, Conget P, Erices A. Biology and clinical utilization of mesenchymal progenitor cells. Braz J Med
Biol Res. 2000;33:881-7. 259. Minguell JJ, Erices A, Conget P. Mesenchymal stem cells. Exp Biol Med (Maywood). 2001;226:507-20. 260. Miranville A, Heeschen C, Sengenes C, Curat CA, Busse R, Bouloumie A. Improvement of postnatal neovas-
cularization by human adipose tissue-derived stem cells. Circulation. 2004;110:349-55. 261. Mitchell JB, McIntosh K, Zvonic S, Garrett S, Floyd ZE, Kloster A, Di Halvorsen Y, Storms RW, Goh B, Kilroy
G, Wu X, Gimble JM. Immunophenotype of human adipose-derived cells: temporal changes in stromal-associated and stem cell-associated markers. Stem Cells. 2006;24:376-85.
262. Miyahara Y, Nagaya N, Kataoka M, Yanagawa B, Tanaka K, Hao H, Ishino K, Ishida H, Shimizu T, Kangawa K, Sano S, Okano T, Kitamura S, Mori H. Monolayered mesenchymal stem cells repair scarred myocardium after myocardial infarction. Nat Med. 2006;12:459-65.
263. Mizuno H, Zuk PA, Zhu M, Lorenz HP, Benhaim P, Hedrick MH. Myogenic differentiation by human processed lipoaspirate cells. Plast Reconstr Surg. 2002;109:199-209; discussion 210-1.
264. Mohr JC, de Pablo JJ, Palecek SP. 3-D microwell culture of human embryonic stem cells. Biomaterials. 2006;27:6032-42.

104
265. Montague CT, Prins JB, Sanders L, Digby JE, O'Rahilly S. Depot- and sex-specific differences in human leptin mRNA expression: implications for the control of regional fat distribution. Diabetes. 1997;46:342-7.
266. Moon MH, Kim SY, Kim YJ, Kim SJ, Lee JB, Bae YC, Sung SM, Jung JS. Human adipose tissue-derived mes-enchymal stem cells improve postnatal neovascularization in a mouse model of hindlimb ischemia. Cell Physiol Biochem. 2006;17:279-90.
267. Morange PE, Bastelica D, Bonzi MF, Van Hoef B, Collen D, Juhan-Vague I, Lijnen HR. Influence of t-pA and u-PA on adipose tissue development in a murine model of diet-induced obesity. Thromb Haemost. 2002;87:306-10.
268. Morimura M, Ishiko O, Sumi T, Yoshida H, Ogita S. Angiogenesis in adipose tissues and skeletal muscles with rebound weight-gain after diet-restriction in rabbits. Int J Mol Med. 2001;8:499-503.
269. Morroni M, Barbatelli G, Zingaretti MC, Cinti S. Immunohistochemical, ultrastructural and morphometric evi-dence for brown adipose tissue recruitment due to cold acclimation in old rats. Int J Obes Relat Metab Disord. 1995;19:126-31.
270. Motoshima H, Wu X, Mahadev K, Goldstein BJ. Adiponectin suppresses proliferation and superoxide genera-tion and enhances eNOS activity in endothelial cells treated with oxidized LDL. Biochem Biophys Res Com-mun. 2004;315:264-71.
271. Moustaid N, Lasnier F, Hainque B, Quignard-Boulange A, Pairault J. Analysis of gene expression during adipogenesis in 3T3-F442A preadipocytes: insulin and dexamethasone control. J Cell Biochem. 1990;42:243-54.
272. Murphy-Mg, Negrel-R, Ailhaud-G. Lipoprotein lipase and monoacylglycerol lipase activities during maturation of ob17 preadipocytes. Biochim-Biophys-Acta. 1981;664:240-248.
273. Nagel T, Resnick N, Atkinson WJ, Dewey CF, Jr., Gimbrone MA, Jr. Shear stress selectively upregulates inter-cellular adhesion molecule-1 expression in cultured human vascular endothelial cells. J Clin Invest. 1994;94:885-91.
274. Nakagami H, Maeda K, Morishita R, Iguchi S, Nishikawa T, Takami Y, Kikuchi Y, Saito Y, Tamai K, Ogihara T, Kaneda Y. Novel autologous cell therapy in ischemic limb disease through growth factor secretion by cul-tured adipose tissue-derived stromal cells. Arterioscler Thromb Vasc Biol. 2005;25:2542-7.
275. Nakagami H, Morishita R, Maeda K, Kikuchi Y, Ogihara T, Kaneda Y. Adipose tissue-derived stromal cells as a novel option for regenerative cell therapy. J Atheroscler Thromb. 2006;13:77-81.
276. Nakajima I, Muroya S, Tanabe R, Chikuni K. Extracellular matrix development during differentiation into adi-pocytes with a unique increase in type V and VI collagen. Biol Cell. 2002;94:197-203.
277. Nakajima I, Yamaguchi T, Ozutsumi K, Aso H. Adipose tissue extracellular matrix: newly organized by adipo-cytes during differentiation. Differentiation. 1998;63:193-200.
278. Nakamura Y, Shimada K, Fukuda D, Shimada Y, Ehara S, Hirose M, Kataoka T, Kamimori K, Shimodozono S, Kobayashi Y, Yoshiyama M, Takeuchi K, Yoshikawa J. Implications of plasma concentrations of adiponectin in patients with coronary artery disease. Heart. 2004;90:528-33.
279. Nathan S, Das De S, Thambyah A, Fen C, Goh J, Lee EH. Cell-based therapy in the repair of osteochondral defects: a novel use for adipose tissue. Tissue Eng. 2003;9:733-44.
280. Neels JG, Thinnes T, Loskutoff DJ. Angiogenesis in an in vivo model of adipose tissue development. Faseb J. 2004;18:983-5.
281. Negrel R, Grimaldi P, Forest C, Ailhaud G. Establishment and characterization of fibroblast-like cell lines derived from adipocytes with the capacity to redifferentiate into adipocyte-like cells. Methods Enzymol. 1985;109:377-85.
282. Ng CW, Poznanski WJ, Borowiecki M, Reimer G. Differences in growth in vitro of adipose cells from normal and obese patients. Nature. 1971;231:445.
283. Nicosia RF, Bonanno E, Smith M, Yurchenco P. Modulation of angiogenesis in vitro by laminin-entactin com-plex. Dev Biol. 1994;164:197-206.
284. Nicosia RF, Ottinetti A. Modulation of microvascular growth and morphogenesis by reconstituted basement membrane gel in three-dimensional cultures of rat aorta: a comparative study of angiogenesis in matrigel, collagen, fibrin, and plasma clot. In Vitro Cell Dev Biol. 1990;26:119-28.
285. Noth U, Osyczka AM, Tuli R, Hickok NJ, Danielson KG, Tuan RS. Multilineage mesenchymal differentiation potential of human trabecular bone-derived cells. J Orthop Res. 2002;20:1060-9.
286. Ntambi JM, Young-Cheul K. Adipocyte differentiation and gene expression. J Nutr. 2000;130:3122S-3126S. 287. Nuttall ME, Patton AJ, Olivera DL, Nadeau DP, Gowen M. Human trabecular bone cells are able to express
both osteoblastic and adipocytic phenotype: implications for osteopenic disorders. J Bone Miner Res. 1998;13:371-82.
288. O'Connor KC, Song H, Rosenzweig N, Jansen DA. Extracellular matrix substrata alter adipocyte yield and lipogenesis in primary cultures of stromal-vascular cells from human adipose. Biotechnol Lett. 2003;25:1967-72.
289. Oda A, Taniguchi T, Yokoyama M. Leptin stimulates rat aortic smooth muscle cell proliferation and migration. Kobe J Med Sci. 2001;47:141-50.
290. Oedayrajsingh-Varma MJ, van Ham SM, Knippenberg M, Helder MN, Klein-Nulend J, Schouten TE, Ritt MJ, van Milligen FJ. Adipose tissue-derived mesenchymal stem cell yield and growth characteristics are affected by the tissue-harvesting procedure. Cytotherapy. 2006;8:166-77.
291. Ogawa R, Mizuno H, Watanabe A, Migita M, Shimada T, Hyakusoku H. Osteogenic and chondrogenic differen-tiation by adipose-derived stem cells harvested from GFP transgenic mice. Biochem Biophys Res Commun. 2004;313:871-7.
292. Oh H, Takagi H, Suzuma K, Otani A, Matsumura M, Honda Y. Hypoxia and vascular endothelial growth factor selectively up-regulate angiopoietin-2 in bovine microvascular endothelial cells. J Biol Chem. 1999;274:15732-9.
293. Okamoto Y, Arita Y, Nishida M, Muraguchi M, Ouchi N, Takahashi M, Igura T, Inui Y, Kihara S, Nakamura T, Yamashita S, Miyagawa J, Funahashi T, Matsuzawa Y. An adipocyte-derived plasma protein, adiponectin, ad-heres to injured vascular walls. Horm Metab Res. 2000;32:47-50.
294. Oswald J, Boxberger S, Jorgensen B, Feldmann S, Ehninger G, Bornhauser M, Werner C. Mesenchymal stem cells can be differentiated into endothelial cells in vitro. Stem Cells. 2004;22:377-84.

105
295. Ouchi N, Kihara S, Arita Y, Nishida M, Matsuyama A, Okamoto Y, Ishigami M, Kuriyama H, Kishida K, Nishi-zawa H, Hotta K, Muraguchi M, Ohmoto Y, Yamashita S, Funahashi T, Matsuzawa Y. Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation. 2001;103:1057-63.
296. Ouchi N, Kihara S, Arita Y, Okamoto Y, Maeda K, Kuriyama H, Hotta K, Nishida M, Takahashi M, Muraguchi M, Ohmoto Y, Nakamura T, Yamashita S, Funahashi T, Matsuzawa Y. Adiponectin, an adipocyte-derived plasma protein, inhibits endothelial NF-kappaB signaling through a cAMP-dependent pathway. Circulation. 2000;102:1296-301.
297. Owen M. Marrow stromal stem cells. J Cell Sci Suppl. 1988;10:63-76. 298. Pairault J, Green H. A study of the adipose conversion of suspended 3T3 cells using glycerophosphate dehy-
drogenase as differentiation marker. Proc Natl Acad Scie. 1979;76:5138-5142. 299. Papetti M, Herman IM. Mechanisms of normal and tumor-derived angiogenesis. Am J Physiol Cell Physiol.
2002;282:C947-70. 300. Paradisi G, Steinberg HO, Shepard MK, Hook G, Baron AD. Troglitazone therapy improves endothelial func-
tion to near normal levels in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2003;88:576-80.
301. Park C, Afrikanova I, Chung YS, Zhang WJ, Arentson E, Fong Gh G, Rosendahl A, Choi K. A hierarchical order of factors in the generation of FLK1- and SCL-expressing hematopoietic and endothelial progenitors from embryonic stem cells. Development. 2004;131:2749-62.
302. Park HY, Kwon HM, Lim HJ, Hong BK, Lee JY, Park BE, Jang Y, Cho SY, Kim HS. Potential role of leptin in angiogenesis: leptin induces endothelial cell proliferation and expression of matrix metalloproteinases in vivo and in vitro. Exp Mol Med. 2001;33:95-102.
303. Patrick CW. Breast tissue engineering. Annu Rev Biomed Eng. 2004;6:109-30. 304. Patrick CW, Jr. Adipose tissue engineering: the future of breast and soft tissue reconstruction following
tumor resection. Semin Surg Oncol. 2000;19:302-11. 305. Patrick CW, Jr. Tissue engineering strategies for adipose tissue repair. Anat Rec. 2001;263:361-6. 306. Paulauskis JD, Sul HS. Cloning and expression of mouse fatty acid synthase and other specific mRNAs.
Developmental and hormonal regulation in 3T3-L1 cells. J Biol Chem. 1988;263:7049-54. 307. Peer L. The neclected free fat graft. Am J Surg. 1956;18:233-250. 308. Peister A, Mellad JA, Larson BL, Hall BM, Gibson LF, Prockop DJ. Adult stem cells from bone marrow (MSCs)
isolated from different strains of inbred mice vary in surface epitopes, rates of proliferation, and differentia-tion potential. Blood. 2004;103:1662-8.
309. Pelton PD, Zhou L, Demarest KT, Burris TP. PPARgamma activation induces the expression of the adipocyte fatty acid binding protein gene in human monocytes. Biochem Biophys Res Commun. 1999;261:456-8.
310. Pepper MS. Extracellular proteolysis and angiogenesis. Thromb Haemost. 2001;86:346-55. 311. Pepper MS. Role of the matrix metalloproteinase and plasminogen activator-plasmin systems in angiogene-
sis. Arterioscler Thromb Vasc Biol. 2001;21:1104-17. 312. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S,
Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143-7. 313. Planat-Benard V, Menard C, Andre M, Puceat M, Perez A, Garcia-Verdugo JM, Penicaud L, Casteilla L. Spon-
taneous cardiomyocyte differentiation from adipose tissue stroma cells. Circ Res. 2004;94:223-9. 314. Planat-Benard V, Silvestre JS, Cousin B, Andre M, Nibbelink M, Tamarat R, Clergue M, Manneville C, Saillan-
Barreau C, Duriez M, Tedgui A, Levy B, Penicaud L, Casteilla L. Plasticity of human adipose lineage cells to-ward endothelial cells: physiological and therapeutic perspectives. Circulation. 2004;109:656-63.
315. Poissonet CM, Burdi AR, Bookstein FL. Growth and development of human adipose tissue during early gesta-tion. Early Hum Dev. 1983;8:1-11.
316. Poissonnet CM, Burdi AR, Garn SM. The chronology of adipose tissue appearance and distribution in the human fetus. Early Hum Dev. 1984;10:1-11.
317. Poznanski WJ, Waheed I, Van R. Human fat cell precursors: Morphologic and metabolic differentiation in culture. Lab Invest. 1973;29:570-576.
318. Prelle K, Zink N, Wolf E. Pluripotent stem cells--model of embryonic development, tool for gene targeting, and basis of cell therapy. Anat Histol Embryol. 2002;31:169-86.
319. Rahmouni K, Haynes WG. Endothelial effects of leptin: implications in health and diseases. Curr Diab Rep. 2005;5:260-6.
320. Rangappa S, Fen C, Lee EH, Bongso A, Sim EK. Transformation of adult mesenchymal stem cells isolated from the fatty tissue into cardiomyocytes. Ann Thorac Surg. 2003;75:775-9.
321. Rasulov MF, Vasilchenkov AV, Onishchenko NA, Krasheninnikov ME, Kravchenko VI, Gorshenin TL, Pidtsan RE, Potapov IV. First experience of the use bone marrow mesenchymal stem cells for the treatment of a pa-tient with deep skin burns. Bull Exp Biol Med. 2005;139:141-4.
322. Ratcliffe PJ, O'Rourke JF, Maxwell PH, Pugh CW. Oxygen sensing, hypoxia-inducible factor-1 and the regula-tion of mammalian gene expression. J Exp Biol. 1998;201:1153-62.
323. Reaven G. Syndrome X: 10 years after. Drugs. 1999;58 Suppl 1:19-20; discussion 75-82. 324. Reed-Bc, Lane-Md. Expression of insulin receptors during preadipocyte differentiation. Adv Enzyme Regul.
1980;18:97-117. 325. Rehman J, Traktuev D, Li J, Merfeld-Clauss S, Temm-Grove CJ, Bovenkerk JE, Pell CL, Johnstone BH, Con-
sidine RV, March KL. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation. 2004;109:1292-8.
326. Reyes M, Lund T, Lenvik T, Aguiar D, Koodie L, Verfaillie CM. Purification and ex vivo expansion of postnatal human marrow mesodermal progenitor cells. Blood. 2001;98:2615-25.
327. Richter W. Cell-based cartilage repair: illusion or solution for osteoarthritis. Curr Opin Rheumatol. 2007;19:451-6.
328. Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671-4. 329. Risau W, Flamme I. Vasculogenesis. Annu Rev Cell Dev Biol. 1995;11:73-91. 330. Rodbell M. Metabolism of isolated fat cells. J Biol Chem. 1964;239:375-380.

106
331. Rodriguez AM, Elabd C, Delteil F, Astier J, Vernochet C, Saint-Marc P, Guesnet J, Guezennec A, Amri EZ, Dani C, Ailhaud G. Adipocyte differentiation of multipotent cells established from human adipose tissue. Biochem Biophys Res Commun. 2004;315:255-63.
332. Rodriguez AM, Pisani D, Dechesne CA, Turc-Carel C, Kurzenne JY, Wdziekonski B, Villageois A, Bagnis C, Breittmayer JP, Groux H, Ailhaud G, Dani C. Transplantation of a multipotent cell population from human adi-pose tissue induces dystrophin expression in the immunocompetent mdx mouse. J Exp Med. 2005;201:1397-405.
333. Romagnani P, Annunziato F, Liotta F, Lazzeri E, Mazzinghi B, Frosali F, Cosmi L, Maggi L, Lasagni L, Schef-fold A, Kruger M, Dimmeler S, Marra F, Gensini G, Maggi E, Romagnani S. CD14+CD34low cells with stem cell phenotypic and functional features are the major source of circulating endothelial progenitors. Circ Res. 2005;97:314-22.
334. Ron D, Brasier AR, McGehee RE, Jr., Habener JF. Tumor necrosis factor-induced reversal of adipocytic phe-notype of 3T3-L1 cells is preceded by a loss of nuclear CCAAT/enhancer binding protein (C/EBP). J Clin In-vest. 1992;89:223-33.
335. Rosen ED, Spiegelman BM. Molecular regulation of adipogenesis. Annu Rev Cell Dev Biol. 2000;16:145-71. 336. Ross R. Atherosclerosis--an inflammatory disease. N Engl J Med. 1999;340:115-26. 337. Rubin CS, Hirsch A, Fung C, Rosen OM. Development of hormone receptors and hormonal responsiveness in
vitro. Insulin receptors and insulin sensitivity in the preadipocyte and adipocyte forms of 3T3-L1 cells. J Biol Chem. 1978;253:7570-7578.
338. Rupnick MA, Panigrahy D, Zhang CY, Dallabrida SM, Lowell BB, Langer R, Folkman MJ. Adipose tissue mass can be regulated through the vasculature. Proc Natl Acad Sci U S A. 2002;99:10730-5.
339. Safford KM, Hicok KC, Safford SD, Halvorsen YD, Wilkison WO, Gimble JM, Rice HE. Neurogenic differentia-tion of murine and human adipose-derived stromal cells. Biochem Biophys Res Commun. 2002;294:371-9.
340. Safford KM, Rice HE. Stem cell therapy for neurologic disorders: therapeutic potential of adipose-derived stem cells. Curr Drug Targets. 2005;6:57-62.
341. Safford KM, Safford SD, Gimble JM, Shetty AK, Rice HE. Characterization of neuronal/glial differentiation of murine adipose-derived adult stromal cells. Exp Neurol. 2004;187:319-28.
342. Saillan-Barreau C, Cousin B, Andre M, Villena P, Casteilla L, Penicaud L. Human adipose cells as candidates in defense and tissue remodeling phenomena. Biochem Biophys Res Commun. 2003;309:502-5.
343. Saito S, Liu B, Yokoyama K. Animal embryonic stem (ES) cells: self-renewal, pluripotency, transgenesis and nuclear transfer. Hum Cell. 2004;17:107-15.
344. Sakaguchi Y, Sekiya I, Yagishita K, Muneta T. Comparison of human stem cells derived from various mesen-chymal tissues: superiority of synovium as a cell source. Arthritis Rheum. 2005;52:2521-9.
345. Samad F, Yamamoto K, Loskutoff DJ. Distribution and regulation of plasminogen activator inhibitor-1 in murine adipose tissue in vivo. Induction by tumor necrosis factor-alpha and lipopolysaccharide. J Clin Invest. 1996;97:37-46.
346. Sarugaser R, Lickorish D, Baksh D, Hosseini MM, Davies JE. Human umbilical cord perivascular (HUCPV) cells: a source of mesenchymal progenitors. Stem Cells. 2005;23:220-9.
347. Sata M, Saiura A, Kunisato A, Tojo A, Okada S, Tokuhisa T, Hirai H, Makuuchi M, Hirata Y, Nagai R. Hemato-poietic stem cells differentiate into vascular cells that participate in the pathogenesis of atherosclerosis. Nat Med. 2002;8:403-9.
348. Schuster MD, Kocher AA, Seki T, Martens TP, Xiang G, Homma S, Itescu S. Myocardial neovascularization by bone marrow angioblasts results in cardiomyocyte regeneration. Am J Physiol Heart Circ Physiol. 2004;287:H525-32.
349. Sen A, Lea-Currie YR, Sujkowska D, Franklin DM, Wilkison WO, Halvorsen YD, Gimble JM. Adipogenic poten-tial of human adipose derived stromal cells from multiple donors is heterogeneous. J Cell Biochem. 2001;81:312-9.
350. Sengenes C, Lolmede K, Zakaroff-Girard A, Busse R, Bouloumie A. Preadipocytes in the human subcutane-ous adipose tissue display distinct features from the adult mesenchymal and hematopoietic stem cells. J Cell Physiol. 2005;205:114-22.
351. Seo MJ, Suh SY, Bae YC, Jung JS. Differentiation of human adipose stromal cells into hepatic lineage in vitro and in vivo. Biochem Biophys Res Commun. 2005;328:258-64.
352. Shahparaki A, Grunder L, Sorisky A. Comparison of human abdominal subcutaneous versus omental preadi-pocyte differentiation in primary culture. Metabolism. 2002;51:1211-5.
353. Shi Q, Rafii S, Wu MH, Wijelath ES, Yu C, Ishida A, Fujita Y, Kothari S, Mohle R, Sauvage LR, Moore MA, Storb RF, Hammond WP. Evidence for circulating bone marrow-derived endothelial cells. Blood. 1998;92:362-7.
354. Shi YY, Nacamuli RP, Salim A, Longaker MT. The osteogenic potential of adipose-derived mesenchymal cells is maintained with aging. Plast Reconstr Surg. 2005;116:1686-96.
355. Shigematsu M, Watanabe H, Sugihara H. Proliferation and differentiation of unilocular fat cells in the bone marrow. Cell Struct Funct. 1999;24:89-100.
356. Shore EM, Ahn J, Jan de Beur S, Li M, Xu M, Gardner RJ, Zasloff MA, Whyte MP, Levine MA, Kaplan FS. Pa-ternally inherited inactivating mutations of the GNAS1 gene in progressive osseous heteroplasia. N Engl J Med. 2002;346:99-106.
357. Shumakov VI, Onishchenko NA, Rasulov MF, Krasheninnikov ME, Zaidenov VA. Mesenchymal bone marrow stem cells more effectively stimulate regeneration of deep burn wounds than embryonic fibroblasts. Bull Exp Biol Med. 2003;136:192-5.
358. Sierra-Honigmann MR, Nath AK, Murakami C, Garcia-Cardena G, Papapetropoulos A, Sessa WC, Madge LA, Schechner JS, Schwabb MB, Polverini PJ, Flores-Riveros JR. Biological action of leptin as an angiogenic fac-tor. Science. 1998;281:1683-6.
359. Silva J, Chambers I, Pollard S, Smith A. Nanog promotes transfer of pluripotency after cell fusion. Nature. 2006;441:997-1001.
360. Silverman KJ, Lund DP, Zetter BR, Lainey LL, Shahood JA, Freiman DG, Folkman J, Barger AC. Angiogenic activity of adipose tissue. Biochem Biophys Res Commun. 1988;153:347-52.
361. Skoog T, Dichtl W, Boquist S, Skoglund-Andersson C, Karpe F, Tang R, Bond MG, de Faire U, Nilsson J, Eriksson P, Hamsten A. Plasma tumour necrosis factor-alpha and early carotid atherosclerosis in healthy middle-aged men. Eur Heart J. 2002;23:376-83.

107
362. Smahel J. Problematik des Fettgewebes in der Plastischen Chirurgie. Handchir Mikrochir Plast Chir. 1984;16:111-114.
363. Smahel J, Meyer V, Schütz K. Vascular augmentation of free adipose tissue grafts. Eur J Plast Surg. 1990;13:163-168.
364. Song HJ, Stevens CF, Gage FH. Neural stem cells from adult hippocampus develop essential properties of functional CNS neurons. Nat Neurosci. 2002;5:438-45.
365. Soukas A, Socci ND, Saatkamp BD, Novelli S, Friedman JM. Distinct transcriptional profiles of adipogenesis in vivo and in vitro. J Biol Chem. 2001;276:34167-74.
366. Spiegelman BM. PPAR-gamma: Adipogenic regulator and thiazolidinedione receptor. Diabetes. 1998;47:507-514.
367. Spiegelman BM, Farmer SR. Decreases in tubulin and actin gene expression prior to morphological differen-tiation of 3T3 adipocytes. Cell. 1982;29:53-60.
368. Spiegelman BM, Frank M, Green H. Molecular cloning of mRNA from 3T3 adipocytes. Regulation of mRNA content for glycerophosphate dehydrogenase and other differentiation-dependent proteins during adipocyte development. J Biol Chem. 1983;258:10083-9.
369. Spiegelman BM, Ginty CA. Fibronectin modulation of cell shape and lipogenic gene expression in 3T3 adipo-cytes. Cell. 1983;35:657-666.
370. Stacker SA, Runting AS, Caesar C, Vitali A, Lackmann M, Chang J, Ward L, Wilks AF. The 3T3-L1 fibroblast to adipocyte conversion is accompanied by increased expression of angiopoietin-1, a ligand for tie2. Growth Factors. 2000;18:177-91.
371. Stary HC, Chandler AB, Dinsmore RE, Fuster V, Glagov S, Insull W, Jr., Rosenfeld ME, Schwartz CJ, Wagner WD, Wissler RW. A definition of advanced types of atherosclerotic lesions and a histological classification of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, Ameri-can Heart Association. Circulation. 1995;92:1355-74.
372. Strem BM, Hicok KC, Zhu M, Wulur I, Alfonso Z, Schreiber RE, Fraser JK, Hedrick MH. Multipotential differen-tiation of adipose tissue-derived stem cells. Keio J Med. 2005;54:132-41.
373. Strem BM, Zhu M, Alfonso Z, Daniels EJ, Schreiber R, Beygui R, MacLellan WR, Hedrick MH, Fraser JK. Ex-pression of cardiomyocytic markers on adipose tissue-derived cells in a murine model of acute myocardial injury. Cytotherapy. 2005;7:282-91.
374. Suganami E, Takagi H, Ohashi H, Suzuma K, Suzuma I, Oh H, Watanabe D, Ojima T, Suganami T, Fujio Y, Nakao K, Ogawa Y, Yoshimura N. Leptin stimulates ischemia-induced retinal neovascularization: possible role of vascular endothelial growth factor expressed in retinal endothelial cells. Diabetes. 2004;53:2443-8.
375. Sugaya K. Possible use of autologous stem cell therapies for Alzheimer's disease. Curr Alzheimer Res. 2005;2:367-76.
376. Sugaya K, Alvarez A, Marutle A, Kwak YD, Choumkina E. Stem cell strategies for Alzheimer's disease therapy. Panminerva Med. 2006;48:87-96.
377. Sugihara H, Yonemitsu N, Miyabara S, Toda S. Proliferation of unilocular fat cells in the primary culture. J Lidid Res. 1987;28:1038-1045.
378. Sugihara H, Yonemitsu N, Miyabara S, Yun K. Primary cultures of unilocular fat cells: characteristics of growth in vitro and changes in differentiation properties. Differentiation. 1986;31:42-49.
379. Sugihara H, Yonemitsu N, Toda S, Miyabara S, Funatsumaru S, Matsumoto T. Unilocular fat cells in three-dimensional collagen gel matrix culture. J Lipid Res. 1988;29:691-7.
380. Sumi M, Sata M, Toya N, Yanaga K, Ohki T, Nagai R. Transplantation of adipose stromal cells, but not mature adipocytes, augments ischemia-induced angiogenesis. Life Sci. 2007;80:559-65.
381. Talens-Visconti R, Bonora A, Jover R, Mirabet V, Carbonell F, Castell JV, Gomez-Lechon MJ. Hepatogenic differentiation of human mesenchymal stem cells from adipose tissue in comparison with bone marrow mes-enchymal stem cells. World J Gastroenterol. 2006;12:5834-45.
382. Talens-Visconti R, Bonora A, Jover R, Mirabet V, Carbonell F, Castell JV, Gomez-Lechon MJ. Human mesen-chymal stem cells from adipose tissue: Differentiation into hepatic lineage. Toxicol In Vitro. 2006.
383. Talens-Visconti R, Bonora-Centelles A, Castell JV, Gomez-Lechon MJ. [Alternative sources of hepatocytes for cell therapy]. Gastroenterol Hepatol. 2006;29:366-76.
384. Tchkonia T, Tchoukalova YD, Giorgadze N, Pirtskhalava T, Karagiannides I, Forse RA, Koo A, Stevenson M, Chinnappan D, Cartwright A, Jensen MD, Kirkland JL. Abundance of two human preadipocyte subtypes with distinct capacities for replication, adipogenesis, and apoptosis varies among fat depots. Am J Physiol Endo-crinol Metab. 2005;288:E267-77.
385. Temple S, Alvarez-Buylla A. Stem cells in the adult mammalian central nervous system. Curr Opin Neurobiol. 1999;9:135-41.
386. Tengku-Muhammad T, Hughes T, Cryer A, Ramji D. Differential regulation of lipoprotein lipase in the macro-phage J774.2 cell line by cytokines. Cytokine. 1996;8:525-533.
387. Tezel TH, Del Priore LV. Serum-free media for culturing and serial-passaging of adult human retinal pigment epithelium. Exp Eye Res. 1998;66:807-15.
388. Tholpady SS, Katz AJ, Ogle RC. Mesenchymal stem cells from rat visceral fat exhibit multipotential differen-tiation in vitro. Anat Rec. 2003;272A:398-402.
389. Tille JC, Wood J, Mandriota SJ, Schnell C, Ferrari S, Mestan J, Zhu Z, Witte L, Pepper MS. Vascular endothe-lial growth factor (VEGF) receptor-2 antagonists inhibit VEGF- and basic fibroblast growth factor-induced an-giogenesis in vivo and in vitro. J Pharmacol Exp Ther. 2001;299:1073-85.
390. Timchenko NA, Wilde M, Nakanishi M, Smith JR, Darlington GJ. CCAAT/enhancer-binding protein alpha (C/EBP alpha) inhibits cell proliferation through the p21 (WAF-1/CIP-1/SDI-1) protein. Genes Dev. 1996;10:804-15.
391. Tokunaga O, Satoh T, Yu S. Multinucleated variant endothelial cells (MVECs) have a greater capacity for LDL cholesterol uptake than typical mononuclear endothelial cells (TECs). J Atheroscler Thromb. 2002;9:35-41.
392. Tontonoz P, Hu E, Devine J, Beale EG, Spiegelman BM. PPAR gamma 2 regulates adipose expression of the phosphoenolpyruvate carboxykinase gene. Mol Cell Biol. 1995;15:351-7.
393. Tontonoz P, Hu E, Graves RA, Budavari AI, Spiegelman BM. mPPAR gamma 2: tissue-specific regulator of an adipocyte enhancer. Genes Dev. 1994;8:1224-34.

108
394. Trzaska KA, Kuzhikandathil EV, Rameshwar P. Specification of a dopaminergic phenotype from adult human mesenchymal stem cells. Stem Cells. 2007;25:2797-808.
395. Tuan RS, Boland G, Tuli R. Adult mesenchymal stem cells and cell-based tissue engineering. Arthritis Res Ther. 2003;5:32-45.
396. Umek RM, Friedman AD, McKnight SL. CCAAT-enhancer binding protein: a component of a differentiation switch. Science. 1991;251:288-92.
397. Urbich C, Dimmeler S. Endothelial progenitor cells functional characterization. Trends Cardiovasc Med. 2004;14:318-22.
398. Urbich C, Dimmeler S. Endothelial progenitor cells: characterization and role in vascular biology. Circ Res. 2004;95:343-53.
399. Van Harmelen V, Rohrig K, Hauner H. Comparison of proliferation and differentiation capacity of human adi-pocyte precursor cells from the omental and subcutaneous adipose tissue depot of obese subjects. Metabo-lism. 2004;53:632-7.
400. van Hinsbergh VW, Havekes L, Emeis JJ, van Corven E, Scheffer M. Low density lipoprotein metabolism by endothelial cells from human umbilical cord arteries and veins. Arteriosclerosis. 1983;3:547-59.
401. Vanden Bossche L, Vanderstraeten G. Heterotopic ossification: a review. J Rehabil Med. 2005;37:129-36. 402. Varzaneh FE, Shillabeer G, Wong KL, Lau DC. Extracellular matrix components secreted by microvascular
endothelial cells stimulate preadipocyte differentiation in vitro. Metabolism. 1994;43:906-912. 403. Vats A, Tolley NS, Polak JM, Buttery LD. Stem cells: sources and applications. Clin Otolaryngol Allied Sci.
2002;27:227-32. 404. Verma S, Li SH, Wang CH, Fedak PW, Li RK, Weisel RD, Mickle DA. Resistin promotes endothelial cell activa-
tion: further evidence of adipokine-endothelial interaction. Circulation. 2003;108:736-40. 405. von Heimburg D, Hemmrich K, Haydarlioglu S, Staiger H, Pallua N. Comparison of viable cell yield from ex-
cised versus aspirated adipose tissue. Cells Tissues Organs. 2004;178:87-92. 406. von Heimburg D, Hemmrich K, Zachariah S, Staiger H, Pallua N. Oxygen consumption in undifferentiated
versus differentiated adipogenic mesenchymal precursor cells. Respir Physiol Neurobiol. 2005;146:107-16. 407. von Heimburg D, Heschel I, Zachariah S, Kühling H, Hafemann B, Pallua N. Human preadipocytes seeded on
freeze-dried collagen scaffolds investigated in vitro and in vivo. Cells Tissues Organs. 2000;166:69. 408. von Heimburg D, Pallua N. Two-year histological outcome of facial lipofilling. Ann Plast Surg. 2001;46:644-46. 409. Wang CH, Ciliberti N, Li SH, Szmitko PE, Weisel RD, Fedak PW, Al-Omran M, Cherng WJ, Li RK, Stanford WL,
Verma S. Rosiglitazone facilitates angiogenic progenitor cell differentiation toward endothelial lineage: a new paradigm in glitazone pleiotropy. Circulation. 2004;109:1392-400.
410. Wang H, Scott RE. Inhibition of distinct steps in the adipocyte differentiation pathway in 3T3 T mesenchymal stem cells by dimethyl sulphoxide (DMSO). Cell Prolif. 1993;26:55-66.
411. Weiner FR, Smith PJ, Wertheimer S, Rubin CS. Regulation of gene expression by insulin and tumor necrosis factor alpha in 3T3-L1 cells. Modulation of the transcription of genes encoding acyl-CoA synthetase and stearoyl-CoA desaturase-1. J Biol Chem. 1991;266:23525-8.
412. Weissman IL. Translating stem and progenitor cell biology to the clinic: barriers and opportunities. Science. 2000;287:1442-6.
413. Weyer C, Funahashi T, Tanaka S, Hotta K, Matsuzawa Y, Pratley RE, Tataranni PA. Hypoadiponectinemia in obesity and type 2 diabetes: close association with insulin resistance and hyperinsulinemia. J Clin Endocri-nol Metab. 2001;86:1930-5.
414. Wickham MQ, Erickson GR, Gimble JM, Vail TP, Guilak F. Multipotent stromal cells derived from the infrapa-tellar fat pad of the knee. Clin Orthop Relat Res. 2003:196-212.
415. Wier M, Scott R. Regulation of the terminal event in cellular differentiation: biological mechanisms of the loss of proliferative potential. J Cell Biol. 1986;102:1955-1964.
416. Wiese C, Kania G, Rolletschek A, Blyszczuk P, Wobus AM. Pluripotency: capacity for in vitro differentiation of undifferentiated embryonic stem cells. Methods Mol Biol. 2006;325:181-205.
417. Winter A, Breit S, Parsch D, Benz K, Steck E, Hauner H, Weber RM, Ewerbeck V, Richter W. Cartilage-like gene expression in differentiated human stem cell spheroids: a comparison of bone marrow-derived and adipose tissue-derived stromal cells. Arthritis Rheum. 2003;48:418-29.
418. Woodbury D, Reynolds K, Black IB. Adult bone marrow stromal stem cells express germline, ectodermal, endodermal, and mesodermal genes prior to neurogenesis. J Neurosci Res. 2002;69:908-17.
419. Woodbury D, Schwarz EJ, Prockop DJ, Black IB. Adult rat and human bone marrow stromal cells differentiate into neurons. J Neurosci Res. 2000;61:364-70.
420. Wu Z, Bucher NL, Farmer SR. Induction of peroxisome proliferator-activated receptor gamma during the conversion of 3T3 fibroblasts into adipocytes is mediated by C/EBPbeta, C/EBPdelta, and glucocorticoids. Mol Cell Biol. 1996;16:4128-36.
421. Xu Y, Malladi P, Wagner DR, Longaker MT. Adipose-derived mesenchymal cells as a potential cell source for skeletal regeneration. Curr Opin Mol Ther. 2005;7:300-5.
422. Yancopoulos GD, Davis S, Gale NW, Rudge JS, Wiegand SJ, Holash J. Vascular-specific growth factors and blood vessel formation. Nature. 2000;407:242-8.
423. Yang LY, Liu XM, Sun B, Hui GZ, Fei J, Guo LH. Adipose tissue-derived stromal cells express neuronal phe-notypes. Chin Med J (Engl). 2004;117:425-9.
424. Yin AH, Miraglia S, Zanjani ED, Almeida-Porada G, Ogawa M, Leary AG, Olweus J, Kearney J, Buck DW. AC133, a novel marker for human hematopoietic stem and progenitor cells. Blood. 1997;90:5002-12.
425. Yokota T, Oritani K, Takahashi I, Ishikawa J, Matsuyama A, Ouchi N, Kihara S, Funahashi T, Tenner AJ, Tomi-yama Y, Matsuzawa Y. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood. 2000;96:1723-32.
426. Yoon YS, Park JS, Tkebuchava T, Luedeman C, Losordo DW. Unexpected severe calcification after transplan-tation of bone marrow cells in acute myocardial infarction. Circulation. 2004;109:3154-7.
427. Yoshimura K, Shigeura T, Matsumoto D, Sato T, Takaki Y, Aiba-Kojima E, Sato K, Inoue K, Nagase T, Koshima I, Gonda K. Characterization of freshly isolated and cultured cells derived from the fatty and fluid portions of liposuction aspirates. J Cell Physiol. 2006;208:64-76.

109
428. Young HE, Steele TA, Bray RA, Hudson J, Floyd JA, Hawkins K, Thomas K, Austin T, Edwards C, Cuzzourt J, Duenzl M, Lucas PA, Black AC, Jr. Human reserve pluripotent mesenchymal stem cells are present in the connective tissues of skeletal muscle and dermis derived from fetal, adult, and geriatric donors. Anat Rec. 2001;264:51-62.
429. Zangani D, Darcy KM, Masso-Welch PA, Bellamy ES, Desole MS, Ip MM. Multiple differentiation pathways of rat mammary stromal cells in vitro: acquisition of a fibroblast, adipocyte or endothelial phenotype is depend-ent on hormonal and extracellular matrix stimulation. Differentiation. 1999;64:91-101.
430. Zhang B, Berger J, Hu E, Szalkowski D, White-Carrington S, Spiegelman BM, Moller DE. Negative regulation of peroxisome proliferator-activated receptor-gamma gene expression contributes to the antiadipogenic ef-fects of tumor necrosis factor-alpha. Mol Endocrinol. 1996;10:1457-66.
431. Zhang F, Chen Y, Heiman M, Dimarchi R. Leptin: structure, function and biology. Vitam Horm. 2005;71:345-72. 432. Zhang QX, Magovern CJ, Mack CA, Budenbender KT, Ko W, Rosengart TK. Vascular endothelial growth factor
is the major angiogenic factor in omentum: mechanism of the omentum-mediated angiogenesis. J Surg Res. 1997;67:147-54.
433. Zuk PA, Zhu M, Ashjian P, De Ugarte DA, Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P, Hedrick MH. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002;13:4279-95.
434. Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P, Lorenz HP, Hedrick MH. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng. 2001;7:211-28.

110
PUBLICATIONS
FULL PAPERS
1. H. Ott, M. Wosnitza, H.F. Merk. Immunologic control parameters during specific immuno-
therapy, Hautarzt – 2008 Jul;59(7):551-6
2. M. Wosnitza, C. Blazek, M. Megahed. Pemphigus herpetiformis, Hautarzt – 2008
Jun;59(6):459-460
3. M. Wosnitza, K. Hemmrich, A. Groger, S. Gräber, N. Pallua. Plasticity of human adipose
stem cells to perform adipogenic and endothelial differentiation, Differentition - 2007
Jan;75(1):12-23
4. K. Hemmrich, G.P.L. Thomas, M. Wosnitza, D. von Heimburg, W. A. Morrison, N. Pallua.
The Isolation and Expansion of Primary Preadipocytes, International Journal of Adipose
Tissue – 2007;1(1):24-33
PUBLISHED ABSTRACTS
1. K. Hemmrich, M. Wosnitza, A. Gröger, N. Pallua. Aus Präadipozyten werden Endothel-
zellen, aus Endothelzellen werden Fettzellen – die Plastizität der Fettgewebsvorläuferzel-
len, Plastische Chirurgie 09/2006; 6, Suppl. 1; 46
2. K. Hemmrich, M. Wosnitza, A. Groger, S. Gräber, N. Pallua. Plasticity of human adipose
stem cells to perform adipogenic and endothelial differentiation, JPRAS 09/2006; 59; S1-
12
SCIENTIFIC PRESENTATIONS
1. M. Wosnitza, C. Blazek, M. Megahed, Aachener Dermatologen Abend, Aachen,
06/2008: Pemphigus herpetiformis
2. M. Wosnitza, K. Hemmrich, A. Gröger, N. Pallua, 14th International Congress of the In-
ternational Confederation for Plastic, Reconstructive and Aesthetic Surgery (IPRAS), Ber-
lin, 06/2007: Adipose tissue is a source of endothelial cells that have the potency to un-
dergo adipogenic differentiation – benefit or pitfall?
3. M. Wosnitza, K. Hemmrich, A. Gröger, N. Pallua, 124. Kongress der Deutschen Gesell-
schaft für Chirurgie, München, 05/2007: Humane Fettgewebsvorläuferzellen ermöglichen
die kombinierte Züchtung von Fettgewebe und endothelialen Strukturen

111
4. K. Hemmrich, M. Wosnitza, C. Gummersbach, D. von Heimburg, N. Pallua. Bernard
O’Brien Institute of Microsurgery Alumni Meeting, Melbourne, Australia, 03/2007: The use
of preadipocytes for adipose tissue engineering
5. K. Hemmrich, M. Wosnitza, A. Gröger, N. Pallua, 37. Jahrestagung der Vereinigung der
Deutschen Plastischen Chirurgen und 11. Jahrestagung der Vereinigung der Deutschen
Ästhetischen Chirurgen, Aachen, 09-10/2006: Aus Präadipozyten werden Endothelzellen,
aus Endothelzellen werden Fettzellen – die Plastizität der Fettgewebsvorläuferzellen
6. K. Hemmrich, M. Wosnitza, A. Gröger, S. Gräber, N. Pallua, European Conference of
Scientists and Plastic Surgeons (ECSAPS), London, England, 09/2006: Plasticity of hu-
man adipose stem cells to perform adipogenic and endothelial differentiation
7. M. Wosnitza, K. Hemmrich, A. Gröger, N. Pallua, Würzburg, 2nd International Conference
"Strategies in Tissue Engineering", Würzburg, 05-06/2006: Human adipose stromal vas-
cular fraction cells allow combined engineering of fat tissue and endothelial structures
8. M. Wosnitza, K. Hemmrich, A. Gröger, S. Gräber, N. Pallua, 41. Congress of the Euro-
pean Society of Surgical Research, Rostock, 05/2006: Potency of human adipose stro-
mal vascular fraction to perform adipogenic as well as endothelial differentiation

112
ACKNOWLEDGEMENT
During the time of my studies working on this thesis I had the fortune to be supported by
many wonderful people whom I owe thanks and gratitude.
Without the grateful support of Prof. Dr. Dr. Norbert Pallua my work on this thesis would not
have been possible at all. His good judgement and patience guided me during the work on
this thesis. Gratitude to him for being a wonderful teacher, for always showing much interest
in my work and for his grateful support. As this is not for granted I owe him special thanks for
all the encouragement and help.
Further I want to thank Christoph Suschek and Karsten Hemmrich for supplying special ad-
vice, supporting my ideas and for making it possible to perform all these experiments. They
were the major force behind this work.
I would also like to acknowledge the wonderful laboratory staff, Tanja Wollersheim and An-
drea Fritz, who thaught and trained me in the basic methods of cell culture and laboratory
methods. They were always available to help me to achieve my goals with excellent advice
and very helpful experience.
I want to thank my parents for always supporting me, for their constant motivation and en-
couragement. Without their help I would never have been able to achieve all this.

113
ERKLÄRUNG § 5 ABS. 1 ZUR DATENAUFBEWARUNG
Hiermit erkläre ich, dass die dieser Dissertation zu Grunde liegenden Originaldaten
bei mir,
Melanie Wosnitza, Wallstr. 15-17, 52064 Aachen,
hinterlegt sind.

114
CURRICULUM VITAE Persönliche Daten geboren am 08.07.1980 in Düsseldorf, deutsch, ledig
Eltern Sigrid Wosnitza, geborene Dierdorf, PA, Ass. Geschäftsleitung, Unifrax GmbH, Düsseldorf Achim Wosnitza, Abteilungs- und Projektleiter, RWE AG, Essen/ Ruhestand
Schulausbildung Grundschule Montessorie-Grundschule, Düsseldorf 1987-1991 Gymnasium Städtisches Gymnasium Koblenzer Straße, Düsseldorf 1991-2000 Abitur 06/2000 Universitäre Ausbildung Studium der Elektrotechnik RWTH Aachen WS 2000/01 Studium der Humanmedizin RWTH Aachen WS 2001/02-
WS 2007/08 Examina Physikum 09/2003 Zweiter Abschnitt der Ärztlichen Prüfung 10/2007 Famulaturen Klinik für Plastische Chirurgie, Hand- und Verbrennungschirurgie,
Universitätsklinikum RWTH Aachen 03-04/2004
Hautklinik Universitätsklinikum RWTH Aachen 03-04/2005 Department of Dermatology, University of Tartu, Tartu, Estland 08/2005 Department of Dermatology, University of Melbourne,
Royal Melbourne Hospital, Melbourne, Australien 03-04/2006
Praktisches Jahr 1. Tertial Department of Internal Medicine, Box Hill Hospital,
Monash University, Melbourne, Australien 08-12/2006
2. Tertial Chirurgische Klinik, Stadtspital Waid, Zürich, Schweiz 12/2006-04/2007 3.Tertial Hautklinik Universitätsklinikum RWTH Aachen 04-07/2007 Forschung Dissertation Klinik für Plastische Chirurgie, Hand- und Verbrennungschirurgie,
Universitätsklinikum RWTH Aachen, Univ.-Prof. Dr. Dr. med. Prof. h.c. (RC) N. Pallua
03/2004-08/2006
Forschungstätigkeit Forschung in der Klinik für Plastische Chirurgie, Hand- und Ver-
brennungschirurgie/ IZKF BIOMAT als studentische Hilfskraft, Uni-versitätsklinikum RWTH Aachen, Univ.-Prof. Dr. Dr. med. Prof. h.c. (RC) N. Pallua
07/2005-12/2006
Forschung in der Hautklinik, Universitätsklinikum RWTH Aachen,
Prof. Dr. med. J. M. Baron 11/2005-08/2006

115
Ärztliche Tätigkeit Approbation als Ärztin
Universitätsklinikum RWTH Aachen 14.11.2007
Assistenzärztin Hautklinik Universitätsklinikum RWTH Aachen, Univ.-Prof. Dr. med. H. F. Merk
Seit 11/2007




















