
Differential response of indica rice genotypes to NaCl stress in relation to physiological and...
13
This article was downloaded by:[INFLIBNET, India order 2005] On: 31 August 2007 Access Details: [subscription number 772695191] Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Archives of Agronomy and Soil Science Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713453776 Differential response of indica rice genotypes to NaCl stress in relation to physiological and biochemical parameters Online Publication Date: 01 October 2007 To cite this Article: Kumar, Vinay, Shriram, Varsha, Jawali, Narendra and Shitole, Mahadev Ganpat (2007) 'Differential response of indica rice genotypes to NaCl stress in relation to physiological and biochemical parameters', Archives of Agronomy and Soil Science, 53:5, 581 - 592 To link to this article: DOI: 10.1080/03650340701576800 URL: http://dx.doi.org/10.1080/03650340701576800 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material. © Taylor and Francis 2007
-
Upload
manpowerforncr -
Category
Documents
-
view
2 -
download
0
Transcript of Differential response of indica rice genotypes to NaCl stress in relation to physiological and...

This article was downloaded by:[INFLIBNET, India order 2005]On: 31 August 2007Access Details: [subscription number 772695191]Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Archives of Agronomy and SoilSciencePublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713453776
Differential response of indica rice genotypes to NaClstress in relation to physiological and biochemicalparameters
Online Publication Date: 01 October 2007To cite this Article: Kumar, Vinay, Shriram, Varsha, Jawali, Narendra and Shitole,Mahadev Ganpat (2007) 'Differential response of indica rice genotypes to NaClstress in relation to physiological and biochemical parameters', Archives ofAgronomy and Soil Science, 53:5, 581 - 592To link to this article: DOI: 10.1080/03650340701576800URL: http://dx.doi.org/10.1080/03650340701576800
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.
© Taylor and Francis 2007

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7
Differential response of indica rice genotypes to NaCl stressin relation to physiological and biochemical parameters
VINAY KUMAR1, VARSHA SHRIRAM1, NARENDRA JAWALI2, &
MAHADEV GANPAT SHITOLE1
1Department of Botany, University of Pune, Ganeshkhind, and 2Bhabha Atomic Research Centre,
Molecular Biology Division, Trombay, Mumbai, India
(Received 8 May 2007; accepted 17 July 2007)
AbstractThe effect of NaCl stress was studied in indica rice cultivars, Panvel-3 (tolerant), Kalarata (moderatelytolerant) and Karjat-3 (sensitive) under various levels (50 – 300 mM) of salinity besides control (0 mM).Salinity stress decreased germination percentage, plant growth, biomass production and chlorophyllpigments, and increased malondialdehyde level (lipid peroxidation) and free proline accumulation.Salinity-induced decrease in germination percentage, biomass production, chlorophyll and total proteincontents were significantly higher in Karjat-3 than Panvel-3. The free proline content at all the levels aswell as the magnitude of increase in accumulation with increasing salinity was also highest in Panvel-3,as at 300 mM NaCl concentration, it was 8 times more than control, while it was about 5 times more inKarjat-3 and around 4 times in Kalarata. The level of lipid peroxidation was lowest at all levels of salinityin Panvel-3 as compared with Karjat-3, however, Kalarata showed intermediate results. Results showedsalinity tolerance of Panvel-3 was manifested by lower decrease in germination, plant growth andchlorophyll pigments and associated with higher proline accumulation and lower lipid peroxidationunder high salinity stress.
Keywords: NaCl stress, proline, lipid peroxidation, chlorophyll, biomass
Introduction
Soil salinity is a major abiotic stress problem around the globe especially in arid and semi-arid
regions and irrigation areas. Salinity affects nearly 20% of the world’s cultivated area and
about half of the world’s total irrigated lands (Zhu 2001; FAO 2006). In Asia alone, 21.5
million ha of land area is thought to be salt-affected, with India having 8.6 million ha of such
area which constitutes a major part of problem soils in India (Sahi et al. 2006).
Salinity adversely affects the quantity and quality of crop produce (Sahi et al. 2006). Salt
stress affects almost every aspect of plant physiology at both whole plant and cellular levels
through osmotic and ionic stress (Hasegawa et al. 2000; Murphy & Durako 2003). Salinity is
Correspondence: Prof. Mahadev Ganpat Shitole, PhD, Department of Botany, University of Pune, Ganeshkhind, Pune-411007,
India. E-mail: [email protected]
Archives of Agronomy and Soil Science
October 2007; 53(5): 581 – 592
ISSN 0365-0340 print/ISSN 1476-3567 online � 2007 Taylor & Francis
DOI: 10.1080/03650340701576800

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7 detrimental to the various processes of crops such as seed germination, seedling growth and
vigour, vegetative growth, flowering and fruit set and ultimately it causes diminished
economic yield and also quality of produce (Sairam & Tyagi 2004).
Rice is the world’s most important cereal crop (Bajaj & Mohanty 2005) with around
610 million tonnes production globally (IRRI 2005). Rice with its relatively small genome size
(*430 Mb), ease of transformation, well developed and known detailed genetics (Sahi et al.
2006), availability of a dense physical map and molecular markers (Wu et al. 2002),
full-genome transcription profiling using high density oligonucleotide tiling microarrays
(Li et al. 2006) together with its complete genome sequence (Sasaki et al. 2005) is considered
as a model monocot system for various biotechnological, metabolic, genetic engineering and
functional genomics development studies worldwide (Shimamoto & Kyozuka 2002; Bajaj &
Mohanty 2005; Ge et al. 2006). In addition, rice shares extensive synteny among the other
cereals thereby increasing the utility of this system.
Although India is second only to China with more than 136 million tonnes rice production
from 43 million ha lands (FAOSTAT 2005), the average yield in India is very low (3 t/ha) as
compared to some other Asian rice-producing countries like China, Korea and Japan. The
main constraints for the low yield are dependency on monsoon (floods and draughts),
improper irrigation and drainage facilities, overuse of fertilizers and soil-related problems
including salinity and alkalinity. The yield of rice especially Asian rice (sativa) is susceptible to
salinity (Flowers & Yeo 1981; Anil et al. 2005).
Increased salt tolerance of crops including rice is needed to sustain food production in
various regions of the world (Munns et al. 2006). As saline soils and saline waters are
common around the world, great effort has been devoted to understand physiological aspects
of tolerance to salinity in plants, as a basis for plant breeders to develop salinity-tolerant
genotypes. In spite of this great effort, only a small number of cultivars, partially tolerant to
salinity, have been developed. Interaction of salt with the plants may depend upon salt
concentration and genotype. Therefore, screening for salt tolerance of genotypes that could
withstand extreme soil salinity will ensure future crop production (Jogeswar et al. 2006).
In the present investigation, we have studied the differential response of indica rice cultivars
to NaCl stress in terms of physiological and biochemical parameters including plant
germination and growth, chlorophyll contents, proline, total proteins and lipid peroxidation.
Materials and methods
Plant material
Indica rice (Oryza sativa L.) cultivars namely Karjat-3, Kalarata and Panvel-3 were selected
for this study and certified seeds were obtained from The Regional Rice Research Station,
Karjat, Maharashtra, India and The Saline Land Research Station, Panvel, Maharashtra,
India.
Salinity treatment and culture conditions
Seeds of all three cultivars were surface sterilised with 0.1% mercuric chloride for 10 min and
then washed several times with sterile distilled water. Twenty seeds of each cultivar (cv.) were
sown in a Petri dish (10 cm diameter) containing germination paper and the experiment was
carried out in triplicate. Every day 10 ml Yoshida’s nutrient solution (with or without varying
NaCl levels, i.e. 50, 100, 150, 200, 250 and 300 mM) was applied per Petri dish and all the
observations were recorded on the 14th day after sowing.
582 V. Kumar et al.

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7 Plant growth analysis
The germination percentage, root length, shoots length, root/shoot ratio, fresh weight (FW)
and dry weight (DW) of seedlings were recorded on the 14th day of germination. The DW
was observed by heating the samples in a hot air oven at 608C for 48 h.
Biochemical analysis
Determination of chlorophyll. Chlorophyll contents were estimated by extracting 100 mg of the
fresh leaf material in 10 ml dimethyl sulfoxide (DMSO) (Hiscox & Israelstam 1979). The
samples were heated at 658C for 4 h and than the absorbance of extract recorded at 665 and
645 nm on UV-VIS spectrophotometer (Shimadzu- 1601, Japan). Chlorophyll contents were
calculated as per standard method (Arnon 1949).
Determination of lipid peroxidation. The level of lipid peroxidation was measured in terms of
malondialdehyde (MDA) contents (Heath & Packer 1968). Fresh shoot and root samples
(500 mg) were homogenized in 10 ml of 0.1% trichloro-acetic acid (TCA). The
homogenate was centrifuged at 15,000 g for 5 min. To 2 ml of aliquot of the supernatant,
4 ml of 0.5% thiobarbituric acid (TBA) in 20% TCA was added. The mixture was heated
at 958C for 30 min and then quickly cooled in ice bath. This was followed by centri-
fugation at 10,000 g for 10 min to remove suspended turbidity and then the absorbance
was recorded at 532 nm. The value for non-specific absorption at 600 nm was
subtracted and the MDA content was calculated using its absorption coefficient of
155 mmol71 cm71.
Determination of proline. The proline content was determined separately in shoots and roots
following the method of Bates et al. (1973). Samples (500 mg) were homogenized in 5 ml of
sulphosalicylic acid (3%) using mortar and pestle. Two ml aliquot of supernatant was mixed
with an equal volume of glacial acetic acid and acid ninhydrin. The reaction mixture was
boiled in water bath at 1008C for 30 min. The reaction was terminated in an ice bath
following by addition of 4 ml toluene and then transferring to a separating funnel. After
thorough mixing, the chromophore containing toluene was separated and absorbance was
recorded at 520 nm in spectrophotometer against toluene blank. Concentration of proline
was estimated by referring to a standard curve of proline.
Protein estimation. Proteins were estimated using Lowry et al. (1951) method. Fresh
samples (250 mg) were homogenized in 2.5 ml of phosphate buffer (pH 7.0). The extract
was centrifuged at 5000 g for 15 min at 48C and the supernatant was transferred to a
tube containing a mixture of 20 ml acetone and 14 ml b-Mercaptoethanol for pre-
cipitation of protein. The sample tubes were stored at 08C for 5 h and then centrifuged at
10,000 g for 20 min. The supernatant was discarded and the pellet was dissolved in 2.5 ml
1 N sodium hydroxide solution. Aliquot of 0.2 ml from this sample was used to prepare
the reaction mixture. The intensity of blue colour developed was recorded at 660 nm and
protein concentration was measured using bovine serum albumin as standard.
Statistical analysis. Each Petri dish was considered as replicate and all of the treatments
were repeated three times and data are expressed as+ standard error (SE). The data was
subjected to analysis of variance (ANOVA) to detect significant difference between means.
The level of significance was also checked for least significant differences (LSD) at
Response of rice genotypes to NaCl stress 583

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7 p� 0.05. All the statistical analyses were done by using MSTAT-C statistical software
package.
Results
Effect of NaCl stress on germination and growth parameters
The seed germination decreased with increasing salt concentration from 50 – 300 mM NaCl.
However, the effect of salt stress on seed germination varied between the cultivars (see
Table I). Up to 50 mM NaCl, 100% germination was observed in all the cultivars; however,
beyond that level noticeable reduction in germination percentage was evident, with more
pronounced reduction at more than 100 mM NaCl concentrations. In Panvel-3, germination
was 100% up to 150 mM NaCl and was comparably least affected at higher salt
concentrations. The increase in salinity decreased the shoot and root lengths considerably
in all the cultivars, with more reduction in shoot growth than the roots. Rice genotypes
Panvel-3 and Kalarata showed lesser decline in shoot length while Karjat-3 showed more
stunted shoots under high salinity stress (see Table I). The root/shoot ratio was increased
markedly with salinity in Karjat-3 followed by Kalarata; however, contrasting to these results,
there was a decline in root/shoot ratio beyond 100 mM NaCl and at 300 mM NaCl, it was
even less than the control. The results of the root length shoot length and root/shoot ratio
makes it clear that Panvel-3 was less affected by salinity stress followed by Kalarata, while
these parameters were markedly affected in Karjat-3.
Table I. Effect of NaCl stress on germination percentage, root length, shoot length and root/shoot ratio in rice
cultivars. Results are mean of three replicates+ standard error (SE). LSD for cultivars and treatments were
significant at p�0.05.
Cultivar
NaCl stress
(mM)
Germination
percentage (%)
Root length
(cm) Mean+SE
Shoot length
(cm) Mean+SE
Root/shoot
ratio
Karjat-3 0 (Control) 100+ 0.5 5.40+ 0.18 4.06+0.18 1.33
50 100+ 1.1 5.18+ 0.30 3.93+0.13 1.32
100 96+ 1.5 6.10+ 0.42 3.13+0.09 1.95
150 90+ 1.5 4.72+ 0.22 2.42+0.29 1.95
200 75+ 1.1 2.72+ 0.15 1.66+0.16 1.64
250 70+ 1.3 1.88+ 0.04 0.78+0.03 2.39
300 25+ 1.5 1.48+ 0.12 0.41+0.02 3.56
Kalarata 0 (Control) 100+ 0.9 6.65+ 0.39 4.28+0.31 1.55
50 100+ 1.4 7.52+ 0.36 4.18+0.08 1.58
100 96+ 1.2 7.03+ 0.53 2.51+0.28 1.75
150 94+ 1.3 4.47+ 0.34 1.07+0.16 1.74
200 90+ 1.1 3.14+ 0.17 0.81+0.07 1.88
250 90+ 2.1 1.67+ 0.23 0.92+0.10 1.76
300 73+ 1.6 1.67+ 0.20 0.90+0.10 1.68
Panvel-3 0 (Control) 100+ 1.0 6.13+ 0.56 5.21+0.13 1.18
50 100+ 0.9 6.10+ 0.20 4.55+0.12 1.34
100 100+ 1.2 6.27+ 0.31 4.47+0.13 1.40
150 100+ 1.1 5.10+ 0.26 3.83+0.16 1.33
200 96+ 1.3 4.14+ 0.22 3.33+0.14 1.24
250 90+ 1.6 2.16+ 0.14 1.93+0.10 1.12
300 85+ 1.9 2.03+ 0.24 1.75+0.05 1.16
584 V. Kumar et al.

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7 A negative correlation between biomass production and salinity stress was evident from the
results obtained (Table II) as FW and DW gradually decrease in all the cultivars with increase
in NaCl concentration. The reduction in biomass under salinity stress was noticeably higher
in Karjat-3, and lowest in Panvel-3, while intermediate results were obtained in the cv.
Kalarata.
Chlorophyll content
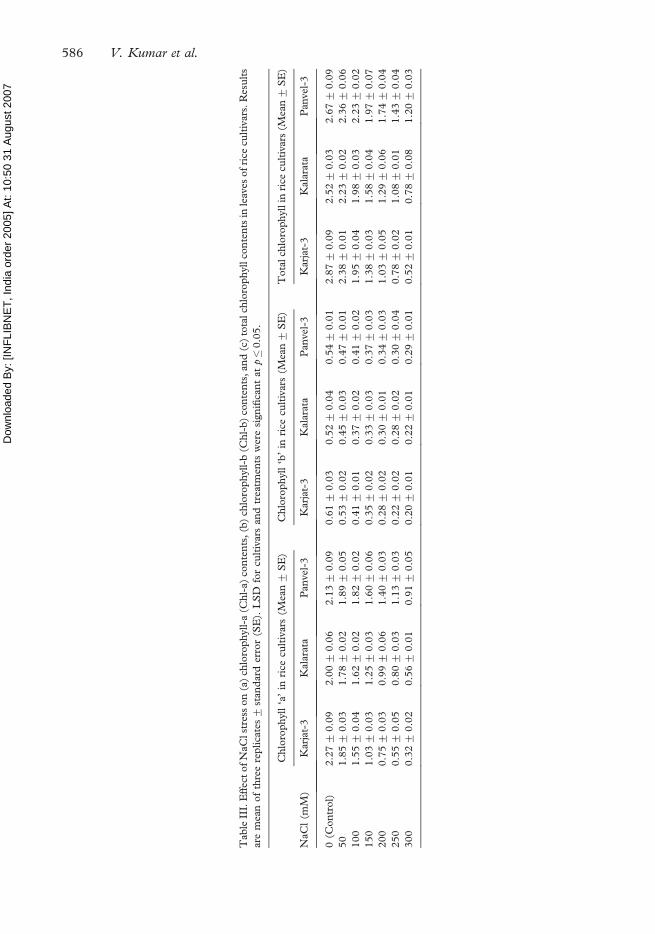
The Chl a, b and total Chl content were influenced significantly by NaCl stress and these
pigments were gradually decreased in leaves with increasing salt concentrations in all the
cultivars irrespective of their salt tolerance (see Table III). Comparably higher Chlorophyll
contents were observed in Karjat-3 cv. under control conditions than other genotypes.
However, the decrease under salt stress was also higher in Karjat-3. The rate of decrease in
Chl with increase in salinity stress was lowest in Panvel-3 and highest in Karjat-3.
Lipid peroxidation
The results depicted in Figure 1 clearly revealed that increased lipid peroxidation (in terms of
MDA content) was observed with increasing NaCl concentrations in all the cultivars. When
exposed to 300 mM NaCl, MDA level in Karjat-3 was about 5 times higher than control, in
Karjat-3, it was 3.5, while in contrast to these genotypes, lesser extent of lipid peroxidation
was noticed in Panvel-3, where about 2.5 times more MDA content than control was
recorded at 300 mM NaCl. This indicates that Panvel-3 is able to tolerate salinity-induced
oxidative damage better than other two cultivars.
Proline content
Amongst the three rice cultivars, comparably very high proline content was observed in
Panvel-3 than the other two cultivars, at non-saline condition. NaCl stress induced an
elevation in proline contents in all cultivars irrespective of their salt stress tolerance, as evident
from the results presented in Figure 2. There was a steep increase in proline content in shoots
of Panvel-3 under salt stress ranging from 50 – 300 mM and 36.47 m mol g71 fresh weight
proline was recorded at 300 mM salinity level, i.e. about 8 times more than the non-saline
conditions followed by approximately 5 times more proline at this salinity concentration
Table II. Effect of NaCl stress on fresh and dry weights of shoots of rice cultivars. Results are mean of three
replicates+ standard error (SE). LSD for cultivars and treatments were significant at p�0.05.
NaCl (mM)
Fresh weight of shoots (mg) in rice
cultivars (Mean+SE)
Dry weight of shoots (mg) in rice
cultivars (Mean+SE)
Karjat-3 Kalarata Panvel-3 Karjat-3 Kalarata Panvel-3
0 (Control) 139.3+ 3.5 133.7+ 5.8 135.2+3.9 25.2+ 1.0 24.4+ 3.0 24.5+2.0
50 121.5+ 4.0 129.0+ 1.2 135.5+4.7 24.5+ 1.2 24.3+ 1.1 24.7+1.5
100 111.6+ 3.0 122.5+ 2.3 122.7+2.2 22.7+ 1.6 20.3+ 1.0 23.5+1.7
150 93.8+ 2.0 121.2+ 4.2 121.5+2.6 19.0+ 1.2 19.7+ 0.8 20.3+1.0
200 85.6+ 2.5 103.5+ 2.8 115.8+1.9 17.3+ 1.1 17.2+ 1.5 18.9+0.7
250 82.5+ 2.8 88.4+ 4.0 101.9+2.5 13.5+ 1.0 18.1+ 1.7 18.5+0.5
300 31.3+ 2.0 80.9+ 3.0 95.5+1.8 9.1+ 0.9 16.3+ 1.3 17.2+0.6
Response of rice genotypes to NaCl stress 585
Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7
Tab
leII
I.E
ffec
to
fN
aClst
ress
on
(a)
chlo
rop
hyl
l-a
(Ch
l-a)
con
ten
ts,(b
)ch
loro
ph
yll-
b(C
hl-
b)
con
ten
ts,an
d(c
)to
talch
loro
ph
yllco
nte
nts
inle
aves
of
rice
cult
ivar
s.R
esu
lts
are
mea
no
fth
ree
rep
lica
tes+
stan
dar
der
ror
(SE
).L
SD
for
cult
ivar
san
dtr
eatm
ents
wer
esi
gn
ifica
nt
atp�
0.0
5.
NaC
l(m
M)
Ch
loro
ph
yll
‘a’
inri
cecu
ltiv
ars
(Mea
n+
SE
)C
hlo
rop
hyl
l‘b
’in
rice
cult
ivar
s(M
ean+
SE
)T
ota
lch
loro
ph
yllin
rice
cult
ivar
s(M
ean+
SE
)
Kar
jat-
3K
alar
ata
Pan
vel-
3K
arja
t-3
Kal
arat
aP
anve
l-3
Kar
jat-
3K
alar
ata
Pan
vel-
3
0(C
on
tro
l)2
.27+
0.0
92.0
0+
0.0
62
.13+
0.0
90
.61+
0.0
30
.52+
0.0
40
.54+
0.0
12
.87+
0.0
92
.52+
0.0
32
.67+
0.0
9
50
1.8
5+
0.0
31.7
8+
0.0
21
.89+
0.0
50
.53+
0.0
20
.45+
0.0
30
.47+
0.0
12
.38+
0.0
12
.23+
0.0
22
.36+
0.0
6
10
01
.55+
0.0
41.6
2+
0.0
21
.82+
0.0
20
.41+
0.0
10
.37+
0.0
20
.41+
0.0
21
.95+
0.0
41
.98+
0.0
32
.23+
0.0
2
15
01
.03+
0.0
31.2
5+
0.0
31
.60+
0.0
60
.35+
0.0
20
.33+
0.0
30
.37+
0.0
31
.38+
0.0
31
.58+
0.0
41
.97+
0.0
7
20
00
.75+
0.0
30.9
9+
0.0
61
.40+
0.0
30
.28+
0.0
20
.30+
0.0
10
.34+
0.0
31
.03+
0.0
51
.29+
0.0
61
.74+
0.0
4
25
00
.55+
0.0
50.8
0+
0.0
31
.13+
0.0
30
.22+
0.0
20
.28+
0.0
20
.30+
0.0
40
.78+
0.0
21
.08+
0.0
11
.43+
0.0
4
30
00
.32+
0.0
20.5
6+
0.0
10
.91+
0.0
50
.20+
0.0
10
.22+
0.0
10
.29+
0.0
10
.52+
0.0
10
.78+
0.0
81
.20+
0.0
3
586 V. Kumar et al.

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7
in Karjat-3 and more than 4 times higher in Kalarata. These results clearly indicated that
Panvel-3 has higher salt-induced proline accumulation than other two cultivars, which may
have played a role in salt stress tolerance.
Figure 1. Effect of different concentrations of NaCl stress on lipid peroxidation in rice cultivars. Each value represents
mean of three replications and vertical bars indicate+SE. Data are statistically significant at p50.05.
Figure 2. Effect of different concentrations of NaCl stress on proline content in rice cultivars. Each value represents
mean of three replications and vertical bars indicate+SE. Data are statistically significant at p50.05.
Response of rice genotypes to NaCl stress 587
Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
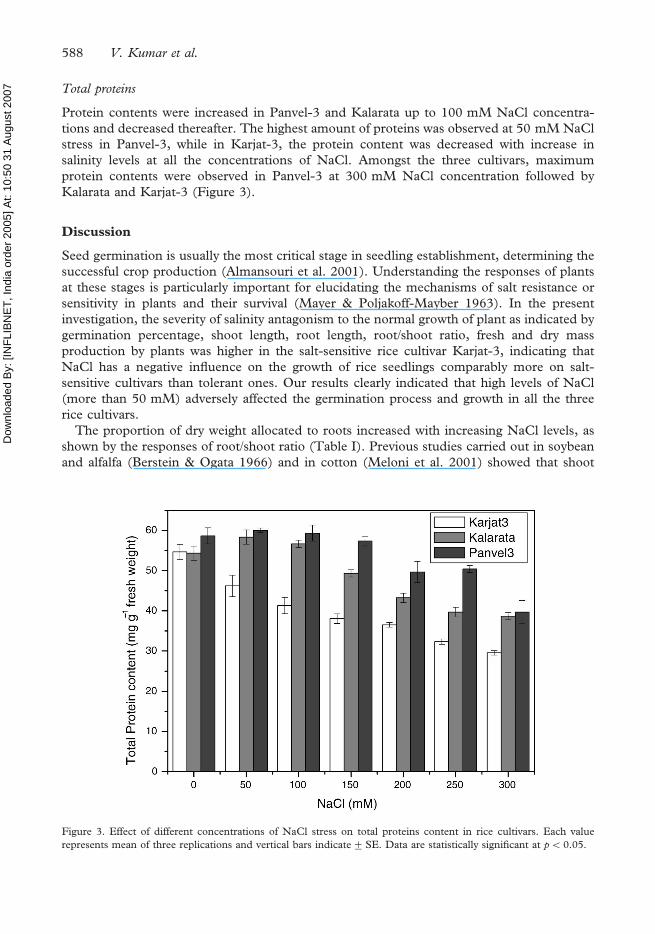
7 Total proteins
Protein contents were increased in Panvel-3 and Kalarata up to 100 mM NaCl concentra-
tions and decreased thereafter. The highest amount of proteins was observed at 50 mM NaCl
stress in Panvel-3, while in Karjat-3, the protein content was decreased with increase in
salinity levels at all the concentrations of NaCl. Amongst the three cultivars, maximum
protein contents were observed in Panvel-3 at 300 mM NaCl concentration followed by
Kalarata and Karjat-3 (Figure 3).
Discussion
Seed germination is usually the most critical stage in seedling establishment, determining the
successful crop production (Almansouri et al. 2001). Understanding the responses of plants
at these stages is particularly important for elucidating the mechanisms of salt resistance or
sensitivity in plants and their survival (Mayer & Poljakoff-Mayber 1963). In the present
investigation, the severity of salinity antagonism to the normal growth of plant as indicated by
germination percentage, shoot length, root length, root/shoot ratio, fresh and dry mass
production by plants was higher in the salt-sensitive rice cultivar Karjat-3, indicating that
NaCl has a negative influence on the growth of rice seedlings comparably more on salt-
sensitive cultivars than tolerant ones. Our results clearly indicated that high levels of NaCl
(more than 50 mM) adversely affected the germination process and growth in all the three
rice cultivars.
The proportion of dry weight allocated to roots increased with increasing NaCl levels, as
shown by the responses of root/shoot ratio (Table I). Previous studies carried out in soybean
and alfalfa (Berstein & Ogata 1966) and in cotton (Meloni et al. 2001) showed that shoot
Figure 3. Effect of different concentrations of NaCl stress on total proteins content in rice cultivars. Each value
represents mean of three replications and vertical bars indicate+SE. Data are statistically significant at p50.05.
588 V. Kumar et al.

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7 growth was more inhibited by NaCl than root growth. Our results are in harmony of all these
reports and root/shoot was increased with salt concentration, with lower to higher root/shoot
ratio in sensitive to tolerant cultivars.
Likewise, the biomass production was influenced negatively by NaCl stress significantly.
However, highest fresh weight and dry weight was recorded in Panvel-3 while lowest dry
weight of was observed in Karjat-3 at 300 mM NaCl. Thus, it was clear from the results that
the Panvel-3, a salt-tolerant rice cultivar could withstand high NaCl stress comparably better
than other rice cultivars. Earlier reports by Yeo and Flowers (1983); Garcia et al. (1995) and
Anil et al. (2005) observed similar trends and reported lesser effect of NaCl stress on
germination process and better survival of salt-tolerant rice cultivars than the salt-sensitive
rice cultivars. Furthermore, the effect of salinity stress on biomass production in rice has been
described by Khan and Panda (2002). Our results are in agreement of these reports.
Chlorophyll concentration in stressed tissues has been advocated as an index of tissue
tolerance to NaCl (Lutts et al. 1996). Chlorophyll contents in our study were markedly
reduced under NaCl stress in leaves of rice cultivars at vegetative phase. Salinity-sensitive rice
genotype Karjat-3 showed significantly higher decline in Chl a, b and total Chl contents than
tolerant genotypes Panvel-3 and Kalarata under increasing salt stress. Similar observations are
reported earlier and chlorophyll contents were reduced more in salt-sensitive cultivars than
the tolerant ones. Lutts et al. (1996) observed a highly significant decrease in chlorophyll
concentration with increasing salinity in salt-sensitive rice genotypes, while a much lesser
effect was recorded in salt-tolerant genotypes. Misra et al. (1997) reported that the
chlorophyll contents were affected heavily by NaCl stress in salt-sensitive rice cultivar Jaya,
while in the salt-tolerant cultivar Damodar, more chlorophyll accumulation was observed
than the control plants. According to Choudhury and Choe (1996), a decline in photo-
synthetic rate with increased concentration of NaCl may be associated with decreased
pigmentation. Our results clearly supported this hypothesis.
Our results clearly indicated that amongst the rice cultivars, the damage to the lipid
membranes was comparatively less severe in the salt-tolerant cultivar Panvel-3. The severe
damage was observed in Karjat-3 while intermediate responses were observed in Kalarata.
The results of the present investigation are in agreement of earlier reports, in which higher
lipid peroxidation in sensitive cultivars than the tolerant ones have been reported.
Vaidyanathan et al. (2003) and Demiral and Turkan (2005) reported greater lipid peroxi-
dation under salinity stress in salt-sensitive rice genotypes than in salt-tolerant ones. The
lower level in shoots and roots of Panvel-3 than of Karjat-3 and Kalarata suggests that it may
have better protection against oxidative damage under salt stress.
Proline is one of the most important and widely studied osmolytes found in plants and is
related with abiotic stress tolerance. Proline is generally assumed to serve as a physiologically
compatible solute that increases as needed to maintain a favourable osmotic potential between
the cell and its surroundings (Pollard & Wyn Jones 1979). In addition to this, proline is an
important osmoprotectant in plants and can protect the photosynthetic machinery against
salt-induced damage (Sivakumar et al. 2002). Rapid accumulation of free proline is a typical
response to salt stress (Demiral & Turkan 2005). When exposed to drought or a high salt
content in soil, many plants have been observed to accumulate high amounts of proline, in
some cases several times the sum of all other amino acids (Mansour 2000). In the present
investigation, the free proline content was significantly increased in the stressed plants over
control plants at all stress regimes in all the cultivars. Significantly higher accumulation of
proline was observed at 300 mM NaCl in salt tolerant cv. Panvel-3 compared to comparably
sensitive cultivars Kalarata and Karjat-3. Similar to our results, Sairam et al. (2002) in
wheat and Jogeswar et al. (2006) in sorghum observed much higher proline accumulation in
Response of rice genotypes to NaCl stress 589

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7 the salt-tolerant genotype than the salt-sensitive genotype. Contrary to these results,
Vaidyanathan et al. (2003) and Demiral and Turkan (2005) concluded higher proline
accumulation in salt-sensitive rice cultivars than salt-tolerant cultivars under high salinity
stress. A large number of researchers have advocated the major role played by the proline in
salt stress tolerance and post-stress recovery of the plants. Proline has been found to protect
cell membranes of onion against salt injury (Mansour 1998). Furthermore, Sultana et al.
(1999) have suggested that proline accumulation in both salinized leaves and grains of rice
plants is implicated in osmotic adjustment to salinity. It is suggested that proline may be the
major source of energy and nitrogen during immediate post-stress metabolism and the
accumulated proline apparently supplies energy for growth and survival thereby inducing
salinity tolerance (Ahmad & Jhon 2005). The results of the present investigation suggested
that comparably higher amount of salt-induced synthesis and accumulation of proline in
Panvel-3 under salinity stress may play a role in combating salt stress in this cultivar.
The protein contents were increased up to 100 mM NaCl concentration than the control in
salt-tolerant cultivars Panvel-3 and Kalarata; however, the protein levels were highly reduced
at all levels of salinity from 50 mM to 300 mM NaCl in Karjat-3. Similar trends were
reported by Lutts et al. (1996) and Misra et al. (1997) and observed comparably higher
reduction in protein contents in salt-sensitive rice cultivars than the tolerant ones under NaCl
stress. However, contrary to these results, Ahmad & Jhon (2005) concluded that NaCl stress
had a positive impact on total protein contents in Pisum sativum and observed a gradual
increase in protein contents with increasing salinity.
Conclusion
From the results of the present investigation, we can conclude that NaCl stress affected
germination and plant growth significantly, with lesser affects on Panvel-3, while Karjat-3 was
affected most. Comparably higher amounts as well as accumulation of proline and lowest
peroxidation of lipid membrane under increasing salt stress in Panvel-3 as compared with
other cultivars, suggested that this cultivar may have a better antioxidant machinery to combat
the salt stress induced injuries in this cultivar.
Acknowledgements
This work was supported by grant from Board of Research in Nuclear Sciences, Department
of Atomic Energy, Government of India (Grant No. 2004/37/28/BRNS) to MGS and
research fellowship to the first author. The authors also acknowledge the facilities used for this
work at the Department of Botany, University of Pune created under ASIST and DRS-SAP
programs of University Grants Commission, India.
References
Ahmad P, Jhon R. 2005. Effect of salt stress on growth and biochemical parameters of Pisum sativum L. Arch Agron
Soil Sci 51:665 – 672.
Almansouri M, Kinet JM, Lutts S. 2001. Effect of salt and osmotic stresses on germination in durum wheat (Triticum
durum Desf.). Plant Soil 23:243 – 254.
Anil VS, Krishnamurthy P, Kuruvilla S, Sucharitha P, Thomas G, Mathew MK. 2005. Regulation of the uptake and
distribution of Naþ in shoots of rice (Oryza sativa) variety pokkali: Role of Ca2þ in salt tolerance response.
Physiol Plant 124:451 – 464.
Arnon DI. 1949. Copper enzymes in isolated chloroplasts. Polyphenol oxidase in Beta vulgaris. Plant Physiol
24:1 – 15.
590 V. Kumar et al.

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7
Bajaj S, Mohanty A. 2005. Recent advances in rice biotechnology – towards genetically superior transgenic rice. Plant
Biotechnol J 3:275 – 307.
Bates L, Waldren RP, Teare JD. 1973. Rapid determination of free proline for water stress studies. Plant Soil
39:205 – 207.
Berstein L, Ogata G. 1966. Effects of salinity on nodulation, nitrogen fixation and growth of soybean and alfalfa.
Agron J 58:201 – 203.
Choudhury NK, Choe HT. 1996. Photoprotective effect of pigment content and photochemical activities of wheat
chloroplasts aging in vitro. Biol plant 38:61 – 69.
Demiral T, Turkan I. 2005. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots
of two rice cultivars in salt tolerance. Environ Exp Bot 53:247 – 257.
FAO. 2006. Global network on integrated soil management for sustainable use of salt affected soils. Rome, Italy: FAO
Land and Plant Nutrition Management Service. http://www.fao.org/ag/agI/agII/spush
FAOSTAT. 2005. http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID¼567
Flowers TJ, Yeo AR. 1981. Variability in the resistance of sodium chloride salinity within rice (Oryza sativa L.)
varieties. New Phytol 88:363 – 373.
Garcia A, Senadhira D, Flowers TJ, Yeo AR. 1995. The effects of selection for sodium transport and of selection for
agronomic characteristics upon salt resistance in rice (Oryza sativa L.). Theor Appl Genet 90:1106 – 1111.
Ge X, Chu Z, Lin Y, Wang S. 2006. A tissue culture system for different germplasms of indica rice. Plant Cell Rep
25:392 – 402.
Hasegawa M, Bressan R, Zhu JK, Bhonert H. 2000. Plant cellular and molecular responses to high salinity. Ann. Rev
Plant Physiol 51:463 – 499.
Heath RL, Packer L. 1968. Photooxidation in isolated chloroplast. I. Kinetics and stoichiometry of fatty acid
peroxidation. Arch Biochem Biophys 125:189 – 198.
Hiscox JD, Israelstam GF. 1979. A method of extraction of chloroplast from leaf tissue without maceration. Can J Bot
57:1332 – 1334.
IRRI. 2005. Atlas of Rice & World Rice Statistics: Rice around the World. http://www.irri.org/science/ricestat/pdfs/
WRS2005-Table01.pdf
Jogeswar G, Pallela R, Jakka NM, Reddy PS, Rao JV, Sreeniwasulu N, Kavi Kishor PB. 2006. Antioxidative response
in different sorghum species under short term salinity stress. Acta Physiol Plant 28:465 – 475.
Khan MH, Panda SK. 2002. Induction of oxidative stress in roots of Oryza sativa L. in response to salt stress.
Biol Plant 45:625–627.
Li L, Wang X, Stolc V, Li X, Zhang D, Su N, Tongprasit W, Li S, Chang Z, Wang J, Deng XW. 2006. Genome-wide
transcription analyses in rice using tiling microarrays. Nature Genet 38:124 – 129.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Protein measurement with the folin phenol reagent. J Biol
Chem 193:265 – 275.
Lutts S, Kinet JM, Bouharmont J. 1996. Effects of salt stress on growth, mineral nutrition and proline accumulation
in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity tolerance. Plant Growth
Regul 19:207 – 218.
Mansour MMF. 1998. Protection of plasma membrane of onion epidermal cells by glycinebetaine and proline against
NaCl stress. Plant Physiol Biochem 36:767 – 772.
Mansour MMF. 2000. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol Plant
43:491 – 500.
Mayer AM, Poljakoff-Mayber A. 1963. The germination of seeds. Oxford, London: Pergamon Press.
Meloni DA, Oliva MA, Ruiz HA, Martinez CA. 2001. Contribution of proline and inorganic solutes to osmotic
adjustment in cotton under salt stress. J Plant Nutr 24:599 – 612.
Misra AN, Sahu SM, Misra M, Singh P, Meera I, Das N, Kar M, Sahu P. 1997. Sodium chloride induced changes in
leaf growth and pigment and protein contents in two rice cultivars. Biol Plant 39:257 – 262.
Munns R, James AR, Lauchli A. 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J Exp
Bot 57:1025 – 1043.
Murphy KST, Durako MJ. 2003. Physiological effects of short-term salinity changes on Ruppia maritima. Aquat Bot
75:293 – 309.
Pollard A, Wyn Jones RG. 1979. Enzyme activities in concentrated solutions of glycienebetaine and other solutes.
Planta 144:291 – 298.
Sahi C, Singh A, Kumar K, Blumwald E, Grover A. 2006. Salt stress response in rice: Genetics, molecular biology
and comparative genomics. Funct Integr Genomics 6:263 – 284.
Sairam RK, Rao KV, Srivastava GC. 2002. Differential response of wheat genotypes to long term salinity
stress in relation to oxidative stress, antioxidant activity and osmolytes concentration. Plant Sci
163:1037 – 1046.
Response of rice genotypes to NaCl stress 591

Dow
nloa
ded
By:
[IN
FLIB
NE
T, In
dia
orde
r 200
5] A
t: 10
:50
31 A
ugus
t 200
7
Sairam RK, Tyagi A. 2004. Physiology and molecular biology of salinity stress tolerance in plants. Curr Sci
86:407 – 421.
Sasaki T, Matsumoto T, Antonio BA, Nagamura Y. 2005. From mapping to sequencing, post-sequencing and
beyond. Plant Cell Physiol 46:3 – 13.
Shimamoto K, Kyozuka J. 2002. Rice as a model for comparative genomics of plants. Annu Rev Plant Biol 53:399–
419.
Sivakumar P, Sharmila P, Vikas J, Pardhasardhi P. 2002. Sugars have potential to curtail oxygenase activity of
Rubisco. Biochem Biophys Res Commun 298:247 – 250.
Sultana N, Ikeda T, Itoh R. 1999. Effect of NaCl salinity on photosynthesis and dry matter accumulation in
developing rice grains. Environ Exp Bot 42:211 – 220.
Vaidyanathan H, Sivakumar P, Chakrabarty R, Thomas G. 2003. Scavenging of reactive oxygen species in NaCl
stressed rice (Oryza sativa L.) – differential response in salt tolerant and sensitive cultivars. Plant Sci
165:1411 – 1418.
Wu J, Maehara T, Shimokawa T, Yamamoto S, Harada C, Takazaki Y, Ono N, Mukai Y, Koike K, Yazaki J, Fujii F,
Shomura A, Ando T, Kono I, Waki K, Yamamoto K, Yano M, Matsumoto T, Sasaki T. 2002. A comprehensive
rice transcript map containing 6591 expressed sequence tag sites. Plant Cell 14:525 – 535.
Yeo AR, Flowers TJ. 1983. Varietal differences in the toxicity of sodium ions in rice leaves. Physiol Plant
59:189 – 195.
Zhu JK. 2001. Over expression of a delta-pyrolline-5carboxylate synthetase gene and analysis of tolerance to water
and salt stress in transgenic rice. Trends Plant Sci 6:66 – 72.
592 V. Kumar et al.




















