
Antioxidant enzyme activities and protein profiling under salt stress in indica rice genotypes...
17
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Shitole, Mahadeo G.] On: 18 August 2009 Access details: Access Details: [subscription number 913989668] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Archives of Agronomy and Soil Science Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713453776 Antioxidant enzyme activities and protein profiling under salt stress in indica rice genotypes differing in salt tolerance Vinay Kumar a ; Varsha Shriram a ; T. D. Nikam a ; Narendra Jawali b ; Mahadeo G. Shitole a a Department of Botany, University of Pune, Pune b Molecular Biology Division, Bhabha Atomic Research Centre, Mumbai, India Online Publication Date: 01 August 2009 To cite this Article Kumar, Vinay, Shriram, Varsha, Nikam, T. D., Jawali, Narendra and Shitole, Mahadeo G.(2009)'Antioxidant enzyme activities and protein profiling under salt stress in indica rice genotypes differing in salt tolerance',Archives of Agronomy and Soil Science,55:4,379 — 394 To link to this Article: DOI: 10.1080/03650340802595543 URL: http://dx.doi.org/10.1080/03650340802595543 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Antioxidant enzyme activities and protein profiling under salt stress in indica rice genotypes...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Shitole, Mahadeo G.]On: 18 August 2009Access details: Access Details: [subscription number 913989668]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Archives of Agronomy and Soil SciencePublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713453776
Antioxidant enzyme activities and protein profiling under salt stress in indicarice genotypes differing in salt toleranceVinay Kumar a; Varsha Shriram a; T. D. Nikam a; Narendra Jawali b; Mahadeo G. Shitole a
a Department of Botany, University of Pune, Pune b Molecular Biology Division, Bhabha Atomic ResearchCentre, Mumbai, India
Online Publication Date: 01 August 2009
To cite this Article Kumar, Vinay, Shriram, Varsha, Nikam, T. D., Jawali, Narendra and Shitole, Mahadeo G.(2009)'Antioxidant enzymeactivities and protein profiling under salt stress in indica rice genotypes differing in salt tolerance',Archives of Agronomy and SoilScience,55:4,379 — 394
To link to this Article: DOI: 10.1080/03650340802595543
URL: http://dx.doi.org/10.1080/03650340802595543
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Antioxidant enzyme activities and protein profiling under salt stress in indica
rice genotypes differing in salt tolerance
Vinay Kumara, Varsha Shrirama{, T.D. Nikama, Narendra Jawalib andMahadeo G. Shitolea*
aDepartment of Botany, University of Pune, Pune; bMolecular Biology Division, Bhabha AtomicResearch Centre, Mumbai, India
(Received 15 May 2008; final version received 3 November 2008)
The effect of NaCl stress (0–300 mM) was investigated in terms of antioxidant enzymesactivity and their isozymatic pattern and SDS-PAGE proteins banding pattern in threerice cultivars (cvs), Panvel-3 (tolerant), Kalarata (moderately tolerant) and Karjat-3(sensitive). Interestingly, superoxide dismutase activity in roots and glutathionereductase activity in both shoots and roots were decreased significantly under highsalinity levels in Karjat-3 and Kalarata, while, a sharp increase was observed inPanvel-3. Catalase and peroxidase activities were increased with salt stress of all cvs,with highest magnitude in Panvel-3. There was an induction of two new POX isoformsin Panvel-3 and Kalarata in stressed plants. Four SOD isoforms were observed in all thegenotypes, irrespective of non/saline conditions. Total 33 proteins bands ranging from17–154.5 kDa were either expressed de novo or up/down-accumulated due to NaClstress. In Panvel-3, three new bands, one of 32 kDa in shoots while two of 37 and116.7 kDa appeared in roots under salt stress. A new band of 37 kDa was also observedin roots of Kalarata under salinity stress. Thus, salinity tolerance nature of Panvel-3may be correlated with higher enzyme activities and expression of some newpolypeptides under salt stress.
Keywords: isozymes; superoxide dismutase; catalase; peroxidase; glutathione reductase;indica rice; salt stress; salt stress-proteins
Introduction
High salinity of soil and irrigation water is a major abiotic stress, which severely affectscrop production worldwide (Kumar et al. 2008). Several rice growing countries, both inthe tropics and in temperate regions, are facing high soil salinity as a major problem witharid, semi-arid, and coastal areas of the tropics affected with a greater extent as salineconditions are more common in rice-lands of these areas (Lee et al. 2003). In India, morethan 8.6 m ha of lands are salt affected, which constitutes a major part of problem soils inIndia (Sahi et al. 2006). Rice is the most important crop of India being the staple food ofover 65% of the population (International Rice Research Institute [IRRI] 2006). Despitethe advances in the increase of plant productivity and resistance to a number of pests anddiseases, improvement in salt tolerance of crop plants remain elusive and this is due to thefact that salinity is a complex trait which affects almost every aspect of the physiology and
*Corresponding author. Email: [email protected]{Present address: Department of Botany, Annasaheb Magar College, Hadapsar, Pune, India.
Archives of Agronomy and Soil Science
Vol. 55, No. 4, August 2009, 379–394
ISSN 0365-0340 print/ISSN 1476-3567 online
� 2009 Taylor & Francis
DOI: 10.1080/03650340802595543
http://www.informaworld.com
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

biochemistry of plants (Cuartero et al. 2006) at both whole plant and cellular levelsthrough osmotic and ionic stresses (Murphy and Durako 2003).
An unfortunate consequence of salinity stress in plants is the excessive generation ofreactive oxygen species (ROS)/intermediates such as super oxide radical (O2
7), hydrogenperoxide (H2O2) and hydroxyl radical (OH7) (Mittler 2002; Sairam et al. 2005). ROS canbe generated in the chloroplast by direct transfer of excitation energy from chlorophyll toproduce singlet oxygen or by univalent oxygen reduction at photo system I in the Mehlerreaction (Allen 1995). Vaidyanathan et al. (2003) suggested that excess production of ROSduring salinity stress results from impaired electron transport processes in chloroplast andmitochondria as well as from pathways such as photorespiration. The overproduction ofROS damage membranes and other essential macromolecules such as photosyntheticpigments, proteins, DNA and lipids (Gomez et al. 2004; Demiral and Turkan 2005) andplay an important role in inhibiting photosynthesis (Price and Hendry 1991).
However, in response, plants have developed various combating mechanisms to copewith the deleterious effects of salinity stress including both enzymatic and non-enzymaticantioxidant mechanisms. Several antioxidant enzymes like superoxide dismutases (SOD),catalases (CAT) and Peroxidase (POX), and Glutathion reductase (GR) (Apse andBlumwald 2002; Mittler 2002; Apel and Hirt 2004) participate in the detoxification ofROS. These antioxidant systems can be divided into two categories: One that reacts withROS and keeps them at low levels (POX, SOD and CAT), and one that regenerates theoxidized antioxidants (ascorbate peroxidase [APX] and glutathione reductase) (Jebaraet al. 2005). There is enough evidence that alleviation of oxidative damage and increasedsalinity tolerance is often correlated with an efficient antioxidant defence system in plants(Hasegawa et al. 2000; Bor et al. 2003; Sairam et al. 2005).
In addition, salinity affects the metabolism of plants adversely and causes importantmodifications in gene expression in plants. Such modifications may lead to accumulationor depletion of certain metabolites resulting in an imbalance in the levels of cellularproteins, which could increase, decrease, appear or disappear after salt treatment (Kong-Ngern et al. 2005). Several proteins may synthesized and accumulate in plant tissues understress conditions, referred to as stress-proteins and information on such proteins isimportant for various reasons such as assessment of these proteins may help in revealingpossible number of genes that are altered in response to the salt stress and identification ofsuch protein markers for stress tolerance could provide an easy tool to plant breeders foranalyzing tolerance traits, which in turn may help in producing salt tolerant genotypes(Pareek et al. 1999). Over the past few years, much attention has been concentrated onresolving the identity of salt stress proteins, in order to identify and understand the role ofproteins in rice salt tolerance. However, at present their functions are still unclear.Studying differential responses of genotypes in terms of antioxidant defence system andproteins alteration, with contrasting stress tolerance will help to dissect the underlying saltstress tolerance mechanisms.
This work investigates the effects of varying salt stress on activities of variousantioxidant enzymes and their isozymatic analysis, and differential electrophoretic proteinprofiling in indica rice genotypes, contrasting in terms of salt stress tolerance, under non-saline and saline conditions.
Materials and methods
Plant material and salt treatments
Indica rice (Oryza sativa L.) cvs, namely, Karjat-3 (salt sensitive), Kalarata (moderatelysalt tolerant) and Panvel-3 (salt tolerant) (Kumar et al. 2007) were selected for this study.
380 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

Certified seeds of Karjat-3 were obtained from The Regional Rice Research Station,Karjat, Maharashtra, India; while seeds of Kalarata and Panvel-3 were obtained from TheSaline Land Research Station, Panvel, Maharashtra, India.
Twenty seeds of each cv were sown in the insulated (with no outlet) plastic pots(15 6 15 6 12 cm) to ensure the constant level of desired salinity stress. The pots werefilled with garden soil mixed with vermiculite and sand (1:1:1) after passing through a10-mm mesh screen, at the Botanical Garden, Department of Botany, University ofPune, Pune, India. Plants were irrigated to meet evapotranspiration demand, and anequal amount of water was applied in all the pots. Plants were irrigated with NaCl-freewater for first 14 days after sowing (DAS). Six salinity treatments were then appliedusing irrigation waters containing 50, 100, 150, 200, 250 and 300 mM NaCl, and salinitylevels were maintained by applying the salt solutions on alternate days for seven daysfrom the 14th to the 21st day after sowing (DAS). The enzyme activities were studiedunder all the salinity levels while electrophoretic protein profiling and isozymaticanalyses was carried out at 100 mM and 200 mM NaCl stress besides controlled plants.All the observations were recorded on the 21st DAS and all the experiments were carriedout in triplicate.
Extraction and antioxidant enzyme assay
SOD activity was estimated by recording the decrease in absorbance of formazanproduced by superoxide-nitro blue tetrazolium (NBT) complex by the enzyme as given byDhindsa et al. (1981). Fresh 500 mg shoot and root samples were homogenized separatelyin 0.1 M phosphate buffer (pH 7.5) with the help of pre-chilled mortar and pestle. Threeml of the reaction mixture containing 0.1 ml of 1.5 M sodium carbonate, 0.2 ml of200 mM methionine, 0.1 ml of 3 mM EDTA, 0.1 ml of 2.25 mM NBT, 1.5 ml of 100 mMpotassium phosphate buffer, 0.95 ml distilled water and 0.05 ml of enzyme were taken inthe test tubes in duplicate from each enzyme sample. Two tubes without enzyme extractwere taken as control. Reaction was started by adding 0.1 ml riboflavin (60 mM) andplacing the tubes below a light source of two 15 W florescent lamps for 15 min. Thereaction was stopped by switching off the light and covering the tubes with black cloth.The tube without enzyme developed maximal colour while the tube without colour servedas blank. Absorbance was recorded at 560 nm and one unit of enzyme activity was takenas the quantity of enzyme which reduced the absorbance reading of samples by 50% incomparison with tubes lacking enzymes.
CAT activity was assayed according to Luck (1974). Fresh shoot and root samples(250 mg) were homogenized in 3 ml of 0.1M phosphate buffer (pH 7) in pre-chilled mortarand pestle. The extract was centrifuged at 15,000 g for 20 min at 48C and the supernatantwas used as enzyme source. The assay mixture contained 0.5 ml of 0.2 M phosphate buffer(pH 7), 0.3 ml of H2O2 and 0.1 ml enzyme and final volume was made 3 ml by addingdistilled water. The assay was carried out a 27 + 28C. The reaction was started by addingenzyme and change in absorbance was measured at 240 nm.
POX activity was assayed by following Putter (1974). Fresh shoot and root samples(250 mg) were homogenized with pre-chilled mortar and pestle, in 5 ml of 0.1M phosphatebuffer (pH 7.0). The extract was centrifuged in cold centrifuge at 15,000 g for 20 min at48C and the supernatant was used as enzyme source. The assay mixture of 3 ml contained1.8 ml of 0.1 M phosphate buffer (pH 7.0), 1 ml freshly prepared 10 mM guaicol, 0.1 mlenzyme extract and 0.1 ml of 12.3 mM H2O2. Initial absorbance was read at 436 nm andthen increase in absorbance was noted at the intervals of 30 sec.
Archives of Agronomy and Soil Science 381
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

GR activity was assayed by following Smith et al. (1988) by recording the increase in thepresence of oxidized glutathione and DTNB (5,50-dithiobis-2-nitrobenzoic acid). Thereaction mixture contained 1 ml of 0.2 M potassium phosphate buffer (pH 7.5) containing0.1 mM EDTA, 0.5 ml of 3 mM DTNB in 0.01 M potassium phosphate buffer (pH 7.5),0.1 ml of 2 mM NADPH, 0.1 ml enzyme extract and final volume of 2.9 ml was made upby adding distilled water. The increase in absorbance at 412 nm was recorded over a periodof 5 min.
Isozymatic analysis
SOD isozymes were separated on 8% non-denaturing polyacrylamide gels at 48C usingMonokin Electrophoresis Unit (Technosource, India). A protein concentration of 50 mgfrom shoots under non-saline and saline conditions was applied into each well and thenelectrophoresed at 30 mA for 2 h. SOD activity staining was done by following themodified method of Beauchamp and Fridovich (1971) and inhibitor studies to distinguishvarious SOD isoforms as per Sandalio et al. (1987). The gel was soaked in 50 ml of100 mM potassium phosphate buffer, pH 7.5 containing 1.23 mM NBT, 0.02 mMriboflavin, 28 mM TEMED for 20 min in the dark and then illuminated under light sourceto initiate the photochemical reaction.
CAT isoenzymes were separated on 10% non-denaturating acrylamide gel run at30 mA for 24 h at 48C and were visualized by following the method of Woodbury et al.(1971). Gels were rinsed in double distilled water (DDW) followed by incubation in 0.01%H2O2, for 5 min, washed twice with DDW and then stained in a solution of 0.5%potassium ferricyanide and 0.5% ferric chloride. After development of bands, reaction wasterminated with 1% HCl.
POX isozymes separation and staining was done as described by Anderson et al.(1995). POX isozymes were separated on 8% non-denaturing polyacrylamide gel and theelectrophoresis was done at 30 mA for 150 min at 48C. Isoforms of POX were visualizedby staining the gel in 50 ml solution containing 100 mM potassium phosphate buffer, pH6.4, 20 mM guaiacol and 5.55 mM H2O2.
Separation of proteins by SDS-PAGE
Proteins of shoots and roots were separated by sodium dodecyl sulphate-poly acrylamidegel electrophoresis (SDS-PAGE) on the basis of their molecular mass by following theLaemmli (1970) method. Fresh plant tissues (200 mg) were ground to powder in liquidnitrogen, reconstituted in 1 ml chilled phosphate extraction buffer (pH 7) with the help ofmortar and pestle and were centrifuged at 10,000 g for 15 min at 48C and extracts wereimmediately stored at 7208C. Estimation of proteins was done prior to loading byfollowing the method of Bradford (1976). An equal amount of protein samples were mixedwith 20 ml of sample buffer (pH 6.8) in eppendorf tubes and then the samples were kept inhot water (1008C) for 1 min to denature the samples. An aliquot of 20 mg proteins sampleswere loaded onto the wells of the gel that consist of two gel layers, upper being the 5%stacking gel and lower 12% (w/v) resolving gel containing 0.1% SDS and 10% ammoniumper sulphate in the vertical electrophoresis apparatus (Technosource, India). Markerproteins (Bangalore Genei, India) were loaded in separate well. The gels were stained with0.2% (w/v) silver nitrate solution and the gels were stored in 50% (v/v) methanol. Theimage analysis of gels was done using the gel documentation system (Alpha Innotech
382 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

Corporation, USA), and molecular weight of polypeptide bands were calculated by thesoftware package Alpha Ease FC 4.0.
Statistical analysis
The experiments were repeated three times for checking the reproducibility of resultsbefore conducting the statistical analyses. The data were subjected to analysis of variance(ANOVA) to detect significant difference between means. Means differing significantlywere compared using Duncan’s Multiple Range test (DMRT) at p � 0.05. All thestatistical analyses were done by using MSTAT-C statistical software package.
Results and discussion
Effect of NaCl stress on antioxidant enzyme activities and isozymatic patterns in ricecultivars
Abiotic stresses including salinity, affects plants via various mechanisms, causing ionicimbalance, osmotic stress, in addition to inducing oxidative stress. The aim of the presentstudy was to study the comparative enzymatic ROS scavenging mechanisms operational inrice cvs with differential salt stress tolerance. This may be helpful to shed the light on the keycomponents employed by salt-tolerant cvs to combat salinity induced oxidative stress. This inturn may be helpful in the identification of candidate genes for salt-stress tolerance ofimportant crop plants. Thus, the present work reports activities and in-gel assay of importantantioxidant enzymes separately from shoots and roots in the rice cvs Panvel-3, Kalarata andKarjat-3 to NaCl stress.
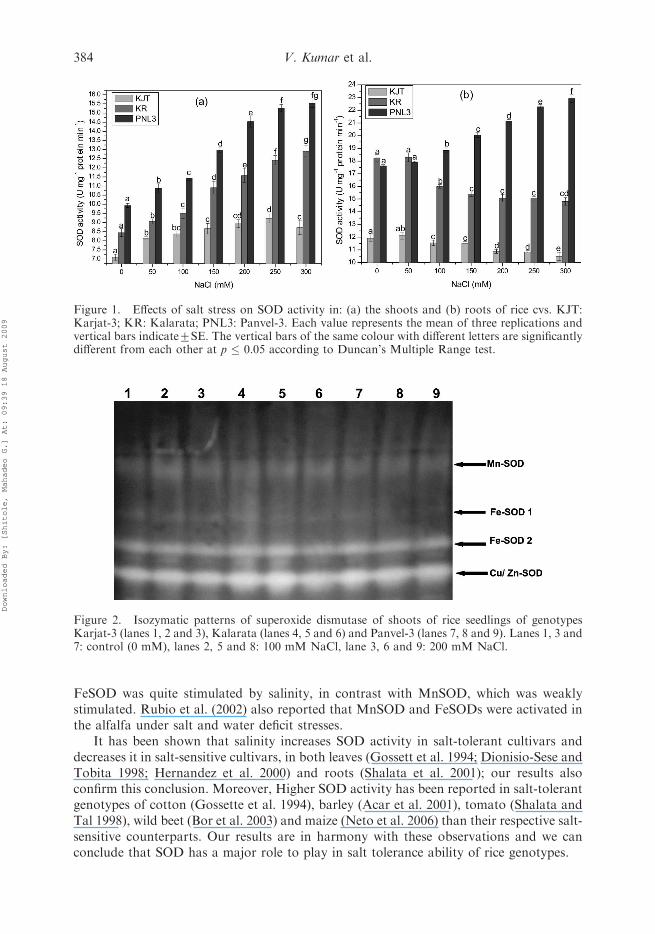
Interestingly, SOD activity in control as well as stress seedlings of cv Panvel-3 wassignificantly higher than those of cv Kalarata and Karjat-3. SOD activity was increasedconsiderably with increasing salinity levels in the shoots of all the cvs, however, with theconsiderable variation in the rate of increase in activity amongst the cvs. An increase of156% was observed in SOD activity in Panvel-3 cv as compared to non-saline plants, and152% increase was recorded in Kalarata cv at this salinity level. On the contrary to theseresults, the rate of increase in SOD activity was much lesser in Karjat-3 and 122% increasewas seen in shoots at 300 mM NaCl stress (Figure 1a).
SOD is one of the ubiquitous enzymes, which play a key role in cellular defensemechanisms against ROS. Its activity modulates the relative amounts of O2
�7and H2O2,the two Haber-Weiss reaction substrates. It also decreases the risk of OH
�
radicalformation, which is highly reactive and may cause severe damage to membranes, proteinsand DNA (Bowler et al. 1992; Neto et al. 2006). The contrasting results were observed forSOD activity in roots of these rice cvs. SOD activity was either remained unchanged ordecreased sharply in roots of Karjat-3 and Kalarata, respectively. In the roots of Panvel-3,on the other hand, SOD activity was increased with increased salinity stress levels, and athighest level of stress, the SOD activity was increased by around 130% as compared withthe controlled plants (Figure 1b), suggesting that the salt-tolerant genotype has a betterO2
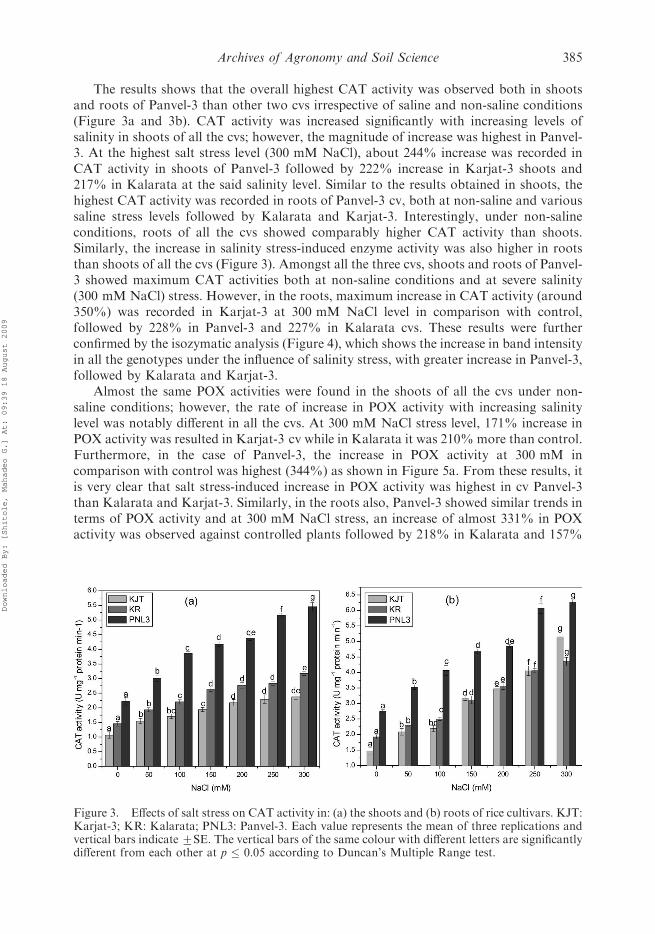
�7 radical scavenging ability. Similar results have been reported by Neto et al. (2006),and they observed that SOD actvity was reduced more pronouncely in salt-sensitive maizecv than the tolerant one. The activity results were well supported by the SOD isozymaticpatterns of shoots (Figure 2). The Fe-SOD and Cu/Zn SOD bands were more intensed intolerant genotypes than sensitive one under salinity stress, which suggested that specificactivity of Fe-SOD and Cu/Zn SOD seems to contribute to a noticed increase in overallSOD activities. Similar results are reported by Jebara et al. (2005), and concluded that the
Archives of Agronomy and Soil Science 383
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009
FeSOD was quite stimulated by salinity, in contrast with MnSOD, which was weaklystimulated. Rubio et al. (2002) also reported that MnSOD and FeSODs were activated inthe alfalfa under salt and water deficit stresses.
It has been shown that salinity increases SOD activity in salt-tolerant cultivars anddecreases it in salt-sensitive cultivars, in both leaves (Gossett et al. 1994; Dionisio-Sese andTobita 1998; Hernandez et al. 2000) and roots (Shalata et al. 2001); our results alsoconfirm this conclusion. Moreover, Higher SOD activity has been reported in salt-tolerantgenotypes of cotton (Gossette et al. 1994), barley (Acar et al. 2001), tomato (Shalata andTal 1998), wild beet (Bor et al. 2003) and maize (Neto et al. 2006) than their respective salt-sensitive counterparts. Our results are in harmony with these observations and we canconclude that SOD has a major role to play in salt tolerance ability of rice genotypes.
Figure 2. Isozymatic patterns of superoxide dismutase of shoots of rice seedlings of genotypesKarjat-3 (lanes 1, 2 and 3), Kalarata (lanes 4, 5 and 6) and Panvel-3 (lanes 7, 8 and 9). Lanes 1, 3 and7: control (0 mM), lanes 2, 5 and 8: 100 mM NaCl, lane 3, 6 and 9: 200 mM NaCl.
Figure 1. Effects of salt stress on SOD activity in: (a) the shoots and (b) roots of rice cvs. KJT:Karjat-3; KR: Kalarata; PNL3: Panvel-3. Each value represents the mean of three replications andvertical bars indicate+SE. The vertical bars of the same colour with different letters are significantlydifferent from each other at p � 0.05 according to Duncan’s Multiple Range test.
384 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009
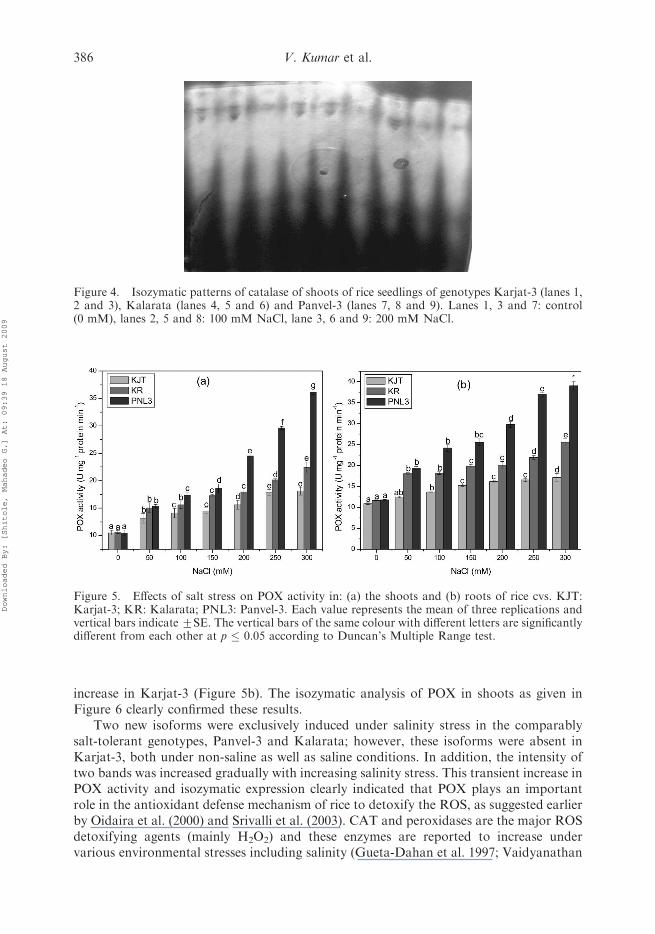
The results shows that the overall highest CAT activity was observed both in shootsand roots of Panvel-3 than other two cvs irrespective of saline and non-saline conditions(Figure 3a and 3b). CAT activity was increased significantly with increasing levels ofsalinity in shoots of all the cvs; however, the magnitude of increase was highest in Panvel-3. At the highest salt stress level (300 mM NaCl), about 244% increase was recorded inCAT activity in shoots of Panvel-3 followed by 222% increase in Karjat-3 shoots and217% in Kalarata at the said salinity level. Similar to the results obtained in shoots, thehighest CAT activity was recorded in roots of Panvel-3 cv, both at non-saline and varioussaline stress levels followed by Kalarata and Karjat-3. Interestingly, under non-salineconditions, roots of all the cvs showed comparably higher CAT activity than shoots.Similarly, the increase in salinity stress-induced enzyme activity was also higher in rootsthan shoots of all the cvs (Figure 3). Amongst all the three cvs, shoots and roots of Panvel-3 showed maximum CAT activities both at non-saline conditions and at severe salinity(300 mM NaCl) stress. However, in the roots, maximum increase in CAT activity (around350%) was recorded in Karjat-3 at 300 mM NaCl level in comparison with control,followed by 228% in Panvel-3 and 227% in Kalarata cvs. These results were furtherconfirmed by the isozymatic analysis (Figure 4), which shows the increase in band intensityin all the genotypes under the influence of salinity stress, with greater increase in Panvel-3,followed by Kalarata and Karjat-3.
Almost the same POX activities were found in the shoots of all the cvs under non-saline conditions; however, the rate of increase in POX activity with increasing salinitylevel was notably different in all the cvs. At 300 mM NaCl stress level, 171% increase inPOX activity was resulted in Karjat-3 cv while in Kalarata it was 210% more than control.Furthermore, in the case of Panvel-3, the increase in POX activity at 300 mM incomparison with control was highest (344%) as shown in Figure 5a. From these results, itis very clear that salt stress-induced increase in POX activity was highest in cv Panvel-3than Kalarata and Karjat-3. Similarly, in the roots also, Panvel-3 showed similar trends interms of POX activity and at 300 mM NaCl stress, an increase of almost 331% in POXactivity was observed against controlled plants followed by 218% in Kalarata and 157%
Figure 3. Effects of salt stress on CAT activity in: (a) the shoots and (b) roots of rice cultivars. KJT:Karjat-3; KR: Kalarata; PNL3: Panvel-3. Each value represents the mean of three replications andvertical bars indicate+SE. The vertical bars of the same colour with different letters are significantlydifferent from each other at p � 0.05 according to Duncan’s Multiple Range test.
Archives of Agronomy and Soil Science 385
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009
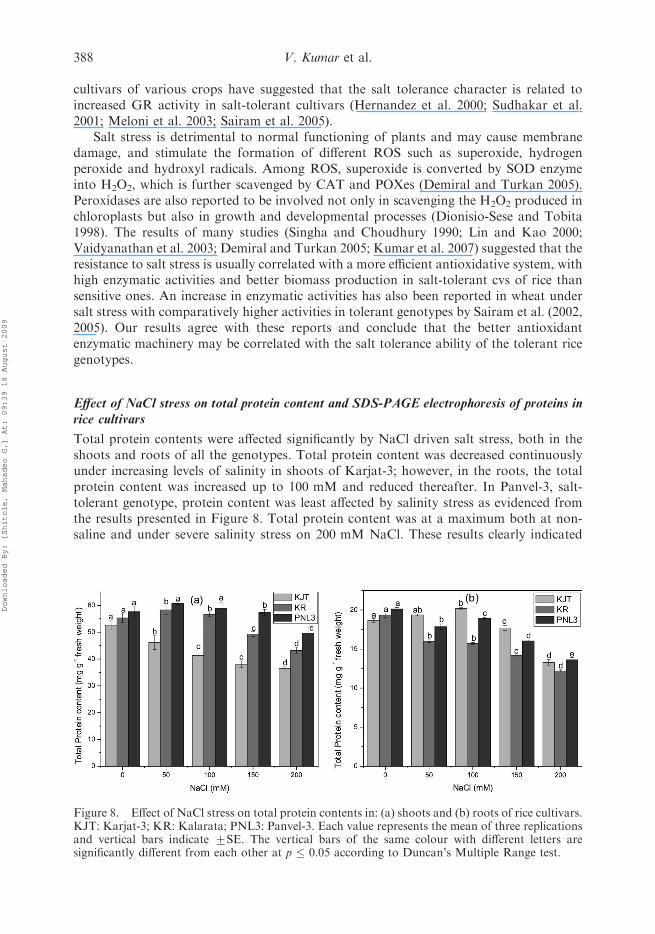
increase in Karjat-3 (Figure 5b). The isozymatic analysis of POX in shoots as given inFigure 6 clearly confirmed these results.
Two new isoforms were exclusively induced under salinity stress in the comparablysalt-tolerant genotypes, Panvel-3 and Kalarata; however, these isoforms were absent inKarjat-3, both under non-saline as well as saline conditions. In addition, the intensity oftwo bands was increased gradually with increasing salinity stress. This transient increase inPOX activity and isozymatic expression clearly indicated that POX plays an importantrole in the antioxidant defense mechanism of rice to detoxify the ROS, as suggested earlierby Oidaira et al. (2000) and Srivalli et al. (2003). CAT and peroxidases are the major ROSdetoxifying agents (mainly H2O2) and these enzymes are reported to increase undervarious environmental stresses including salinity (Gueta-Dahan et al. 1997; Vaidyanathan
Figure 4. Isozymatic patterns of catalase of shoots of rice seedlings of genotypes Karjat-3 (lanes 1,2 and 3), Kalarata (lanes 4, 5 and 6) and Panvel-3 (lanes 7, 8 and 9). Lanes 1, 3 and 7: control(0 mM), lanes 2, 5 and 8: 100 mM NaCl, lane 3, 6 and 9: 200 mM NaCl.
Figure 5. Effects of salt stress on POX activity in: (a) the shoots and (b) roots of rice cvs. KJT:Karjat-3; KR: Kalarata; PNL3: Panvel-3. Each value represents the mean of three replications andvertical bars indicate+SE. The vertical bars of the same colour with different letters are significantlydifferent from each other at p � 0.05 according to Duncan’s Multiple Range test.
386 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

et al. 2003). The enhanced activities of these enzymes have been correlated with the role inimparting tolerance of plants against environmental stresses (Sairam et al. 2002). In thisbackground, higher CAT and POX activities in Panvel-3 under increasing salt stresssignifies its relative tolerance to salt stress, while other two cvs were inferior on this count.
High salinity stress significantly reduced GR activity in salt-sensitive cv (Karjat-3) andsalt moderately-tolerant cv Kalarata. On the other hand, contrasting observations werenoticed in Panvel-3, as activity was increased markedly due to salt stress both in shootsand roots (Figure 7a and 7b, respectively). In the present investigation, the reduction inGR activity in comparably salt-sensitive cvs suggests a decreased GSH turnover rate.Several authors investigating antioxidant responses of salt-tolerant and salt-sensitive
Figure 6. Isozymatic patterns of peroxidase of shoots of rice seedlings of genotypes Karjat-3 (lanes1, 2 and 3), Kalarata (lanes 4, 5 and 6) and Panvel-3 (lanes 7, 8 and 9). Lanes 1, 3 and 7: control(0 mM), lanes 2, 5 and 8: 100 mM NaCl, lane 3, 6 and 9: 200 mM NaCl.
Figure 7. Effects of salt stress on GR activity in: (a) the shoots and (b) roots of rice cvs. KJT:Karjat-3; KR: Kalarata; PNL3: Panvel-3. Each value represents the mean of three replications andvertical bars indicate+SE. The vertical bars of the same colour with different letters are significantlydifferent from each other at p � 0.05 according to Duncan’s Multiple Range test.
Archives of Agronomy and Soil Science 387
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009
cultivars of various crops have suggested that the salt tolerance character is related toincreased GR activity in salt-tolerant cultivars (Hernandez et al. 2000; Sudhakar et al.2001; Meloni et al. 2003; Sairam et al. 2005).
Salt stress is detrimental to normal functioning of plants and may cause membranedamage, and stimulate the formation of different ROS such as superoxide, hydrogenperoxide and hydroxyl radicals. Among ROS, superoxide is converted by SOD enzymeinto H2O2, which is further scavenged by CAT and POXes (Demiral and Turkan 2005).Peroxidases are also reported to be involved not only in scavenging the H2O2 produced inchloroplasts but also in growth and developmental processes (Dionisio-Sese and Tobita1998). The results of many studies (Singha and Choudhury 1990; Lin and Kao 2000;Vaidyanathan et al. 2003; Demiral and Turkan 2005; Kumar et al. 2007) suggested that theresistance to salt stress is usually correlated with a more efficient antioxidative system, withhigh enzymatic activities and better biomass production in salt-tolerant cvs of rice thansensitive ones. An increase in enzymatic activities has also been reported in wheat undersalt stress with comparatively higher activities in tolerant genotypes by Sairam et al. (2002,2005). Our results agree with these reports and conclude that the better antioxidantenzymatic machinery may be correlated with the salt tolerance ability of the tolerant ricegenotypes.
Effect of NaCl stress on total protein content and SDS-PAGE electrophoresis of proteins inrice cultivars
Total protein contents were affected significantly by NaCl driven salt stress, both in theshoots and roots of all the genotypes. Total protein content was decreased continuouslyunder increasing levels of salinity in shoots of Karjat-3; however, in the roots, the totalprotein content was increased up to 100 mM and reduced thereafter. In Panvel-3, salt-tolerant genotype, protein content was least affected by salinity stress as evidenced fromthe results presented in Figure 8. Total protein content was at a maximum both at non-saline and under severe salinity stress on 200 mM NaCl. These results clearly indicated
Figure 8. Effect of NaCl stress on total protein contents in: (a) shoots and (b) roots of rice cultivars.KJT: Karjat-3; KR: Kalarata; PNL3: Panvel-3. Each value represents the mean of three replicationsand vertical bars indicate +SE. The vertical bars of the same colour with different letters aresignificantly different from each other at p � 0.05 according to Duncan’s Multiple Range test.
388 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009
that Panvel-3 could withstand salt stress in a better way than the other two genotypes interms of total proteins content.
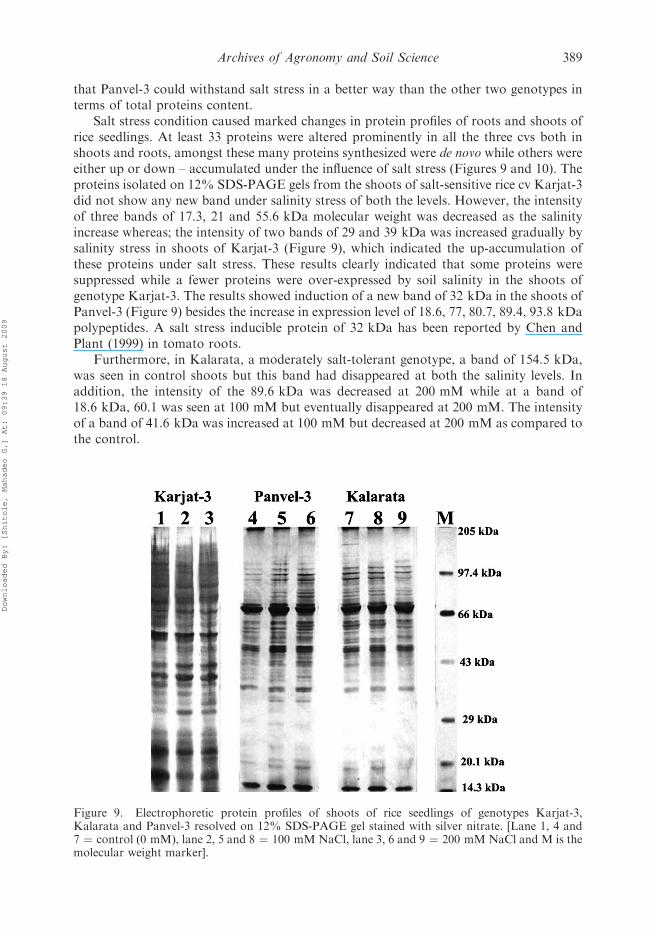
Salt stress condition caused marked changes in protein profiles of roots and shoots ofrice seedlings. At least 33 proteins were altered prominently in all the three cvs both inshoots and roots, amongst these many proteins synthesized were de novo while others wereeither up or down – accumulated under the influence of salt stress (Figures 9 and 10). Theproteins isolated on 12% SDS-PAGE gels from the shoots of salt-sensitive rice cv Karjat-3did not show any new band under salinity stress of both the levels. However, the intensityof three bands of 17.3, 21 and 55.6 kDa molecular weight was decreased as the salinityincrease whereas; the intensity of two bands of 29 and 39 kDa was increased gradually bysalinity stress in shoots of Karjat-3 (Figure 9), which indicated the up-accumulation ofthese proteins under salt stress. These results clearly indicated that some proteins weresuppressed while a fewer proteins were over-expressed by soil salinity in the shoots ofgenotype Karjat-3. The results showed induction of a new band of 32 kDa in the shoots ofPanvel-3 (Figure 9) besides the increase in expression level of 18.6, 77, 80.7, 89.4, 93.8 kDapolypeptides. A salt stress inducible protein of 32 kDa has been reported by Chen andPlant (1999) in tomato roots.
Furthermore, in Kalarata, a moderately salt-tolerant genotype, a band of 154.5 kDa,was seen in control shoots but this band had disappeared at both the salinity levels. Inaddition, the intensity of the 89.6 kDa was decreased at 200 mM while at a band of18.6 kDa, 60.1 was seen at 100 mM but eventually disappeared at 200 mM. The intensityof a band of 41.6 kDa was increased at 100 mM but decreased at 200 mM as compared tothe control.
Figure 9. Electrophoretic protein profiles of shoots of rice seedlings of genotypes Karjat-3,Kalarata and Panvel-3 resolved on 12% SDS-PAGE gel stained with silver nitrate. [Lane 1, 4 and7 ¼ control (0 mM), lane 2, 5 and 8 ¼ 100 mMNaCl, lane 3, 6 and 9 ¼ 200 mM NaCl and M is themolecular weight marker].
Archives of Agronomy and Soil Science 389
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009
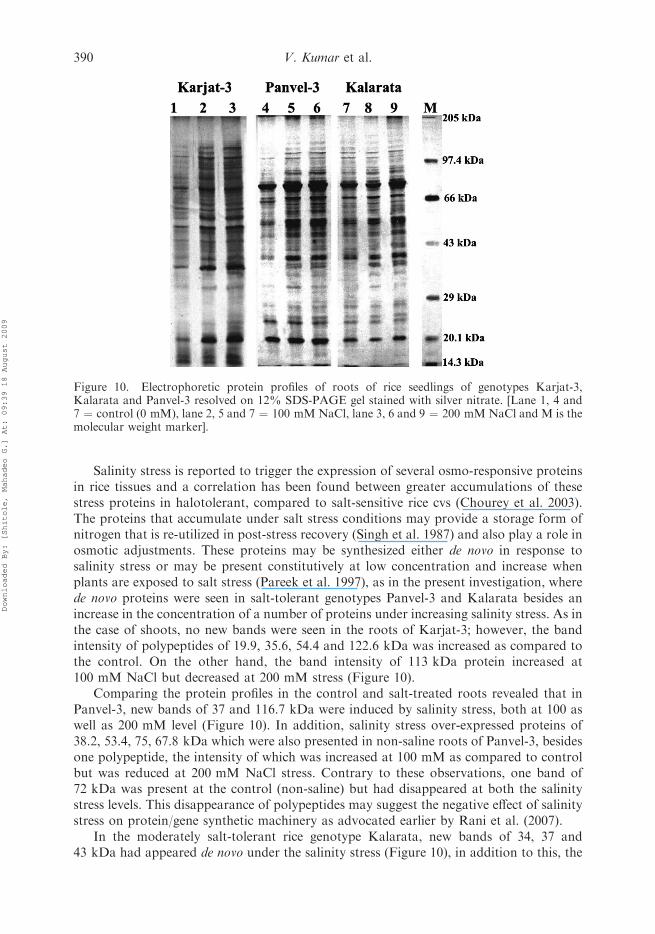
Salinity stress is reported to trigger the expression of several osmo-responsive proteinsin rice tissues and a correlation has been found between greater accumulations of thesestress proteins in halotolerant, compared to salt-sensitive rice cvs (Chourey et al. 2003).The proteins that accumulate under salt stress conditions may provide a storage form ofnitrogen that is re-utilized in post-stress recovery (Singh et al. 1987) and also play a role inosmotic adjustments. These proteins may be synthesized either de novo in response tosalinity stress or may be present constitutively at low concentration and increase whenplants are exposed to salt stress (Pareek et al. 1997), as in the present investigation, wherede novo proteins were seen in salt-tolerant genotypes Panvel-3 and Kalarata besides anincrease in the concentration of a number of proteins under increasing salinity stress. As inthe case of shoots, no new bands were seen in the roots of Karjat-3; however, the bandintensity of polypeptides of 19.9, 35.6, 54.4 and 122.6 kDa was increased as compared tothe control. On the other hand, the band intensity of 113 kDa protein increased at100 mM NaCl but decreased at 200 mM stress (Figure 10).
Comparing the protein profiles in the control and salt-treated roots revealed that inPanvel-3, new bands of 37 and 116.7 kDa were induced by salinity stress, both at 100 aswell as 200 mM level (Figure 10). In addition, salinity stress over-expressed proteins of38.2, 53.4, 75, 67.8 kDa which were also presented in non-saline roots of Panvel-3, besidesone polypeptide, the intensity of which was increased at 100 mM as compared to controlbut was reduced at 200 mM NaCl stress. Contrary to these observations, one band of72 kDa was present at the control (non-saline) but had disappeared at both the salinitystress levels. This disappearance of polypeptides may suggest the negative effect of salinitystress on protein/gene synthetic machinery as advocated earlier by Rani et al. (2007).
In the moderately salt-tolerant rice genotype Kalarata, new bands of 34, 37 and43 kDa had appeared de novo under the salinity stress (Figure 10), in addition to this, the
Figure 10. Electrophoretic protein profiles of roots of rice seedlings of genotypes Karjat-3,Kalarata and Panvel-3 resolved on 12% SDS-PAGE gel stained with silver nitrate. [Lane 1, 4 and7 ¼ control (0 mM), lane 2, 5 and 7 ¼ 100 mMNaCl, lane 3, 6 and 9 ¼ 200 mM NaCl and M is themolecular weight marker].
390 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

expression levels of two bands of 54.5 and 75 kDa had increased at 100 and 200 mMNaCl, besides two proteins, whose expression levels were increased at 100 mM NaCl butdecreased at 200 as compared to the controlled plants. A number of workers have reportedalterations in protein profiles in various crops under salinity stress. Wimmer et al. (2003)observed induction of 25 kDa polypeptide while an increase in level of 33 kDa protein wasseen in wheat under combined stress of salt and boron. Sousa et al. (2004) reported thatcowpea seedlings subjected to NaCl stress showed increased concentration of nineproteins, decreased concentration of one and de novo synthesis of one 21.2 kDa protein.Kim et al. (2003) found an increase in the concentration or expression level of a 33.8 kDaprotein in rice under salinity stress and identified it as a salt inducible protein. Our resultsagree with these claims and suggest that 34 kDa protein may serve as a salt stress-inducedprotein which may be correlated with the salt stress tolerance nature of Kalarata genotype.It is clear that these stress proteins may have a role in providing plants with the ability tocross-adapt.
Conclusions
The results clearly indicated that more polypeptides were either induced de novo or theirexpression level was increased under salt stress in comparably salt-tolerant rice genotypesthan the sensitive one, which may play a protective role in combating higher salt stress.Further characterization of these proteins and the use of data for understanding themolecular basis of salinity stress tolerance will be worthwhile.
Our earlier results (Kumar et al. 2007) showed that the salt-tolerant nature of cvPanvel-3 may be attributed to its high proline accumulation and lower lipid peroxidationrate, lesser extent of reduction in chlorophyll contents along with the lowest reduction ingermination and growth rate under the influence of salinity stress, as compared to othertwo genotypes at germination level. Furthermore, Panvel-3 showed noticeably lower Na/K, Na/Ca and Na/N ratios with much lower extent than the salt-sensitive genotypeKarjat-3 (Kumar et al. 2008). These observations are well supported by the results of thepresent investigation in which the capacity of Panvel-3 to withstand salinity stress may becorrelated to its significantly higher active antioxidant enzymatic machinery andexpression of particular polypeptides under varying levels of salinity stress.
Acknowledgements
This work was supported by a grant from Board of Research in Nuclear Sciences, Department ofAtomic Energy (DAE-BRNS), Government of India (Grant No. 2004/37/28/BRNS) to M.G. Shitoleand a research fellowship to the first author. The authors also acknowledge the facilities used for thiswork at the Department of Botany, University of Pune, created under the UGC-ASIST and UGC-DRS-SAP programs.
References
Acar O, Turkan I, Ozdemir F. 2001. Superoxide dismutase and peroxidase activities in droughtsensitive and resistant barley (Hordeum vulgare L.) varieties. Acta Physiol Plant. 23:351–356.
Allen R. 1995. Dissection of oxidative stress tolerance using transgenic plants. Plant Physiol.107:1049–1054.
Anderson MD, Prasad TK, Stewart CR. 1995. Changes in isoenzymes profile of catalase, peroxidaseand glutathione reductase during acclimation to chilling in mesocotyls of maize seedlings. PlantPhysiol. 109:1247–1257.
Apel K, Hirt H. 2004. Reactive oxygen species: metabolism, oxidative stress and signal transduction.Ann Rev Plant Biol. 55:373–399.
Apse MP, Blumwald E. 2002. Engineering salt tolerance in plants. Curr Opin Biotech. 13:146–150.
Archives of Agronomy and Soil Science 391
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

Beauchamp C, Fridovich I. 1971. Superoxide dismutase. Improved assays and an assay applicable toacrylamide gels. Anal Biochem. 44:276–287.
Bor M, Ozdemir F, Turkan I. 2003. The effect of salt stress on lipid peroxidation and anti-oxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritia L. Plant Sci. 164:77–84.
Bowler C, Van Montagu M, Inze D. 1992. Superoxide dismutase and stress tolerance. Annu RevPlant Physiol Plant Mol Biol. 43:83–116.
Bradford MM. 1976. A rapid and sensitive method for the quantification of microgram quantities ofprotein utilizing the principle of protein-dye binding. Ann Biochem. 72:248–254.
Chen CCS, Plant AL. 1999. Salt-induced protein synthesis in tomato roots: the role of ABA. J ExpBot. 50:677–687.
Chourey K, Ramani S, Apte SK. 2003. Accumulation of LEA proteins in salt (NaCl) stressed youngseedlings of rice (Oryza sativa L.) cv Bura Rata and their degradation during recovery fromsalinity stress. J Plant Physiol. 160:1165–1174.
Cuartero J, Boların MC, Asıns MJ, Moreno V. 2006. Increasing salt tolerance in the tomato. J ExpBot. 57:1045–1058.
Demiral T, Turkan I. 2005. Comparative lipid peroxidation, antioxidant defence systems and prolinecontent in roots of two rice cultivars in salt tolerance. Env Exp Bot. 53:247–257.
Dhindsa RS, Plumb-Dhindsa P, Thorne TA. 1981. Leaf senescence: correlated with increased levelsof membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase.J Exp Bot. 32:93–101.
Dionisio-Sese ML, Tobita S. 1998. Antioxidant responses of rice seedlings to salinity stress. PlantSci. 135:1–9.
Gomez JM, Jimenez A, Olmos E, Sevilla F. 2004. Location and effects of long term NaCl stress onsuperoxide dismutase and ascorbate peroxidase isoenzymes of pea (Pisum sativum cv. Puget)chloroplasts. J Exp Bot. 55:119–130.
Gossett DR, Millhollon EP, Lucas MC. 1994. Antioxidant response to NaCl stress in salt-tolerantand salt-sensitive cultivars of cotton. Crop Sci. 34:706–714.
Gueta-Dahan Y, Yaniv Z, Zilinskas BA, Ben-Hayyim G. 1997. Salt and oxidative stress: similar andspecific responses and their relation to salt tolerance in citrus. Planta. 203:460–469.
Hernandez JA, Jimenez A, Mullineaux P, Sevilla F. 2000. Tolerance of pea (Pisum sativum L.) tolong-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ.23:853–862.
Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ. 2000. Plant cellular and molecular responses tohigh salinity. Ann Rev Plant Physiol Plant Mol Biol. 51:463–499.
International Rice Research Institute (IRRI). 2006. International Rice Research Institute,Philippines. Available from the website: http://www.irri.org
Jebara S, Jebara M, Limam F, Aouani ME. 2005. Changes in ascorbate peroxidase, catalase,guaiacol peroxidase and superoxide dismutase activities in common bean (Phaseolus vulgaris)nodules under salt stress. J Plant Physiol. 162:929–936.
Kim ST, Cho KS, Yu S, Kim SG, Hong JC, Han C, Bae DW. 2003. Proteomic analysis ofdifferentially expressed proteins induced by rice blast fungus and elicitor in suspension-culturedrice cells. Proteomics. 3:2368–2378.
Kong-Ngern K, Daduang S, Womggkham C, Bunnag S, Kosittrakun M, Theerakulpisut P. 2005.Protein profiles in response to salt stress in leaf sheaths of rice seedlings. Sci Asia. 31:403–408.
Kumar V, Shriram V, Jawali N, Shitole MG. 2007. Differential response of indica rice genotypes toNaCl stress in relation to physiological and biochemical parameters. Arch Agron Soil Sci.53:581–592.
Kumar V, Shriram V, Nikam TD, Jawali N, Shitole MG. 2008. Sodium chloride-induced changes inmineral nutrients and proline accumulation in indica rice cultivars differing in salt tolerance.J Plant Nutr. 31:1999–2017.
Laemmli UK. 1970. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature. 227:680–685.
Lee K-S, Chol W-Y, Ko J-C, Kim T-S, Gregorio GB. 2003. Salinity tolerance of japonica and indicarice (Oryza sativa L.) at the seedling stage. Planta. 216:1043–1046.
Lin CC, Kao CH. 2000. Effect of NaCl stress on H2O2 metabolism in rice leaves. Plant GrowthRegul. 30:151–155.
392 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

Luck H. 1974. Catalase. In: Bergmeyer HU, Gawahen K, editors. Methods of enzymatic analysis.Vol. 2. New York: Academic Press. p. 885–894.
Meloni DA, Oliva MA, Martinez CA, Cambraia J. 2003. Photosynthesis and activity of superoxidedismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ Exp Bot.49:69–76.
Mittler R. 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 7:405–410.Murphy KST, Durako MJ. 2003. Physiological effects of short-term salinity changes on Ruppia
maritima. Aquat Bot. 75:293–309.Neto AD de A, Prisco JT, Eneas-Filho J, Braga de Abreu CE, Gomes-Filho E. 2006. Effect of salt
stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant andsalt-sensitive maize genotypes. Environ Exp Bot. 56:87–94.
Oidaira H, Sano S, Koshiba T, Ushimaru T. 2000. Enhancement of antioxidative emzyme activitiesin chilled rice seedlings. J Plant Physiol. 156:811–813.
Pareek A, Singla SL, Grover A. 1997. Salt responsive proteins/genes in crop plants. In: Jaiwal PK,Singh RP, Gulati A, editors. Strategies for improving salt tolerance in higher plants. New Delhi:Oxford and IBH Publication Co. pp 365–391.
Pareek A, Singla SL, Grover A. 1999. Proteins alterations associated with salinity, desiccation, highand low temperature stresses and abscisic acid application in seedlings of Pusa 169 – a highyielding rice (Oryza sativa L.) cultivar. Curr Sci. 75:1023–1035.
Price A, Hendry G. 1991. Ion-catalysed oxygen radical formation and its possible contribution todrought damage in nine native grasses and three cereals. Plant Cell Environ. 14:477–484.
Putter J. 1974. Peroxidase. In: Bergmeyer HU, Gawahen K, editors. Methods of enzymatic analysis.Vol. 2. New York: Academic Press. p. 685.
Rani C, Bishnoi SK, Kumar B, Sheoran IS, Angrish R, Datta KS. 2007. Changes in protein profileunder sodium chloride and boron toxicity stress in seedlings of two wheat cultivars. Indian JPlant Physiol. 12:13–17.
Rubio MC, Gonzalez EM, Minchin FR, Webb KJ, Arrez-Igor C, Ramos J, Becana M. 2002. Effectsof water stress on antioxidant enzymes of leaves and nodules of transgenic alfalfa overexpressingsuperoxide dismutase. Physiol Plantarum. 115:531–540.
Sahi C, Singh A, Kumar K, Blumwald E, Grover A. 2006. Salt stress response in rice: genetics,molecular biology, and comparative genomics. Funct Integ Genomics. 6:263–284.
Sairam RK, Srivastava GC, Agarwal S, Meena RC. 2005. Differences in response to salinity stress intolerant and susceptible wheat genotypes. Biol Plant. 49:85–91.
Sairam RK, Rao KV, Srivastava GC. 2002. Differential response of wheat genotypes to long-termsalinity stress in relation to oxidative stress, antioxidant activity and osmolytes concentration.Plant Sci. 163:1037–1046.
Sandalio LM, Palma JM, del-Rio LA. 1987. Localization of manganese superoxide dismutase inperoxisomes isolated from Pisum sativum L. Plant Sci. 51:1–8.
Shalata A, Tal M. 1998. The effect of salt stress on lipid peroxidation and antioxidants in the leaf ofthe cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiol Plant.104:169–174.
Shalata A, Mittova V, Volokita M, Guy M, Tal M. 2001. Response of the cultivated tomato and itswild salt-tolerant relative Lycopersicon pennellii to salt-dependent oxidative stress: the rootantioxidative system. Physiol Plant. 112:487–494.
Singh NK, Bracken CA, Hasegawa PM, Handa AK, Buckel S, Hermodson MA, Pfankoch F,Regnier FE, Bressan RA. 1987. Characterization of osmotin: a thaumatin-like protein associatedwith osmotic adjustment in plant cells. Plant Physiol. 85:529–536.
Singha S, Choudhury MA. 1990. Effect of salinity (NaCl) stress on H2O2 metabolism in Vigna andOryza seedlings. Biochem Physiol Pflanzen. 186:69–74.
Smith IK, Vierheller TL, Thorpe CA. 1988. Assay of glutathione reductase in crude tissuehomogenates using 5,50-dithiobis (2-nitrobenzoic acid). Ann Biochem. 175:408–413.
Sousa MF, Campos FAP, Prisco JT, Eneas-Filho J, Gomes-Filho E. 2004. Growth and proteinpattern in cowpea seedlings subjected to salinity. Biol Plant. 47:341–346.
Srivalli B, Sharma G, Khanna-Chopra R. 2003. Antioxidative defense system in an upland ricecultivar subjected to increasing intensity of water stress followed by recovery. Physiol Plantarum.119:503–512.
Sudhakar C, Lakshmi A, Giridarakumar S. 2001. Changes in the antioxidant enzyme efficacy in twohigh yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci. 161:613–619.
Archives of Agronomy and Soil Science 393
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009

Vaidyanathan H, Sivakumar P, Chakrabarty R, Thomas G. 2003. Scavenging of reactive oxygenspecies in NaCl stressed rice (Oryza sativa L.)-differential response in salt tolerant and sensitivecultivars. Plant Sci. 165:1411–1418.
Wimmer MA, Muhling KH, Lauchli A, Brown PH, Goldbach HE. 2003. The interaction betweensalinity and boron toxicity affects the sub-cellular distribution of ions and proteins in wheatleaves. Plant Cell Environ. 26:1267–1274.
Woodbury W, Spencer AK, Stahmann MA. 1971. An improved procedure using ferricyanide forcatalase isozymes. Anal Biochem. 44:301–305.
394 V. Kumar et al.
Downloaded By: [Shitole, Mahadeo G.] At: 09:39 18 August 2009




















