
Physiological aspects of tolerance in Atriplex halimus L. to NaCl and drought
11
ORIGINAL PAPER Physiological aspects of tolerance in Atriplex halimus L. to NaCl and drought Mamdouh M. Nemat Alla • Abdel-Hamid A. Khedr • Mamdouh M. Serag • Amina Z. Abu-Alnaga • Reham M. Nada Received: 22 December 2009 / Revised: 29 June 2010 / Accepted: 23 July 2010 Ó Franciszek Go ´rski Institute of Plant Physiology, Polish Academy of Sciences, Krako ´w 2010 Abstract Forty-day-old seedlings of Atriplex halimus were treated either with NaCl (50, 300 and 550 mM) for the subsequent 30 days or with 15% PEG for the sub- sequent 10 days. As much as 50 mM of NaCl significantly increased shoot fresh and dry weight and height; never- theless, 300 or 550 mM NaCl seemed to have no effect. On the other hand, these growth parameters were not affected by drought after 3 or 6 days, but were reduced after 10 days. The gas exchange parameters (photosynthetic rate, stomatal conductance and transpiration rate) were increased by 50 mM NaCl, but decreased by 300 and 550 mM. These parameters were decreased in response to drought only after 10 days of withholding water. In con- trast to Na ? ,K ? was significantly decreased by NaCl but not by drought. The time course effect revealed that phosphoenol pyruvate carboxylase (PEPC) protein was doubled in response to NaCl after 1 and 5 h and continued thereafter, higher than control, while drought had no sig- nificant effect. Rubisco seemed unchanged by NaCl or drought. It could be concluded that the decrease in fresh weight might be attributed to the decrease in water content. Moreover, the decrease in photosynthesis could result from a decrease in stomatal conductance, a protective mecha- nism against water loss to improve water use efficiency. These findings indicate that Atriplex halimus tolerates NaCl and drought through decreasing growth, reducing gas exchange parameters to improve water use efficiency, uptake Na ? and saving, if any, the photosynthetic enzyme particularly PEPC. Keywords Atriplex halimus Á Drought Á Gas exchange Á Growth Á NaCl Á PEPC Á Tolerance Introduction Stresses cause a reduction in plant growth. One cause of growth rate reduction is inefficient photosynthesis. Salinity affects adversely seed germination by osmotic effect and/or ionic toxicity that could result in molecular damage, growth arrest and even death of plants (Duan et al. 2007). Three physiological mechanisms are suggested to be involved in the reduction of plant growth resulting from osmotic stress caused by salinity as well as drought stress: (1) reduction in turgor pressure, (2) reduction in photo- synthetic activity and (3) effects of accumulated salts on the metabolic activities (Neumann 1997). Stress can inhibit cell expansion in both leaves and roots (Neumann 1995). Cell expansion is powered by inward water flow. It depends on the hydraulic conductivity of water uptake pathway, the uptake of solutes to maintain osmotic potential and the volume of the surrounding cells. The reduction occurs due to nutrient deficiency (Hu et al. 2007) and ion toxicity. The gas exchange parameters (photosynthetic rate, transpiration rate and stomatal conductance) are affected by salinity and drought stress (Chaves 1991; Zhu and Meinzer 1999; Munns et al. 2006; Redondo-Gomez et al. 2007). Photosynthetic activity is affected due to malfunc- tion of Rubisco, the key enzyme in Calvin cycle, and of PEPC, the key enzyme of C4 plants. PEPC catalyzes the carboxylation of PEP to form oxaloacetate in mesophyll Communicated by K. Trebacz. M. M. Nemat Alla (&) Á A.-H. A. Khedr Á M. M. Serag Á A. Z. Abu-Alnaga Á R. M. Nada Botany Department, Faculty of Science at Damietta, Mansoura University, Damietta, Egypt e-mail: [email protected] 123 DOI 10.1007/s11738-010-0578-7 Acta Physiol Plant (2011) 33:547–557 / Published online: 2010 13 August
-
Upload
dibrugarhuniversity -
Category
Documents
-
view
2 -
download
0
Transcript of Physiological aspects of tolerance in Atriplex halimus L. to NaCl and drought

ORIGINAL PAPER
Physiological aspects of tolerance in Atriplex halimus L. to NaCland drought
Mamdouh M. Nemat Alla • Abdel-Hamid A. Khedr •
Mamdouh M. Serag • Amina Z. Abu-Alnaga •
Reham M. Nada
Received: 22 December 2009 / Revised: 29 June 2010 / Accepted: 23 July 2010
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2010
Abstract Forty-day-old seedlings of Atriplex halimus
were treated either with NaCl (50, 300 and 550 mM) for
the subsequent 30 days or with 15% PEG for the sub-
sequent 10 days. As much as 50 mM of NaCl significantly
increased shoot fresh and dry weight and height; never-
theless, 300 or 550 mM NaCl seemed to have no effect. On
the other hand, these growth parameters were not affected
by drought after 3 or 6 days, but were reduced after
10 days. The gas exchange parameters (photosynthetic
rate, stomatal conductance and transpiration rate) were
increased by 50 mM NaCl, but decreased by 300 and
550 mM. These parameters were decreased in response to
drought only after 10 days of withholding water. In con-
trast to Na?, K? was significantly decreased by NaCl but
not by drought. The time course effect revealed that
phosphoenol pyruvate carboxylase (PEPC) protein was
doubled in response to NaCl after 1 and 5 h and continued
thereafter, higher than control, while drought had no sig-
nificant effect. Rubisco seemed unchanged by NaCl or
drought. It could be concluded that the decrease in fresh
weight might be attributed to the decrease in water content.
Moreover, the decrease in photosynthesis could result from
a decrease in stomatal conductance, a protective mecha-
nism against water loss to improve water use efficiency.
These findings indicate that Atriplex halimus tolerates
NaCl and drought through decreasing growth, reducing gas
exchange parameters to improve water use efficiency,
uptake Na? and saving, if any, the photosynthetic enzyme
particularly PEPC.
Keywords Atriplex halimus � Drought � Gas exchange �Growth � NaCl � PEPC � Tolerance
Introduction
Stresses cause a reduction in plant growth. One cause of
growth rate reduction is inefficient photosynthesis. Salinity
affects adversely seed germination by osmotic effect and/or
ionic toxicity that could result in molecular damage,
growth arrest and even death of plants (Duan et al. 2007).
Three physiological mechanisms are suggested to be
involved in the reduction of plant growth resulting from
osmotic stress caused by salinity as well as drought stress:
(1) reduction in turgor pressure, (2) reduction in photo-
synthetic activity and (3) effects of accumulated salts on
the metabolic activities (Neumann 1997). Stress can inhibit
cell expansion in both leaves and roots (Neumann 1995).
Cell expansion is powered by inward water flow. It
depends on the hydraulic conductivity of water uptake
pathway, the uptake of solutes to maintain osmotic
potential and the volume of the surrounding cells. The
reduction occurs due to nutrient deficiency (Hu et al. 2007)
and ion toxicity.
The gas exchange parameters (photosynthetic rate,
transpiration rate and stomatal conductance) are affected
by salinity and drought stress (Chaves 1991; Zhu and
Meinzer 1999; Munns et al. 2006; Redondo-Gomez et al.
2007). Photosynthetic activity is affected due to malfunc-
tion of Rubisco, the key enzyme in Calvin cycle, and of
PEPC, the key enzyme of C4 plants. PEPC catalyzes the
carboxylation of PEP to form oxaloacetate in mesophyll
Communicated by K. Trebacz.
M. M. Nemat Alla (&) � A.-H. A. Khedr �M. M. Serag � A. Z. Abu-Alnaga � R. M. Nada
Botany Department, Faculty of Science at Damietta, Mansoura
University, Damietta, Egypt
e-mail: [email protected]
123
DOI 10.1007/s11738-010-0578-7
Acta Physiol Plant (2011) 33:547–557
/ Published online: 201013 August

tissue, from which CO2 is released again in bundle sheath
tissue for the decarboxylation of ribulose-1,5-bisphosphate,
the first enzymatic reaction in Calvin cycle that is catalyzed
by Rubisco. Halophytes and xerophytes undertake the same
measures to counter osmotic stress (Kefu et al. 2003). In
this study, the effects of salinity (NaCl) and drought
(polyethyleneglycol, PEG) in the xero-halophyte Atriplex
halimus were studied to relate tolerance to the harsh con-
ditions with some physiological aspects such as gas
exchange parameters and contents of Na?, K?, PEPC and
Rubisco.
Materials and methods
Plant material and growth conditions
The seeds of Atriplex halimus used in this study were
collected from the Baltim area in Egypt. The seeds were
collected from one bush, mixed properly and stored in
paper bags at room temperature until use. The seeds were
germinated in 10 9 15 cm trays containing vermiculite
and watered daily with Long Ashton nutrient solution. The
trays were incubated in a growth room at 28/20�C day/
night temperature, 16-h photoperiod, 60% RH and
300 lmol m-2 s-1 light intensity. After 19 days, seedlings
with uniform sizes were selected for treatments. For NaCl
stress treatment, the seedlings were transplanted into
4 9 6 cm pots (one seedling per pot) containing vermic-
ulite and the pots were kept in 50 9 35 cm trays contain-
ing Long Ashton nutrient solution (3 L per tray). For
drought treatment, the seedlings were transplanted into
10 9 10 cm pots (one seedling per pot) containing com-
post (150 g per pot) and watered daily with deionized
water.
The salt stress treatment started when the plants were
40 days old. Four sets of plants were used and each set
included 50 plants. The control set was watered with Long
Ashton nutrient solution, while the other three sets were
treated with 50, 300 or 550 mM NaCl for 10 days. For
drought stress treatment, two sets of 40-day-old seedlings
were used. The control set was watered with deionized
water up to the end of experiment and the other set was
exposed to drought stress by withholding water. Twenty
plants from each set were harvested 10 days after applying
NaCl or 3, 6 or 10 days after withholding water. The
leaves were frozen immediately in liquid nitrogen and
then stored at -80�C until use for further analyses. After
harvest, the fresh weights of shoots were recorded and
dried in an oven at 80�C for 2 days and the dry weights
were then recorded. The water contents were calculated on
fresh weight basis. Five plant replicates were used for
each treatment.
Determination of Na? and K?
Na? and K? were extracted as described by Hansen and
Munns (1988). About 100 mg of frozen leaf tissue was first
homogenized in liquid nitrogen and then 2 ml of boiled
water was added. The mixture was kept in a water bath at
100�C for 1 h. The residue was removed by centrifugation
at 14,0009g for 20 min. The diluted supernatant (1:10 with
H2O) was used to measure Na? and K? by flame pho-
tometry. Three replicates were used for each treatment.
Measurement of photosynthesis, transpiration
and stomatal conductance
Photosynthetic rate, transpiration rate and stomatal con-
ductance of the third leaves were measured by using
LCA-4 portable gas exchange system (Analytical Devel-
opment Company Limited, Hertfordshire, England). Leaf
area was measured by using leaf area meter (Delta-T,
Cambridge, UK). Leaf was fitted into a 625-cm2 PLC4 leaf
chamber. Gas exchange measurements were made in the
growth chamber under saturating light conditions (light
sources were filtered using a small heat absorbing filter)
and adjusted to an intensity of 1,000 lmol m-2 s-1. For
each treatment, three replicates were used.
Time course effect of NaCl and drought on Rubisco
and PEPC contents
Another experiment was conduct in which NaCl (0, 50, 200
and 500 mM) or osmotic stress (by using 15% PEG alone
or combined with 50 mM NaCl) were applied to 40-day-
old seedlings. Third leaf samples from each treatment were
harvested after 1, 5, 24, 48 h and 10 days. The leaves were
frozen immediately in liquid nitrogen and then stored at
-80�C until use for further analyses (the protein levels of
Rubisco and PEPC contents).
Extraction of total proteins for SDS-PAGE
The extraction buffer used in protein dissociation was: 4%
SDS, 2% b-mercaptoethanol and 20% glycerol in 100 mM
Tris–HCl (pH 8.5). About 100 mg of frozen leaves was
ground to a fine powder in a pre-cooled mortar and 4 ll of
extraction buffer was added for each 1 mg of leaf material.
The homogenate was heated up to 80�C for 3 min and the
debris was removed by centrifugation at 14,0009g for
10 min. The proteins were resolved as described by
Laemmlli (1970) by using the Bio-Rad Mini Protean 3
(Bio-Rad laboratories, Hercules, CA, USA). The resolving
gel contained 11% acrylamide and the stacking gel con-
tained 5% acrylamide. Proteins from 3 mg leaf fresh
weight were loaded onto each lane. The proteins were
123
Acta Physiol Plant (2011) 33:547–557548

resolved at 100 V for 90 min. The gels were stained with
brilliant blue R-250 (Bio-Rad) and then destained with 20%
methanol. The gels were dried using Bio-Rad gel drier,
scanned and used for Rubisco quantification. Rubisco bands
were quantified using ‘‘Lab Image V 2.7.2’’ program.
Western blotting and immunochemical detection
of PEPC
Proteins were resolved on 11% acrylamide as described
before by loading 3 mg of fresh weight in a volume of
15 ll onto each lane and then transferred to PVDF mem-
branes (Bio-Rad) using Bio-Rad mini protean 3 transblot-
ter. The transfer buffer consisted of 5.8 g l-1 Tris base and
2.9 g l-1 glycine. The proteins were transferred for 1 h at
100 V and 4�C. The membranes were then blocked in 2.5%
dry non-fat dried milk in TBS (Tris-buffered saline:
150 mM NaCl in 20 mM Tris–HCl, pH 7.4). The blots
were used for immunochemical detection of PEPC. PEPC
was detected by using a primary antibody developed in
rabbit. The blots were incubated in 1:1,000 dilution of the
primary antibody in 2.5% milk TBS for 2 h. After five
washes (3 min each) in TBS, the membranes were incu-
bated in 1:1,000 dilution of the secondary antibody (goat
anti-rabbit IgG-horseradish peroxidase conjugate) (Sigma)
for 90 min and washed as before. The signals were
detected by ECL kit (Pharmacia Amersham) and visualized
according to the manufacturer’s instructions.
The full data were statistically analyzed using SPSS V
12.0.1 for ANOVA test.
Results
Figure 1 shows that the soil water content remained at
81%. Withholding water after the addition of PEG resulted
in significant decreases in soil water content. This decrease
was augmented as the time of withholding water elapsed.
Soil water content decreased from 81% in the control to 70,
49 and 22% after 3, 6 and 10 days of withholding water,
respectively.
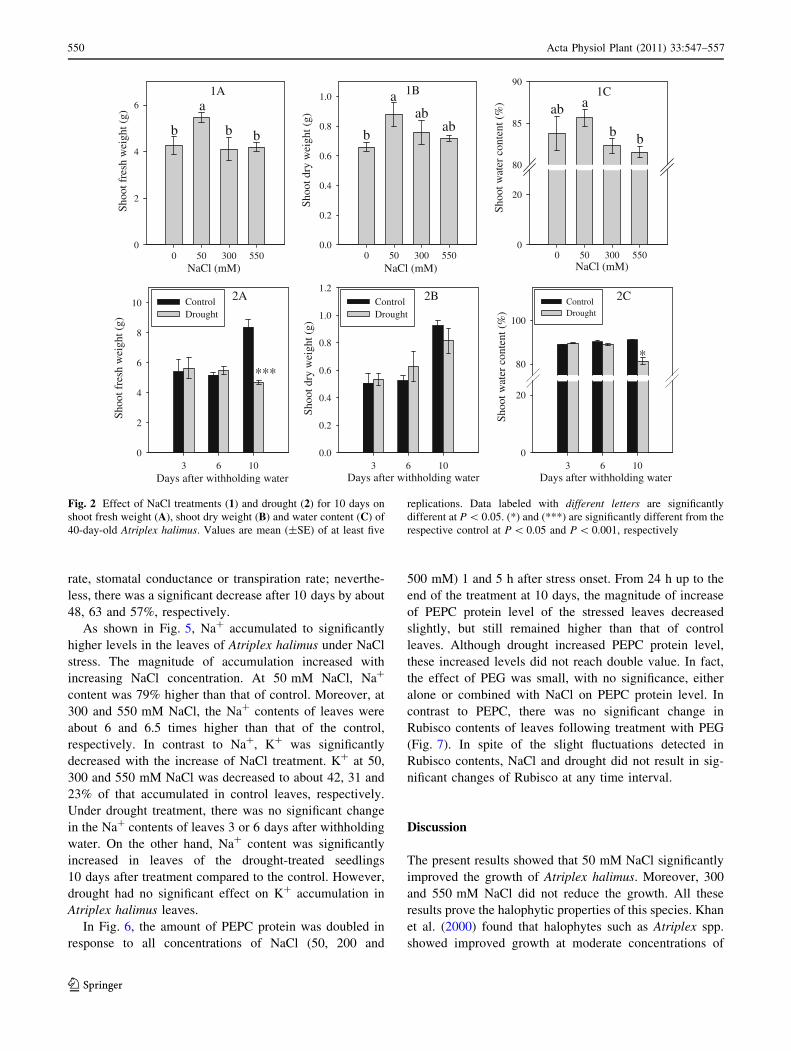
Shoot fresh weight of Atriplex halimus seedlings was
maximum at 50 mM NaCl (Fig. 2). However, the other
concentrations showed significant reduction in weight
compared to 50 mM, but had no effects relative to control.
Similarly, the shoot dry weight was significantly greater in
the presence of 50 mM NaCl than that of the control.
However, shoot dry weights remained unchanged with
other concentrations. On the other hand, water content at
all concentrations of NaCl was comparable to control.
Nevertheless, water content at 300 and 550 mM was sig-
nificantly lower than that at 50 mM NaCl. On the other
hand, drought stress had no significant effect on fresh and
dry weight 3 and 6 days after withholding water. On the
contrary, shoot fresh weight significantly decreased by
about 45%, 10 days after withholding water; however,
shoot dry weight was not significantly changed. On the
other hand, shoot water content did not show significant
reduction due to drought 3 or 6 days after withholding
water. Nevertheless, shoot water content significantly
decreased 10 days after withholding water to reach 81%
compared to that of the well-watered seedlings.
In Fig. 3, NaCl at 300 and 550 mM significantly
decreased the shoot height by about 15 and 27%, respec-
tively; nevertheless, 50 mM had no effect. Also, there was
no significant change in leaf numbers of seedlings under 50
or 300 mM NaCl, but 550 mM resulted in a significant
decrease up to 31% compared to the control. Also in Fig. 3,
there was no significant effect of drought on the shoot
height 3 or 6 days after withholding water. However, shoot
height was significantly reduced by 15% of the control
after 10 days. The same trend was observed for the number
of leaves; there was a significant decrease in leaf number
by about 37% compared to the well-watered seedlings only
after 10 days.
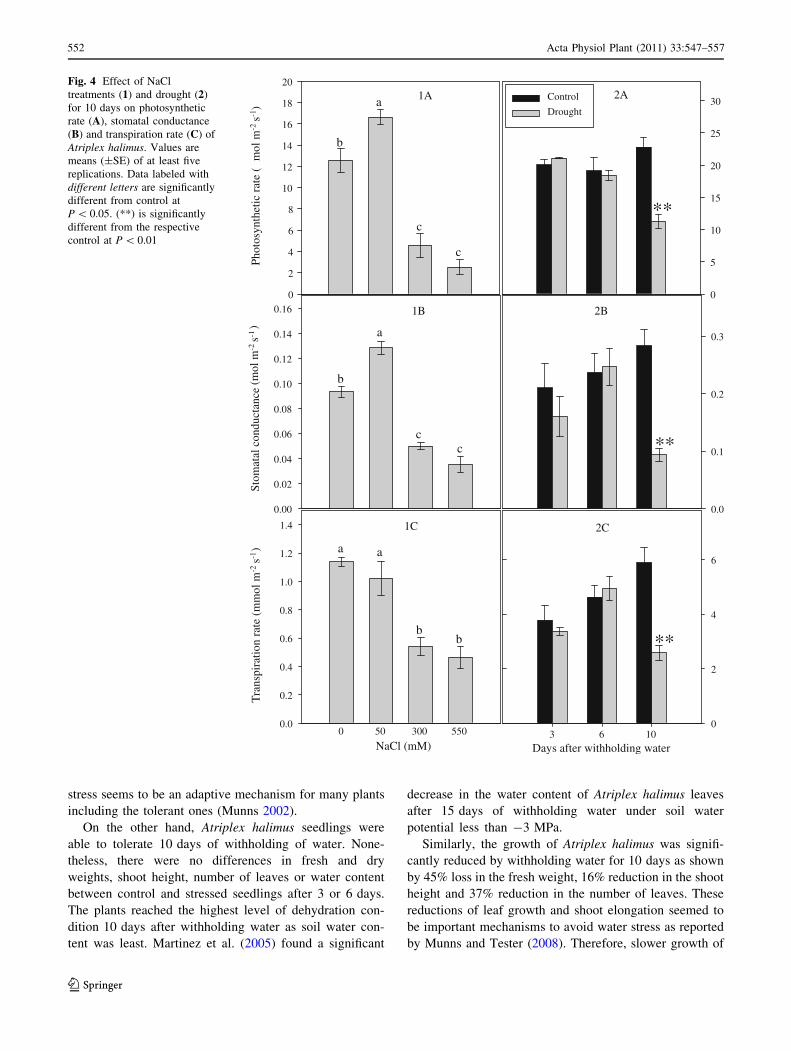
The photosynthetic rate increased significantly in the
presence of 50 mM NaCl compared to the control (Fig. 4).
However, it was significantly reduced by about three and
fourfolds at 300 and 550 mM NaCl, respectively. The same
trend was observed for stomatal conductance where the
highest value was shown at 50 mM NaCl, but decreased at
300 and 550 mM of NaCl by 45 and 62% of the control,
respectively. The transpiration rate remained similar to the
control at 50 mM NaCl, but decreased at 300 and 550 mM
of NaCl, by about three and fourfolds as compared to the
control, respectively. At 3 and 6 days after withholding
water, there was no significant change in photosynthetic
Days after withholding water3 6 10
Soil
wat
er c
onte
nt (
%)
0
20
40
60
80
100
120ControlDrought
a
b
c
d
aa
Fig. 1 Soil water content 3, 6 and 10 days after withholding water
from 40-day-old Atriplex halimus. Values are mean (±SE) of at least
five replications. Data labeled with different letters are significantly
different at P \ 0.05
123
Acta Physiol Plant (2011) 33:547–557 549
rate, stomatal conductance or transpiration rate; neverthe-
less, there was a significant decrease after 10 days by about
48, 63 and 57%, respectively.
As shown in Fig. 5, Na? accumulated to significantly
higher levels in the leaves of Atriplex halimus under NaCl
stress. The magnitude of accumulation increased with
increasing NaCl concentration. At 50 mM NaCl, Na?
content was 79% higher than that of control. Moreover, at
300 and 550 mM NaCl, the Na? contents of leaves were
about 6 and 6.5 times higher than that of the control,
respectively. In contrast to Na?, K? was significantly
decreased with the increase of NaCl treatment. K? at 50,
300 and 550 mM NaCl was decreased to about 42, 31 and
23% of that accumulated in control leaves, respectively.
Under drought treatment, there was no significant change
in the Na? contents of leaves 3 or 6 days after withholding
water. On the other hand, Na? content was significantly
increased in leaves of the drought-treated seedlings
10 days after treatment compared to the control. However,
drought had no significant effect on K? accumulation in
Atriplex halimus leaves.
In Fig. 6, the amount of PEPC protein was doubled in
response to all concentrations of NaCl (50, 200 and
500 mM) 1 and 5 h after stress onset. From 24 h up to the
end of the treatment at 10 days, the magnitude of increase
of PEPC protein level of the stressed leaves decreased
slightly, but still remained higher than that of control
leaves. Although drought increased PEPC protein level,
these increased levels did not reach double value. In fact,
the effect of PEG was small, with no significance, either
alone or combined with NaCl on PEPC protein level. In
contrast to PEPC, there was no significant change in
Rubisco contents of leaves following treatment with PEG
(Fig. 7). In spite of the slight fluctuations detected in
Rubisco contents, NaCl and drought did not result in sig-
nificant changes of Rubisco at any time interval.
Discussion
The present results showed that 50 mM NaCl significantly
improved the growth of Atriplex halimus. Moreover, 300
and 550 mM NaCl did not reduce the growth. All these
results prove the halophytic properties of this species. Khan
et al. (2000) found that halophytes such as Atriplex spp.
showed improved growth at moderate concentrations of
NaCl (mM)0 50 300 550
Shoo
t fre
sh w
eigh
t (g)
0
2
4
6 a
b b b
Shoo
t dry
wei
ght (
g)
NaCl (mM)0 50 300 550
Shoo
t wat
er c
onte
nt (
%)
0
20
80
85
90
a
b
ab
b
1A 1C
Shoo
t dry
wei
ght (
g)
NaCl (mM)0 50 300 550
0.0
0.2
0.4
0.6
0.8
1.0 a
bab
ab
1B
Days after withholding water3 6 10
0.0
0.2
0.4
0.6
0.8
1.0
1.2ControlDrought
Days after withholding water3 6 10
0
20
80
100
ControlDrought
*
C2
Shoo
t wat
er c
onte
nt (
%)
B2
Days after withholding water3 6 10
Shoo
t fre
sh w
eigh
t (g)
0
2
4
6
8
10 ControlDrought
***
A2
Fig. 2 Effect of NaCl treatments (1) and drought (2) for 10 days on
shoot fresh weight (A), shoot dry weight (B) and water content (C) of
40-day-old Atriplex halimus. Values are mean (±SE) of at least five
replications. Data labeled with different letters are significantly
different at P \ 0.05. (*) and (***) are significantly different from the
respective control at P \ 0.05 and P \ 0.001, respectively
123
Acta Physiol Plant (2011) 33:547–557550

NaCl, which seem to be inhibitory for the growth of non-
halophyte plants. They concluded that Atriplex griffithii
showed little inhibition in seedling growth in a medium
containing up to 180 mM NaCl. They, moreover, con-
firmed that deleterious effects of salt stress on plants are
thought to result from low water potentials, ion toxicities,
nutrient deficiencies or a combination of all of these
factors.
Salinity imposed by 50 mM NaCl increased shoot fresh
and dry weights of Atriplex halimus, but shoot height and
the number of leaves were unchanged. The higher con-
centrations seemed to have no effect on fresh and dry
weights, but decreased the shoot height and leaf number.
These results indicate that high concentrations of NaCl
seemed to have no dramatic effect on growth of Atriplex
halimus. Moreover, the optimal growth of Atriplex halimus
appeared to require low concentration of NaCl. Atriplex
halimus, like other halophytes such as Suaeda salsa (Kefu
et al. 2003), could adapt to conditions of low salinity
without reduction in dry matter by using Na? in shoot
growth, accumulating it inside the vacuole to maintain a
lower water potential (Nobel 1988). Kefu et al. (2003)
found that Suaeda salsa adapts very well to saline soil,
concluding that it could absorb quite a lot of Na and Cl
from soil and compartmentalize them in the vacuoles to
lower the plant water potential, hence it can absorb water to
maintain life. They related this adaptation to the relatively
small contribution of organic osmotica to osmotic adjust-
ment and also to the high activity of H?-ATPase and H?-
PPase in the tonoplast, which endow the plant with more
energy to translocate Na into vacuoles through Na?/H?
antiports. Moreover, the water content of Atriplex halimus
decreased significantly at 300 and 550 mM NaCl. There-
fore, the decrease in shoot fresh weight might be attributed
to the decrease in water content rather than to decrease in
the dry weight. The decrease in water content during salt
NaCl (mM)0 50 300 550
Shoo
t hei
ght (
cm)
0
10
20
30 a
bc
a
NaCl (mM)0 50 300 550
Num
ber
of le
aves
0
20
40
60
80
100
a
bab
a
Days after withholding water3 6 10
Shoo
t hei
ght (
cm)
0
5
10
15
20
25 Control
Drought
*
Days after withholding water3 6 10
Num
ber
of le
aves
0
20
40
60
80
100 Control
Drought
***
1B1A
2A 2B
Fig. 3 Effect of NaCl
treatments (1) and drought (2)
for 10 days on shoot height
(A) and number of leaves (B) of
Atriplex halimus. Values are
mean (±SE) of at least five
replications. Data labeled with
different letters are significantly
different at P \ 0.05. (*) and
(***) are significantly different
from the respective control at
P \ 0.05 and P \ 0.001,
respectively
123
Acta Physiol Plant (2011) 33:547–557 551
stress seems to be an adaptive mechanism for many plants
including the tolerant ones (Munns 2002).
On the other hand, Atriplex halimus seedlings were
able to tolerate 10 days of withholding of water. None-
theless, there were no differences in fresh and dry
weights, shoot height, number of leaves or water content
between control and stressed seedlings after 3 or 6 days.
The plants reached the highest level of dehydration con-
dition 10 days after withholding water as soil water con-
tent was least. Martinez et al. (2005) found a significant
decrease in the water content of Atriplex halimus leaves
after 15 days of withholding water under soil water
potential less than -3 MPa.
Similarly, the growth of Atriplex halimus was signifi-
cantly reduced by withholding water for 10 days as shown
by 45% loss in the fresh weight, 16% reduction in the shoot
height and 37% reduction in the number of leaves. These
reductions of leaf growth and shoot elongation seemed to
be important mechanisms to avoid water stress as reported
by Munns and Tester (2008). Therefore, slower growth of
NaCl (mM)
Tra
nspi
ratio
n ra
te (
mm
ol m
-2 s
-1 )
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
a
b
a
b
Phot
osyn
thet
ic r
ate
(µm
ol m
-2 s
-1)
0
2
4
6
8
10
12
14
16
18
20
a
b
c
c
Stom
atal
con
duct
ance
(m
ol m
-2 s
-1 )
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
a
b
cc
1A
1B
1C
0
5
10
15
20
25
30Control
Drought
**
0.0
0.1
0.2
0.3
**
Days after withholding water
0 50 300 550 3 6 100
2
4
6
**
2B
2C
2AFig. 4 Effect of NaCl
treatments (1) and drought (2)
for 10 days on photosynthetic
rate (A), stomatal conductance
(B) and transpiration rate (C) of
Atriplex halimus. Values are
means (±SE) of at least five
replications. Data labeled with
different letters are significantly
different from control at
P \ 0.05. (**) is significantly
different from the respective
control at P \ 0.01
123
Acta Physiol Plant (2011) 33:547–557552

Atriplex halimus under salt or drought stress could be an
adaptive feature that enables plants to rely on other
resources as building blocks (e.g. producing compounds
that can be used as compatible solutes and/or osmolytes to
maintain its normal status) and save energy to affect
adaptation under these harsh conditions (Zhu 2001).
Days after withholding water3 6 10
Lea
f N
a+ co
nten
t (µm
ol g
-1 F
Wt)
0
50
100
150
200
250
Control
Drought**
Days after withholding water3 6 10
Lea
f K
+co
nten
t (µm
ol g
-1 F
Wt)
0
50
100
150
200
Control
Drought
NaCl (mM)
Lea
f N
a+co
nten
t (µm
ol g
-1 F
Wt)
0
200
400
600
800
1000
1200
1400
1600
1800
a
b
c
a
NaCl (mM)0 50 300 550 0 50 300 550
Lea
f K
+ c
onte
nt (
µmol
g-1
FW
t)
0
20
40
60
80
100
120
140
160
180
200
a
b
bc
c
1B1A
2B2A
Fig. 5 Effect of NaCl
treatments (1) and drought (2)
for 10 days on Na? (A) and K?
(B) content in Atriplex halimusleaves. Values are mean (±SE)
of at least five replications. Data
labeled with different letters are
significantly different at
P \ 0.05. (**) is significantly
different from the respective
control at P \ 0.01
Fig. 6 Time course effect of NaCl treatments (1) and drought (2) on PEPC protein level of Atriplex halimus. A Western blot and B the protein
level (±SE) of at least four replications from two independent experiments
123
Acta Physiol Plant (2011) 33:547–557 553

Gas exchange parameters were affected by both salt and
drought stresses. There was a significant increase in pho-
tosynthetic rate and stomatal conductance in plants treated
with 50 mM NaCl and a significant decrease by 300 and
550 mM of NaCl. The transpiration rate was reduced only
under higher concentrations. This is consistent with pre-
vious findings. Zhu and Meinzer (1999) found that 4 weeks
after imposing stress, photosynthetic rate and stomatal
conductance were moderately enhanced at 50 and 100 mM
NaCl, but significantly decreased at higher levels (400, 500
and 600 mM NaCl). Na? has a positive impact on C4
photosynthesis, where it is found to increase the conversion
of pyruvate into phosphoenolpyruvate in the light condition
(Murata et al. 1992). Conversely, Redondo-Gomez et al.
(2007) found significant decreases in net photosynthetic
rate and stomatal conductance of Atriplex portulacoides
with increasing NaCl from 20 to 700 mM.
Photosynthesis is among the primary processes affected
by drought (Chaves 1991) and salinity (Munns et al. 2006).
The effect can be direct through decrease in CO2 assimi-
lation and diffusion from the stomata to mesophyll cells
(Flexas et al. 2007) or through the alteration in the pho-
tosynthetic mechanism (Lawlor and Cornic 2002) and
indirect through oxidative stress that impairs the photo-
synthetic machinery (Chaves et al. 2009). The decrease in
photosynthetic rate in Atriplex halimus under salt and
drought treatment might be attributed to the decrease in the
availability of CO2 resulting from reduction of stomatal
conductance, but not due to injuries in the photosynthetic
apparatus (Demmig and Bjorkman 1987).
The gas exchange parameters of Atriplex halimus did
not show significant changes 3 and 6 days after withhold-
ing water, but decreased after 10 days. Brown and Pezeshki
(2007) reported that the stomatal conductance and the net
photosynthetic rate of dehydrated Spartina alterniflora
significantly declined after 8 days of drought stress. Simi-
larly, the stomatal conductance of Carrizo citrange was
negatively affected either after a dehydration period of
8–18 days or after exposure to 100 mM NaCl (Perez-Perez
et al. 2007).
Plants rely on reducing their stomatal conductance as a
protective mechanism against excessive water loss (Chaves
et al. 2009). The present study indicates that the decline in
stomatal conductance under high concentrations of NaCl
was not accompanied with change in water content.
Redondo-Gomez et al. (2007) observed that the decrease in
stomatal conductance was not accompanied with a decline
in the relative water content in the salt-stressed Atriplex
portulacoides. They attributed the decline in stomatal
conductance to the signaling process rather than to the loss
of turgor. Therefore, Atriplex portulacoides was able to
increase water use efficiency under high salinities. There-
fore, the decline in the transpiration rate of Atriplex hali-
mus subjected to 300 and 550 mM NaCl as well as to
drought for 10 days could be considered as one of the
tolerance mechanisms, as supported by Veselov et al.
(2008).
Chen et al. (2003) reported that lowering the transpira-
tion rates under salt stress contributes to improving their
ability to control Na? influx into roots and subsequently to
the shoots. However, Atriplex halimus actively accumu-
lates Na? to high concentrations in its shoots at high
concentrations; therefore, the decrease in the transpiration
rate does not seem to be essentially for the purpose of
avoidance of excessive Na? influx. The reduction in the
transpiration rate in Atriplex halimus could be an avoidance
mechanism to prevent water loss under salt and drought
stress, i.e. an improvement in the water use efficiency.
Fig. 7 Time course effect of NaCl treatments (1) and drought (2) on Rubisco protein level of Atriplex halimus. A SDS-Page and B the protein
level (±SE) of at least four replications from two independent experiments
123
Acta Physiol Plant (2011) 33:547–557554

Similar responses were reported in Argyranthemum cor-
onopifolium (De Herralde et al. 1998).
Vacuoles represent about 90% of mature cell volume, so
ions can act as ‘‘cheap’’ osmolytes, if they are sequestered
properly in the vacuole. Ions, particularly Na?, require less
energy compared to other organic compounds such as
proline and glycine betaine used by plants for osmotic
adjustment (Raven 1985). Growth and survival of halo-
phytes is attributed to their capacity to accumulate high
concentrations of Na? in their vacuoles and use it in the
maintenance of turgor and osmotic adjustment (Flowers
1972).
The stimulation of Atriplex halimus growth at 50 mM
NaCl was associated with Na? accumulation, indicating its
ability to resist salt stress by accumulating Na? and not to
restrict its uptake as in glycophytes (Lutts et al. 1996).
Similar results were obtained by Silveira et al. (2009) who
found that Na? concentrations gradually increased inside
Atriplex nummularia leaves with increasing NaCl concen-
trations from 0 to 600 mM.
In some Atriplex species, the increased sequestration of
Na? appears to be mainly into leaf vacuoles and special-
ized leaf structures such as trichomes and vesicles, where
toxic concentrations of Na? and Cl- are kept away from
the other biochemically active cell compartments (Bradley
and Morris 1991). The accumulation of Na? inside the
vacuole requires an active transport against a concentration
gradient (Silveira et al. 2009). The accumulation of high
concentrations of Na?, in the present study, was accom-
panied by a slight decline in water content. Conversely, the
leaf succulence of Cakile maritima was significantly
enhanced by NaCl concentrations up to 500 mM (Debez
et al. 2006). Also, the accumulation of Na? inside the
leaves of Atriplex halimus improved the water status of the
leaves and had positive consequences on the metabolic and
osmotic processes (Debez et al. 2003). Therefore, leaf
succulence and/or the ability to conserve hypertony are
considered to be one of the important features that char-
acterize the halophytes.
Under water deficit conditions, Na? was increased only
10 days after starting dehydration. As in salt stress, Na?
might be involved in osmotic adjustment; nevertheless, the
insignificant change in Na? content after 3 and 6 days might
be attributed to the absence of stress at these points as
revealed by unchanged growth and soil water content. Under
water deficit, the plant is exposed to a combination of water
stress and ionic stress as the soil matrix potential decreases
simultaneously with decreasing soil moisture. It was found
that the dehydrated Atriplex halimus accumulated more Na?
than the control plants even without the addition of NaCl to
the stressed plants (Martinez et al. 2003).
According to these findings, specific Na? uptake could
be concluded as an integral part of the response of Atriplex
halimus to salt stress as well as to drought stress. Despite
accumulating high concentrations of Na? in leaves of
Atriplex halimus, its growth did not show a consequent
dramatic decrease as reported in glycophytes in which
growth was considerably inhibited by exposure to 200 mM
NaCl (Zhang et al. 2001). Glenn and Brown (1998) con-
cluded that tolerance of Atriplex canescens to water and
salt stress was linked through a common mechanism of
accumulating Na? for osmotic adjustment.
In contrast to Na? accumulation inside Atriplex halimus
leaves, K? concentrations were significantly decreased by
NaCl concentrations. Bajji et al. (1998) found that K?
decreased with the increasing NaCl concentration in the
external medium from 0 to 600 mM. Similar results were
detected in Atriplex nummularia (Silveira et al. 2009). This
decrease in K? content in the salt-stressed Atriplex halimus
may be attributed to the competitive uptake of Na? and K?
(Pitman 1981), the depletion of K? in leaves in which Na?
replaces K? (Redondo-Gomez et al. 2007), depolarization
of the plasma membrane with a consequent K? loss
(Shabala et al. 2005) and down-regulation of the genes
involved in K? transport (Zhu 2003).
On the other hand, drought had no effect on the K?
content. These findings are consistent with the unchanged
K? content found in Atriplex halimus exposed to water
stress for 22 days (Martinez et al. 2003). Such unchanged
K? content in dehydrated Atriplex halimus leaves in the
present study might be attributed to the limited Na?
already found in the soil that compete with the K? uptake
by the plant. This conclusion could be supported by the
data from salt experiment, in which K? content decreased
with increasing application of external Na?.
Photosynthetic rate could be decreased as a result of
stomatal closures (Garcia-Legaz et al. 1993) as well as
stress-induced damage to Rubisco and other enzymes in the
Calvin cycle (Rivelli et al. 2002). In this study, there was
no significant change in the Rubisco content (not the
activity) under all concentrations of NaCl, whereas the
content of PEPC was doubled by 50, 200 and 500 mM
NaCl within 1 and 5 h after treatment and remained
thereafter higher than control. Bouraima et al. (1987) stated
that the response of PEPC in pearl millet to mild treatment
with NaCl depends on the tolerance of the seedlings to the
salt. PEPC was enhanced in the salt-tolerant genotype and
decreased in the most sensitive one. They concluded that
PEPC may be effective markers of salt tolerance in the C4
plant. Moreover, Li and Chollet (1994) found that treat-
ment of the common ice plant (Mesembryanthemum crys-
tallinum) with high salinity caused an increase in PEPC
protein. Contrarily, Debez et al. (2006) reported that
Rubisco activity of Cakile maritime significantly increased
at salinity levels of 100–200 mM NaCl but declined at
higher levels, while PEPC activity did not show any
123
Acta Physiol Plant (2011) 33:547–557 555

difference from that of control plants. Also, Zhu and
Meinzer (1999) found a decrease in Rubisco and PEPC
activities in response to salt stress in Atriplex lentiformis.
On the other hand, the contents (not the activities) of
both enzymes were not significantly affected by PEG either
alone or combined with NaCl. In this regard, Majumdar
et al. (1991) stated that exposing Glycine max to drought
for 8 days sharply inhibited the activity of Rubisco within
2 days. They, moreover, found that although PEPC activity
did not show any significant change after 2 days, it
declined to almost zero value within the 5th day. In
dehydrated sugarcane (Saccharum officinarum), Rubisco
activity remained the same as that of a well-watered plant,
but PEPC activity increased with decreasing availability of
water (Saliendra et al. 1996).
The present results indicate that the decrease in stomatal
conductance at high concentration was not accompanied by
a decrease in Rubisco, suggesting that the reduction in
photosynthetic rate was due to the limitation of CO2 rather
than photosynthetic enzyme activity. Moreover, the
unchanged Rubisco content in Atriplex halimus under salt
and/or drought stress is a proof of the ability of this plant to
save the most important enzyme in the photosynthetic
machinery to resume its growth under this harsh environ-
ment. PEPC could be involved in increasing the efficiency
of photosynthesis. Moreover, PEPC content increased
during severe conditions to refix CO2 that passively leaked
from the bundle sheath under salt conditions (Hatch and
Osmond 1976).
In conclusion, the present findings showed that 50 mM
NaCl improved the growth parameters of Atriplex halimus.
On the other hand, growth was reduced by drought treat-
ment after withholding water for only 10 days. The gas
exchange parameters were increased by 50 mM NaCl, but
decreased by drought after 10 days. Despite increasing
Na?, all treatments decreased K?. The insignificant change
in Na? content after 3 and 6 days might be attributed to the
absence of stress. However, the accumulated Na? could be
considered as an integral part of the response of Atriplex
halimus to stress. PEPC protein was increased by NaCl but
was unaffected by drought, while Rubisco seemed
unchanged by all treatments. These results revealed that the
decrease in fresh weight might be attributed to the decrease
in water content. Moreover, the decrease in stomatal con-
ductance was not accompanied by a decrease in Rubisco
suggesting that the reduced photosynthesis was due to
limitation of CO2 rather than inhibited photosynthetic
enzyme activity. However, the decrease in photosynthesis
might be attributed to the decrease in the availability of
CO2 resulting from reduction of stomatal conductance. So,
plants rely on reducing their stomatal conductance as a
protective mechanism against water loss. Such reduction
could improve their water use efficiency. These findings
indicate that Atriplex halimus can tolerate low NaCl con-
centration and also short-term drought. Such tolerance
might be regulated by reducing growth, to save energy, and
gas exchange parameters, to improve water use efficiency,
as well as increasing Na? uptake and PEPC protein to
efficiently refix CO2 that passively leaked from the bundle
sheath under stress conditions.
References
Bajji M, Kinet JM, Lutts S (1998) Salt stress effects on roots and
leaves of Atriplexhalimus L. and their corresponding callus
cultures. Plant Sci 137:131–142
Bouraima S, Vidal J, Lavergne D, Hoarau A, Champigny M-L (1987)
Effects of sodium chloride stress on phosphoenolpyruvate
carboxylase, NADP-malic enzyme and ribulose-1, 5-bisphos-
phate carboxylase in shoots of pearl millet. Phytochemistry
26:1329–1332
Bradley PM, Morris JT (1991) Relative importance of ion exclusion,
secretion and accumulation in Spartina alterniflora Loisel. J Exp
Bot 42:1525–1532
Brown CE, Pezeshki SR (2007) Threshhold for recovery in the marsh
halophyte Spartina alterniflora grown under the combined
effects of salinity and soil drying. J Plant Physiol 164:274–282
Chaves MM (1991) Effects of water deficits on carbon assimilation.
J Exp Bot 42:1–46
Chaves MM, Flexas J, Pinheiro C (2009) Photosynthesis under
drought and salt stress: regulation mechanisms from whole plant
to cell. Ann Bot 103:551–560
Chen S, Li J, Wang S, Fritz E, Huttermann A, Altman A (2003)
Effects of NaCl on shoot growth, transpiration, ion compart-
mentation, and transport in regenerated plants of Populuseuphratica and Populus tomentosa. Can J For Res 33:967–975
De Herralde F, Biel C, Save R, Morales MA, Torrecillas A, Alarcon
JJ, Sanchez-Blanco MJ (1998) Effect of water and salt stress on
the growth, gas exchange and water relations in Argyranthemumcoronopifolium plants. Plant Sci 139:9–17
Debez A, Chaibi W, Bouzid S (2003) Physiological responses and
structural modifications in Atriplex halimus L. plants exposed to
salinity. In: Lieth H, Moschenko M (eds) Cash crop halophytes:
recent studies: 10 years after the Al Ain meeting. Kluwer
Academic Publishers, Dordrecht, The Netherlands, pp 19–30
Debez A, Saadaoui D, Ramani B, Ouerghi Z, Koyro H-W, Huchzer-
meyer B, Abdelly C (2006) Leaf H?-ATPase activity and
photosynthetic capacity of Cakile maritima under increasing
salinity. Environ Exp Bot 57:285–295
Demmig B, Bjorkman O (1987) Comparison of the effect of
excessive light on chlorophyll fluorescence (77 K) and photon
yield of O2 evolution in leaves of higher plant. Planta
171:171–174
Duan DY, Li WQ, Liu XJ, Ouyang H, An P (2007) Seed germination
and seedlings growth of Suaeda salsa under salt stress. Ann Bot
Fennici 44:161–169
Flexas J, Diaz-Espejo A, Galme’s J, Kaldenhoff R, Medrano H,
Ribas-Carbo M (2007) Rapid variations of mesophyll conduc-
tance in response to changes in CO2 concentration around leaves.
Plant Cell Environ 30:1284–1298
Flowers TJ (1972) Salt tolerance in Suaeda maritima (L.) Dum. J Exp
Bot 23:310–321
Garcia-Legaz MF, Ortiz JM, Garcia-Lindon A, Cerda A (1993) Effect
of salinity on growth, ion content and CO2 assimilation rate in
123
Acta Physiol Plant (2011) 33:547–557556

lemon varieties on different rootstocks. Physiol Plant
89:127–142
Glenn EP, Brown JJ (1998) Effects of soil salt levels on the growth
and water use efficiency of Atriplex canescens (Chenopodiaceae)
varieties in drying soil. Am J Bot 85:10–16
Hansen EH, Munns DN (1988) Effect of CaSO4 and NaCl on mineral
content of Leucaena leucocephala. Plant Soil 7:101–105
Hatch MD, Osmond CB (1976) Compartmentation and transport in
C4 photosynthesis. In: Stocking CR, Heber U (eds) Transport in
plants. III. Intracellular interactions and transport processes.
Encyclopedia of Plant Physiology, New series 3:144–184
Hu Y, Burucs Z, von Tucher S, Schmidhalter U (2007) Short-term
effects of drought and salinity on mineral nutrient distribution
along growing leaves of maize seedlings. Environ Exp Bot
60:268–275
Kefu Z, Hai F, San Z, Jie S (2003) Study on the salt and drought
tolerance of Suaeda salsa and Kalanchoe claigremontiana under
iso-osmotic salt and water stress. Plant Sci 165:837–844
Khan MA, Ungar IA, Showalter AM (2000) Effects of salinity on
growth, water relations and ion accumulation of the subtropical
perennial halophyte, Atriplex griffithii var. stocksii. Ann Bot
85:225–232
Laemmlli EK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680–685
Lawlor DW, Cornic G (2002) Photosynthetic carbon assimilation and
associated metabolism in relation to water deficits in higher
plants. Plant Cell Environ 25:275–294
Li B, Chollet R (1994) Salt induction and the partial purification/
characterization of phosphoenolpyruvate carboxylase protein-
serine kinase from an inducible crassulacean-acid-metabolism
(CAM) plant, Mesembryanthemum crystallinum L. Arch Bio-
chem Biophys 314:247–254
Lutts S, Kinet JM, Bouharmont J (1996) NaCl-induced senescence in
leaves of rice (Oryza sativa L.) cultivars differing in salinity
resistance. Ann Bot 78:389–398
Majumdar S, Ghosh S, Glick BR, Erwin B, Dumbrof T (1991)
Activities of chlorophyllase, phosphoenolpyruvate carboxylase
and ribulose-l,5-bisphosphate carboxylase in the primary leaves
of soybean during senescence and drought. Physiol Plant
8:473–480
Martınez JP, Ledent JF, Bajji M, Kinet JM, Lutts S (2003) Effect of
water stress on growth, Na and K accumulation and water use
efficiency in relation to osmotic adjustment in two populations of
Atriplex halimus L. Plant Growth Regul 41:63–73
Martınez JP, Kinet JM, Bajji M, Lutts S (2005) NaCl alleviates
polyethylene glycol-induced water stress in the halophyte
species Atriplex halimus L. J Exp Bot 56:2421–2431
Munns R (2002) Comparative physiology of salt and water stress.
Plant Cell Environ 25:239–250
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Ann
Rev Plan Biol 59:651–681
Munns R, James RA, Lauchli A (2006) Approaches to increasing the
salt tolerance of wheat and other cereals. J Exp Bot
57:1025–1043
Murata S, Kobayashi M, Matoh T, Sekiya J (1992) Sodium stimulates
regeneration of phosphoenolpyruvate in mesophyll chloroplasts
of Amaranthus tricolor. Plant Cell Physiol 33:1247–1250
Neumann PM (1995) The role of cell wall adjustment in plant
resistance to water deficits. Crop Sci 35:1258–1266
Neumann PM (1997) Salinity resistance and plant growth revisited.
Plant Cell Environ 20:1193–1198
Nobel PS (1988) Environmental biology of Agaves and Cocti.Cambridge University Press, Cambridge, NY
Perez-Perez JG, Syvertsen JP, Botia P, Garcia-Sanchez F (2007) Leaf
water relations and net gas exchange responses of salinized
Carrizo Citrange seedlings during drought stress and recovery.
Ann Bot 100:335–345
Pitman MG (1981) Ion uptake. In: Paleg LG, Aspinall DD (eds)
Physiology and biochemistry of drought resistance in plants.
Academic Press, New York, pp 71–96
Raven JA (1985) Regulation of pH and generation of osmolarity in
vascular plants: a cost-benefit analysis in relation to efficiency of
use of energy, nitrogen and water. New Phytol 101:25–77
Redondo-Gomez S, Mateos-Naranjo E, Davy AJ, Fernandez-Munoz
F, Castellanos EM, Luque T, Figueroa MA (2007) Growth and
photosynthetic responses to salinity of the salt-marsh shrub
Atriplex portulacoides. Ann Bot 100:555–563
Rivelli AR, Lovelli S, Perniola M (2002) Effects of salinity on gas
exchange, water relations and growth of sunflower (Helianthusannuus). Funct Plant Biol 29:1405–1415
Saliendra NZ, Meinzer FC, Perry M, Thom M (1996) Associations
between partitioning of carboxylase activity and bundle sheath
leakiness to CO2, carbon isotope discrimination, photosynthesis
and growth in sugarcane. J Exp Bot 47:907–914
Shabala L, Cuin TA, Newman IA, Shabala S (2005) Salinity-induced
ion flux patterns from the excised roots of Arabidopsis sos
mutants. Planta 222:1041–1050
Silveira JAG, Araujo SAM, Lima GPMS, Viegas RA (2009) Roots
and leaves display contrasting osmotic adjustment mechanisms
in response to NaCl-salinity in Atriplex nummularia. Environ
Exp Bot 66:1–8
Veselov DS, Sharipova GV, Veselov SU, Kudoyarova GR (2008) The
effects of NaCl treatment on water relations, growth, and ABA
content in barley cultivars differing in drought tolerance. J Plant
Growth Regul 27:380–386
Zhang HX, Hodson J, Williams JP, Blumwald E (2001) Engineering
salt tolerant Brassica plants: characterization of yield and seed
oil quality in transgenic plants with increased vacuolar Na
accumulation. Proc Natl Acad Sci USA 98:12832–12836
Zhu JK (2001) Plant salt tolerance. Trends Plant Sci 6:66–71
Zhu JK (2003) Regulation of ion homeostasis under salt stress. Curr
Opin Plant Biol 6:441–445
Zhu J, Meinzer FC (1999) Efficiency of C4 photosynthesis in Atriplexlentiformis under salinity stress. Aust J Plant Physiol 26:79–86
123
Acta Physiol Plant (2011) 33:547–557 557




















