
Differential Requirement of ZAP-70 for CD2-Mediated Activation Pathways of Mature Human T Cells
7
of June 13, 2013. This information is current as Mature Human T Cells CD2-Mediated Activation Pathways of Differential Requirement of ZAP-70 for Pirzer, Luis Barata and Claire Hivroz Edgar Meinl, Doris Lengenfelder, Norbert Blank, Rainer http://www.jimmunol.org/content/165/7/3578 2000; 165:3578-3583; ; J Immunol References http://www.jimmunol.org/content/165/7/3578.full#ref-list-1 , 17 of which you can access for free at: cites 34 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2000 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on June 13, 2013 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Differential Requirement of ZAP-70 for CD2-Mediated Activation Pathways of Mature Human T Cells

of June 13, 2013.This information is current as
Mature Human T CellsCD2-Mediated Activation Pathways of Differential Requirement of ZAP-70 for
Pirzer, Luis Barata and Claire HivrozEdgar Meinl, Doris Lengenfelder, Norbert Blank, Rainer
http://www.jimmunol.org/content/165/7/35782000; 165:3578-3583; ;J Immunol
Referenceshttp://www.jimmunol.org/content/165/7/3578.full#ref-list-1
, 17 of which you can access for free at: cites 34 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

Differential Requirement of ZAP-70 for CD2-MediatedActivation Pathways of Mature Human T Cells1
Edgar Meinl,2*† Doris Lengenfelder,* Norbert Blank,‡ Rainer Pirzer,* Luis Barata, § andClaire Hivroz ¶
This study addresses the role of the tyrosine kinase ZAP-70 in CD2-mediated T cell activation. Patients lacking ZAP-70 have fewmature CD81 T cells and high numbers of CD41 T cells that are nonfunctional upon TCR triggering. Such a patient with ahomozygous deletion in thezap-70gene that resulted in the complete absence of ZAP-70 protein expression has been identified.Expression of the tyrosine kinases Lck, Fyn, and Syk was normal. The patient’s T cells were activated with two different pairs ofmitogenic mAbs. CD2-induced phosphorylation of thez-chain and influx of Ca21 was defective in the ZAP-70-deficient T cells,whereas CD2-induced phosphorylation of several other proteins, including Syk, was not affected. CD2-induced proliferation aswell as production of TNF-a and IFN-g was abrogated in ZAP-70-deficient T cells, whereas PMA plus ionomycin induced normalactivation of these cells. Together, this study shows that CD2-activation triggers ZAP-70-dependent and -independent pathways.Deletion of ZAP-70 affected CD2- and CD3-mediated proliferation and cytokine production in a similar way, suggesting that oneof the different CD2 pathways converges with a CD3 pathway at or upstream of the activation of ZAP-70.The Journal ofImmunology,2000, 165: 3578–3583.
CD2 is expressed on the surface of thymocytes, mature Tcells, and NK cells. Its ligand in humans, CD58, is foundon many different cell types. CD2-CD58 interactions
strengthen the binding to APCs, increase the efficiency of Ag rec-ognition (1), and induce cytoskeletal polarization of T cells in con-tact with APCs (2). Engagement of CD2 also augments the T cellresponse to cytokines (3). Simultaneous triggering of two distinctepitopes on CD2 by mAbs induces T cells to proliferate and se-crete lymphokines in the absence of Ag and APCs. This has beenreferred to as the alternative pathway of T cell activation (4). Thepossibility to induce anergy (5), apoptosis (6), or even to reverseanergy (7) indicates potent effects of CD2.
T cell activation via CD2 or CD3 triggers a cascade of signalingevents, which seem to be very similar (8, 9). In some studies,stimulation via CD2 or CD3 induced tyrosine phosphorylation ofindistinguishable patterns of polypeptides (10). Despite these sim-ilarities, a number of differences between these two pathways havealso been described (11–13).
Optimal CD2-mediated signaling in T cells or NK cells requiresthe presence of thez-chain, which is a component of both the CD3complex and the CD16 complex on NK cells (14, 15). The precise
molecular mechanism through which thez-chain mediates CD2signaling is unknown. Upon TCR stimulation, thez-chain interactswith the Src-family tyrosine kinases Lck and Fyn, becomes phos-phorylated on its immunoreceptor tyrosine-based activation motifs(ITAM), 3 and recruits the Syk-family protein tyrosine kinase(PTK) ZAP-70 (16). CD2 stimulation also induces activation ofLck (17) and phosphorylation ofz (8). However, several studieshave led to conflicting data regarding the involvement ofz andZAP-70 in CD2-induced signaling: CD2-mediated signaling isalso possible in TCR-positive T cells that lackz-chain-specificITAMs (18). Evidence has been presented that stimulation viaCD2 can bypass tyrosine phosphorylation and recruitment ofZAP-70 to the CD3-TCR complex for induction of phospholipaseC-g1 activation (19) and activation of mitogen-activated proteinkinases (20).
ZAP-70 is solely expressed in thymocytes, T cells, and NK cellsand therefore shows a pattern of expression similar to CD2.ZAP-70 is essential for TCR-mediated activation of mature T cells,but it also plays a role in T cell maturation (16). Lack of ZAP-70expression leads to severe T cell immunodeficiency (21–23). ZAP-70-deficient patients have very few CD81 T cells in the periphery,whereas the number of peripheral CD41 T cells is increased. How-ever, these CD41 T cells are nonfunctional, showing no prolifer-ation in response to TCR triggering. Remarkably, lack of ZAP-70affects T cell maturation differently in humans and in mice. InZAP-702/2 mice, T cell maturation is arrested at the double-pos-itive stage of differentiation, and, therefore, no mature CD41 orCD81 T cells are found in the periphery (24).
The aim of this study was to analyze the role of ZAP-70 inCD2-mediated activation of mature T cells. We have identified apatient with a severe immunodeficiency due to a homozygous de-letion in thezap-70gene leading to a lack of expression of ZAP-70protein. Analysis of mature T cells from this patient revealed adifferential requirement of ZAP-70 for CD2-mediated signaling
*Institute for Clinical and Molecular Virology, University Erlangen-Nurnberg,Erlangen, Germany;†Department of Neuroimmunology, Max-Planck-Institute ofNeurobiology, Martinsried, Germany and Institute for Clinical Neuroimmunology,Ludwig-Maximilians-University, Munich, Germany;‡Department of Internal Medi-cine III and Institute for Clinical Immunology, University of Erlangen-Nurnberg,Erlangen, Germany;§Primary Immunodeficiencies Unit, Department of Allergy andClinical Immunology, Coimbra Paediatric Hospital, Coimbra, Portugal; and¶InstitutNational de la Sante et de la Recherche Medicale Unit 520, Institut Curie, Paris,France
Received for publication February 25, 2000. Accepted for publication July 6, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Deutsche Forschungsgemeinschaft (SFB 466) andby the Wilhelm Sander-Stiftung (97.081.1). The Institute for Clinical Neuroimmu-nology is supported by the Hermann and Lilly Schilling Foundation.2 Address correspondence and reprint requests to Dr. Edgar Meinl, Department ofNeuroimmunology, Max-Planck-Institute of Neurobiology, D-82152 Martinsried,Germany. E-mail address: [email protected]
3 Abbreviations used in this paper: ITAM, immune receptor tyrosine-based activationmotif; PTK, protein tyrosine kinase.
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

events: ZAP-70 was absolutely required for CD2-mediated Ca21
influx, for persistent phosphorylation ofz, for proliferation, andcytokine production. However, several proteins, including Syk,were phosphorylated after CD2 activation in the patient’s T cells,indicating that CD2 also induces ZAP-70-independent signalingevents.
Materials and MethodsCase report
The patient described in this study, a second child of unrelated parents,developed pneumonia at 3 mo of age due toPneumocystis cariniiand, at6 mo, developed an extensive varicella infection. Severe combined immu-nodeficiency was diagnosed at the age of 7 mo. The lymphocyte bloodcount was high (15,000–20,000 cells/ml). Less than 1% of the CD31 cells,either TCRab or TCRgd, expressed CD8. B cells and NK cells were foundat a normal proportion.
zap-70 sequencing and segregation of the mutation
Total RNA and cDNA from the PBMC of the patient and from a normaldonor and genomic DNA of the patient, her parents, and a control wereprepared. Amplification of cDNAs was performed by PCR usingzap-70-specific pairs of primers. The upper primer, 59-TTTGCCTGGACATCCACCTGTACGTCC-39, is located upstream of the ATG, and the lowerprimer, 59-CAGCTGTGTGTGGAGACAACCAA-39, binds 39of the stopcodon. PCR products were subcloned into the pGemTvector before DNAsequencing using the Thermosequenase kit (Amersham, Braunschweig, Ger-many). Four independent clones were sequenced. Analysis of the segregationof the mutation was performed on genomic DNA using one intronic primerupstream of the deletion, 59-CCTGATCCAGCAGCATCTCCC-39, and oneexonic primer downstream of the deletion, 59-CTTGCCCTGCTCGATGAAGGC-39. PCR were conducted in the presence of 2mCi of [a-32P]dCTPper reaction. Samples were separated using a 6% denaturing PAGE gel andexposed to autoradiography. The experiments determining the segregation ofthe mutation were done three times.
T cell culture and activation
PBMC from the ZAP-70-deficient patient and two healthy donors werestimulated with 10 ng/ml PMA (Sigma, Deisenhofen, Germany) and 1mg/ml ionomycin (Sigma). Culture medium consisted of 45% Panserin 401(Pan, Aidenbach, Germany), 45% RPMI 1640, and 10% FCS (BoehringerMannheim, Mannheim, Germany) supplemented with 2 mM glutamine and50 mg/ml gentamicin (Life Technologies, Berlin, Germany). Recombinanthuman IL-2 (Chiron, Ratingen, Germany) was added at a concentration of100 U/ml 2 days later. The growing cells were expanded in the IL-2-containing medium for at least 2 wk.
Flow cytometry
mAbs directed to CD3, CD4, CD8, TCRab, TCRgd, HLA-DR, and la-beled isotype controls were obtained from Becton Dickinson (Heidelberg,Germany). CD2 expression was detected with supernatant of the hybrid-oma TS2/18.1.1 (American Type Culture Collection, Manassas, VA) and aFITC-labeled goat-anti-mouse-IgG F(ab9)2 fragment (Dianova, Hamburg,Germany). Each surface marker was analyzed two to three times.
Activation via CD2 and CD3, assessment of proliferation, andcytokine production
For CD2-mediated activation, two different pairs of mAbs were used: ei-ther a combination of the mAb 39C1.5, which recognizes the T11.1epitope, and the mAb 6F10.3, which recognizes the T11.2 epitope (bothfrom Coulter Immunotech, Hamburg, Germany), or a combination of themAb X11, which recognizes the T11.1 epitope of CD2, and the mAb D66,which recognizes the CD2R cryptic epitope (kindly provided by Dr. L.Boumsel, INSERM U448, Créteil, France). For CD3-mediated activation,96-well U-bottom plates were coated overnight with 1mg/ml of purifiedmAb OKT3. Additionally PMA (10 ng/ml) plus ionomycin (1mg/ml) wereused to activate T cells. About 73 104 T cells were seeded per well in avolume of 200ml medium without IL-2. All experiments were performedin triplicates. The supernatant was collected 24 h after activation. Produc-tion of IFN-g and TNF-a was determined by ELISA. Two days after ac-tivation, 0.2 mCi of [3H]thymidine (Amersham) was added for another16 h. Cultures were harvested using a Matrix TM96 (Packard, Frankfurt,Germany). The filters were dried, exposed overnight on a [3H]thymidine-sensitive screen, and analyzed with a BAS2000 imaging system (Fuji Ray-
test, Straubenhardt, Germany). Comparative measurements of the samefilters with the direct beta-counter (Packard) and the BAS2000 showed thatthese two evaluation systems had a correlation of.0.95 and a comparablesensitivity. Cpm values obtained with the direct beta-counter are about20% of those obtained with a liquid scintillation counter. To analyze cy-tokine production and proliferation after CD2 and CD3 activation, threeindependent experiments were performed with two ZAP-70-expressingcontrol cell lines included in each experiment.
Immunoprecipitation and Western blot analysis
For detection of ZAP-70, two different Abs were applied, a mAb (no. Z24820; Transduction Laboratories, Lexington, KY) and a polyclonal rabbitAb (kindly provided by Dr. B. Malissen, Institut National de la Santé et dela Recherche Médicale-Centre National de la Recherche Scientifique, Mar-seille, France). Syk was detected with a mAb (no. 05-434) from UpstateBiotechnology (Biozol, Eching, Germany) or a polyclonal Ab (Santa CruzBiotechnology, Santa Cruz, CA), p56lck and p59fyn were detected withpolyclonal Abs (Santa Cruz Biotechnology). Cleared cell lysates were sep-arated under reducing conditions on an 8% SDS-PAGE and electroblottedon Immobilon P membrane (Millipore, Eschborn, Germany). Blots wereblocked with PBS containing 5% low-fat milk and 0.05% Tween 20, in-cubated with 1mg/ml of the primary mAbs, a peroxidase-conjugated sec-ondary Ab, and developed by using the ECL detection system (Amer-sham). Immunoprecipitation for ZAP-70 and Syk was performed fourtimes.
Purified primary T cells, obtained by rosetting with neuraminidase-treated sheep RBC or T cell blasts were left unactivated or activated for 3min at 37°C in the presence of UCHT1 (ascitic fluid at 1/1000) or a com-bination of the mAbs X11 and D66, each used at 10mg/ml. Cells werelysed in lysis buffer (20 mM Tris-HCl, pH 7.4, 140 mM NaCl, 2 mMEDTA, 50 mM NaF, 1% Nonidet P-40, 0.5% Na DOC, 0.1% SDS, 100mMNa3VO4, protease inhibitors) for 20 min at 4°C. Nuclei and cell debris wereremoved by centrifugation. Protein concentrations were determined in thepostnuclear lysates using a Bio-Rad kit (Bio-Rad, Richmond, U.K.). Thesame amount of lysates was precleared at 4°C by rocking with mouse orrabbit purified IgG for 1 h at 4°C. Then, protein G-Sepharose beads wereadded, and the nonspecific immunoprecipitates were recovered by centrif-ugation. After this preclearing, lysates were incubated overnight with anti-Syk Abs (Santa Cruz Biotechnology; sc-1077), anti-z mAb (Santa CruzBiotechnology; sc-1239), or anti-ZAP-70 Abs (polyclonal rabbit anti-serum, kind gift from Dr. B. Malissen). Specific immunoprecipitates wererecovered by addition of protein G-Sepharose beads for 1 h and washedthree times in lysis buffer. Immunoprecipitates were then run on standardSDS-PAGE, and transferred to a polyvinylidene difluoride membrane (Im-mobilon-P; Millipore). Nonspecific binding was blocked with 5% BSA inPBS/0.05% Tween, then the anti-phosphotyrosine mAb 4G10, an anti-SykmAb (Upstate Biotechnology), an anti-z mAb, or anti-ZAP-70 Abs (SantaCruz Biotechnology; sc-574) were applied. The Ab/Ag complexes werevisualized by an enhanced chemiluminescence detection system accordingto the manufacturer’s instruction (ECL; Amersham) using anti-mouse oranti-rabbit Ig Abs coupled to HRP as secondary Abs. Experiments detect-ing activation induced tyrosine phosphorylation and specific phosphoryla-tion of Syk andz were performed twice.
Measurement of Ca21 influx
The procedure of measuring [Ca21]i was adapted from previously de-scribed protocols (25, 26). In brief, 13 107 cells were incubated in 1 mlof their culture medium with Fluo3-AM (4mM) on an agitator for 30 minat room temperature. To facilitate dye equilibration, 4 ml culture mediumwas added and cells were incubated for another 30 min (27). Rapid dyeexclusion was prevented by the use of sulfinpyrazone (250mM) in eachsolution. After the loading procedure was completed, cells were split insamples of 13 106 cells for each experiment, spun down, and resuspendedin a simplified saline solution containing 1 mM Ca21. The fluorescence ofthe cells was detected in a PTI (South Brunswick, NJ) spectrometer at 37°Cunder constant stirring. The excitation wavelength was 505 nm and emis-sion wavelength was 530 nm. Experiments were started when the baselinewas constant over 2–3 min. TheKd of Fluo3-AM for Ca21 binding at 37°Cwas 864 nM as was described (26) and cytosolic Ca21 was calculated as[Ca21]i 5 Kd (F 2 Fmin)/(Fmax 2 F ). The fluorescence maximum (Fmax)was calibrated using Ionomycin (5mM). A control sample of 13 106 cellswas not incubated with dye to determine autofluorescence (Fmin) (26). Theexperiments determining Ca influx were performed three times.
3579The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

ResultsMutation in thezap-70gene and expression of tyrosine kinasesin the patient’s T cells
Immunological findings in this child were reminiscent of a defectin ZAP-70 (23) and prompted us to study the coding region ofzap-70. We amplified the complete coding sequence for ZAP-70from cDNA and sequenced independent clones. A 13-bp deletioninvolving nucleotides 1719–1731 was found (Fig. 1,top). Thismutation has already been detected in another, unrelated patientand has been shown to induce instability of the protein (28). Seg-regation analysis revealed that the patient was omozygous, but theparents were heterozygous for this mutation (Fig. 1,bottom).
This mutation leads to a total absence of ZAP-70 protein ex-pression as shown on immunoprecipitates of total lysates (Fig. 2).The absence of ZAP-70 was seen with a commercially availablemAb and also with a polyclonal anti-ZAP-70 Ab (a kind gift of Dr.B. Malissen). Levels of expression of Syk, p56lck, and p59fyn werecompared in postnuclear lysates of CD41 T cells from a controland from the patient. As shown in Fig. 2, these three tyrosinekinases were expressed at the same level in both T cellpopulations.
Culture and phenotype of ZAP-70-defective T cells
PBMC from the patient described above were stimulated withPMA plus ionomycin and cultured with IL-2. Under these condi-tions, the proliferative response of the patient’s T cells was similar
to the one observed with control T cells. After at least 2 wk of culture,these cells had the phenotype of mature activated T cells. They werelargely CD41CD82 (71%), but contained a CD42CD82 (15%) anda CD81CD42 (13%) subpopulation. Activation markers such asCD69, CD25, and HLA-DR were expressed. The outgrowing cellswere homogeneously (98%) positive for CD3, 81% expressed anabTCR, whereas agd TCR was found on 18%. CD2 was strongly ex-pressed, as on T cells from healthy controls (Fig. 3).
FIGURE 1. Determination of thezap-70mutation and its segregation.Top, Sequence of clonedzap-70cDNA obtained by RT-PCR from thepatient’s mRNA or a control. A deletion of 13 bp between codons 1718 and1730 was observed in two independent cDNA clones from the patient.Bottom, An informative region of thezap-70gene of the patient, of theparents, and of an unrelated control was amplified in the presence of[a-32P]dCTP. Radioactive products were separated by PAGE and exposedto autoradiography. WT, Wild type.
FIGURE 2. Tyrosine kinase expression in patients’ T cells.A, ZAP-70was precipitated from lysates obtained from control CD41 T lymphocytesor patient T lymphocytes. The immunoprecipitated ZAP-70 was revealedby Western blot analysis using a ZAP-70 polyclonal Ab.B, Expression ofZAP-70 in total cell lysates was analyzed by Western blot with a commer-cially available mAb. Postnuclear lysates of T cells from a control or fromthe patient were also analyzed by Western blot with an anti-Syk mAb (C),anti-p56lck (D), or anti-p59fyn (E). Bands corresponding to Syk, p56lck,p59fyn, or the heavy chains of the ZAP-70 immunoprecipitating Ab (IgH)are indicated by arrows.
FIGURE 3. Phenotype of cultured ZAP-70-deficient T cells. T cellblasts from the ZAP-70-deficient patient were activated with PMA plusionomycin and expanded in IL-2 for 2 wk. In the histograms, the open linerepresents the isotype control and the closed graph represents the specificstaining for the indicated marker. The number below the marker gives thepercentage of cells scored positive. In the dot blot graph, the lower leftquadrant represents the negative control. The numbers in each quadrantrepresent the percentage of positive cells.
3580 ZAP-70 AND CD2-MEDIATED T CELL ACTIVATION
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

Lack of ZAP-70 affects CD2 signaling
To study the role of ZAP-70 in CD2 signaling, we activated T cellblasts obtained from the patient described herein with a combina-tion of the CD2 mAbs X11 and D66. We first analyzed the proteintyrosine phosphorylation in total cell lysates induced by triggeringof CD3/TCR or CD2. As shown in Fig. 4A, CD3 and CD2 acti-vation led to tyrosine phosphorylation of several proteins in T cellblasts from both the control and the ZAP-70-deficient patient. Thebasal level and CD3- or CD2-induced tyrosine phosphorylationswere reproducibly slightly reduced in the patient’s T cell blasts,but the pattern of tyrosine phosphorylated proteins was grosslysimilar in normal and ZAP-70-deficient blasts. However, we con-sistently noted that a phosphorylated band around 21 kDa inducedby both CD3 and CD2 was absent in the ZAP-70-deficient blasts.Because this molecular mass was reminiscent of one of the phos-phorylated forms ofz, we immunoprecipitatedz and studied itsphosphorylation status in quiescent or activated T cells. As shownin Fig. 4C, we observed tyrosine phosphorylatedz in the quiescentcontrol cells; this phosphorylation was enhanced by CD3 or CD2triggering. In marked contrast, we did not see anyz phosphoryla-tion, neither in quiescent nor in activated blasts from the patient.
We then analyzed whether the lack of ZAP-70 affects the acti-vation-induced phosphorylation of Syk. To this end, we immuno-precipitated Syk in unstimulated, CD2- or CD3-activated T cellblasts. As shown in Fig. 4B, the level of Syk expression was thesame in the patient’s and control’s cells. Analysis of the tyrosinephosphorylation status of Syk revealed that CD3 and CD2 activa-tion induced Syk phosphorylation in both normal and ZAP-70-deficient T cells. Finally, we measured the activation-inducedCa21 influx. These experiments showed that ZAP-70 was essentialfor CD2-mediated Ca21 influx (Fig. 5).
Lack of ZAP-70 affects CD2-mediated proliferation and cytokineproduction
Activation via CD2 triggered normal T cells to produce TNF-aand IFN-g, whereas the ZAP-70-deficient T cells showed no re-sponse (Fig. 6). Cells were also stimulated with solid-phase boundanti-CD3 and with PMA plus ionomycin. Although ZAP-70-defi-
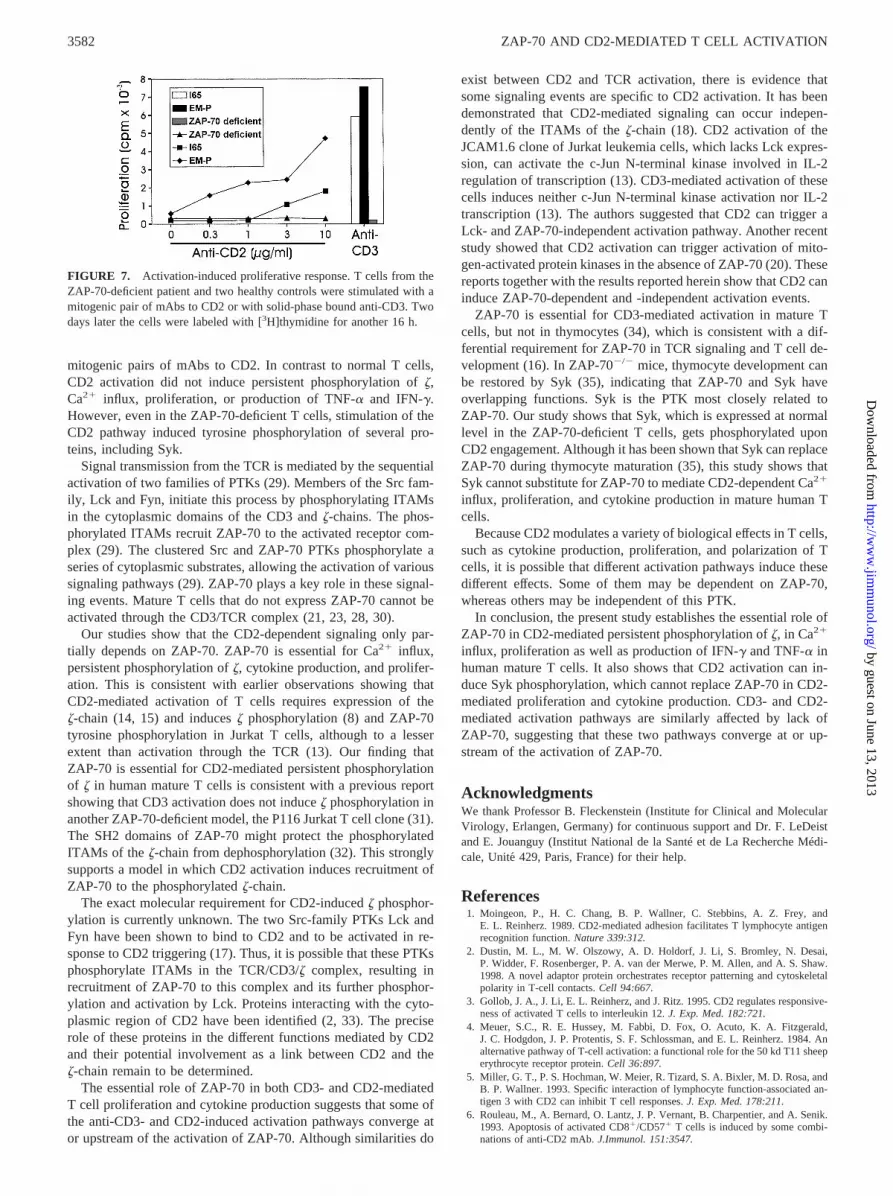
cient T cells did not respond to activation via CD3, stimulationwith PMA plus ionomycin induced IFN-g and TNF-a secretion.These results confirm that ZAP-70-deficient T cells can be acti-vated by signals bypassing the TCR membrane signaling (Fig. 6).Measurement of the proliferative response induced by CD2 trig-gering showed that no proliferation of the ZAP-70-deficient T cellscould be induced in contrast to what was observed with T cellsobtained from two control donors (Fig. 7). The ZAP-70-deficientcells did not show any proliferative response at any concentrationof anti-CD2 mAbs. By contrast, at 10mg/ml of anti-CD2 mAbs,the control cells EM-P gave a stimulation index of 8, the controlcells I-65, which were triggered to a lower proliferation and had alower background of spontaneous proliferation, displayed a stim-ulation index of 9 (Fig. 7).
DiscussionThis study shows that ZAP-70 has a differential effect on differentsignaling pathways activated upon CD2-engagement. We haveidentified a patient with a homozygous deletion in the gene codingfor ZAP-70 resulting in complete lack of ZAP-70 protein expres-sion. T cells from this patient were stimulated with two different
FIGURE 6. Activation-induced secretion of IFN-g and TNF-a. Cul-tured short-term T cell lines from the ZAP-70-deficient patient and twohealthy controls (EM-P and I65) were stimulated with a mitogenic pair ofanti-CD2 mAbs (Aand C), solid-phase bound anti-CD3, or PMA plusionomycin (Band D). Supernatants were collected 24 h after activation,and the amount of secreted IFN-g or TNF-a was determined.
FIGURE 4. Tyrosine phosphorylation induced by CD2 activation. Tcell blasts from the ZAP-70-deficient patient and one healthy control wereleft unstimulated (0) or activated by the combination of anti-CD2 mAbsX11 and D66 (CD2) or by the anti-CD3 mAb UCHT1 (CD3) for 3 min. at37°C, before lysis and immunoprecipitation with either anti-Syk or anti-z.Postnuclear lysates (A) or immune complexes (BandC) were resolved bySDS-PAGE. Subsequently, a Western blot analysis (WBA) was performedwith a mAb directed to phosphotyrosine (4G10), Syk, orz. Bands corre-sponding to Syk,z, phospho-z (P z), or the heavy chain of the immuno-precipitating Ab (IgH) are indicated by arrows.
FIGURE 5. Activation-induced [Ca21]i. T cell blasts from a control (Aand C) and the ZAP-70-deficient patient (Band D) were labeled withFluo3-AM and analyzed for [Ca21]i after CD2 stimulation using 1–1.53106 cells per cuvette in a volume of 1 ml. Two different pairs of mAbs wereused. Arrows in each figure indicate the addition of Abs.A andB, The AbsX11 (dilution 1:50), D66 (dilution 1:50), and goat-anti-mouse Ig (10mg/ml, dilution 1:150 in 6.6ml) were used.C andD, The Abs 6F10.3 (4mg/ml,dilution 1:50), 39C1.5 (4mg/ml, dilution 1:50), and goat-anti-mouse-Ig (10mg/ml, dilution 1:150) were added. Addition of 20ml solution with Abs(dilution 1:50) induced a drop of the baseline due to dilution of the buffersolution.
3581The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
mitogenic pairs of mAbs to CD2. In contrast to normal T cells,CD2 activation did not induce persistent phosphorylation ofz,Ca21 influx, proliferation, or production of TNF-a and IFN-g.However, even in the ZAP-70-deficient T cells, stimulation of theCD2 pathway induced tyrosine phosphorylation of several pro-teins, including Syk.
Signal transmission from the TCR is mediated by the sequentialactivation of two families of PTKs (29). Members of the Src fam-ily, Lck and Fyn, initiate this process by phosphorylating ITAMsin the cytoplasmic domains of the CD3 andz-chains. The phos-phorylated ITAMs recruit ZAP-70 to the activated receptor com-plex (29). The clustered Src and ZAP-70 PTKs phosphorylate aseries of cytoplasmic substrates, allowing the activation of varioussignaling pathways (29). ZAP-70 plays a key role in these signal-ing events. Mature T cells that do not express ZAP-70 cannot beactivated through the CD3/TCR complex (21, 23, 28, 30).
Our studies show that the CD2-dependent signaling only par-tially depends on ZAP-70. ZAP-70 is essential for Ca21 influx,persistent phosphorylation ofz, cytokine production, and prolifer-ation. This is consistent with earlier observations showing thatCD2-mediated activation of T cells requires expression of thez-chain (14, 15) and inducesz phosphorylation (8) and ZAP-70tyrosine phosphorylation in Jurkat T cells, although to a lesserextent than activation through the TCR (13). Our finding thatZAP-70 is essential for CD2-mediated persistent phosphorylationof z in human mature T cells is consistent with a previous reportshowing that CD3 activation does not inducez phosphorylation inanother ZAP-70-deficient model, the P116 Jurkat T cell clone (31).The SH2 domains of ZAP-70 might protect the phosphorylatedITAMs of the z-chain from dephosphorylation (32). This stronglysupports a model in which CD2 activation induces recruitment ofZAP-70 to the phosphorylatedz-chain.
The exact molecular requirement for CD2-inducedz phosphor-ylation is currently unknown. The two Src-family PTKs Lck andFyn have been shown to bind to CD2 and to be activated in re-sponse to CD2 triggering (17). Thus, it is possible that these PTKsphosphorylate ITAMs in the TCR/CD3/z complex, resulting inrecruitment of ZAP-70 to this complex and its further phosphor-ylation and activation by Lck. Proteins interacting with the cyto-plasmic region of CD2 have been identified (2, 33). The preciserole of these proteins in the different functions mediated by CD2and their potential involvement as a link between CD2 and thez-chain remain to be determined.
The essential role of ZAP-70 in both CD3- and CD2-mediatedT cell proliferation and cytokine production suggests that some ofthe anti-CD3- and CD2-induced activation pathways converge ator upstream of the activation of ZAP-70. Although similarities do
exist between CD2 and TCR activation, there is evidence thatsome signaling events are specific to CD2 activation. It has beendemonstrated that CD2-mediated signaling can occur indepen-dently of the ITAMs of thez-chain (18). CD2 activation of theJCAM1.6 clone of Jurkat leukemia cells, which lacks Lck expres-sion, can activate the c-Jun N-terminal kinase involved in IL-2regulation of transcription (13). CD3-mediated activation of thesecells induces neither c-Jun N-terminal kinase activation nor IL-2transcription (13). The authors suggested that CD2 can trigger aLck- and ZAP-70-independent activation pathway. Another recentstudy showed that CD2 activation can trigger activation of mito-gen-activated protein kinases in the absence of ZAP-70 (20). Thesereports together with the results reported herein show that CD2 caninduce ZAP-70-dependent and -independent activation events.
ZAP-70 is essential for CD3-mediated activation in mature Tcells, but not in thymocytes (34), which is consistent with a dif-ferential requirement for ZAP-70 in TCR signaling and T cell de-velopment (16). In ZAP-702/2 mice, thymocyte development canbe restored by Syk (35), indicating that ZAP-70 and Syk haveoverlapping functions. Syk is the PTK most closely related toZAP-70. Our study shows that Syk, which is expressed at normallevel in the ZAP-70-deficient T cells, gets phosphorylated uponCD2 engagement. Although it has been shown that Syk can replaceZAP-70 during thymocyte maturation (35), this study shows thatSyk cannot substitute for ZAP-70 to mediate CD2-dependent Ca21
influx, proliferation, and cytokine production in mature human Tcells.
Because CD2 modulates a variety of biological effects in T cells,such as cytokine production, proliferation, and polarization of Tcells, it is possible that different activation pathways induce thesedifferent effects. Some of them may be dependent on ZAP-70,whereas others may be independent of this PTK.
In conclusion, the present study establishes the essential role ofZAP-70 in CD2-mediated persistent phosphorylation ofz, in Ca21
influx, proliferation as well as production of IFN-g and TNF-a inhuman mature T cells. It also shows that CD2 activation can in-duce Syk phosphorylation, which cannot replace ZAP-70 in CD2-mediated proliferation and cytokine production. CD3- and CD2-mediated activation pathways are similarly affected by lack ofZAP-70, suggesting that these two pathways converge at or up-stream of the activation of ZAP-70.
AcknowledgmentsWe thank Professor B. Fleckenstein (Institute for Clinical and MolecularVirology, Erlangen, Germany) for continuous support and Dr. F. LeDeistand E. Jouanguy (Institut National de la Santé et de La Recherche Médi-cale, Unite 429, Paris, France) for their help.
References1. Moingeon, P., H. C. Chang, B. P. Wallner, C. Stebbins, A. Z. Frey, and
E. L. Reinherz. 1989. CD2-mediated adhesion facilitates T lymphocyte antigenrecognition function.Nature 339:312.
2. Dustin, M. L., M. W. Olszowy, A. D. Holdorf, J. Li, S. Bromley, N. Desai,P. Widder, F. Rosenberger, P. A. van der Merwe, P. M. Allen, and A. S. Shaw.1998. A novel adaptor protein orchestrates receptor patterning and cytoskeletalpolarity in T-cell contacts.Cell 94:667.
3. Gollob, J. A., J. Li, E. L. Reinherz, and J. Ritz. 1995. CD2 regulates responsive-ness of activated T cells to interleukin 12.J. Exp. Med. 182:721.
4. Meuer, S.C., R. E. Hussey, M. Fabbi, D. Fox, O. Acuto, K. A. Fitzgerald,J. C. Hodgdon, J. P. Protentis, S. F. Schlossman, and E. L. Reinherz. 1984. Analternative pathway of T-cell activation: a functional role for the 50 kd T11 sheeperythrocyte receptor protein.Cell 36:897.
5. Miller, G. T., P. S. Hochman, W. Meier, R. Tizard, S. A. Bixler, M. D. Rosa, andB. P. Wallner. 1993. Specific interaction of lymphocyte function-associated an-tigen 3 with CD2 can inhibit T cell responses.J. Exp. Med. 178:211.
6. Rouleau, M., A. Bernard, O. Lantz, J. P. Vernant, B. Charpentier, and A. Senik.1993. Apoptosis of activated CD81/CD571 T cells is induced by some combi-nations of anti-CD2 mAb.J.Immunol. 151:3547.
FIGURE 7. Activation-induced proliferative response. T cells from theZAP-70-deficient patient and two healthy controls were stimulated with amitogenic pair of mAbs to CD2 or with solid-phase bound anti-CD3. Twodays later the cells were labeled with [3H]thymidine for another 16 h.
3582 ZAP-70 AND CD2-MEDIATED T CELL ACTIVATION
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

7. Boussiotis, V. A., G. J. Freeman, J. D. Griffin, G. S. Gray, J. G. Gribben, andL. M. Nadler. 1994. CD2 is involved in maintenance and reversal of humanalloantigen-specific clonal anergy.J. Exp. Med. 180:1665.
8. Monostori, E., D. Desai, M. H. Brown, D. A. Cantrell, and M. J. Crumpton. 1990.Activation of human T lymphocytes via the CD2 antigen results in tyrosine phos-phorylation of T cell antigen receptorz-chains.J. Immunol. 144:1010.
9. Kanner, S. B., N. K. Damle, J. Blake, A. Aruffo, and J. A. Ledbetter. 1992.CD2/LFA-3 ligation induces phospholipase-Cg1 tyrosine phosphorylation andregulates CD3 signaling.J. Immunol. 148:2023.
10. Ley, S. C., A. A. Davies, B. Druker, and M. J. Crumpton. 1991. The T cellreceptor/CD3 complex and CD2 stimulate the tyrosine phosphorylation of indis-tinguishable patterns of polypeptides in the human T leukemic cell line Jurkat.Eur. J. Immunol. 21:2203.
11. Hubert, P., P. Debre, L. Boumsell, and G. Bismuth. 1993. Tyrosine phosphory-lation and association with phospholipase Cg-1 of the GAP-associated 62-kDprotein after CD2 stimulation of Jurkat T cell.J. Exp. Med. 178:1587.
12. Jin, Y. J., D. R. Kaplan, M. White, G. C. Spagnoli, T. M. Roberts, andE. L. Reinherz. 1990. Stimulation via CD3-Ti but not CD2 induces rapid tyrosinephosphorylation of a 68-kDa protein in the human Jurkat T cell line.J. Immunol.144:647.
13. Sunder-Plassmann, R., and E. L. Reinherz. 1998. A p56lck-independent pathwayof CD2 signaling involves Jun kinase.J. Biol. Chem. 273:24249.
14. Spruyt, L. L., M. J. Glennie, A. D. Beyers, and A. F. Williams. 1991. Signaltransduction by the CD2 antigen in T cells and natural killer cells: requirement forexpression of a functional T cell receptor or binding of antibody Fc to the Fcreceptor, Fc gamma RIIIA (CD16).J. Exp. Med. 174:1407.
15. Howard, F. D., P. Moingeon, U. Moebius, D. J. McConkey, B. Yandava,T. E. Gennert, and E. L. Reinherz. 1992. The CD3z cytoplasmic domain medi-ates CD2-induced T cell activation.J. Exp. Med. 176:139.
16. Chu, D. H., C. T. Morita, and A. Weiss. 1998. The Syk family of protein tyrosinekinases in T-cell activation and development.Immunol. Rev. 165:167.
17. Danielian, S., A. Alcover, L. Polissard, M. Stefanescu, O. Acuto, S. Fischer, andR. Fagard. 1992. Both T cell receptor (TcR)-CD3 complex and CD2 increase thetyrosine kinase activity of p56lck: CD2 can mediate TcR-CD3-independent andCD45-dependent activation of p56lck. Eur. J. Immunol. 22:2915.
18. Steeg, C., A. von Bonin, H. W. Mittrucker, B. Malissen, and B. Fleischer. 1997.CD2-mediated signaling in T cells lacking thez-chain-specific immune receptortyrosine-based activation (ITAM) motif.Eur. J. Immunol. 27:2233.
19. Hubert, P., V. Lang, P. Debre, and G. Bismuth. 1996. Tyrosine phosphorylationand recruitment of ZAP-70 to the CD3-TCR complex are defective after CD2stimulation.J. Immunol. 157:4322.
20. Ueno, H., S. Matsuda, K. Katamura, M. Mayumi, and S. Koyasu. 2000. ZAP-70is required for calcium mobilization but is dispensable for mitogen-activatedprotein kinase (MAPK) superfamily activation induced via CD2 in human Tcells.Eur. J. Immunol. 30:78.
21. Chan, A. C., T. A. Kadlecek, M. E. Elder, A. H. Filipovich, W. L. Kuo,M. Iwashima, T. G. Parslow, and A. Weiss. 1994. ZAP-70 deficiency in an
autosomal recessive form of severe combined immunodeficiency.Science 264:1599.
22. Arpaia, E., M. Shahar, H. Dadi, A. Cohen, and C. M. Roifman. 1994. DefectiveT cell receptor signaling and CD81 thymic selection in humans lackingzap-70kinase.Cell 76:947.
23. Hivroz, C., and A. Fischer. 1994. Immunodeficiency diseases: multiple roles forZAP-70.Curr. Biol. 4:731.
24. Negishi, I., N. Motoyama, K. Nakayama, S. Senju, S. Hatakeyama, Q. Zhang,A. C. Chan, and D. Y. Loh. 1995. Essential role for ZAP-70 in both positive andnegative selection of thymocytes.Nature 376:435.
25. Kao, J. P., A. T. Harootunian, and R. Y. Tsien. 1989. Photochemically generatedcytosolic calcium pulses and their detection by fluo-3.J. Biol. Chem. 264:8179.
26. Merritt, J. E., S. A. McCarthy, M. P. Davies, and K. E. Moores. 1990. Use offluo-3 to measure cytosolic Ca21 in platelets and neutrophils. Loading cells withthe dye, calibration of traces, measurements in the presence of plasma, and buff-ering of cytosolic Ca21. Biochem. J. 269:513.
27. Vandenberghe, P. A., and J. L. Ceuppens. 1990. Flow cytometric measurement ofcytoplasmic free calcium in human peripheral blood T lymphocytes with fluo-3,a new fluorescent calcium indicator.J. Immunol. Methods 127:197.
28. Elder, M. E., D. Lin, J. Clever, A. C. Chan, T. J. Hope, A. Weiss, andT. G. Parslow. 1994. Human severe combined immunodeficiency due to a defectin ZAP-70, a T cell tyrosine kinase.Science 264:1596.
29. Weiss, A., and D. R. Littman. 1994. Signal transduction by lymphocyte antigenreceptors.Cell 76:263.
30. Arpaia, E., M. Shahar, H. Dadi, A. Cohen, and C. M. Roifman. 1994. DefectiveT cell receptor signaling and CD81 thymic selection in humans lackingzap-70kinase.Cell 76:947.
31. Williams, B. L., K. L. Schreiber, W. Zhang, R. L. Wange, L. E. Samelson,P. J. Leibson, and R. T. Abraham. 1998. Genetic evidence for differential cou-pling of Syk family kinases to the T-cell receptor: reconstitution studies in aZAP-70-deficient Jurkat T-cell line.Mol. Cell Biol. 18:1388.
32. Chan, A. C., D. M. Desai, and A. Weiss. 1994. The role of protein tyrosinekinases and protein tyrosine phosphatases in T cell antigen receptor signal trans-duction.Annu. Rev. Immunol. 12:555.
33. Li, J., K. Nishizawa, W. An, R. E. Hussey, F. E. Lialios, R. Salgia,R. Sunder-Plassmann, and E. L. Reinherz. 1998. A cdc15-like adaptor protein(CD2BP1) interacts with the CD2 cytoplasmic domain and regulates CD2-trig-gered adhesion.EMBO J. 17:7320.
34. Gelfand, E. W., K. Weinberg, B. D. Mazer, T. A. Kadlecek, and A. Weiss. 1995.Absence of ZAP-70 prevents signaling through the antigen receptor on peripheralblood T cells but not on thymocytes.J. Exp. Med. 182:1057.
35. Gong, Q., L. White, R. Johnson, M. White, I. Negishi, M. Thomas, andA. C. Chan. 1997. Restoration of thymocyte development and function inzap-702/2 mice by the Syk protein tyrosine kinase.Immunity 7:369.
3583The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from




















