
Chemical denaturation studies of two isozymes of aryl ... - Uncg

ctssIitoaattmCnbiFmimtdiadcR
Ac
u
L
dPE
Experimental Cell Research 249, 131–146 (1999)Article ID excr.1999.4447, available online at http://www.idealibrary.com on
Differential Localization of Protein Kinase A Type II Isozymesin the Golgi–Centrosomal Area
Guy Keryer,*,1 Bjørn S. Skålhegg,† Brynjar F. Landmark,† Vidar Hansson,†Tore Jahnsen,† and Kjetil Tasken†,2
*Faculte des Sciences Pharmaceutiques et Biologiques, INSERM Unite 427, Universite Rene Descartes, F-75270 Paris Cedex 06, France;and †Institute of Medical Biochemistry, University of Oslo, N-0317 Oslo, Norway
hhtfco(moti(euspt
iisioiucWdkapa(lpm(C
atIam
Selectivity in the action of cAMP may be mediated byompartmentalized pools of cyclic AMP-dependent pro-ein kinase (PKA). PKA type II is directed to differentubcellular loci by interaction of the type II regulatoryubunits (RIIa, RIIb) with A-kinase anchoring proteins.n order to separately investigate the subcellular local-zation of PKA type II isozymes, monospecific antibodieso human RIIa and RIIb subunits of PKA were devel-ped. We demonstrate that centrosomes bind both RIIand RIIb. Centrosomes were the preferred intracellularnchoring site for RIIb. However, centrosomal localiza-ion of RIIb was observed only in some highly differen-iated cells such as keratinocytes, granulosa cells, andacrophages and in all neoplastic cell lines examined.entrosomal localization of RIIb was not observed inormal undifferentiated cells such as fibroblasts, myo-lasts, and T and B cells. In contrast, RIIa was abundantn the Golgi area and in the trans-Golgi network (TGN).urthermore, although RIIa appeared to colocalize withicrotubules in the Golgi/TGN, extractions with non-
onic detergent demonstrated that RIIa was mainlyembrane-associated. In addition, alterations of micro-
ubule dynamics with Nocodazole or Taxol affected theistribution of the detergent-extractable pool of RIIa,ndicating that RIIa may localize with microtubule-ssociated vesicles. Thus, RIIa and RIIb clearly localizeifferently in the Golgi–centrosomal region. This indi-ates specific roles for PKA isozymes containing eitherIIa or RIIb. © 1999 Academic Press
Key Words: PKA; cAMP-dependent protein kinase;KAP; subcellular localization; Golgi apparatus;entrosome.
INTRODUCTION
In mammalian cells four different regulatory sub-nits (R) of cAMP-dependent protein kinase (PKA)
1 Present address: Institut Curie, Biologie du Cycle Cellulaire et dea Motilite, 75248 Paris Cedex 05, France.
2 To whom correspondence and reprint requests should be ad-ressed at the Institute of Medical Biochemistry, University of Oslo,.O. Box 1112 Blindern, N-0317 Oslo, Norway. Fax: 147-22 85 14 97.-mail: [email protected].
131
ave been identified as separate gene products andave been demonstrated to exert two main functions inhe cAMP signal transduction pathway. The primaryunction of the R subunit is to bind and inactivate theatalytic subunit (C) of PKA and to control the releasef free active C from the inactive tetramer complexR2C2) upon the positive cooperative binding of four
olecules of cAMP to the R subunit dimer [1, 2]. Sec-nd, certain R subunits target different PKA isozymeso various subcellular loci and specific substrates bynteraction with various A-kinase anchoring proteinsAKAPs) localized in organelles or bound to cytoskel-tal components [3–6]. The diversity of regulatory sub-nits (RIa, RIb, RIIa, and RIIb) and their differentubcellular localizations may explain how PKA viahosphorylation can mediate multiple and to some ex-ent opposite effects of cAMP on cell function.
The intracellular localization of PKA type I or type IIsozymes, containing RI or RII subunits, respectively,s variable from cell to cell. RI is known to be mainlyoluble, but has been demonstrated to localize in prox-mity to membrane receptors such as antigen receptorsn lymphoid cells and nicotinic acetylcholine receptorsn neuromuscular junctions [7–9]. In contrast, RII sub-nits are primarily particulate and associated withytoskeletal elements and a number of organelles.hereas the subcellular targeting of RI via mono- or
ual-specific AKAPs is emerging [7, 10–13], it is wellnown that RII can be specifically bound to AKAPsssociated with microtubules (MAP2 and AKAP150),ostsynaptic densities and cortical actin (AKAP79/75),ctin (AKAP78/ezrin, AKAP-KL), nuclear matrixAKAP95 and nuclear AKAP150), sarcoplasmic reticu-um (AKAP100), endoplasmic reticulum (D-AKAP1),eroxisomes (AKAP220), Golgi apparatus (AKAP85),itochondria (AKAP84/149/D-AKAP1), centrosomes
AKAP450), filopodia (gravin/AKAP250), and L-typea21 channels (AKAP15/18) [14–31].The presence of two different isoforms of RII, RIIa
nd RIIb, raises the question of whether the localiza-ions of these two subunits are similar or different.ntracellular accumulation of RII has been observed inssociation with microtubules in nerve cells and withicrotubule-organizing centers and Golgi complex in
0014-4827/99 $30.00Copyright © 1999 by Academic Press
All rights of reproduction in any form reserved.

st3elscppTsiiLR
htRmadatamdubdcdgacb
Mfwm
b5atnac
i(GDaim
RttWi
PKo
posiaMispEastowmbu
mSltnCiamd
cJpnb0D15AtwfobLf(b
s1PMfPmT
132 KERYER ET AL.
everal mammalian cell types by immunocytochemis-ry and immunogold electron microscopy [25, 28, 32–6]. Whereas RIIa is expressed ubiquitously, RIIb isxpressed mainly in brain, adipocytes, ovarian granu-osa cells, and testicular Sertoli cells [37–42]. Expres-ion of RIIb has also been reported in a number ofancer cell lines and tumors [42–46]. The recently re-orted RIIb “knockout” mice display a distinct leanhenotype due to absence of RIIb in adipose tissue [47].his indicates that in some organ systems, a specific Rubunit is required. In other cells or tissues, R subunitsoforms may be redundant and overlap in function ass demonstrated for modulations of skeletal muscle-type Ca21 channel by PKA type II anchored throughIIa which is rescued by RIa in RIIa 2/2 mice [11, 48].Previous studies of the localization of RIIa and RIIb
ave either not concomitantly examined the localiza-ion of both isoforms or not fully distinguished betweenIIa and RIIb due to lack of suitable high-affinity,onospecific antibodies. Furthermore, a recent study
ddressed the specificity of the interaction of theimerization/AKAP binding domain of RIIa and RIIbnd showed clearly that, whereas some anchoring pro-eins bind both RIIa and RIIb, although with differentffinity, other anchoring molecules like AKAP95 al-ost exclusively bind RIIa [49]. To investigate in more
etail the differential localizations of the two RII sub-nit isoforms, we have generated monospecific anti-odies against human RIIa and human RIIb. It isemonstrated that RIIb, which is preferentially asso-iated with centrosomes, is expressed in terminallyifferentiated cells and in all neoplastic cells investi-ated, but not in normal proliferating cells. In contrast,major proportion of RIIa decorates Golgi-associated
ompartments and vesicles associated with microtu-ules in the trans-Golgi network (TGN) area.
MATERIALS AND METHODS
Chemicals. Nocodazole was purchased from Sigma (St. Louis,O) and dissolved to a concentration of 1022 M in DMSO. Taxol was
rom Sigma and was dissolved to 5 mM in DMSO. Brefeldin A (BFA)as from Sandoz Co. (Basel, Switzerland) and was dissolved to 5g/ml in methanol and subsequently stored at 220°C.Cells. Human osteosarcoma cells (SaOS2) were grown in Dul-
ecco’s modified Eagle’s medium supplemented with 0.1 mM glutamine,0 U/ml penicillin, 50 mg/ml streptomycin, and 10% fetal calf serumnd plated at 104 cells/ml for immunocytochemistry. Human neuroblas-oma cells (SMN-SMS) were grown in RPMI 1640 supplemented withonessential amino acids, 0.1 mM glutamine, 0.47 mM serine, 0.38 mMsparagine, and 10% fetal calf serum. Cell lines were from the Ameri-an Type Culture Collection (Rockville, MD).
Antibodies. Anti-human RIIa and RIIb antisera were made bymmunizing rabbits with hemocyanine-coupled synthetic peptidesRIIa, NH2-TPRQSLGHPPPEPGPDRVA-CONH2; RIIb, NH2-ERK-TARFGHEGRTWGDLG-CONH2; Multiple Peptide Systems, Saniego, CA) corresponding to amino acids 54 to 72 of RIIa and aminocids 44 to 62 of RIIb [38, 50]. Crude antisera were diluted 1/50 formmunoprecipitations of recombinant RII proteins or 1/100 for im-
unoprecipitations of RII subunits from cell extracts. Anti-human
IIa and RIIb antisera were affinity purified on columns with pep-ides coupled to Sepharose-4B (Pharmacia, Stockholm, Sweden) andhe resulting antibodies (hRIIa and hRIIb Abs) were screened on
estern blots and used at a concentration of 100 to 500 ng/ml formmunofluorescence studies.
An anti-human RIIa [21] monoclonal antibody (IgG1; Cat. No.55120) was developed by Transduction Laboratories (Lexington,Y) in collaboration with K.T. and T.J. and used at a concentrationf 1 mg/ml.Monoclonal antibodies CTR453 (IgG2b) and CTR433 (kindly sup-
lied by Dr. Michel Bornens, Curie Institute, Paris, France) werebtained from a library of monoclonal antibodies against centro-omes isolated from human lymphoblasts and have been character-zed as a specific marker of centrosomes (mAb CTR453 [51]) and as
marker of the median Golgi compartment (mAb CTR433 [52]).onoclonal Ab CTR453 was used as purified IgG (140 ng/ml) for
mmunocytochemistry and mAb CTR433 was used as hybridomaupernatant diluted 1/3 for immunofluorescence labeling. Affinity-urified anti-human galactosyl transferase Abs (a kind gift from Dr.ric Berger, Institute of Physiology, Zurich, Switzerland) was usedt a dilution of 1/100. Anti-a-tubulin mAb (Amersham, Buckingham-hire, UK) was used at a concentration of 600 ng/ml for immunocy-ochemistry. Secondary antibodies (Jackson ImmunoResearch Lab-ratories, West Grove, PA) used in immunofluorescence labelingere, unless otherwise stated, Texas red-conjugated anti-rabbit im-unoglobulin and FITC-conjugated anti-mouse immunoglobulin,
oth diluted 1/200. Rhodamine-conjugated anti-mouse immunoglob-lin (Cappell, West Chester, PA) was used at 1/750 dilution.Photoaffinity labeling by 8-azido-[32P]cAMP. Recombinant hu-an RIIa and RIIb were expressed in Escherichia coli as glutathione-transferase fusion proteins, cleaved by thrombin to generate full-
ength RIIa and RIIb, and purified (K. Tasken, unpublished). Pho-oaffinity labeling was accomplished by incubating RII subunits (20g/ml final concentration) with 1 mM 8-azido-[32P]cAMP (sp act 50i/mmol; ICN, Irvine, CA) in 50 mM Tris z Cl, pH 7.4, for 1 h at 4°C
n the dark as described elsewhere [53]. Covalent incorporation wasccomplished by exposure of reactions to UV light (254 nM, UVSL-58ineral lamp; Ultra Violet Products, Inc., San Gabriel, CA) at a
istance of 5 cm for 15 min at 20°C.Immunoprecipitations. Preparation of Staphylococcus aureus
ells expressing protein A (SACs; Pansorbin; Calbiochem Corp., Laolla, CA) and immunoprecipitation of labeled RII subunits wereerformed as described previously for RI subunits [53]. For immu-oprecipitation of RII subunits from cells, cell lysates were preparedy homogenization in RIPA buffer (150 mM NaCl, 1% Nonidet-P40,.5% deoxycholate, 0.1% SDS, and 50 mM Tris z Cl, pH 8.0) using aounce homogenizer, and lysates were subsequently centrifuged at5,000g for 15 min to remove debris. Cell extract (100 mg of protein,0 ml) was precleared by addition of 50 ml of 1:1 suspension of protein–Sepharose (Pharmacia), followed by incubation with anti-RII an-
ibodies for 4 h at 4°C. Subsequently antigen–antibody complexesere precipitated by incubation with 25 ml of protein A–Sepharose
or 2 h at 4°C and thereafter washed five times in RIPA buffer andnce in phosphorylation buffer (10 mM MgCl2, 20 mM cAMP, 10 mM-mercaptoethanol, 50 mM ATP, and 50 mM Tris z Cl, pH 7.4).abeling of immunoprecipitated RII was accomplished by incubation
or 15 min with 5 mg of purified bovine C subunit of PKA type IIkindly provided by Dr. Suzanne M. Lohmann, University of Wurz-urg, Germany) as described previously [28, 54].Immunocytochemistry. Cells were grown on culture-treated
lides for 2 days, rinsed twice in PBS, and fixed directly with cold00% methanol (220°C). Alternatively, cells were washed once withHEM buffer (45 mM Pipes, 45 mM Hepes, 10 mM EGTA, 5 mMgCl2, 1 mM PMSF, pH 6.9), extracted with detergent by incubating
or 1 min with PHEM buffer with 0.5% Triton X-100, washed inHEM buffer, and subsequently fixed with cold 100% methanol for 6in. Following fixation, cells were rinsed with PBS containing 0.1%ween 20 (PBST). Primary antibodies diluted in PBS containing 3%

Bwb1(BFPmwIcu5mcmsoosi
A
s7Ritdarlerp8hmt[(nan1i5lvrhi
wht
abftpaiH(bptmtfdap
eSRr
stfibl5tSS
133RIIa AND RIIb IN THE GOLGI–CENTROSOMAL AREA
SA were added for 1 h at room temperature. Cells were thenashed three times in PBST to remove unbound antibodies followedy incubation with fluorochrome-conjugated secondary antibodies forh at room temperature. Finally, cells were mounted in Citifluor
London, UK), examined, and photographed with either an OlympusX60 or a Zeiss Axiophot microscope (363 plan neofluor objective).or confocal microscopy, cells were fixed in 4% paraformaldehyde inBS for 20 min at 37°C, rinsed twice in PBS, and incubated for 10in with 50 mM ammonium chloride in PBS. Subsequently, cellsere permeabilized with 0.1% Triton X-100 in PBS with 0.2% BSA.
ncubation with antibodies was as described above. Confocal micros-opy was performed on a Sarastro 2000 confocal microscope (Molec-lar Dynamics, Palo Alto, CA) equipped with an argon laser (488- to14-nm wavelength) and mounted on either an Optiphot II Nikonicroscope (360, NA, 1.40, plan objective) or a Zeiss Axiophot mi-
roscope (363 plan neofluor objective). For double-staining experi-ents both fluorescein- and rhodamine- or Texas red-conjugated
econdary antibodies were used. Ten sections of 0.25 mm (averagingn five full frames of the same section) were scanned and stacks ofptical sections for each data set were compiled with Voxel Viewoftware on an IRIS 4D-70 GT graphics workstation (Silicon Graph-cs, Mountain View, CA).
RESULTS
ntibodies to Human RIIa and RIIb
Human RIIa and RIIb antisera were raised againstpecific sequences corresponding to amino acids 54 to2 and 44 to 62 of the N-terminal domain of humanIIa and RIIb, respectively. Oligopeptides were chosen
n the domains showing the greatest differences be-ween the human RIIa and RIIb sequences. Figure 1emonstrates lack of cross-reactivity between hRIIand hRIIb Abs. A sensitive assay for possible cross-eactivity was immunoprecipitation of 8-azido-[32P]cAMP-abeled recombinant human RII subunits followed bylectrophoresis in SDS–polyacrylamide gels and auto-adiography. Figure 1A shows that hRIIa Ab immuno-recipitated increasing amounts (12.5 to 200 ng) of-azido-[32P]cAMP photoaffinity-labeled recombinantRIIa (51 kDa) as well as a 46-kDa proteolytic frag-ent of hRIIa (lanes 1 to 5). Similarly, Fig. 1B shows
hat increasing amounts (12.5 to 200 ng) of 8-azido-32P]cAMP photoaffinity-labeled recombinant hRIIb53 kDa) was precipitated by hRIIb Ab. Importantly,o cross-reactivity was observed with either antibodys 200 ng of photoaffinity-labeled hRIIa or hRIIb wasot precipitated by the opposite antibody (Figs. 1A andB, lane 6 in both). The synthetic peptide used formmunization to produce hRIIa Ab covers amino acids4 to 72 and the position of the peptide is within aow-homology region of RIIa that displays interspeciesariation [38, 55]; thus, the resulting hRIIa Ab did noteact with mouse, rat, or bovine RIIa. In contrast,RIIb Ab reacted well with bovine and rodent RIIb in
mmunoblot analyses (data not shown).Immunoprecipitations using hRIIa and hRIIb Absere carried out on crude cell lysates from severaluman transformed cell lines. Figure 2 (top) showshat the hRIIa Ab precipitated a protein migrating at
pproximately 54 kDa from human SMN-SMS neuro-lastoma cells corresponding to the phosphorylatedorm of RIIa (lanes 1 and 2) after radioactive labeling ofhe immunoprecipitates by the C subunit of PKA. Com-arably lower amounts of a labeled protein migratingt 53 kDa corresponding to RIIb were detected bymmunoprecipitation with hRIIb Ab (lanes 3 and 4).uman SaOS2 osteosarcoma cells contained RIIa
lanes 5 and 6). However, RIIb was not readily detectedy immunoprecipitation (lanes 7 and 8), although atrolonged exposures a weak signal from a 53-kDa pro-ein corresponding to RIIb was observed following im-unoprecipitation with hRIIb Ab from SaOS2 cell ex-
racts (data not shown). Immunoblotting of extractsrom various cell types using the hRIIa and RIIb Absemonstrated distinct bands corresponding to RIIand RIIb with no apparent cross-reactivity to otherroteins (Table 1 and references cited therein).The specificity of hRIIa and hRIIb Abs was further
xamined by immunofluorescence analysis of humanMN-SMS neuroblastoma cells which contained bothIIa and RIIb (Fig. 2, bottom, A to F). hRIIa Ab deco-ated the Golgi–centrosomal area of SMN-SMS neuro-
FIG. 1. Immunoprecipitation of photoaffinity-labeled human RIIubunits using anti-human RII antibodies. Increasing amounts (12.5o 200 ng) of recombinant human RIIa (A) or RIIb (B) were photoaf-nity labeled using 8-azido-[32P]cAMP. Labeled proteins were incu-ated in solution with antibodies against human RIIa (anti-RIIa; A,anes 1 to 5, and B, lane 6) and human RIIb (anti-RIIb; B, lanes 1 to, and A, lane 6). Antigen–antibody complexes were immunoprecipi-ated by addition of S. aureus cells expressing protein A. WashedAC pellets were boiled in SDS sample buffer and subjected toDS–PAGE and subsequent autoradiography.
b2hpats
h(ch1ca
nRia[a(aa
134 KERYER ET AL.
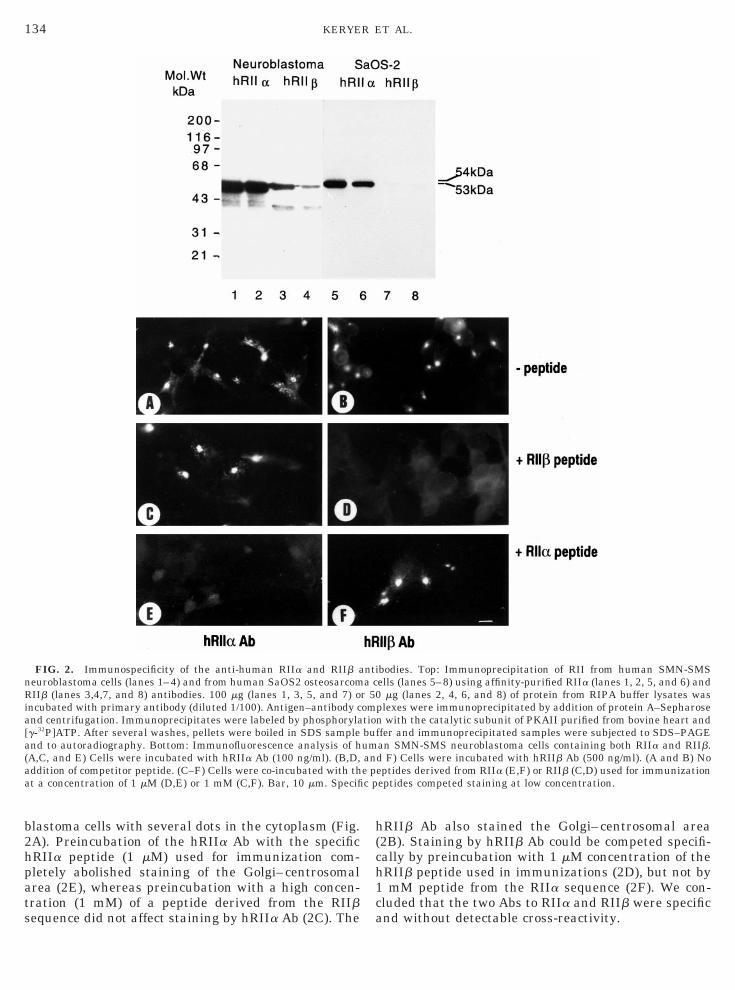
lastoma cells with several dots in the cytoplasm (Fig.A). Preincubation of the hRIIa Ab with the specificRIIa peptide (1 mM) used for immunization com-letely abolished staining of the Golgi–centrosomalrea (2E), whereas preincubation with a high concen-ration (1 mM) of a peptide derived from the RIIbequence did not affect staining by hRIIa Ab (2C). The
FIG. 2. Immunospecificity of the anti-human RIIa and RIIb aeuroblastoma cells (lanes 1–4) and from human SaOS2 osteosarcomIIb (lanes 3,4,7, and 8) antibodies. 100 mg (lanes 1, 3, 5, and 7) o
ncubated with primary antibody (diluted 1/100). Antigen–antibody cnd centrifugation. Immunoprecipitates were labeled by phosphorylag-32P]ATP. After several washes, pellets were boiled in SDS samplend to autoradiography. Bottom: Immunofluorescence analysis of hA,C, and E) Cells were incubated with hRIIa Ab (100 ng/ml). (B,D,ddition of competitor peptide. (C–F) Cells were co-incubated with tht a concentration of 1 mM (D,E) or 1 mM (C,F). Bar, 10 mm. Specifi
RIIb Ab also stained the Golgi–centrosomal area2B). Staining by hRIIb Ab could be competed specifi-ally by preincubation with 1 mM concentration of theRIIb peptide used in immunizations (2D), but not bymM peptide from the RIIa sequence (2F). We con-
luded that the two Abs to RIIa and RIIb were specificnd without detectable cross-reactivity.
bodies. Top: Immunoprecipitation of RII from human SMN-SMSells (lanes 5–8) using affinity-purified RIIa (lanes 1, 2, 5, and 6) and
mg (lanes 2, 4, 6, and 8) of protein from RIPA buffer lysates wasplexes were immunoprecipitated by addition of protein A–Sepharose
with the catalytic subunit of PKAII purified from bovine heart andfer and immunoprecipitated samples were subjected to SDS–PAGEan SMN-SMS neuroblastoma cells containing both RIIa and RIIb.d F) Cells were incubated with hRIIb Ab (500 ng/ml). (A and B) Noeptides derived from RIIa (E,F) or RIIb (C,D) used for immunizationeptides competed staining at low concentration.
ntia c
r 50omtionbufumane pc p
L
Gfl
pcimm
U
D
T
a(
c
i
c
b
(
135RIIa AND RIIb IN THE GOLGI–CENTROSOMAL AREA
ocalization of the RIIa and RIIb Subunits of PKA inHuman SaOS2 Osteosarcoma Cells
The differential localization of RIIa and RIIb in theolgi–centrosomal area was investigated by immuno-uorescence in a number of different cell lines and
TAB
Presence and Subcellular Localization ofDifferentiated, and Tr
Cell type RII subunit Solublea Pa
ndifferentiatedSkin fibroblasts a 1
b 2Myoblasts a 1
b 2T lymphocytes [7, 75] a 1
b 2B lymphocytes [8] a 1
b 2Preovulatory granulosa cells [59] a 1
b 2Cytotrophoblast [58] a 1
b 2ifferentiatedKeratinocytes a 1
b 2Myotube a 1
b 2Diff. granulosa lutein cellsf [59] a 1
b 1Syncytiotrophoblast [58] a 1
b 2Macrophages a 1
b 2ransformed cell linesHTB-17 astrocytoma a 1
b 2SMN-SMS neuroblastoma a 1
b 2SMN-SMS neuroblastoma
differentiatedg a 1b 2
SaOS2 osteosarcoma a 1b 2
KE-37 T lymphoblast [28] a 1b 2
HeLa cells a 1b 2
Note. Localization of RIIa and RIIb was mapped in different primnalysis, immunoprecipitation, or immunocytochemistry as indicated1) denotes a low-level expression not detected by all methods empl
a Soluble indicates observed expression in soluble fraction in the ahemistry.
b Particulate indicates observed expression in extracts solubilizemmunocytochemistry.
c Golgi associated indicates expression in solubilized Golgi prepahemistry and use of Golgi markers.
d Centrosome associated indicates expression in solubilized centroy immunocytochemistry and use of a centrosomal marker, CTR453
e Methods: IB, immunoblotting; IP, immunoprecipitation; IF, immf Differentiation of preovulatory human granulosa cell primary cult
100 ng/ml) and LH (10 ng/ml) for 36 h.g Differentiation of human SMN-SMS neuroblastoma cells was in
rimary cultures. Localization of RIIa in the Golgi–entrosomal area appeared independent of the cell typenvestigated (Table 1 and data not shown). Thus, hu-
an SaOS2 osteosarcoma cells were used to address inore detail the localization of RIIa and RIIb. Figure 3
1
a and RIIb in Human Undifferentiated,formed Human Cells
ulateb Golgi associatedc Centrosome associatedd Methode
1 1 If2 2 If1 1 If2 2 If1 1 If/Ip/Ib2 2 Ip1 1 If/Ip/Ib2 2 If/Ip/Ib1 1 If2 1 If1 1 If/Ib2 2 If/Ib
1 1 If2 1 If1 2 If2 2 If1 1 If1 1 If1 1 If2 2 If1 1 If1 1 If
1 1 If2 1 If1 1 If/Ip/Ib2 1 If/Ip/Ib
1 1 If/Ip/Ib) 1 1 If/Ip/Ib
1 1 If/Ip/Ib2 1 If/Ip/Ib1 1 If/Ip/Ib1 1 If/Ip/Ib1 1 If/Ip/Ib
(1) 1 If/Ip/Ib
y cultures or cell lines using hRIIa and hRIIb Abs by immunoblotor 2 denotes the absence or presence of the indicated RII subunit.
d.nce of detergent or a cytoplasmic localization seen by immunocyto-
ith 0.1% Triton X-100 or a membrane-associated localization by
ons or a Golgi-associated localization as observed by immunocyto-
e preparations or a centrosome-associated localization as observed
fluorescence microscopy.s to granulosa lutein cells was accomplished by treatment with FSH
ed by treatment with 100 mM 8-bromo-cAMP for 20 h.
LE
RIIans
rtic
121212121212
1212111212
1212
1(1121111
ar. 1
oyebse
d w
rati
som.unoure
duc

sw3d(fletssas
waNl
idwTmow
aea
136 KERYER ET AL.
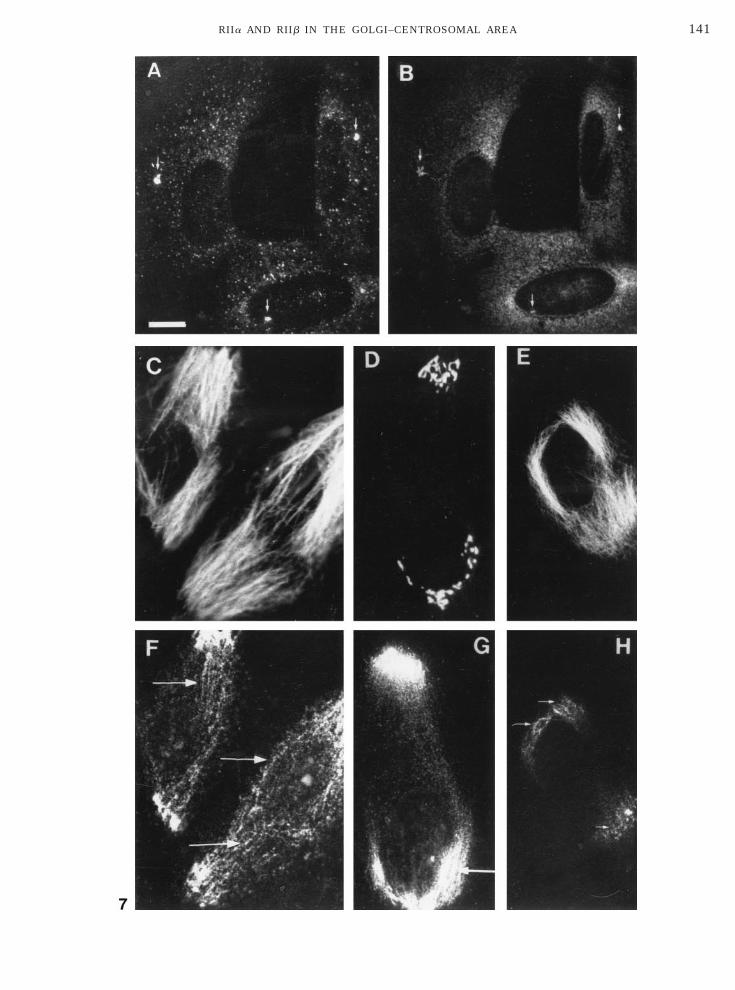
hows immunofluorescence labeling of SaOS2 cellsith hRIIa Ab (Figs. 3A–3C) and hRIIb Ab (Figs. 3D–F). As illustrated in Fig. 3A, hRIIa Ab prominentlyecorated a perinuclear area, including centrosomesarrow), Golgi apparatus (arrowhead), and a granularuorescence extending into the cytoplasm. Followingxtraction of cells with Triton X-100 prior to fixation,he granular fluorescence was lost, whereas the centro-omal- (arrow) and some Golgi-associated labeling per-isted (Fig. 3B). At higher magnification (Fig. 3C), RIIappeared to be associated with filamentous structuresurrounding centrosomes (arrows). This association
FIG. 3. Localization of RIIa and RIIb in human osteosarcoma cend D) or first permeabilized with 0.5% Triton X-100 in PHEM and tither hRIIa Ab (200 ng/ml) (A–C) or hRIIb Ab (500 ng/ml) (D–F). Whnd white arrows indicate staining of centrosomes. Bars: A,B,D,E, 3
as not affected by Triton X-100 extraction (Figs. 3Bnd 3C) nor by subsequent extractions with salt (0.5 MaCl) and/or ATP (10 mM), which is known to solubi-
ize molecular motor proteins as kinesin or dynein [56].Although only very low levels of RIIb were detected
n SaOS2 cells in immunoprecipitation experiments, aistinct staining of centrosomes (Figs. 3D–3F, arrows)as observed with hRIIb Ab, which was not affected byriton X-100 extraction (Figs. 3E and 3F). Further-ore, some staining of small cytoplasmic vesicles was
bserved also with the hRIIb antibody (Fig. 3D) thatas lost during Triton X-100 extraction (Fig. 3E).
SaOS2 osteosarcoma cells were either fixed with cold methanol (Afixed with cold methanol (B,C,E, and F). Cells were incubated with
arrowheads indicate the staining of a Golgi-associated compartmentmm; C,F, 2.5 mm.
lls.henite.5

L
ltrhb(hcimo(itHcsrlA[tntowpFwrwHrfnhn
T
iftws
gaFplmoadtbshifhtc
eRlllp
D
GGmCcG(mmwwes
RGm6c
nmha
h
137RIIa AND RIIb IN THE GOLGI–CENTROSOMAL AREA
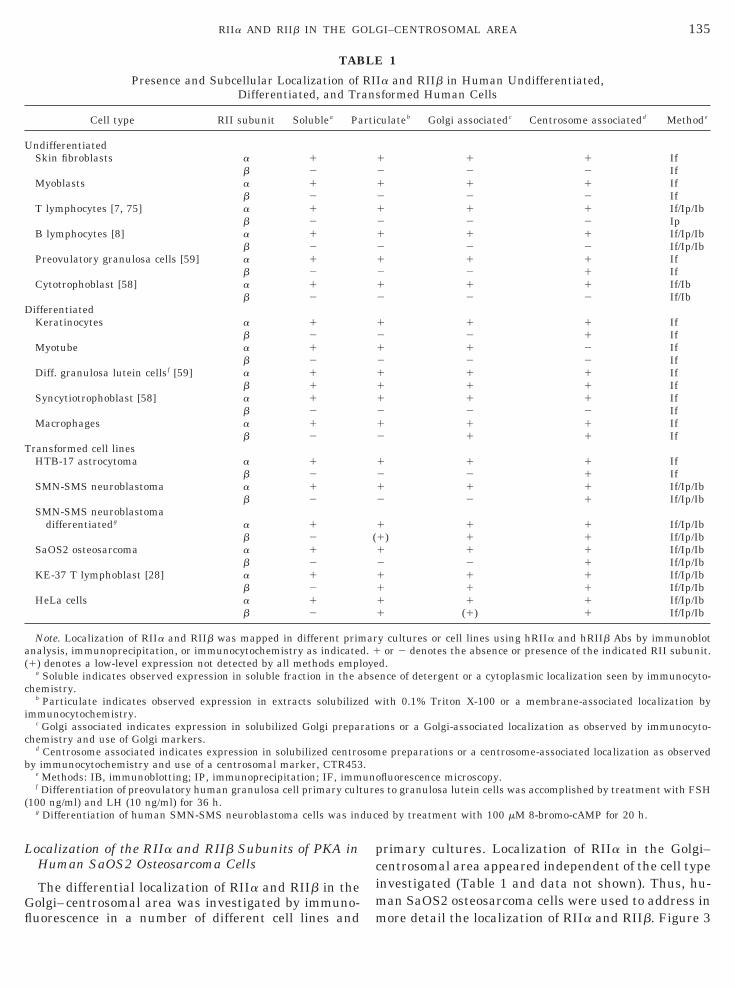
ocalization of RIIb in the Pericentrosomal Area
In order to assess in more detail the centrosomalocalization of RII, we investigated the subcellular dis-ribution of RIIb that preferentially localized to thisegion. Human SMN-SMS neuroblastoma cells, havingigher expression of RIIb than SaOS2 cells, were dou-le labeled with hRIIb Ab and a centrosomal markermAb CTR453). The decoration of centrosomes byRIIb Ab (Fig. 4A) colocalized with the staining of theentrosomal marker, CTR453 (Fig. 4C [51]). A confocalmage reconstruction of sections through the centroso-
al area showed a very precise colocalization (yellow)f hRIIb Ab staining (red) with mAb CTR453 labelinggreen) (Fig. 4B). RIIb appeared also to be radiatingnto the pericentrosomal area. These radiations fromhe centrosome could be microtubule-associated RIIb.owever, RIIb did not colocalize with a-tubulin of mi-
rotubules in a double-labeling experiment (data nothown), ruling out this hypothesis. Furthermore, suchadiating decorations were observed in the pericentrio-ar matrix itself. Double immunostaining with hRIIbb (green) and a median-Golgi marker, mAb CTR433
52] (red), demonstrated that RIIb did not localize tohe Golgi compartment in undifferentiated SMN-SMSeuroblastoma cells (Fig. 4D). A single yellow dot inhe figure represents centrosomal RIIb superimposedn the Golgi staining. Similar observations were madeith primary cultures of human granulosa cells fromreovulatory follicles that expressed RIIb (Table 1).urthermore, modifications of microtubule dynamicsith Nocodazole or Taxol, or disruption of Golgi appa-
atus with BFA, did not alter the localization of RIIb,hich remained strictly centrosomal (data not shown).owever, cAMP treatment of human SMN-SMS neu-
oblastoma, which initiates differentiation and neuriteormation as described elsewhere [57], was accompa-ied by higher levels of RIIb and a redistribution ofRIIb Ab labeling along the neurites (Table 1 and dataot shown).
ransformed Cell Lines Contain Centrosomal RIIb
In order to examine the distribution of RII subunitsn more detail, a number of primary cultures of undif-erentiated and differentiated human cells were inves-igated for the localization of RIIa and RIIb, and thisas compared to the expression and localization of RII
ubunits in transformed cell lines. All cells investi-
FIG. 4. Major centrosomal localization of RIIb. Human SMN-Sg/ml) and with a centrosomal marker (mAb CTR453) or with a mediAb CTR453, respectively. (B) Confocal image reconstruction fromRIIb Ab (red) and mAb CTR453 (green). (D) Staining with hRIIb Abntibodies were (A–C) Texas red-conjugated anti-rabbit Ab and (D)FIG. 5. RIIa colocalization with Golgi compartments. Double lab
uman galactosyl transferase (B) in interphase cells. Bar represents
ated expressed RIIa in soluble, particulate, Golgi-ssociated, and centrosomal compartments (Table 1).urthermore, undifferentiated fibroblast and myoblastrimary cultures as well as peripheral blood B and Tymphocytes did not express RIIb. Granulosa cell pri-
ary cultures isolated from preovulatory follicles dem-nstrated a low-level expression of RIIb localized onlyt centrosomes. Differentiated syncytia of myotubesid not express RIIb nor did the syncytia of syncy-iotrophoblast following differentiation of cytotropho-last [58]. Keratinocytes displayed a low-level expres-ion of RIIb localized only at centrosomes, whereasuman granulosa cell primary cultures differentiatedn vitro to granulosa lutein cells by treatment withollicle-stimulating hormone (FSH) and luteinizingormone [59] exhibited a high-level expression of RIIbhat was present in cytosol, membranes, Golgi, andentrosomes.In contrast to the absence of RIIb in most undiffer-
ntiated primary cultures, a striking finding was thatIIb was present in centrosomes of all transformed cell
ines investigated (Table 1). In some transformed cellsike the KE-37 T lymphoblasts or HeLa cells, RIIb alsoocalized to a Golgi-associated and particulate com-artment.
istribution of RIIa in the Golgi Area
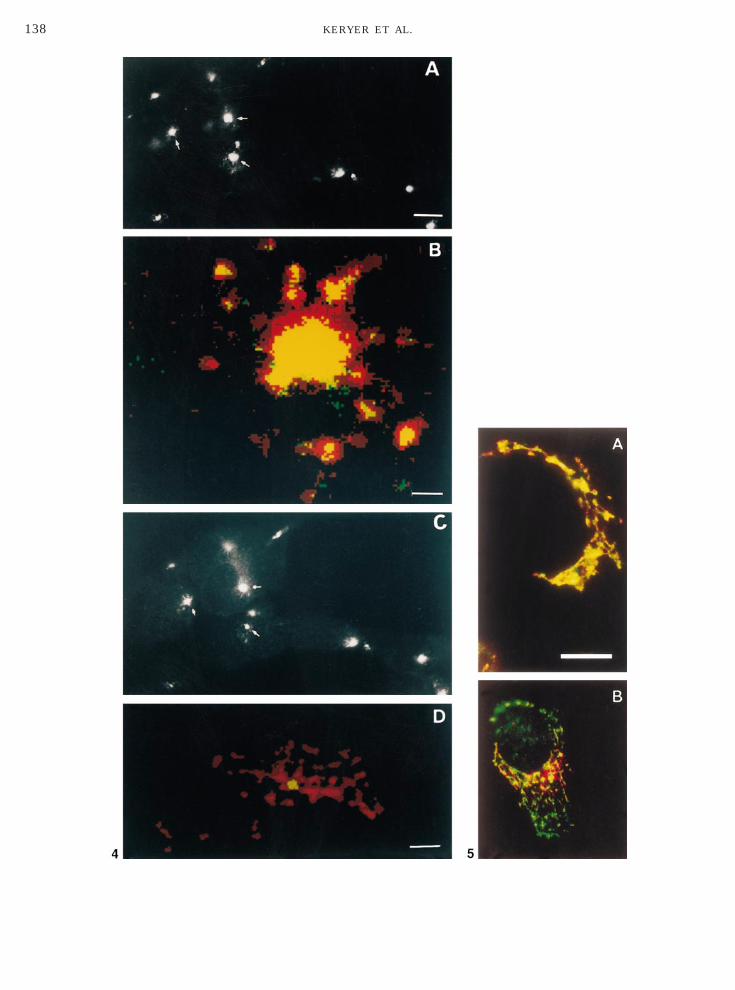
We next investigated the localization of RIIa inolgi-associated compartments. We examined theolgi area of SaOS2 osteosarcoma cells by double im-unofluorescence with a median-Golgi marker (mAbTR433) and hRIIa Ab. Dual labeling (yellow) indi-ated quite extensive overlap throughout the median-olgi area, although some discrimination of RIIa
green) could be seen (Fig. 5A). Partial overlap of RIIaAb labeling (red) with a trans-Golgi compartmentarker (anti-human galactosyl transferase Ab; green)as also detected (Fig. 5B). Furthermore, stainingith a cis-Golgi compartment marker [60] indicatedven lower or no colocalization with RIIa (data nothown).In order to investigate more closely the association ofIIa with Golgi compartments, dual staining of theolgi area of SaOS2 cells with the median-Golgiarker CTR433 (Figs. 6A and 6C) and hRIIa Ab (Figs.
B and 6D) was performed and examined in singleonfocal sections. Surprisingly, comparison of the pho-
neuroblastoma cells were double stained with the hRIIb Ab (500Golgi marker (mAb CTR433). (A and C) Staining with hRIIb Ab anden sections in the centrosomal area following double staining witheen, 500 ng/ml) and median-Golgi marker CTR433 (red). SecondaryC-conjugated anti-mouse. Bars: A,C, 10 mm; B, 0.25 mm; D, 2 mm.g of RIIa (green in A, red in B) and Golgi markers CTR433 (A) and
, 4.8 mm, and B, 7.8 mm.
MSan-sev(gr
FITelin
A
138 KERYER ET AL.
139RIIa AND RIIb IN THE GOLGI–CENTROSOMAL AREA

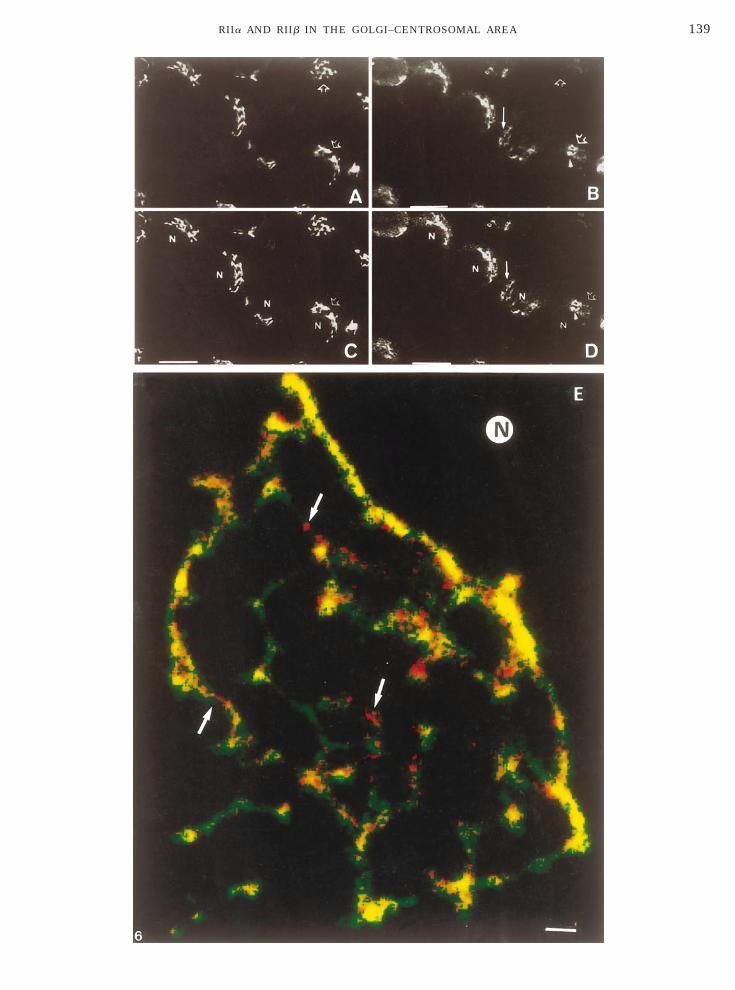
tsC(rvutyaGaaa
mca(a(G3StmaTgactmtcro(FSasa(tci
a
Gtt8vrmct8btlmw(wdd
udsgshcRnpieRmaa
hilthakpt
dAvoi
140 KERYER ET AL.
omicrographs revealed that, whereas some of the RIIataining overlapped with the median-Golgi marker,TR433, a larger proportion of the staining did not
indicated by open and solid arrows). Confocal imageeconstruction of 10 thin optical sections which re-ealed that although RIIa labeling (red) was distrib-ted across the median-Golgi area and colocalized withhe median-Golgi compartment (green label, overlap inellow) (Fig. 6E), a significant proportion of the Golgi-ssociated RIIa was discriminated from the median-olgi marker at this resolution. RIIa appeared morebundant at the trans side of the Golgi apparatus andssociated with small granular structures in the Golgirea (Fig. 6E, arrows).Next, we investigated the redistribution of RIIa byodifications of the microtubule network in SaOS2
ells. Following treatment with Nocodazole [61] to dis-ssemble microtubules (verified by a-tubulin staining)Fig. 7A), a granular hRIIa Ab labeling was observedlong with a centrosomal staining indicated by arrowsFig. 7B). However, granules no longer localized to theolgi–centrosomal area (compare with Figs. 3A andC). To assess the microtubule association of RIIa,aOS2 cells were treated with Taxol to stabilize micro-ubules [62]. This led to a distinct reorganization oficrotubules originating from the periphery of the cell
s demonstrated by a-tubulin staining (Fig. 7C). Afteraxol treatment, hRIIa Ab staining was granular, sug-esting association of RIIa with cytoplasmic vesiclesnd/or a cytoskeletal component redistributed with mi-rotubule tracks (Fig. 7F, arrows) that could be iden-ified by double staining for a-tubulin (Fig. 7C). Dra-atic redistribution of Golgi apparatus subjacent to
he plasma membrane was observed in Taxol-treatedells by mAb CTR433 (Fig. 7D). The redistributed Golgiegion now appeared concentrated with a granular flu-rescence when RIIa was stained for in double labelingFig. 7G), as this region was most intensely labeled.urthermore, Triton X-100 extraction of Taxol-treatedaOS2 cells prior to fixation and double labeling with-tubulin Ab and hRIIa Ab (Figs. 7E and 7H) demon-trated that, whereas a-tubulin organization was notffected (Fig. 7E), RIIa labeling was greatly reducedFig. 7H). This indicates that a distinct proportion ofhe RIIa binding involves membrane structures whichan be solubilized by nonionic detergent and not directnteractions between RIIa and a-tubulin.
To further investigate association of RIIa with Golgi-ssociated compartments, BFA was used to disrupt
FIG. 6. Staining of SaOS2 osteosarcoma cells with hRIIa Ab andescribed above. Ten sections of 0.25 mm were scanned. (A to D) Twoto D indicate differences in Golgi staining; open arrows denote pres
ersa. Arrowhead in B and D indicates centrosome decorated only brescence with mAb CTR433 (green) and hRIIa (red) Ab. Confocal ims shown. N, nucleus. Arrows indicate small vesicular structures sta
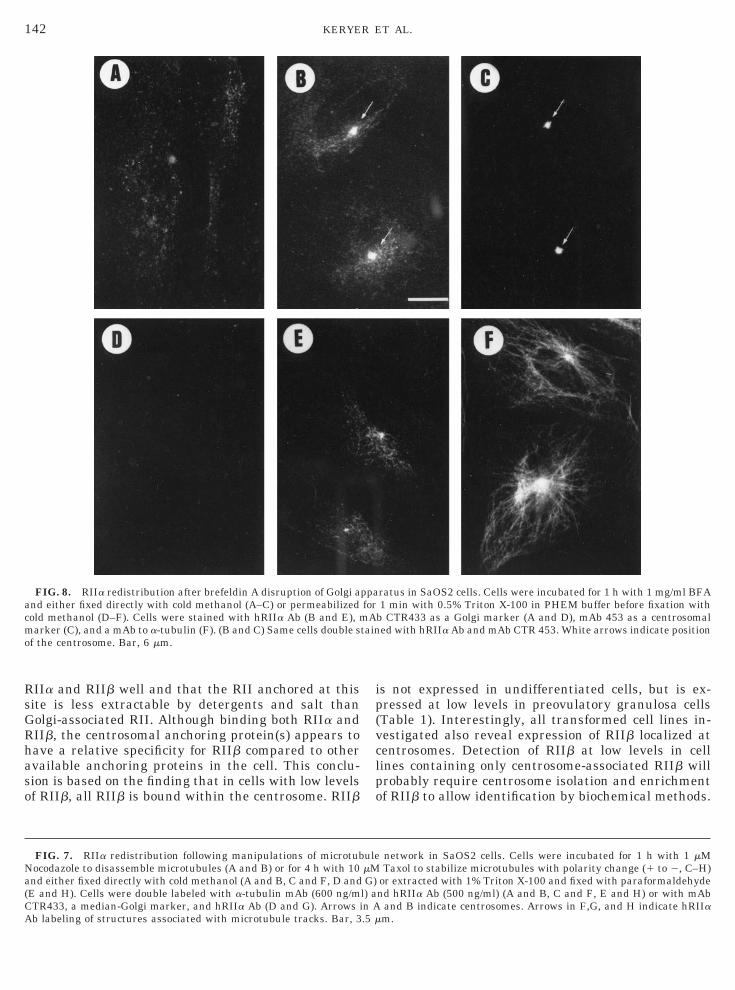
olgi cisternae in SaOS2 cells [63]. Following BFAreatment, a faint, disperse granular staining was de-ected with the median-Golgi marker CTR433 (Fig.A). Labeling of BFA-treated cells with hRIIa Ab re-ealed a prominent centrosomal staining (Fig. 8B, ar-ows) identified by colocalization with a centrosomalarker, CTR453 (Fig. 8C, arrows), as well as some
ytoplasmic granular fluorescence. Triton X-100 ex-raction of BFA-treated cells prior to fixation (Figs.D–8F) demonstrated that the CTR433 marker coulde washed out by detergent (Fig. 8D), whereas neitherhe centrosomal nor the granular cytoplasmic RIIaabel was removed (Fig. 8E). However, although the
icrotubule network was intact following treatmentith BFA as assessed by staining with a-tubulin mAb
Fig. 8F), no exact colocalization of RIIa with tubulinas found. Furthermore, the RIIa-associated structureid not collapse onto centrosomes as parts of the TGNo [64].
DISCUSSION
The present study demonstrates that the RII sub-nit isoforms of PKA are localized to overlapping andifferent subcellular structures in human cells. Mono-pecific anti-human RIIa and RIIb antibodies wereenerated, characterized, and used to investigate theubcellular localization of RII subunits in severaluman cell types. The results show that the Golgi–entrosomal area concentrates a major proportion ofII subunits. In cells containing RIIb (transformed orondividing cells), both RIIb and RIIa localize in theericentriolar matrix of the centrosome. Similar local-zation is observed for RIIa, but not RIIb, in undiffer-ntiated proliferating cells. Furthermore, RIIa, but notIIb, is anchored to several Golgi-associated compart-ents that include the median- and trans-Golgi as well
s structures that appear to be small microtubule-ssociated vesicles in the Golgi/TGN area.Centrosomal anchoring of both RIIa and RIIb is
ighly resistant to nonionic detergent solubilization,ndicating tight binding to a protein(s) not embedded inipid membranes. We have previously identified a cen-rosome-specific anchoring protein, AKAP450, that isighly insoluble [28]. (See note added in proof.) Inddition, centrosomal preparations contained a 160-Da anchoring protein that was also present in Golgireparations [28]. The observations made here indicatehat the centrosomal anchoring protein(s) binds both
edian-Golgi marker (mAb CTR433). SaOS2 cells were prepared asrial sections stained with CTR433 (A, C) and RIIa (B, D). Arrows ine of CTR433 and absence of RIIa, whereas filled arrows indicate viceIIa. Bar, 15 mm. (E) Dual color overlay of double-label immunoflu-reconstruction from 10 optical sections of 0.25 mm in the Golgi area
d with RIIa only. Bar, 1 mm.
a mse
ency Rageine
141RIIa AND RIIb IN THE GOLGI–CENTROSOMAL AREA
RsGRhaso
ip(vclpo
Na(CA
acmo
142 KERYER ET AL.
IIa and RIIb well and that the RII anchored at thisite is less extractable by detergents and salt thanolgi-associated RII. Although binding both RIIa andIIb, the centrosomal anchoring protein(s) appears toave a relative specificity for RIIb compared to othervailable anchoring proteins in the cell. This conclu-ion is based on the finding that in cells with low levelsf RIIb, all RIIb is bound within the centrosome. RIIb
FIG. 7. RIIa redistribution following manipulations of microtubocodazole to disassemble microtubules (A and B) or for 4 h with 10nd either fixed directly with cold methanol (A and B, C and F, D andE and H). Cells were double labeled with a-tubulin mAb (600 ng/mTR433, a median-Golgi marker, and hRIIa Ab (D and G). Arrows ib labeling of structures associated with microtubule tracks. Bar, 3
FIG. 8. RIIa redistribution after brefeldin A disruption of Golgi and either fixed directly with cold methanol (A–C) or permeabilizedold methanol (D–F). Cells were stained with hRIIa Ab (B and E), marker (C), and a mAb to a-tubulin (F). (B and C) Same cells double s
f the centrosome. Bar, 6 mm.
s not expressed in undifferentiated cells, but is ex-ressed at low levels in preovulatory granulosa cellsTable 1). Interestingly, all transformed cell lines in-estigated also reveal expression of RIIb localized atentrosomes. Detection of RIIb at low levels in cellines containing only centrosome-associated RIIb willrobably require centrosome isolation and enrichmentf RIIb to allow identification by biochemical methods.
network in SaOS2 cells. Cells were incubated for 1 h with 1 mMTaxol to stabilize microtubules with polarity change (1 to 2, C–H)
or extracted with 1% Triton X-100 and fixed with paraformaldehydend hRIIa Ab (500 ng/ml) (A and B, C and F, E and H) or with mAb
and B indicate centrosomes. Arrows in F,G, and H indicate hRIIam.
ratus in SaOS2 cells. Cells were incubated for 1 h with 1 mg/ml BFA1 min with 0.5% Triton X-100 in PHEM buffer before fixation with
b CTR433 as a Golgi marker (A and D), mAb 453 as a centrosomaled with hRIIa Ab and mAb CTR 453. White arrows indicate position
ulemMG)
l) an A.5 m
ppaforA
tain

SscssocapcAaeaartstPltic
RaGatrimpwRswGgtcRTcmlcaoemtadi
cdtocpftrsw
sektnGalbpatinActovwmtIAatitia
aevsm7lidtnnIw
143RIIa AND RIIb IN THE GOLGI–CENTROSOMAL AREA
RIIb is expressed at high levels in granulosa andertoli cells that are differentiating by FSH or cAMPtimulation [41, 65–67]. In FSH-treated granulosaells or in cAMP-treated neuroblastoma cells, expres-ion of RIIb is higher and RIIb reveals a differentubcellular localization. RIIb becomes anchored tother loci than centrosomes, including Golgi-associatedompartments. In granulosa lutein cells (Table 1) andlso in rat Sertoli cell primary cultures [68], a majorroportion of the RIIb protein becomes soluble, indi-ating that the binding capacity of the high-affinityKAP in these cells is exceeded. RIIb probably inter-cts with a number of hereto unknown AKAPs. Forxample, RIIb may associate with hormonally inducednchoring proteins such as the FSH-induced 80-kDanchoring protein in granulosa cells [69]. Hormonalegulation, differentiation-specific expression, and dis-inct subcellular localization of RIIb indicate that thisubunit of PKA concentrates catalytic activity to cer-ain sites and may regulate specific functions of cAMP/KA in terminally differentiated or hormone-stimu-
ated cells. In this respect, it is interesting to speculatehat the expression of RIIb in transformed proliferat-ng cells may represent a disturbance of normal growthontrol.
In contrast to the centrosomal localization of RIIb,IIa appears more dispersed. In addition to anchoringt centrosomes, most of RIIa exhibits a perinuclear/olgi-associated localization and is detergent extract-ble. Furthermore, RIIa seems to localize in a filamen-ous network in the Golgi–centrosomal area, alsoesistant to detergent extraction. Analysis of the local-zation of RIIa by double-immunofluorescence confocal
icroscopy using cis-, median-, and trans-Golgi com-artment markers reveals some colocalization of RIIaith the median- and trans-Golgi. Nevertheless, someIIa labeling was also associated with small vesiculartructures in the proximity of the Golgi cisternae,hich could indicate staining of vesicles trafficking theolgi area. RIIa can be removed by nonionic deter-ents, which supports a membrane-associated localiza-ion. In addition, disassembly of microtubules by No-odazole leads to a dispersed granular staining forIIa. Finally, reorganization of microtubules withaxol results in redistribution of RIIa from the Golgi–entrosomal area to regions subjacent to the plasmaembrane with a granular, but detergent-extractable
abeling radiating along microtubule tracks. This indi-ates that RIIa may redistribute with microtubule-ssociated vesicular structures. In vivo relocalizationf the minus end of microtubules toward the nuclearnvelope in myotube nuclei during myoblast–myotubeorphogenesis [70] is also accompanied by RIIa redis-
ribution from a pericentriolar location in myoblasts toperinuclear localization in myotubes (Table 1 and
ata not shown). No RIIa is observed around centriolesn the differentiated myotube. These observations are
ompatible with the notion that RIIa is associated notirectly with microtubules, but with small vesiclesraveling along the minus end of microtubules. Therigin of RIIa vesicles appears to be the trans-Golgiompartment. Furthermore, disruption of the Golgi ap-aratus with BFA separates the distribution of RIIarom that of a median-Golgi marker and from the dis-ribution of trans-Golgi proteins that are known toedistribute to the intermediate compartment (nothown). This suggests that RIIa may be associatedith structures in the TGN.A previous report characterized an 85-kDa Golgi-
pecific anchoring protein, AKAP85, in preparationsnriched in Golgi vesicles [25]. Furthermore, the 160-Da AKAP present in centrosomes was also detected inhe same preparations, but this Golgi-rich fraction didot allow segregation of different compartments in theolgi/TGN. We have also observed that in Golgi prep-rations of several cell lines (HeLa, astrocytoma, KE37ymphoblasts), a low amount of pericentriolar matrix,ut not centrioles, is present. This indicates that theericentriolar matrix is tightly intermingled with TGNnd that AKAPs identified in centrosomal prepara-ions (AKAP160, AKAP450) may copurify with AKAPsn the TGN. In contrast, centrosomal preparations doot contain the Golgi-associated AKAP85. In addition,KAP150 and MAP2 are microtubule-associated an-horing proteins that may be involved in the localiza-ion of RIIa in the TGN region [14, 15]. However, thebservation that the majority of RIIa is associated withesicles does not support a direct interaction of RIIaith microtubule-associated AKAPs. Nevertheless, ainor proportion of the RIIa is not solubilized by de-
ergent and may be associated with the TGN scaffold.n contrast to the detergent-resistant pool of RIIa,KAP85 is an integral membrane protein [25]. Associ-tion of vesicles in the TGN compartment with micro-ubules indicates that elements in these vesicles maynteract with a cognate receptor associated with micro-ubules in the TGN scaffold. Whether either of thesenteracting molecules or associated proteins serves tonchor RIIa is not known.No exact function has yet been associated with RIIa
nd PKA type II in the Golgi/TGN compartment. How-ver, endocytotic function as cellular uptake and re-erse transport of the enterotoxin, ricin, has beenhown to increase upon cAMP treatment [71]. Further-ore, cAMP enhances formation of microtubules [72,
3], and an anchored PKA type II may serve to stabi-ize the minus end of microtubules. In Drosophila, annactivating mutation in the C subunit DC0 of PKAisrupts microtubule distribution along the anteriopos-erior axis of oocytes, indicating that PKA may beecessary for regulation of specific spatial microtubuleucleation and disassembly [73]. In addition, PKA typeI is anchored at centrosomes in proliferating cells asell as in terminally differentiated cells. However,

wnltsdtsmaG
Ph(kgrLNcStCp2
rBvAE
1
1
1
1
1
1
1
1
1
1
2
2
2
2
144 KERYER ET AL.
hereas RIIb present in normal differentiated andeoplastic cells does not detach from its centrosomal
ocalization at mitosis (data not shown), RIIa is redis-ributed and not associated with centrosomes at mito-is [74]. The presence or absence of PKA at centrosomesuring mitosis may have functional consequences dic-ated by the PKA type II isozyme. Future cloning andtructure/function analysis of Golgi/TGN and centroso-al AKAPs may lead to an understanding of the role
nd impact of PKA localized in the centrosome andolgi/TGN compartments.
The authors are grateful to Drs. M. Bornens (Curie Institute,aris, France) and D. Evain-Brion (Inserm U427, Paris, France) forelpful discussions during this work, to Dr. Francoise RaynaudInserm U427, Paris, France) for preparation of skin fibroblast anderatinocyte primary cultures, and to Dr. Philippe Collas (Norwe-ian College of Veterinary Medicine, Oslo, Norway) for criticallyeading the manuscript. We appreciate the expert assistance of Dr.eif Scherven and Randi Væråmoen (The Animal Department, Theational Hospital, Oslo, Norway) in the production of rabbit poly-
lonal antibodies. This work was supported by the Norwegian Cancerociety, the Norwegian Research Council, Anders Jahre’s Founda-ion for the Promotion of Science, the Novo Nordisk Foundationommittee, and the Odd Fellow Medical Foundation. G.K. was sup-orted by the Association pour la Recherche sur le Cancer (ARC No.071) and the Centre National de la Recherche Scientifique.
Note added in proof. While this paper was in review we cloned andeported a centrosomal anchoring protein: Witczak, O., Skalhegg,. S., Keryer, G., Bornens, M., Tasken, K., Jahnsen, T., and Ørsta-ik, S. (1999). Cloning and characterization of a cDNA encoding an-kinase anchoring protein located in the centrosome, AKAP450.MBO J. 18, 1858–1868.
REFERENCES
1. Scott, J. D. (1991). Cyclic nucleotide-dependent protein kinases.Pharmacol. Ther. 50, 123–145.
2. Francis, S. H., and Corbin, J. D. (1994). Structure and functionof cyclic nucleotide-dependent protein kinases. Annu. Rev.Physiol. 56, 237–272.
3. Scott, J. D., and Carr, D. W. (1992). Subcellular localization ofthe type II cAMP-dependent protein kinase. News Physiol. Sci.7, 143–148.
4. Scott, J. D., and McCartney, S. (1994). Localization of A-kinasethrough anchoring proteins. Mol. Endocrinol. 8, 5–11.
5. Rubin, C. S. (1994). A kinase anchor proteins and the intracel-lular targeting of signals carried by cyclic AMP. Biochim. Bio-phys. Acta 1224, 467–479.
6. Faux, M. C., and Scott, J. D. (1996). Molecular glue: Kinaseanchoring and scaffold proteins. Cell 85, 9–12.
7. Skålhegg, B. S., Tasken, K., Hansson, V., Huitfeldt, H. S.,Jahnsen, T., and Lea, T. (1994). Location of cAMP-dependentprotein kinase type I with the TCR/CD3 complex. Science 263,84–87.
8. Levy, F. O., Rasmussen, A. M., Tasken, K., Skålhegg, B. S.,Huitfeldt, H. S., Funderud, S., Smeland, E. B., and Hansson, V.(1996). Cyclic AMP-dependent protein kinase (cAK) in human Bcells: Co-localization of type I cAK (RIa2C2) with the antigenreceptor during anti-immunoglobulin-induced B cell activation.Eur. J. Immunol. 26, 1290–1296.
9. Imaizumi-Scherrer, T., Faust, D. M., Benichou, J. C., Hellio, R.,and Weiss, M. C. (1996). Accumulation in fetal muscle and
localization to the neuromuscular junction of cAMP-dependentprotein kinase A regulatory and catalytic subunits RI alpha andC alpha. J. Cell Biol. 134, 1241–1254.
0. Huang, L. J., Durick, K., Weiner, J. A., Chun, J., and Taylor,S. S. (1997). Identification of a novel protein kinase A anchoringprotein that binds both type I and type II regulatory subunits.J. Biol. Chem. 272, 8057–8064.
1. Burton, K. A., Johnson, B. D., Hausken, Z. E., Westenbroek,R. E., Idzerda, R. L., Scheuer, T., Scott, J. D., Catterall, W. A.,and McKnight, G. S. (1997). Type II regulatory subunits are notrequired for the anchoring-dependent modulation of Ca21 chan-nel activity by cAMP-dependent protein kinase. Proc. Natl.Acad. Sci. USA 94, 11067–11072.
2. Huang, L. J., Durick, K., Weiner, J. A., Chun, J., and Taylor,S. S. (1997). D-AKAP2, a novel protein kinase A anchoringprotein with a putative RGS domain. Proc. Natl. Acad. Sci. USA94, 11184–11189.
3. Angelo, R., and Rubin, C. S. (1998). Molecular characterizationof an anchor protein (AKAPCE) that binds the RI subunit(RCE) of type I protein kinase A from Caenorhabditis elegans.J. Biol. Chem. 273, 14633–14643.
4. Lohmann, S. M., DeCamilli, P., Einig, I., and Walter, U. (1984).High-affinity binding of the regulatory subunit (RII) of cAMP-dependent protein kinase to microtubule-associated and othercellular proteins. Proc. Natl. Acad. Sci. USA 81, 6723–6727.
5. Bregman, D. B., Bhattacharyya, N., and Rubin, C. S. (1989).High affinity binding protein for the regulatory subunit ofcAMP-dependent protein kinase II-B. Cloning, characteriza-tion, and expression of cDNAs for rat brain P150. J. Biol. Chem.264, 4648–4656.
6. Carr, D. W., Stofko-Hahn, R. E., Fraser, I. D., Cone, R. D., and Scott,J. D. (1992). Localization of the cAMP-dependent protein kinase tothe postsynaptic densities by A-kinase anchoring proteins. Char-acterization of AKAP 79. J. Biol. Chem. 267, 16816–16823.
7. Li, Y., Ndubuka, C., and Rubin, C. S. (1996). A kinase anchorprotein 75 targets regulatory (RII) subunits of cAMP-dependentprotein kinase to the cortical actin cytoskeleton in non-neuronalcells. J. Biol. Chem. 271, 16862–16869.
8. Dransfield, D. T., Bradford, A. J., Smith, J., Martin, M., Roy, C.,Mangeat, P. H., and Goldenring, J. R. (1997). Ezrin is a cyclicAMP-dependent protein kinase anchoring protein. EMBO J.16, 35–43.
9. Dong, F., Feldmesser, M., Casadevall, A., and Rubin, C. S.(1998). Molecular characterization of a cDNA that encodes sixisoforms of a novel murine A kinase anchor protein. J. Biol.Chem. 273, 6533–6541.
0. Coghlan, V. M., Langeberg, L. K., Fernandez, A., Lamb, N. J.,and Scott, J. D. (1994). Cloning and characterization of AKAP95, a nuclear protein that associates with the regulatory sub-unit of type II cAMP-dependent protein kinase. J. Biol. Chem.269, 7658–7665.
1. Eide, T., Coghlan, V., Ørstavik, S., Holsve, C., Solberg, R.,Skålhegg, B. S., Lamb, N. J., Langeberg, L., Fernandez, A.,Scott, J. D., Jahnsen, T., and Tasken, K. (1998). Molecularcloning, chromosomal localization, and cell cycle-dependentsubcellular distribution of the A-kinase anchoring protein,AKAP95. Exp. Cell Res. 238, 305–316.
2. Zhang, Q., Carr, D. W., Lerea, K. M., Scott, J. D., and Newman,S. A. (1996). Nuclear localization of type II cAMP-dependentprotein kinase during limb cartilage differentiation is associ-ated with a novel developmentally regulated A-kinase anchor-ing protein. Dev. Biol. 176, 51–61.
3. McCartney, S., Little, B. M., Langeberg, L. K., and Scott, J. D.(1995). Cloning and characterization of A-kinase anchor protein100 (AKAP100). A protein that targets A-kinase to the sarco-plasmic reticulum. J. Biol. Chem. 270, 9327–9333.

2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
5
5
145RIIa AND RIIb IN THE GOLGI–CENTROSOMAL AREA
4. Lester, L. B., Coghlan, V. M., Nauert, B., and Scott, J. D. (1996).Cloning and characterization of a novel A-kinase anchoringprotein. AKAP 220, association with testicular peroxisomes.J. Biol. Chem. 271, 9460–9465.
5. Rios, R. M., Celati, C., Lohmann, S. M., Bornens, M., andKeryer, G. (1992). Identification of a high affinity binding pro-tein for the regulatory subunit RIIb of cAMP-dependent proteinkinase in Golgi enriched membranes of human lymphoblasts.EMBO J. 11, 1723–1731.
6. Lin, R.-Y., Moss, S. B., and Rubin, C. S. (1995). Characteriza-tion of S-AKAP84, a novel developmentally regulated A kinaseanchor protein of male germ cells. J. Biol. Chem. 270, 27804–27811.
7. Trendelenburg, G., Hummel, M., Riecken, E.-O., and Hanski, C.(1996). Molecular characterization of AKAP149, a novel A ki-nase anchor protein with a KH domain. Biochem. Biophys. Res.Commun. 225, 313–319.
8. Keryer, G., Rios, R. M., Landmark, B. F., Skålhegg, B. S.,Lohmann, S. M., and Bornens, M. (1993). A high-affinity bind-ing protein for the regulatory subunit of cAMP-dependent pro-tein kinase II in the centrosome of human cells. Exp. Cell Res.204, 230–240.
9. Nauert, J. B., Klauck, T. M., Langeberg, L. K., and Scott, J. D.(1997). Gravin, an autoantigen recognized by serum from my-asthenia gravis patients, is a kinase scaffold protein. Curr. Biol.7, 52–62.
0. Gray, P. C., Johnson, B. D., Westenbroek, R. E., Hays, L. G.,Yates, J. R., Scheuer, T., Catterall, W. A., and Murphy, B. J.(1998). Primary structure and function of an A kinase anchor-ing protein associated with calcium channels. Neuron 20, 1017–1026.
1. Fraser, I. D., Tavalin, S. J., Lester, L. B., Langeberg, L. K.,Westphal, A. M., Dean, R. A., Marrion, N. V., and Scott, J. D.(1998). A novel lipid-anchored A-kinase anchoring protein fa-cilitates cAMP-responsive membrane events. EMBO J. 17,2261–2272.
2. De Camilli, P., Moretti, M., Donini, S. D., Walter, U., andLohmann, S. M. (1986). Heterogeneous distribution of thecAMP receptor protein RII in the nervous system: Evidence forits intracellular accumulation on microtubules, microtubule-organizing centers, and in the area of the Golgi complex. J. CellBiol. 103, 189–203.
3. Glantz, S. B., Amat, J. A., and Rubin, C. S. (1992). cAMPsignaling in neurons: Patterns of neuronal expression and in-tracellular localization for a novel protein, AKAP 150, thatanchors the regulatory subunit of cAMP-dependent protein ki-nase IIb. Mol. Biol. Cell 3, 1215–1228.
4. Nigg, E. A., Schafer, G., Hilz, H., and Eppenberger, H. M.(1985). Cyclic-AMP-dependent protein kinase type II is associ-ated with the Golgi complex and centrosomes. Cell 41, 1039–1051.
5. Griffiths, G., Hollinshead, R., Hemmings, B. A., and Nigg, E. A.(1990). Ultrastructural localization of the regulatory (RII) sub-unit of cyclic AMP-dependent protein kinase to subcellular com-partments active in endocytosis and recycling of membranereceptors. J. Cell Sci. 96, 691–703.
6. Singh, A. K., Tasken, K., Walker, W., Frizzell, R. A., Watkins,S. C., Bridges, R. J., and Bradbury, N. A. (1998). Characteriza-tion of PKA isoforms and kinase-dependent activation of chlo-ride secretion in T84 cells [in process citation]. Am. J. Physiol.275, C562–C570.
7. Scott, J. D., Glaccum, M. B., Zoller, M. J., Uhler, M. D., Helf-man, D. M., McKnight, G. S., and Krebs, E. G. (1987). Themolecular cloning of a type II regulatory subunit of the cAMP-dependent protein kinase from rat skeletal muscle and mousebrain. Proc. Natl. Acad. Sci. USA 84, 5192–5196.
8. Øyen, O., Myklebust, F., Scott, J. D., Hansson, V., and Jahnsen,T. (1989). Human testis cDNA for the regulatory subunit RIIa ofcAMP-dependent protein kinase encodes an alternate amino-terminal region. FEBS Lett. 246, 57–64.
9. Stein, J. C., Sarkar, D., and Rubin, C. S. (1984). Tryptic peptidemapping studies on the regulatory subunits of type II proteinkinases from cerebral cortex and heart. Evidence for overallstructural divergence and differences in the autophosphoryla-tion and cAMP-binding domains. J. Neurochem. 42, 547–553.
0. Jahnsen, T., Hedin, L., Lohmann, S. M., Walter, U., and Rich-ards, J. S. (1985). The neural type II regulatory subunit ofcAMP-dependent protein kinase is present and regulated byhormones in the rat ovary. J. Biol. Chem. 261, 6637–6639.
1. Øyen, O., Sandberg, M., Eskild, W., Levy, F. O., Knutsen, G.,Beebe, S., Hansson, V., and Jahnsen, T. (1988). Differentialregulation of messenger ribonucleic acids for specific subunitsof cyclic adenosine 39,59-monophosphate (cAMP)-dependentprotein kinase by cAMP in rat Sertoli cells. Endocrinology 122,2658–2666.
2. Schwartz, D. A., and Rubin, C. S. (1985). Identification anddifferential expression of two forms of regulatory subunits (RII)of cAMP-dependent protein kinase II in Friend erythroleuke-mic cells. J. Biol. Chem. 260, 6296–6303.
3. Ally, S., Clair, T., Katsaros, D., Tortora, G., Yokozaki, H., Finch,R. A., Avery, T. L., and Cho-Chung, Y. S. (1989). Inhibition ofgrowth and modulation of gene expression in human lung car-cinoma in athymic mice by site-selective 8-Cl-cyclic adenosinemonophosphate. Cancer Res. 49, 5650–5655.
4. Bradbury, A. W., Carter, D. C., Miller, W. R., Cho-Chung, Y. S.,and Clair, T. (1994). Protein kinase A (PK-A) regulatory sub-unit expression in colorectal cancer and related mucosa. Br. J.Cancer 69, 738–742.
5. Rohlff, C., Clair, T., and Cho-Chung, Y. S. (1993). 8-Cl-cAMPinduces truncation and down-regulation of the RI alpha subunitand up-regulation of the RII beta subunit of cAMP-dependentprotein kinase leading to type II holoenzyme-dependent growthinhibition and differentiation of HL-60 leukemia cells. J. Biol.Chem. 268, 5774–5782.
6. Tortora, G., Clair, T., Katsaros, D., Ally, S., Colamonici, O.,Neckers, L. M., Tagliaferri, P., Jahnsen, T., Robins, R. K., andCho-Chung, Y. S. (1989). Induction of megakaryocytic differen-tiation and modulation of protein kinase gene expression bysite-selective cAMP analogs in K-562 human leukemic cells.Proc. Natl. Acad. Sci. USA 86, 2849–2852.
7. Cummings, D. E., Brandon, E. P., Planas, J. V., Motamed, K.,Idzerda, R. L., and McKnight, G. S. (1996). Genetically leanmice result from targeted disruption of the RII beta subunit ofprotein kinase A. Nature 382, 622–626.
8. Johnson, B. D., Scheuer, T., and Catterall, W. A. (1994). Volt-age-dependent potentiation of L-type Ca21 channels in skeletalmuscle cells requires anchored cAMP-dependent protein ki-nase. Proc. Natl. Acad. Sci. USA 91, 11492–11496.
9. Hausken, Z. E., Dell’Acqua, M. L., Coghlan, V. M., and Scott,J. D. (1996). Mutational analysis of the A-kinase anchoringprotein (AKAP)-binding site on RII. Classification of side chaindeterminants for anchoring and isoform selective associationwith AKAPs. J. Biol. Chem. 271, 29016–29022.
0. Levy, F. O., Øyen, O., Sandberg, M., Tasken, K., Eskild, W.,Hansson, V., and Jahnsen, T. (1988). Molecular cloning, com-plementary deoxyribonucleic acid structure and predicted full-length amino acid sequence of the hormone-inducible regula-tory subunit of 39,59-cyclic adenosine monophosphate-dependent protein kinase from human testis. Mol. Endocrinol.2, 1364–1373.
1. Bailly, E., Doree, M., Nurse, P., and Bornens, M. (1989).p34cdc2 is located in both nucleus and cytoplasm; part is cen-

5
5
5
5
5
5
5
5
6
6
6
6
6
6
6
6
6
6
7
7
7
7
7
7
RR
146 KERYER ET AL.
trosomally associated at G2/M and enters vesicles at anaphase.EMBO J. 8, 3985–3995.
2. Jasmin, B. J., Cartaud, J., Bornens, M., and Changeux, J. P.(1989). Golgi apparatus in chick skeletal muscle: Changes in itsdistribution during end plate development and after denerva-tion. Proc. Natl. Acad. Sci. USA 86, 7218–7222.
3. Tasken, K., Skålhegg, B. S., Solberg, R., Andersson, K. B.,Taylor, S. S., Lea, T., Blomhoff, H. K., Jahnsen, T., and Hans-son, V. (1993). Novel isozymes of cAMP-dependent protein ki-nase exist in human cells due to formation RIa–RIb het-erodimeric complexes. J. Biol. Chem. 268, 21276–21283.
4. Skålhegg, B. S., Landmark, B. F., Foss, K. B., Lohmann, S. M.,Hansson, V., Lea, T., and Jahnsen, T. (1992). Identification,purification, and characterization of subunits of cAMP-depen-dent protein kinase in human testis. Reverse mobilities of hu-man RII alpha and RII beta on sodium dodecyl sulfate–poly-acrylamide gel electrophoresis compared with rat and bovineRIIs. J. Biol. Chem. 267, 5374–5379.
5. Foss, K. B., Solberg, R., Simard, J., Myklebust, F., Hansson, V.,Jahnsen, T., and Tasken, K. (1997). Molecular cloning, up-stream sequence and promoter studies of the human gene forthe regulatory subunit RIIa of cAMP-dependent protein kinase.Biochim. Biophys. Acta 1350, 98–108.
6. Stephens, R. E., and Prior, G. (1992). Dynein from serotonin-activated cilia and flagella: Extraction characteristics and dis-tinct sites for cAMP-dependent protein phosphorylation. J. CellSci. 103, 999–1012.
7. Prasad, K. N. (1991). Differentiation of neuroblastoma cells: Auseful model for neurobiology and cancer. Biol. Rev. CambridgePhilos. Soc. 66, 431–451.
8. Keryer, G., Alsat, E., Tasken, K., and Evain-Brion, D. (1998).Cyclic AMP-dependent protein kinases and human trophoblastcell differentiation in vitro. J. Cell Sci. 111, 995–1004.
9. Saltarelli, D., Crosnier, L., Mallasine, A., and Keryer, G. (1996).Mol. Cell. Biol. 7, 10a.
0. Rios, R. M., Tassin, A. M., Celati, C., Antony, C., Boissier, M. C.,Homberg, J. C., and Bornens, M. (1994). A peripheral proteinassociated with the cis-Golgi network redistributes in the in-termediate compartment upon brefeldin A treatment. J. CellBiol. 125, 997–1013.
1. Kreis, T. E. (1990). Role of microtubules in the organisation ofthe Golgi apparatus. Cell Motil. Cytoskeleton 15, 67–70.
2. De Brabander, M., Geuens, G., Nuydens, R., Willebrords, R.,and De Mey, J. (1981). Taxol induces the assembly of freemicrotubules in living cells and blocks the organizing capacityof the centrosomes and kinetochores. Proc. Natl. Acad. Sci. USA78, 5608–5612.
3. Pelham, H. R. (1991). Multiple targets for brefeldin A. Cell 67,449–451.
4. Banting, G., and Ponnanbalam, S. (1997). TGN38 and its or-thologues: Roles in post-TGN vesicle formation and mainte-
nance of TGN morphology. Biochim. Biophys. Acta 1355, 209–217.
5. Jahnsen, T., Lohmann, S. M., Walter, U., Hedin, L., and Rich-ards, J. S. (1985). Purification and characterization of hormone-regulated isoforms of the regulatory subunit of type II cAMP-dependent protein kinase from rat ovaries. J. Biol. Chem. 260,15980–15987.
6. Ratoosh, S. L., Lifka, J., Hedin, L., Jahnsen, T., and Richards,J. S. (1987). Hormonal regulation of the synthesis and mRNAcontent of the regulatory subunit of cyclic AMP-dependent pro-tein kinase type II in cultured rat ovarian granulosa cells.J. Biol. Chem. 262, 7306–7313.
7. Tasken, K. A., Knutsen, H. K., Attramadal, H., Tasken, K.,Jahnsen, T., Hansson, V., and Eskild, W. (1991). Differentmechanisms are involved in cAMP-mediated induction ofmRNAs for subunits of cAMP-dependent protein kinases. Mol.Endocrinol. 5, 21–28.
8. Landmark, B. F., Fauske, B., Eskild, W., Skålhegg, B. S., Loh-mann, S. M., Hansson, V., Jahnsen, T., and Beebe, S. J. (1991).Identification, characterization, and hormonal regulation of39,59-cyclic adenosine monophosphate-dependent protein ki-nases in rat Sertoli cells. Endocrinology 129, 2345–2354.
9. Carr, D. W., DeManno, D. A., Atwood, A., Hunzicker-Dunn, M.,and Scott, J. D. (1993). Follicle-stimulating hormone regulationof A-kinase anchoring proteins in granulosa cells. J. Biol.Chem. 268, 20729–20732.
0. Tassin, A. M., Maro, B., and Bornens, M. (1985). Fate of micro-tubule-organizing centers during myogenesis in vitro. J. CellBiol. 100, 35–46.
1. Eker, P., Holm, P. K., van Deurs, B., and Sandvig, K. (1994).Selective regulation of apical endocytosis in polarized Madin–Darby canine kidney cells by mastoparan and cAMP. J. Biol.Chem. 269, 18607–18615.
2. Tash, J. S., Means, A. R., Brinkley, B. R., Dedman, J. R., andCox, S. M. (1980). In “Microtubules and Microtubule Inhibitors”(M. De Brabander and J. Demey, Eds.), pp. 269–279, Elsevier–North Holland, Amsterdam.
3. Lane, M. E., and Kalderon, D. (1994). RNA localization alongthe anteroposterior axis of the Drosophila oocyte requires PKA-mediated signal transduction to direct normal microtubule or-ganization. Genes Dev. 8, 2986–2995.
4. Keryer, G., Yassenko, M., Labbe, J. C., Castro, A., Lohmann,S. M., Evain-Brion, D., and Tasken, K. (1998). Mitosis-specificphosphorylation and subcellular redistribution of the RIIalpharegulatory subunit of cAMP-dependent protein kinase. J. Biol.Chem. 273, 34594–34602.
5. Skålhegg, B. S., Landmark, B. F., Døskeland, S. O., Hansson,V., Lea, T., and Jahnsen, T. (1992). Cyclic AMP-dependentprotein kinase type I mediates the inhibitory effects of 39,59-cyclic adenosine monophosphate on cell replication in human Tlymphocytes. J. Biol. Chem. 267, 15707–15714.
eceived October 2, 1998evised version received February 8, 1999
Copyright © 2022 FDOKUMEN




















