
Household income modifies the association of insurance and dental visiting
Upload
independentCategory
view
10download
0
Differential Alteration of Heat Shock Protein 90in Mice Modifies Glucocorticoid Receptor Function
and Susceptibility to Trauma
Hai-Ying Shen,1,2 Yan Zhao,1 Xing-Yun Chen,1 Ren-Ping Xiong,1 Jin-Li Lu,1
Jiang-Fan Chen,2 Li-Yong Chen,3 and Yuan-Guo Zhou1
Abstract
Heat shock protein 90 (Hsp90), encoded by the murine hsp84 and hsp86 genes in mice, is a pivotal regulator ofglucocorticoid receptor (GR) function in the hypothalamus-pituitary-adrenal axis and affords stress protection.To explore the underlying molecular mechanisms of strain susceptibility to traumatic stress, we investigated thealteration by Hsp90 of the function of the glucocorticoid-glucocorticoid receptor (GC-GR) pathway in attenu-ating stress responses in C57BL=6 and BALB=c mice using the whole-body blast injury (WBBI) model. We foundthat C57BL=6 mice had a lower WBBI-induced mortality, higher nuclear GR level, and higher glucocorticoid-response element (GRE) binding activity than BALB=c mice. This study is the first report identifying four geneticvariations of the murine hsp84 gene: 226A>C, 996G>C, 1483G>C, and 2000G>T. These nucleotide changesoccur in the functional domains associated with the nuclear=cytosolic translocation of GR, GR-Hsp90 interaction,ATP binding, and self-dimerization of Hsp90, respectively. Further, we used a specific Hsp90 inhibitor, gelda-namycin (GA), to assess the role of Hsp90 in the discriminative traumatic response in C57BL=6 mice. Pre-treatment with GA reduced nuclear GR levels and GRE binding activity, and enhanced WBBI-induced mortality.These findings suggest that Hsp90 may underlie the strain-selective (C57BL=6 versus BALB=c) susceptibility toWBBI by mediating the nuclear translocation of GRs and GRE binding. Thus, pharmacological manipulation ofHsp90 may represent a therapeutic strategy to modify the function of the GC-GR pathway and traumatic stressresponse.
Key words: geldanamycin; glucocorticoid receptor; heat shock protein; traumatic susceptibility; whole-body blastinjury
Introduction
Disturbances in the body evoke a stress response andlead to restoration of homeostasis and facilitation of
adaptive processes via the hypothalamic-pituitary-adrenal(HPA) axis (De Kloet et al., 1998, 2005), which is modulated byneuronal and endocrine interactions. Corticosteroids, whichare predominant glucocorticoids, facilitate the body’s abilityto cope with, adapt to, and recover from stress by maintainingbasal activity of the HPA system, controlling the sensitivityof the system’s response to stress, and terminating stress-induced HPA activation by a feedback mechanism. Dys-function in any of the stress mediators affecting HPA axisactivity, such as glucocorticoid (GC) levels, glucocorticoid
receptor (GR) function, or GR-related chaperone proteinfunction, may lead to pathological consequences in the stressresponse (Reichardt et al., 2000; Sapolsky et al., 1986). Heatshock protein 90 (Hsp90), a highly abundant ubiquitouschaperon protein, is essential for GR function, includingmaturation, conformation maintenance, ligand conjugation,nuclear=cytosolic translocation, and intranuclear circulationof GR (Csermely et al., 1998; Whitesell and Cook, 1996). Hsp90has been proposed to play a pivotal role in stress protection byinfluencing HPA-GR axis activity through the GC-GR path-way (Liu and DeFranco, 1999; Pearl and Prodromou, 2001;Tago et al., 2004). Mouse Hsp90s are encoded by the hsp84and hsp86 genes with different genomic locations and dif-ferences in tissue distribution of the resulting gene products
1Molecular Biology Center, State Key Laboratory of Trauma, Burn and Combined Injury, and 3Department of Anesthesiology, ResearchInstitute of Surgery and Daping Hospital, Third Military Medical University, Chongqing, P.R. China.
2Department of Neurology, Boston University School of Medicine, Boston, Massachusetts.
JOURNAL OF NEUROTRAUMA 27:373–381 (February 2010)ª Mary Ann Liebert, Inc.DOI: 10.1089=neu.2009.0926
373

(i.e., constitutive Hsp84 is predominantly expressed in thebrain, whereas the more inducible Hsp86 is enriched in pe-ripheral organs) (Csermely et al., 1998; Pearl and Prodromou,2001; Ramaglia and Buck, 2004).
Though the modulation of Hsp90 activity has been dis-cussed as a means to alter phenotype in certain traits andorganisms (Sangster et al., 2008), the importance of Hsp90 indetermining the amplitude of the traumatic stress responsehas not been directly assessed. Further, mutational deletionanalysis of engineered Hsp90 variants on GR functions havebeen extensively discussed at the cellular level (Grenert et al.,1997; Jibard et al., 1999; Nemoto et al., 1995; Soti et al., 2003).In the present study, we employed the whole-body blast in-jury (WBBI) traumatic model to test our hypothesis that al-teration of Hsp90 may result in different traumatic responsesby modifying GR function. Distinctly differing mortality ratesbetween BALB=c and C57BL=6 mice as a consequence ofWBBI led us to further evaluate polymorphisms in the hsp84gene at the genomic DNA level. We also studied the potentialassociation between hsp84 polymorphisms and alterations inthe GC-GR signaling pathway, by determining the expressionand subcellular distribution of GR and Hsp90, and gluco-corticoid-response element (GRE) binding activity afterWBBI. Finally, geldanamycin (GA), a specific Hsp90 inhibitor,was used to validate the effect of Hsp90 inhibition on trau-matic stress responses in mice.
Methods
Animals
A total of 317 pathogen-free, 56- to 70-day-old maleC57BL=6 and BALB=c mice (Shanghai Laboratorial AnimalCenter, China) were used in the present study. Mice (weight22–27 g) were housed individually with free access to foodand water in a temperature- and humidity-controlled roomunder a 12-h light=dark cycle for 10 days prior to WBBI. Allanimal experiments were performed in accordance with theRegulations for the Administration of Affairs ConcerningExperimental Animals Guideline of Third Military MedicalUniversity, and were consistent with the NIH guidelines forthe care and use of laboratory animals.
Drug treatment and WBBI model
Geldanamycin (GA; Sigma, St Louis, MO) was dissolved in10% DMSO and 90% saline for IP injection of different doses(1, 10, or 100 mg=kg), in a volume of 0.1 mL=10 g body weight,3 h prior to WBBI.
The bio-shock tube (BST-I) apparatus was used to produceWBBI as previously described (Cernak et al., 2001a; Wang,1989) with minor modifications. Briefly, prior to WBBI, themice were placed into individual cages (1 mouse=per cage)within a cage-array in which all the cages were positioned atthe same vertical plane inside the BST-I apparatus to ensureequal pressure exposure (Fig. 1A). Specifically, mice of dif-ferent strains or treatments were arranged in a checkerboardfashion to ensure equal exposure (Fig. 1B). For a more realisticeffect, all the mice were unanesthetized and free to moveduring WBBI. The peak of positive pressure of the BST-I was498.76� 23.72 kPa, lasting for 2.75� 0.03 msec (mean� SEMfrom 8 WBBI experiments). The entire high-pressure period ofthe WBBI model lasted for 56.93� 0.38 msec (Fig. 1C). After
WBBI, all mice, except those used for subsequent mortalityassessment, were sacrificed at different time points (0.5 to72 h) to collect blood and tissue samples for further assays.
Serum corticosterone measurement
To evaluate the systemic changes in glucocorticoid levels,serum corticosterone levels were determined with a com-mercial radioimmunoassay kit according to the manufac-turer’s protocol (ICN, Meckenheim, Germany).
Western blot of Hsp90 and GR
Nuclear protein and whole-cell lysate (WCL) were isolatedfrom brain tissue as previously described (Shen et al., 2005)with modifications. Nuclear protein (50mg) or WCL (30mg) waselectrophoresed in a 10–20% gradient Tris-glycine gel. Afterblotting, the membranes were incubated with antibodiesagainst Hsp90 (1:500, sc-7947; Santa Cruz Biotechnology, SantaCruz, CA), or GR (1:500, sc-8992; Santa Cruz Biotechnology),and then incubated with peroxidase-conjugated anti-rabbitantibody (1:5000; Santa Cruz Biotechnology). The opticaldensities of the Hsp90 and GR immunoreactivities were mea-sured by an UVP BioImaging acquisition and image analysissystem (Ultra-Violet Products Ltd., Cambridge, U.K.). Anti-bodies against b-actin (1:5000, A3854; Sigma), and GAPDHantibody (1:5000, ab9484; Abcam, Cambridge, MA) were usedto re-probe the same blot after stripping of the membrane tonormalize Hsp90 and GR immunoreactivity.
Gel shift assay
Gel shift assays were carried out as previously described(Shen et al., 2005) with modifications. Nuclear extracts (15mgprotein) were incubated with a double-stranded GRE oligo-nucleotide probe (50-CCATATGCAGCCACTGTTCTTAGCGGGTTT-30; TaKaRa, Dalian, China) in 10 mL binding buffer(20 mM HEPES pH 7.8, 50 mM KCl, 1 mM EDTA, 10% glyc-erol, 1 mM DTT, and 50 mg=mL poly dI-dC) in an ice=waterbath for 10 min. A 32P-labeled GRE oligonucleotide probe wasadded and the mixture was incubated for another 20 min at258C. Then the protein-DNA complexes were submitted to a5% non-denaturing PAGE gel. To determine the specificity ofthe protein-DNA complex, the samples were incubated with a50-fold molar excess of unlabeled GRE, or 1mg of specific GRantibody (H-300, sc-8992X; Santa Cruz Biotechnology) for15 min before adding labeled GRE probe. Autoradiographywas carried out by exposing the dried gel to film. DNAbinding activities were estimated using the UVP BioImagingAnalyzer as described above.
Rapid amplification of 50 cDNA end (5 0 RACE)and 3 0 cDNA end (30 RACE)
Mouse total RNA was isolated using TRIzol reagent (In-vitrogen, Carlsbad, CA) according to the manufacturer’sprotocol. RNA concentration and integrity were assessed byspectrophotometry and gel electrophoresis, and then the totalRNA was used for reverse transcription reaction to producetotal cDNA of mice. The 50 end and 30 end of the mouse hsp84cDNA were amplified with primers (Table 1) according to themanufacturer’s protocol of the TaKaRa RACE kit, using totalcDNA from the brains of BALB=c or C57BL=6 mice. After de-tection in 1% agarose gels stained with ethidium bromide,
374 SHEN ET AL.

the PCR products were purified and subcloned into the pBlue-script KSþ vector (Strategene, LaJolla, CA) for sequencing.The sequencing was carried out in the DNA Sequencing CoreFacility of TaKaRa and the Massachusetts General HospitalDNA Core Facility (Cambridge, MA). All DNA samples weresequenced from both strands to ensure accuracy.
Identification of polymorphisms of the hsp84 gene
A total of 2452 bp of full-length hsp84 cDNA, including 50
upstream of the authentic ATG translation initiation codonand 30 UTR region, was obtained. Sequence comparisons ofhsp84 cDNA were made with the Basic Local Alignment
Table 1. Primers Used in PR-PCR, gDNA PCR, and RACE
RT-PCR primerssense 50-ACTACTCGGCTTTCTCGT-30 (85-102 of M18186)antisense 50-CCAACCCTGCTATCTGTG-30 (2437-2419 of M18186)
gDNA PCR primersHspcb-f1 50-CCTCCTTGTCAGAGTATGTG-30
Hspcb-r1 50-CGTCAATAGGCTCAGTCAT-30
Hspcb-f2 50-CCAACCGCATCTACCGC-30
Hspcb-r2 50-GGCAGGGCTTCTTCCAG-30
50RACE primersAnchor primer (s-rt3) 50-(P)CCATGAACGCCTTCG-30
First PCR primerss-f1 50-AATTGACATCATCCCCAACCCTC-30
s-r1 50-TTGTTCAGGGCATCTGAAGCATT-30
Second nested PCR primerss-f2 50-GGTGGACACAGGCATTGGCATGA-30
s-r2 50-GGACATGAGCTGGGCAATTTCTG-30
30RACE primerss-30-f 50-CCAAACCCACTCCAACCGCATCT-30
PR-PCR, proofreading polymerase chain reaction; gDNA PCR, genomic DNA polymerase chain reaction; RACE, rapid amplification ofcomplementary DNA ends; RT-PCR real-time polymerase chain reaction.
FIG. 1. The bio-shock tube (BST-I) apparatus mimics the whole-body blast injury (WBBI) model. (A) Photo illustrating theBST-I apparatus and the position of the cage-array in the same vertical plane to ensure equal exposure to WBBI for all mice.(B) Photo illustrating that the mice were arranged in a checkerboard fashion. (C) Graph showing the dynamic pattern ofpressure change during the WBBI model.
HSP90 AFFECTS TRAUMATIC RESPONSES 375

Search Tool (BLASTN) at the National Center for Bio-technology Information (NCBI). The hsp84 cDNA sequence ofC57BL=6 reported in this study has been deposited in theGenBank database (accession no. AF465639).
To confirm the strain-specific variations of the hsp84 allele,the genomic DNA of BALB=c and C57BL=6 mice was isolatedby digestion with proteinase K (0.2 mg=mL), in the presence of1% SDS in TE buffer (0.5 M Tris-HCl [pH 9.0], 20 mM EDTA,and 10 mM NaCl) at 508C overnight, followed by phenol=chloroform extraction and ethanol precipitation. PCR ampli-fication and sequencing were performed as described above.Two sets of primers were used to identify polymorphisms(Table 1).
Statistical analysis
All data are presented as mean� SEM and were derivedusing the Student’s t-test, one-way ANOVA with post-hocBonferroni testing, or log-rank testing, as appropriate. Inall analyses, p< 0.05 was considered to be statistically sig-nificant.
Results
Different severities of WBBI-induced injuries betweenBALB=c and C57BL=6 mice correlate with distinctnuclear GR levels and GRE-binding activities
To demonstrate strain-selective susceptibility to traumaticinjury, the WBBI-induced mortalities of C57BL=6 and BALB=cmice were evaluated at different time points (0.5, 6, 12, 24, 48,and 72 h). The cumulative mortality over a time span of 72 hfollowing WBBI in BALB=c mice (24.7%, 13=54) was signifi-cantly higher than that of C57BL=6 mice (5.66%, 3=53)( p< 0.05; Fig. 2A). Notably, the mortality difference withinthe first 6 h after WBBI between C57BL=6 (3.77%) and BALB=c(16.77%) mice was most prominent ( p< 0.01; Fig. 2A).
The GC-GR pathway function was evaluated by measure-ments of serum levels of corticosterone (a glucocorticoid se-creted in response to stress), total and nuclear GR levels, andGRE-binding activity. Serum corticosterone levels (10–7 M)rose to a significant peak at 0.5 h post-WBBI (8.22� 0.92 inC57BL=6 mice, n¼ 8; 9.36� 1.21 in BALB=c mice, n¼ 7), thendecreased gradually in both strains (n¼ 8) at 6, 12, 24, and 72 hpost-WBBI (Fig. 2B). There was no difference in serum corti-costerone levels between strains at each time point assayed( p> 0.05). Total GR levels in both strains were increased at0.5 h, peaked at 6 h post-WBBI, and returned to basal levels at24 h post-WBBI (n¼ 6 per group; Fig. 2C). No statisticallysignificant difference was observed between strains at eachtime point tested. Nuclear GR levels in both strains of micewere also increased at 0.5 h and 6 h post-WBBI (n¼ 6, p< 0.05;Fig. 2D), inducing nuclear distribution of GR after traumaticinjury. Most interestingly, C57BL=6 mice showed higher nu-clear GR levels at 0.5 h and 6 h post-WBBI (n¼ 6, p< 0.05),which indicates a different stress response on the GC-GRpathway between BALB=c and C57BL=6 mice (Fig. 2D). Fur-ther, C57BL=6 mice showed a higher GRE-binding activity at6 h post-WBBI (n¼ 6, p< 0.05), while the two strains of miceshowed similar basal (pre-WBBI) GRE-binding activity (Fig.2E). The correlation of increased nuclear GR level and en-hanced GRE binding activity, with a lower WBBI-inducedmortality and slight pathological changes, such as swellingof neurons and pulmonary edema as previously reported(Cernak et al., 2001a; Wang, 1989) in C57BL=6 mice, indicatesa relationship between the higher responsiveness of theHPA-GR pathway and reduced severity of traumatic injury.
Genetic variations of hsp84 found in BALB=cand C57BL=6 mice
To explore the underlying role of Hsp90 in the modulationof GR nuclear translocation, total and nuclear Hsp90 levels in
FIG. 2. Whole-body blast injury (WBBI)-induced changes of mortality, serum corticosterone levels and glucocorticoidreceptor (GR) function between mouse strains. (A) The cumulative post-WBBI mortalities at different time points (ap< 0.01and bp< 0.05 at 6 h and 72 h post-WBBI, respectively, by log-rank testing). (B) Serum corticosterone levels at 0.5, 6, 12, and24 h (n¼ 8 C57BL=6 and n¼ 7 BALB=c mice), and 72 h post-WBBI (n¼ 8 per group; §p< 0.05 and Dp< 0.05 compared to thenon-WBBI group within each strain). (C) Total and (D) nuclear GR levels of both strains, at 0 (non-WBBI), 0.5, 6, and 24 hpost-WBBI (n¼ 6 per group). (E) Glucocorticoid-response element (GRE) binding activity at 0 (non-WBBI) and 6 h post-WBBI(n¼ 6; *p< 0.05 compared to the non-WBBI group within each strain; #p< 0.05 compared to the corresponding groups ofBALB=c mice; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; ROD, relative optical density).
376 SHEN ET AL.

the brain were evaluated by Western blot. Total Hsp90 levelsin C57BL=6 mice were increased at 0.5, 6, and 24 h post-WBBIcompared with pre-WBBI (n¼ 6, p< 0.05). BALB=c mice dis-played a similar total Hsp90 induction time course post-WBBI(Fig. 3A). Meanwhile, nuclear Hsp90 levels in both strainswere increased at 24 h post-WBBI, while no difference wasevident between strains at any time point post-WBBI (Fig. 3B).
Since strain-specific changes in nuclear GR (Fig. 2D) couldnot be explained by strain-specific differences in the expres-sion levels of total or nuclear Hsp90, we hypothesized thatgenetic variations in the hsp84 gene, predominantly expressedin the brain, might account for the different functionality oftheir respective gene products. Thus, RT-PCR and RACE wereused to investigate the possible role of genetic variations ofhsp84 in GR-function regulation. Full-length cDNAs of hsp84were cloned and sequenced from brain tissue of C57BL=6 andBALB=c mice. The hsp84 cDNA sequence of BALB=c mice inour study was identical to M18186, a previously publishedhsp84 sequence from BALB=c mice (Moore et al., 1987). Thehsp84 cDNA sequence of the C57BL=6 mice reported in this
study has not been reported previously, and was deposited inthe GenBank (accession no. AF465639). The comparativeanalysis of AF465639 and M18186 revealed 14 nucleotidesubstitutions (Table 2). A total of 13 out of the 14 substitutionsoccurred in the open reading frame of hsp84. One (2358T>A)occurred in the 30 UTR region. Four missense mutations(226A>C, 996G>C, 1483G>C, and 2000G>T) are locatedwithin exons 2, 6, 9, and 10, respectively, so they are unlikelyto be affected by mRNA splicing. These variants are expectedto result in amino acid substitutions (Ile76>Leu, Leu332>Phe,Ala495>Pro, and Gly667>Val), that are located in differentdomains, and interfere with nuclear=cytosolic translocation ofGR, GR-Hsp90 interaction, ATP binding, and self-dimerizationof Hsp90, respectively. The above-mentioned four geneticvariations were also confirmed by PCR using genomic DNAtemplates isolated from brain, liver, and tail tissues, whichindicate that these four substitutions in AF465639 are genomicstrain-specific variations rather than post-transcriptionalmodifications. This finding indicates that the AF465639 geneis a new stable genetic variation of hsp84.
Enhancement of WBBI-induced mortality by GApretreatment correlates with reduced nuclear GR leveland decreased GRE-binding activity
Since the Ile76>Leu variation was found in the principalATP binding domain, which is also the interaction region ofGA with Hsp90, GA was used for impairment of Hsp90function, to further evaluate the effect of Hsp90 dysfunctionvia attenuation of ATP binding to Hsp90 in the GC-GRpathway, as well as the traumatic response after WBBI.
As expected, GA pretreatment increased WBBI-inducedmortality in C57BL=6 mice (Fig. 4A) in a dose-dependentmanner (5.66%þ 3=53, 7.50%þ 3=40, and 12.5%þ 4=32, fromgroups with vehicle only, 10 mg=kg GA, and 100 mg=kg GA,respectively). To explore the potential molecular meanswithin the GC-GR pathway that could account for the in-creased WBBI-induced injury after GA pretreatment, Hsp90levels were quantified after combined WBBI and GA treat-ment. Total and nuclear Hsp90 levels were upregulated byWBBI at 6 h post-WBBI ( p< 0.05), but were not affected byGA pretreatment (Fig. 4B and 4C). Similarly, GA pretreat-ment did not alter the enhancement of total GR levels at 6 hpost-WBBI ( p> 0.05; Fig. 4D). However, WBBI-induced en-hancement of nuclear GR levels was reduced by 10 mg=kg GApretreatment (n¼ 6, p< 0.05; Fig. 4E). GRE-binding activity inthe vehicle-pretreatment groups was enhanced by WBBI at 6 h(n¼ 6, p< 0.05; Fig. 4F), whereas 10-mg=kg GA pretreatmentreduced WBBI-induced enhancement of GRE-binding activity(n¼ 6, p< 0.05), in parallel with the reduction of nuclear GRlevels of GA-pretreated mice. The pharmacological inhibitionof Hsp90 results in changes in both nuclear GR levels and GREbinding, without affecting Hsp90 levels, suggesting a crucialrole for Hsp90-intrinsic properties in the modulation of theGC-GR pathway.
Discussion
The major finding of this study is the demonstration ofdifferent responsiveness of the GC-GR pathway in the HPAaxis, as well as different brain injury patterns and mortalityrates after WBBI between C57BL=6 and BABL=c mice. Thesedifferent WBBI-induced injuries and mortality rates were
FIG. 3. Whole-body blast injury (WBBI)-induced changesof total and nuclear Hsp90. (A) Total and (B) nuclear Hsp90levels of both strains, at 0 (non-WBBI), 0.5, 6, and 24 h post-WBBI (n¼ 6 per group; *p< 0.05 compared to the corre-sponding non-WBBI group within each strain; GAPDH,glyceraldehyde-3-phosphate dehydrogenase; OD, opticaldensity).
HSP90 AFFECTS TRAUMATIC RESPONSES 377
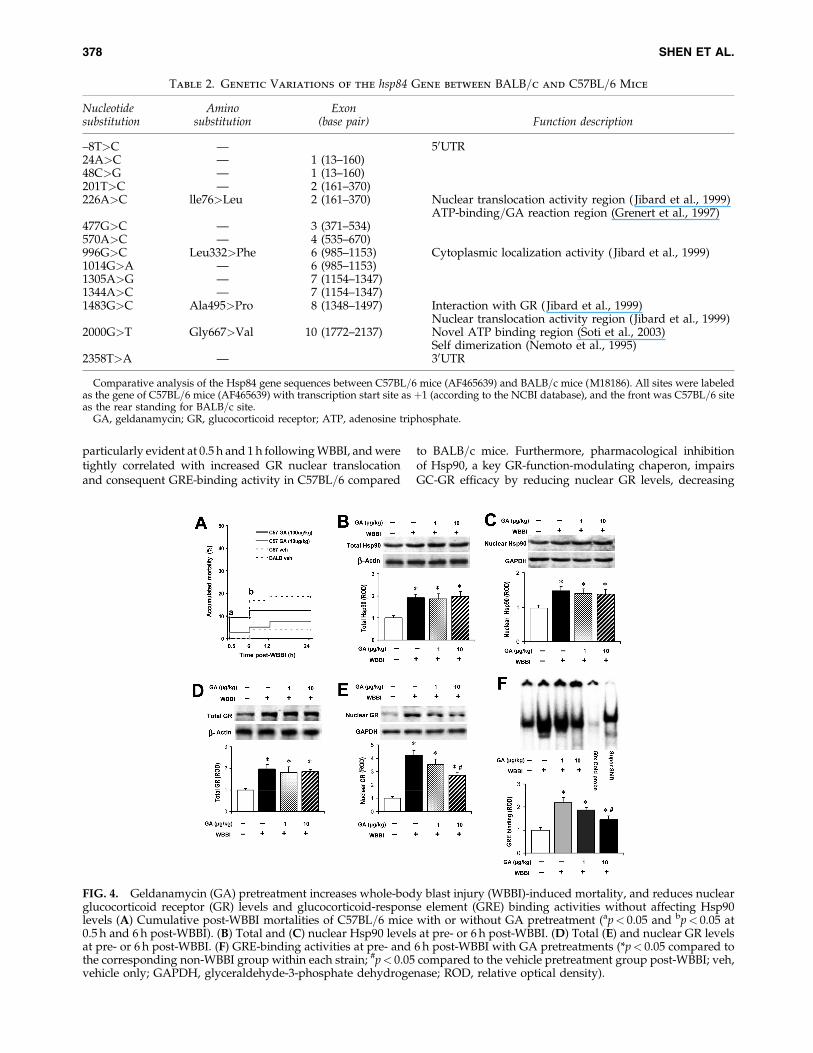
particularly evident at 0.5 h and 1 h following WBBI, and weretightly correlated with increased GR nuclear translocationand consequent GRE-binding activity in C57BL=6 compared
to BALB=c mice. Furthermore, pharmacological inhibitionof Hsp90, a key GR-function-modulating chaperon, impairsGC-GR efficacy by reducing nuclear GR levels, decreasing
Table 2. Genetic Variations of the hsp84 Gene between BALB=c and C57BL=6 Mice
Nucleotidesubstitution
Aminosubstitution
Exon(base pair) Function description
–8T>C — 50UTR24A>C — 1 (13–160)48C>G — 1 (13–160)201T>C — 2 (161–370)226A>C lle76>Leu 2 (161–370) Nuclear translocation activity region ( Jibard et al., 1999)
ATP-binding=GA reaction region (Grenert et al., 1997)477G>C — 3 (371–534)570A>C — 4 (535–670)996G>C Leu332>Phe 6 (985–1153) Cytoplasmic localization activity ( Jibard et al., 1999)1014G>A — 6 (985–1153)1305A>G — 7 (1154–1347)1344A>C — 7 (1154–1347)1483G>C Ala495>Pro 8 (1348–1497) Interaction with GR ( Jibard et al., 1999)
Nuclear translocation activity region (Jibard et al., 1999)2000G>T Gly667>Val 10 (1772–2137) Novel ATP binding region (Soti et al., 2003)
Self dimerization (Nemoto et al., 1995)2358T>A — 30UTR
Comparative analysis of the Hsp84 gene sequences between C57BL=6 mice (AF465639) and BALB=c mice (M18186). All sites were labeledas the gene of C57BL=6 mice (AF465639) with transcription start site as þ1 (according to the NCBI database), and the front was C57BL=6 siteas the rear standing for BALB=c site.
GA, geldanamycin; GR, glucocorticoid receptor; ATP, adenosine triphosphate.
FIG. 4. Geldanamycin (GA) pretreatment increases whole-body blast injury (WBBI)-induced mortality, and reduces nuclearglucocorticoid receptor (GR) levels and glucocorticoid-response element (GRE) binding activities without affecting Hsp90levels (A) Cumulative post-WBBI mortalities of C57BL=6 mice with or without GA pretreatment (ap< 0.05 and bp< 0.05 at0.5 h and 6 h post-WBBI). (B) Total and (C) nuclear Hsp90 levels at pre- or 6 h post-WBBI. (D) Total (E) and nuclear GR levelsat pre- or 6 h post-WBBI. (F) GRE-binding activities at pre- and 6 h post-WBBI with GA pretreatments (*p< 0.05 compared tothe corresponding non-WBBI group within each strain; #p< 0.05 compared to the vehicle pretreatment group post-WBBI; veh,vehicle only; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; ROD, relative optical density).
378 SHEN ET AL.

GRE-binding activity, and leading to increased WBBI-induced mortality. In addition, the present study identifiedfour genetic variations in the hsp84 gene between C57BL=6and BALB=c mice, which are located in several Hsp90 func-tional domains associated with regulation of GR activity (i.e.,GR nuclear=cytosolic translocation, GR-Hsp90 interaction,and self-dimerization and ATP binding activity of Hsp90)(Nemoto et al., 1995; Grenert et al., 1997; Jibard et al., 1999;Soti et al., 2003). Lastly, pretreatment with GA, an Hsp90 in-hibitor, was able to partially blunt strain-related high GC-GRefficiency and lower mortality after WBBI in C57BL=6 mice,via impairment of Hsp90 effects on GR activity. Together,these results suggest that Hsp84=Hsp90 may play an impor-tant role in the difference in susceptibility to traumatic injurybetween C57BL=6 and BALB=c mice via modulation of GRactivity.
C57BL=6 and BALB=c mice have different phenotypes intheir early post-traumatic changes (Radojicic et al., 1990),post-traumatic inflammatory reaction and wound healing (Liet al., 2001), and immune responses (Ulett et al., 2000). Thesedifferent pathological responses to trauma are suggested to bedirectly or indirectly related to the activity of the GC-GRpathway. We previously demonstrated that different WBBI-induced mortalities between C57BL=6 and BALB=c mice wascorrelated with the severity of neuronal pathological changes,such as swelling of neurons, gliosis, myelin debris, and pul-monary edema and hemorrhage (Cernak et al., 2001a, 2001b;Wang, 1989). The parallel post-traumatic changes in the lungand the brain implies that neuronal mechanisms may also beinvolved in WBBI-elicited pulmonary pathophysiologicalchanges (Colice et al., 1984). Indeed, activity of the HPA axisin the brain has been proposed to affect pulmonary responseand traumatic animal death (Bahloul et al., 2006; Rogerset al., 1995). While GC and GR levels have been tightly linkedto stress responses, adaptation, and HPA axis activity intraumatic injury (De Kloet et al., 1998, 2005), the underly-ing molecular mechanisms of strain-selective susceptibility totraumatic stress are not yet clear.
Our data demonstrated a surge of endogenous corticoste-rone after WBBI in two mouse strains, in line with the well-documented common post-traumatic response (De Kloet et al.,1996). Meanwhile, total GR levels were found to be upregu-lated in both mouse strains after WBBI. This temporal profileof GR changes was also observed in another trauma study, inwhich it was found that GR expression sharply increased at4 h and peaked at 8 h, with an increased GRE binding activitythat peaked at 4 h post-injury (Yan et al., 1999). However, ourfurther analysis displayed a similar dynamic pattern in post-traumatic serum corticosterone levels and total cellular GRlevels between the two strains of mice. This suggests thatchanges other than serum GC and total cellular GR levels areresponsible for the differences in GC-GR responsiveness in theHPA-GR pathway between these two mouse strains (Murphyet al., 2002; Rosenhagen et al., 2001). While total GR levelsmay differ depending on the nature and extent of traumaticexposure=stress stimulation (Liu et al., 2002; McCullers et al.,2002; Yan et al., 1999), nuclear GR levels (resulting from nu-clear translocation of GR) may better reflect HPA-GR functionthan total GR levels ( Joels, 2006; Liu and DeFranco, 1999).Interestingly, we demonstrated that nuclear GR levels areclosely correlated with the different mortality rates seen inthese two mouse strains.
Hsp90, a highly abundant ubiquitous chaperon protein,universally increases under most types of stress (Csermelyet al., 1998; Hashiguchi et al., 2001; Zhao et al., 2007). Themolecular structure, expression, and protein interaction ofHsp90 are believed to modulate GR functions, includingmaturation, ligand conjugation and nuclear=cytosolic trans-location of GR, and GR intranuclear circulation (Csermelyet al., 1998; Whitesell and Cook, 1996). The indistinguishablepost-traumatic dynamic pattern of cellular and nuclear Hsp90levels in brain between these two mouse strains suggests thatan intrinsic property of Hsp90, other than protein level, maybe responsible for their differences in post-traumatic GC-GRresponsiveness. Indeed, we also investigated the level ofHsp90 in liver and heart tissues after WBBI, and found thatthere was no significant difference between the protein ex-pression levels of Hsp90 in these two mouse strains afterWBBI (data not shown). Furthermore, we showed that inhi-bition of Hsp90 causes impairment of GR activity withoutaffecting Hsp90 expression levels. Rather, we postulated thatit is likely that the different functional and intrinsic propertiesof Hsp90 may account for the different nuclear GR levels seenin these two strains. The different functions in these strainsmay be partly due to different structural and biochemicalproperties of Hsp90 as result of Hsp90 genetic variations. Forexample, genetic variations in the ATP binding site of Hsp90may alter ATP binding affinity, and thus affect Hsp90’schaperon function, as suggested by some in-vitro studies(Grenert et al., 1997; Soti et al., 2003). Other post-translationalchanges such as Hsp90 hyperacetylation, without changingthe expression of Hsp90 and GR, can decrease Hsp90’s abilityto assemble stable GR-Hsp90 heterocomplexes, and thus itssubsequent steroid-binding activity, leading to reductions inGR-ligand binding, nuclear translocation, and gene activationin vitro (Murphy et al., 2005). Thus the different nuclear GRlevels and regulators affecting the nucleocytoplasmic traf-ficking of GR, as well as dynamic GR-Hsp90 heterocomplexassembly=disassembly, may contribute to the different levelsof efficiency of the GC-GR pathway seen in these two mousestrains. Further, the different mortality rates, predominantlydisplayed during the early post-traumatic phase (the first 6 hpost-WBBI), indicate that constitutively-expressed factorsplay dominant roles in the modulation of HPA-GR axis re-sponsiveness. Different temporal and anatomical features(Csermely et al., 1998; Pearl and Prodromou, 2001) of consti-tutive hsp84, which is predominantly expressed in the brain,may preferentially contribute to differences in the modulationof HPA activity post-WBBI.
Additionally, the genetic variations described here for thefirst time, occur in several crucial domains of Hsp90 related tothe regulation of GR activity and self-dimerization, and to theATP-binding activity of Hsp90 (Grenert et al., 1997; Jibardet al., 1999; Nemoto et al., 1995; Soti et al., 2003). TheIle76>Leu substitution in the principal ATP binding site, andthe Gly667>Val substitution in the supplemental ATP bind-ing site of Hsp90 (Scheibel et al., 1999; Soti et al., 2003), mayaffect GR ligand binding and GA-Hsp90 interactions, andmay thus destabilize GR, leading to a depletion of GR viadegradation, rather than reductions in Hsp90. This notion issupported by our preliminary finding from in-vitro culturedcells that cultured BALB=c cells expressing plasmid contain-ing the C57BL=6 hsp84 gene (pcD3-CH84) have an elevatedlevel of nuclear GRs compared with cultured BALB=c cells
HSP90 AFFECTS TRAUMATIC RESPONSES 379

expressing plasmid containing the BALB=c hsp84 gene (pcD3-BH84) (unpublished data, Zhao and Zhou). GA pretreatmentin this study provided direct evidence that impaired ATPbinding in Hsp90 led to altered HPA-GR function, reductionsin nuclear GR levels, and decreases in GRE-binding activity,as well as consequently higher WBBI-induced mortality. Thisis parallel with the post-traumatic phenotype of BALB=c micewith genetic variations in ATP-binding domains of Hsp84=Hsp90. Notably, GA-enhanced mortality mainly occurred inthe first 6 h post-WBBI, similarly to the time course of the dif-ferent mortality rates for BALB=c and C57BL=6 mice. Regard-ing the other three alterations of the murine hsp84 gene, thesevariations in C57BL=6 mice may act synergistically or sepa-rately to alter the HPA-GR pathway, resulting in higher nuclearGR levels and GRE-binding activity, allowing them to moreeasily adapt to WBBI-induced traumatic stress. Alternatively, itis possible that expression of Hsp90-associated proteins inthese strains may be different, and this may further amplify thedifferences in biochemical and functional properties of Hsp90,leading to different levels of GRs in the two strains. Additionalstudies are needed to address the exact mechanisms underly-ing the differences seen in these two mouse strains.
The data presented here reveal different GC-GR respon-siveness between two mouse strains, which may indicate dif-ferent WBBI-induced traumatic phenotypes. Different GC-GRfunctions may reflect different Hsp90 biochemical and struc-tural properties, such as genetic variations in critical functionaldomains. These findings help us to better understand the dif-fering traumatic=stress-related phenotypes of BALB=c andC57BL=6 mice (i.e., early post-traumatic responses) (Radojicicet al., 1990), that may affect post-traumatic inflammation,wound healing (Li et al., 2001), and immunoreactions to ecto-genic immunogens (Ulett et al., 2000). It also suggests thattargeting Hsp90 could be a potential means of modulating theefficiency of glucocorticoid therapy (i.e., in situations of glu-cocorticoid resistance and modulation of functional discrimi-nation of GR). Further, our results from strain-specific geneticvariations suggest that the genetic analysis of Hsp90=Hsp84may help diagnose the severity of traumatic injury. Last, phar-macological manipulation of Hsp90 indicates that it may bea potential target to modify the traumatic stress response.
Acknowledgments
This work was supported by grants from National NaturalScience Foundation of China (to H.Y.S., no. 30871170; and toY.G.Z., no. 30470988), the Foundation for the Author ofNational Excellent Doctoral Dissertation of P.R. China (no.200156), and National Basic Research Program of China (No.2005CB522602).
The authors thank Xiao-Yan Li and Xin Ning for the WBBImodel, Yan Lin and Pin Liu for technical assistance, and Dr.Joana E. Coelho and Dr. Samaneh Maysami for editing themanuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Bahloul, M., Chaari, A.N., Kallel, H., Khabir, A., Ayadi, A.,Charfeddine, H., Hergafi, L., Chaari, A.D., Chelly, H.E., Ben
Hamida, C., Rekik, N., and Bouaziz, M. (2006). Neurogenicpulmonary edema due to traumatic brain injury: evidence ofcardiac dysfunction. Am. J. Crit. Care 15, 462–470.
Cernak, I., Wang, Z., Jiang, J., Bian, X., and Savic, J. (2001a).Cognitive deficits following blast injury-induced neuro-trauma: possible involvement of nitric oxide. Brain Inj. 15,593–612.
Cernak, I., Wang, Z., Jiang, J., Bian, X., and Savic, J. (2001b).Ultrastructural and functional characteristics of blast injury-induced neurotrauma. J. Trauma 50, 695–706.
Colice, G.L., Matthay, M.A., Bass, E., and Matthay, R.A. (1984).Neurogenic pulmonary edema. Am. Rev. Respir. Dis. 130,941–948.
Csermely, P., Schnaider, T., Soti, C., Prohaszka, Z., and Nardai,G. (1998). The 90-kDa molecular chaperone family: structure,function, and clinical applications. A comprehensive review.Pharmacol. Ther. 79, 129–168.
De Kloet, E.R., Joels, M., and Holsboer, F. (2005). Stress and thebrain: from adaptation to disease. Nat. Rev. Neurosci. 6, 463–475.
De Kloet, E.R., Korte, S.M., Rots, N.Y., and Kruk, M.R. (1996).Stress hormones, genotype, and brain organization. Implica-tions for aggression. Ann. N.Y. Acad. Sci. 794, 179–191.
De Kloet, E.R., Vreugdenhil, E., Oitzl, M.S., and Joels, M. (1998).Brain corticosteroid receptor balance in health and disease.Endocr. Rev. 19, 269–301.
Grenert, J.P., Sullivan, W.P., Fadden, P., Haystead, T.A., Clark,J., Mimnaugh, E., Krutzsch, H., Ochel, H.J., Schulte, T.W.,Sausville, E., Neckers, L.M., and Toft, D.O. (1997) The amino-terminal domain of heat shock protein 90 (hsp90) that bindsgeldanamycin is an ATP=ADP switch domain that regulateshsp90 conformation. J. Biol. Chem. 272, 23843–23850.
Hashiguchi, N., Ogura, H., Tanaka, H., Koh, T., Aoki, M.,Shiozaki, T., Matsuoka, T., Shimazu, T., and Sugimoto, H.(2001). Enhanced expression of heat shock proteins in leuko-cytes from trauma patients. J. Trauma 50, 102–107.
Jibard, N., Meng, X., Leclerc, P., Rajkowski, K., Fortin, D.,Schweizer-Groyer, G., Catelli, M.G., Baulieu, E.E., and Cade-pond, F. (1999). Delimitation of two regions in the 90-kDa heatshock protein (Hsp90) able to interact with the glucocorticos-teroid receptor (GR). Exp. Cell Res. 247, 461–474.
Joels, M. (2006). Corticosteroid effects in the brain: U-shape it.Trends Pharmacol. Sci. 27, 244–250.
Liu, D.H., Su, Y.P., Zhang, W., Lou, S.F., Ran, X.Z., Gao, J.S., andCheng, T.M. (2002). Downregulation of glucocorticoid recep-tors of liver cytosols and the role of the inflammatory cyto-kines in pathological stress in scalded rats. Burns 28, 315–320.
Liu, J., and DeFranco, D.B. (1999). Chromatin recycling of glu-cocorticoid receptors: implications for multiple roles of heatshock protein 90. Mol. Endocrinol. 13, 355–365.
Li, X., Gu, W., Masinde, G., Hamilton-Ulland, M., Xu, S., Mohan,S., and Baylink, D.J. (2001). Genetic control of the rate ofwound healing in mice. Heredity 86, 668–674.
McCullers, D.L., Sullivan, P.G., Scheff, S.W., and Herman, J.P.(2002). Traumatic brain injury regulates adrenocorticosteroidreceptor mRNA levels in rat hippocampus. Brain Res. 947, 41–49.
Moore, S.K., Kozak, C,. Robinson, E.A., Ullrich, S.J., and Ap-pella, E. (1987). Cloning and nucleotide sequence of the mu-rine hsp84 cDNA and chromosome assignment of relatedsequences. Gene 56, 29–40.
Murphy, E.K., Spencer, R.L., Sipe, K.J., and Herman, J.P. (2002).Decrements in nuclear glucocorticoid receptor (GR) proteinlevels and DNA binding in aged rat hippocampus. En-docrinology 143, 1362–1370.
380 SHEN ET AL.

Murphy, P.J., Morishima, Y., Kovacs, J.J., Yao, T.P., and Pratt,W.B. (2005). Regulation of the dynamics of hsp90 action on theglucocorticoid receptor by acetylation=deacetylation of thechaperone. J. Biol. Chem. 280, 33792–33799.
Nemoto, T., Ohara-Nemoto, Y., Ota, M., Takagi, T., andYokoyama, K. (1995). Mechanism of dimer formation of the90-kDa heat-shock protein. Eur. J. Biochem. 233, 1–8.
Pearl, L.H., and Prodromou, C. (2001). Structure, function, andmechanism of the Hsp90 molecular chaperone. Adv. ProteinChem. 59, 157–186.
Radojicic, C., Andric, B., Simovic, M., Dujic, A., and Marinkovic,D. (1990). Genetic basis of resistance to trauma in inbredstrains of mice. J. Trauma 30, 211–213.
Ramaglia, V., and Buck, L.T. (2004). Time-dependent expressionof heat shock proteins 70 and 90 in tissues of the anoxicwestern painted turtle. J. Exp. Biol. 207, 3775–3784.
Reichardt, H.M., Umland, T., Bauer, A., Kretz, O., and Schutz, G.(2000). Mice with an increased glucocorticoid receptor genedosage show enhanced resistance to stress and endotoxicshock. Mol. Cell Biol. 20, 9009–9017.
Rogers, F.B., Shackford, S.R., Trevisani, G.T., Davis, J.W.,Mackersie, R.C., and Hoyt, D.B. (1995). Neurogenic pulmo-nary edema in fatal and nonfatal head injuries. J. Trauma 39,860–866; discussion 866–868.
Rosenhagen, M.C., Young, J.C., Wochnik, G.M., Herr, A.S.,Schmidt, U., Hartl, F.U., Holsboer, F., and Rein, T. (2001).Synergistic inhibition of the glucocorticoid receptor by radi-cicol and benzoquinone ansamycins. Biol. Chem. 382, 499–504.
Sangster, T.A., Salathia, N., Undurraga, S., Milo, R., Schellenberg,K., Lindquist, S., and Queitsch, C. (2008). HSP90 affects theexpression of genetic variation and developmental stability inquantitative traits. Proc. Natl. Acad. Sci. U.S.A. 105, 2963–2968.
Sapolsky, R.M., Krey, L.C., and McEwen, B.S. (1986). The neu-roendocrinology of stress and aging: the glucocorticoid cas-cade hypothesis. Endocr. Rev. 7, 284–301.
Scheibel, T., Siegmund, H.I., Jaenicke, R., Ganz, P., Lilie, H., andBuchner, J. (1999). The charged region of Hsp90 modulates thefunction of the N-terminal domain. Proc. Natl. Acad. Sci.U.S.A. 96, 1297–1302.
Shen, H.Y., He, J.C., Wang, Y., Huang, Q.Y., and Chen, J.F.(2005). Geldanamycin induces heat shock protein 70 andprotects against MPTP-induced dopaminergic neurotoxicity inmice. J. Biol. Chem. 280, 39962–39969.
Soti, C., Vermes, A., Haystead, T.A., and Csermely, P. (2003).Comparative analysis of the ATP-binding sites of Hsp90 by nu-cleotide affinity cleavage: a distinct nucleotide specificity of theC-terminal ATP-binding site. Eur. J. Biochem. 270, 2421–2428.
Tago, K., Tsukahara, F., Naruse, M., Yoshioka, T., and Takano,K. (2004). Regulation of nuclear retention of glucocorti-coid receptor by nuclear Hsp90. Mol. Cell Endocrinol. 213,131–138.
Ulett, G.C., Ketheesan, N., and Hirst, R.G. (2000). Cytokine geneexpression in innately susceptible BALB=c mice and relativelyresistant C57BL=6 mice during infection with virulent Bur-kholderia pseudomallei. Infect. Immun. 68, 2034–2042.
Wang, Z.G. (1989). [An experimental study of blast injury].Zhonghua Yi Xue Za Zhi 69, 7–11, 12.
Whitesell, L,. and Cook, P. (1996) Stable and specific binding ofheat shock protein 90 by geldanamycin disrupts glucocorticoidreceptor function in intact cells. Mol. Endocrinol. 10, 705–712.
Yan, P., Xu, J., Li, Q., Chen, S., Kim, G.M., Hsu, C.Y., and Xu, X.M.(1999). Glucocorticoid receptor expression in the spinal cordafter traumatic injury in adult rats. J. Neurosci. 19, 9355–9363.
Zhao, X.H., Peng, Y.Z., Wang, Y.Y., and Huang, Y.S. (2007).[Influence of heat shock protein 90 on protein serine threoninekinases expression in hypoxic cardiomyocytes]. ZhonghuaShao Shang Za Zhi 23, 265–268.
Address correspondence to:Yuan-Guo Zhou, Ph.D.
Molecular Biology CenterResearch Institute of Surgery and Daping Hospital
10 Changjiang ZhiluChongqing 400042, P.R. China
E-mail: [email protected]
or
Li-Yong Chen, M.D.Department of Anesthesiology
Research Institute of Surgery and Daping Hospital10 Changjiang Zhilu
Chongqing 400042, P.R. China
E-mail: [email protected]
HSP90 AFFECTS TRAUMATIC RESPONSES 381

Copyright © 2022 FDOKUMEN




















