Differences in Iron Nutrition Strategies of Two Calcifuges,Carex piluliferaL. andVeronica...
7
Annals of Botany 80 : 553–559, 1997 Differences in Iron Nutrition Strategies of Two Calcifuges, Carex pilulifera L. and Veronica officinalis L. ANGELIKA ZOHLEN* and GERMUND TYLER Department of Ecology, Soil-Plant Research, Ecology Building, Lund Uniersity, S-223 62 Lund, Sweden Received : 6 January 1997 Accepted : 3 July 1997 Veronica officinalis and Carex pilulifera, widespread calcifuge plants in Europe, were cultivated in acid and calcareous soils to study differences in Fe aquisition strategies indicated in previous studies. The experiments were performed in a computer-controlled glasshouse at a soil solution moisture content of 50–60 % water holding capacity ; additional light was supplied at 70 W m -# if ambient light was ! 100 W m -# between 0600 and 1800h. Both species developed chlorosis when grown in the calcareous soil. C. pilulifera proved unable to translocate sufficient amounts of Fe to the leaves when cultivated in calcareous soil, but much Fe accumulated in, and especially as a precipitate on the surface of roots. In contrast, V. officinalis tended to increase Fe taken up into the leaves of plants grown on calcareous soil, but a much greater proportion of the leaf tissue Fe was accumulated as less active forms not extracted by Fe complexing agents, e.g. 1,10-phenanthroline, than was the case in acid-soil grown plants. Considerably less Fe was accumulated in the root biomass of V. officinalis compared to C. pilulifera. It is concluded that chlorosis in C. pilulifera is related to insufficient Fe uptake in the leaves, whereas leaf immobilization of Fe in physiologically less active forms is the problem in V. officinalis. # 1997 Annals of Botany Company Key words : Iron, chlorosis, calcifuge, iron immobilization, leaf tissue, fractionation, Carex pilulifera, Veronica officinalis. INTRODUCTION The majority of plants in central and northern Europe are able to grow and compete successfully on calcareous soils. However, an appreciable minority of species, the so called calcifuges, are unable to grow in calcareous soils because of limitations in their mineral nutrition. Often, the primary limitation for these species is an inability to render the indigenous forms of soil phosphate available for uptake (Tyler, 1992), but there are also species that are limited by a deficiency of micronutrients, especially iron. The typical symptom of lime-induced chlorosis is a yellowing of leaves. The veins may remain more or less green because Fe is poorly redistributed within the tissue (Chaney, 1984). Several calcifuges, e.g. Veronica officinalis L., Carex pilulifera L. and Rumex acetosella L., develop chlorotic symptoms, probably due to Fe deficiency, when cultivated in limestone soils (Tyler, 1996 a). This is particularly evident when the primary growth limitation by phosphate is reversed by phosphate addition to the soil. In C. pilulifera, but not in V. officinalis, chlorosis is related to a decreased uptake rate of Fe. Chlorosis caused by a physiological Fe deficiency may be due to an immobilization of this element in metabolically inactive forms in the tissues (Abadı ! a et al., 1989). Interactions with pH, bicarbonate, phosphate, and low-molecular organic acids may be involved (Kinzel, 1982), but mechanisms are not well understood. The aim of this study was to elucidate the difference in the * E-mail Angelika.Zohlena!planteco.lu.se Fe nutrition strategies of the two calcifuges C. pilulifera and V. officinalis using tissue fractionation methods to char- acterize the Fe solubility in leaves and roots. It was hypothesized that ‘ lime ’ chlorosis in C. pilulifera may be due to insufficient uptake or transport of Fe to the shoots, whereas in V. officinalis it may be due to an immobilization of tissue Fe in non-metabolic forms. MATERIALS AND METHODS Two cultivation experiments were performed in a glasshouse in three different soils : an acid soil (the original substrate), a calcareous soil and a calcareous soil with added phosphate to prevent growth limitations caused by phosphorus deficiency. The two soil types used were a Rendzic leptosol (pH-BaCl # 7–5) from the Ordovician limestone ‘ alvar ’ of O $ land (5625« N, 1630« E), approx. 10 cm deep, and the top 10 cm (A h horizon) of a dystric cambisol (pH BaCl # 4–0), developed from a silicate-rock moraine in central Scania (5557« N, 1350« E), south Sweden. We sieved the soils (through a 6 mm mesh) and stored them for 2–3 d at 5 C before further treatment. To half of the samples of limestone soil we added 5 mmol dm - $ of water-soluble CaHPO % [2H # O to provide additional phosphate. For the first experiment we collected adult, well-developed plants of V. officinalis and C. pilulifera from the same area as the acid soil. Equal-sized plants with roots and shoots cut down to approx. 3 cm were planted in 3 l polythene vessels with basal drainage, using five replicates per species and treatment, and five plants per vessel. 0305-7364}97}100553›07 $25.00}0 bo970493 # 1997 Annals of Botany Company
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Differences in Iron Nutrition Strategies of Two Calcifuges,Carex piluliferaL. andVeronica...

Annals of Botany 80 : 553–559, 1997
Differences in Iron Nutrition Strategies of Two Calcifuges,
Carex pilulifera L. and Veronica officinalis L.
ANGELIKA ZOHLEN* and GERMUND TYLER
Department of Ecology, Soil-Plant Research, Ecology Building, Lund Uni�ersity, S-223 62 Lund, Sweden
Received: 6 January 1997 Accepted: 3 July 1997
Veronica officinalis and Carex pilulifera, widespread calcifuge plants in Europe, were cultivated in acid and calcareoussoils to study differences in Fe aquisition strategies indicated in previous studies. The experiments were performed ina computer-controlled glasshouse at a soil solution moisture content of 50–60% water holding capacity ; additionallight was supplied at 70 W m−# if ambient light was ! 100 W m−# between 0600 and 1800h. Both species developedchlorosis when grown in the calcareous soil. C. pilulifera proved unable to translocate sufficient amounts of Fe to theleaves when cultivated in calcareous soil, but much Fe accumulated in, and especially as a precipitate on the surfaceof roots. In contrast, V. officinalis tended to increase Fe taken up into the leaves of plants grown on calcareous soil,but a much greater proportion of the leaf tissue Fe was accumulated as less active forms not extracted by Fecomplexing agents, e.g. 1,10-phenanthroline, than was the case in acid-soil grown plants. Considerably less Fe wasaccumulated in the root biomass of V. officinalis compared to C. pilulifera. It is concluded that chlorosis in C. piluliferais related to insufficient Fe uptake in the leaves, whereas leaf immobilization of Fe in physiologically less active formsis the problem in V. officinalis. # 1997 Annals of Botany Company
Key words : Iron, chlorosis, calcifuge, iron immobilization, leaf tissue, fractionation, Carex pilulifera, Veronicaofficinalis.
INTRODUCTION
The majority of plants in central and northern Europe areable to grow and compete successfully on calcareous soils.However, an appreciable minority of species, the so calledcalcifuges, are unable to grow in calcareous soils because oflimitations in their mineral nutrition. Often, the primarylimitation for these species is an inability to render theindigenous forms of soil phosphate available for uptake(Tyler, 1992), but there are also species that are limited bya deficiency of micronutrients, especially iron. The typicalsymptom of lime-induced chlorosis is a yellowing of leaves.The veins may remain more or less green because Fe ispoorly redistributed within the tissue (Chaney, 1984).
Several calcifuges, e.g. Veronica officinalis L., Carexpilulifera L. and Rumex acetosella L., develop chloroticsymptoms, probably due to Fe deficiency, when cultivatedin limestone soils (Tyler, 1996a). This is particularly evidentwhen the primary growth limitation by phosphate is reversedby phosphate addition to the soil. In C. pilulifera, but not inV. officinalis, chlorosis is related to a decreased uptake rateof Fe. Chlorosis caused by a physiological Fe deficiencymay be due to an immobilization of this element inmetabolically inactive forms in the tissues (Abadı!a et al.,1989). Interactions with pH, bicarbonate, phosphate, andlow-molecular organic acids may be involved (Kinzel,1982), but mechanisms are not well understood.
The aim of this study was to elucidate the difference in the
* E-mail Angelika.Zohlena!planteco.lu.se
Fe nutrition strategies of the two calcifuges C. pilulifera andV. officinalis using tissue fractionation methods to char-acterize the Fe solubility in leaves and roots. It washypothesized that ‘ lime’ chlorosis in C. pilulifera may bedue to insufficient uptake or transport of Fe to the shoots,whereas in V. officinalis it may be due to an immobilizationof tissue Fe in non-metabolic forms.
MATERIALS AND METHODS
Two cultivation experiments were performed in a glasshousein three different soils : an acid soil (the original substrate),a calcareous soil and a calcareous soil with added phosphateto prevent growth limitations caused by phosphorusdeficiency. The two soil types used were a Rendzic leptosol(pH-BaCl
#7±5) from the Ordovician limestone ‘alvar ’ of
O$ land (56°25« N, 16°30« E), approx. 10 cm deep, and thetop 10 cm (A
hhorizon) of a dystric cambisol (pH BaCl
#4±0),
developed from a silicate-rock moraine in central Scania(55°57« N, 13°50« E), south Sweden. We sieved the soils(through a 6 mm mesh) and stored them for 2–3 d at5 °C before further treatment. To half of the samples oflimestone soil we added 5 mmol dm−$ of water-solubleCaHPO
%[2 H
#O to provide additional phosphate.
For the first experiment we collected adult, well-developedplants of V. officinalis and C. pilulifera from the same areaas the acid soil. Equal-sized plants with roots and shoots cutdown to approx. 3 cm were planted in 3 l polythene vesselswith basal drainage, using five replicates per species andtreatment, and five plants per vessel.
0305-7364}97}10055307 $25.00}0 bo970493 # 1997 Annals of Botany Company

554 Zohlen and Tyler—Iron Strategies in Calcifuges
The experiment began on 8 May 1996 and finishedbetween 5–8 July. The glasshouse was computer-controlled,with a mean minimum temperature at night of 16±5³2±2 °C(95% confidence limit) and mean maximum temperature byday of 26±4³3±5 °C. The soil moisture was maintained at50–60% of the water holding capacity (WHC) during theexperiment (which corresponded to 15±5–18±6% d.wt of theacid soil and 30±0–36±0% d.wt of the calcareous soil), byadding H
#O to the soil surface every 24–48 h, as necessary.
Additional light, 70 W m−#, was provided by high pressuresodium lamps between 0600h and 1800h if ambient lightwas less than 100 W m−#.
We continuously removed all flower buds produced bythe plants during the experiment. Before harvesting, weestimated the leaf colour of the plants, using the Swedishstandard colour atlas (sheet 2, SIS 1989; transformed into a15 degree scale from yellow to deep green).
At harvest, the above-ground biomass of plants was cutnear the soil surface with titanium scissors and washed byvigorous shaking in polythene vessels for 30 s with 100 ml0±1 HCl and H
#O, respectively. Excess water was removed
by blotting. Leaves produced during the experiment werefinely chopped with the same scissors. Within a few hours ofstorage at 7 °C, we extracted 0±30 g of the freshly choppedmaterial with 7 ml of a 1±5% 1,10-phenanthroline (Katyaland Sharma, 1980, 1984) and a 1 HCl solution (Takkarand Kaur, 1984), respectively, shaken continuously for 17 hin 36 ml plastic tubes. Before use, all equipment had beensoaked overnight in 0±1 HCl and rinsed in H
#O. 1,10-
phenanthroline is an Fe complexing agent considered tochelate mainly Fe#+ and used to characterize differences inmetabolically active Fe between chlorotic and non-chloroticplants. We solubilized 15 g 1,10-phenanthroline in 850 mlH
#O, adjusted the pH to 3±0 with 1 HCl and made the
volume to 1 l. The 1 HCl extractable fraction is consideredto reflect the Fe fraction that is active in chlorophyllformation (Oserkowsky, 1933).
To measure the pH of the plant tissue, 0±30 g freshchopped material was extracted with 7 ml 0±2 KCl, pH 7for 17 h. Before chopping, the plant material was preparedas described above, but washed by shaking for 2¬30 s withH
#O only.Subsamples of the chopped plant material were used for
dry weight determination at 85 °C and determination oftotal Fe by wet digestion (conc. HNO
$) for complete
destruction of organic matter. We evaporated excess acid to2 ml and added H
#O to 25 ml.
The concentrations of Ca, Fe, Al and Mg in theexperimental soils, exchangable with 0±2 BaCl
#(10 g soil
at field moisture and 100 ml extractant, 1 h), were de-termined by plasma emission (ICP-ES) and exchangable Kwas determined by flame AAS in 1000 mg l−" of Cs as CsCl.We also extracted soil Fe (10 g soil at field moisture plus50 ml extractant for 1 h) with two complexing agents,0±02 Na citrate (pH 8±5) and 0±02 Na EDTA (pH 4±7),respectively. The extraction by organic ligands shouldsimulate a possible influence of root exudates on Femobilization. For easily exchangable phosphate, the Sb}Moblue method was used with ascorbic acid reductant onfiltered and centrifuged extracts obtained by shaking 10 g
soil at field moisture for 20 min with 100 ml unbufferedneutral 0±05 Na
#SO
%plus 0±02 NaF (Olsen and
Sommers, 1982; Tyler 1992).We determined the pH of the soil solution obtained by
high-speed centrifugation (12000 rpm) of approx. 130 g soilat field moisture for 1 h, in a specially equipped coolingcentrifuge at 10 °C (Falkengren-Grerup and Tyler, 1993).
The organic matter content of the soil was estimated asloss on ignition (550 °C for 3 h) and calculated as apercentage of dry weight. Clay content was not determined,but did not exceed 5% in any of the soils according toprevious studies using the same soils (Tyler, 1996b).
A second growth experiment was performed in order toconfirm the results of the first one using the same soils andthe sameplant species but somedifferent extractionmethods.Eight replicate vessels of each species and treatment wereused in the second experiment which ran between 9 Aug.and 24 Oct. 1996, using the same growing conditions as inthe first experiment. The mean minimum temperature atnight was 15±5³0±4 °C (95% confidence limit) and the meanmaximum temperature by day was 24±6³0±1 °C.
For the extractions, we washed the plants in 0±1 HCl asabove, followed by three washes in H
#O. After removing
surplus water between blotting paper, we extracted 0±5 gfresh chopped sample with 5 ml 1,10-phenanthroline sol-ution for 17 h. We also used TPTZ (2,4,6-tris-(2-pyridyl)-1,3,5-triazine) as an alternative extractant, being consideredmore specific for Fe#+ than 1,10-phenanthroline (Piersonand Clark 1984a). A 0±02 TPTZ solution was prepared bydissolving 6±25 g in 700 ml H
#O, adding HCl until the TPTZ
was dissolved and making the volume to 1 l with dilutedHCl. The pH of this solution was 2±1. To 0±5 g freshlychopped plant material we added 5 ml TPTZ solution andthe sample was shaken continuously at room temperaturefor 17 h. We performed the total digestion as in the firstexperiment. Before use, we had rinsed all equipment in10−& TPTZ and H
#O.
We also measured the Fe contents of the roots in thesecond experiment. We distinguished between the Fe‘plaque’, i.e. the insoluble amorphous and crystalline Fe-oxides and oxyhydrates precipitated on the root surface(Crowder and St Cyr, 1991) using reducing extraction withDCB (dithionite-citrate-bicarbonate), and the remaining Feinside the root determined by wet digestion. The DCBsolution was prepared according to St Cyr and Campbell(1996) and Otte et al. (1989). It contained 17±8 ml 0±03 Nacitrate solution, 2±2 ml 0±125 Na bicarbonate solution and1±5 g Na dithionite per 20 ml. After shaking the roots withapprox. 300 ml H
#O in 500 ml flasks overnight, we cleaned
them in running H#O, washed them again with approx.
300 ml H#O, shaken continuously overnight and, before
extraction, removed surplus water between blotting paper.We then added 20 ml of DCB solution and extracted for30 min.
All extracts of samples were cleaned by micropore filtering(Gelman Acrodisc PF 0±8 µm prefilter}0±2 µm supor) andanalysed directly by flame atomic absorption spectrophoto-metry (AAS; Varian SpectraAA-400) against standardsprepared in the appropriate extractant.
Statistical treatment comprised the calculation of mean
Zohlen and Tyler—Iron Strategies in Calcifuges 555
values and 95% confidence limits, Pearson correlationcoefficientswith P-values, Tukey test and P-values accordingto one-way Anova.
RESULTS
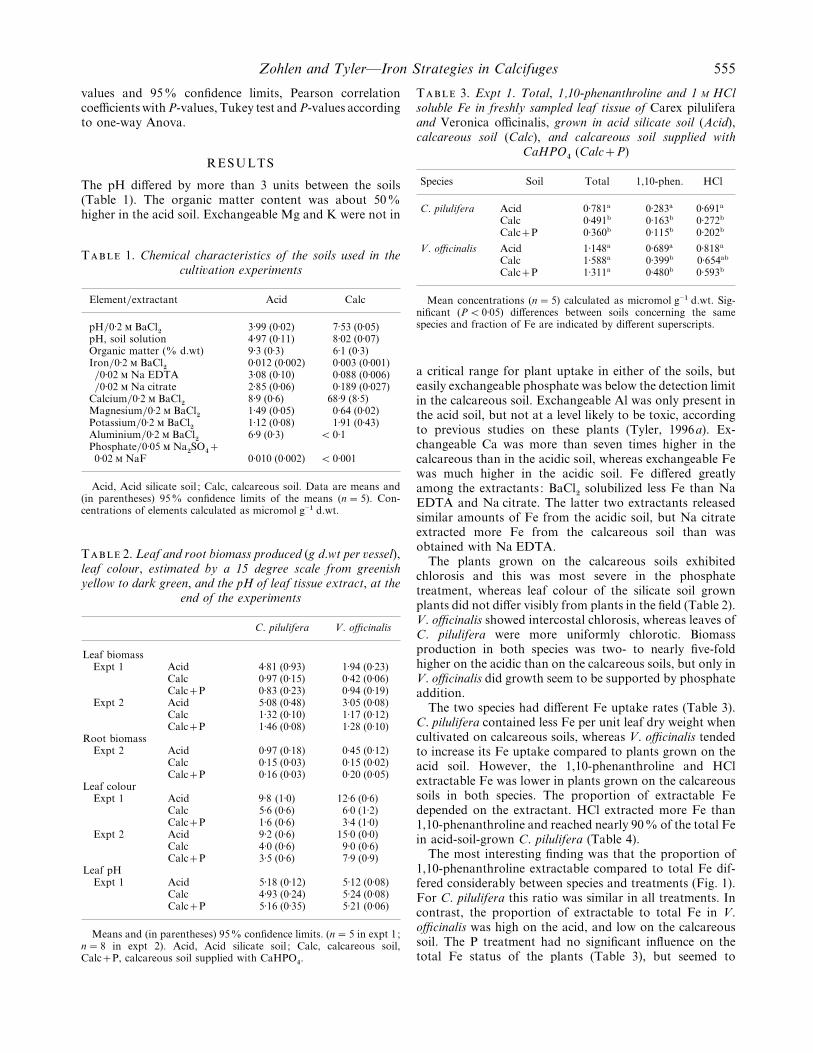
The pH differed by more than 3 units between the soils(Table 1). The organic matter content was about 50%higher in the acid soil. Exchangeable Mg and K were not in
T 1. Chemical characteristics of the soils used in theculti�ation experiments
Element}extractant Acid Calc
pH}0±2 BaCl#
3±99 (0±02) 7±53 (0±05)pH, soil solution 4±97 (0±11) 8±02 (0±07)Organic matter (% d.wt) 9±3 (0±3) 6±1 (0±3)Iron}0±2 BaCl
#0±012 (0±002) 0±003 (0±001)
}0±02 Na EDTA 3±08 (0±10) 0±088 (0±006)}0±02 Na citrate 2±85 (0±06) 0±189 (0±027)
Calcium}0±2 BaCl#
8±9 (0±6) 68±9 (8±5)Magnesium}0±2 BaCl
#1±49 (0±05) 0±64 (0±02)
Potassium}0±2 BaCl#
1±12 (0±08) 1±91 (0±43)Aluminium}0±2 BaCl
#6±9 (0±3) ! 0±1
Phosphate}0±05 Na#SO
%
0±02 NaF 0±010 (0±002) ! 0±001
Acid, Acid silicate soil ; Calc, calcareous soil. Data are means and(in parentheses) 95% confidence limits of the means (n¯ 5). Con-centrations of elements calculated as micromol g−" d.wt.
T 2. Leaf and root biomass produced (g d.wt per �essel),leaf colour, estimated by a 15 degree scale from greenishyellow to dark green, and the pH of leaf tissue extract, at the
end of the experiments
C. pilulifera V. officinalis
Leaf biomassExpt 1 Acid 4±81 (0±93) 1±94 (0±23)
Calc 0±97 (0±15) 0±42 (0±06)CalcP 0±83 (0±23) 0±94 (0±19)
Expt 2 Acid 5±08 (0±48) 3±05 (0±08)Calc 1±32 (0±10) 1±17 (0±12)CalcP 1±46 (0±08) 1±28 (0±10)
Root biomassExpt 2 Acid 0±97 (0±18) 0±45 (0±12)
Calc 0±15 (0±03) 0±15 (0±02)CalcP 0±16 (0±03) 0±20 (0±05)
Leaf colourExpt 1 Acid 9±8 (1±0) 12±6 (0±6)
Calc 5±6 (0±6) 6±0 (1±2)CalcP 1±6 (0±6) 3±4 (1±0)
Expt 2 Acid 9±2 (0±6) 15±0 (0±0)Calc 4±0 (0±6) 9±0 (0±6)CalcP 3±5 (0±6) 7±9 (0±9)
Leaf pHExpt 1 Acid 5±18 (0±12) 5±12 (0±08)
Calc 4±93 (0±24) 5±24 (0±08)CalcP 5±16 (0±35) 5±21 (0±06)
Means and (in parentheses) 95% confidence limits. (n¯ 5 in expt 1;n¯ 8 in expt 2). Acid, Acid silicate soil ; Calc, calcareous soil,CalcP, calcareous soil supplied with CaHPO
%.
T 3. Expt 1. Total, 1,10-phenanthroline and 1 M HClsoluble Fe in freshly sampled leaf tissue of Carex piluliferaand Veronica officinalis, grown in acid silicate soil (Acid),calcareous soil (Calc), and calcareous soil supplied with
CaHPO%
(CalcP)
Species Soil Total 1,10-phen. HCl
C. pilulifera Acid 0±781a 0±283a 0±691a
Calc 0±491b 0±163b 0±272b
CalcP 0±360b 0±115b 0±202b
V. officinalis Acid 1±148a 0±689a 0±818a
Calc 1±588a 0±399b 0±654ab
CalcP 1±311a 0±480b 0±593b
Mean concentrations (n¯ 5) calculated as micromol g−" d.wt. Sig-nificant (P! 0±05) differences between soils concerning the samespecies and fraction of Fe are indicated by different superscripts.
a critical range for plant uptake in either of the soils, buteasily exchangeable phosphate was below the detection limitin the calcareous soil. Exchangeable Al was only present inthe acid soil, but not at a level likely to be toxic, accordingto previous studies on these plants (Tyler, 1996a). Ex-changeable Ca was more than seven times higher in thecalcareous than in the acidic soil, whereas exchangeable Fewas much higher in the acidic soil. Fe differed greatlyamong the extractants : BaCl
#solubilized less Fe than Na
EDTA and Na citrate. The latter two extractants releasedsimilar amounts of Fe from the acidic soil, but Na citrateextracted more Fe from the calcareous soil than wasobtained with Na EDTA.
The plants grown on the calcareous soils exhibitedchlorosis and this was most severe in the phosphatetreatment, whereas leaf colour of the silicate soil grownplants did not differ visibly from plants in the field (Table 2).V. officinalis showed intercostal chlorosis, whereas leaves ofC. pilulifera were more uniformly chlorotic. Biomassproduction in both species was two- to nearly five-foldhigher on the acidic than on the calcareous soils, but only inV. officinalis did growth seem to be supported by phosphateaddition.
The two species had different Fe uptake rates (Table 3).C. pilulifera contained less Fe per unit leaf dry weight whencultivated on calcareous soils, whereas V. officinalis tendedto increase its Fe uptake compared to plants grown on theacid soil. However, the 1,10-phenanthroline and HClextractable Fe was lower in plants grown on the calcareoussoils in both species. The proportion of extractable Fedepended on the extractant. HCl extracted more Fe than1,10-phenanthroline and reached nearly 90% of the total Fein acid-soil-grown C. pilulifera (Table 4).
The most interesting finding was that the proportion of1,10-phenanthroline extractable compared to total Fe dif-fered considerably between species and treatments (Fig. 1).For C. pilulifera this ratio was similar in all treatments. Incontrast, the proportion of extractable to total Fe in V.officinalis was high on the acid, and low on the calcareoussoil. The P treatment had no significant influence on thetotal Fe status of the plants (Table 3), but seemed to
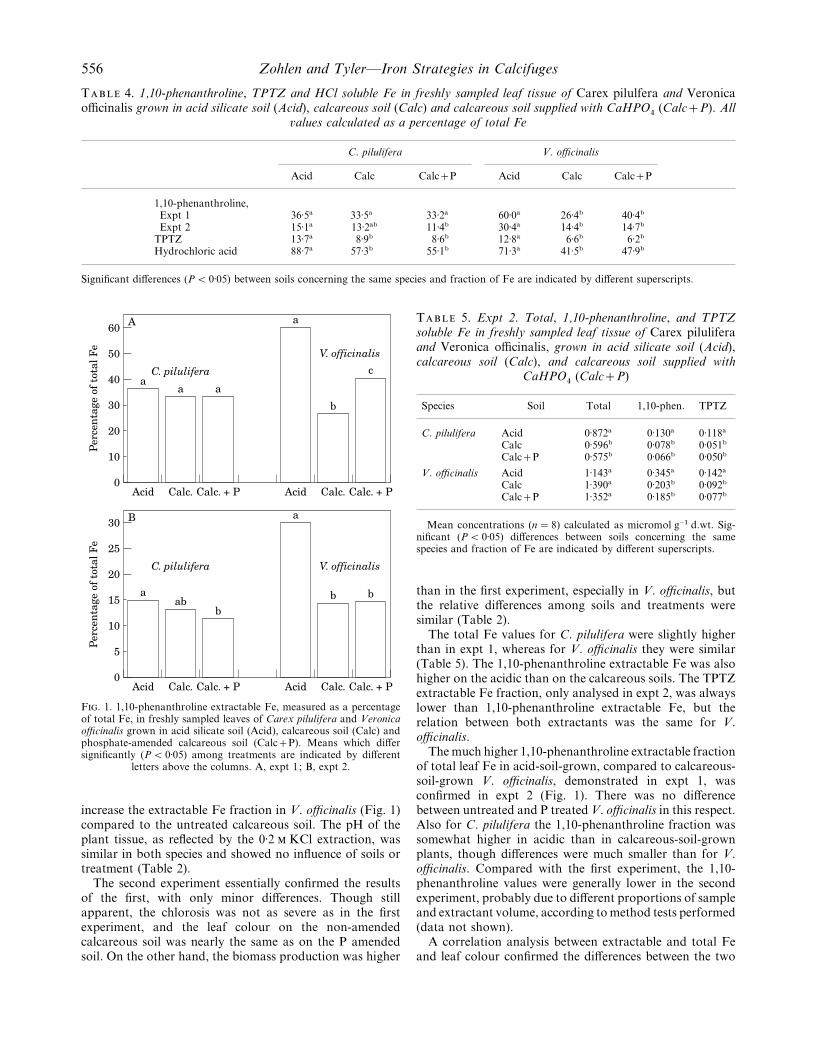
556 Zohlen and Tyler—Iron Strategies in Calcifuges
T 4. 1,10-phenanthroline, TPTZ and HCl soluble Fe in freshly sampled leaf tissue of Carex pilulfera and Veronicaofficinalis grown in acid silicate soil (Acid), calcareous soil (Calc) and calcareous soil supplied with CaHPO
%(CalcP). All
�alues calculated as a percentage of total Fe
C. pilulifera V. officinalis
Acid Calc CalcP Acid Calc CalcP
1,10-phenanthroline,Expt 1 36±5a 33±5a 33±2a 60±0a 26±4b 40±4b
Expt 2 15±1a 13±2ab 11±4b 30±4a 14±4b 14±7b
TPTZ 13±7a 8±9b 8±6b 12±8a 6±6b 6±2b
Hydrochloric acid 88±7a 57±3b 55±1b 71±3a 41±5b 47±9b
Significant differences (P! 0±05) between soils concerning the same species and fraction of Fe are indicated by different superscripts.
30
0Acid
B
Per
cen
tage
of
tota
l Fe
5
25
20
15
10
Calc. Calc. + P Acid Calc. Calc. + P
aab
b
a
b b
C. pilulifera V. officinalis
60
0Acid
A
Per
cen
tage
of
tota
l Fe
10
50
40
30
20
Calc. Calc. + P Acid Calc. Calc. + P
aa a
a
b
cC. pilulifera
V. officinalis
F. 1. 1,10-phenanthroline extractable Fe, measured as a percentageof total Fe, in freshly sampled leaves of Carex pilulifera and Veronicaofficinalis grown in acid silicate soil (Acid), calcareous soil (Calc) andphosphate-amended calcareous soil (CalcP). Means which differsignificantly (P! 0±05) among treatments are indicated by different
letters above the columns. A, expt 1; B, expt 2.
increase the extractable Fe fraction in V. officinalis (Fig. 1)compared to the untreated calcareous soil. The pH of theplant tissue, as reflected by the 0±2 KCl extraction, wassimilar in both species and showed no influence of soils ortreatment (Table 2).
The second experiment essentially confirmed the resultsof the first, with only minor differences. Though stillapparent, the chlorosis was not as severe as in the firstexperiment, and the leaf colour on the non-amendedcalcareous soil was nearly the same as on the P amendedsoil. On the other hand, the biomass production was higher
T 5. Expt 2. Total, 1,10-phenanthroline, and TPTZsoluble Fe in freshly sampled leaf tissue of Carex piluliferaand Veronica officinalis, grown in acid silicate soil (Acid),calcareous soil (Calc), and calcareous soil supplied with
CaHPO%
(CalcP)
Species Soil Total 1,10-phen. TPTZ
C. pilulifera Acid 0±872a 0±130a 0±118a
Calc 0±596b 0±078b 0±051b
CalcP 0±575b 0±066b 0±050b
V. officinalis Acid 1±143a 0±345a 0±142a
Calc 1±390a 0±203b 0±092b
CalcP 1±352a 0±185b 0±077b
Mean concentrations (n¯ 8) calculated as micromol g−" d.wt. Sig-nificant (P! 0±05) differences between soils concerning the samespecies and fraction of Fe are indicated by different superscripts.
than in the first experiment, especially in V. officinalis, butthe relative differences among soils and treatments weresimilar (Table 2).
The total Fe values for C. pilulifera were slightly higherthan in expt 1, whereas for V. officinalis they were similar(Table 5). The 1,10-phenanthroline extractable Fe was alsohigher on the acidic than on the calcareous soils. The TPTZextractable Fe fraction, only analysed in expt 2, was alwayslower than 1,10-phenanthroline extractable Fe, but therelation between both extractants was the same for V.officinalis.
The much higher 1,10-phenanthroline extractable fractionof total leaf Fe in acid-soil-grown, compared to calcareous-soil-grown V. officinalis, demonstrated in expt 1, wasconfirmed in expt 2 (Fig. 1). There was no differencebetween untreated and P treated V. officinalis in this respect.Also for C. pilulifera the 1,10-phenanthroline fraction wassomewhat higher in acidic than in calcareous-soil-grownplants, though differences were much smaller than for V.officinalis. Compared with the first experiment, the 1,10-phenanthroline values were generally lower in the secondexperiment, probably due to different proportions of sampleand extractant volume, according to method tests performed(data not shown).
A correlation analysis between extractable and total Feand leaf colour confirmed the differences between the two

Zohlen and Tyler—Iron Strategies in Calcifuges 557
T 6. Linear correlation coefficients (r �alues) for leafcolour on tissue Fe �ariables
C. pilulifera V. officinalis
Expt 1Total Fe 0±880*** ®0±267Fe-phen. 0±882*** 0±707**Fe-1 HCl 0±904*** 0±592*
Expt 2Total Fe 0±891*** ®0±489*Fe-phen. 0±893*** 0±893***Fe-TPTZ 0±913*** 0±822***Total Fe—Fe-phen. 0±854*** ®0±657**Total Fe—Fe-TPTZ 0±836*** ®0±416
***P! 0±001, **P! 0±01, *P! 0±05.
T 7. Expt 2. DCB soluble ‘plaque ’ Fe, remaining rootFe, and ‘plaque ’ Fe as a percentage of total Fe in the roots ofCarex pilulifera and Veronica officinalis, grown in acidsilicate soil (Acid), calcareous soil (Calc) and calcareous
soil supplied with CaHPO%
(CalcP)
Species Soil ‘Plaque’ Root ‘Plaque’ % total
C pilulifera Acid 10±27a 3±04a 77±3a
Calc 4±50b 3±03a 59±2b
CalcP 3±03b 4±92a 40±6c
V. officinalis Acid 5±55a 1±02a 84±5a
Calc 1±38b 1±12a 55±1b
CalcP 1±95b 1±19a 60±0b
Mean concentrations (n¯ 8) calculated as micromol g−" d.wt. Sig-nificant (P! 0±05) differences between soils concerning the samespecies and fraction of Fe are indicated by different superscripts.
species (Table 6). For C. pilulifera, both total and extractableFe were significantly positively correlated with leaf colour.For V. officinalis this was only true for the extractablefractions. Total Fe actually had a negative relation to leafcolour in this species, which was particularly valid for the Fefraction not extracted by 1,10-phenanthroline (Table 6).
The root biomass, only measured in expt 2, varied amongtreatments in a similar way to the leaf biomass and washigher on the acid soil (Table 2). An analysis of the DCBsoluble ‘plaque’ Fe on the roots demonstrated much highervalues in the acid soil in both species. However, the Feremaining in the roots after DCB extraction did not varyamong treatments, but differed considerably between thespecies (Table 7). In contrast to leaf Fe, the ‘plaque’ and theroot Fe were higher in C. pilulifera than in V. officinalis.However, the ratio of ‘plaque’ to total root Fe (‘plaque’Feroot Fe) did not differ greatly between species exceptfor the phosphorus treatment.
DISCUSSION
The Fe complexing reagent, 1,10-phenanthroline is notperfectly selective for Fe#+, though it has higher affinity forFe#+ than for Fe$+ (Pierson and Clark 1984a, b). Moreover,if the extracted solution is measured spectrophotometrically,
the Fe values can easily be overestimated because of thebackground colour of other extracted compounds (Abadı!aet al., 1984, Katyal and Sharma, 1984; Takkar and Kaur,1984). Therefore the Fe determination was done by atomicabsorption spectrometry, though a discrimination betweenthe two oxidation states is not possible. On the other hand,even in the dark Fe$+ is rapidly reduced to Fe#+ by leafcompounds such as ascorbic acid, phenols or organic acids(Abadı!a et al., 1984, Mehrotra and Gupta 1990, 1991b) and,therefore, it is hardly possible to discriminate between Fe#+
and Fe$+ using the usual extraction methods. 2,4,6-tris-(2-pyridyl)-1,3,5-triazine (TPTZ) is quite selective for Fe#+,especially at low concentrations (Collins and Diehl, 1959;Pierson and Clark, 1984a). HCl is not selective for Fe#+ orFe$+ but Takkar and Kaur (1984) found a better correlationbetween HCl extractable Fe and chlorosis than with 1,10-phenanthroline.
In our experiment the correlation between leaf colour and1,10-phenanthroline or TPTZ extraction did not differgreatly, but both values were higher than for the HClextracts (Table 6). This is not in agreement with Takkar andKaur (1984) who did not find a strong relation to chlorosisusing the 1,10-phenanthroline extraction. The finding that1,10-phenanthroline accounted somewhat better than TPTZfor the variability in chlorosis in fresh plant material wasalso reported by Mehrothra and Gupta (1991a). However,Benton Jones (1991) could not distinguish between chloroticand non-chlorotic plants using 1,10-phenanthroline. Thedifferent extraction methods do not provide a direct measureof physiologically ‘active’ Fe (Chaney, 1984) but they aregenerally better related to chlorosis than total tissue Fe(Oserkowsky, 1933; Abadı!a et al., 1984; Mehrotra andGupta, 1991a). However, the origin and localization of theextracted Fe remains undetected (Abadı!a, 1992). Total Fe inchlorotic plants is often higher than in non-chlorotic plants(Oserkowsky, 1933; Mengel and Buebl, 1983; Takkar andKaur, 1984). This was supported by the results for V.officinalis in our study. There was no positive correlationbetween total Fe and the degree of chlorosis or betweentotal and extractable Fe in this species. In contrast, for C.pilulifera both total and extractable Fe were proportionallylower in calcareous-soil-grown plants and both fractionswere highly interrelated and well correlated with chlorosis.Ao et al. (1987) found decreased total Fe contents in plantssuffering from lime-induced chlorosis. Dicotyledonousspecies are more effective in Fe solubilization than mono-cotyledonous plants (Olsen and Brown, 1980a). This couldexplain the difference between C. pilulifera and V. officinalisforced to grow on calcareous soil.
Plants mainly take up Fe in the form of Fe#+ (Chaney,Brown and Tiffin, 1972; Olsen and Brown, 1980b), but inwell-drained soils most Fe is present as Fe$+ (Olsen andBrown, 1980b ; Jolley and Brown, 1994) and has to bereduced by the root before uptake (Chaney et al., 1972).During translocation in the plant, Fe#+ is reoxidized to Fe$+
and transported mainly as citrate or malate (Tiffin, 1970),and most of the Fe in plants is present as Fe$+ (Machold,Meisel and Schnorr, 1968; Jolley and Brown, 1994).
Dicotyledons and non-gramineous monocotyledons areregarded as ‘strategy I’ plants (Marschner, Roemheld and

558 Zohlen and Tyler—Iron Strategies in Calcifuges
Acid
Leaves
Roots
Root'plaque'
58%
17%
25%
Calc.
35%
24%
41%
Acid
39%
7%
54%
Calc.
10%9%
81%
Carex pilulifera Veronica officinalis
F. 2. Percentage distribution of Fe between leaves, roots and root ‘plaque’ (8) in Carex pilulifera and Veronica officinalis, grown in acid silicatesoil (Acid) and unamended calcareous soil (Calc). The area of each compartment is proportional to the percentage.
Kissel, 1986; Marschner, 1995). Under Fe deficiency theyexhibit several mechanisms to maintain an adequate level ofFe. Many plants release H+ ions, extruded by an enhancedactivity of a plasma membrane-bound reductase. Reduc-tants}chelators are often released as well, but this dependson the plant species (Jolley, Brown and Nugent, 1991). Thereduction of Fe$+ to Fe#+ at the root plasmalemma isenhanced, and citrate is accumulated in root tissues,accelerating the transport of solubilized Fe to the upperparts of the plant (De Vos, Lubberding and Bienfait, 1986;Jolley and Brown, 1994). That citrate is capable ofsolubilizing Fe in soils was shown in our experiment by theincreased Fe extraction, especially from the calcareous soil(Table 1).
A high HCO$
− content of the soil or plant tissue isregarded as the main cause of lime-induced chlorosis,whereas P is considered to be of little influence (Boxma,1972; Pissaloux, Morard and Bertoni, 1995; Mengel,Breininger and Buebl, 1984, Romera, Alca! ntara and de laGuardia, 1991). Due to HCO
$
−, a high pH inhibits rootsresponses, including neutralization of the H+ and reductionof Fe$+ at the plasmamembrane (Marschner and Ro$ mheld,1994). Our results might indicate some influence ofphosphorus on Fe dynamics (Fig. 1), but the differencesbetween non-amended and phosphate-amended calcareoussoil were not distinct enough for consistent conclusions tobe drawn. Leaf P concentrations were not determined in ourstudy, but previous work has demonstrated considerableincreases in P uptake by these species when grown inphosphate-amended limestone soil (Tyler, 1996b). A highCa content does probably not induce chlorosis, but canimpair plant growth (Pissaloux et al., 1995). Also severe Fe-deficiency may inhibit cell growth and thus leaf growth(Abbott, 1967). This was evident for both experimentalplants, especially for the root biomass, though phosphateaddition apparently promoted growth of calcareous-soil-grown V. officinalis. In contrast to Mengel (1994), we foundno correlation between leaf colour and tissue pH (Table 2)but this could be due to the rather coarse method for pHdetermination.
Shoot and root biomass exhibited a significant shift
towards shoot biomass on calcareous soils in C. pilulifera,whereas V. officinalis had a similar shoot:root ratio in alltreatments. Mengel (1994) reported increased levels of Fe inroots under Fe deficiency, but in our experiments the ratioof total shoot Fe and root Fe was similar for each species inall treatments. Comparing the Fe distribution among leaves,roots and root ‘plaque’, a relatively greater proportion ofthe Fe was located in the leaves of the calcareous-soil-grownplants, compared to the acid-soil-grown plants (Fig. 2). C.pilulifera accumulated more ‘plaque’ than V. officinalis. Thestrategy of V. officinalis was to translocate most of the totalbiomass Fe to the leaves, whereas most of the Fe in C.pilulifera was recovered as ‘plaque’ from the root surface.
These differences in Fe distribution strategy between thetwo species were particularly apparent in the calcareous-soil-grown plants. In C. pilulifera from the calcareous soilalmost 60% of the total plant Fe was located in or on theroot biomass, whereas this value was less than 20% for V.officinalis. This, together with the low total uptake of Fefrom calcareous soil in the leaves of C. pilulifera, and thehigh leaf uptake but immobilization of Fe in forms notextractable by 1,10-phenanthroline in V. officinalis, consti-tutes a major difference between these species in Fe nutritionstrategy and calcifuge behaviour.
ACKNOWLEGEMENT
We thank Maj-Britt Larsson for her skilful technicalassistance.
LITERATURE CITED
Abadı!a J. 1992. Leaf responses to Fe deficiency: a review. Journal ofPlant Nutrition 15 : 1699–1713.
Abadı!a J, Monge E, Montanes L, Heras L. 1984. Extraction of ironfrom plant leaves by Fe (II) chelators. Journal of Plant Nutrition 7 :777–784.
Abadı!a A, Sanz M, de las Rivas J, Abadı!a J. 1989. Photosyntheticpigments and mineral composition of iron deficient pear leaves.Journal of Plant Nutrition 12 : 827–838.
Abbott AJ. 1967. Physiological effects of micronutrient deficiencies inisolated roots of Lycopersicon esculentum. The New Phytologist66 : 419–437.

Zohlen and Tyler—Iron Strategies in Calcifuges 559
Ao TY, Chaney RL, Korcak RF, Fan F, Faust M. 1987. Influence of soilmoisture level on apple iron chlorosis development in a calcareoussoil. Plant and Soil 104 : 85–92.
Benton Jones J Jr. 1991. Plant tissue analysis in micronutrients. In :Mortvedt JJ et al., eds. Micronutrients in agriculture. Madison:Soil Science Society of America Book Series 477–521.
Boxma R. 1972. Bicarbonate as the most important soil factor in lime-induced chlorosis in the Netherlands. Plant and Soil 37 : 233–243.
Chaney RL. 1984. Diagnostic practices to identify iron deficiency inhigher plants. Journal of Plant Nutrition 7 : 47–67.
Chaney RL, Brown JC, Tiffin LO. 1972. Obligatory reduction of ferricchelates in iron uptake by soybean. Plant Physiology 50 : 208–213.
Collins PF, Diehl H. 1959. 2,4,6-Tripyridyl-s-triazine as reagent for iron.Determination of iron in limestone, silicates, and refractories.Analytical Chemistry 11 : 1862–1867.
Crowder A, St Cyr L. 1991. Iron oxide plaques on wetland roots.Trends in Soil Science 1 : 315–329.
De Vos RC, Lubberding HJ, Bienfait HF. 1986. Rhizosphere acidifi-cation as a response to iron deficiency in bean plants. PlantPhysiology (Bethesda) 81 : 842–846.
Falkengren-Grerup U, Tyler G. 1993. The importance of soil aciditymoisture, exchangeable cation pools and organic matter solubilityto the cationic composition of beech forest (Fagus syl�atica L.) soilsolution. Zeitschrift fuX r PflanzenernaX hrung und Bodenkunde 156 :365–370.
Jolley VD, Brown JC. 1994. Genetically controlled uptake and use ofiron by plants. In: Manthey JA et al., eds. Biochemistry of metalmicronutrients in the rhizosphere. Boca Raton: CRC Press Inc.,251–265.
Jolley D, Brown JC, Nugent PE. 1991. A genetically related response toiron deficiency stress in muskmelon. Plant and Soil 130 : 87–92.
Katyal JC, Sharma BD. 1980. A new technique of plant analysis toresolve iron chlorosis. Plant and Soil 55 : 105–119.
Katyal JC, Sharma BD. 1984. Some modification in the assay of Fe#+
in 1-10,o-phenanthroline extracts of fresh plant tissues. Plant andSoil 79 : 449–450.
Kinzel H. 1982. PflanzenoX kologie und Mineralstoffwechsel. Stuttgart :Eugen Ulmer.
Machold O, Meisel W, Schnorr H. 1968. Determination of the bindingform of iron in the leaves through mossbauer spectrometry.Naturwissenschaften 55 : 499–500.
Marschner H. 1995. Mineral nutrition of higher plants. London:Academic Press.
Marschner H, Roemheld V. 1994. Strategies of plants for acquisition ofiron. Plant and Soil 165 : 261–274.
Marschner H, Roemheld V, Kissel M. 1986. Different strategies inhigher plants in mobilization and uptake of iron. Journal of PlantNutrition 9 : 695–713.
Mehrotra SC, Gupta P. 1990. Reduction of iron by leaf extracts and itssignificance for the assay of Fe(II) iron in plants. Plant Physiology(Bethesda) 93 : 1017–1020.
Mehrotra SC, Gupta P. 1991a. A comparative study of some methodsof active Fe estimation in maize leaves. Indian Journal ofExperimental Biology 29 : 1051–1053.
Mehrotra SC, Gupta P. 1991b. Studies on evaluation of Fe(II) ironassay procedures in plants : pH, extractant and reducing activity indifferent plant parts. Indian Journal of Experimental Biology 29 :661–663.
Mengel K. 1994. Iron availability in plant tissues—iron chlorosis on
calcareous soils. Plant and Soil 165 : 275–283.
Mengel K, Breininger MT, Buebl W. 1984. Bicarbonate, the most
important factor inducing iron chlorosis in vine grapes on
calcareous soil. Plant and Soil 81 : 333–344.
Mengel K, Buebl W. 1983. Distribution of iron in vine leaves with
HCO$
− induced chlorosis. Zeitschrift fuX r PflanzenernaX hrung und
Bodenkunde 146 : 560–571.
Olsen RA, Brown JC. 1980a. Factors related to iron uptake by
dicotyledonous and monocotyledonous plants 1. pH and re-
ductant. Journal of Plant Nutrition 2 : 629–645.
Olsen RA, Brown JC. 1980b. Factors related to iron uptake by
dicotyledonous and monocotyledonous plants 2. The reduction of
Fe$+ as influenced by roots and inhibitors. Journal of Plant
Nutrition 2 : 647–660.
Olsen SR, Sommers LE. 1982. Phosphorus. In : Methods of soil analysis.
2. Chemical and microbiological properties. Agronomy 9. Madison:
American Society of Agronomy, Inc., 403–430.
Oserkowsky J. 1933. Quantitative relation between chlorophyll and
iron in green and chlorotic pear leaves. Plant Physiology 8 :
449–468.
Otte ML, Rozema J, Koster L, Haarsma MS. 1989. Iron plaque on
roots of Aster tripolium L.: interaction with zinc uptake. New
Phytologist 111 : 309–317.
Pierson EE, Clark RB. 1984a. Ferrous iron determination in plant
tissue. Journal of Plant Nutrition 7 : 107–116.
Pierson EE, Clark RB. 1984b. Chelating agent differences in ferrous
iron determinations. Journal of Plant Nutrition 7 : 91–106.
PissalouxA,MorardP,BertoniG. 1995.Alkalinity-bicarbonate-calcium
effects on iron chlorosis in white lupine in soilless culture. In:
Abadı!a J, ed. De�elopments in plant and soil sciences, 59. Iron
nutrition in soils and plants: Se�enth International Symposium on
Iron Nutrition and Interactions in Plants, Zaragoza, Spain, June
27–July 2, 1993. Dordrecht : Kluwer Academic Publishers,
127–133.
Romera FJ, Alca! ntara E, de la Guardia MD. 1991. Characterization of
the tolerance to iron chlorosis in different peach rootstocks grown
in nutrient solution. I. Effect of bicarbonate and phosphate. Plant
and Soil 130 : 115–119.
St Cyr L, Campbell PGC. 1996. Metals (Fe, Mn, Zn) in the root plaque
of submerged aquatic plants collected in situ : Relations with metal
concentrations in the adjacent sediments and in the root tissue.
Biogeochemistry 33 : 45–76.
Takkar PN, Kaur NP. 1984. HCl method for Fe#+ estimation to resolve
iron chlorosis in plants. Journal of Plant Nutrition 7 : 81–90.
Tiffin LO. 1970. Translocation of iron citrate and phosphorus in xylem
exudate of soy bean-d. Plant Physiology 45 : 280–283.
Tyler G. 1992. Inability to solubilize phosphate in limestone soils—key
factor controlling calcifuge habit of plants. Plant and Soil 145 :
65–70.
Tyler G. 1996a. Soil chemical limitations to growth and development
of Veronica officinalis L. and Carex pilulifera L. Plant and Soil 184 :
281–289.
Tyler G. 1996b. Mineral nutrient limitations of calcifuge plants in
phosphate sufficient limestone soil. Annals of Botany 77 : 649–656.