
The isotopic ecology of vertebrates and conservation biology

D E V O N I A N V E R T E B R A T E S F R O M C O L O M B I A
by P . J A N V I E R and C . V I L L A R R O E L
ABSTRACT. Vertebrate remains are reported from the Emsian±Eifelian Floresta Formation and the Late Devonian(?Frasnian) Cuche Formation of north-eastern Colombia. The material from the Floresta Formation is associated with amarine invertebrate fauna and includes an arthrodire and probably a rhenanid. Several vertebrate-bearing localities arerecorded from the Cuche Formation; vertebrates occur with plant remains and lingulid fragments. They include anacanthodian (Cheiracanthoides? sp.), a chondrichthyan (Antarctilamna? sp.), placoderms (Bothriolepis sp., Asterolepis?sp. and an undetermined groenlandaspidid or primitive brachythoracid arthrodire), a stegotrachelid actinopterygian,and three sarcopterygians (a cosmine-covered form tentatively referred to an osteolepidid, the porolepiform Holoptychiussp., and the rhizodontid Strepsodus? sp.). This assemblage suggests a Late Frasnian age and is surprisingly similar toLate Devonian vertebrate assemblages found in similar facies of Europe and North America, notwithstanding thepresence of the Gondwanan chondrichthyan Antarctilamna?.
KEY WORDS: Vertebrata, Devonian, Colombia, South America, biostratigraphy, palaeobiogeography.
P R A C T I C A L L Y no pre-Carboniferous vertebrate remains, except for a few early records from Brazil andthe Falkland Islands (see LelieÁvre et al. 1993 for a historical review), were known from South Americauntil the 1970s and 1980s, when a number of Ordovician±Devonian ®sh-bearing localities werediscovered in Bolivia (Janvier 1976, 1978a, 1987, 1992; Goujet et al. 1984, 1985, 1993; Gagnier et al.1986, 1988, 1989; Janvier and Suarez-Riglos 1986, 1989; Janvier and Dingerkus 1991; Gagnier 1992,1993a, b; Gagnier and Blieck 1992; Blieck et al. 1996; Diaz-Martinez et al. 1996). Other Ordovician±Devonian occurrences have also been reported from Brazil (Janvier and Melo 1987, 1988, 1992; Janvier1993), Argentina (Albanesi et al. 1995), and Venezuela (Thery 1982; Young 1993a), and indeterminate®sh remains have also been found in the Upper Devonian of Colombia (Mojica and Villarroel 1984).
Agnathans are only represented by the thelodont Turinia gondwana from the Lower Devonian ofSeripona, Bolivia (Turner, in Gagnier et al. 1988). Most of the Devonian ®sh remains known to date fromSouth America are gnathostomes. Among these, chondrichthyans and acanthodians predominate incollections from Bolivia and Brazil. Placoderms, which are the most abundant and diverse Devonianvertebrates elsewhere, were hitherto poorly represented in South America (Goujet et al. 1985; Diaz-Martinez et al. 1996; Young 1993a). The situation was even worse with osteichthyans, which were onlyknown from a few actinopterygian and sarcopterygian remains from Bolivia and Venezuela (Janvier andSuarez-Riglos 1986; Gagnier et al. 1989; Young 1993a). The Colombian Devonian vertebrate localitiesdescribed in this paper greatly increase our knowledge of the South American vertebrates from thisinterval.
Schmidt (1938, pl. 1, ®g. 12) mentioned a `crossopterygian' scale (identi®ed by Walter Gross) fromsupposedly Lower Carboniferous `Kulm-like' beds, now known as the GuatiquõÂa Red Beds of the GachalaÂarea (Quetame Massif) of Colombia, but the ®rst record of Devonian vertebrate remains from Colombiawas made by Mojica and Villarroel (1984), who noticed `®sh scales' in various levels of the CucheFormation. A sample from the Cuche Formation (at Potrero RincoÂn 1) containing an interestingassemblage of actinopterygian, acanthodian and chondrichthyan remains was noted in 1995 byChristopher Berry (University of Wales, Cardiff) who passed it on to one of us (PJ) for further study.This discovery triggered a joint project that included prospection of both the Cuche Formation and theunderlying Floresta Formation in November 1996 by the authors of the present paper, and was supported
[Palaeontology, Vol. 43, Part 4, 2000, pp. 729±763, 5 pls] q The Palaeontological Association

by the Universidad Nacional de Colombia (BogotaÂ), the MuseÂum National d'Histoire Naturelle (Paris),and the Institut FrancËais d'Etudes Andines (Lima). A short, preliminary ®eld survey resulted in thediscovery of several vertebrate-bearing localities, mostly in the middle and upper parts of the LateDevonian Cuche Formation, one of which is remarkably rich in, and yields a wide diversity of taxa. Duringthe same survey some vertebrate remains were also discovered in the Emsian-Eifelian Floresta Formation.These Colombian occurrences are, to date, the richest Devonian vertebrate assemblage from SouthAmerica, although the Late Devonian ®sh fauna recorded from western Venezuela (Young 1993a) maywell turn out to be of similar importance.
At this stage of the study, the vertebrate fauna of the Cuche Formation appears to be composed of somewidespead taxa, some typically Gonwanan taxa, and some taxa that have hitherto been recorded only fromNorth America and Europe. Since most of these vertebrates are generally bound to continental margins,this unusual association raises the possibility of close relations between Euramerica and westernGondwana by Late Devonian times.
G E O G R A P H I C A L A N D G E O L O G I C A L S E T T I N G
The Floresta Massif is situated in the north-central part of Colombia, north of Sogamoso, in theDepartment of Boyaca (Text-®g. 1). The massif consists of an anticlinorium that belongs to the centralpart of the Oriental Cordillera, and is bounded by two major faults, the Boyaca fault in the west and theSoapaga fault in the east (Mojica and Villarroel 1984, ®g. 2). The core of the anticlinorium is orientedNNE-SSW and consists of metamorphic rocks (Silgara Formation sensu Etayo Serna et al. 1986) ofPrecambrian?±Cambrian? age, and granites dated at 471 6 22 Ma (Stock de Chuscales, in Etayo Sernaet al. 1986). These metamorphic and granitic rocks are overlain by marine and continental sediments oflate Early±early Late Devonian, Mesozoic (Late Jurassic?±Cretaceous) and Cenozoic (Paleogene) ages,lying on either side of the core.
The geological history of this structure is not yet clearly understood, although it is supposed that themassif acted as a positive element that was exposed three times during the Phanerozoic. First, from the MidOrdovician to the Emsian, as evidenced by the lack of any sediment dated for this interval, although it isstill uncertain whether these were ever deposited, or were eroded during the Caledonian orogeny, asadvocated by some authors (BuÈrgl 1961). Second, from the Famennian to the Jurassic or Cretaceous,although this interval is recorded in the neighbouring regions. However, in this case, there is no doubt thatit is linked with some phases of the Hercynian orogeny (Cediel 1972) because the GiroÂn Formation, whichis overlain conformably by the Hauterivian Tibasosa Formation, lies unconformably upon all the pre-Palaeozoic and Palaeozoic units. Finally, the third period is the most poorly known, and the role of themassif during the Andean orogeny, after the regression of the Cretaceous sea, has not been established.
Lithostratigraphical units
Since the subject of the present study is the record of Devonian ®shes, this stratigraphical introductionfocuses on the Palaeozoic formations of the Floresta Massif and the vertebrate-bearing localities, withparticular reference to the palaeoenvironments that they represent. More general and detailed studies ofthis massif can be found in, for example, works by Botero-Restrepo (1950), Cediel (1969) and Mojica andVillarroel (1984). The major units involved in this study, from the oldest to the youngest, are as follows(Text-®g. 2):
El Tibet Formation (Text-®g. 2B). The dominant lithology of the El Tibet Formation consists of ®ne- tocoarse-grained quartzitic and kaolinitic sandstones, locally interbedded with thin (about 10 cm) claystoneand mudstone layers. These layers are normally tabular, but may be bevelled laterally. Their base is erosiveand may include clay intraclasts. When the sandstones are overlain by claystones and mudstones, thetransition is only a few millimetres thick. Usually, the sandstones are massive, but normal or invertedbedding, graduated strati®cation, and cross- or channel-bedding, always of low angle, also occur.
The unit unconformably overlies either the metamorphic or the granitoid rocks of the core of the
730 P A L A E O N T O L O G Y , V O L U M E 4 3

anticline, and this is re¯ected in the components of its basal conglomerates. The top part of the formationgrades progressively into the Floresta Formation (Text-®g. 2B). This transition zone includes thin layers ofmicaceous, ®ne-grained sandstone, that display a plano-parallel lamination, and are interbedded withyellowish and light yellowish brown mudstones and claystones.
The age of the El Tibet Formation has usually been established on the basis of the Emsian±Eifelian ageof the overlying Floresta Formation, well dated by its rich marine invertebrate fauna (see below).However, recently GroÈsser and ProÈssl (1994) directly con®rmed the Emsian age of the El Tibet Formationon the basis of a spore assemblage.
In the area of the Floresta Massif, the El Tibet Formation has a restricted distribution and seems to occuras fringes on the ¯anges of the core of the anticline, mainly in the east; it becomes bevelled to the south andeast. For example, in Horno y Vivas, south of Floresta, the formation is missing and the Floresta Formationrests directly on the cordieritic schists of the core. North of this area, the El Tibet Formation is 20 m thick,and west of Otenga and further north, it reaches 327 m and even 425 m, so that its base no longer crops out(Paz and Urrutia 1996). Paz and Urrutia (1996) considered that the formation was deposited in acontinental environment (from alluvial fans to active braided rivers), and in coastal to subcoastalenvironments during the interval corresponding to the transition zone to the Floresta Formation.
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 731
TEXT-FIG. 1. Locality map.

Floresta Formation (Text-®g. 2A±B). Except for its upper and basal parts, the Florestra Formation consistsof a thick sequence of ®ne-grained, micaceous mudstones and claystones that are off-white to lightyellowish brown in colour. Most of this sequence is so strongly jointed that it is dif®cult to trace thebedding planes. According to Barrett (1988), the formation is 500±600 m thick, although this includes theEl Tibet Formation, as the `El Tibet Member'. In the Rio Pargua area, Cuevas and Sanchez (1991)measured a 598-m-thick sequence.
The Floresta Formation conformably overlies the El Tibet Formation. The lowermost 15 m consists ofconglomerates and coarse-grained, quartzitic sandstones, recalling the latter formation; these are interbeddedwith micaceous and feldspathic, well-sorted, ®ne-grained sandstones. The quartzitic sandstones are moreabundant at the base, and become more and more feldspathic, and interbedded with mudstones andclaystones towards the top. In some places, limestone lenses may occur in the transition zone of sandstonesand claystone (Text-®g. 2A). These lenses are located within the lowermost 50 m and are very fossiliferous.They contain the same fauna as that found in the more fossiliferous, upper horizons. In general, theargillaceous part, which represents 85 per cent of the formation, is devoid of fossils.
732 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 2. Synthetic logs of the El Tibet, Floresta and Cuche formations. A, Floresta Formation and transition to theCuche Formation; B, El Tibet Formation and transition to the Floresta Formation (the ®sh symbol indicates remains atthe base of the Floresta Formation at Alto El Uche); C, Cuche Formation (PR1±3, ®sh-bearing localities of Potrero
RincoÂn 1±3). cl, claystone; st, siltstone; sd, sandstone; cg, conglomerate.

The Cuche Formation lies conformably on the Floresta Formation. The transition between the two isshown by changes in the colour of sediments, from yellowish to purple or reddish, alternating in the formof lenses of medium size and thickness; that is, about 1 m thick and 10±15 m broad (Text-®g. 2A).Lithologically, the transition zone is marked by an increase in grain-size, passing from claystones andmudstones to feldspathic sandstones, often in the form of lenticular layers with wavy to parallellamination, sometimes bearing undetermined ichnofossils. The Floresta Formation seems to be anepicontinental marine deposit, corresponding to the high point of the Devonian transgression, whichprobably progressed from north to south over an area now occupied by the Oriental Cordillera of Colombiaand the Girardot Sub-basin of the Upper Magdalena valley, and reached the Quetame Massif.
The age of the Floresta Formation is Emsian±Eifelian. This was established on the basis of its fossils,essentially brachiopods (Caster 1939; McNair 1940; Royo y Gomez 1942; Morales 1965; Barrett 1985, 1988).
Cuche Formation (Text-®g. 2C). This formation overlies the Floresta Formation and is unconformabyoverlain by conglomerates of the Jurassic GiroÂn Formation (Text-®g. 2C). However, in some places, it isalso overlain by the conglomeratic sandstones of the Early Cretaceous Tobasosa Formation, as, forexample, in `Puerto Arepas', about 200 m east of Duitama, along the road to Sogamoso. Since the CucheFormation is always truncated at the top, its entire thickness remains unknown. In Potrero RincoÂn, belowthe discordance with the GiroÂn Formation, it is 540 m thick.
The Cuche Formation consists of medium-sized (c. 1 m) to thin (c. 10 cm) layers and lenses offeldspathic sandstones, claystones and micaceous mudstones. At its base are brownish to pink, argillac-eous mudstones with leaf remains and traces of roots. The basal section of the middle part of the formationconsists of a series of sandstones, strati®ed into medium-sized (1 m) to thick (5 m) layers and lenses; theserepresent palaeo-channels. In the middle and uppermost levels of the middle section lenses of yellowishand light yellowish brown, silty claystone and clayey, ®nely micaceous claystone frequently occur, someof which contain ostracods, bivalves, plants, and ®sh remains. Abundant ®sh remains and isolated lingulidsalso occur in sandstone levels at the top of the middle section and the base of the upper section.
A sedimentological study of the Cuche Formation by Carrillo (1992) provided evidence for depositionin a coastal environment along a clastic shore, with backshore deposits, tidal inlets, tidal ¯ats and mud¯ats.According to this author, the lenses of fossil-rich claystone and clay may indicate a drop in energy levels inthe environment and, hence, the accumulation of deposits that were particularly favourable for thedevelopment of marine life.
The age of the Cuche Formation could not hitherto be reliably established. As pointed out by Mojica andVillarroel (1984), previous palaeontological studies were preliminary and incomplete and did not matchthe stratigraphical observations. In this respect, the present study, based on ®shes, provides strong supportfor a Late Devonian age (possibly Late Frasnian), which agrees with the stratigraphical position of thisformation, above the Floresta Formation.
Geographical and stratigraphical position of the vertebrate-bearing localities
Alto El Uche. This locality is situated 5 km due south-east of BeleÂn (Text-®g. 1), along the BeleÂn-Florestaroad passing by the Las Puentes school (58 550 4000 N, 728 500 4800 W). Stratigraphically, the fossiliferouslevel occurs at the base of the Floresta Formation (Text-®g. 2B), which consists here of interbedded,medium- to ®ne-grained feldspathic sandstones and grey to yellowish claystones. The ®sh remains arefound in association with brachiopod and crinoid remains.
Santa Rosa de Viterbo-Floresta roadcut. This exposure is situated 2´5 km due east-south-east of SantaRosa de Viterbo (58 500 3000 N, 728 550 3600 W), along the road to Floresta (Text-®g. 1). The ®sh remains(the only fossils found here) are rhizodontid scales. They occur in levels belonging to the middle part of theCuche Formation, which consists of reddish sandstones and mudstones.
Tunguaquita. This outcrop is in an old quarry situated 7 km due south-south-east of Santa Rosa de Viterbo(58 450 4600 N, 728 550 4500 W), along the road which starts from the Sagamoso-Duitama highway and leads
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 733

to Santa Rosa de Viterbo, passing by the villages of Cuche and Cachavita (Text-®g. 1). The stratigraphicposition of the exposure is uncertain, yet its lithology suggests that it is within the upper part of the CucheFormation. In addition to ®sh remains, it yields well-preserved fossil plants, including Archaeopteris(C. Berry, pers. comm. 1997).
Quebrada Potrero RincoÂn. The most important fossiliferous levels of the Cuche Formation, not only for®shes but also for bivalves, ostracods, lingulids and plants, are in this section (58 450 5200 N, 728 550 4600 W),which is situated 5´2 km due south-east of Floresta. Fish remains occur in three, closely set outcrops,referred to here as Potrero RincoÂn 1±3 (PR-1±3, Text-®g.1). The ®sh-bearing lenses may correspondlithologically to grey±yellowish claystones, as in the case of Potrero RincoÂn 1 (PR-1, Text-®g. 2C), orgreyish sandstones, as in Potrero RincoÂn 2 (PR-2, Text-®g. 2C). Potrero RincoÂn 3 (PR-2, Text-®g. 2C) isabout 250 m north of Potrero RincoÂn 1. Potrero RincoÂn 1 and 3 are approximately at the same level withinthe section, but Potrero RincoÂn 2 is somewhat higher up (c. 10 m) in the section. Stratigraphically, the threeexposures are in the upper part of the Cuche Formation. Potrero RincoÂn 1 and 3 also yield abundant plantremains.
S Y S T E M A T I C P A L A E O N T O L O G Y
The material from the Floresta and Cuche Formation is generally preserved in ®ne-grained sandstones orclaystones. Most of the exposures are deeply weathered and the bone of the vertebrate remains isextremely fragile. The specimens have, therefore, been studied by removing the altered bone with a brush,and making elastomer casts of their natural impression. The material is housed in the collections of theDepartamiento de Geociencias, Universidad Nacional de Colombia, Bogota (UNC-DG). Casts of theillustrated material and some ®sh-bearing sediment samples are deposited in the collection of theLaboratoire de PaleÂontologie, MuseÂum National d'Histoire Naturelle, Paris.
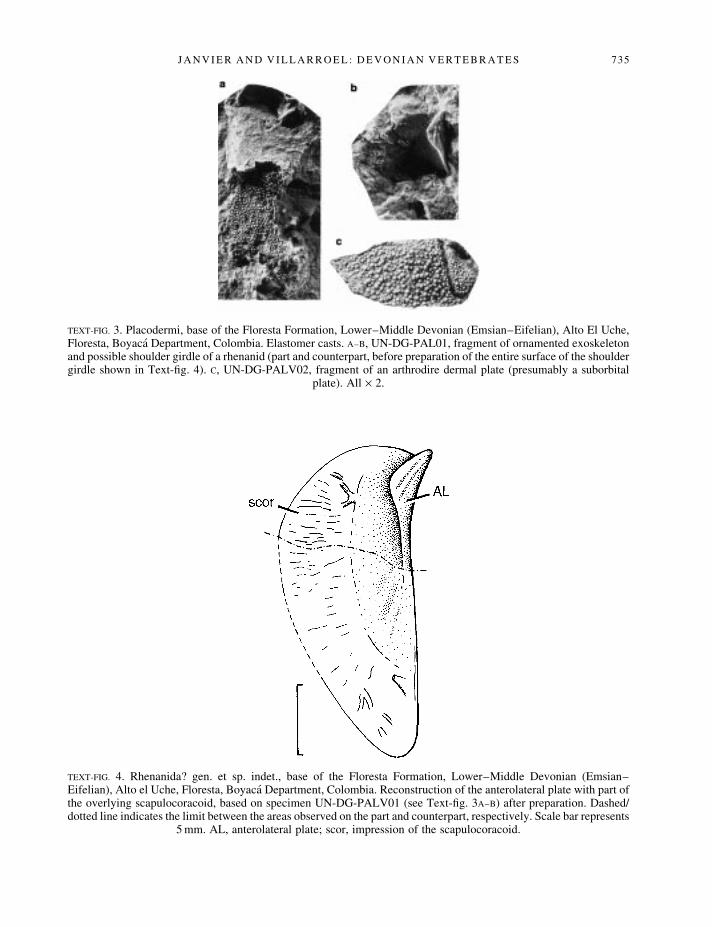
Vertebrates from the Floresta Formation. Placoderm remains have been found at the base of the FlorestaFormation, in the pink sandstone layers that immediately underly the lowermost marine invertebratefaunas. One consists of a small fragment of an arthrodire plate ornamented with large, slightly costulatetubercles, and bearing a recurved sensory-line groove (Text-®g. 3C). It possibly belongs to an earlybrachythoracid arthrodire and might be a portion of a suborbital plate. Two other, closely associatedfragments may belong to a rhenanid. One of these is a dermal bone fragment bearing pointed andsomewhat stellate tubercles (Text-®g. 3A). It overlies a peculiar, elongated oval element that bearsnumerous blood vessel impressions on part of its surface (Text-®gs 3A±B, 4), and whose overall aspectrecalls the scapulocoracoid of rhenanids (Goujet et al. 1993, ®g. 7).
Vertebrates from the Cuche Formation
Class CHONDRICHTHYES Huxley, 1880Subclass ELASMOBRANCHII Bonaparte, 1838
Order XENACANTHIDA Glikman, 1964
Genus ANTARCTILAMNA Young, 1982?
Antarctilamna? sp. indet.
Plate 1, ®gure 1; Text-®gure 5A
Description and remarks. A single chondrichthyan spine was found at Potero RincoÂn 1. It is a broad-based spine,hollowed by a broad central canal, that is widely open posteriorly (Pl. 1, ®g.1). Its ornamentation consists of ®ve,slightly nodose ridges, located on the anterior surface of the spine, the rest of its surface being unornamented. The
734 P A L A E O N T O L O G Y , V O L U M E 4 3
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 735
TEXT-FIG. 3. Placodermi, base of the Floresta Formation, Lower±Middle Devonian (Emsian±Eifelian), Alto El Uche,Floresta, Boyaca Department, Colombia. Elastomer casts. A±B, UN-DG-PAL01, fragment of ornamented exoskeletonand possible shoulder girdle of a rhenanid (part and counterpart, before preparation of the entire surface of the shouldergirdle shown in Text-®g. 4). C, UN-DG-PALV02, fragment of an arthrodire dermal plate (presumably a suborbital
plate). All ´ 2.
TEXT-FIG. 4. Rhenanida? gen. et sp. indet., base of the Floresta Formation, Lower±Middle Devonian (Emsian±Eifelian), Alto el Uche, Floresta, Boyaca Department, Colombia. Reconstruction of the anterolateral plate with part ofthe overlying scapulocoracoid, based on specimen UN-DG-PALV01 (see Text-®g. 3A±B) after preparation. Dashed/dotted line indicates the limit between the areas observed on the part and counterpart, respectively. Scale bar represents
5 mm. AL, anterolateral plate; scor, impression of the scapulocoracoid.

overall morphology and ornamentation are suggestive of Antarctilamna, an elasmobranch that is widely distributed inthe Devonian of Gondwanan regions. Initially described from Australia and Antarctica (Young 1982; Long and Young1995), Antarctilamna has also been recorded from the Emsian of Iran (Blieck et al. 1980) and Saudi Arabia (LelieÁvreet al. 1994) and in the Emsian±Frasnian of Bolivia (Janvier and Suarez-Riglos 1986; Gagnier et al. 1988, 1989; Goujetet al. 1993). This spine differs, however, from typical Antarctilamna spines, which have numerous nodose ridges, byits scarce and comparatively smooth ridges. The only chondrichthyan tooth, from the same outcrop, appears to beincomplete and is preserved as a natural impression. It has a broadly expanded and ventrally concave base, and asingle, oblique cusp (Text-®g. 5A). It may belong to Antarctilamna, but its morphology also shows similarities toanother xenacanthiform elasmobranch, Portalodus, from the Upper Devonian of Antarctica (Long and Young 1995).
Class ACANTHODII Owen, 1846Order CLIMATIIFORMES Berg, 1940
Genus CHEIRACANTHOIDES Wells, 1944?
Cheiracanthoides? sp. indet.
Text-®gure 5B±C
Description and remarks. The acanthodian scales from Potrero RincoÂn 1 have a ¯at, backwardly directed crownornamented with parallel or slightly convergent, rounded ridges (Text-®g. 5B). This type of ornamentation is found inmany acanthodians, in particular climatiiforms and diplacanthids. These scales bear an overall resemblance to scalesusually referred to the Lochkovian±Frasnian climatiid genus Cheiracanthoides (e.g. C. comptus, Wells; Gross 1973,pl. 27, ®gs 3, 5). Owing to their poor preservation, no histologic study of their structure can be made and, therefore, no
736 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 5. Chondrichthyes, Acanthodii, Cuche Formation, Upper Devonian, Potrero RincoÂn 1, Floresta, BoyacaÂDepartment, Colombia. A, Antarctilamna sp. indet., UN-DG-PALV40d; tooth in lingual view, camera lucida drawingof elastomer cast. B±C, Cheiracanthoides? sp. indet.; scale in external (B) and lateral (C) view; reconstruction based on
several imperfect specimens in the sample UN-DG-PALV40. Scale bars represent 1 mm.
E X P L A N A T I O N O F P L A T E 1
Fig. 1. Antarctilamna? sp. indet., UN-DG-PALV04; Cuche Formation, Upper Devonian, Potrero RincoÂn 1, Floresta,Boyaca Department, Colombia; dorsal ®n spine in lateral view, elastomer cast; ´ 2.
Figs 2±14. Bothriolepis sp. indet.; Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta, BoyacaÂDepartment, Colombia; elastomer casts. 2, UN-DG-PALV05; nuchal plate. 3, UN-DG-PALV06; nuchal plate. 4,UN-DG-PALV07; right lateral plate, external view. 5, UN-DG-PALV08; left lateral plate, external view. 6, UN-DG-PALV09; left lateral plate, internal view. 7, UN-DG-PALV10; left lateral plate with elliptical preorbital recess,internal view. 8, UN-DG-PALV11; left lateral plate with pentagonal preorbital recess, internal view. 9, UN-DG-PALV12; anterior median dorsal plate, internal view. 10, UN-DG-PALV13; anterior median dorsal plate, externalview. 11, UN-DG-PALV14; anterior median dorsal and anterior dorsolateral plates, internal view. 12, UN-DG-PALV15; imperfect anterior median dorsal plate with narrow levator fossa, internal view. 13±14, UN-DG-PALV16;posterior median dorsal plate in internal (13) and external (14) view. All ´ 2.

P L A T E 1
JANVIER and VILLARROEL, Antarctilamna?, Bothriolepis

more precise determination can be given. It should be kept in mind that only the external morphology of these scalesrecalls Cheiracanthoides, and that similar, ribbed scales also occur in several Late Devonian acanthodians, such as theacanthodiform Homalacanthus and various diplacanthids.
Class PLACODERMI McCoy, 1848Subclass ANTIARCHA Cope, 1885
Family BOTHRIOLEPIDIDAE Cope, 1886
Genus BOTHRIOLEPIS von Eichwald, 1840
Bothriolepis sp. indet.
Plate 1, ®gures 2±14; Plates 2±3; Text-®gures 6±9
Description and remarks. The majority of the vertebrate remains collected at Potrero RincoÂn 2 and 3 are dermal platesof a bothriolepidid antiarch whose characters match the de®nition of the genus Bothriolepis (Young 1988); that is,reticulate ornamentation in juvenile and adult, anterior median dorsal plate broadest across lateral corners, andmixilateral plate broadest through its dorsal corner.
Among the three, apparently undistorted, lateral plates of Bothriolepis from Potrero RincoÂn 2, showing the internalsurface and the lateral part of the preorbital recess, two suggest a roughly elliptical recess (pror, Text-®g. 7A; Pl. 1, ®gs6±7), while the third indicates a pentagonal recess (pror, Text-®g. 7B; Pl. 1, ®g. 8). This, and the slightly differentproportions between the two types of lateral plates, suggests that they represent two different species. Long (1983) andYoung (1988) regarded the elliptical (`simple') shape as the plesiomorphic condition for Bothriolepis. In the lateralplates with an elliptical preorbital recess, there may be an enlarged pit (lp, Text-®g. 7A) anterior to the transverse lateralgroove (spiracular groove, Young and Zhang 1992; spg, Text-®g. 7A), as in the four Australian and Antarctic speciesB. barretti, B. cullodenensis, B. fergusoni, and B. gippslandiensis. In both types of lateral plates, the anteriorattachment area for the submarginal plate is of normal type; that is, not separated from the area for the prelateral plate(aSM� PrL, Text-®g. 7A). In this respect, these lateral plates are closely similar to that of Bothriolepis sp. described byYoung (1984, pl. 57) from the Frasnian of Gogo (Australia); that is, with a small, rounded anterior articular knob forthe submarginal plate (aSM�PrL, Text-®g. 7A); but they differ from that of, for example, B. canadensis Whiteaves(StensioÈ 1948, ®g. 12), which has separate attachment areas for the submarginal and prelateral plates.
The external surface of the nuchal and lateral plates are as in other Bothriolepis species (Nu, L, Text-®g. 9A; Pl. 1,®gs 2±5). Apart from these isolated skull-roof plates (all nuchal and lateral plates), a poorly preserved skull-roof hasbeen found in Potrero RincoÂn 2 (Text-®g. 6) and another, more imperfect skull roof can be seen to the left of the AMDplate in Plate 3, ®g. 2 (Skr.).
All the anterior median dorsal plates (AMD, Text-®g. 9A) from Potrero RincoÂn 2 and 3 are rather homogeneous inaspect and proportion (Pl. 1, ®gs 9±11; Pl. 3, ®gs 2±3; Text-®g. 7C), with the exception of a single, incompletespecimen (Pl. 1, ®g. 12), that has a narrow levator fossa, and in which the postlevator cristae form a narrower angle. Allother AMD plates display classic Bothriolepis features, and are broadest at the level of the lateral corners (lc, Text-®g.7C). The lateral corners are situated in the posterior third of the total length of the plate. The postlevator cristae form anangle of approximately 70 degrees (plc, Text-®g. 7C), and the anterior margin of the plate is broad and slightly convex.There is a distinct anterior ventral pit (pt1, Text-®g. 7C), situated at the level of the anterior third of the total platelength. The external surface shows a shallow median ridge on the posterior two thirds of the plate length.
738 P A L A E O N T O L O G Y , V O L U M E 4 3
E X P L A N A T I O N O F P L A T E 2
Figs 1±11. Bothriolepis sp. indet.; Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta, BoyacaÂDepartment, Colombia; elastomer casts. 1, UN-DG-PALV17; right anterior dorsolateral plate, external view. 2±3, UN-DG-PALV18; left anterior ventrolateral plate in external (2) and internal (3) views. 4, UN-DG-PALV19;right anterior ventrolateral plate, external view. 5, UN-DG-PALV20; brachial process of right anterior ventrolateralplate. 6, UN-DG-PALV23; right mixilateral plate, external view. 7, UN-DG-PALV24; median ventral plate,external view. 8, UN-DG-PALV25; left posterior ventrolateral plate, internal view. 9±10, UN-DG-PALV26;marginal lateral plate 2 of the left pectoral ®n, in dorsal (9) and ventral (10) views. 11, UN-DG-PALV27; rightanterior dorsolateral plate, internal view. All ´ 2.

P L A T E 2
JANVIER and VILLARROEL, Bothriolepis

The posterior median dorsal plate (PMD, Text-®g. 9A; Pl. 1, ®gs 13±14) also bears a slight median ridge that issomewhat sharper anteriorly. Its ventral surface is similar to that in other Bothriolepis species, with a distinct posteriorventral pit (pt2, Text-®g. 7D), a ventral tuberosity (tb, Text-®g. 7D) and a sharp posterior internal transverse crest (citp,Text-®g. 7D).
The anterior dorsolateral plate (ADL, Text-®g. 9A; Pl. 2, ®gs 1, 11) has a relatively deep lateral lamina and broaddorsal lamina. Otherwise it is similar to that of most other Bothriolepis species.
The mixilateral plate (MxL, Text-®g. 9A; Pl. 2, ®g. 6) is virtually indistinguishable from that of Bothriolepiscanadensis (StensioÈ 1948, ®g. 153), except that its posterior oblique dorsal sensory-line groove (dlg2, Text-®g. 9A)runs somewhat closer to the suture with the PMD plate.
The internal surface of the anterior ventrolateral plate (AVL, Text-®g. 9B) has a sharp main anterior transverseinternal crest (cita, Text-®g. 8A; AVL, Pl. 3, ®g. 1), sometimes doubled posteriorly by a second, shallower crest(cita 1±2, Text-®g. 8C; Pl. 1, ®g. 3). This variation is frequently oberved in other Bothriolepis species (e.g. Young1988, ®g. 50B). The brachial process (brp, Text-®g. 8A±B; Pl. 2, ®gs 4±5) and associated structures are as in otherBothriolepis species. The axillary foramen is small, oval in shape or slightly longer than high (axf, Text-®g. 8), andresembles that of B. canadensis. The posterior ventrolateral plate (PVL, Text-®g. 9B; Pl. 2, ®g. 8) has a relatively longposterior ventral lamina and moderately deep lateral lamina. The median ventral plate (MV, Text-®g. 9B; Pl. 2, ®g. 7)has the same size and proportions as in B. canadensis.
Although isolated pectoral ®n plates are abundant in Potrero RincoÂn 2, we have reconstructed the proximal portionof the pectoral ®n on the basis of a single specimen, in which the proximal central dorsal (Cd1, Text-®g. 9A; Pl. 3,®g. 1) and marginal lateral (Ml2, Text-®g. 9A; Pl. 3, ®g. 1) plates are partly articulated. This shows that the pectoral®ns were relatively slender, with a rather even series of lateral marginal spines (Ml2, Pl. 3, ®g. 1). However, an isolatedMl2 plate (Pl. 2, ®gs 9±10) shows a strongly convex lateral margin, armed with prominent lateral spines. Thismorphology, when present, occurs only in adult individuals (Werderlin and Long 1986), yet this plate is smaller thatthe Ml2 plate of the specimen used for reconstruction (Ml2, Pl. 3, ®g. 1), which does not show this particular feature.Again, this may be an indication that at least two species of Bothriolepis occur in Potrero RincoÂn 2.
Most of the Bothriolepis plates from Potrero RincoÂn 2 and 3 are found as isolated elements, but there are some partiallyarticulated thoracic armours, showing ensembles of plates which clearly belong to the same individual (Pl. 1, ®g. 11; Pl.3, ®g. 1). This suggests that complete, articulated specimens may eventually be found in this locality. In Text-®gure 9, wepropose a synthetic reconstruction of one of the two presumed species. The outline of the lateral plate is based on thespecimens with an oval preorbital recess. Although mainly restored from isolated plates (except for the proximal part ofthe pectoral ®n, which is based on the partly articulated specimen shown in Plate 3, ®g. 1), the good correspondencebetween the overlap areas of the similar-sized plates suggests that this reconstruction is relatively accurate.
There are presently about 80 species included in the genus Bothriolepis, a large number of them being de®ned on thebasis of isolated and often undiagnostic plates. This is because Bothriolepis remains are extremely abundant in Middle
740 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 6. Bothriolepis sp. indet., UN-DG-PALV29c; Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta,Boyaca Department, Colombia. Imperfect skull-roof in dorsal view (camera lucida drawing of elastomer cast). Scalebar represents 10 mm. L, lateral plate; obtnu, obtected nuchal area; onf, orbitonasal fenestra; Pm, postmarginal plate;
PNu, paranuchal plate.

and Late Devonian marginal facies, in particular in Old Red Sandstone-like facies, and display a high rate of individualvariability, and differences due to allometric growth. Nevertheless, some species (e.g. B. gippslandiensis, B.cullodenensis) are readily recognized by conspicuous, unique characters, such as a prominent crest on the mediandorsal plates. After a ®rst attempt by Long (1983), Young (1988) made a thorough analysis of the interrelationships ofthe Bothriolepis species on the basis of the 37 best known species. He managed to identify some characters that
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 741
TEXT-FIG. 7. Bothriolepis sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta, BoyacaÂDepartment, Colombia. A±B; UN-DG-PALV10 and UN-DG-PALV11, respectively; left lateral plates in internalview, possibly belonging to two different species with elliptical (A) and pentagonal (B) preorbital recess (cameralucida drawings of elastomer casts; see also Pl. 2, ®gs 7±8). C, reconstruction of the anterior median dorsal plate ininternal view, based on UN-DG-PALV12 (see also Pl. 2, ®g. 9). D, reconstruction of the posterior median dorsal platein internal view, based on UN-DG-PALV16 (see also Pl. 2, ®gs 13±14). Scale bar represents 10 mm. aSM, posteriorarea for articulation of the submarginal plate; aSM� PRL, anterior area for articulation of the submarginal plate andfor the prelateral plate; citp, posterior internal transverse crest; lc, lateral corner; lf, levator fossa; lp, lateral pit;oaADL, overlap area for the anterior dorsolateral plate; oaAMD, overlap area for the anterior median dorsal plate;oaMxL, overlap area for the mixilateral plate; plc, postlevator crest; pror, preorbital recess; pt1, anterior median dorsal
pit; pt2, posterior median dorsal pit; tb, internal tuberosity.

allowed de®nition of a number of clades. Within these clades, however, the relationships between the species remainlargely unresolved. One of Young's major clades includes all species with a pentagonal preorbital recess. In turn, thisclade includes a clade in which the pentagonal preorbital recess has a tri®d shape, and another in which the axillaryforamen is longer than high and the preorbital region of the skull-roof is elongated. Besides the large cladecharacterized by a pentagonal (to tri®d) preorbital recess, two other minor clades are de®ned. One is characterized
742 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 8. Bothriolepis sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta, BoyacaÂDepartment, Colombia. Anterior ventrolateral (AVL) plates, camera lucida drawings of elastomer casts. A, UN-DG-PALV21; right AVL plate in medial, internal view. B, UN-DG-PALV18a (see also Pl. 2, ®g. 2); left AVL plate inlateral view. C, UN-DG-PALV18b; left AVL plate in dorsomedial, internal view (see also Pl. 2; ®g. 3). Scale barrepresents 10 mm. axf, axillary foramen; brp, branchial process; cita, anterior internal transverse crest (cita 1, 2, when
doubled); oa ADL, overlap area for anterior dorsolateral plate.

by the arrangement of the pectoral ®n plates, and the other by the presence of an enlarged pit on the internal surface ofthe lateral plate. The latter includes a small group of Gondwanan species characterized by a trilobate preorbital recess.Of the two Bothriolepis species from the Cuche Formation, one may belong to the group characterized by a pentagonalpreorbital recess, while the other may belong to the group characterized by an enlarged internal pit of the lateral plate.
This brief survey of the Bothriolepis material from the Cuche Formation illustrates once more the dif®culty inde®ning and identifying the Bothriolepis species that display no conspicuous, unique characters. De®nitions based onplate proportions are extremely suspect, considering the wide range of individual variation and allometric growth(Graham-Smith 1978; Werdelin and Long 1986; Luksevics 1995). Practically all the plates found in Potrero RincoÂn 2match, in ornamentation, shape or proportions, their homologue in one or another individual of B. canadensis. Only thetwo different shapes of the preorbital recess, inferred from the lateral plates of the skull-roof, and the lateral marginalplates 2 of the proximal paired ®n segment suggest the presence of two different species. However, no such differencecan be inferred from the plates of the trunk armour. Instead of further complicating the systematics of Bothriolepis bynaming species on the basis of isolated plates, we prefer here to postpone the identi®cation of the ColombianBothriolepis species until articulated material is discovered in these localities.
Family ASTEROLEPIDIDAE Traquair, 1888
Genus ASTEROLEPIS von Eichwald, 1840?
Asterolepis? sp. indet.
Text-®gure 10
Description and remarks. Some large and fragmentary antiarch plates are tentatively referred here to the familyAsterolepididae on the basis of their vermiculate ornamentation, which tends to develop into parallel ridges, a type ofornamentation that frequently occurs in the Late Devonian asterolepidids Asterolepis and Remigolepis, but may alsooccur in some other antiarchs (e.g. Bothriolepis fergusoni Long and Werdelin 1986). A fragment of a large plate from
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 743
TEXT-FIG. 9. Bothriolepis sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta, BoyacaÂDepartment, Colombia. Attempted reconstruction of the dermal armour of one of the species from the locality,based on isolated plates except for the proximal part of the pectoral ®n and the posterolateral part of the skull roof.Unknown plate limits indicated by dashed lines. A, dorsal view. B, ventral view of the trunk armour only. Scale barrepresents 10 mm. ADL, anterior dorsolateral plate; AMD, anterior median dorsal plate; AVL, anterior ventrolateralplate; brp, brachial process; Cd1, dorsal central plate 1; dlg2, posterior oblique dorsal sensory-line groove; L, lateralplate; Ml2, lateral marginal plate 2; MV, median ventral plate; MxL, mixilateral plate; Nu, nuchal plate; Pm,postmarginal plate; PMD, posterior median dorsal plate; PNu, paranuchal plate; PVL, posterior ventrolateral plate.

Tunguaquita (Text-®g. 10C) is most probably the right posterolateral portion of an anterior median dorsal plate. Itsornamentation is strikingly similar to that of Asterolepis radiata Rohon from the Early Frasnian Amata beds of Estoniaand Latvia (Karatajute-Talimaa 1963, pl. 15). Two other fragments, from Potrero RincoÂn 1, can also be referred to theAsterolepididae. One of the specimens is a posterior median part of the skull roof of a large antiarch, whoseornamentation is also similar to that of Asterolepis (Text-®g. 10A). It seems to show two divergent central sensory-linegrooves, which would rather suggest a bothriolepid pattern, but in some Asterolepis species (e.g. A. radiata), thelineation of the ornamentation in this region of the skull roof may also be developed in the same way. The otherspecimen is a portion of an anterior dorsolateral plate (Text-®g. 10B) that is somewhat similar to that of A. radiata®gured by Karatajute-Talimaa (1963, pl. 17, ®g. 2).
Subclass ARTHRODIRA Woodward, 1891
Arthrodira gen. et sp. indet.
Plate 4; Text-®gure 11
Description and remarks. Arthrodires are represented in this fauna by some poorly preserved plates from PotreroRincoÂn 3. All these plate fragments bear the same type of ornamentation, which consists of very large, scatteredtubercles with a slightly ridged apex. Only two plates can be clearly identi®ed: a nuchal plate (Pl. 4, ®gs 1±2; Text-®g.11), and an anterior portion of a spinal plate (Pl. 4, ®g. 3). Other undetermined fragments show large overlap areas thatare suggestive of brachythoracid plates. One of them (Pl. 4, ®g. 5) may possibly be the anterolateral portion of aposterior ventrolateral plate, with part of the overlap area for the anterior ventrolateral plate and an unornamentedlateral margin. If this interpretation is correct, this plate would not have possessed any dorsal process contacting theposterolateral plate, and its pectoral fenestra may have been widely open posteriorly. Another possibility is that it ispart of a very deep posterior dorsolateral plate of groenlandaspidid type. Other fragments, although showing overlapareas, cannot be identi®ed with certainty (Pl. 4, ®gs 4, 6).
The nuchal plate is the most informative specimen. It is strongly arched transversally, and its internal surface showslong, parallel overlap areas for the paranuchal plates (oaPNu, Text-®g. 11B). On its posterior part, the ventral surfacebears paired, elongated inranuchal pits (inup, Text-®g. 11B), separated by a sharp ridge, while the anterior part isslightly convex. This structure is somewhat suggestive of phlyctaeniid arthrodires (Goujet, 1984, ®g. 32), andgroenlandaspidids, but more so of the primitive brachythoracids (Heintz, 1934, ®g. 5; Bystrov 1957, ®g. 9; White1978; Young 1981, ®g. 6; LelieÁvre 1984, ®g. 3; Otto 1992). The spinal plate, though fragmentary, differs from that ofthe phlyctaeniids (including the Late Devonian groenlandaspidids) in being gently rounded transversally, and is moresimilar to that of holonematids (Miles 1971, ®g. 85).
These arthrodire remains represent only the second occurrence of this widespread placoderm group in the UpperDevonian of South America, following the discovery of a dinichthyid in the Famennian of Bolivia (Diaz-Martinezet al. 1996).
Class OSTEICHTHYES Huxley, 1880Subclass ACTINOPTERYGII Klein, 1885
Family STEGOTRACHELIDAE Gardiner, 1963
Stegotrachelidae gen. et sp. indet.
Text-®gures 12±14
Description and remarks. Devonian actinopterygians from South America were hitherto represented by just a few
744 P A L A E O N T O L O G Y , V O L U M E 4 3
E X P L A N A T I O N O F P L A T E 3
Figs 1±5. Bothriolepis sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta, Boyaca Department,Colombia; elastomer casts. 1, UN-DG-PALV28a-d; right anterior ventrolateral plate (AVL) in internal view,articulated to the central dorsal plate 1 (Cd1) and the lateral marginal plate 2 (Ml2) of the right pectoral ®n, andisolated lateral marginal plate 2 (Ml2, to the left). 2, UN-DG-PALV29a,b; anterior median dorsal plate (AMD) ininternal view, partly overlying an imperfect skull-roof (Skr) in external view, and isolated central dorsal plate (Cd1).3, UN-DG-PALV30; anterior median dorsal plate, internal view. 4±5, UN-DG-PALV31; dorsal central plate 1 ofright pectoral ®n in external (4) and internal (5) view. All ´ 2.

P L A T E 3
JANVIER and VILLARROEL, Bothriolepis

isolated scales from the Famennian Iquiri Formation of southern Bolivia (Janvier and Suarez-Riglos 1986), an isolatedcleithrum from the Givetian of the Sica Sica Formation of the Bolivian Altiplano (Gagnier et al. 1989), and isolatedscales from deposits of supposed Late Famennian age in eastern Brazil (Janvier and Melo 1987). The Bolivianspecimens belong to a rather generalized actinopterygian type, as represented by the Late Devonian generaMoythomasia and Mimia (Jessen 1968; Gardiner 1984). All the actinopterygian remains described here are fromthe clayey mudstone and claystone levels of Potrero RincoÂn 1.
The best specimen is represented by the right half of a skull roof that certainly comprises the parietal, postparietal,intertemporal and supratemporal, although no suture can be traced with certainty, even by immersion in aniseed oil(Skr, Text-®g. 12; Text-®g. 13A). The ornamentation consists of smooth, sinuous ganoine ridges, as in most earlyactinopterygians. Interruptions in these ridges or changes in their orientation sometimes indicate the position of thesutures between the dermal bones, in particular between the parietal and postparietal. Laterally, the specimen gives riseto a broad, descending expansion that probably includes the dermosphenotic. Its anterolateral limit thus represents the
746 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 10. Asterolepis? sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 1 (A±B) and Tunguaquita (C),Boyaca Department, Colombia. Elastomer casts. A, UN-DG-PALV32; nuchal and postpineal regions of a large skull-roof. B, UN-DG-PALV33, imperfect anterior dorsolateral plate. C, UN-DG-PALV34; fragment of a large mixilateral
plate. All ´ 1´5.

orbital margin and suggests the presence of a rather large orbit. Medially, the parietal is embayed by an irregular notchthat corresponds to the right half of the pineal fenestra, which included the pineal foramen (pi, Text-®g. 13A). Theposterior edge of the skull-roof is deeply embayed, and its unornamented margin was overlapped by the extrascapulars.
The anterior, middle and posterior pit lines (ppl, mpl, Text-®g. 13A), as well as the supraorbital sensory-line canal,are visible (soc, Text-®g. 13A), but only a small part of the cephalic portion of the infraorbital canal can be traced onthe supratemporal (ll, Text-®g. 13A).
Adjacent to the skull-roof, the external impression of an incomplete left lateral gular plate (gul, Text-®g. 12) showsthe same ornamentation as the skull roof and bears a well-marked gular pit-line (gpl, Text-®g. 13B). Its anterior end isgently ¯exed and unornamented, possibly for the overlap by the median gular plate (oamg, Text-®g. 13B). This gularplate seems to be remarkably large if regarded as belonging to the same individual as the skull roof. However, suchlarge lateral gulars, as long as the length of the parietal� postparietal, are known in Devonian actinopterygians such asHowqualepis (Long 1988) and Cheirolepis (Pearson and Westoll 1979). By contrast, Moythomasia, Mimia and mostother post-Devonian actinopterygians have rather short lateral gulars or no gulars at all (Gardiner 1984).
The proportions of the skull-roof can be compared with those of the best known Devonian actinopterygians. Theonly suture that can be inferred from the changes in the orientation of the ridges of the ornamentation is that betweenthe parietal and postparietal, and it suggests that the postparietal is slightly shorter than the parietal. Such a proportionof the parietal relative to the postparietal is clearly more similar to that of Middle±Late Devonian actinopterygians(e.g. Cheirolepsis, Howqualepis, Mimia, Moythomasia, and Osorioichthys; Jessen 1968; Pearson 1982; Gardiner 1984;Long 1988; Taverne 1997), than to that of the Lower Devonian Dialipina (Schultze 1992), where the postparietal islonger than the parietal. Laterally, the supratemporal and intertemporal are almost certainly present, but it is uncertainif the dermosphenotic is included in the skull-roof. This is, however, suggested by its large, lateral expansion though,conversely, there is no evidence for a spiracular slit, which normally opens between the dermosphenotic, intertemporaland supratemporal. The ornamentation, the sensory-line pattern and the very large pineal fenestra of the specimenaccord best with the condition in Mimia, Moythomasia (Gardiner 1984), and an unnamed Middle Devonianactinopterygian from Antarctica (Young 1991). None of the characters or character states used by Long (1988) andGardiner and Schaeffer (1989) in reconstructing early actinopterygian interrelationships can be observed in thespecimen. Nevertheless, the relative size of the parietal and postparietal, and the large size of the pineal fenestra wouldenable this form to be placed below the node of the `Kentuckia group' and all other actinopterygians in the cladogramproposed by Gardiner and Schaeffer (1989).
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 747
TEXT-FIG. 11. Arthrodira gen. et sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 3, Floresta, BoyacaÂDepartment, Colombia. A±B, UN-DG-PALV35; nuchal plate in external (A) and internal (B) aspects (camera lucida;see also Pl. 4, ®gs 1±2). Scale bar represents 10 mm. inup, infranuchal pits; oaPNu, overlap area for the paranuchal
plate.

Other bones found in the same level include the posterior part of a lower jaw (Text-®g. 14B), whose dentary bearsminute teeth with an acrodine cap. The chevron-shaped ornamentation of the posterior part of the dentary is similar tothat in Mimia (Gardiner 1984, ®g. 90). A right maxillary (Text-®g. 14A) is of generalized actinopterygian type, yet thestrong ventral curvature and the ornamentation are more suggestive of Mimia than Moythomasia (Gardiner 1984,®g. 60).
Many actinopterygian scales found at Potrero RincoÂn 1 are of a generalized actinopterygian type, with a prominentand pointed dorsal peg (Text-®g. 14C). The ganoine ridges on their surface are smooth and tend to fuse into acontinuous ganoine layer pierced by large, slit-shaped pores. Some scales are more or less rounded in shape andsuggestive of the ventral scales of Moythomasia (Jessen 1972, ®g. 1E). Some asymmetrical, arrowhead-shaped scalescan be regarded as fringing fulcra of paired ®ns (Text-®g. 14D), and thus evidence for an actinopteran af®nity.
On the basis of these dermal bones and scales, and under the assumption that they all belong to the same species, onecan ascertain that this actinopterygian is an actinopteran, and possibly closer to Mimia than to any other form.
Subclass SARCOPTERYGII Romer, 1955Infraclass RHIPIDISTIA Cope, 1887
Order POROLEPIFORMES Jarvik, 1942Family HOLOPTYCHIIDAE Owen, 1860
Genus HOLOPTYCHIUS Agassiz, 1839
Holoptychius sp. indet.
Plate 5, ®gures 9±10
Description and remarks. Some large, rounded scales are ornamented with sinuous ridges that develop anteriorly intorows of small, pointed or spoon-shaped tubercles (Pl. 5, ®gs 9±10). These scales are remarkably similar to those ofsome Late Devonian species of the porolepiform Holoptychius, in particular H. jarviki Cloutier and Schultze (1996)from the Frasnian of Quebec. Various species of this genus have been erected on the basis of scale ornamentation but,as pointed out by Cloutier and Schultze (1996), there are doubts as to the validity of this character, as it may vary withthe age of individuals.
Order OSTEOLEPIFORMES Berg, 1937Family OSTEOLEPIDIDAE Cope, 1889
Osteolepididae gen. et sp. indet.
Text-®gure 15A
Description and remarks. Some rare, cosmine-covered dermal bone and scale fragments and an isolated opercular(Text-®g. 15A) occur in Potrero RincoÂn 2. Although the cosmine is extremely weathered and does not allow reliablehistological study, its surface does not show any trace of Westoll-lines and thus rules out the possibility that it belongsto a dipnoan, despite the fact that the overall shape of the opercular is quite similar to that of the opercular of certaindipnoans, such as Howidipterus and Barwickia (Long 1992, ®g.6). The shape of this opercular is also suggestive of that
748 P A L A E O N T O L O G Y , V O L U M E 4 3
E X P L A N A T I O N O F P L A T E 4
Figs. 1±6. Arthrodira gen. et sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 3, Floresta, BoyacaÂDepartment, Colombia; elastomer casts. 1±2, UN-DG-PALV35; nuchal plate in external (1) and internal (2) view. 3,UN-DG-PALV36; portion of a spinal plate, external view. 4, UN-DG-PALV37; undetermined plate fragment withoverlap area, external view. 5, UN-DG-PALV38; undetermined plate fragment, possibly a posterior ventrolateralplate, external view. 6, UN-DG-PALV39; undetermined plate fragment, external view. All ´ 2.

P L A T E 4
JANVIER and VILLARROEL, Arthrodira

bone in generalized, Middle Devonian osteolepidids (Jarvik 1948), rather than that of Late Devonian andCarboniferous forms, such as the Megalichthyinae.
Order RHIZODONTIDA Andrews and Westoll, 1970Family RHIZODONTIDAE Traquair, 1881
Genus STREPSODUS? Huxley and Etheridge, 1865
Strepsodus? sp. indet.
Plate 5, ®gures 1±8; Text-®gure 15B±C
Description and remarks. A rhizodontid is present in the three exposures of Potrero RincoÂn and represented by a largenumber of scales, a dentary and a cleithrum, which all resemble those of Strepsodus more than any other knownrhizodontid, except for the unornamented surface of the dentary.
The dentary bears no distinct ornamentation, but the marginal teeth show a slight lingual curvature, with anenlarged, sigmoid parasymphysial tusk, as in Strepsodus? anculonamensis Andrews (1985, ®g. 9C), Barameda
750 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 12. Actinopterygii gen. et sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 1, Floresta, BoyacaÂDepartment, Colombia. Right half of a skull-roof and impression of a gular plate, associated with the impression of abivalve (UN-DG-PALV40, sample initially labelled as cche-III-84-002); ´ 5. biv, bivalve; gul, gular plate; Skr, skull-
roof.

decipiens Long (1989, ®g. 7B) and Gooloogongia loomesi Johanson and Ahlberg (1998, ®g. 1a). The structure of theteeth could not be observed as they are mostly preserved as impressions, but the base of the parasymphysial dentarytusk is distinctly grooved, thereby suggesting some basal infolding of the dentine (Pl. 5, ®g. 2; dt, Text-®g. 15B±C). Inaddition, the enamel of the tooth surface shows extremely thin striae, as in Strepsodus. The parasymphysial tusk isrounded in section and thus different from the laterally compressed tusks of Rhizodus, Screbinodus and Notorhizodon(Andrews 1985; Young et al. 1992). The dentary differs from that of Carboniferous rhizodontids known to date inbeing somewhat deeper. The symphysial region bears a shallow depression that extends anteriorly to the largeparasymphysial tusk (sym, Text-®g. 15B). Posterior to this area there is a deep depression, bounded anteriorly by aprominent, curved ridge.
The cleithrum is preserved as an internal impression only (Pl. 5, ®g. 8). However, it clearly shows the characteristic`waisted' and twisted shape, with a narrow, paddle-shaped dorsal blade and a large, strongly ¯exed, ventral lamina. Itis quite similar to the cleithrum of S.? anculonamensis (Andrews 1985, ®g. 11) and Strepsodus sp. from the Famennianof south-eastern Turkey (Janvier et al. 1984, ®g. 9A).
The scales are rounded in shape and generally show a distinct median `boss' in the centre of the internal surface (Pl.5, ®gs 3±4), which frequently occurs in rhizodontids and tristichopterids (Jarvik 1980; Andrews 1985; Long 1989).The external ornamentation of these scales is variable. Some are ornamented with very thin, sinuous ridges, like thoseof S.? anculonamensis (Andrews 1985, ®g. 5A), while others show larger, branching ridges (Pl. 5, ®g. 7). Some scalesexposed in internal view show the secondary tubules of the lateral line canals and sometimes large, scattered tuberclesor outgrowths of the basal surface (Pl. 5, ®gs 5±6). In these respects, they are extremely similar to the unnamedrhizodontid scales described by Andrews (1985, ®g. 5g±h) from the Carboniferous of Foulden, Scotland. Such scaleshave also been found in the red sandstones and siltstones of the roadcut of the Santa Rosa de Viterbo-Floresta road,which probably belong to the middle part of the Cuche Formation.
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 751
TEXT-FIG. 13. Actinopterygii gen. et sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 1, Floresta, BoyacaÂDepartment, Colombia. A, UN-DG-PALV40a; right half of a skull-roof. B, UN-DG-PALV40b; left gular plate. Cameralucida drawings based on the specimens in Text-®g. 12. Scale bar represents 1 mm. apl, anterior pit line; gpl, gular pitline; ll, main lateral sensory line; mpl, middle pit line; oamg, overlap area for the median gular; ppl, posterior pit line;
soc, supraorbital sensory-line canal.

By its unornamented and rather deep dentary, this Colombian rhizodontid recalls Gooloogongia from thesupposedly Early Famennian locality of Canowindra, Australia, a form which is, however, distinguished from allother rhizodontids by its posteriorly tilted parasymphysial tusks, protruding lower jaw, and untwisted and unwaistedcleithrum (Johanson and Ahlberg 1998). However, the very ®nely striated surface on the tusks, strongly waistedcleithrum, and scale morphology are characters in which the Colombian rhizodontid is most similar to Strepsodus?anculonamensis, the only species referred (with some doubt) to Strepsodus, and known from an articulated specimen.
D I S C U S S I O N
These two vertebrate assemblages from the Floresta and Cuche Formation, respectively, yield a number ofhigher taxa that were hitherto unrecorded from the Devonian of South America, except for a brief mentionby Young (1993a). As for the two placoderm remains from Floresta Formation, rhenanids were previouslyreported from Bolivia (Goujet et al. 1985, 1993), but no arthrodire had ever been recorded from the LowerDevonian of South America. With regard to the vertebrate assemblage from the Cuche Formation, theantiarchs and sarcopterygians were hitherto undescribed from the Devonian of South America (see,however, Young 1993a), and the other taxa (Antarctilamna, acanthodians, arthrodires and actinopter-ygians) were known from the Middle±Upper Devonian of Bolivia (Gagnier et al. 1989; Janvier andSuarez-Riglos 1986, 1989; Diaz-Martinez et al. 1996). These assemblages provide information about theages and palaeoenvironments of the two formations, but mainly for the Cuche Formation, and also new
752 P A L A E O N T O L O G Y , V O L U M E 4 3
TEXT-FIG. 14. Actinopterygii gen. et sp. indet., Cuche Formation, Upper Devonian, Potrero RincoÂn 1, Floresta, BoyacaÂDepartment, Colombia. A, UN-DG-PALV41; imperfect right maxillary, with a section through its posterior part. B,UN-DG-PALV42, posterior portion of a right dentary in external view. C, UN-DG-PALV40d; body scale in external
aspect. D, UN-DG-PALV43, paired ®n fulcrum in anterior view. Scale bars represent 5 mm (A) and 1 mm (B±D).

insights regarding the faunal relationships between western Gondwana and Euramerica in Late Devoniantimes.
Biostratigraphy
The basal layers of the Floresta Formation are well dated by a rich invertebrate fauna that indicates a LateEmsian±Eifelian age. Moreover, the El Tibet Formation, which comformably underlies the FlorestaFormation, is now dated as Emsian on the basis of palynology. The Devonian series described by Boinetet al. (1986) in north-eastern Colombia, east of the Santander Massif, comprises some plant- and marineinvertebrate-bearing levels that are restricted to the base of a thick, reddish siliciclastic series. Thesefossiliferous beds have been regarded as early Middle Devonian in age, and may correspond to thefossiliferous levels of the Floresta Formation. The few ®sh remains from the Floresta Formation(doubtfully referred here to a brachythoracid arthrodire and a rhenanid) do not add any further informationas to its age. They only indicate that a ®sh fauna is present in this formation, which has the potential toprovide more detailed biostratigraphical information.
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 753
TEXT-FIG. 15. Sarcopterygii, Cuche Formation, Upper Devonian, Potrero RincoÂn 2, Floresta, Boyaca Department,Colombia. A, UN-DG-PALV44; Osteolepididae? gen. et sp. indet., right opercular in internal view, camera lucidadrawing. B±C, Strepsodus? sp. indet., UN-DG-PALV45; detail of the symphysial region in medial view (B) andreconstruction of a right lower jaw in lateral view (C) (see also Pl. 9, ®gs 1±2). Scale bars represent 5 mm (A±B) and
10 mm (C). dt, dentary tusk; sym, symphysial area.

The ®sh assemblage of the Cuche Formation is, by contrast, far more informative, as it representspractically the only fossils from this formation, along with a few lingulids, bivalves, ostracods,conchostracans, and relatively abundant plant remains.
Bothriolepis is ®rst known from the Early Givetian in Australia and Antarctica (Young 1993b) and fromthe Mid Eifelian in China (Wang 1993), but only from the Late Givetian in Euramerica (Dineley andLoef¯er 1993). It appears to have become extinct at the end of the Famennian, although there are somedoubtful Tournaisian records. The other, larger antiarch from this fauna, Asterolepis? (given that thisidenti®cation is correct) is known from the Eifelian to the Late Frasnian. The arthrodire from PotreroRincoÂn 3 is possibly a new taxon, but its systematic position remains uncertain. It may be a primitivebrachythoracid, but some of its features also recall groenlandaspidids. Therefore, it cannot be regarded asstratigraphically informative. Nevertheless, the arthrodires range from the Lochkovian (Eifelian forbrachythoracids) to Late Famennian.
The chondrichthyan Antarctilamna is known from the Givetian of Australia, but occurs in rocks as oldas Late Emsian in Iran and Saudi Arabia. The Bolivian records (Gagnier et al. 1988, 1989) possibly rangefrom the Emsian up to the Frasnian.
The acanthodian scales from Potrero RincoÂn 1 are rather undiagnostic, although their crownmorphology is suggestive of the climatiiform Cheiracanthoides, recorded from the Lochkovian±Frasnian(De Pomroy 1996). However, similarly ribbed scales occur in several Late Devonian forms, such as theacanthodid Homalacanthus. The absence of data regarding their histology makes them indeterminable at amore precise level.
The actinopterygian remains are more similar to Mimia than to any other actinopterygian genus.However, this rests largely on shared primitive actinopteran characters. In terms of evolutionary grade,however signi®cant this may or may not be, the occurrence of this Colombian form is consistent with aFrasnian age.
Among sarcopterygians, Holoptychius is known from the Frasnian to the latest Famennian, butTournaisian records are doubtful. Holoptychius has formerly been recorded from the Givetian, but thisis based on material now referred to Glyptolepis. The cosmine-covered osteolepidid is too poorly known toprovide any useful information. Cosmine-covered osteolepidids become rare after the Frasnian, butnevertheless survive until the Early Permian. The rhizodontid questionably referred here to Strepsodus isperhaps the most informative taxon, since rhizodontids are known only from the Famennian to the MiddleCarboniferous (Bashkirian). Rhizodontids occur relatively frequently in the Famennian, and the earliestrecord of this group is thought to be Sauripteris taylori, from the Late Frasnian? or Early FamennianCatskill delta facies of New York State (Schultze 1993). An Early Frasnian age has been suggested forNotorhizodon, from the Pambulaspis Zone of the Aztec Siltstone of Antarctica (Young et al. 1992), butthis remains uncertain (Young 1993b). Strepsodus is essentially Carboniferous, but also occurs in the latestFamennian, in association with placoderms and cosmine-covered dipnoans, in the locality of Cukurca,south-eastern Turkey (Janvier et al. 1984).
Notwithstanding the fact that the faunal list given here is provisional, and that additional material, inparticular cranial material, is necessary to reach a more precise level of speci®c determination, the age-rangeshared by taxa that can be identi®ed at the familial or generic level is latest Frasnian or earliest Famennian.
754 P A L A E O N T O L O G Y , V O L U M E 4 3
E X P L A N A T I O N O F P L A T E 5
Figs 1±8. Strepsodus? sp. indet. Cuche Formation, Upper Devonian, Boyaca Department, Colombia; elastomer casts,except for ®gure 1. 1±2, UN-DG-PALV45; incomplete right lower jaw, natural cast of the external surface (1) andelastomer cast of the internal surface, showing the symphysial region (2); Potrero RincoÂn 2. 3, UN-DG-PALV46;scale in internal view; Potrero RincoÂn 2. 4, UN-DG-PALV47; scale in internal view; Potrero RincoÂn 2. 5±6, UN-DG-PALV60-61; scales in internal view, showing internal, scattered tubercles (5), and branching, lateral-line canals(6); roadcut along the Santa Rosa de Viterbo-Floresta road. 7, UN-DG-PALV48; scale in external view, PoteroRincoÂn 2. 8, UN-DG-PALV49; cleithrum in internal view, Potrero RincoÂn 2.
Figs 9±10. Holoptychius sp. indet.; Cuche Formation, Upper Devonian, Potrero RincoÂn 3, Boyaca Department,Colombia; UN-DG-PALV50-51; scales in external view. All ´ 2.

P L A T E 5
JANVIER and VILLARROEL, Strepsodus?, Holoptychius

It is possible that the ®sh assemblage brie¯y recorded by Young (1993a) from the Devonian of theMaracaibo area in Venezuela (presently being studied by G. Young and J. Moody) will turn out to beclosely similar to that of the Cuche Formation. It is known to include chondrichthyans, acanthodians,bothriolepidid antiarchs, and osteolepidid sarcopterygians (Young 1993a). These Venezuelan localitiesalso yield a rich Late Devonian ¯ora (Berry et al. 1993; Young 1993a; Berry 1994; Berry and Edwards1996) that will have to be compared to that of the Cuche Formation, where the presence of Archaeopterisin the ®sh-bearing outcrops (C. Berry, pers. comm. 1997) is strongly suggestive of a Late Devonian age.Berry and Edwards (1996) also mentioned the presence of Archaeopteris in the upper member of theCampo Chico Formation of Venezuela, where most of the ®shes also occur (Young 1993a).
Palaeoenvironment
Some of the ®sh-bearing levels of the Cuche Formation, in particular Potrero RincoÂn 2 and Tunguaquita,contain a typical assemblage that is found in the Late Devonian localities of `Old Red Sandstone' facies ineastern North America (e.g. Catskills), Greenland, Scotland and the Baltic states. In the Potrero RincoÂnsection, most of the Cuche Formation was deposited in a subaerial environment, as evidenced by numerousimprints of roots and other plant remains in the basal part and the lack of any marine organisms. Thesediments deposited under marine in¯uence can be easily distinguished from the continental deposits bytheir yellowish, instead of reddish colour, and assemblages of conchostracans, ostracods (Walleria? sp.),bivalves, lingulids, and occasional plant remains. It is clear that the ®sh-bearing levels accumulated inbrackish±marine environments, as shown by the presence of small inarticulate brachiopods (lingulids).The deposits of Potrero RincoÂn 1 show the strongest marine in¯uence, the presence of a Mimia-likeactinopterygian, known elsewhere from typically marine Devonian successions. The chondrichthyanAntarctilamna, which also occurs at Potrero RincoÂn 1, is recorded from both marine and supposedly freshwater deposits. Moreover, the accumulation of fragmented placoderm plates in Potrero RincoÂn 1 and 3 issuggestive of reworking by waves. By contrast, the presence of partly articulated armour of Bothriolepis inPotrero RincoÂn 2 suggests a low energy environment of deposition. The Cuche Formation can thus beregarded as a marginal facies of a delta and lagoons with frequent, but temporary, invasions of sea water.The depth of deposition of such ®sh assemblages was probably no more than 10 metres. Whether therhizodontid from this assemblage was a marine or freshwater form is unclear. Most Carboniferousrhizodontids are regarded as freshwater indicators and are probably the largest freshwater ®shes (up to 7 m)that have ever lived (Jeffery 1997).
It is possible that the ®sh bone-beds of the Cuche Formation represent mass mortalities of marine ®shes,killed by sudden in¯uxes of fresh water arriving from the continent. It is worth noting that the small(juvenile) inarticulate brachiopods are only found in the ®sh-bearing levels, and it is known that recentlingulids are among the few marine invertebrates that can tolerate signi®cant variations in salinity (Emig1981; Goujet and Emig 1985). Fishes exhibit a similar degree of tolerance. The absence of other marineinvertebrates from these levels also suggests an environment that experienced strong variations in salinity.
Palaeobiogeography
At this stage of the investigation, the most intriguing feature of this assemblage from Colombia is itsstriking resemblance to assemblages from Late Devonian (in particular Late Frasnian) localities of Europeand North America. Among antiarchs, Bothriolepis is admittedly a widespread genus, now known from allthe major Devonian continental blocks, but Asterolepis is unknown outside Euramerica, except for asingle, doubtful record from the Frasnian of Iran (Janvier 1978b). Other asterolepidid antiarchs, however,are known from China and Australia. The two Bothriolepis species from Colombia do not belong to thegroup de®ned by a trilobate preorbital recess, which is considered as endemic to Gondwana (Young 1988).By contrast, Antarctilamna is not recorded with certainty outside Gondwana, and can be regarded as theonly reliable Gondwanan endemic from this assemblage. Holoptychius is very rare outside Euramerica,but Young (1993b) recorded Holoptychius remains from the Famennian Jemalong-Canowindra andWorange Point faunas in Australia. Rhizodontids are unknown outside of Euramerica in rocks older than
756 P A L A E O N T O L O G Y , V O L U M E 4 3

latest Famennian (with the possible exception of Notorhizodon; Young et al. 1992), but they are generallyrare and poorly preserved ®shes, and this may re¯ect sampling biases.
Although Antarctilamna is a good Gondwanan `signature' for this assemblage, its overall resemblanceto the Late Frasnian faunas of Euramerica deserves an explanation. It is worth noting that antiarchs aregenerally found in marginal and even possibly freshwater facies, and often display very clear patterns ofendemism (Young 1993c). Moreover, antiarchs were probably poor swimmers, their dispersal requiringvery shallow waters. In addition, rhizodontid sarcopterygians are also generally found in marginal depositsand became almost exclusively restricted to fresh water in the Carboniferous. These observations suggest aclose geographical relationship between western Gondwana and Euramerica in Late Frasnian times;alternatively occurrences in the northern part of South America may be allochthonous and derived fromNorth America, but there is no clear evidence for this model to date.
This Colombian vertebrate assemblage is, by contrast, quite different from that described from theMiddle and Upper Devonian of Bolivia (Janvier 1992), the only common taxon being Antarctilamna. Thisdisparity may admittedly be a result of facies differences, since the Devonian vertebrate faunas of Boliviaclearly re¯ect deeper, marine environments than the Colombian assemblage. However, the Middle±UpperDevonian of the Bolivian Altiplano (BeleÂn, Sica Sica and Colpacucho formations) also includes somesiliciclastic, shallow water members with plant remains and inarticulate brachiopods, that have yet to yieldany antiarchs or sarcopterygian remains. Only a large, marine dinichthyid arthrodire has been found inthese facies in the Colpacucho Formation (Diaz-Martinez et al. 1996). So far, it is unclear whether thedifference between the Bolivian and Colombian assemblages is a re¯ection of geographical isolation orclimate; Bolivia was farther to the south and, thus, presumably cooler than Colombia in Devonian times.
The distribution of the continent-bound (i.e. related to marginal marine and freshwater environments)Devonian vertebrate assemblages indicates well-marked patterns of endemism in the Early Devonian, andan increasing homogeneity towards the Late Devonian. Only the eastern Asian areas (correspondingessentially to the South and North China blocks, and the Indochina Block) seem to retain largely endemictaxa until the Givetian±Late Devonian (Young 1993c; Young and Janvier 1999). This increasinghomogeneity of the vertebrate assemblages is interpreted as the result of extensive interchanges betweenpreviously isolated major continental units during the formation of Pangaea, but how and under exactlywhat conditions these interchanges occurred remains uncertain. It may be an effect of the increasinglyimportant Middle±Late Devonian marine transgression, but this explanation hardly holds for manyplacoderm groups, such as antiarchs. Another explanation is that Euramerica, Siberia, Kazakhstan and theChinese-Indochinese blocks came into closer contact with Gondwana along the tropical belt. It is nowquite clear that the Chinese-Indochinese blocks had continental links with eastern Gondwana by LateDevonian times, as shown by the presence of an endemic Chinese antiarch group, the Sinolepidae, in theFamennian of Australia (Ritchie et al. 1992; Young 1993c), but there is no evidence that the numerousMiddle±Late Devonian taxa shared by eastern Gondwana and Euramerica�Siberia dispersed through theChinese-Indochinese blocks. Only the Kazakhstan block (however isolated it may have been earlier),seems to have had some contact with the Chinese-Indochinese blocks (Young 1993c). Young (1990) andLelieÁvre et al. (1993) suggested that the Gondwana-Euramerica contact may have occurred at the level ofthe Arabian platform, which might have been extended northward by a number of microplates (Turkey andIran). This assumption was based on the presence of a somewhat mixed Euramerican and Gondwananfauna in the Frasnian of this area (LelieÁvre et al. 1993). Another possibility is that it occurred through aclose contact between south-western Euramerica and western Gondwana: the Venezuelian-Colombianvertebrate faunas would provide such a biogeographic link. This would imply a much closer position ofEuramerica to Gondwana than in the palaeomagnetism-based world maps proposed by, for example.Scotese and McKerrow (1990, ®g. 16) and Kent and Van der Voo (1990, ®g. 6) for the Late Devonian. Infact, the hypothesis of a Late Devonian Gondwana-Euramerica interchange through northern SouthAmerica would agree most closely with Kent and Van der Voo's (1990, ®gs 4±5) model for the LateSilurian and Early Devonian, where eastern North America is adjacent to north-western South America.
In sum, among the forms that could be identi®ed at the generic level in the Cuche Formation, only one,the antiarch Bothriolepis, has a widespread Late Devonian distribution. The others are endemic to, orparticularly abundant in either Gondwana (Antarctilamna?, the Mimia-like actinopterygian) or Euramerica
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 757

(Asterolepis?, Strepsodus?, Holoptychius). This mix of Gondwanan and Euramerican forms, which areessentially related to marginal marine±deltaic environments, supports close biogeographic links betweennorthern South America and Euramerica in Devonian times, as previously suggested on the basis of marineinvertebrates (Boucot and Racheboeuf 1993).
C O N C L U S I O N S
The Devonian vertebrate remains recovered from the Floresta Massif occur in rocks dated as Emsian±Eifelian and Late Devonian, presumably Late Frasnian or Early Famennian. The Emsian±Eifelianoccurrence at the base of the Floresta Formation is not particularly informative, although it does provideevidence for placoderms (probably arthrodires and rhenanids). The Late Devonian occurrence, from theCuche Formation, yields a remarkably diverse assemblage of chondrichthyans, acanthodians, placoderms(antiarchs and arthrodires), actinopterygians, and sarcopterygians. Except for the still undeterminedarthrodire, which may be either a primitive brachythoracid or a groenlandaspidid, all the vertebrates of thisassemblage are either closely related to, or belong to, genera known from other continents. However, inmost cases (antiarchs, sarcopterygians), this is the ®rst record of their occurrence in South America.
Besides the presence of some widespread taxa, such as Bothriolepis, the ®sh assemblage from the CucheFormation includes a mix of taxa that are reputedly endemic to Gondwana and taxa that are either endemicto, or more abundant in, Euramerica. This suggests close biogeographic links between western Gondwanaand Euramerica in Late Devonian times.
Acknowledgements. We are grateful to the Universidad Nacional de Colombia (BogotaÂ), the Institut FrancËais d'EtudesAndines (Lima), and the MuseÂum National d'Histoire Naturelle (Paris) for their ®nancial support. The photographswere prepared by D. Serrette (Paris). Jon Jeffery (Cambridge) is warmly thanked for his advice on the rhizodontidmaterial and references. We are most grateful to Christopher Berry (Cardiff) for having called our attention to the ®rst®sh-bearing sample from Potrero RincoÂn (UN-DG-PALV40, initially labelled as `cche-III-84-002'). The anonymousreviewers and Gavin C. Young (Canberra) are gratefully acknowledged for their helpful comments.
R E F E R E N C E S
AGASSIZ, J. L. R. 1839. Fishes of the Upper Ludlow Rock. 605±607. In MURCHISON, R. I. (ed.). The Silurian System,Volume 2. John Murray, London, 576 pp.
ALBANESI, G. L., BENEDETTO, J. L. and GAGNIER, P.-Y. 1995. Sacabambaspis janvieri (Vertebrata) y conodontes delLlandeiliano temprano en la Formacion La Cantera, Precordillera de San Juan, Argentina. Boletin de la Academia deCiencias, Cordova, 60, 519±543.
ANDREWS, S. M. 1985. Rhizodont crossopterygian ®sh from the Dinantian of Foulden, Berwickshire, Scotland, with are-evaluation of this group. Transactions of the Royal Society of Edinburgh, 76, 67±95.
ÐÐ and WESTOLL, T. S. 1970. The postcranial skeleton of rhipidistian ®shes, excluding Eusthenopteron. Transactionsof the Royal Society of Edinburgh, 68, 391±489.
BARRETT, S. F. 1985. Paleoecology and stratigraphy of Devonian sediments in the northern Andes, Colombia:paleogeographic implications. Unpublished PhD thesis, Department of Geophysical Sciences, University ofChicago.
ÐÐ 1988. The Devonian system in Colombia. 705±717. In MCMILLAN, N. J., EMBRY, A. F. and GLASS, D. J. (eds).Devonian of the world. Canadian Society of Petroleum Geologists, Regional Syntheses, Volume 1, Calgary,795 pp.
BERG, L. S. 1937. A classi®cation of ®sh-like vertebrates. Bulletin de l'AcadeÂmie des Sciences de l'URSS, Classe deMatheÂmatiques et de Sciences Naturelles, SeÂrie Biologie, 1937, 1277±1280. [In Russian and English].
ÐÐ 1940. Classi®cation of ®shes, both Recent and fossil. Trudy Zoologicheskogo Instituta, Akademya Nauk SSSR,Leningrad, 5, 87±517.
BERRY, C. M. 1994. First record of the lycophyte Leclecqia from South America. Geological Magazine, 131, 269±272.ÐÐ and EDWARDS, D. 1996. The herbaceous lycophyte Haskinsia Grierson and Banks from the Devonian of western
Venezuela, with observations on leaf morphology and fertile specimens. Botanical Journal of the Linnean Society,122, 103±122.
ÐÐ CASAS, H. E. and MOODY, J. M. 1993. Diverse Devonian plant assemblages from Venezuela. Documents duLaboratoire de GeÂologie de Lyon, 125, 29±42.
758 P A L A E O N T O L O G Y , V O L U M E 4 3

BLIECK, A., GOLSHANI, F., GOUJET, D., HAMDI, A., JANVIER, P., MARK-KURIK, E. and MARTIN, M. 1980. A new vertebrate localityin the Eifelian of the Khush-Yeilagh Formation, eastern Alborz, Iran. Palaeovertebrata, 9, 133±154.
ÐÐ GAGNIER, P.-Y., BIGEY, F. P., EDGECOMBE, G. D., JANVIER, P., LOBOZIAK, S., RACHEBOEUF, P. R., SEMPEÂ REÂ , T. and STEEMANS,
P. 1996. New Devonian fossil localities in Bolivia. Journal of South American Earth Sciences, 9, 295±308.BOINET, T., BABIN, C., BOURGOIS, J., BROUTIN, J., LARDEUX, H., PONS, D. and RACHEBOEUF, P. 1986. Les grandes eÂtapes de
l'eÂvolution paleÂozoõÈque du Massif de Santander (Andes de Colombie): signi®cation de la discordance du DeÂvonienmoyen. Compte Rendus de l'AcadeÂmie des Sciences, Paris, 303, 707±712.
BONAPARTE, C. L. 1838. Synopsis vertebratorum systematis. Nuovi Annali delle Scienze Naturali, Bologna, 11, 105±133.
BOTERO-RESTREPO, G. 1950. Reconocimiento geologico del area comprendida por los Municipios de BeleÂn, Cerinza,Corrales, Floresta, Nobsa y Santa Rosa de Viterbo (Departamento de BoyacaÂ). Compilacion de los EstudiosGeologicos O®ciales en Colombia. Ministerio de Minas y Petroleos, Servicio Geologico Nacional, 8, 245±311.
BOUCOT, A. J. and RACHEBOEUF, P. 1993. Biogeographic summary of the Malvinokaffric Realm Silurian and Devonianfossils. 71±75. In SUAREZ-SORUCO, R. (ed.). Fosiles y facies de Bolivia. II, Invertebrados y Paleobotanica. RevistaTecnica de Yacimientos Petroliferos Fiscales Bolivianos, Santa Cruz, 13±14, 1±193.
BUÈ RGL, H. 1961. Historia geologica de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Fisicas yNaturales, 11 (43), 137±191.
BYSTROV, A. P. 1957. The microstructure of dermal bones in arthrodires. Acta Zoologica, Stockholm, 38, 9±275.CARRILLO, F. 1992. Estudio estratigra®co y sedimentologica de la Formacion Cuche en la vereda Horno y Vivas
(Macizo de Floresta, Depto de BoyacaÂ). Unpublished PhD thesis, Universidad Nacional de Colombia, Facultad deCiencias, Departamento de Geociencias, BogotaÂ.
CASTER, K. E. 1939. A Devonian fauna from Colombia. Bulletins of American Paleontology, 24, 101±318.CEDIEL, F. 1969. Geologia del Macizo de Floresta. Memoria, Primer Congresso Colombiano de Geologia, 17±29.ÐÐ 1972. Movimientos tectonicos en el intervalo Paleozoico-Mesozoico en Colombia y su in¯uencia en
reconstrucciones paleogeogra®cas. Simposio Internacional sobre os Sistemas Carbonifero e Permiano na AmeÂricado Sul. Anais da Academia Brasileira de Ciencias (Supplemento), 44, 87±93.
CLOUTIER, R. and SCHULTZE, H.-P. 1996. Porolepiform ®shes (Sarcopterygii). 248±270. In SCHULTZE, H.-P. and CLOUTIER,
R. (eds). Devonian ®shes and plants of Miguasha, Quebec, Canada. Pfeil, Munich, 374 pp.COPE, E. D. 1885. The position of Pterichthys in the system. American Naturalist, 9, 1±9.ÐÐ 1886. An interesting connecting genus of Chordata. American Naturalist, 20, 1027±1031.ÐÐ 1887. Zittel's manual of palaeontology. American Naturalist, 21, 1014±1019.ÐÐ 1889. Synopsis of the families of Vertebrata. American Naturalist, 23, 849±877.CUEVAS, R. D. and SANCHEZ, J. 1991. Analisis estratigra®co y facial de una seccion de la Formacion Floresta a lo largo del
rio Pargua entre los Municipios de BeleÂn, Tutaza y Paz del Rio, en el Departamento de BoyacaÂ. Unpublished PhDthesis, Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Geociencias, BogotaÂ.
DE POMROY, A. M. 1996. Biostratigraphy of Devonian microvertebrates from Broken River, North Queensland. Recordsof the Western Australian Museum, 17, 417±437.
DINELEY, D. L. and LOEFFLER, E. J. 1993. Biostratigraphy of the Silurian and Devonian gnathostomes of the EuramericanProvince. 104±138. In LONG, J. A. (ed.). Palaeozoic vertebrate biostratigraphy and biogeography. Belhaven Press,London, 369 pp.
DIAZ-MARTINEZ, E., SUAREZ-RIGLOS, M., LELIEÁVRE, H. and JANVIER, P. 1996. PremieÁre deÂcouverte d'Arthrodires (Vertebrata,Placodermi) dans le DeÂvonien d'AmeÂrique du Sud. Compte Rendus de l'AcadeÂmie des Sciences, Paris, 323, 349±356.
EICHWALD, E. I. VON 1840. Die Thier- und P¯anzenreste des alten rothen Sandsteins und Bergkalks in NovgorodschenGouvernement. Bulletin de l'AcadeÂmie ImpeÂriale des Sciences de Saint-Petersbourg, 7, 78±91.
EMIG, C. C. 1981. Implications de donneÂes reÂcentes sur les Lingules actuelles dans les interpreÂtations paleÂoeÂcologiques.Lethaia, 14, 151±156.
ETAYO SERNA, F. and 20 others 1986. Mapa de terrenos geologicos de Colombia. Publicaciones Geologicas Especialesdel Ingeominas, 14, 1±235.
GAGNIER, P.-Y. 1992. Ordovician vertebrates from Bolivia. 371±379. In SUAREZ-SORUCO, R. (ed.). Fosiles y facies deBolivia. I. Vertebrados. Revista Tecnica de Yacimientos Petroliferos Fiscales Bolivianos, Santa Cruz, 12, 359±718.
ÐÐ 1993a. Sacabambaspis janvieri, VerteÂbre ordovicien de Bolivie. 1, Analyse morphologique. Annales dePaleÂontologie (VerteÂbreÂs-InverteÂbreÂs), 79, 19±69.
ÐÐ 1993b. Sacabambaspis janvieri, VerteÂbre ordovicien de Bolivie. 2. Analyse phylogeÂneÂtique. Annales dePaleÂontologie (VerteÂbreÂs-InverteÂbreÂs), 79, 119±166.
ÐÐ and BLIECK, A. 1992. On Sacabambaspis janvieri and the vertebrate diversity in Ordovician seas. 9±20. In MARK-
KURIK, E. (ed.). Fossil ®shes as living animals. Academia, 1, 1±299.ÐÐ BLIECK, A. and RODRIGO, G. 1986. First Ordovician vertebrate from South America. Geobios, 19, 629±634.
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 759

ÐÐ TURNER, S., FRIMAN, L., SUAREZ-RIGLOS, M. and JANVIER, P. 1988. The Devonian vertebrate and mollusc fauna fromSeripona (Chuquisaca, Bolivia). Neues Jahrbuch fuÈr Geologie und PalaÈontologie, Abhandlungen, 176, 269±297.
ÐÐ PARIS, F., RACHEBOEUF, P., JANVIER, P. and SUAREZ-RIGLOS, M. 1989. Les VerteÂbreÂs deÂvoniens de Bolivie: donneÂesbiostratigraphiques et anatomiques compleÂmentaires. Bulletin de l'Institut FrancËais d'Etudes Andines, Lima, 18,75±93.
GARDINER, B. G. 1963. Certain palaeoniscid ®shes, a review based on new specimens of Mimia and Moythomasia fromthe Upper Devonian of Western Australia. Bulletin of the British Museum (Natural History), Geology Series, 8,255±325.
ÐÐ 1984. The relationships of the palaeoniscid ®shes, a review based on new specimens of Mimia and Moythomasiafrom the Upper Devonian of Western Australia. Bulletin of the British Museum (Natural History), Geology Series,37, 173±428.
ÐÐ and SCHAEFFER, B., 1989. Interrelationships of lower actinopterygian ®shes. Zoological Journal of the LinneanSociety, 97, 135±187.
GLIKMAN, L. S. 1964. [Subclass Elasmobranchii (sharks).] 292±352. In OBRUCHEV, D. V. (ed.) [Fundamentals ofpaleontology, 11. Agnatha, Pisces.] Nauka, Moscow. [In Russian; English translation, 1967, Israel program forscienti®c translations].
GOUJET, D. 1984. Les poissons Placodermes du Spitsberg. Arthrodires Dolichothoraci de la Formation de Wood Bay(DeÂvonien inferieur). Cahiers de PaleÂontologie, Centre National de la Recherche Scienti®que, Paris, 284 pp.
ÐÐ and EMIG, C. 1985. Des Lingula fossiles, indicateurs de modi®cations de l'environnement dans un gisement duDeÂvonien infeÂrieur du Spitsberg. Compte Rendus de l'AcadeÂmie des Sciences, Paris, Series 2, 301, 945±948.
ÐÐ JANVIER, P. and RACHEBOEUF, P. 1993. Placoderm and chondrichthyan remains from the Middle Devonian ofChacoma, La Paz area, Bolivia. Neues Jahrbuch fuÈr Geologie und PalaÈontologie, Monatshefte, 1993, 355±372.
ÐÐ ÐÐ and SUAREZ-RIGLOS, M. 1984. Devonian vertebrates of South America. Nature, 312, 311.ÐÐ ÐÐ ÐÐ 1985. Un nouveau rheÂnanide (Vertebrata, Placodermi) de la Formation de BeleÂn (DeÂvonien moyen),
Bolivie. Annales de PaleÂontologie (VerteÂbreÂs-InverteÂbreÂs), 71, 35±53.GRAHAM-SMITH, W. 1978. On some variations on the latero-sensory lines of the placoderm ®sh Bothriolepis.
Philosophical Transactions of the Royal Society of London, Series B, 282, 1±39.GROSS, W. 1973. Kleinschuppen, Flossenstacheln und ZaÈhne von Fischen aus europaÈischen und nordamerikanischen
Bonebeds des Devons. Palaeontographica, Abteilung A, 142, 151±155.GROÈ SSER, J. R. and PROÈ SSL, K. F. 1994. Palynologische Untersuchungen der Devonbasis in Floresta Massiv, Ostkordillere,
Kolumbien, S.A. Giessener Geologische Schriften, 51, 105±121.HEINTZ, A. 1934. A revision of the Estonian Arthrodira. Part 1, Family Homosteidae. Arkhiv fuÈr die Naturkunde
Estlands, 10 (4), 1±114.HUXLEY, T. H. 1880. On the application of the laws of evolution to the arrangement of the Vertebrata and more
particularly of the Mammalia. Proceedings of the Zoological Society of London, 1880, 649±662.ÐÐ and ETHERIDGE, R. 1865. A catalogue to the collections of fossils in the Museum of Practical Geology, with an
explanatory introduction. United Kingdom Geological Survey, British Regional Geology Handbooks, G. E. Eyre &W. Spottiswoode for HMSO, London, 381 pp.
JANVIER, P. 1976. Description de restes d'eÂlasmobranches (Pisces) du DeÂvonien moyen de Bolivie. Palaeovertebrata, 7,126±132.
ÐÐ 1978a. Los peces devonicos de Bolivia. Programma internacional de correlacion geologica (n8 42, 44, 120).Academia Nacional de Ciencias, La Paz, 13 pp.
ÐÐ 1978b. Les VerteÂbreÂs deÂvoniens de l'Iran Central. 3: Antiarches. Geobios, 12, 718±720.ÐÐ 1987. Les VerteÂbreÂs siluriens et deÂvoniens de Bolivie: remarques particulieÁres sur les Chondrichthyens. 4e
Congresso Latinoamericano de Paleontologia, Santa Cruz, A, 159±178.ÐÐ 1992. Silurian and Devonian vertebrates. Permian and Triassic vertebrates. 381±392. In SUAREZ-SORUCO, R. (ed.).
Fosiles y facies de Bolivia. I. Vertebrados. Revista Tecnica de Yacimientos Petroliferos Fiscales Bolivianos, SantaCruz, 12, 359±718.
ÐÐ 1993. Os peixes mais antiguos do Brasil. Ciencia Hoje, Rio de Janeiro, 15, 110.ÐÐ and DINGERKUS, G. 1991. Le synarcual de Pucapampella Janvier & Suarez-Riglos: une preuve de l'existence
d'HoloceÂphales deÁs le DeÂvonien moyen. Compte Rendus de l'AcadeÂmie des Sciences, Paris, 312, 549±552.ÐÐ LETHIERS, F., MONOD, O. and BALKAS, OÈ . 1984. Discovery of a vertebrate fauna at the Devonian-Carboniferous
boundary in S.E. Turkey (Hakkari Province). Journal of Petroleum Geology, 7, 147±168.ÐÐ and MELO, J. H. G. 1987. Late Devonian actinopterygian scales from the Amazon Basin, northwestern Brazil.
Annais da Academia Brasileira de CieÃncias, 59, 213±218.ÐÐ ÐÐ 1988. Acanthodian ®sh remains from the Upper Silurian or Lower Devonian of the Amazon Basin, Brazil.
Palaeontology, 31, 771±777.
760 P A L A E O N T O L O G Y , V O L U M E 4 3

ÐÐ ÐÐ 1992. New acanthodian and chondrichthyan remains from the Lower and Middle Devonian of Brazil.Neues Jahrbuch fuÈr Geologie und PalaÈontologie, Monatshefte, 1992, 193±206.
ÐÐ and SUAREZ-RIGLOS, M. 1986. The Silurian and Devonian vertebrates of Bolivia. Bulletin de l'Institut FrancËaisd'Etudes Andines, Lima, 15, 73±114.
ÐÐ ÐÐ 1989. Los primeros Vertebrados devonicos de Sud America. Revista Tecnica de Yacimientos PetroliferosFiscales Bolivianos, Santa Cruz, 10, 229±231.
JARVIK, E. 1942. On the structure of the snout of crossopterygians and lower gnathostomes in general. ZoologiskaBidrag, Uppsala, 21, 235±675.
ÐÐ 1948. On the morphology and taxonomy of the Middle Devonian osteolepid ®shes of Scotland. KungligaSvenska Vetenskapsakademiens Handlingar, 25, 1±301.
ÐÐ 1980. Basic structure and evolution of vertebrates. Volume 1. Academic Press, London, 575 pp.JEFFERY, J. E. 1997. Rhizodontiformes; giant, predatory sarcopterygian ®shes from the Upper Devonian and Lower
Carboniferous. Journal of Vertebrate Paleontology, 17 (Supplement 3), 55A.JESSEN, H. L. 1968. Moythomasia nitida Gross und M. cf. striata, devonische Palaeonisciden aus dem Oberen
Plattenkalk der Bergisch-Gladbach-Paffrather Mulde (Rheinisches Schiefergebirge). Palaeontographica, AbteilungA, 128, 87±114.
ÐÐ 1972. Die Bauchschuppen von Moythomasia nitida Gross (Pisces, Actinopterygii. PalaÈontologische Zeitschrift,46, 121±132.
JOHANSON, Z. and AHLBERG, P. E. 1998. A complete primitive rhizodont from Australia. Nature, 394, 569±573.KARATAJUTE-TALIMAA, V. I. 1963. [The genus Asterolepis from the Devonian of the Russian Platform.] Leituvos TSR
Mokslu Akademija, Geologijos ir Geogra®jos Institutas, Vilnius, 1963, 65±223. [In Russian].KENT, D. V. and VAN DER VOO, R. (1990). Palaeozoic palaeogeography from palaeomagnetism of the Atlantic-bordering
continents. 49±56. In MCKERROW, W. S. and SCOTESE, C. R. (eds). Palaeozoic palaeogeography and biogeography.Memoir of the Geological Society, London, 12, 1±435.
KLEIN, E. F. 1885. BeitraÈge zur Bildung des SchaÈdels der Knochen®sche, 2. Jahreshefte Vereins VaterlandischerNaturkunde in WuÈrtenberg, 42, 205±300.
LELIEÁ VRE, H. 1984. Antineosteus lehmani, n. g., n. sp., nouveau Brachythoraci du DeÂvonien infeÂrieur du MarocpreÂsaharien. Annales de PaleÂontologie (VerteÂbreÂs), 70, 115±158.
ÐÐ JANJOU, D., HALAWANI, M., JANVIER, P., MUAMEL, M. S. A., WYNNS, R. and ROBELIN, C. 1994. Nouveaux VerteÂbreÂs(Placodermes, Acanthodiens, Chondrichthyens et SarcopteÂrygiens) de la Formation de Jauf (DeÂvonien infeÂrieur,reÂgion de Al Huj, Arabie Saoudite). Compte Rendus de l'AcadeÂmie des Sciences, Paris, 319, 1247±1254.
ÐÐ JANVIER, P. and BLIECK, A. 1993. Silurian-Devonian vertebrate biostratigraphy of Western Gondwana and relatedterranes (South America, Africa, Armorica±Bohemia, Middle-East). 139±173. In LONG, J. A. (ed.). Palaeozoicvertebrate biostratigraphy and biogeography. Belhaven Press, London, 369 pp.
LONG, J. A. 1983. New bothriolepid ®sh from the Late Devonian of Victoria, Australia. Palaeontology, 26, 295±320.ÐÐ 1988. New palaeoniscoid ®shes from the Late Devonian and Early Carboniferous of Victoria. Memoir of the
Association of Australasian Palaeontologists, 7, 1±64.ÐÐ 1989. A new rhizodontiform ®sh from the Early Carboniferous of Victoria, Australia, with remarks on the
phylogenetic position of the group. Journal of Vertebrate Paleontology, 9, 1±17.ÐÐ 1992. Cranial anatomy of two new Late Devonian lung®shes (Pisces: Dipnoi) from Mount Howitt, Victoria.
Records of the Australian Museum, 44, 299±318.ÐÐ and WERDELIN, L. 1986. A new Late Devonian bothriolepid (Placodermi, Antiarchi) from Victoria, with
descriptions of other species from the state. Alcheringa, 10, 355±399.ÐÐ and YOUNG, G. C. 1995. Sharks from the Middle±Late Devonian Aztec Siltstone, southern Victoria Land,
Antarctica. Records of the Western Australian Museum, 17, 287±308.LUKSEVICS, E. 1995. Variability in bothriolepid antiarchs (Placodermi) from the Main Devonian Field (East European
Platform). 117±120. In LELIEÁ VRE, H., WENZ, S., BLIECK, A. and CLOUTIER, R. (eds). Premiers VerteÂbreÂs et VerteÂbreÂsInfeÂrieurs. Geobios, MeÂmoire SpeÂcial, 19, 1±409.
MCCOY, F. 1848. On some new fossil ®shes of the Carboniferous period. Annals and Magazine of Natural History,2, 1±10.
MCNAIR, A. H. 1940. Devonian bryozoa from Colombia. Bulletins of American Paleontology, 25 (93), 113±146.MILES, R. S. 1971. The Holonematidae (placoderm ®shes), a review based on new specimens of Holonema from the
Upper Devonian of Western Australia. Philosophical Transactions of the Royal Society of London, Series B, 263,191±234.
MOJICA, J. and VILLARROEL, C. 1984. Contribucion al conicimiento de las unidades paleozoicas del area de Floresta(Cordillera oriental colombiana; Departamento de BoyacaÂ) y en especial al de la Formacion Cuche. GeologiaColombiana, 13, 55±79.
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 761

MORALES, P. A. 1965. A contribution to the knowledge of the Devonian faunas of Colombia. Boletin de Geologia, 19,51±111.
OTTO, M. 1992. Ein Neufund der brachythoracen Arthrodiren Tityosteus rieversi aus dem unterdevonischenHunsruÈckschiefer des rheinischen Schiefergebirges. Neues Jahrbuch fuÈr Geologie und PalaÈontologie,Abhandlungen, 187, 53±82.
OWEN, R. 1846. Lectures on the comparative anatomy and physiology of the vertebrate animals, delivered at the RoyalCollege of Surgeons of England. 1, Fishes. Longman, Brown, Green and Longmans, London, 308 pp.
ÐÐ 1860. Palaeontology, or a systematic summary of extinct animals and their geological relations. Black,Edinburgh, 420 pp.
PAZ, M. G. and URRUTIA, A. D. 1996. Estratigra®a y ambientes de deposito de la Formacion El Tibet (Macizo de Floresta,BoyacaÂ). Unpublished PhD thesis, Universidad Nacional de Colombia, Facultad de Ciencias, Departamento deGeociencias, BogotaÂ.
PEARSON, D. M. 1982. Primitive bony ®shes, with especial reference to Cheirolepis and palaeonisciform actino-pterygians. Zoological Journal of the Linnean Society, London, 74, 35±67.
ÐÐ and WESTOLL, T. S. 1979. The Devonian actinopterygian Cheirolepis Agassiz. Transactions of the Royal Society ofEdinburgh, 70, 337±399.
RITCHIE, A., WANG, S., YOUNG, G. C. and ZANG, G. 1992. The Sinolepidae, a family of antiarchs (placoderm ®shes) from theDevonian of South China and eastern Australia. Records of the Australian Museum, 44, 319±370.
ROMER, A. S. 1955. Herpetichthyes, Amphibioidei or Sarcopterygii? Nature, 176, 126.ROYO Y GOMEZ, J. 1942. Fosiles devonicos de Floresta (Departamento de BoyacaÂ). Compilacion de los Estudios
Geologicos O®ciales en Colombia. Ministerio de Minas y Petroleos, Servicio Geologico Nacional, Bogota , 5, 389±396.
SCHMIDT, W. E. 1938. Fossiles paleozoicos de la Cordillera Oriental de Colombia (Cordillera de BogotaÂ). 1±15. InEstudios geologicos y paleontologicos sobre la Cordillera Oriental de Colombia, Parte Secunda. Ministerio desIndustrias y Trabajo, Departamento de Minas y Petroleo, BogotaÂ, 80 pp.
SCHULTZE, H. P. 1992. Early Devonian actinopterygians (Osteichthyes, Pisces) from Siberia. 233±243. In MARK-KURIK,
E. Fossil ®shes as living animals. Academia, 1, 1±299.ÐÐ 1993. Osteichthyes, Sarcopterygii. 657±663. In BENTON, M. J. (ed.). The fossil record 2. Chapman & Hall,
London, 841 pp.SCOTESE, S. R. and MCKERROW, W. S. 1990. Revised world maps and introduction. 1±21. In MCKERROW, W. S. and SCOTESE,
C. R. (eds). Palaeozoic palaeogeography and biogeography. Memoir of the Geological Society, London, 12, 1±435.STENSIOÈ , E. A. 1948. On the Placodermi of the Upper Devonian of East Greenland. II. Antiarchi: subfamily
Bothriolepidae. Meddelelser om Grùnland, 139, 1±622.TAVERNE, L. 1997. Osorioichthys marginis, `PaleÂonisciforme' du Famennien de Belgique, et la phylogeÂnie des
ActinopteÂrygiens deÂvoniens (Pisces). Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de laTerre, 67, 57±78.
THERY, J. M. 1982. Constitution du nord-ouest du continent sud-ameÂricain avant les tectoniques andines. UnpublishedPhD thesis, University of Bordeaux III.
TRAQUAIR, R. H. 1881. On the cranial osteology of Rhizodopsis. Transactions of the Royal Society of Edinburgh, 30,167±179.
ÐÐ 1888. Notes on the nomenclature of the ®shes of the Old Red Sandstone of Great Britain. Geological Magazine,(3), 5, 507±517.
WANG SHITAO 1993. Vertebrate biostratigraphy of the Middle Palaeozoic of China. 252±276. In LONG, J. A. (ed.).Palaeozoic vertebrate biostratigraphy and biogeography. Belhaven Press, London, 369 pp.
WELLS, J. W. 1944. Fish remains from the Middle Devonian bonebeds of the Cincinnati Arch Region. Palaeonto-graphica Americana, 3, 3±62.
WERDERLIN, L. and LONG, J. A. 1986. Allometry in the placoderm Bothriolepis canadensis and its signi®cance to antiarchevolution. Lethaia, 19, 161±169.
WHITE, E. I. 1978. The larger athrodiran ®shes from the area of Burrinjuck Dam, NSW. Transactions of the ZoologicalSociety of London, 34, 149±262.
WOODWARD, A. S. 1891. Catalogue of the fossil ®shes in the British Museum (Natural History), Part 2. British Museum(Natural History), London, 567 pp.
YOUNG, G. C. 1981. New Early Devonian brachythoracids (placoderm ®shes) from the Taemas-Wee Jasper region ofNew South Wales. Alcheringa, 5, 247±271.
ÐÐ 1982. Devonian sharks from south-eastern Australia and Antarctica. Palaeontology, 25, 87±89.ÐÐ 1984. Reconstruction of the jaws and braincase in the Devonian placoderm ®sh Bothriolepis. Palaeontology, 27,
625±661.
762 P A L A E O N T O L O G Y , V O L U M E 4 3

ÐÐ 1988. Antiarchs (placoderm ®shes) from the Devonian Aztec sandstone, southern Victoria Land, Antarctica.Palaeontographica, Abteilung A, 202, 1±125.
ÐÐ 1990. Devonian vertebrate distribution patterns and cladistic anaysis of palaeogeographical hypotheses. 243±255. In MCKERROW, W. S. and SCOTESE, R. C. (eds). Palaeozoic palaeogeography and biogeography. Memoir of theGeological Society, London, 12, 1±435.
ÐÐ 1991. Fossil ®shes from Antarctica. 538±567. In TINGEY, R. J. (ed.). The geology of Antarctica. OxfordMonographs in Geology and Geophysics, 17, Oxford University Press, Oxford, 579 pp.
ÐÐ 1993a. Untitled. Ichthyolith Issues, 11, 43±44.ÐÐ 1993b. Middle Palaeozoic macrovertebrate biostratigraphy of eastern Gondwana. 208±251. In LONG, J. A. (ed.).
Palaeozoic vertebrate biostratigraphy and biogeography. Belhaven Press, London, 369 pp.ÐÐ 1993c. Vertebrate faunal provinces in the Middle Palaeozoic. 293-323. In LONG, J. A. (ed.). Palaeozoic vertebrate
biostratigraphy and biogeography. Belhaven Press, London, 369 pp.ÐÐ and JANVIER, P. 1999. Early±Middle Palaeozoic vertebrate faunas in relation to Gondwana dispersion and Asian
accretion. 115±140. In METCALFE, I. (ed.). Gondwana dispersion and Asian accretion in the eastern Tethys andwestern Circum-Paci®c region (IGCP 321 ®nal report). Balkema, Rotterdam.
ÐÐ LONG, J. A. and RITCHIE, A. 1992. Crossopterygian ®shes from the Devonian of Antarctica: systematics,relationships and biogeographic signi®cance. Records of the Australian Museum, Supplement, 14, 1±77.
ÐÐ and ZHANG, G. R. 1992. Structure and function of the pectoral joint and operculum in antiarchs, Devonianplacoderm ®shes. Palaeontology, 35, 443±464.
PHILIPPE JANVIER
URA12 du CNRSLaboratoire de PaleÂontologie
MuseÂum National d'Histoire Naturelle8, rue Buffon
75005 Paris, [email protected]
CARLOS VILLARROEL
Departamiento de GeocienciasUniversidad Nacional de Colombia
Aptdo AeÂreo 56833BogotaÂ, Colombia
[email protected] received 11 February 1998Revised typescript received 31 January 2000
J A N V I E R A N D V I L L A R R O E L : D E V O N I A N V E R T E B R A T E S 763
Copyright © 2022 FDOKUMEN




















