LIM Homeodomain Factors Lhx3 and Lhx4 Assign Subtype Identities for Motor Neurons

21Development 125, 21-31 (1998)Printed in Great Britain © The Company of Biologists Limited 1998DEV2241
δEF1, a zinc finger and homeodomain transcription factor, is required for
skeleton patterning in multiple lineages
Tsuyoshi Takagi, Hiroki Moribe, Hisato Kondoh and Yujiro Higashi*
Institute for Molecular and Cellular Biology, Osaka University, 1-3 Yamadaoka, Suita, Osaka 565, Japan*Author for correspondence (e-mail: [email protected])
Accepted 28 October 1997; published on WWW 8 December 1997
δEF1 is a DNA binding protein containing a homeodomainand two zinc finger clusters, and is regarded as a vertebratehomologue of zfh-1 (zinc finger homeodomain-containingfactor-1) in Drosophila. In the developing embryo, δEF1 isexpressed in the notochord, somites, limb, neural crestderivatives and a few restricted sites of the brain and spinalcord. To elucidate the regulatory function of δEF1 in mouseembryogenesis, we generated δEF1 null mutant(δEF1null(lacZ)) mice. The δEF1null(lacZ) homozygotesdeveloped to term, but never survived postnatally. Inaddition to severe T cell deficiency of the thymus, theδEF1null(lacZ) homozygotes exhibited skeletal defects ofvarious lineages. (1) Craniofacial abnormalities of neuralcrest origin: cleft palate, hyperplasia of Meckel’s cartilage,
dysplasia of nasal septum and shortened mandible. (2)Limb defects: shortening and broadening of long bones,fusion of carpal/tarsal bone and fusion of joints. (3) Fusionof ribs. (4) Sternum defects: split and asymmetricossification pattern of the sternebrae associated withirregular sternocostal junctions. (5) Hypoplasia ofintervertebral discs. These results indicate that δEF1 hasan essential role in regulating development of these skeletalstructures. Since the skeletal defects were not observed inδEF1∆C727 mice, δEF1 bears distinct regulatory activitieswhich are dependent on different domains of the molecule.
Key words: δEF1, Gene targeting, Skeletal development, DNAbinding protein, Mouse
SUMMARY
nws
indof3).owiall.,f
r,
ivel.,do1).-
glfd
smh
INTRODUCTION
Skeletons of vertebrates develop from three distinct linea(Erlebacher et al., 1995). The neural crest gives rise to branchial arch derivatives of the craniofacial skeletons. Tsclerotome generates most of the axial skeleton includvertebrae and ribs. The lateral plate mesoderm contributes tolimb and sternum elements. Correct patterning and developmof these skeletal elements is fundamental to the divemorphology of vertebrate skeletons, and is a model systemunderstanding the pattern formation during animal developm
Hoxgenes, the vertebrate counterpart of DrosophilaHOM/Cgenes, have been considered to be master regulators of plan in vertebrate development. Their essential role development and patterning of skeletal elements has bdemonstrated (Duboule, 1994). Recent studies employinggene targeting method, however, have also shown equimportant involvement of other transcription factor genes skeletal development (Erlebacher et al., 1995). As presentethis report, δEF1 is one such new member participating skeleton patterning.
δEF1 was originally identified as an enhancer binding facof the chicken δ1-crystallin gene, but was found to bexpressed in a variety of tissues (Funahashi et al., 1991, 19δEF1 contains two Krüppel type-C2H2 zinc finger clusters
gestheheing theentrse forent.
bodyineen theallyind in
in
tore93).
located close to N- and C-termini with a homeodomain ibetween (Funahashi et al., 1993). The homeodomain shohighest similarity to those in LIM proteins (Bürglin, 1994).δEF1 is regarded as a vertebrate homologue of the Drosophilazinc finger homeodomain factor, zfh-1 (Fortini et al., 1991; Laet al., 1991), based on the similarity of the sequence aorganization of zinc-finger clusters and homeodomain, and exon-intron organization of the genes (Funahashi et al., 199Other vertebrate homologues have been reported, which shhighly conserved amino acid sequences; nil-2-a (a partsequence; Williams et al., 1991), AREB6 (Watanabe et a1993) and ZEB (Genetta et al., 1994) of human, MEB1 omouse (Genetta and Kadesch, 1996) identical to mouse δEF1(Sekido et al., 1996), BZP (Franklin et al., 1994) of hamsteand zfhep (Cabanillas and Darling, 1996) of rat.
The δEF1 homologues have been shown to have repressactivities on transcription (Franklin et al., 1994; Genetta et a1994; Kamachi and Kondoh, 1993; Postigo et al., 1997; Sekiet al., 1994, 1997; Watanabe et al., 1993; Williams et al., 199δEF1 preferentially binds to the CACCT sequence. The Cproximal zinc finger clusters are essential for DNA bindinactivity (Sekido et al., 1994), while the homeodomain by itsedoes not show an appreciable DNA binding activity (Ikeda anKawakami, 1995; Sekido et al., 1997). One possible mechaniin which δEF1 functions as a repressor is to compete wit

22
sehe 1
to
eomicceshe,
thethector
tingd byern
byack-pesainneith
;
T. Takagi and others
various bHLH activators for binding to the E2 box (CACCTG)sequence which overlaps with the δEF1 DNA binding sequence(Sekido et al., 1994). Overexpression of δEF1 in the 10T1/2cells actually inhibited the MyoD-induced muscldifferentiation (Postigo and Dean, 1997; Sekido et al., 1994
δEF1 is expressed in various anlages of developing tissbesides lens cells. In chick embryos, δEF1 is first detectedduring the postgastrulation period in the mesodermal tissuinitially in the notochord, followed by somites, heart, limb buand other components. In addition, δEF1 is also expressed inthe nervous system and neural crest derivatives suchcephalic mesenchyme (Funahashi et al., 1993). A simexpression pattern was observed in the mouse (this paper
To investigate the function of δEF1 during mouseembryogenesis, we took advantage of gene targeting. previously generated δEF1 mutant mice (δEF1∆C727) lackingthe C-terminal portion downstream of the 727th amino aresidue, which included the C-proximal zinc-finger clusteδEF1∆C727 mutant mice showed severe hypocellularity in ththymus, resulting from the depletion of c-kit+ early T cellprecursors, while many other tissues were normal in spiteexpression of δEF1 (Higashi et al., 1997).
To approach the entire function of δEF1, and to gain insightinto the possible domain-specific regulatory functions, wgenerated δEF1null mutant (δEF1null(lacZ)) mice which lackedalmost the entire δEF1 coding sequence. As presented in threport, skeletal defects of craniofacial, limb, and vertebcolumn development were observed in these mice as wethe same thymus defect as found in the δEF1∆C727mutant. Theresults demonstrate the essential role of δEF1 in skeletondevelopment, and suggest differential usage of its functiodomains in different tissues.
Fig. 1. Generation ofδEF1null(lacZ) mutant mice.(A) Structure of the δEF1geneand targeting vector. Openboxes represent exons. Exon 2is not shown since its exactlocation has not beendetermined. Vertical bars abovethe line indicate EcoRI siteswhile those below the line NcoIsites. EH and BN are the 5′ and3′ flanking sequence probes,respectively, used foridentification of thehomologous recombinants andfor the subsequent genotyping.(B) Confirmation ofhomologous recombination andgenotyping of mutant mice bySouthern analysis. The blot ofEcoRI-digested DNAs from theES (R1) cells, the targeted ESclones #292 and #355, and theembryos of each genotype werehybridized with the EH probe.(C) Northern blot analysis ofδEF1expression inδEF1null(lacZ) mutant embryos atE13.5. 5 µg of whole embryo RNA was hybridized with 2.5 kb δEF1cDHeterozygotes had half the amount of δEF1RNA compared with wild-
e).
ues
es:d
asilar).
We
cidrs.e
of
e
isralll as
nal
MATERIALS AND METHODS
Construction of targeting vectorUsing mouse δEF1 cDNA (Sekido et al., 1996) as a probe, a mougenomic DNA library of 129/Sv strain was screened to obtain tmouse δEF1 gene sequence. The DNA fragment containing exonwas used for construction of the targeting vector (Fig. 1).
A 2.5 kb genomic fragment spanning the region from upstreamthe middle of the exon 1 of δEF1, the 3.8 kb BamHI fragment ofpMoZtk (Ueno et al., 1987) coding for β-galactosidase, the XhoI-BamHI fragment of pSTneoB (Kato et al., 1987) containing thneomycin-resistance gene in reverse orientation, and a 7.8 kb genfragment of intron I were joined using appropriate linker sequenof pBluescrjpt II SK+. The combined fragment was inserted into tDT-A (Diphtheria Toxin A-fragment) cassette vector (Yagi et al.1993), generating the targeting vector pTVδEF1null(lacZ)-DT. The lacZfusion protein coded by this vector contains 15 amino acids of exon 1 and 12 amino acids from the linker sequence followed by β-galactosidase coding sequence. The resulting targeting vecontains a 2.5 (5′) and a 7.8 kb (3′) sequence homologous to thegenomic sequence (Fig. 1A).
Gene targetingES cells (R1; Nagy et al., 1993) were electroporated with the targevector and selected for G418 resistance (200 µg/ml) as describeSawai et al. (1991). The targeted clones were identified by Southhybridization (Fig. 1B). Germ-line chimera mice were obtained blastocyst injection of the targeted ES cells. Heterozygotes were bcrossed to C57BL/6 for five generations before analysis. Genotywere determined by Southern hybridization or PCR (polymerase chreaction) analysis of DNAs from ear, tail or yolk sac. PCR was doat 94°C for 30 seconds, 65°C for 1 minute and 72°C for 1 minute w35 cycles, using: primer J, TAGGTGTTAGGAAGGTGATGTCGprimer I, AACCGTGCATCTGCCAGTTTGAG.
NA as probe. The same membrane was rehybridized with G3PDH.type, and there was no mRNA in the homozygotes.
23Skeletal abnormalities in δEF1-knockout mice
the
ialFig.s
,
he
In situ hybridizationWhole-mount in situ hybridization was done according to Wilkinso(1992) using digoxigenin-labeled RNA probes. δEF1, 0.8 kb SalI-ApaI cDNA fragment; Hoxd-13, a 253 bp cDNA fragment providedby Dr D. Duboule.
Skeleton stainingBone and cartilage were stained according to a standard prot(Hogan et al., 1994), except for E12.5-15.5 embryos which were fifixed in Bouin’s fixative for 3 hours, and then washed in 70% ethawith frequent change of the solution for 2 days or until the yellowicolor disappeared.
Histology Specimens were fixed in Bouin’s fixative, embedded in paraffsectioned at a thickness of 7 µm, and stained with hematoxylin eosin (HE).
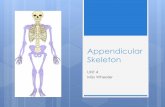
Fig. 2. δEF1expression in early embryos. δEF1expression in embryowith whole embryos (A-G, I-K, N), with a transverse section at thewas examined for comparison with the δEF1expression pattern by in was also examined by X-gal staining of δEF1null(lacZ) heterozygous emstaining and in-situ hybridization were partly ascribable to the fact localized in the nucleus (data not shown), while in situ hybridizatioforelimb bud; h, headfold; hl, hindlimb bud; lm, lateral myotome; mcells; ps, presomitic mesoderm; tb, tail bud; v, ventricular zone of nin B and G indicate non-specific signals produced by probes trappScale bars: (A) 160 µm, (B,E,O) 320 µm, (C,D,F,I) 250 µm, (G,N,P) 50
n
ocolrst
nolsh
in,and
RESULTS
Expression pattern of mouse δEF1 during earlyembryogenesisThe expression pattern of mouse δEF1 was examined atvarious developmental stages by in situ hybridization.
In embryos at E8.5, strong δEF1 expression was firstdetected in the headfold and moderate expression in presomitic and lateral plate mesoderm (Fig. 2A).
At E9.5, δEF1was expressed in the derivatives of the cranneural crest, such as branchial arches and cranial ganglia (2B). In individual somites δEF1 expression was homogenouand at moderate levels. Strong δEF1 expression also occurredin the limb buds (Fig. 2B). In hindlimbs of the E9.5 embryowhich was just at the beginning of limb outgrowth, δEF1 wasexpressed evenly in the entire limb bud (Fig. 2C), while in t
was examined by in situ hybridization of the δEF1probe (A-E,G,H,I-M) trunk level (H) and with sections of forelimb (L,M). The Hoxd-13expressionsitu hybridization of the Hoxd-13(F,N) probe. The δEF1expression patternbryos (O,P). Notable differences in the tone of staining between X-galthat the β-galactosidase had the δEF1N-terminal 15 amino acids mostlyn signals were in the cytoplasm. cg, cranial ganglia; d, dorsal root ganglia; fl,m, medial myotome; my, migrated myotome; n, neural tube; nc, neural cresteural tube. A and P indicate the direction of anterior-posterior axis. Asterisksed in the brain ventricles and observed with sense probes (data not shown).0 µm, (H) 250 µm, (J,K) 400 µm, (L,M) 320 µm.
24
dyorewn
l-
lye
m
nisat
to intheas
nt
the
ryos
etd
theino
reblot
ut
T. Takagi and others
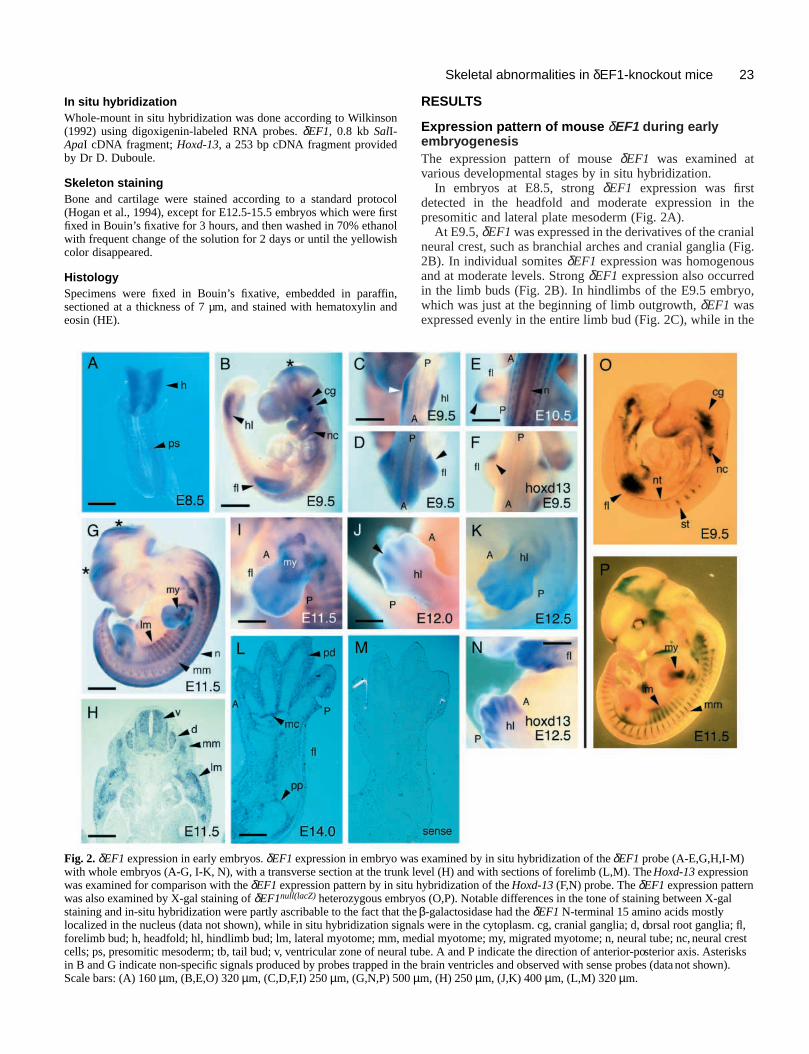
Fig. 3. External morphology of δEF1null(lacZ) homozygous mutantembryo at E15.5. Control (A) and mutant (B,C) embryos of the samelitter were compared. Note that the mutant embryos were stoopedand were shorter in the body. The embryo in B had short limbs andmandible (arrowheads). The phenotype of the embryo in C was moresevere and included edema and a curled tail (additional arrowheads).Growth and development were apparently retarded in this embryo.Scale bar, 1.6 mm.
Fig. 4. Craniofacial defects in the δEF1null(lacZ) mutant.(A-D) δEF1null(lacZ) mutant mice had a cleft in the secondary palate.(A,B) Ventral view of the base of the skull of control (A) and mutant(B) embryos at E18.5. (C,D) Frontal sections of the heads of control (C)and mutant (D) embryos at E18.5 were stained with HE.(E,F) Transverse sections of the head of control (E) and mutant (F)embryos at E18.5 stained with HE. Ectopic cavity structure (arrow) wasformed in the nasal septum of the mutant. The number of serous glands(sg) was less in the mutant. (G,H) Skeletal preparation of control (G)and mutant (H) mandibles at E18.5. (I,J) Cartilage staining of control(I) and mutant (J) embryos at stage E14.5. a, angular process; bs,basisphenoid bone; c, coronoid process; co, condylar process; d, dentalprocess; m, Meckel’s cartilage; ns, nasal septum; sg, serous gland;pl, palatal bone; ps, palatal shelf; pt, pterygoid bone. Scale bars: (A,B)570 µm, (C,D) 320 µm, (E,F) 320 µm, (G,H) 640 µm, (I,J) 250 µm.
forelimb of the same embryo, where the limb bud was alreaprominent, δEF1 expression was confined to the posteriportion (Fig. 2D). δEF1 expression included the region of thHoxd-13expression (Fig. 2F) or other genes which are knoto be expressed posteriorly (e.g. Shh, Bmp2, Fgf4). Later atE10.5, δEF1expression in the limb was restricted to the distaposterior region (Fig. 2E).
At E11.5, δEF1 expression in the neural tube was cleardetected (Fig. 2E,G,H), being particularly prominent in thventricular zone (Fig. 2H). Dorsal root ganglia derived frothe neural crest also expressed δEF1 (Fig. 2H). In the somitesof the trunk, δEF1 was confined to the myotome. Expressiowas particularly high in the lateral myotome (Fig. 2G,H). Thmyotomal δEF1 expression was maintained in those cells thmigrated in the limb (Fig. 2G,I). In limb, δEF1expression waslost from the condensation of mesenchyme which gives risecartilaginous skeleton (Fig. 2I), then expressed transientlythe interdigit mesenchyme at E12.0 (Fig. 2J), and finally in perichondrium of the cartilage when the skeletal pattern walready formed (Fig. 2K,L). At this stage of limb developmealso, overlap of expression of δEF1 and Hoxd-13was noted(Fig. 2K,N).
The results described above in general agreed with immunohistological observations of chicken δEF1 (Funahashiet al., 1993). In contrast to the chicken, however, δEF1expression was not detected in lens cells of the mouse emb(data not shown)
Generation of δEF1 null mutant miceA targeting vector was designed so that the STneoB (Katoal., 1987) and lacZ gene, each carrying a poly(A) addition antermination signals, were inserted into exon 1 of the δEF1gene(Fig 1A). The resulting mutated allele was expected to lack δEF1 activity but encoded a fusion protein of the first 15 amacids of δEF1 with a β-galactosidase. (Fig. 1A).
A total of 10 clones of homologous recombinants weidentified among 324 G418 resistant clones by Southern analysis using 5′ and 3′ sequences as probes (Fig. 1B and danot shown). Four clones were used to generate chimeric mby blastocyst injection. Three of them gave rise to machimeras which transmitted the mutation to their offsprin
taiceleg.
Heterozygous mutants were fertile and healthy withoappreciable abnormalities in morphology and behavior.
The X-gal staining pattern of δEF1null(lacZ) heterozygotes at
25Skeletal abnormalities in δEF1-knockout mice
dThetal
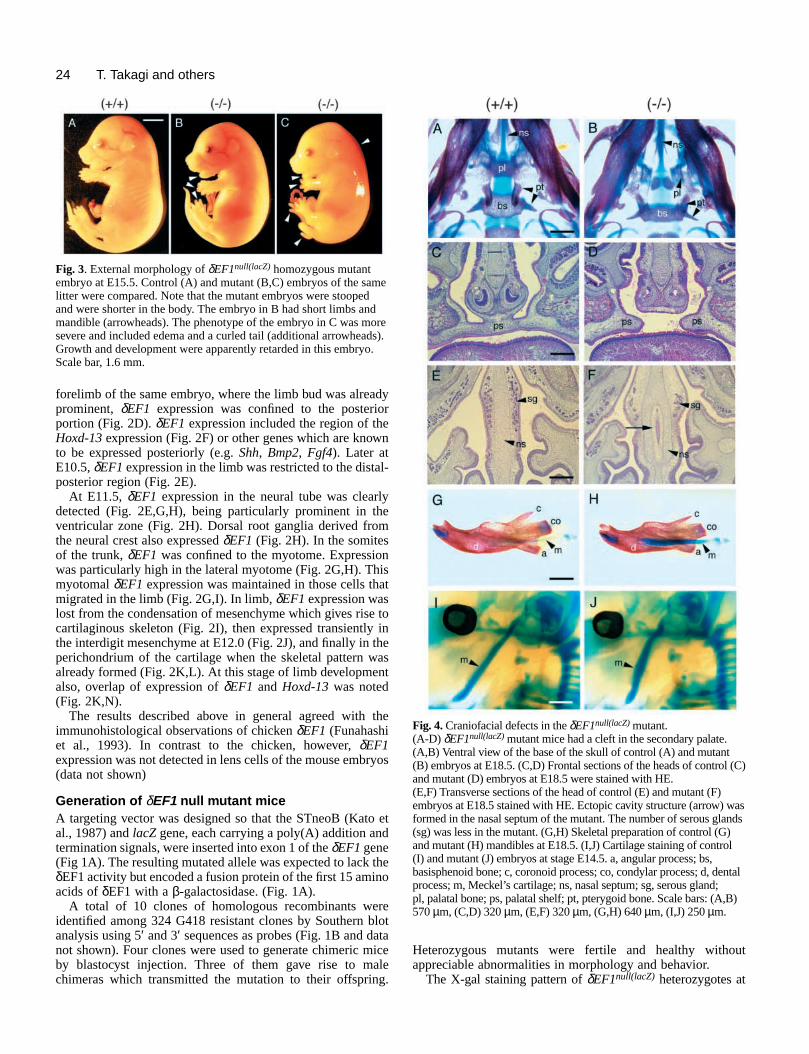
ts in the limbs of δEF1null(lacZ) mutant mice. (A-D) Forelimb (A,B) andntrol (A,C) and mutant (B,D) embryos at E18.5. Arrowheads indicate the tibia (t) in the mutant. (E-H) Autopod of forelimb (E,F) and hindlimb) and mutant (F,H) embryos at E18.5. Arrowheads in F indicate absenceal end of P3 and P1. Arrowheads in H indicate a fusion of metatarsal (c1) in the mutant. (I-L) Magnification of distal part of carpal/tarsal
J) and hindlimb (K,L) of control (I,K) and mutant (J,L) embryos. L indicate the malformed carpal and tarsal bones, respectively, in thedetails. The navicular (n) and talus (t) were also fused in the mutant (L).elimb (M,N) and hindlimb (O,P) of control (M,O) and mutant (N,P)alformation of carpal/tarsal bones was caused by mislocation ofsors in the mutant. Scale bars: (A-D) 800 µm, (E-H) 270 µm, (I-L) 160
E9.5 (Fig. 2O) and E11.5 (Fig. 2P) mimicked the in shybridization data. One large deviation was the δEF1expressionin the notochord which was clearly indicated by the X-gstaining of the δEF1null(lacZ) heterozygotes (Fig. 2O), but wabarely detectable by in situ hybridization (Fig. 2B,H). Themight be some specific mechanisms regulating completion stability of δEF1 transcript in thenotochord.
Northern analysis revealed that δEF1transcripts in heterozygotes werereduced to half the level of wild-type,while they were undetectable in thehomozygotes (Fig. 1C). Thus, thehomozygote was regarded as null, andthe allele was termed δEF1null(lacZ).
Morphology of δEF1null(lacZ)
mutant miceδEF1null(lacZ) homozygous mutantembryos developed to term, but diedshortly after birth (data not shown). UponCaesarean section at E18.5, thehomozygote did not start breathing, anddied of cyanosis. The cause(s) of therespiratory failure has not been clarified.
Earlier in embryogenesis, thehomozygotes could be distinguished fromother genotypes by their morphology (Fig.3): growth retardation which becameevident from the stage around E15.0; shortand dumpy limbs (E14.5-18.5,100%=67/67); shortened distal maxillaand mandible; curled tails (E12.5-18.5,89%=73/82); frequent edema atmidgestation (E13.5-16.5 52%=34/65).Also observed, though less frequently, wasexencephaly and internal bleeding in thenasal region (E13.5-18.5, 3%=3/80 and5%=4/80, respectively). Failure of spinalcord closure was observed in cranial andcaudal ends, 1% (1/80) and 4% (3/80), ofthe homozygotes, respectively, while nosuch abnormalities were observed insibling heterozygotes or wild types.
A series of sections of homozygotes atE14.5 and E16.5 showed no grossabnormalities other than those in skeletalelements which are discussed below, andthe infrequent neural tube closure defectand exencephaly. Sections of E18.5homozygotes, however, revealed smallhypocellular thymi with no distinction ofmedulla and cortex, as observedpreviously in theδEF1∆C727mutant mice(Higashi et al., 1997). The total numberof thymocytes in E18.5 embryos wasreduced to 1/10 that of the control. FACSanalysis showed significant reduction ofc-kit+ cells in the CD4−CD8− thymocytecell population, indicating thatδEF1null(lacZ)mutation caused exactly the
Fig. 5. Skeletal defechindlimb (C,D) of cocurved radius (r) and(G,H) of control (E,Gof ossification of dist(mt) and cuneiform 1bones of forelimb (I,Arrowheads in J andmutant. See text for (M-P) Autopod of forembryos at E14.5. Mchondrogenic precurµm, (M-P) 160 µm.
itu
alsreand
same phenotype in thymus as seen in δEF1∆C727 mutant mice(data not shown).
Skeletal analysis of the δEF1null(lacZ) mutants by stainingwith alcian blue for cartilage and with alizarin red S for ossifiebones revealed abnormalities in various skeletal elements. mutant phenotype, including these abnormalities of skele

26
thetroledInnd
tionr atf
d1, arely.de
ed
rate
T. Takagi and others
elements, are tabulated and compared with those of omutant mice (Table 1).
Craniofacial skeletonsWhen examined at E18.5, δEF1null(lacZ) mutants had a cleft inthe secondary palate (Fig. 4A-D). Hypoplasia of the palbones as well as deformation of basisphenoid and pterygbones were noted (Fig. 4A,B). Dysplasia of the nasal sepwas also observed in the mutants (56%=5/9; Fig. 4A,Sections revealed formation of an ectopic cavity in the naseptum (Fig. 4E,F).
The mandible and premaxilla were shortened in the muta(Fig. 4G,H and data not shown). The coronoid, condylar, aangular processes in the mutant mandible were hypopla(Fig. 4G,H), while Meckel’s cartilage was hyperplastic (Fi4G,H), which was already noticeable at early chondrogestage, E14.5 (Fig. 4I,J).
Table 1. Summary of δEF1 expression sitesδEF1expression site Phenotype
Neuroectoderm derivativesbrain infrequent exencephalyspinal cord (ventricular zone) infrequent neural tube
curled tail
Neural crest derivativesganglia
dorsal root ganglia n.d.cranial ganglia n.d.sympathetic ganglia n.d.
craniofacial bonesMeckel’s cartilage (hypemaxyllary bone (hypoplamandibular bone (hypoppalatine bone (absent)basisphenoid bone (defopterygoid bone (deformenasal septum (cavity)
Mesoderm derivativeshead fold mesenchymelateral mesoderm sternebrae (split ossific
asymmetric sternocosxiphoid process (absent)
somite rib (fusion)anulus fibrosus (IVD) (re
notochord nucleus pulposus (IVD)limb bud mesenchyme limb skeleton
long bones (shortenedradius/tibia (curved)carpal/tarsal bones (fuP1, P2 (delayed or noP3, (no ossification)joint (elbow/hip) (fusedpatella (displaced)
myotome/muscle system n.d.
mesenchymal cellsheart n.d.kidney n.d.lung n.d.
n.d.: not detected. n.f.: not found.
aPenetrance of the phenotype is not considered.bEpstein et al. (1991); cSatokata and Maas, (1994); dMartin et al. (1995); eMo e
al. (1993); iCondie and Capecchi, (1993); jKingsley et al. (1992); kSuemori et aloDavis and Capecchi, (1996); pStorm and Kingsley, (1996); qDolle et al. (1993).
ther
ateoid
tumB).sal
ntsndsticg.nic
Limb skeletonsLong bones in the stylopod and zeugopod were shorter inmutants, and appeared to be slightly thicker than the con(Fig. 5A-D). The radius and tibia in the mutants were curv(Fig. 5A-D). The scapula was also shortened (Fig. 5A,B). the autopod, growth of phalanges (P1, P2, P3) in fore- ahindlimbs was severely attenuated: endochondral ossificaof P1 and ossification at the distal end of P3 did not occuE18.5 (Fig. 5E-H). Fusion of P1 and P2 in digits III and IV oforelimb was detected in some cases (Fig. 5F).
In normal embryos, there are 5 distal carpal bones calledd2, d3, d4 and a central carpal bone (c). d1, d2 and d3located proximal to the metacarpi, I, II, and III, respectived4 is generated by fusion of d4a and d4b which are maproximal to metacarpal IV and V, respectively. c is producby a split of d2. In most of the δEF1null(lacZ) mutant embryos,three of the carpal bones (d2, d3 and c) did not exist as sepa
and phenotype of δEF1null(lacZ) mutant miceSimilarity to known mutantsa
Pax3b
closure defect Pax3b
Pax3b
rplasia) n.f.sia) Msx1c, Mhoxd, Gli2e
lasia) Msx1c, Mhoxd, Gli2e
Hoxa-2f, Msx1c, Mhoxd, Gli2,3e
rmed) Gli2e
d) Mhoxd
n.f.
ation centers andtal junctions) Hoxa-4g, Hoxa-5h, Hoxd-3i
Bmp5j
Hoxc-9k
duced) Gli2e
(reduced) Gli2e
and thickened) Mhoxd
Hoxa-11l
sed) Hoxa-13m, Hoxd-11n, Hoxd-13o, GDF5p
ossification) Hoxd-13q, Gli2e
n.f.) n.f.
GDF5p
t al. (1997); fGendron-Maguire et al. (1993); gHoran et al. (1994); hJeannotte et. (1995); lDavis et al. (1995); mMortlock et al. (1996); nFavier et al. (1996);
27Skeletal abnormalities in δEF1-knockout mice
ts,ow, the6B,hipata
ay: thentsed
istalthe
ut5.0ibion
e
structures but as a single large fused bone (stage E15.5-94%=16/17; Fig. 5I,J). d4a and d4b, which normally fuseform d4 at stage E15.0, were often separated in the mutantstage E15.5-18.5 56%=9/16; Fig. 5I,J).
There are also 5 distal tarsal bones, cuneiform 1 tocuboideum and navicular. In all the δEF1null(lacZ) mutantsexamined, cuneiform 2 and 3 were replaced by a large b(stage E15.5-18.5 100%=16/16; Fig. 5K,L). Fusion navicular and talus, and that of metatarsal I and cuneiformin digit I were also observed in all the mutants examinedE18.5 (100%=18/18; Fig. 5G,H,K,L).
The distal carpal/tarsal bones were already affected atchondrogenic precursor stage of E14.5 embryos. In the muforelimbs, the central carpal bone precursor was reduced wd4 was hyperplastic. d2-c was fused with d3 (Fig. 5M,N). the hindlimb, cuneiform 2 was hypoplastic and already fuswith cuneiform 3. The navicular was also hypoplastic, adislocated near the talus (Fig. 5O,P). The navicular becafused to the talus at later stages (data not shown). Comparof the mutant carpal/tarsal bones of different stages indicathat the morphological alterations in the mutant bones wcaused mainly by the fusion of primary cartilaginouprecursors at an early stage, rather than by deletion ofprecursors.
and/15;edionn,
thec
Fig. 6. Joint defects of the δEF1null(lacZ) mutant. (A,B) Elbow joint ofthe control (A) and mutant (B) embryos at E18.5. The humerus (hand ulna (u) were fused in the mutant (arrowhead). (C,D) Sectionthe elbow of control (C) and mutant (D) embryos at E18.5. Theradius (r) and humerus (h) were fused in this mutant (D). (E,F) Thposition of the patella (p) was shifted more medially in the mutant(F) as compared to the control (E). t, tibia; f, fibula. Scale bars: (A230 µm, (C,D) 320 µm, (E,F) 640 µm.
18.5, tos (at
3,
oneof 1
at
thetanthileInedndmeisontederes
the
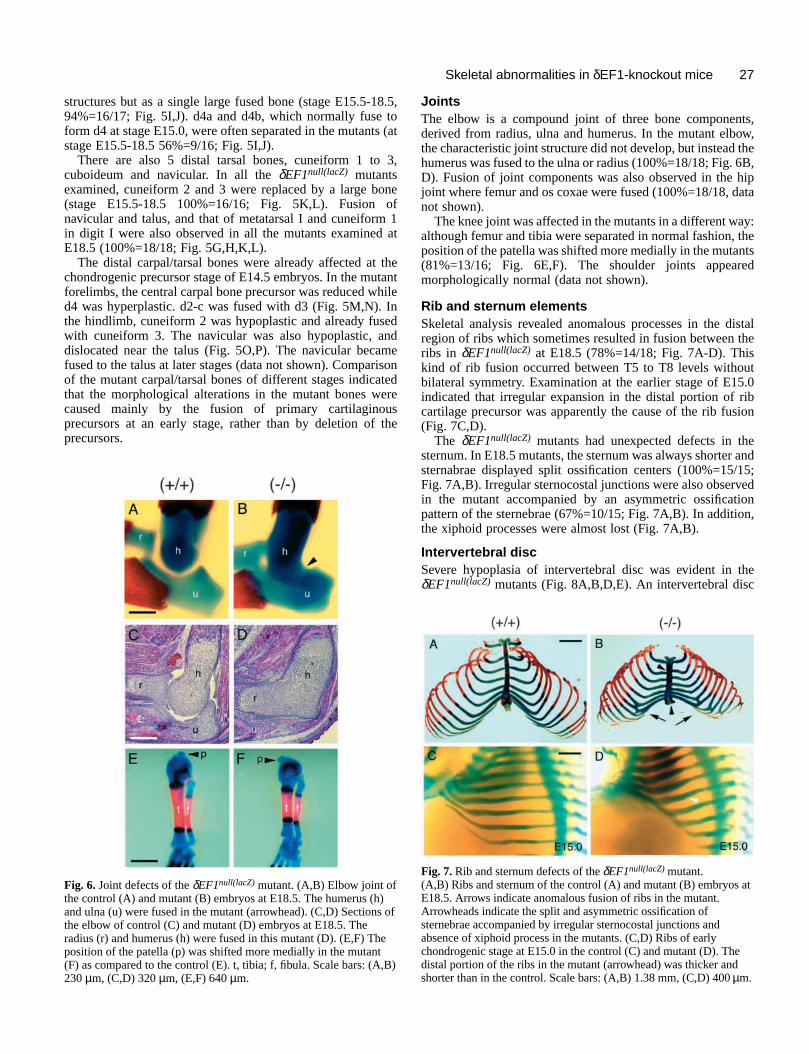
Joints The elbow is a compound joint of three bone componenderived from radius, ulna and humerus. In the mutant elbthe characteristic joint structure did not develop, but insteadhumerus was fused to the ulna or radius (100%=18/18; Fig. D). Fusion of joint components was also observed in the joint where femur and os coxae were fused (100%=18/18, dnot shown).
The knee joint was affected in the mutants in a different walthough femur and tibia were separated in normal fashion,position of the patella was shifted more medially in the muta(81%=13/16; Fig. 6E,F). The shoulder joints appearmorphologically normal (data not shown).
Rib and sternum elementsSkeletal analysis revealed anomalous processes in the dregion of ribs which sometimes resulted in fusion between ribs in δEF1null(lacZ) at E18.5 (78%=14/18; Fig. 7A-D). Thiskind of rib fusion occurred between T5 to T8 levels withobilateral symmetry. Examination at the earlier stage of E1indicated that irregular expansion in the distal portion of rcartilage precursor was apparently the cause of the rib fus(Fig. 7C,D).
The δEF1null(lacZ) mutants had unexpected defects in thsternum. In E18.5 mutants, the sternum was always shortersternabrae displayed split ossification centers (100%=15Fig. 7A,B). Irregular sternocostal junctions were also observin the mutant accompanied by an asymmetric ossificatpattern of the sternebrae (67%=10/15; Fig. 7A,B). In additiothe xiphoid processes were almost lost (Fig. 7A,B).
Intervertebral discSevere hypoplasia of intervertebral disc was evident in δEF1null(lacZ) mutants (Fig. 8A,B,D,E). An intervertebral dis
)s of
e
,B)
Fig. 7. Rib and sternum defects of the δEF1null(lacZ) mutant.(A,B) Ribs and sternum of the control (A) and mutant (B) embryos atE18.5. Arrows indicate anomalous fusion of ribs in the mutant.Arrowheads indicate the split and asymmetric ossification ofsternebrae accompanied by irregular sternocostal junctions andabsence of xiphoid process in the mutants. (C,D) Ribs of earlychondrogenic stage at E15.0 in the control (C) and mutant (D). Thedistal portion of the ribs in the mutant (arrowhead) was thicker andshorter than in the control. Scale bars: (A,B) 1.38 mm, (C,D) 400 µm.

28
areiousthesitun,tly
ll
theeds
velsnot
d in
ate.,esgh
T. Takagi and others
lastic intervertebral disc of the δEF1null(lacZ) mutant. Lateral (A,D) andview of vertebral columns of the control (A,B) and mutant (D,E)18.5. Note that the intervertebral disc (iv) was severely hypoplastic in,E arrowheads). (C,F) Sagittal sections of the control (C) and mutant
at E18.5 were stained with HE. Nucleus pulposus (np) in thel disc was clearly hypoplastic in the mutant (arrows). Arrowheadsnnulus fibrosus (af) surrounding the nucleus pulposus. Scale bars: (A,D)
) 250 µm, (C,F) 160 µm.
is composed of two tissues, nucleus pulposus, which is derifrom notochord, and annulus fibrosus, the cartilaginous cederived from intervertebral sclerotomal cells (Fig. 8B,CHistological analysis of the mutant intervertebral discrevealed that the cell number of nucleus pulposus wsignificantly reduced (Fig. 8C,F), while the surroundincartilaginous cells of annulus fibrosus did not appear muaffected (Fig. 8F).
Deviation from normal morphology of the disc had alreadbeen apparent in the mutant at E14.5 (data not showExpression of collagen type II(Cheah et al., 1991), Pax-1(Wallin et al., 1994) and Ndr-1 (Shimono et. al., unpublisheddata) in the intervertebral discs was not affected in the mutaanalyzed at E15.5 by in situ hybridization (data not shown)
DISCUSSION
Concordance between δEF1 expression and themutant phenotypeWe generated δEF1 null mutant mice to investigate thefunction of δEF1 in vivo. The δEF1null(lacZ) mutant miceexhibited skeletal defects in craniofacial bones, ribintervertebral disc, sternum and limb skeletons. These skelelements are derived from different primordia, the cranneural crest, somite, notochord and lateral plate mesodermthese tissue primordia express δEF1 (Fig. 2A,B). Thus, the results indicate a goodconcordance between the tissues affected by themutation and earlier expression of δEF1 in thosetissues (Table 1).
The case of rib abnormality in δEF1null(lacZ)
mutants, however, is not straightforward. It isintriguing that δEF1 is expressed in the entireearly somite but not in the sclerotome, the directprecursor of the ribs. An analogous situation isfound in the mice mutant for myogenic factors,myogenin, myf5 and MRF4, all having certainrib defects, although the genes are notappreciably expressed in the sclerotome (Braunet al., 1992; Hasty et al., 1993; Zhang et al.,1995). Expression of these myogenic factorsand δEF1 in an early somite stage or inmyotome of a later stage seems essential forproper development of the ribs. Considering thepossible interaction of these myogenic factorswith δEF1 as discussed below, a rib phenotypemay be developed when there is an imbalanceamong the interacting factors in the early somiteor the myotome.
Major deviation from the concordancebetween the δEF1 expression site and mutantphenotypic manifestation are noted in themuscular and neuronal cell lineages. In musclecells, possible interaction of δEF1 withmyogenic factors has been suggested, sinceδEF1 can counteract the function of bHLHproteins, including myoD, by competitivebinding to common cis-element E2-box intransfected cultured cells (Postigo and Dean,1997; Sekido et al., 1994). δEF1 is indeed
Fig. 8. Hypopfrontal (B,E) embryos at Ethe mutant (D(F) embryos intervertebraindicate the a250 µm, (B,E
vedlls
).sasgch
yn).
nts.
s,etalial; all
highly expressed in tissues where myogenic bHLH proteins expressed (Fig. 2; Funahashi et al., 1993); however, no obvabnormality in the muscular system was detected in mutants. Overall levels and distribution of mRNAs, aexamined by northern blot at E13.5 and whole mount in shybridization at E10-11, respectively, of myoD, myogenimuscle creatine kinase and desmin were not significanaffected in δEF1null(lacZ) mutant embryos (data not shown).
α4 integrin is also activated during muscle cedifferentiation (Rosen et al., 1992). In addition, α4 integrinpromoter is reported to be repressed in cultured cells by δEF1 homologue, ZEB (Postigo et al., 1997). We reportrecently that α4 integrin expression in immature thymocytewas significantly increased in the δEF1∆C727 mutant mice(Higashi et al., 1997). We, therefore, examined the overall leof α4 integrin mRNA at E13.5 and E15.5, but no obviouchange was detected by northern blot analysis (data shown).
It is possible that some other factors which are expressemyogenic lineage cells and have a function similar to δEF1could compensate for the effects of deficiency of δEF1. Themouse Snais such a candidate gene which could compensfor the loss of δEF1 function (see Discussion in Higashi et al1997). An analogous situation may exist in neural tissuwhere no morphological defect was found in spite of the hiexpression of δEF1.

29Skeletal abnormalities in δEF1-knockout mice
ns.
m
her
l.ich
deenernn
7).the
d inalnd eted
toy,t in
n. Itnal
lyus
es,
δEF1∆C727
δEF1null(lacZ) (β-galactosidase)
Thymus
Thymus andskeletal elements
Protein structure Sites affected
None
region I region II
Required for normal development of the skeletal elements. Region I :
Region II : Required for thymus development, but is not for skeletal development.
N-fin C-finHD
Wild-type
(Higashi et al., 1997)
Fig. 9. A model for domain-specific regulatoryfunction of δEF1. Protein encoded by each allele ofδEF1 (wild-type, δEF1∆C727and δEF1null(lacZ)) isschematically shown, and correspondence with thephenotype is indicated. N- and C-fin indicate thezinc finger clusters proximal to N- and C-termini,respectively. HD indicates the homeodomainsequence.
Analogies between skeletal phenotypes of theδEF1null(lacZ) and Hox mutant miceIt is interesting to note that many of the skeletal defeobserved in the δEF1null(lacZ) mutants recapitulated those inmice mutant for some of the Hox genes (Table 1): cleft palatein Hoxa-2mutant (Fig. 4A-D; Gendron-Maguire et al., 1993Rijli et al., 1993), irregularities in the sternocostal junctions Hoxa-4, Hoxa-5and Hoxd-3mutants (Fig. 7A,B; Condie andCapecchi, 1993; Horan et al., 1994; Jeannotte et al., 1993)fusion of ribs in Hoxc-9 mutants (Fig. 7A,B; Suemori et al.,1995). The shared mutant phenotypes between δEF1 and Hoxgenes were particularly remarkable in the limb. The radiusthe δEF1null(lacZ) mutant was curved, showing a closresemblance to theHoxa-11−/− mutant (Davis et al., 1995). Acharacteristic of the δEF1null(lacZ) mutant limb was fusion ofthe three (d2, d3 and c) carpal bones (Fig. 5J). Variationsthe carpal bone phenotype similar to those of the δEF1null(lacZ)
mutants were observed in single or compound mutants of soof the Hox genes: fusion of d1 and d2 in heterozygotes spontaneous Hoxa-13mutation, hypodactyly(Mortlock et al.,1996); fusion of c and d3 in Hoxd-11−/− mutants (Favier et al.,1996); fusion of d3 and d4, and indentation of d4 in compouheterozygotes, [Hoxd-11+/−; Hoxd-13+/−] and [Hoxd-12+/−;Hoxd-13+/−] (Davis and Capecchi, 1996); split of d4 in Hoxd-13−/− mutant and compound heterozygote [Hoxa-13+/− ; Hoxd-13+/−] (Dolle et al., 1993; Fromental-Ramain et al., 1996).
Major expression domains of δEF1 and 5′ proximal Hoxdgenes are superimposed in the limbs (Fig. 2C-F,I-N): in ealimb bud, δEF1 is expressed uniformly throughout the limbbud as is Hoxd-9and 10 (Dolle et al., 1989). As the limb budgrows, δEF1 became localized posteriorly, and then confineto the distal-posterior region (Fig. 2D,E). Similarly, Hoxd-11,12 and 13 are known to be expressed in the posterior regi(Fig. 2F; Dolle et al., 1989). When the skeletal pattern is onformed (Fig. 2J, K), δEF1 expression is observed in theperichondrium of the cartilage as are the Hoxd-12and 13genes(Fig. 2N; Dolle et al., 1989, 1993). Thus, the spacio-tempopattern of δEF1 expression during the limb developmensignificantly overlaps that of 5′ proximal Hoxdgenes.
The shared mutant phenotype and expression patbetween the δEF1 and Hox genes, particularly in the limb, led
cts
;in
and
ofe
of
meof
nd
rly
d
once
ralt
tern
us to postulate possible models for their genetic interactioSince expression of the 5′ proximal Hoxd and Hoxa genes inlimb were not significantly affected in δEF1null(lacZ) mutants(data not shown), the Hox genes are probably not downstreaof δEF1, but epistatic to the δEF1 gene in a stage of limbdevelopment. It would thus be interesting to examine whetδEF1 expression is affected in the Hox mutants. Anotherintriguing possibility is direct interaction at the protein leveδEF1 contains a homeodomain (Funahashi et al., 1993), whitself does not show DNA binding activity (Ikeda anKawakami, 1995; Sekido et al., unpublished data). It has bindicated that Hox proteins heterodimerize with othhomeodomain-containing proteins and function in vivo (Maand Chan, 1996). It is also to be noted that the δEF1∆C727
mutants did not exhibit skeletal defects (Higashi et al., 199Considering the fact that the homeodomain is retained in δEF1∆C727mutant protein, direct interaction of δEF1 with Hoxproteins is an attracting model.
Other mouse mutants which show some similaritiesto δEF1null(lacZ) miceRecently reported GDF5 (growth and differentiation factor 5)mutant mice also showed the skeletal phenotype observeδEF1null(lacZ) mutant mice: the fusion of some carpal and tarsbones; shortening of the long bones in fore- and hindlimb; amedial displacement of the patella in the knee joint (Stormal., 1994; Storm and Kingsley, 1996). GDF5 and its relatmolecules, GDF6 and 7, belong to the TGF-β superfamily, andare expressed specifically in the regions which give risejoints of the limb (Hatterley et al., 1995; Storm and Kingsle1996). Hence, these molecules are believed to be importandevelopment of proper joint structure. δEF1 is also expressedin developing elbow and carpals (Fig. 2L), and δEF1null(lacZ)
mutant mice actually showed defects of joint formatioresulting in the fused elbow and carpal bone malformationwould be interesting to see whether there is any functiointeraction between GDFs and δEF1.
Mhox, a paired-like homeobox-containing gene, is wideexpressed in the mesenchymal cells including the varioskeletal precursors (Kuratani et al., 1994). Mhox mutant micealso showed defects of skeletogenesis in multiple lineag

30
ofndasn
In
3-
t.
he
of
the
s
d
ne
is.
ed
T. Takagi and others
some of which are shared with the δEF1null(lacZ) mutant(Martin et al., 1995), i.e., the morphology of the maxillary anmandibular processes, and radius and tibia.
Mo et al. (1997) recently reported that Gli mutant mice alsoshowed multiple skeletal abnormalities including cleft palatooth defects, absence of vertebral body and intervertebdiscs, shortened limb and sternum. Among these, lareduction of intervertebral discs and delayed ossification digit bones observed in Gli2−/− mutants are reminiscent of theaffected morphology in these tissues of δEF1null(lacZ) mutantmice. The similarity among δEF1null(lacZ), Mhox and Glimutants may suggest genetic interactions among these ge
Domain-specific regulatory function of δEF1Previously, we generated δEF1∆C727 mutant mice (Higashi etal., 1997). Comparison of the phenotype of δEF1null(lacZ) andδEF1∆C727mutants revealed the unique function of the interndomain of δEF1 protein. Impairment of thymus developmenis shared by mutants of both alleles (Higashi et al., 1997 adata not shown), but other phenotypes described in this paare unique to the δEF1null(lacZ) allele. These results clearlyshow that the C-proximal portion of δEF1 protein including azinc finger cluster is essential for gene regulation leadingproper T cell development, while function of the remaininportion of δEF1 is required for normal skeleton developme(Fig. 9). Of particular interest is the homeodomain in thmiddle of the δEF1 protein since genetic interaction betweeδEF1 and Hox genes is suggested. Other δEF1 alleles with amutation in the homeodomain or the N-proximal zinc fingcluster are being introduced into the mouse. Phenotypesthese mutant mice will further distinguish them from onanother, which will further clarify a specific functional domaiof the δEF1 protein.
Evolutional conservation of the δEF1 functionδEF1 is thought to be a vertebrate homologue of Drosophilazfh-1 (Fortini et al., 1991; Funahashi et al., 1993) becausethe similarity of zinc-finger clusters and homeodomain, andtheir gene organization, although conservation of sequeother than the putative DNA binding domains is low. Not onis there similarity in the protein structure between δEF1 andzfh-1, but there is also similarity in their expression patternIn the Drosophilaembryo, expression of zfh-1is first observedin the presumptive procephalic mesoderm, then in tmesodermal anlages and later in the CNS including moneurons and in mesoderm-derived structures including admuscle precursors (Lai et al., 1991). δEF1 expression is firstdetected in the headfold, then at an early stage in mesodederivatives (e.g., somite, notochord, limb bud mesenchymand at a later stage strongly in CNS, PNS and muscle cells (2).
The phenotype of the zfh-1loss-of-function mutation (Lai etal., 1993) shows similarity to that of δEF1null(lacZ)mutant mice:early histogenesis of primitive mesodermal tissues and nervous system proceeds normally, but in later stagmispositioning and cell fate alteration of mesodermal tissubecome evident. These observations are consistent withidea that δEF1 is a structural and functional counterpart Drosophilazfh-1.
We thank Dr A. Nagy for ES R1, and Dr D. Duboule for Hoxd
d
te,ral
rgeof
nes.
altndper
tognten
er ofen
of ofncely
s.
hetorult
rmale),Fig.
theeses
theof
sequences. This work was supported by grants from the MinistryEducation, Science and Culture of Japan to T. T., Y. H. and H. K., afrom the Science and Technology Agency of Japan to Y. H. T. T. wthe recipient of a fellowship from the Japan Society for the Promotioof Science for Japanese Junior Scientists.
REFERENCES
Braun, T., Rudnicki, M. A., Arnold, H.-H. and Jaenisch, R.(1992). Targetedinactivation of the muscle regulatory gene Myf-5 results in abnormal ribdevelopment and perinatal death. Cell 71,369-382.
Bürglin, T. R. (1994). A comprehensive classification of homeobox genes. Guidebook to the Homeobox Genes(ed. D. Duboule), pp.25-71. Oxford:Oxford University Press
Cabanillas, A. M. and Darling, D. S.(1996). Alternative splicing gives rise totwo isoforms of Zfhep, a zinc finger/homeodomain protein that binds Tresponse element. DNA Cell Biol.15,643-651.
Cheah, K. S., Lau, E. T., Au, P. K. and Tam, P. P.(1991). Expression of themouse α1(II) collagengene is not restricted to cartilage during developmenDevelopment111,945-953.
Condie, B. G. and Capecchi, M. R.(1993). Mice homozygous for a targeteddisruption of Hoxd-3(Hox-4.1) exhibit anterior transformations of the first andsecond cervical vertebrae, the atlas and the axis. Development119,579-595.
Davis, A. P. and Capecchi, M. R.(1996). A mutational analysis of the 5′ HoxDgenes: dissection of genetic interactions during limb development in tmouse. Development122,1175-1185.
Davis, A. P., Witte, D. P., Hsieh-Li, H. M., Potter, S. S. and Capecchi, M. R.(1995). Absence of radius and ulna in mice lacking hoxa-11and hoxd-11.Nature375,791-795.
Dolle, P., Dierich, A., LeMeur, M., Schimmang, T., Schuhbaur, B.,Chambon, P. and Duboule, D.(1993). Disruption of the Hoxd-13geneinduces localized heterochrony leading to mice with neotenic limbs. Cell 75,431-441.
Dolle, P., Izpisúa-Belmonte, J. C., Falkenstein, H., Renucci, A. andDuboule, D. (1989). Coordinate expression of the murine Hox-5 complexhomeobox-containing genes during limb pattern formation. Nature342,767-772.
Duboule, D. (1994). Temporal coliniarity and the phylotypic progression: abasis for the stability of a vertebrate Bauplan and the evolution morphologies through heterochrony. Development120, Supplement 135-142.
Epstein, D. J., Vekemans, M. and Gros, P.(1991). splotch(Sp2H), a mutationaffecting development of the mouse neural tube, shows a deletion within paired homeodomain of Pax3. Cell 67,767-774.
Erlebacher, A., Filvaroff, E. H., Gitelman, S. E. and Derynck, R.(1995).Toward a molecular understanding of skeletal development. Cell 80, 371-378.
Favier, B., Rijli, F. M., Fromental-Ramain, C., Fraulob, V., Chambon, P.and Pascal, D.(1996). Functional cooperation between the non-paralogougenes Hoxa-10and Hoxd-11in the developing forelimb and axial skeleton.Development122,449-460.
Fortini, M. E., Lai, Z. and Rubin, G. M. (1991). The Drosophila zfh-1andzfh-2 genes encode novel proteins containing both zinc-finger anhomeodomain motifs. Mech. Dev.34,113-122.
Franklin, A. J., Jelton, T. L., Shelton, K. D. and Magnuson, M. A.(1994).BZP, a novel serum-responsive zinc finger protein that inhibits getranscription. Mol. Cell. Biol.14,6773-6788.
Fromental-Ramain, C., Warot, X., Messadecq, N., LeMeur, M., Pascal, D.and Chambon, P.(1996). Hoxa-13and Hoxd-13play a crucial role in thepatterning of the limb autopod. Development122,2997-3011.
Funahashi, J.-I., Kamachi, Y., Goto, K. and Kondoh, H. (1991).Identification of nuclear factor δEF1 and its binding site essential for lens-specific activity of the δ1-crystallin enhancer. Nucl. Acids Res.19, 3543-3547.
Funahashi, J.-I., Sekido, R., Murai, K., Kamachi, Y. and Kondoh, H.(1993). δ-crystallin enhancer binding protein δEF1 is a zinc fingerhomeodomain protein implicated in postgastrulation embryogenesDevelopment119,433-446.
Gendron-Maguire, M., Mallo, M., Zhang, M. and Gridley, T. (1993). Hoxa-2 mutant mice exhibit homeotic transformation of skeletal elements derivfrom cranial neural crest. Cell 75,1317-1331.
Genetta, T. and Kadesch, T.(1996). Cloning of a cDNA encoding a mouse

31Skeletal abnormalities in δEF1-knockout mice
by
ial
.
s.
s.
ytein
he
e
se
n.
transcriptional repressor displaying striking conservation across vertebrGene169,289-290.
Genetta, T., Ruezinsky, D. and Kadesch, T.(1994). Displacement of an E-box-binding repressor by basic helix-loop-helix proteins: Implication for cell specificity of the immunoglobulin heavy-chain enhancer. Mol. Cell. Biol.14,6153-6163.
Hasty, P., Bradley, A., Morris, J. H., Edmondson, D. G., Venuti, J. M.,Olson, E. N. and Klein, W. H.(1993). Muscle deficiency and neonatal deain mice with a targeted mutation in the myogeningene. Nature364,501-506.
Hatterley, G., Hewick, R. and Rosen, V.(1995). In situ localization and invitro activity of BMP-13. J. Bone Min. Res.10,S163.
Higashi, Y., Moribe, H., Takagi, T., Sekido, R., Kawakami, K., Kikutani, H.and Kondoh, H. (1997). Impairment of T cell development in δEF1 mutantmice. J. Exp. Med.185,1467-1480.
Hogan, B., Beddington, R., Constantini, F. and Lacy, E.(1994). Stainingembryos for cartilage and bone. In Manipulating the Mouse Embryo: aLaboratory Manual, pp.325-379. Cold Spring Harbor, New York: ColSpring Harbor Laboratory Press.
Horan, G. S., Wu, K., Wolgemuth, D. J. and Behringer, R. R.(1994).Homeotic transformation of cervical vertebrae in Hoxa-4mutant mice. Proc.Natl. Acad. Sci. USA91,12644-12648.
Ikeda, K. and Kawakami, K. (1995). DNA binding through distinct domainsof zinc-finger-homeodomain protein AREB6 has different effects on gtranscription. Eur. J. Biochem.233,73-82.
Jeannotte, L., Lemieux, M., Charron, J., Poirier, F. and Robertson, E. J.(1993). Specification of axial identity in the mouse: role of the Hoxa-5(Hox1.3) gene. Genes Dev.7,2085-2096.
Kamachi, Y. and Kondoh, H. (1993). Overlapping positive and negativregulatory elements determine lens-specific activity of the δ1-crystallinenhancer. Mol. Cell. Biol.13,5206-5215.
Kato, K., Takahashi, Y., Hayashi, S. and Kondoh, H.(1987). Improvedmammalian vectors for high expression of G418 resistance. Cell Struct.Func.12,575-580.
Kingsley, D., M., Bland, A. E., Grubber, J. M., Marker, P. C., Russell, L. B.,Copeland, N. C. and Jenkins, N. A.(1992). The mouse short ear skeletmorphogeneis locus is associated with defects in a bone morphogemember of the TGFβ superfamily. Cell 71,399-410.
Kuratani, S., Martin, J. F., Wawersik, S., Lilly, B., Eichele, G. and Olson, E.N. (1994). The expression pattern of the chick homeobox gene gMhoxsuggests a role in patterning of the limbs and face and compartmentalization of somites. Dev. Biol.161,357-369.
Lai, Z., Fortini, M. E. and Rubin, G. M. (1991). The embryonic expressionpatterns of zfh-1and zfh-2, twoDrosophilagenes encoding novel zinc-fingehomeodomain proteins. Mech. Dev.34,123-134.
Lai, Z.-C., Rushton, E., Bate, M. and Rubin, G. M.(1993). Loss of functionof the Drosophila zfh-1 gene results in abnormal development mesodermally derived tissues. Proc. Natl. Acad. Sci. USA90,4122-4126.
Mann, R. S. and Chan, S.-k.(1996). Extra specificity from extradenticle; thpartnership between HOX and PBX/EXD homeodomain proteins. TrendsGenet.12,258-262.
Martin, J. F., Bradley, A. and Olson, E. N.(1995). The paired-like homeo boxgene Mhoxis required for early events of skeletogenesis in multiple lineagGenes Dev.9,1237-1249.
Mo, R., Freer, A. M., Zinyk, D. L., Crackower, M. A., Michaud, J., Heng, H.H.-Q., Chik, X. W., Shi, X.-M., Tsui, L.-C., Cheng, S. H., Joyner, A. L. andHui, C.-c. (1997). Specific and redundant function of Gli2 and Gli3 zinc fingergenes in skeletal patterning and development. Development124,113-123.
Mortlock, D. P., Post, L. C. and Innis, J. W.(1996). The molecular basis ohypodactyly (Hd): a deletion in Hoxa13 leads to arrest of digital archformation. Nature Genet.13,284-289.
Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W. and Roder, J. C(1993). Derivation of completely cell culture-derived mice from earpassage embryonic stem cells. Proc. Natl. Acad. Sci. USA90,8424-8428.
ates.
B-
th
d
ene
e
alnetic
in
r
of
e
es.
f
.ly-
Postigo, A. A. and Dean, D. C.(1997). ZEB, a vertebrate homologue ofDrosophilaZfh-1, is a negative regulator of muscle differentiation. EMBO J.16,3935-3943.
Postigo, A. A., Sheppard, A. M., Mucenski, M. L. and Dean, D. C.(1997). c-Myb and Ets proteins synergize to overcome transcriptional repressionZEB. EMBO J.16,3924-3934.
Rijli, F. M., Mark, M., Lakkaraju, S., Dierich, A., Pascal, D. and Chambon,P. (1993). A homeotic transformation is generated in the rostral branchregion of the head by disruption of Hoxa-2, which acts as a selector gene. Cell75,1333-1349.
Rosen, G. D., Sanes, J. R., LaChance, R., Cunningham, J. M., Roman, Jand Dean, D. C.(1992). Roles for the integrin VLA-4 and its counterreceptor VCAM-1 in myogenesis. Cell 69,1107-19.
Satokata, I. and Maas, R.(1994). Msx1deficient mice exhibit cleft palate andabnormalities of craniofacial and tooth development. Nature Genet6, 349-355.
Sawai, S., Shimono, A., Hanaoka, K. and Kondoh, H.(1991). Embryoniclethality resulting from disruption of both N-myc alleles in mouse zygoteThe New Biologist3,861-869.
Sekido, R., Murai, K., Funahashi, J., Kamachi, Y., Fujisawa-Sehara, A.,Nabeshima, Y. and Kondoh, H.(1994). The δ-crystallin enhancer-bindingprotein δEF1 is a repressor of E2-box-mediated gene activation. Mol. Cell.Biol. 14,5692-5700.
Sekido, R., Takagi, T., Okanami, M., Moribe, H., Yamamura, M., Higashi,Y. and Kondoh, H. (1996). Organization of the gene encodingtranscriptional repressor δEF1 and cross-species conservation of its domainGene173,227-232.
Sekido, R., Murai, K., Kamachi, Y. and Kondoh, H. (1997). Twomechanisms in the action of repressor δEF1: binding site competition with anactivator and active repression. Genes to Cells 2 (in press).
Storm, E. E., Huyuh, T. Y., Copeland, N. G., Jenkins, N. A., Kingsley, D. M.and Lee, S.-L. (1994). Limb alterations in brachypodism mice due tomutations in a new menber of the TGF-beta super family. Nature368,639-643.
Storm, E. E. and Kingsley, D. M.(1996). Joint patterning defects caused bsingle and double mutations in members of the bone morphogenetic pro(BMP) family. Development122,3969-3979.
Suemori, H., Takahashi, N. and Noguchi, S.(1995). Hoxc-9 mutant miceshow anterior transformation of the vertebrae and malformation of tsternum and ribs. Mech. Dev.51,265-273.
Ueno, K., Hiramoto, Y., Hayashi, S. and Kondoh, H.(1987). Introductionand expression of recombinant β-galactosidase genes in cleavage stagmouse embryos. Dev. Growth Differ.30,61-73.
Wallin, J., Wilting, J., Koseki, H., Fritsch, R., Christ, B. and Balling, R.(1994). The role of Pax-1in axial skeleton development. EMBO J.120,1109-1121.
Watanabe, Y., Kawakami, K., Hirayama, Y. and Nagano, K.(1993).Transcription factors positively and negatively regulating the Na, K-ATPaα1 subunit gene. J. Biochem.114,849-855.
Wilkinson, D. G. (1992). Whole mount in situ hybridization of vertebrateembryos. In In Situ Hybridization: A Practical Approach, (ed. D. G.Wilkinson), pp.75-84. Oxford: IRL Press.
Williams, T. M., Moolten, D., Burlein, J., Romano, J., Bhaerman, R.,Godillot, A., Mellon, M., Rauscher III, F. J. and Kant, J. A. (1991).Identification of a zinc finger protein that inhibits IL-2 gene expressioScience254,1791-1794.
Yagi, T., Nada, S., Watanabe, N., Tamemoto, H., Kohmura, N., Ikawa, Y.and Aizawa, S. (1993). A novel negative selection for homologousrecombination using diphtheria toxin A fragment gene. Anal. Biochem.214,77-86.
Zhang, W., Behringer, R. R. and Olson, E. N.(1995). Inactivation of themyogenic bHLH gene MRF4 results in up-regulation of myogenin and ribanomalies. Genes Dev.9,1388-1399.
Copyright © 2022 FDOKUMEN