
Chronic difluoromethylornithine treatment impairs spatial learning and memory in rats
Upload
lmu-munichCategory
view
1download
0
DC
VA
*MFCN
Bcid(ccCtavcmcpdcciiraiIcrtcti
Mapma
GASTROENTEROLOGY 2009;137:1415–1424
eletion of TRPC4 and TRPC6 in Mice Impairs Smooth Muscleontraction and Intestinal Motility In Vivo
OLODYMYR V. TSVILOVSKYY,* ALEXANDER V. ZHOLOS,‡ THOMAS ABERLE,* STEPHAN E. PHILIPP,*LEXANDER DIETRICH,§ MICHAEL X. ZHU,� LUTZ BIRNBAUMER,¶ MARC FREICHEL,* and VEIT FLOCKERZI*
Experimentelle und Klinische Pharmakologie und Toxikologie, Universität des Saarlandes, Homburg, Germany; ‡Centre for Vision & Vascular Science, School ofedicine, Dentistry and Biomedical Sciences, Queens’s University Belfast, Belfast, United Kingdom; §Institut für Pharmakologie und Toxikologie, Medizinische
akultät, Universität Marburg, Marburg, Germany; �Department of Neuroscience, Biochemistry, and Center for Molecular Neurobiology, The Ohio State University,olumbus, Ohio; and ¶Signal Transduction and Neurobiology Laboratories, Division of Intramural Research, National Institute of Environmental Health Sciences,
ational Institutes of Health, Research Triangle Park, North CarolinaciTtCmAocaCd
dtmlcbtnoscuotmmtwct
Gpc
BA
SIC–
ALI
MEN
TARY
TRA
CT
See editorial on page 1211.
ACKGROUND & AIMS: Downstream effects of mus-arinic receptor stimulation in intestinal smooth musclenclude contraction and intestinal transit. We thought toetermine whether classic transient receptor potential
TRPC) channels integrate the intracellular signaling cas-ades evoked by the stimulated receptors and therebyontribute to the control of the membrane potential,a-influx, and cell responses. METHODS: We created
rpc4-, trpc6-, and trpc4/trpc6-gene– deficient mice and an-lyzed them for intestinal smooth muscle function initro and in vivo. RESULTS: In intestinal smooth muscleells TRPC4 forms a 55 pS cation channel and underliesore than 80% of the muscarinic receptor-induced cation
urrent (mICAT). The residual mICAT depends on the ex-ression of TRPC6, indicating that TRPC6 and TRPC4etermine mICAT channel activity independent of otherhannel subunits. In TRPC4-deficient ileal myocytes thearbachol-induced membrane depolarizations are dimin-shed greatly and the atropine-sensitive contraction elic-ted by acetylcholine release from excitatory motor neu-ons is reduced greatly. Additional deletion of TRPC6ggravates these effects. Intestinal transit is slowed downn mice lacking TRPC4 and TRPC6. CONCLUSIONS:n intestinal smooth muscle cells TRPC4 and TRPC6hannels are gated by muscarinic receptors and areesponsible for mICAT. They couple muscarinic recep-ors to depolarization of intestinal smooth muscleells and voltage-activated Ca2�-influx and contrac-ion, and thereby accelerate small intestinal motilityn vivo.
any visceral smooth muscles, including those ofthe gastrointestinal tract, typically co-express M2
nd M3 muscarinic receptor subtypes, which mediate thehysiological action of the parasympathetic neurotrans-itter acetylcholine in evoking smooth muscle excitation
nd contraction.1–3 Stimulation of muscarinic receptors
auses the opening of nonselective cationic channelsn smooth muscle cells of the gastrointestinal tract.1–9
hereby depolarization is produced and it is assumedhat these electrical events in turn result in increaseda2� influx via voltage-dependent Ca2� channels, smoothuscle contraction, and promotion of intestinal motility.ccordingly, it has been hypothesized that the generationf these muscarinic receptor–induced nonselective cationurrents (mICAT) is the mechanism coupling acetylcholinections to membrane depolarization, voltage-dependenta2� influx, and cell responses. However, decisive evi-ence on this mechanism still is lacking.Transient receptor potential (TRPC) channels are
ownstream effectors of G-protein– coupled recep-ors10,11 including muscarinic receptors, and in smooth
uscle cells these channels are widely assumed to under-ie nonselective cation currents.12,13 However, their pre-ise role in physiological processes often is uncertainecause channels bearing biophysical and regulatory fea-ures similar to the overexpressed TRPC channels haveot been identified unequivocally in native cells. The lackf appropriate channel blockers and agonists as well asuitable antibodies to identify TRPC proteins in primaryells has only escalated this problem. In this study wesed isolated cells and tissues prepared from several linesf TRPC knockout mice to define the proteins that formhe molecular basis of channel activity responsible for
ICAT and to analyze their impact on intestine smoothuscle contraction in vitro and in vivo. Our data show
hat channel activity depends on TRPC4 and TRPC6,hich contribute essentially to neurogenic cholinergic
ontraction of ileal smooth muscle strips and gastroin-estinal smooth muscle activity in vivo.
Abbreviations used in this paper: EFS, electrical field stimulation;ABA, �-aminobutyric acid; GTP�S, guanosine 5=-(3-thio) triphos-hate; I-V, current-voltage; mICAT, muscarinic receptor-induced cationurrent; TRPC, classic transient receptor potential.
© 2009 by the AGA Institute0016-5085/09/$36.00
doi:10.1053/j.gastro.2009.06.046

dtWCpbupta
paLp(
t
da
a
tips
CawutUwmac
Fctriknc
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1416 TSVILOVSKYY ET AL GASTROENTEROLOGY Vol. 137, No. 4
Materials and MethodsAnimal ModelsAll animal experiments were performed in accor-
ance with the Universität des Saarlandes Ethic Regula-ions and the animal welfare committees of the Saarland.
ild-type mice (129SvJ or F1 generation of 129SvJ and57Bl6/J), TRPC4�/� mice,14 TRPC6�/� mice,15 and com-ound TRPC4/TRPC6 double knockout mice obtainedy intercrossing of TRPC4�/� with TRPC6�/� mice weresed in this study. Methods used for genotyping (Sup-lementary Figure 1) and smooth muscle isolation fromhese mice are described in the Supplementary Materialsnd Methods section.
ElectrophysiologyStandard whole-cell patch-clamp recordings were
erformed using borosilicate patch pipettes (2–3 M�)16
nd an EPC-9 patch-clamp amplifier (HEKA, Elektronic,ambrecht, Germany). For single-channel recordingsatch pipettes (5–10 M�) were coated with Sylgard
World Precision Instruments, Sarasota, FL).
Contraction RecordingsLongitudinal layer muscle strips (5–7 mm) were
ied at both ends by silk suture, suspended from a force-
igure 1. TRPC4 and TRPC6 underlie mICAT in ileal smooth muscle cells isolated from ileum of wild-type (�/�, black), (A) TRPC4�/� (red), (Bime courses of mICAT evoked by carbachol (CCh, 100 �mol/L) applielationships (right panels) are shown. (D) Density of carbachol (left) andsolated from wild-type (black), TRPC4�/� (red), TRPC6�/� (blue), andnockout animals were significantly different from those in wild-type ceumber of cells. (E and F) TRPC4�, (E) TRPC4�, and (F) TRPC6 are
orresponding cells from TRPC-deficient mice. Loading controls: (E) CaV�3isplacement transducer (ADInstruments), and placed inn isolated tissue bath.
Small Intestine TransitSmall intestine transit essentially was performed
s described by Chang et al17 and Torjman et al.18
A full description of cell isolation techniques, solu-ions, and protocols used for electrophysiological exper-ments, contraction recordings, and intestinal transit isrovided in the Supplementary Materials and Methodsection.
Statistical AnalysisThe data were analyzed and plotted using Micro-
al Origin software (Micro-Cal Software, Inc., North-mpton, MA); for statistical tests GraphPad Prism soft-are (GraphPad Software, Inc, San Diego, CA) wassed. Values are given as the means � standard error ofhe mean; n represents the number of cells tested.nless otherwise noted, for comparison of 2 groupse used 2-tailed unpaired Student’s t tests and forore than 2 groups we used 1-way analysis of vari-
nce followed by the Dunnett’s test for multipleomparison.
uscarinic receptor–induced cation currents (mICAT) in smooth muscleC6�/� (blue), and (C) TRPC4�/�/TRPC6�/� mice (green). (A–C) Typical(arrowhead) recorded at �50 mV (left panels) and steady-state I-V
S-induced mICAT (right) at �50 mV and �50 mV in smooth muscle cells4�/�/TRPC6�/� mice (green). Current densities in cells isolated from� .01; ***P � .001), and differed between each other (P � .0085). n,
table in ileal myocyte lysates from wild-type mice (wt), but not in the
ells. M) TRP
cationGTP�TRPClls (**Pdetec
and (F) TRPC4 protein expression.

iwmvt(s�o
aripotddsoamgid
ipgT(rit
ecTc�(wwg1bwva
orbw1�dACcpTpnifTtif
icp2Utrtc1pTab
cdstvrasm(
2mptscwp
BA
SIC–
ALI
MEN
TARY
TRA
CT
October 2009 TRPC IN SMOOTH MUSCLE SIGNAL TRANSDUCTION 1417
ResultsMuscarinic Cation Currents (mICAT) inMurine Ileal Smooth Muscle CellsExternal application of carbachol (100 �mol/L) to
solated single smooth muscle myocytes caused an in-ard current (Figure 1A) with the typical features ofICAT characterized in guinea pig myocytes. Its current-
oltage (I-V) relationship was U-shaped at negative po-entials and the reversal potential (Erev) was close to 0 mVFigure 1 and Supplementary Figure 2). The mean den-ity of this current amounts to �11.8 � 0.4 pA/pF at50 mV and 11.0 � 1.0 pA/pF at �50 mV (n � 53 cells
btained from 14 mice) (Figure 1D, left).In guinea pig myocytes mICAT depends on synergistic
ctivation by Gi/o-coupled and Gq-coupled muscariniceceptors.19 –26 Likewise, in murine ileal cells mICAT isnhibited both after pertussis toxin pretreatment (Sup-lementary Figure 2B) and by increasing concentrationsf the phospholipase C inhibitor U73122 (Supplemen-ary Figure 2C and D). Phospholipase C inhibition re-uces formation of inositol 1,4,5-triphosphate andiacylglycerol while preventing the breakdown of pho-phatidylinositol 4,5-bisphosphate. Similar to previ-us studies25,26 we found that inositol 1,4,5-triphosphatend diacylglycerol seem not to play an important role inICAT regulation (Supplementary Figure 3) but similar to
uinea pig ileal myocytes24 agonist-activated mICAT isnhibited strongly by intracellularly applied phosphati-ylinositol 4,5-bisphosphate (Supplementary Figure 2E).
TRPC4�/TRPC4� Constitute Channels ThatAccount for the Greater Part of mICAT
These initial results indicate that the carbachol-nduced currents of murine ileal myocytes share the bio-hysical and regulatory properties with the mICAT ofuinea pig myocytes. They also resemble expressedRPC4 currents.24,27,28 The 2 TRPC4 isoforms TRPC4�
�100 kilodaltons) and TRPC4� (�93 kilodaltons)eadily are detectable in microsomal membranes fromleal myocytes isolated from wild-type mice but not inhose from TRPC4�/� mice (Figure 1E).
Compared with the wild-type myocytes, the carbachol-voked current was reduced considerably in TRPC4-defi-ient cells (Figure 1A). The resulting I-V obtained fromRPC4-deficient cells is almost linear (Figure 1A) and theurrent densities at �50 mV (�1.9 � 0.3 pA/pF) and50 mV (1.6 � 0.4 pA/pF; n � 10 cells) amount to 16.1%
�50 mV) and 15.3% (�50 mV) of the current densities inild-type cells (Figure 1D). Similar results were obtainedhen mICAT was activated by intracellular infusion ofuanosine triphosphate-�S (GTP�S) (200 �mol/L, FigureD, right) or the fluoroaluminate AlF4
� (data not shown),oth directly activate G proteins.29 After formation of thehole-cell configuration an inward current gradually de-
eloped in the presence of GTP�S that was indistinguish-
ble from carbachol-induced mICAT in terms of the shape Tf the I-V curve and maximal current amplitude, whicheached a maximum level within 3–10 minutes afterreakthrough. Current densities at �50 mV and �50 mVere �14.7 � 1.0 pA/pF and 13.7 � 1.0 pA/pF (FigureD, right) in wild-type cells (n � 24, from 4 animals) and2.3 � 0.2 pA/pF and 2.1 � 0.4 pA/pF in TRPC4-
eficient cells (n � 35, from 5 animals), respectively.lthough mICAT can be potentiated by an increase ina2�,30 the remaining current in TRPC4-deficient myo-
ytes, IC4�/�, no longer is affected when Ca2� in theipette solution is increased to 0.5 �mol/L (not shown).he whole-cell capacitances of wild-type cells (28.0 � 0.9F, n � 122) and TRPC4-deficient cells (29.2 � 0.9 pF,� 133) are not different (P � .38). In addition, carbachol-
nduced Ca2� release from intracellular stores is not af-ected by the lack of TRPC4 (wild-type, n � 10 cells,RPC4�/�, n � 14; Supplementary Figure 4), indicating
hat possible changes of phospholipase C activity andnositol 1,4,5-triphosphate formation could not accountor the greatly reduced mICAT.
Single-channel studies of outside-out patches excisedn the presence of carbachol revealed activity of 3 types ofationic channels with unitary conductances of 7.2 � 0.4S (n � 43), 55 � 8 pS (n � 14), and 116 � 14 pS (n �6) in ileal myocytes from wild-type mice (Figure 2).nitary currents through these channels reversed close to
he Cs� equilibrium potential at 0 mV. These valuesoughly correspond to the 10-, 57-, and 130-pS conduc-ances in guinea pig myocytes31 and the 20, 70, and 140onductances in murine myocytes.23,32 The activity of the16-pS channel (Figure 2C) was detected in 18 of 47atches and consisted of bursts of very brief openings.he longest open times were about some millisecondsnd this channel displayed only a very low open proba-ility (PO) (PO � .1; Figure 2C).
In contrast, the PO of the 7-pS and 55-pS conductancehannels (Figure 2A and B) strongly were potentialependent. At positive potentials their PO increasedtrongly with membrane depolarization, whereas at po-entials less than �60 mV the PO remarkably decreased,ery similarly to the carbachol-induced whole-cell cur-ent. However, the PO of the 7-pS conductance channellso was low (PO � .1); and together with its relativelymall unitary current amplitude this channel would
ake only a small contribution to the whole-cell currentFigure 2A).
The intermediate 55-pS conductance channel (FigureB) was present in 14 of 47 patches taken from wild-typeyocytes but a similar activity was not detectable in 27
atches from TRPC4-deficient myocytes, indicating thathe 55-pS channels are formed by TRPC4. In contrast, themall- and large-conductance channels were detectable inells of both genotypes at a similar frequency (7 pS:ild-type, 27 in 47 [57%]; TRPC4�/�, 17 in 27 [63%]; 116S: wild-type, 18 in 47 [38%]; TRPC4�/�, 8 in 27 [30%]).
aken together these data allow for the firm conclusion
tc(c
m(TtcT1mli(mamiam
rp.0ad
fppdbmmlvmDt0tts
ricAtts
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1418 TSVILOVSKYY ET AL GASTROENTEROLOGY Vol. 137, No. 4
hat TRPC4 underlies the intermediate-conductancehannels, which in turn account for the major part�80%) of the steady-state whole-cell I-V relationship,haracteristically U-shaped at negative potentials.31
TRPC6 Channels Contribute to mICAT
Another TRPC channel could account for the re-aining mICAT in TRPC4-deficient myocytes (mIC4�/�)
Figure 1A). In addition to TRPC4, both TRPC6 andRPC7 consistently were detectable by reverse-transcrip-
ion polymerase chain reaction in intestinal smooth mus-les33 (Supplementary Figure 5). By Western blotting theRPC6 protein was detectable in ileal myocytes (FigureF) as an approximately 95-kilodalton protein and higherolecular weight bands, which might represent glycosy-
ated versions34 of TRPC6 because they also are absentn the same type of cells from TRPC6 knock-out miceFigure 1F). Bath application of TRPC6 activator,35 the
embrane-permeable analogue of diacylglycerol, 1-oleoyl-2-cetyl-sn-glycerol (50 �mol/L), activates a current (Supple-entary Figure 6) indistinguishable from the carbachol-
nduced mIC4�/�, being almost linear at negative potentialsnd slightly outwardly rectifying at positive potentials (�50V, �1.5 � 0.2 pA/pF; �50 mV, 1.6 � 0.2 pA/pF; n � 13).In TRPC6�/� myocytes the characteristic U-shaped I-V
elationship of mICAT still was preserved (Figure 1B, rightanel), but currents were slightly but significantly (P �
002) smaller than in wild-type myocytes (�50 mV, �9.6 �.5 pA/pF; �50 mV, 7.8 � 0.7 pA/pF; n� 28 cells from 7nimals) (Figure 1B and D). The 7-pS and 116-pS con-
uctance channels still were detectable in the myocytes trom TRPC6-deficient mice (7 pS, 14 in 27 patches; 116S, 11 in 27 patches) as was the 55-pS channel (10 in 27atches), indicating that TRPC4 and TRPC6 underlieifferent channels, with TRPC4 channels being responsi-le for the remaining current in TRPC6-deficient cells,IC6�/�, whereas TRPC6 channels might underlieIC4�/�. To test this assumption we generated a mouse
ine deficient in both TRPC4 and TRPC6. These mice areiable and fertile, allowing us to isolate ileal smoothuscle cells to record mICAT. As shown in Figure 1C and, carbachol-induced currents hardly were detectable in
hese myocytes (�50 mV, �0.30 � 0.04 pA/pF; 50 mV,.26 � 0.06 pA/pF; n � 24 cells from 6 animals). Fromhese results we conclude that TRPC4 and TRPC6 pro-eins are the only essential components of channels re-ponsible for mICAT in ileal smooth muscle cells.
The Muscarinic Cation CurrentmICAT/mITRPC Couples Muscarinic ReceptorStimulation to Membrane Depolarization inIleal Smooth Muscle CellsTo assess the physiological function of mICAT we
ecorded membrane potentials in current clamp moden wild-type, TRPC4-deficient, and TRPC4-/TRPC6-defi-ient myocytes after application of carbachol (Figure 3).part from rarely occurring spontaneous hyperpolariza-
ions (�10% of cells) myocytes showed no electrical ac-ivity. The mean resting membrane potentials were notignificantly different in cells from the 3 genotypes (wild-
Figure 2. Single-channel cur-rents recorded from murine ilealmyocytes in the presence of car-bachol. Single-channel currentsof (A) 7-pS, (B) 55-pS, and(C) 116-pS conductance cationchannels recorded in the pres-ence of carbachol (100 �mol/L)in outside-out patches. Exam-ples of single-channel currenttraces (left panels), correspon-ding I-V relationships (middlepanels), and amplitude histo-grams of the records at �50 mV(right panels) are shown; the7-pS channel activity also is vis-ible in panel B.
ype cells, �58 � 1 mV, n � 33; TRPC4�/�, �55 � 1
m(cmtftclmi�mwomcstd�n�namPTm
lmtlTi
tcnpsdndmmFpau
FdT(p1osimTToir(mi1mta(ge(ffT
BA
SIC–
ALI
MEN
TARY
TRA
CT
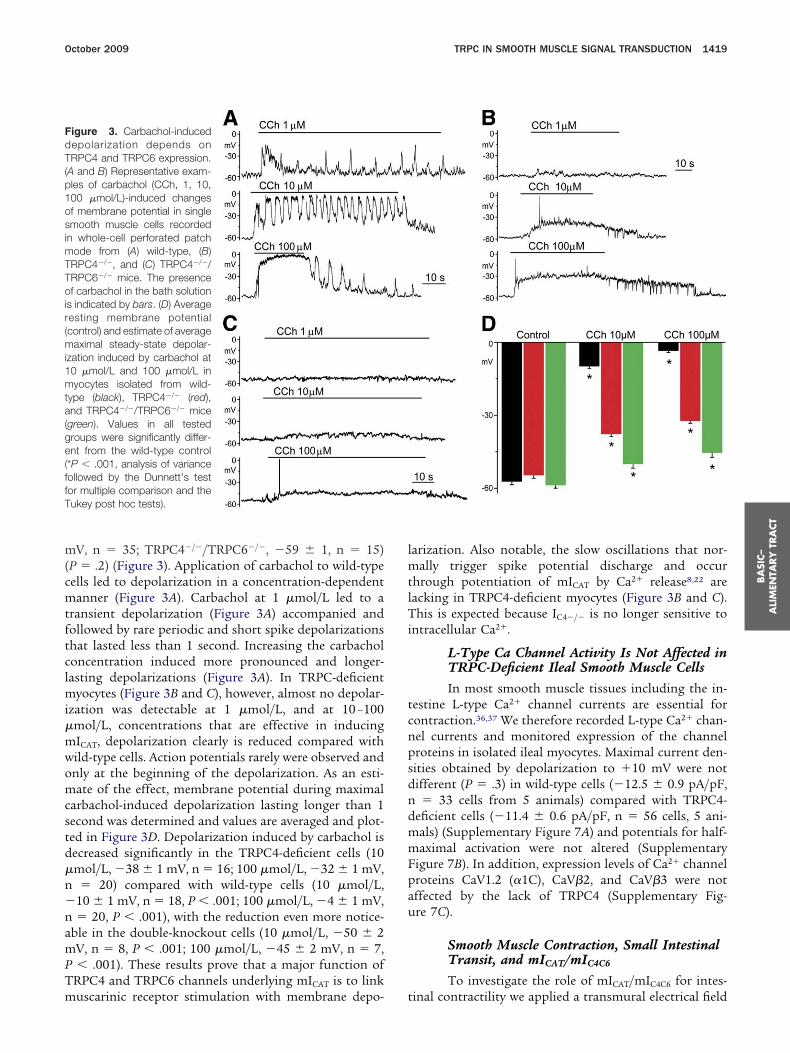
October 2009 TRPC IN SMOOTH MUSCLE SIGNAL TRANSDUCTION 1419
V, n � 35; TRPC4�/�/TRPC6�/�, �59 � 1, n � 15)P � .2) (Figure 3). Application of carbachol to wild-typeells led to depolarization in a concentration-dependentanner (Figure 3A). Carbachol at 1 �mol/L led to a
ransient depolarization (Figure 3A) accompanied andollowed by rare periodic and short spike depolarizationshat lasted less than 1 second. Increasing the carbacholoncentration induced more pronounced and longer-asting depolarizations (Figure 3A). In TRPC-deficient
yocytes (Figure 3B and C), however, almost no depolar-zation was detectable at 1 �mol/L, and at 10 –100mol/L, concentrations that are effective in inducingICAT, depolarization clearly is reduced compared withild-type cells. Action potentials rarely were observed andnly at the beginning of the depolarization. As an esti-ate of the effect, membrane potential during maximal
arbachol-induced depolarization lasting longer than 1econd was determined and values are averaged and plot-ed in Figure 3D. Depolarization induced by carbachol isecreased significantly in the TRPC4-deficient cells (10mol/L, �38 � 1 mV, n � 16; 100 �mol/L, �32 � 1 mV,� 20) compared with wild-type cells (10 �mol/L,
10 � 1 mV, n � 18, P � .001; 100 �mol/L, �4 � 1 mV,� 20, P � .001), with the reduction even more notice-
ble in the double-knockout cells (10 �mol/L, �50 � 2V, n � 8, P � .001; 100 �mol/L, �45 � 2 mV, n � 7,� .001). These results prove that a major function of
RPC4 and TRPC6 channels underlying mICAT is to link
igure 3. Carbachol-inducedepolarization depends onRPC4 and TRPC6 expression.
A and B) Representative exam-les of carbachol (CCh, 1, 10,00 �mol/L)-induced changesf membrane potential in singlemooth muscle cells recorded
n whole-cell perforated patchode from (A) wild-type, (B)RPC4�/�, and (C) TRPC4�/�/RPC6�/� mice. The presencef carbachol in the bath solution
s indicated by bars. (D) Averageesting membrane potentialcontrol) and estimate of average
aximal steady-state depolar-zation induced by carbachol at0 �mol/L and 100 �mol/L inyocytes isolated from wild-
ype (black), TRPC4�/� (red),nd TRPC4�/�/TRPC6�/� mice
green). Values in all testedroups were significantly differ-nt from the wild-type control
*P � .001, analysis of varianceollowed by the Dunnett’s testor multiple comparison and theukey post hoc tests).
uscarinic receptor stimulation with membrane depo- t
arization. Also notable, the slow oscillations that nor-ally trigger spike potential discharge and occur
hrough potentiation of mICAT by Ca2� release8,22 areacking in TRPC4-deficient myocytes (Figure 3B and C).his is expected because IC4�/� is no longer sensitive to
ntracellular Ca2�.
L-Type Ca Channel Activity Is Not Affected inTRPC-Deficient Ileal Smooth Muscle CellsIn most smooth muscle tissues including the in-
estine L-type Ca2� channel currents are essential forontraction.36,37 We therefore recorded L-type Ca2� chan-el currents and monitored expression of the channelroteins in isolated ileal myocytes. Maximal current den-ities obtained by depolarization to �10 mV were notifferent (P � .3) in wild-type cells (�12.5 � 0.9 pA/pF,� 33 cells from 5 animals) compared with TRPC4-
eficient cells (�11.4 � 0.6 pA/pF, n � 56 cells, 5 ani-als) (Supplementary Figure 7A) and potentials for half-aximal activation were not altered (Supplementary
igure 7B). In addition, expression levels of Ca2� channelroteins CaV1.2 (�1C), CaV�2, and CaV�3 were notffected by the lack of TRPC4 (Supplementary Fig-re 7C).
Smooth Muscle Contraction, Small IntestinalTransit, and mICAT/mIC4C6
To investigate the role of mICAT/mIC4C6 for intes-
inal contractility we applied a transmural electrical field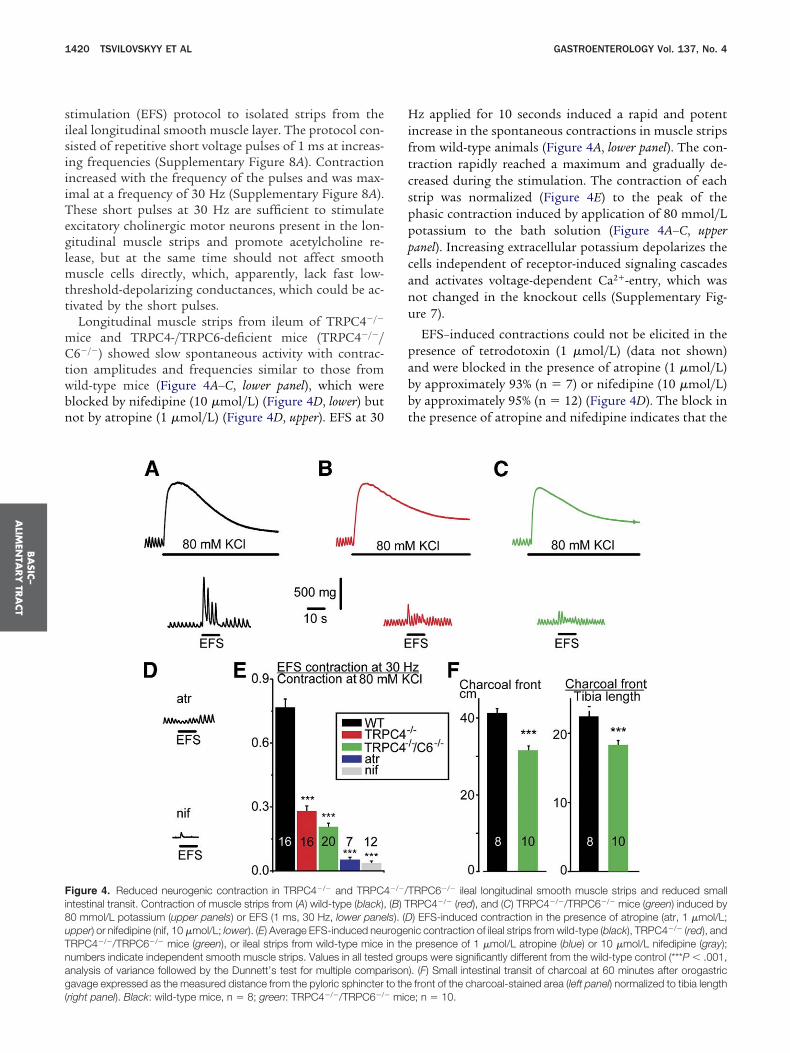
sisiiiTeglmtt
mCtwbn
Hiftcspppcanu
pabbt
Fi8uTnag(
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1420 TSVILOVSKYY ET AL GASTROENTEROLOGY Vol. 137, No. 4
timulation (EFS) protocol to isolated strips from theleal longitudinal smooth muscle layer. The protocol con-isted of repetitive short voltage pulses of 1 ms at increas-ng frequencies (Supplementary Figure 8A). Contractionncreased with the frequency of the pulses and was max-mal at a frequency of 30 Hz (Supplementary Figure 8A).hese short pulses at 30 Hz are sufficient to stimulatexcitatory cholinergic motor neurons present in the lon-itudinal muscle strips and promote acetylcholine re-ease, but at the same time should not affect smooth
uscle cells directly, which, apparently, lack fast low-hreshold-depolarizing conductances, which could be ac-ivated by the short pulses.
Longitudinal muscle strips from ileum of TRPC4�/�
ice and TRPC4-/TRPC6-deficient mice (TRPC4�/�/6�/�) showed slow spontaneous activity with contrac-
ion amplitudes and frequencies similar to those fromild-type mice (Figure 4A–C, lower panel), which werelocked by nifedipine (10 �mol/L) (Figure 4D, lower) butot by atropine (1 �mol/L) (Figure 4D, upper). EFS at 30
igure 4. Reduced neurogenic contraction in TRPC4�/� and TRPC4ntestinal transit. Contraction of muscle strips from (A) wild-type (black),0 mmol/L potassium (upper panels) or EFS (1 ms, 30 Hz, lower panepper) or nifedipine (nif, 10 �mol/L; lower). (E) Average EFS-induced neuRPC4�/�/TRPC6�/� mice (green), or ileal strips from wild-type miceumbers indicate independent smooth muscle strips. Values in all testenalysis of variance followed by the Dunnett’s test for multiple compaavage expressed as the measured distance from the pyloric sphincter
right panel). Black: wild-type mice, n � 8; green: TRPC4�/�/TRPC6�/� mic
z applied for 10 seconds induced a rapid and potentncrease in the spontaneous contractions in muscle stripsrom wild-type animals (Figure 4A, lower panel). The con-raction rapidly reached a maximum and gradually de-reased during the stimulation. The contraction of eachtrip was normalized (Figure 4E) to the peak of thehasic contraction induced by application of 80 mmol/Lotassium to the bath solution (Figure 4A–C, upperanel). Increasing extracellular potassium depolarizes theells independent of receptor-induced signaling cascadesnd activates voltage-dependent Ca2�-entry, which wasot changed in the knockout cells (Supplementary Fig-re 7).
EFS–induced contractions could not be elicited in theresence of tetrodotoxin (1 �mol/L) (data not shown)nd were blocked in the presence of atropine (1 �mol/L)y approximately 93% (n � 7) or nifedipine (10 �mol/L)y approximately 95% (n � 12) (Figure 4D). The block inhe presence of atropine and nifedipine indicates that the
RPC6�/� ileal longitudinal smooth muscle strips and reduced smallRPC4�/� (red), and (C) TRPC4�/�/TRPC6�/� mice (green) induced by) EFS-induced contraction in the presence of atropine (atr, 1 �mol/L;ic contraction of ileal strips from wild-type (black), TRPC4�/� (red), andpresence of 1 �mol/L atropine (blue) or 10 �mol/L nifedipine (gray);
ups were significantly different from the wild-type control (***P � .001,). (F) Small intestinal transit of charcoal at 60 minutes after orogastricfront of the charcoal-stained area (left panel) normalized to tibia length
�/�/T(B) Tls). (Drogenin thed grorisonto the
e; n � 10.
ivcosaTa(
cTi8taatTb
livmodsw1(aiTti
FscTcTasTddn
tati
vtioTTscGccwis
F(cT((aocoC
BA
SIC–
ALI
MEN
TARY
TRA
CT
October 2009 TRPC IN SMOOTH MUSCLE SIGNAL TRANSDUCTION 1421
ncreased contraction involves muscarinic receptors andoltage-activated dihydropyridine-sensitive L-type Ca2�
hannels most probably located in the plasma membranef the smooth muscle cells whereas tetrodotoxin acts onodium channels. Compared with wild-type (n �16) themplitude of EFS-induced contraction was reduced inRPC4�/� strips by approximately 64% (Figure 4B and E)nd in TRPC4�/�/C6�/� strips by approximately 72%Figure 4C and E).
A similar reduction was observed when EFS-inducedontraction of muscle strips from TRPC4- and TRPC4/RPC6-deficient mice were monitored in the presence of
nhibitors of neurotransmission (Supplementary FigureB and the Supplementary Materials and Methods sec-ion), which might have been co-released together withcetylcholine during EFS or formed as a consequence ofcetylcholine release, supporting the conclusion that ofhe EFS-induced contraction which is reduced in theRPC4- and TRPC4/TRPC6-deficient mice is mediatedy acetylcholine.
To determine whether this impairment of acetylcho-ine-induced contraction translates into changes of smallntestine motility in vivo, 0.25 mL of charcoal (10% wt/ol) were applied to mice by orogastric gavage. After 60inutes mice were killed for exposure and examination
f the gastrointestinal tract and transit of charcoal wasetermined as the measured distance from the pyloricphincter to the leading edge of the charcoal-stained areaithin the intestine (Figure 4F) and amounted to 41.3 �.2 cm (wild-type mice; n � 8) and 31.6 � 1.1 cmTRPC4�/�/C6�/�-mice; n � 10) (P � .000025). A similarnd significant reduction also was observed after normal-zing these values to tibia length (wild-type, 2.25 � 0.07;RPC4�/�/C6�/�, 1.83 � 0.06) (P � .00046), showing
hat TRPC4/TRPC6 channels are important in promot-ng intestinal motility in vivo.
DiscussionIn a first series of experiments (Supplementary
igure 2) we show that mICAT in mouse ileal myocyteshares the regulatory and biophysical properties of theurrent in guinea pig cells. We then show that TRPC4�,RPC4�, and TRPC6 proteins are expressed in ileal myo-ytes and that mICAT is no longer detectable in TRPC4-/RPC6-deficient myocytes. Resting membrane potentialnd spontaneous contractile activity of longitudinalmooth muscle fibers are not affected by the lack ofRPC4 and TRPC6, whereas muscarinic receptor–in-uced depolarizations and contractile responses are re-uced significantly and, as a consequence, small intesti-al transit of charcoal is slowed down in vivo.Members of the TRPC subfamily have been implicated
o constitute cationic channels in smooth muscle and bynalyzing genetically modified mice it recently was shownhat TRPC6 channels underlie hypoxia-induced cation
nflux in pulmonary smooth muscle cells38 and are in- *olved in the regulation of vascular smooth muscleone,15 whereas pressure-induced and store-operated cat-on influx in vascular smooth muscle,39 similar to store-perated cation influx in platelets,40 is independent ofRPC1. Genetic inactivation of another TRPC gene,RPC4, resulted in impaired endothelium-dependent va-
orelaxation and loss of a Ca2�-selective store-operatedurrent,14 changes in microvascular permeability,41 andABA release from thalamic interneurons.42 TRPC4
hannels also have been implicated in carbachol-inducedationic currents in stomach smooth muscle cells, buthether the TRPC4 protein is expressed in these cells and
ts impact on smooth muscle contraction has not beenhown.43
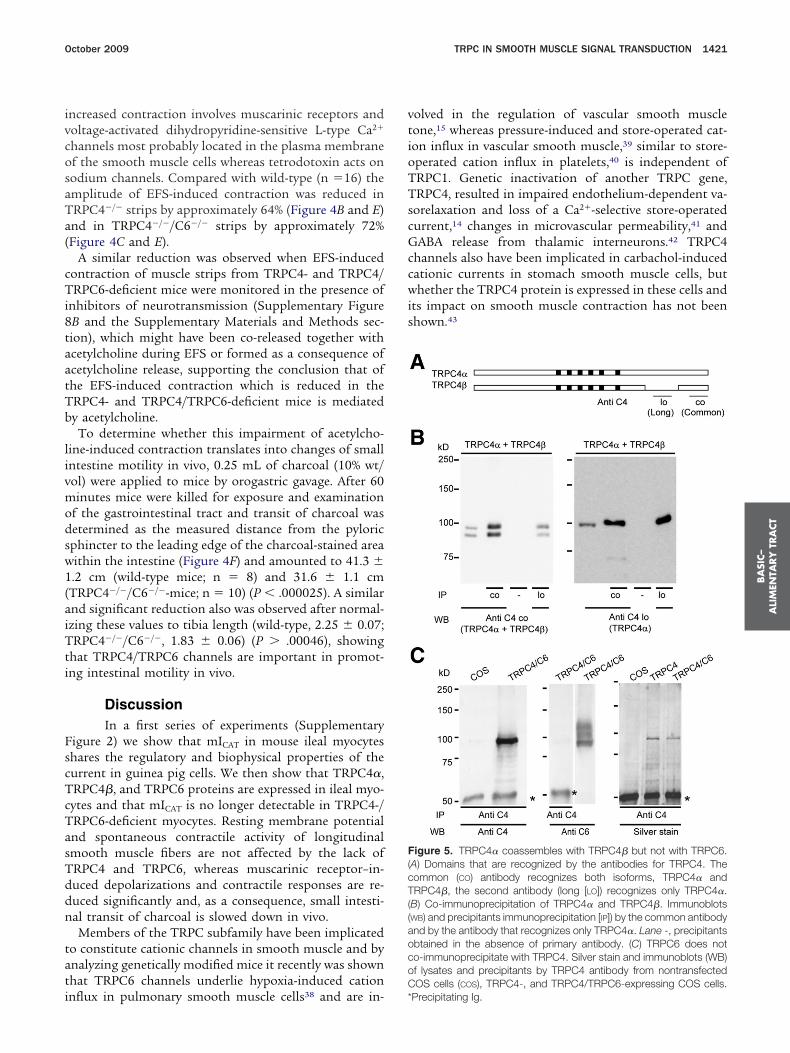
igure 5. TRPC4� coassembles with TRPC4� but not with TRPC6.A) Domains that are recognized by the antibodies for TRPC4. Theommon (CO) antibody recognizes both isoforms, TRPC4� andRPC4�, the second antibody (long [LO]) recognizes only TRPC4�.
B) Co-immunoprecipitation of TRPC4� and TRPC4�. ImmunoblotsWB) and precipitants immunoprecipitation [IP]) by the common antibodynd by the antibody that recognizes only TRPC4�. Lane -, precipitantsbtained in the absence of primary antibody. (C) TRPC6 does noto-immunoprecipitate with TRPC4. Silver stain and immunoblots (WB)f lysates and precipitants by TRPC4 antibody from nontransfectedOS cells (COS), TRPC4-, and TRPC4/TRPC6-expressing COS cells.
Precipitating Ig.

1tbomiabactt
rmpectnD8capc4Tmrc�2Tc
td[tMpmTs
cdcTacar
it
ftprmaTbTr
aG1
Fcb
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1422 TSVILOVSKYY ET AL GASTROENTEROLOGY Vol. 137, No. 4
We show here that 2 TRPC4 proteins of approximately00 kilodaltons (TRPC4�) and approximately 93 kilodal-ons (TRPC4�) are expressed in murine ileal myocytesut are absent in the same cell type from TRPC4 knock-ut mice; they also are co-expressed in brain44 and inacrovascular endothelium.14 When the longer TRPC4�
s precipitated, the TRPC4� is retained (Figure 5A and B),lthough it is not recognized by the precipitating anti-ody (Figure 5A). This result is in line with the co-ssembly of TRPC4� and TRPC4� to TRPC4�/TRPC4�hannels, whereas there is no evidence of direct interac-ion of TRPC4 and TRPC6, even after overexpressingheir complementary DNAs in COS7 cells (Figure 5C).
In ileal myocytes stimulation of M2 and M3 muscariniceceptors provide concurrent, but different, signals for
ICAT channel opening,19,45,46 which appear to requireertussis toxin–sensitive Gi/o-proteins, phospholipase Cnzymatic activity, and an increase in [Ca2�]i, which in-reases the channel’s open probability (for discussion ofhe Ca2�-dependence of mICAT and ion selectivity of chan-els underlying mICAT see the Supplementary materials).eletion of the TRPC4 gene in mice reduced mICAT by
4%. In addition, the 55-pS channel recorded on carba-hol stimulation in wild-type cells was no longer detect-ble in the TRPC4 knockout cells, indicating that TRPC4roteins are essential components of the responsiblehannel. This 55-pS conductance channel resembles the2-pS and 57-pS conductance channels recorded fromRPC4 expressing HEK 293 cells47 and guinea pig ilealyocytes31; their mean patch I-V relationships closely
esemble the muscarinic whole-cell current in ileal myo-ytes, showing the reduced PO at potentials of less than60 mV and a reversal potential close to 0 mV (Figure
B). For a discussion of single-channel conductance ofRPC4 in HEK 293 cells and in ileum smooth muscleells see the Supplementary materials.
TRPC4 channel activity in HEK cells depends on in-racellular Ca2�24,27 and it is this aspect of TRPC4 mo-ulation that may explain concurrent oscillations of
Ca2�]i, mICAT, and membrane potential observed in wild-ype cells but not in TRPC4-deficient myocytes (Figure 3).
embrane potential oscillations, often preceded by spikeotentials, normally occur through potentiation byICAT by Ca2� release events,48,49 but are absent inRPC4-deficient myocytes because IC4�/� is no longer
ensitive to intracellular Ca2�.The contribution of TRPC6 to mICAT is significant but
onsiderably smaller than that of TRPC4. In myocyteseficient in both genes no mICAT could be evoked byarbachol, indicating that the remaining currents inRPC6-deficient myocytes (IC6�/�) are TRPC4 currentsnd that the remaining currents in TRPC4-deficient myo-ytes (IC4�/�) are TRPC6 currents (ITRPC6). ITRPC6, can bectivated in the presence of 1-oleoyl-2-acetyl-sn-glycerol,
esembling the remaining 1-oleoyl-2-acetyl-sn-glycerol–nduced cationic current observed in pertussis toxin–reated mouse ileal cells.50
In summary, TRPC4 and TRPC6 proteins are essentialor muscarinic receptor–induced channel activities in thisype of smooth muscle independent of the other channelroteins. By integrating downstream effects of musca-inic receptor stimulation they are pivotal in coupling
uscarinic receptors to membrane potential, Ca2� influx,nd cell responses (Figure 6). The decisive role of TRPC4/RPC6 shown here to occur in smooth muscle also coulde relevant in other cell types including neurons, whereRPC channels also should respond to metabotropic
eceptors and cause membrane depolarization.
Supplementary Data
Note: To access the supplementary materialccompanying this article, visit the online version ofastroenterology at www.gastrojournal.org, and at doi:0.1053/j.gastro.2009.06.046.
References
1. Kuriyama H, Kitamura K, Itoh T, et al. Physiological features ofvisceral smooth muscle cells, with special reference to receptorsand ion channels. Physiol Rev 1998;78:811–920.
2. Bolton TB, Prestwich SA, Zholos AV, et al. Excitation-contractioncoupling in gastrointestinal and other smooth muscles. Annu RevPhysiol 1999;61:85–115.
3. Sanders KM. Invited review: mechanisms of calcium handling insmooth muscles. J Appl Physiol 2001;91:1438–1449.
4. Bolton TB. The depolarizing action of acetylcholine or carbachol inintestinal smooth muscle. J Physiol 1972;220:647–671.
5. Klevets MY, Shuba MF. Mechanisms of adrenaline, noradrenalineand acetylcholine effects on the electrophysiological propertiesof smooth muscle cells. 92-107. Transactions of the 2nd Sym-posium on General Physiology: Synaptic Processes [in Russian].Kiev: Naukova Dumka, 1968.
6. Inoue R, Isenberg G. Acetylcholine activates nonselective cationchannels in guinea pig ileum through a G protein. Am J Physiol1990;258:C1173–C1178.
7. Wang YX, Fleischmann BK, Kotlikoff MI. M2 receptor activation ofnonselective cation channels in smooth muscle cells: calcium
igure 6. In ileal smooth muscle TRPC4 and TRPC6 (TRPC) channelsouple the extracellular chemical signal acetylcholine (AGONIST) to mem-rane depolarization, Ca2�-influx, and cell responses.
and Gi/G(o) requirements. Am J Physiol 1997;273:C500–C508.

1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
BA
SIC–
ALI
MEN
TARY
TRA
CT
October 2009 TRPC IN SMOOTH MUSCLE SIGNAL TRANSDUCTION 1423
8. Unno T, Kwon SC, Okamoto H, et al. Receptor signaling mecha-nisms underlying muscarinic agonist-evoked contraction in guinea-pig ileal longitudinal smooth muscle. Br J Pharmacol 2003;139:337–350.
9. So I, Kim KW. Nonselective cation channels activated by thestimulation of muscarinic receptors in mammalian gastricsmooth muscle. J Smooth Muscle Res 2003;39:231–247.
0. Ramsey IS, Delling M, Clapham DE. An introduction to TRP chan-nels. Annu Rev Physiol 2006;68:619–647.
1. Venkatachalam K, Montell C. TRP channels. Annu Rev Biochem2007;76:387–417.
2. Beech DJ, Muraki K, Flemming R. Non-selective cationic channelsof smooth muscle and the mammalian homologues of DrosophilaTRP. J Physiol 2004;559:685–706.
3. Inoue R, Okada T, Onoue H, et al. The transient receptor potentialprotein homologue TRP6 is the essential component of vascularalpha(1)-adrenoceptor-activated Ca(2�)-permeable cation chan-nel. Circ Res 2001;88:325–332.
4. Freichel M, Suh SH, Pfeifer A, et al. Lack of an endothelialstore-operated Ca2� current impairs agonist-dependent vasore-laxation in TRP4�/� mice. Nat Cell Biol 2001;3:121–127.
5. Dietrich A, Mederos YS, Gollasch M, et al. Increased vascularsmooth muscle contractility in TRPC6�/� mice. Mol Cell Biol2005;25:6980–6989.
6. Hamill OP, Marty A, Neher E, et al. Improved patch-clamp tech-niques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch 1981;391:85–100.
7. Chang FY, Doong ML, Chen TS, et al. Vasoactive intestinalpolypeptide appears to be one of the mediators in misoprostol-enhanced small intestinal transit in rats. J Gastroenterol Hepatol2000;15:1120–1124.
8. Torjman MC, Joseph JI, Munsick C, et al. Effects of isoflurane ongastrointestinal motility after brief exposure in rats. Int J Pharm2005;294:65–71.
9. Zholos AV, Bolton TB. Muscarinic receptor subtypes controllingthe cationic current in guinea-pig ileal smooth muscle. Br J Phar-macol 1997;122:885–893.
0. Rhee JC, Rhee PL, Park MK, et al. Muscarinic receptors control-ling the carbachol-activated nonselective cationic current inguinea pig gastric smooth muscle cells. Jpn J Pharmacol 2000;82:331–337.
1. Okamoto H, Prestwich SA, Asai S, et al. Muscarinic agonistpotencies at three different effector systems linked to the M(2) orM(3) receptor in longitudinal smooth muscle of guinea-pig smallintestine. Br J Pharmacol 2002;135:1765–1775.
2. Unno T, Matsuyama H, Komori S. Interaction between the M2-and M3-receptor subtypes in muscarinic electrical and mechani-cal responses of intestinal smooth muscles. Neurophysiology2003;35[3/4]:290–301.
3. Sakamoto T, Unno T, Kitazawa T, et al. Three distinct muscarinicsignalling pathways for cationic channel activation in mouse gutsmooth muscle cells. J Physiol 2007;582:41–61.
4. Otsuguro K, Tang J, Tang Y, et al. Isoform-specific inhibition ofTRPC4 channel by phosphatidylinositol 4,5-bisphosphate. J BiolChem 2008;283:10026–10036.
5. Zholos AV, Tsytsyura YD, Gordienko DV, et al. Phospholipase C,but not InsP3 or DAG, -dependent activation of the muscarinicreceptor-operated cation current in guinea-pig ileal smooth mus-cle cells. Br J Pharmacol 2004;141:23–36.
6. Okamoto H, Unno T, Arima D, et al. Phospholipase C involvementin activation of the muscarinic receptor-operated cationic currentin Guinea pig ileal smooth muscle cells. J Pharmacol Sci 2004;95:203–213.
7. Plant TD, Schaefer M. TRPC4 and TRPC5: receptor-operatedCa2�-permeable nonselective cation channels. Cell Calcium
2003;33:441–450.8. Strubing C, Krapivinsky G, Krapivinsky L, et al. TRPC1 and TRPC5form a novel cation channel in mammalian brain. Neuron 2001;29:645–655.
9. Bigay J, Deterre P, Pfister C, et al. Fluoroaluminates activatetransducin-GDP by mimicking the gamma-phosphate of GTP in itsbinding site. FEBS Lett 1985;191:181–185.
0. Inoue R, Isenberg G. Intracellular calcium ions modulate acetyl-choline-induced inward current in guinea-pig ileum. J Physiol1990;424:73–92.
1. Zholos AV, Zholos AA, Bolton TB. G-protein-gated TRP-like cat-ionic channel activated by muscarinic receptors: effect of poten-tial on single-channel gating. J Gen Physiol 2004;123:581–598.
2. Dresviannikov AV, Bolton TB, Zholos AV. Muscarinic receptor-activated cationic channels in murine ileal myocytes. Br J Phar-macol 2006;149:179–187.
3. Walker RL, Hume JR, Horowitz B. Differential expression andalternative splicing of TRP channel genes in smooth muscles.Am J Physiol Cell Physiol 2001;280:C1184–C1192.
4. Dietrich A, Schnitzler M, Emmel J, et al. N-linked protein glyco-sylation is a major determinant for basal TRPC3 and TRPC6channel activity. J Biol Chem 2003;278:47842–47852.
5. Hofmann T, Obukhov AG, Schaefer M, et al. Direct activation ofhuman TRPC6 and TRPC3 channels by diacylglycerol. Nature1999;397:259–263.
6. Bolton TB. Mechanisms of action of transmitters and other sub-stances on smooth muscle. Physiol Rev 1979;59:606–718.
7. Wegener JW, Schulla V, Koller A, et al. Control of intestinalmotility by the Cav1.2 L-type calcium channel in mice. FASEB J2006.
8. Weissmann N, Dietrich A, Fuchs B, et al. Classical transientreceptor potential channel 6 (TRPC6) is essential for hypoxicpulmonary vasoconstriction and alveolar gas exchange. Proc NatlAcad Sci U S A 2006;103:19093–19098.
9. Dietrich A, Kalwa H, Storch U, et al. Pressure-induced and store-operated cation influx in vascular smooth muscle cells is inde-pendent of TRPC1. Pflugers Arch 2007;20:1260–1262.
0. Varga-Szabo D, Authi KS, Braun A, et al. Store-operated Ca(2�)entry in platelets occurs independently of transient receptor po-tential (TRP) C1. Pflugers Arch 2008;455:465–477.
1. Tiruppathi C, Freichel M, Vogel SM, et al. Impairment of store-operated Ca2� entry in TRPC4(�/�) mice interferes with in-crease in lung microvascular permeability. Circ Res 2002;91:70–76.
2. Munsch T, Freichel M, Flockerzi V, et al. Contribution of transientreceptor potential channels to the control of GABA release fromdendrites. Proc Natl Acad Sci U S A 2003;100:16065–16070.
3. Lee KP, Jun JY, Chang IY, et al. TRPC4 is an essential componentof the nonselective cation channel activated by muscarinic stim-ulation in mouse visceral smooth muscle cells. Mol Cell 2005;20:435–441.
4. Flockerzi V, Jung C, Aberle T, et al. Specific detection and semi-quantitative analysis of TRPC4 protein expression by antibodies.Pflugers Arch 2005;451:81–86.
5. Komori S, Kawai M, Takewaki T, et al. GTP-binding protein in-volvement in membrane currents evoked by carbachol and hista-mine in guinea-pig ileal muscle. J Physiol 1992;450:105–126.
6. Yan HD, Okamoto H, Unno T, et al. Effects of G-protein-specificantibodies and G beta gamma subunits on the muscarinic recep-tor-operated cation current in guinea-pig ileal smooth musclecells. Br J Pharmacol 2003;139:605–615.
7. Schaefer M, Plant TD, Obukhov AG, et al. Receptor-mediatedregulation of the nonselective cation channels TRPC4 andTRPC5. J Biol Chem 2000;275:17517–17526.
8. Unno T, Inaba T, Ohashi H, et al. Role of Ca2� mobilizationin muscarinic receptor-mediated membrane depolarization inguinea pig ileal smooth muscle cells. Jpn J Pharmacol 2000;84:
431–437.
4
5
R
EUH1
A
BM
Sb
VXcptL
C
F
NHHFVF
BA
SIC–
ALIM
ENTA
RY
TRA
CT
1424 TSVILOVSKYY ET AL GASTROENTEROLOGY Vol. 137, No. 4
9. Kohda M, Komori S, Unno T, et al. Carbachol-induced oscillationsin membrane potential and [Ca2�]i in guinea-pig ileal smoothmuscle cells. J Physiol 1998;511:559–571.
0. Sakamoto T, Unno T, Matsuyama H, et al. Characterization ofmuscarinic receptor-mediated cationic currents in longitudinalsmooth muscle cells of mouse small intestine. J Pharmacol Sci2006;100:215–226.
Received November 23, 2008. Accepted June 18, 2009.
eprint requestsAddress requests for reprints to: Marc Freichel, MD,
xperimentelle und Klinische Pharmakologie und Toxikologie,niversität des Saarlandes, Kirrberger Str. Gebäude 46 66421omburg, Germany. e-mail: [email protected]; fax: (49) 6841-626402.
cknowledgmentsThe authors thank Christine Wesely, Kerstin Fischer, Stefanie
uchholz, and Katrin Schumacher for their valued assistance;
arcel Meissner and Adolfo Cavalié for helpful discussions; and Cabine Pelvay, Ramona Gölzer, and Martin Jung for immunizing andleeding rabbits.Volodymyr V. Tsvilovskyy, Alexander V. Zholos, Marc Freichel, and
eit Flockerzi contributed to all aspects of the manuscript, Michael. Zhu contributed to acquisition of data from HEK 293 cells andritical revision of the manuscript, Thomas Aberle contributed torotein chemistry, Stephan E. Philipp contributed to reverse-ranscription polymerase chain reaction; and Alexander Dietrich andutz Birnbaumer provided TRPC6-deficient mice.
onflicts of interestThe authors disclose no conflicts.
undingSupported in part by the Intramural Research Programs of the
ational Institutes of Health, National Institute of Environmentalealth Sciences (Z01-ES-101684) to (L.B.), by National Institutes ofealth grant DK081654 (M.X.Z. and A.V.Z.), by Homburgerorschungsförderung sprogramm (HOMFOR) (M.F., V.V.T., S.E.P., and.F.), by Deutsche Forschungsgemeinschaft andorschungskommission der UdS (M.F. and V.F.), and by Fonds der
hemischen Industrie (V.F.).Copyright © 2022 FDOKUMEN




















