
Anisotropic contraction in forisomes: Simple models won't fit
11
Anisotropic Contraction in Forisomes: Simple Models Won’t Fit Winfried S. Peters, 1 * Michael Knoblauch, 2 Stephen A. Warmann, 3 William F. Pickard, 4 and Amy Q. Shen 3 1 Department of Biology, Indiana University/Purdue University Fort Wayne, Fort Wayne, Indiana 2 School of Biological Science, Washington State University, Pullman, Washington 3 Department of Mechanical and Aerospace Engineering, Washington University, St. Louis, Missouri 4 Department of Electrical and Systems Engineering, Washington University, St. Louis, Missouri Forisomes are ATP-independent, Ca 21 -driven contractile protein bodies act- ing as reversible valves in the phloem of plants of the legume family. Fori- some contraction is anisotropic, as shrinkage in length is associated with radial expansion and vice versa. To test the hypothesis that changes in length and width are causally related, we monitored Ca 21 - and pH-dependent defor- mations in the exceptionally large forisomes of Canavalia gladiata by high- speed photography, and computed time-courses of derived geometric parame- ters (including volume and surface area). Soybean forisomes, which in the resting state resemble those of Canavalia geometrically but have less than 2% of the volume, were also studied to identify size effects. Calcium induced sixfold volume increases in forisomes of both species; in soybean, responses were completed in 0.15 s, compared to about 0.5 s required for a rapid response in Canavalia followed by slow swelling for several minutes. This size-dependent behavior supports the idea that forisome contractility might rest on similar mechanisms as those of polyelectrolyte gels, a class of artificial ‘‘smart’’ materials. In both species, time-courses of forisome length and diameter were variable and lacked correlation, arguing against a simple causal relationship between changes in length and width. Moreover, changes in the geometry of soybean forisomes differed qualitatively between Ca 21 - and pH-responses, suggesting that divalent cations and protons target differ- ent sites on the forisome proteins. Cell Motil. Cytoskeleton 65: 368–378, 2008. ' 2008 Wiley-Liss, Inc. Key words: Ca 21 -dependent contractility; contractile protein; contraction kinetics; pH-dependent contractility; Canavalia gladiata [Jacq.] DC.; Glycine max L. *Correspondence to: Winfried S. Peters, Indiana/Purdue University Fort Wayne, 2101 East Coliseum Boulevard, Fort Wayne, Indiana 46805-1499, USA. E-mail: [email protected] Contract grant sponsor: Indiana/Purdue University Fort Wayne; Con- tract grant sponsor: Federal Ministry of Education and Research, Ger- many; Contract grant number: 0312014A; Contract grant sponsor: National Science Foundation’s Sensors and Sensor Networks pro- gram; Contract grant number: 0510941. Received 31 July 2007; Accepted 15 January 2008 Published online 10 March 2008 in Wiley InterScience (www. interscience.wiley.com). DOI: 10.1002/cm.20266 ' 2008 Wiley-Liss, Inc. Cell Motility and the Cytoskeleton 65: 368–378 (2008)
Transcript of Anisotropic contraction in forisomes: Simple models won't fit

Anisotropic Contraction in Forisomes: SimpleModels Won’t Fit
Winfried S. Peters,1* Michael Knoblauch,2 Stephen A. Warmann,3
William F. Pickard,4 and Amy Q. Shen3
1Department of Biology, Indiana University/Purdue University Fort Wayne,Fort Wayne, Indiana
2School of Biological Science, Washington State University, Pullman,Washington
3Department of Mechanical and Aerospace Engineering,Washington University, St. Louis, Missouri
4Department of Electrical and Systems Engineering, Washington University,St. Louis, Missouri
Forisomes are ATP-independent, Ca21-driven contractile protein bodies act-ing as reversible valves in the phloem of plants of the legume family. Fori-some contraction is anisotropic, as shrinkage in length is associated withradial expansion and vice versa. To test the hypothesis that changes in lengthand width are causally related, we monitored Ca21- and pH-dependent defor-mations in the exceptionally large forisomes of Canavalia gladiata by high-speed photography, and computed time-courses of derived geometric parame-ters (including volume and surface area). Soybean forisomes, which in theresting state resemble those of Canavalia geometrically but have less than2% of the volume, were also studied to identify size effects. Calciuminduced sixfold volume increases in forisomes of both species; in soybean,responses were completed in 0.15 s, compared to about 0.5 s required for arapid response in Canavalia followed by slow swelling for several minutes.This size-dependent behavior supports the idea that forisome contractilitymight rest on similar mechanisms as those of polyelectrolyte gels, a class ofartificial ‘‘smart’’ materials. In both species, time-courses of forisome lengthand diameter were variable and lacked correlation, arguing against a simplecausal relationship between changes in length and width. Moreover, changesin the geometry of soybean forisomes differed qualitatively between Ca21-and pH-responses, suggesting that divalent cations and protons target differ-ent sites on the forisome proteins. Cell Motil. Cytoskeleton 65: 368–378,2008. ' 2008 Wiley-Liss, Inc.
Key words: Ca21-dependent contractility; contractile protein; contraction kinetics; pH-dependent
contractility; Canavalia gladiata [Jacq.] DC.; Glycine max L.
*Correspondence to: Winfried S. Peters, Indiana/Purdue University
Fort Wayne, 2101 East Coliseum Boulevard, Fort Wayne, Indiana
46805-1499, USA. E-mail: [email protected]
Contract grant sponsor: Indiana/Purdue University Fort Wayne; Con-
tract grant sponsor: Federal Ministry of Education and Research, Ger-
many; Contract grant number: 0312014A; Contract grant sponsor:
National Science Foundation’s Sensors and Sensor Networks pro-
gram; Contract grant number: 0510941.
Received 31 July 2007; Accepted 15 January 2008
Published online 10 March 2008 in Wiley InterScience (www.
interscience.wiley.com).
DOI: 10.1002/cm.20266
' 2008 Wiley-Liss, Inc.
Cell Motility and the Cytoskeleton 65: 368–378 (2008)

INTRODUCTION
In higher plants, the products of photosynthesis aredistributed by pressure-driven mass flow in the continu-ous sieve tube network of the phloem, a natural micro-fluidics system [Evert, 1982; Behnke and Sjolund, 1990;Sjolund, 1997]. Sieve tubes are formed by specializedcells, the sieve elements (SE), which during differentia-tion loose their cytoskeleton while developing a multi-tude of specific proteins of unknown function. Straßbur-ger [1891] described peculiar protein bodies in the sievetubes of legumes. While these ‘‘nondispersive P-proteinbodies’’ [Behnke, 1991] were subjected to detailed ultra-structural studies, it took over a century before theirATP-independent contractility [Knoblauch et al., 2003]and function as sieve tube valves [Knoblauch et al.,2001] were discovered. To underline their unique prop-erties and function, we now refer to these structures asforisomes (‘‘gate bodies’’).
Forisomes represent a proteinaceous ‘‘smart mate-rial’’ [Mavroidis and Dubey, 2003; Knoblauch andPeters, 2004a] which consists of fibrils that in the restingstate are coaligned with the main body’s longitudinalaxis [Palevitz and Newcomb, 1971; Lawton, 1978]. Cal-cium ion concentrations above a threshold of about 50lM [Knoblauch et al., 2005] induce a reversible aniso-tropic deformation: a longitudinal shortening with simul-taneous radial expansion result in a drastic volumeincrease [Knoblauch et al., 2003]. Optical properties offorisomes suggest that a reversible self-assembly of or-dered fibril arrays occurs during the contraction/expan-sion cycle [Peters et al., 2007b]. Nonphysiologicallyhigh and low pH values trigger parts of the Ca21
response [for review, see Knoblauch and Peters, 2004b].While we had discovered forisome dynamics first inVicia faba, we recently established Canavalia gladiataas a new model system since it possesses exceptionallylarge forisomes (>40 lm length) with two tail-like pro-trusions which facilitate manipulation of isolated speci-mens [Peters et al., 2007a]. In this species, the Ca21-response consists of a rapid initial phase followed byslow swelling over a period of about 2 min, whereas thechelator-induced reverse reaction is complete in lessthan 5 s. However, available data did not allow fordetailed kinematic analysis due to a lack of temporalresolution [Peters et al., 2007a].
Spasmonemes are motor-organelles of sessile cili-ates that resemble forisomes as their contractility isATP-independent and Ca21-driven [Amos et al., 1976].However, the proteins forming spasmonemes on onehand and forisomes on the other are not related [Macie-jewski et al., 1999; Noll et al., 2007; and own unpub-lished results]. Furthermore, differences in the contract-ile mechanisms are indicated by the geometric isotropyof spasmoneme contraction, where a longitudinal short-
ening is accompanied by a proportional decrease inwidth [Moriyama et al., 1999]. In forisomes, the anisot-ropy of the Ca21-response can become lost while con-tractility as such still persists; we had found that speci-mens which had lost their elongate shape during isola-tion from living cells were capable of isotropic but notanisotropic swelling and shrinking [Knoblauch et al.,2003]. This suggests that the molecules mediating Ca21-dependent contractility might be active regardless oftheir supramolecular arrangement, but that in intact fori-somes, they may be interlinked in ways that enforce thecoupling of contraction along one axis with a concurrentperpendicular expansion. In such an assembly, Ca21 ionsmight drive radial expansion only, with a passive longi-tudinal shortening being a mere consequence—or viceversa. If so, Ca21-dependent changes in forisome shapeshould follow constant temporal patterns, with causallyrelated sequences of shrinkage and expansion in lengthand diameter, respectively.
We examined the dynamic geometry of Canavaliaforisomes during contraction/expansion cycles inducedby Ca21 or pH steps by high-speed video recording fol-lowed by single-frame analysis. Our geometric analysesutilized the highly versatile quantitative 3D model[Peters et al., 2007a] that we had derived from an olderdescriptive model of soybean forisomes [Wergin andNewcomb, 1970]. Soybean (Glycine max) forisomes,which have less than 2% of the Canavalia forisome vol-ume, were studied to identify size effects.
MATERIALS AND METHODS
Plant Material and Forisome Preparation
Soybean (Glycine max L. ‘‘Williams 82’’; KennethGerloff, New Florence, MO) and sword bean (Canavaliagladiata [Jacq.] DC ‘‘Akanata Mame’’; Kitazawa Seed,Oakland, CA) were greenhouse grown, Glycine in pottingmix in 10 cm pots and Canavalia in potting mix supple-mented with bark bits in 35 cm pots. Plants were main-tained on a 14 h day length under sunlight supplementedby metal halide illumination, watered twice daily, andfertilized once a week. Temperature was kept between 25and 308C (day) or between 23 and 308C (night).
Canavalia samples were collected from plants sev-eral months old as segments of maturing runners 3–4mm in diameter. Glycine samples were juvenile stemsegments collected at 3–5 weeks after germination. Inboth species, the peripheral stem tissues were peeled offand were bathed in Ca21-free medium (10 mMNa2EDTA, 100 mM KCl, 10 mM HEPES pH 7.3) for15 min. Then, the phloem tissue was scratched off of theinner face of the peels, incubated in Ca21-free mediumfor 10 min, and ground in liquid nitrogen with pestle and
Forisome Contraction Kinetics 369

mortar. The resulting frozen tissue powder was thawedin Ca21-free medium and passed through a 60 lm meshfilter, to yield a crude forisome preparation.
Microscopy
A drop of a crude forisome preparation was posi-tioned on a microscope slide. The Ca21-free mediumwas exchanged just before the experiment by low saltCa21-free medium (10 lM Na2EDTA, 100 lM KCl, 10lM HEPES pH 7.3) to prevent a protraction of the accessof Ca21 by an excess of EDTA. Our light microscope(Leica DM IRB, Leica, Wetzlar, Germany) wasequipped with Leica HCX APO water immersion objec-tives. Forisomes were manipulated using glass micropip-ettes (pulled from borosilicate capillaries, 1 mm O.D.)mounted on a micromanipulator (MX-1, Narashige,Tokyo, Japan) as described previously [Peters et al.,2007a]. To test Ca21-responses, Na2EDTA in the Ca21-free medium was replaced by 10 mM CaCl2. To studyeffects of increased pH, HEPES in the Ca21-free me-dium was replaced by 10 mM CAPS and the pH wasadjusted to 10.5. Media were exchanged by shooting thenew solution onto the slide with a micropipette, whileremoving medium continuously from the slide with aPasteur pipette connected to a vacuum pump.
Data Acquisition and Analysis
Responses were monitored with a Photron Fastcam1280 PCI camera controlled by the Fastcam viewer soft-ware v.2.4.3.2 (Photron, San Diego, CA) to producehigh-speed movies of 2–4 s length at 1000 or 2000frames per second. Depending on the velocity of theresponse captured, forisome geometry was analyzed onframes at intervals of 3, 4, 6, or 8 milliseconds. Length(L) of the forisome main body, its diameter in the center(D), and diameter at the tip (d) were measured using thefreeware ImageJ (v 1.33u, http://rsb.info.nih.gov/ij/). Anextended version of the flexible ‘‘step-stool’’ function[Peters and Baskin, 2006] that, in its full 9-coefficientversion, allowed for two linear portions within the transi-tion, was fitted to the data. Fitting was done first man-ually, then automatically, and individual coefficientswere eliminated depending on the characteristics of thedataset given (for a detailed description of these proce-dures, see [Peters and Baskin, 2006]). In cases in whichthe data showed more complex trends, overlapping poly-nomials of varying degree were fitted to portions of thedataset. No inherent biological meaning was assigned toa function’s coefficients. The curves fitted served amerely descriptive purpose in facilitating the determina-tion of derived geometric parameters which were calcu-lated according to our previously derived model [Peterset al., 2007a]. To express forisome volume (V), forisomemain body surface area (S), equatorial circumference of
the forisome main body (CFE; the circumference meas-ured around the center of the main body), and polar cir-cumference of the forisome main body (CFP; the circum-ference measured across the tips of the forisome mainbody) in terms of the measured parameters D, d, and Lonly, we define for clarity
r ¼ 1
4
L2
D� dþ D� d
� �
a ¼ r arcsinL
2 r
� �
b ¼ 1
2
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffir2 � L2
4
r
and arrive at
V ¼ 0:93L D2 � 4 r Dþ 8 r2 � L2
3
�
þð4D� 8rÞ bþ a r
L
� �i
S ¼ 0:907LD
2� r þ bþ a r
L
� �
CFE ¼ 3:63D
CFP ¼ 4aþ 2d
We wish to stress that the complexity of this modeldoes not reflect specific assumptions regarding the geom-etry of one particular type of forisome. Rather, it is themathematical price, as it were, one has to pay for thehigh degree of flexibility that allows for an accuratequantitative description of all forisome shapes we haveobserved so far. This flexibility has the desirable conse-quence that the relative errors of the estimates of geo-metric parameters remain similar at all stages of fori-some deformation such as those described below.
As a characteristic measure of the response veloc-ity in individual forisomes, the intervals between thetimes at which the rapid volume increase or decrease hadreached one and nine tenths of its final value (10–90%response times) were calculated.
All numerical results given in the text are mean6 SD.
RESULTS
Kinetics of Rapid Ca21- and pH-Responsesin Canavalia Forisomes
We studied the Ca21-induced deformation of fori-somes from Canavalia by high-speed photography. Fori-some length (L), central diameter (D), and tip diameter(d) were measured on individual frames taken at 1000
370 Peters et al.
frames per second, and derived geometric parameterswere calculated from these measurements. Most Cana-valia forisomes appear elongate box-shaped in side viewand essentially retain this shape during the Ca21-response [Peters et al., 2007a]. Figures 1A and 1B dem-onstrate an extreme case of forisome ‘‘rounding’’; werarely observed a Canavalia forisome approaching aspherical shape as closely as shown here. However, thevariability in shape had little impact on the accuracy of
our estimates of forisome volume, surface area, circum-ferences, and their time-courses, because the model usedto calculate these derived parameters describes all shapesobserved equally well [Peters et al., 2007a].
The short-term time-courses of D, d, and L showedgreat variability, of which the three examples in Fig. 2are on the whole representative. In these graphs, time 0corresponds to the first frame in which the forisome waswell enough focused following Ca21-addition to allowunambiguous measurements; the addition of Ca21
actually occurred between 0.6 and 1.4 s prior to thattime. The lengths of these phase lags between Ca21-application and the first visible reaction probably repre-sent the time required for the Ca21-medium to reach theforisome rather than any inherent phase lags. On theother hand, the kinetics of the responses after their initia-tion were fully resolved. The response curves appearedsigmoidal with smooth (e.g. L in Fig. 2E) or abrupt (e.g.L in Fig. 2A) changes in slope, but no general patternbecame evident. The starts of the responses in the threeparameters followed no particular sequence. The longitu-dinal contraction could require a shorter (Fig. 2A), a sim-ilar (Fig. 2C), or a longer length of time than the radialexpansion (Fig. 2E). The radial expansion could proceedin parallel at the forisome tip and center (Fig. 2A) or not(Figs. 2C and 2E); obviously, the degree of curvature ofthe longitudinal forisome surfaces changed in the lattercase, but not in the former. Curves fitted to the datasetswere used to compute time-courses of forisome mainbody volume (Figs. 2B, 2D, and 2F). Although our high-speed movies were too short to include the slow Ca21-induced swelling documented in long-term observations[Peters et al., 2007a], the drastic decrease in the rate ofswelling over time let most short-term time-courses ofvolume appear as if the complete response had been cap-tured (Figs. 2B, 2D, and 2F). In this situation we found ituseful to determine the apparent 10–90% response timeof volume as a measure of the velocity of the rapidresponse to Ca21; on average, the reaction took 0.52 60.23 s (n5 8; compare Figs. 2B, 2D, and 2F).
In the responses to replacing calcium by Ca21-freemedium, it became even more obvious that the onsets ofaxial and radial length changes were independent ofeach other (Fig. 3). The start of longitudinal extensioncoincided with (Fig. 3A), lagged behind (Fig. 3C), orpreceded (Fig. 3E) that of radial contraction. Frequently,the time-courses appeared to be polyphasic with two lin-ear portions (e.g. L and d in Fig. 3C), but no general pat-tern was apparent. In agreement with the conclusiondrawn from long-term studies [Peters et al., 2007a], theoverall reaction to Ca21-removal appeared to proceedfaster than the Ca21-driven volume increase, as the appa-rent volumetric 10–90% response time was 0.41 60.16 s (n 5 6; compare Figs. 3B, 3D, and 3F). Notably,
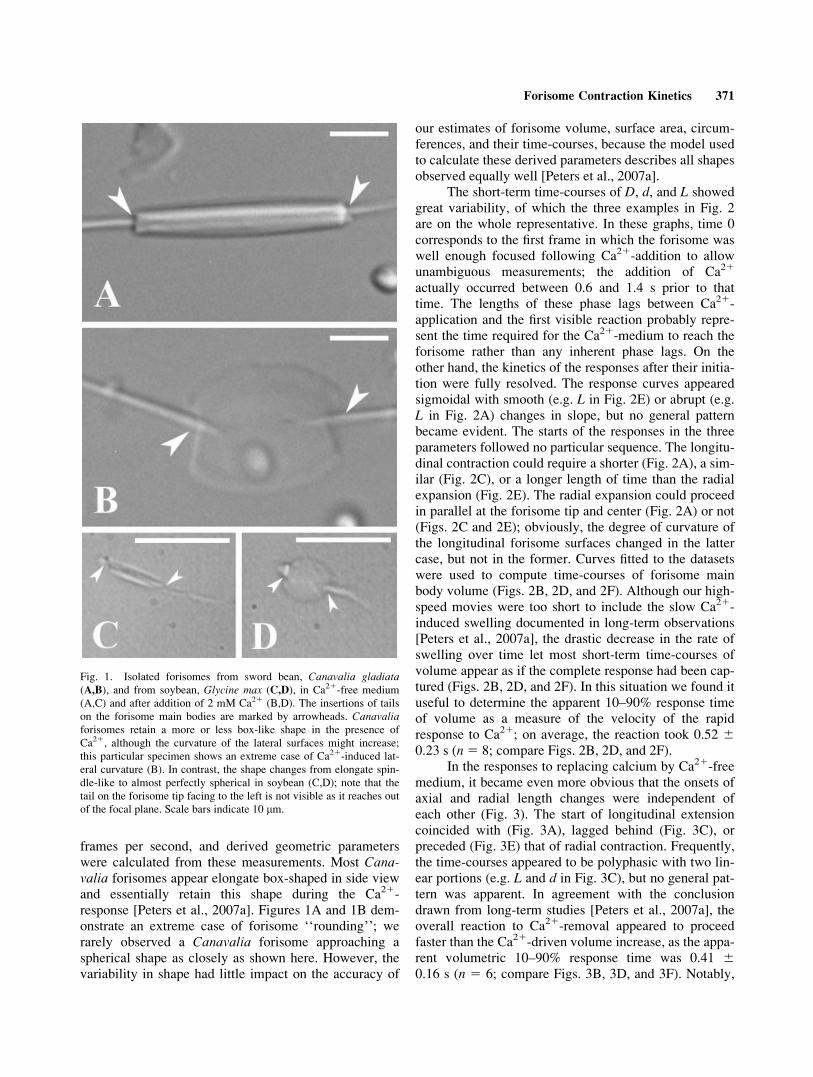
Fig. 1. Isolated forisomes from sword bean, Canavalia gladiata(A,B), and from soybean, Glycine max (C,D), in Ca21-free medium
(A,C) and after addition of 2 mM Ca21 (B,D). The insertions of tails
on the forisome main bodies are marked by arrowheads. Canavaliaforisomes retain a more or less box-like shape in the presence of
Ca21, although the curvature of the lateral surfaces might increase;
this particular specimen shows an extreme case of Ca21-induced lat-
eral curvature (B). In contrast, the shape changes from elongate spin-
dle-like to almost perfectly spherical in soybean (C,D); note that the
tail on the forisome tip facing to the left is not visible as it reaches out
of the focal plane. Scale bars indicate 10 lm.
Forisome Contraction Kinetics 371

the variability of the responses to Ca21 or chelators wasnot based on different properties of individual forisomes;if repeated reaction cycles were evoked in one forisome(not shown), the consecutive time-courses of changes inlength and width showed no higher degree of similaritybetween them than that observed between time-coursesfrom different forisomes.
On average, forisome volume increased threefoldduring the Ca21-responses monitored by high-speedphotography (from 900 6 270 lm3 to 2950 6 390 lm3;n 5 8). During the chelator response, on the other hand,which was induced in forisomes that had been equili-brated in Ca21-medium for at least 3 min, the volumedecreased sixfold, from 5310 6 1090 lm3 to 860 6 140lm3 (n 5 6). Thus, the volume of forisomes in the Ca21-expanded state was significantly greater at the start of
the Ca21-depletion tests than at the end of the high-speedmovies showing the Ca21-response. This had to beexpected because of the retarded swelling documented inlong-term experiments [Peters et al., 2007a].
During the short-term responses, time-courses ofthe surface area of the forisome main body and of itsequatorial circumference (Fig. 4) roughly paralleledthose of volume (Figs. 2 and 3). In contrast, polar cir-cumference showed only slight changes (Fig. 4). Thegreatest cross-sectional area of the forisomes is physio-logically significant because it determines the proportionof the sieve tube’s cross-sectional area that can be occu-pied by the forisome plug. It changed by factors between5 and 15 during the responses analyzed (not shown), inagreement with our previous analysis of long-termresponses [Peters et al., 2007a].
Fig. 2. Kinetics of the response to 10 mM Ca21 of three representa-
tive forisomes isolated from Canavalia gladiata. Graphs in the upper
row (A,C,E) show measurements of forisome main body length (L;&), central diameter (D; l), and tip diameter (d; *) taken on each
6th frame of movies produced at a frame rate of 1000 s-1. A versatile
function allowing for poly-phasic sigmoidal transitions was fitted to
the original data (black or white lines in A,C,E), from which continu-
ous time-courses of the volume of the forisome main body were com-
puted (B,D,F). Shaded areas in B, D, and F indicate the 10–90%
response times of forisome volume. In all graphs, time 0 corresponds
to the first frame in which L, D, and d could be determined unambigu-
ously following transient movements of the forisomes out of the
microscope’s focal plane due to turbulences caused by the injection of
the new medium at 0.6–1.4 s before time 0.
372 Peters et al.

Forisomes from Vicia faba swell in response tounphysiologically high pH [Knoblauch et al., 2003].Canavalia forisomes treated with a Ca21-free incubationmedium of pH 10.5 reacted analogously and underwentan anisotropic deformation (Fig. 5A). In contrast to theCa21-responses, the time-courses of L, D, and d showedlengthened periods of change with lower and occasion-ally wobbling rates of change (Fig. 5B), resulting inincreased 10–90% response times of volume (rangingfrom 0.68 to 1.01 s in the three specimens analyzed).The polar circumference of the forisome main bodyremained constant during the pH-response (Fig. 5B) as itdid in the Ca21-reaction (Fig. 4).
Ca21- and pH-Responses in Glycine Forisomes
Isolated soybean forisomes, which have only about2% of the volume of Canavalia forisomes, showed allthe typical reactions. In contrast to Canavalia forisomes,
they always changed their main body side-view appear-ance from elongate spindle-like to spherical upon Ca21-induced longitudinal contraction (Fig. 1). Owing to theirsmall size, we found it impossible to mount soybean fori-somes on micropipettes to verify that their cross-sec-tional shape is a rounded square (for head-view imagesof Canavalia forisomes, see [Peters et al., 2007a]). How-ever, forisomes that were visibly rotating around theirlongitudinal axis during medium exchange in the setupfor high-speed photography, showed regular oscillationsof their central diameter, D (Fig. 6A), in agreement withthe assumption of a square cross-section.
While the tip diameter, d, did not change inresponse to the addition or removal of Ca21 in soybeanforisomes (tip diameter d, in this case, was taken to bethe diameter of the tail at the point of insertion), thetime-courses of L and D exhibited similar variability asin Canavalia. The responses showed biphasic kinetics
Fig. 3. Kinetics of the response to the removal of Ca21 of three rep-
resentative Canavalia gladiata forisomes. Upper row graphs (A,C,E)give forisome main body length (L; &), central diameter (D; l), and
tip diameter (d; *) as measured on each 6th frame of movies taken at
1000 frames per second. Functions fitted to the original data (black or
white lines in A,C,E) were used to compute continuous time-courses
of forisome main body volume (B,D,F). Shaded areas in B, D, and F
show the 10–90% response times of forisome volume. For definition
of time 0, see Fig. 2.
Forisome Contraction Kinetics 373

(e.g. D in Fig. 6G) or not (D in Fig. 6A), and the changesof L and D appeared well correlated over time in somecases (e.g. Fig. 6G) but not in others (e.g. Figs. 6C and6E). The overall responses proceeded three to four timesfaster in soybean; the apparent 10–90% response timesof volume were identical for the Ca21-addition (0.14 60.02 s, n 5 4) and the Ca21-removal reaction (0.13 60.06 s, n 5 6). Though we did not study the long-termkinetics of the Ca21-response in soybean, a comparisonof forisome volumes at the start and end of the high-speed movies provided no evidence for a retarded, slowswelling analogous to that observed in Canavalia. Insoybean, forisome volumes differed between the start ofCa21-addition tests and the end of Ca21-removal experi-ments (10.6 6 0.6 lm3, n 5 4 [group A], compared to12.3 6 1.5, n 5 6 [group B]). This difference reflects ac-cidental variation in average size between the two groupsof forisomes. The volumes of the same groups also dif-fered between the end of high-speed movies showing theCa21-response (when forisomes had been exposed to
Ca21 for not more than 1 s; 56.1 6 9.1 lm3, n 5 4[group A]) and the start of clips showing the reaction toCa21-removal (when forisomes had been preincubatedin Ca21-solution for 3 min; 69.7 6 9.8 lm3, n 5 6[group B]). If the latter difference were significantlygreater than the former, the data would indicate that a re-tarded swelling, similar to that described for Canavalia,had occurred in these soybean forisomes. We normalizedthe data to compare the differences and found theminsignificantly different (P 5 0.39 in the two-tailedt-test). Thus the differences of forisome volumeobserved between the two relatively small groups of fourand six specimens probably are due to statistical happen-stance rather than to a retarded swelling.
During 3 min after the onset of Ca21-induced de-formation, the volume of soybean forisomes increasedsixfold (from 11.6 6 1.5 lm3, n 5 10, to 69.7 6 9.8lm3, n5 6). This is the same factor as in Canavalia fori-somes, which in terms of volume were about 75 timeslarger. The physiologically important greatest cross-
Fig. 4. Time-courses of polar circumference (dashed black lines; left ordinate), equatorial circumference
(solid black lines; left ordinate), and total surface area (gray lines; right ordinate) of the main bodies of
representative Canavalia gladiata forisomes during the short-term responses to the addition (A,B,C) andthe removal (D,E,F) of Ca21. A, B, and C correspond to the original data presented in Figs. 2A, 2C, and
2E, respectively. Similarly, D, E, and F correspond to Figs. 3A, 3C, and 3E, respectively.
374 Peters et al.

sectional area stepped up eightfold from 2.5 6 0.4 lm2
(n5 10) to reach 21.1 6 2.0 lm2 (n 5 6).As observed in Canavalia, the surface area of the
soybean forisome main body and its equatorial circum-ference changed strongly during the Ca21-relatedresponses (not shown), while the polar circumferenceremained more or less constant (Figs. 6B, 6D, 6F, and6H). The latter finding would not necessarily have beenexpected because of the distinct changes in geometrywhich forisomes of the two species undergo during theresponse. Canavalia forisomes change from parallelepi-peds of high aspect ratio to ones of markedly lower as-pect ratio when exposed to Ca21 ([Peters et al., 2007a];Fig. 1), implying that changes of the length of the lateralface are balanced by reverse changes of tip diameter. In
contrast, soybean forisomes transform from elongatespindle-shaped to spherical without detectable change indiameter at the points of tail insertion (Fig. 1), so thatunlike in Canavalia, no changes in the length of the lat-eral faces occur.
The reaction of soybean forisomes to an increaseof the medium pH to 10.5 differed qualitatively fromthat to Ca21 ions. At increased pH, the length of the fori-some main body remained constant throughout, whileboth D and d increased (Figs. 7A and 7B). As a result,forisome volume increased rather more strongly than inresponse to Ca21 (Fig. 7C; the final value was 132 655.2 lm3, n 5 5, as compared to 69.7 6 9.8 lm3, n 5 6,in the Ca21-response). The polar circumference alwaysbecame larger since the length of the lateral facesremained constant while both diameters (D and d)increased markedly. It should be noted that this type ofreaction was fully reversible (not shown), similar to theCa21-response.
DISCUSSION
The Ca21-driven but ATP-independent contractil-ity of forisome has been discovered only recently [Kno-blauch et al., 2003]. At this early stage, very simple mod-els have to serve as research-guiding hypotheses of fori-some function. One of these models aimed to explainpH-induced forisome swelling by postulating that Ca21
and protons acted on the same target domains on fori-some proteins [Knoblauch and Peters, 2004b]. Anothersimple model triggered the present study which was con-ducted to test the hypothesis that during the anisotropicdeformations in response to the addition and removal ofcalcium ions, changes in forisome length and diameterwere causally related and therefore strictly correlated.This notion was based on our previous finding that theanisotropy of the responses, but not Ca21-inducibleswelling as such was lost in mechanically damaged fori-somes [Knoblauch et al., 2003]. We speculated that iso-tropic swelling was a property of unorganized agglomer-ates of forisome proteins, whereas the anisotropicresponse with its opposite changes in length and diame-ter depended on defined supramolecular linkagesbetween the fibrils of the forisome. This idea led to asimplifying model in which the forisome main body wasenvisaged as a bundle of fibrils firmly attached to eachother only at the main body’s tips; such a forisomewould resemble a pair of umbrellas with their hubspointing in opposite directions, connected by pairwiselinkages between the free ends of the spokes. If the con-nected umbrellas open, the distances between the ribs(fibrils) will increase the most at the equator. A longitu-dinal shortening of the assembly will necessarily result;the umbrella poles (forisome tips) will move toward
Fig. 5. Kinetics of the response of a representative Canavalia gladi-ata forisome to changing the pH of the incubation medium from 7.3 to
10.5. (A) Time-courses of forisome main body length (L; &), central
diameter (D; l), and tip diameter (d; *) taken on each 8th frame of a
movie produced at a frame rate of 1000 s-1. Overlapping polynomials
were fitted piecewise to describe the complex trends in the original
data (black and gray lines in A), and continuous time-courses of the
volume of the forisome main body and its polar circumference were
computed from these fits (B). For definition of time 0, see Fig. 2.
Forisome Contraction Kinetics 375

each other, even though the lengths of the ribs (forisomefibrils) remain constant. Thus, changes in length wouldcoincide with radial swelling. Consequently, temporallyhighly resolved analyses of Ca21-induced forisome de-formation would be expected to reveal a strict correlationof the reverse changes in length and diameter. The natureof the causal relationship, however, would not necessar-ily be resolved; probably it would remain unclearwhether a passive longitudinal contraction occurred as aconsequence of a Ca21-driven radial expansion orwhether a Ca21-initiated longitudinal contraction drovea passive radial expansion. Moreover, if the umbrellamodel were to hold, the polar circumference of the fori-some main body would remain constant during theresponse, since the ribs (forisome fibrils) do not contract.In fact, we had found this to be the case in studies of thelong-term kinetics of forisome deformation in Canavalia[Peters et al., 2007a].
The corresponding short-term kinetics, however,did not unequivocally support the model. While changes
in polar circumference seemed insignificant during bothCa21- and pH-responses (Figs. 4 and 5), Ca21-dependentchanges in forisome length and diameter were not relatedin a way that would allow one to distinguish cause andeffect, as changes in one dimension did not obligatorilyprecede or follow those in the other (Figs. 2 and 3).Although the geometries of Ca21-dependent deformationdiffered significantly between Glycine and Canavalia(Fig. 1), Glycine forisomes resembled those of Canava-lia in undergoing contraction-expansion cycles withpractically constant polar circumferences (Figs. 6B, 6D,6F, and 6H), as required by the umbrella model. How-ever, the unexpected response of Glycine forisomes toalkaline pH during which they approached a box-shapedrather than spherical form and significantly increasedtheir polar circumference (Fig. 7) argues against the gen-eral validity of the umbrella model. The peculiar pH-response in Glycine cannot be explained by a pH-induced destruction of the proposed connections of theforisome fibrils at the tips of the main body, since the
Fig. 6. Kinetics of the responses to the addition (A–D) or removal
(E–H) of Ca21 of Glycine max forisomes; two representative exam-
ples are given for each type of reaction. Upper row graphs (A,C,E,G)
give forisome main body length (L; &), central diameter (D; l), and
tip diameter (d; *) as measured on each 3rd or 4th frame of movies
taken at 1000 frames per second; generally, d remained constant dur-
ing the experiments. Functions fitted to the original data (black or
gray lines in A,C,E,G) were used to compute continuous time-courses
of forisome main body volume (solid lines, left ordinate) and polar
circumference (dashed lines, right ordinate) shown in the lower row of
graphs (B,D,F,H). There, shaded areas indicate the 10–90% response
times of forisome volume. For definition of time 0, see Fig. 2. In A,
two arrows delimit a portion of the time-course of D in which the
main development is overlain by a regular oscillation of about 55 ms
periodicity. This oscillation in D corresponds to a rotation of the fori-
some around its longitudinal axis that is visible in the original movie,
supporting the idea that the forisome’s cross-section is not circular
(see text for details).
376 Peters et al.
response was fully reversible and repeatable, andbecause typical Ca21-responses could be induced after-wards (not shown). In support of the idea of a causalrelationship between changes in longitudinal and radialdimensions, one may argue that in contrast to the vari-able sequences of the onsets of changes in length and
diameter observed in Canavalia forisomes (Figs. 2 and3), the starts of the responses in Glycine almost alwaysappeared to be simultaneous (Fig. 6), and that the stricttemporal pattern implied by the model was present butnot resolved. While this may be true, it should be notedthat the rates of change in length and diameter did notusually coincide in soybean. We observed polyphasickinetics with sudden rate changes between phases ofapparently constant rate, but the times at which theserate changes occurred in forisome length and diameteroften differed (e.g. Fig. 6E). Taken together, our resultsdo not support the idea of a simple cause-effect relation-ship between changes in forisome length and diameter;the anisotropy of forisome contraction certainly repre-sents a more complex phenomenon. At this stage, thepossibility cannot be dismissed that forisome contractil-ity includes two or more basically independent molecu-lar mechanisms acting in parallel.
Compared to forisomes from Vicia faba [Kno-blauch et al., 2003] and Canavalia gladiata (this study),the behavior of soybean forisomes is baffling. Respond-ing to Ca21, their volume increased through changes incentral diameter and length at constant tip diameter (Fig.7). In contrast, greater changes in volume were achievedin alkaline pH when changes in diameter occurred alongthe complete forisome at constant length (Fig. 7). Thisqualitative variance argues against another simplemodel, namely our previous hypothesis that pH and diva-lent cations act on identical target sites [Knoblauch andPeters, 2004b].
Generally, forisome contractility is accompaniedby dramatic changes in forisome volume (Figs. 2, 3, 5, 6,and 7; [Knoblauch et al., 2003]) which are hard toexplain other than by fluxes of water into or out of theforisomes [Pickard et al., 2006]. They therefore resemblestimuli-sensitive hydrogels [Gehrke, 1993; Ball, 1997].More specifically, polyelectrolyte gels, a class of smartmaterials intensively studied at present [de Rossi andOsada, 1999], may provide a working hypothesis for themechanism of forisome contractility. Polyelectrolytepolymers carry groups that can be reversibly ionized,leading to the accumulation of counterions and subse-quent water fluxes which let the material swell [Smela,2003]. In technical systems, the ionization state of thepolymer is controlled electrically. It is conceivable thatforisomes may function analogously and that their swel-ling state is controlled either by their ionization state,which in a proteinaceous material obviously depends onthe medium pH, or by the availability of suitable coun-terions such as Ca21. As the rate of swelling in materialsof this type mostly depends on diffusive processes, mac-roscopic specimens of artificial gels react relativelyslowly [Smela, 2003]. In microscopic specimen of differ-ent size but similar shape, the time required to complete
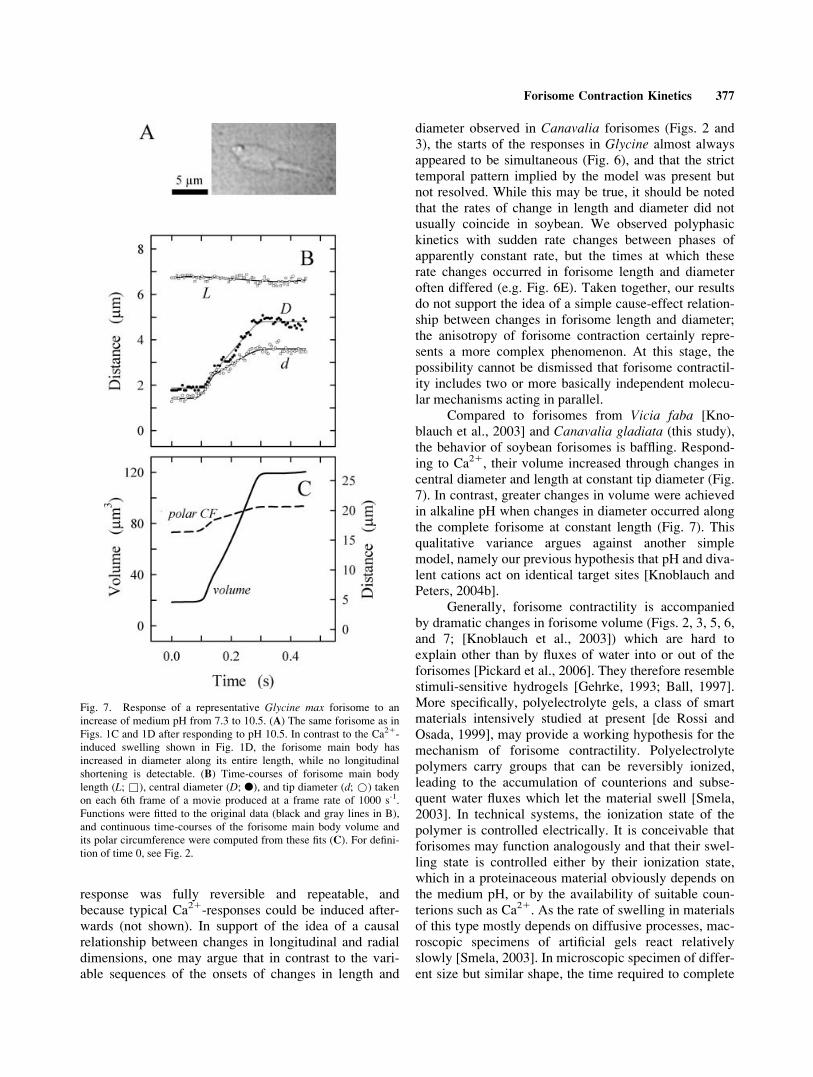
Fig. 7. Response of a representative Glycine max forisome to an
increase of medium pH from 7.3 to 10.5. (A) The same forisome as in
Figs. 1C and 1D after responding to pH 10.5. In contrast to the Ca21-
induced swelling shown in Fig. 1D, the forisome main body has
increased in diameter along its entire length, while no longitudinal
shortening is detectable. (B) Time-courses of forisome main body
length (L; &), central diameter (D; l), and tip diameter (d; *) taken
on each 6th frame of a movie produced at a frame rate of 1000 s-1.
Functions were fitted to the original data (black and gray lines in B),
and continuous time-courses of the forisome main body volume and
its polar circumference were computed from these fits (C). For defini-tion of time 0, see Fig. 2.
Forisome Contraction Kinetics 377

the swelling process should correlate with size. The factsthat Canavalia forisomes (resting state volume roughly900 lm3) required substantially longer periods to com-plete the swelling process than forisomes from soybean(resting state volume about 12 lm3), and that the formershowed a retarded swelling at low rates over periods ofminutes [Peters et al., 2007a] which we could not detectin the latter, were in line with this expectation.
Despite all differences between the species studiedso far, the uniformity of the contractile responses interms of dependency on divalent cations and pH but notATP, geometric anisotropy (at least in response toCa21), as well as ultrastructural similarities (longitudinalfibrils with regular cross-striation) and phylogenetic con-siderations (forisomes have been claimed to be a synapo-morphic trait of the faboid legumes; [Behnke and Pop,1981]) suggest that the basic molecular mechanisms ofcontractility are qualitatively identical in all forisomes.The conspicuous differences between forisomes fromdifferent species would then appear attributable to dis-tinct supramolecular architectures in which their ele-ments are arranged in the protein bodies. The disentan-glement of the proposed common molecular mechanismsand the taxon-specific supramolecular structures is achallenge for future studies.
REFERENCES
Amos WB, Routledge LM, Weis-Fogh T, Yew FF. 1976. The spasmo-
neme and calcium-dependent contraction in connection with
specific calcium binding proteins. In: Duncan CJ, editor. Cal-
cium in Biological Systems. Cambridge: Cambridge University
Press. pp 273–301.
Ball P. 1997. Made to Measure: New Materials for the 21st Century.
Princeton: Princeton University Press.
Behnke HD. 1991. Nondispersive protein bodies in sieve elements: A
survey and review of their origin, distribution and taxonomic
significance. IAWA Bull 12:143–175.
Behnke HD, Pop L. 1981. Sieve-element plastids and crystalline
p(hloem)-protein in Leguminosae: Micromorphological charac-
ters as an aid in the circumscription of the family and subfamilies.
In: Polhill RM, Raven PH, editors. Advances in Legume System-
atics 2. Kew (UK): Royal Botanical Gardens. pp 707–715.
Behnke HD, Sjolund RD, editors. 1990. Sieve Elements. Berlin:
Springer.
de Rossi D, Osada Y. 1999. Polymer Sensors and Actuators. Berlin:
Springer.
Evert RF. 1982. Sieve-tube structure in relation to function. Bio-
science 32:789–795.
Gehrke SH. 1993. Synthesis, equilibrium swelling, kinetics, perme-
ability and application of environmentally responsive gels.
Adv Polym Sci 110:81–144.
Knoblauch M, Peters WS. 2004a. Biomimetic actuators: Where tech-
nology and cell biology merge. Cell Mol Life Sci 61:2497–
2509.
Knoblauch M, Peters WS. 2004b. Forisomes, a novel type of Ca21-de-
pendent protein motor. Cell Motil Cytoskel 58:137–142.
Knoblauch M, Peters WS, Ehlers K, van Bel AJE. 2001. Reversible
calcium-regulated stopcocks in legume sieve tubes. Plant Cell
13:1221–1230.
Knoblauch M, Noll GA, Muller T, Prufer D, Schneider-Huther I,
Scharner D, van Bel AJE, Peters WS. 2003. ATP-independent
contractile proteins from plants. Nature Mater 2:600–603.
Knoblauch M, Noll GA, Muller T, Prufer D, Schneider-Huther I,
Scharner D, van Bel AJE, Peters WS. 2005. ATP-independent
contractile proteins from plants. Nature Mater 4:353.
Lawton DM. 1978. Ultrastructural comparison of the tailed and tail-
less P-protein crystals respectively of runner bean (Phaseolusmultiflorus) and garden pea (Pisum sativum) with tilting stage
electron microscopy. Protoplasma 97:1–11.
Maciejewski JJ, Vacchiano EJ, McCutcheon SM, Buhse HE, Jr. 1999.
Cloning and expression of a cDNA encoding a Vorticella con-vallaria spasmin: An EF-hand calcium-binding protein. J
Eukaryot Microbiol 46:165–173.
Mavroidis C, Dubey A. 2003. From pulses to motors. Nature Mater
2:573–574.
Moriyama Y, Okamoto H, Asai H. 1999. Rubber-like elasticity and vol-
ume changes in the isolated spasmoneme of giant Zoothamniumsp. under Ca21-induced contraction. Biophys J 76:993–1000.
Noll GA, Fontanellaz ME, Ruping B, Ashoub A, van Bel AJE, Fischer
R, Knoblauch M, Prufer D. 2007. Spatial and temporal regula-
tion of the forisome gene for1 in the phloem during plant
development. Plant Mol Biol 65:285–294.
Palevitz BA, Newcomb EH. 1971. The ultrastructure and development
of tubular and crystalline P-protein in the sieve elements of cer-
tain papilionaceous legumes. Protoplasma 72:399–425.
Peters WS, Baskin TI. 2006. Tailor-made composite functions as tools
in model choice: The case of sigmoidal vs bi-linear growth pro-
files. Plant Methods 2:11.
Peters WS, Knoblauch M, Warmann SA, Shen AQ, Pickard WF.
2007a. Tailed forisomes of Canavalia gladiata: A new model to
study Ca21-driven protein contractility. Ann Bot 100:101–109.
Peters WS, Schnetter R, Knoblauch M. 2007b. Reversible birefrin-
gence suggests a role for molecular self-assembly in forisome
contractility. Funct Plant Biol 34:302–306.
Pickard WF, Knoblauch M, Peters WS, Shen AQ. 2006. Prospective
energy densities in the forisome, a new smart material. Mater
Sci Eng C 26:104–112.
Sjolund RD. 1997. The phloem sieve element: A river runs through it.
Plant Cell 9:1137–1146.
Smela E. 2003. Conjugated polymer actuators for biomedical applica-
tions. Adv Mater 15:481–494.
Straßburger E. 1891. Ueber den Bau und die Verrichtungen der Lei-
tungsbahnen in den Pflanzen (Histologische Beitrage 3). Jena:
Fischer.
Wergin WP, Newcomb EH. 1970. Formation and dispersal of crystal-
line P-protein in sieve elements of soybean (Glycine max L.).
Protoplasma 71:365–388.
378 Peters et al.




















