
Defining the selectivity of processes along the auxin response chain: a study using auxin analogues
15
Defining the selectivity of processes along the auxin response chain: a study using auxin analogues Sibu Simon 1,2,3 , Martin Kube s 1 , Pawel Baster 2,3 , St ephanie Robert 2,4 , Petre Ivanov Dobrev 1 , Ji r ı Friml 2,3,5 , Jan Petr a sek 1 and Eva Za z ımalov a 1 1 Institute of Experimental Botany, The Academy of Sciences of the Czech Republic, Rozvojov a 263, 16502, Prague 6, Czech Republic; 2 Department of Plant Systems Biology, VIB and Department of Plant Biotechnology and Genetics, Ghent University, 9052, Ghent, Belgium; 3 Developmental and Cell Physiology of Plants, Institute of Science and Technology (IST Austria), 3400, Klosterneuburg, Austria; 4 SLU/Ume a Plant Science Center, 90183, Ume a, Sweden; 5 Department of Functional Genomics and Proteomics, Central European Institute of Technology (CEITEC), Masaryk University, 62500, Brno, Czech Republic Author for correspondence: Eva Za z ımalov a Tel: +420 225106429 Email: [email protected] Received: 6 May 2013 Accepted: 25 June 2013 New Phytologist (2013) 200: 1034–1048 doi: 10.1111/nph.12437 Key words: auxin analogues, auxin signalling, auxin transport, indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), naphthalene-1-acetic acid (NAA), 2,4-dichlorophenoxyacetic acid (2,4-D). Summary The mode of action of auxin is based on its non-uniform distribution within tissues and organs. Despite the wide use of several auxin analogues in research and agriculture, little is known about the specificity of different auxin-related transport and signalling processes towards these compounds. Using seedlings of Arabidopsis thaliana and suspension-cultured cells of Nicotiana tabacum (BY-2), the physiological activity of several auxin analogues was investigated, together with their capacity to induce auxin-dependent gene expression, to inhibit endocyto- sis and to be transported across the plasma membrane. This study shows that the specificity criteria for different auxin-related processes vary widely. Notably, the special behaviour of some synthetic auxin analogues suggests that they might be useful tools in investigations of the molecular mechanism of auxin action. Thus, due to their differential stimulatory effects on DR5 expression, indole-3-propionic (IPA) and 2,4,5- trichlorophenoxy acetic (2,4,5-T) acids can serve in studies of TRANSPORT INHIBITOR RESPONSE 1/AUXIN SIGNALLING F-BOX (TIR1/AFB)-mediated auxin signalling, and 5-flu- oroindole-3-acetic acid (5-F-IAA) can help to discriminate between transcriptional and non- transcriptional pathways of auxin signalling. The results demonstrate that the major determinants for the auxin-like physiological potential of a particular compound are very complex and involve its chemical and metabolic stability, its ability to distribute in tissues in a polar manner and its activity towards auxin sig- nalling machinery. Introduction The auxin class of plant growth regulatory compounds controls different growth and developmental events in plants. Charles and Francis Darwin first noted the existence of an endogenous sub- stance with biological activity in plants (Darwin & Darwin, 1980). This substance was characterized and reported as indole-3- acetic acid (IAA) (K€ ogl et al., 1934; Went & Thimann, 1937) and it has been proved to be the major endogenous auxin present in plants. Other compounds with auxin characteristics, such as indole-3-butyric acid (IBA) (Zimmerman & Wilcoxon, 1935), phenylacetic acid (PAA) (Koepfli et al., 1938) and 4-chloroin- dole-3-acetic acid (4-Cl-IAA) (Porter & Thimann, 1965), were later identified as endogenous auxins. Chemically more stable synthetic auxins, such as naphthalene-1-acetic acid (NAA) and 2,4-dichlorophenoxyacetic acid (2,4-D), are used as auxins in var- ious biological applications, and some of these auxins and their structural analogues, such as, for example, polychloroaromatic acids or chlorinated picolinic acids, are efficient herbicides (Walsh et al., 2006; Grossmann, 2010; M€ uller & Appleby, 2010). A number of other substances, mostly various carboxy derivatives of benzene, phenol, naphthalene or indole, show varying degrees of auxin-like activity in different bioassays (Koepfli et al., 1938; Porter & Thimann, 1965; Ferro et al., 2006; Christian et al., 2008). The coordinated growth and development of plants require the maintenance of optimum concentration gradients of active auxin(s) in specific organs, tissues and/or cell types, where they often act in co-operation with other plant hormones (Depuydt & Hardtke, 2011; Leyser, 2011; Rosquete et al., 2012). The regu- lated development and maintenance of appropriate auxin con- centration gradients in tissues are achieved by local metabolic processes, including biosynthesis, conjugation/deconjugation and degradation (Ljung et al., 2002; Chandler, 2009; Ljung, 2013), and/or by polar transport and intracellular auxin compartmental- ization (Grunewald & Friml, 2010). These processes result in the 1034 New Phytologist (2013) 200: 1034–1048 Ó 2013 The Authors New Phytologist Ó 2013 New Phytologist Trust www.newphytologist.com Research
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Defining the selectivity of processes along the auxin response chain: a study using auxin analogues

Defining the selectivity of processes along the auxin responsechain: a study using auxin analogues
Sibu Simon1,2,3, Martin Kube�s1, Pawel Baster2,3, St�ephanie Robert2,4, Petre Ivanov Dobrev1, Ji�r�ı Friml2,3,5,
Jan Petr�a�sek1 and Eva Za�z�ımalov�a1
1Institute of Experimental Botany, The Academy of Sciences of the Czech Republic, Rozvojov�a 263, 16502, Prague 6, Czech Republic; 2Department of Plant Systems Biology, VIB and
Department of Plant Biotechnology and Genetics, Ghent University, 9052, Ghent, Belgium; 3Developmental and Cell Physiology of Plants, Institute of Science and Technology (IST Austria),
3400, Klosterneuburg, Austria; 4SLU/Ume�a Plant Science Center, 90183, Ume�a, Sweden; 5Department of Functional Genomics and Proteomics, Central European Institute of Technology
(CEITEC), Masaryk University, 62500, Brno, Czech Republic
Author for correspondence:Eva Za�z�ımalov�a
Tel: +420 225106429Email: [email protected]
Received: 6 May 2013
Accepted: 25 June 2013
New Phytologist (2013) 200: 1034–1048doi: 10.1111/nph.12437
Key words: auxin analogues, auxinsignalling, auxin transport, indole-3-aceticacid (IAA), indole-3-butyric acid (IBA),naphthalene-1-acetic acid (NAA),2,4-dichlorophenoxyacetic acid (2,4-D).
Summary
� The mode of action of auxin is based on its non-uniform distribution within tissues and
organs. Despite the wide use of several auxin analogues in research and agriculture, little is
known about the specificity of different auxin-related transport and signalling processes
towards these compounds.� Using seedlings of Arabidopsis thaliana and suspension-cultured cells of Nicotiana
tabacum (BY-2), the physiological activity of several auxin analogues was investigated,
together with their capacity to induce auxin-dependent gene expression, to inhibit endocyto-
sis and to be transported across the plasma membrane.� This study shows that the specificity criteria for different auxin-related processes vary
widely. Notably, the special behaviour of some synthetic auxin analogues suggests that they
might be useful tools in investigations of the molecular mechanism of auxin action. Thus, due
to their differential stimulatory effects on DR5 expression, indole-3-propionic (IPA) and 2,4,5-
trichlorophenoxy acetic (2,4,5-T) acids can serve in studies of TRANSPORT INHIBITOR
RESPONSE 1/AUXIN SIGNALLING F-BOX (TIR1/AFB)-mediated auxin signalling, and 5-flu-
oroindole-3-acetic acid (5-F-IAA) can help to discriminate between transcriptional and non-
transcriptional pathways of auxin signalling.� The results demonstrate that the major determinants for the auxin-like physiological
potential of a particular compound are very complex and involve its chemical and metabolic
stability, its ability to distribute in tissues in a polar manner and its activity towards auxin sig-
nalling machinery.
Introduction
The auxin class of plant growth regulatory compounds controlsdifferent growth and developmental events in plants. Charles andFrancis Darwin first noted the existence of an endogenous sub-stance with biological activity in plants (Darwin & Darwin,1980). This substance was characterized and reported as indole-3-acetic acid (IAA) (K€ogl et al., 1934; Went & Thimann, 1937)and it has been proved to be the major endogenous auxin presentin plants. Other compounds with auxin characteristics, such asindole-3-butyric acid (IBA) (Zimmerman & Wilcoxon, 1935),phenylacetic acid (PAA) (Koepfli et al., 1938) and 4-chloroin-dole-3-acetic acid (4-Cl-IAA) (Porter & Thimann, 1965), werelater identified as endogenous auxins. Chemically more stablesynthetic auxins, such as naphthalene-1-acetic acid (NAA) and2,4-dichlorophenoxyacetic acid (2,4-D), are used as auxins in var-ious biological applications, and some of these auxins and theirstructural analogues, such as, for example, polychloroaromatic
acids or chlorinated picolinic acids, are efficient herbicides (Walshet al., 2006; Grossmann, 2010; M€uller & Appleby, 2010). Anumber of other substances, mostly various carboxy derivatives ofbenzene, phenol, naphthalene or indole, show varying degrees ofauxin-like activity in different bioassays (Koepfli et al., 1938;Porter & Thimann, 1965; Ferro et al., 2006; Christian et al.,2008).
The coordinated growth and development of plants requirethe maintenance of optimum concentration gradients of activeauxin(s) in specific organs, tissues and/or cell types, where theyoften act in co-operation with other plant hormones (Depuydt &Hardtke, 2011; Leyser, 2011; Rosquete et al., 2012). The regu-lated development and maintenance of appropriate auxin con-centration gradients in tissues are achieved by local metabolicprocesses, including biosynthesis, conjugation/deconjugation anddegradation (Ljung et al., 2002; Chandler, 2009; Ljung, 2013),and/or by polar transport and intracellular auxin compartmental-ization (Grunewald & Friml, 2010). These processes result in the
1034 New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research

establishment of optimum auxin concentration gradients in spe-cific organs, tissues, cells and their compartments, and, in turn,transduce an auxin signal into appropriate biochemical and phys-iological responses.
Among plant hormones, active polar cell-to-cell transport is aunique property of auxin. The specificity of transport routes forfrequently used auxins has been characterized in many plant sys-tems, including maize coleoptiles (Hertel et al., 1969) and roots(Martin & Pilet, 1986), zucchini hypocotyl segments (Depta &Rubery, 1984), Arabidopsis inflorescence stalks (Parry et al.,2001), Arabidopsis root, hypocotyls and inflorescence (Rashotteet al., 2003), suspension-cultured crown gall cells (Rubery,1977), soybean root cells (Loper & Spanswick, 1991), suspen-sion-cultured tobacco cells of Nicotiana tabacum L. cv XanthiXHFD8 (Delbarre et al., 1996), cv Bright Yellow-2 (BY-2)(Petr�a�sek et al., 2006) and cv Virginia Bright Italia (VBI-0)(Petr�a�sek et al., 2002), and zucchini and lupin microsomal vesi-cles (Sabater & Rubery, 1987). At the molecular level, the move-ment of auxin across the plasma membrane (PM) is mediated bythe AUXIN-RESISTANT1/LIKE AUX1 (AUX1/LAX) auxininflux carriers (Bennett et al., 1996; Swarup et al., 2008) andPIN-FORMED (PIN) auxin efflux carriers (K�re�cek et al., 2009).Moreover, plant orthologues of the mammalian ATP-bindingcassette subfamily B (ABCB)-type transporters also transportauxin (Petr�a�sek & Friml, 2009; Yang & Murphy, 2009). Theaffinity of components of the auxin transport machinery formajor auxins and their specific interactions differ in different celltypes (Delbarre et al., 1996; Paciorek et al., 2005), and a detailedstudy of auxin transport specificity is still lacking.
It has been reported that auxin itself can inhibit endocytosis ofsome PM-localized proteins, including PINs (Paciorek et al.,2005). Recently, a putative auxin receptor, the AUXIN BINDINGPROTEIN 1 (ABP1), has been shown to act as a positive regulatorof clathrin-mediated endocytosis of PINs and the binding of auxinrestricts this effect (Robert et al., 2010). Even though the narrowspecificity of PM-associated ABP1 towards auxin-like compoundswas suggested decades ago (Hertel et al., 1972; L€obler & Kl€ambt,1985; Za�z�ımalova & Kuta�cek, 1985), details of the specificityof the auxin-dependent inhibition of endocytosis are not yetknown.
In addition to ABP1, there is also a well-characterizedTRANSPORT INHIBITOR RESPONSE 1 (TIR1)-dependentauxin signalling pathway (Dharmasiri et al., 2005; Kepinski &Leyser, 2005). It is based on the degradation of Aux/IAA tran-scriptional repressors, mediated by the SCFTIR1 ubiquitin ligasecomplex in contact with auxin. The involvement of auxin is con-centration dependent and auxin acts as a ‘molecular glue’,enhancing the interaction between TIR1 and Aux/IAA (Tanet al., 2007). Recently, the joint action of TIR1/AFBs and Aux/IAAs has been reported for differential auxin sensing, suggestingthat the TIR1/AFBs and Aux/IAAs function as auxin co-receptors(Calderon Villalobos et al., 2012). On the basis of the structureof the promoter part of auxin-responsive genes, the ‘synthetic’auxin-responsive promoter DR5 was constructed (Ulmasov et al.,1997) and, in fusion with reporter genes, it is used widely to visu-alize the level of response to auxin.
It is obvious that the final mode of auxin action will dependon many partial auxin-related actions; however, to date there hasbeen no comprehensive study of the auxin specificity of theseprocesses. To address this deficiency and to obtain an insight intothe complex auxin-related regulatory mechanism(s) in plantdevelopment, we have selected a group of compounds (‘auxinanalogues’) that are structurally related to the three major auxins,that is, the native IAA and the synthetic NAA and 2,4-D. Inorder to determine their auxin-like effectiveness and to investi-gate the selectivity of processes along the auxin response chainfrom the whole plant to the organ, tissue and cellular level, weanalysed the capacity of these auxin analogues to be transportedand to induce DR5-driven gene expression. We also investigatedtheir ability to inhibit endocytosis of PM-related PIN proteins.These assays, together with the characterization of the auxin-likephysiological potential of selected auxin analogues, showed thesimilarity of the structural requirements for auxin-like substancesfor all these processes; however, they also revealed the specialbehaviour of some compounds.
Materials and Methods
Auxins and auxin-like compounds used
IAA, indole-3-propionic acid (IPA), IBA, PAA, NAA, naphtha-lene-2-acetic acid (2-NAA), 2,4-D, 2,4,5-T, 2-(2,4-dichloro-phenoxy) propionic acid (2,4-DP) and indole-3-lactic acid (ILA)were all obtained from Sigma-Aldrich Inc. (St Louis, MO, USA).5-Fluoroindole-3-acetic acid (5-F-IAA), 4-Cl-IAA, 5-chloroin-dole-3-acetic acid (5-Cl-IAA), 5-bromoindole-3-acetic acid(5-Br-IAA) and indole-3-acetyl alanine (IAAla) were purchasedfrom OlChemIm (Olomouc, Czech Republic). Benzoic acid(BA), indole and naphthalene were obtained from Fluka.6-Chloroindole-3-acetic acid (6-Cl-IAA) was a generous gift fromProfessor Kjeld Engvild (Riso National Laboratory, Roskilde,Denmark). 5-Cl-IAA stock solution (50 mM) was prepared indimethylsulfoxide (DMSO) and all other compounds were dis-solved in ethanol. [3H]NAA and [3H]2,4-D (both with a molar(specific) radioactivity of 20 Ci mmol�1) and [3H]IBA (molar(specific) radioactivity of 25 Ci mmol�1) were purchased fromAmerican Radiolabeled Chemicals Inc. (St Louis, MO, USA).
Plant materials and growth conditions
Wild-type Arabidopsis thaliana Heynh. seeds and tir1-1 mutant(Ruegger et al., 1998) (both ecotype Columbia) were surface ster-ilized with 70% (v/v) ethanol and 0.05% (v/v) Triton X-100 for10 min, shaking occasionally. The liquid from the first wash wasreplaced with the same volume of 100% ethanol for 5 min. Seedswere dried on sterile Whatman filter paper in a sterile hood.Dried seeds were plated on medium containing 0.6% (w/v) agar,2.3 g l�1 Murashige and Skoog salts, 0.5 g l�1 Mes (pH 5.8), 1%(w/v) sucrose and 0.1 g l�1 myo-inositol. Seeds were kept in thedark for 24 h at 4°C. Plates were wrapped with surgical tape andplaced vertically in racks under a 16 h : 8 h, light : dark photope-riod at room temperature (22°C).
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 1034–1048
www.newphytologist.com
NewPhytologist Research 1035

Cells of tobacco (Nicotiana tabacum L., cv Bright Yellow-2)line BY-2 (Nagata et al., 1992) were cultivated as described previ-ously (Petr�a�sek et al., 2006) and subcultured weekly. Arabidopsisthaliana, ecotype Landsberg erecta, suspension-cultured cells(May & Leaver, 1993) were maintained in the same medium asBY-2 cells and subcultured weekly.
Arabidopsis root elongation and lateral root formationassays
Wild-type and tir1 Arabidopsis seeds were germinated and grownfor 4 d as described above. Seedlings were transferred to solidmedium supplemented with a determined concentration of thecompound to be analysed and the position of the root tip wasmarked. The seedlings were grown for an additional 4 d in con-tinuous light under a yellow filter (Stasinopoulos & Hangarter,1990) and the root length was measured from the marked pointto the tip of the root.
For lateral root numbers, the primary root length was mea-sured as explained above, but not from the marked point, butrather from just below the hypocotyl part to the root tip, andthen photographed. Lateral root numbers were determined withthe image analysis software ImageJ (http://rsb.info.nih.gov/nih-image) by marking every emerged lateral root, and the num-ber of lateral roots per centimetre of primary length was calcu-lated with respect to the primary root length data.
Determination of BY-2 cell division activity
From 1-wk-old BY-2 suspension culture, 7.5 ml was transferredto 500 ml of fresh auxin-free BY-2 medium. This culture was ali-quoted (30 ml each) to 100-ml conical flasks, and the specifiedcompound to be tested was added to each flask from the stock togive the required final concentration. The culture was furtherincubated in the dark at 27°C on an orbital shaker at 120 rpmfor 1 wk. Samples of the culture were collected every day and thecell density (number of cells per 1 ml of cell suspension) wasdetermined by counting cells in at least six aliquots of each sam-ple using a Fuchs–Rosenthal haemocytometer slide.
DR5rev::GFP induction assay
Five to six, 4-d-old DR5rev::GFP seedlings (Benkov�a et al., 2003)were transferred to sterile liquid medium (2.3 g l�1 Murashige andSkoog salts, 0.5 g l�1 Mes (pH 5.8), 1% (w/v) sucrose, 0.1 g l�1
myo-inositol). The required concentration (1 or 5 lM) of experi-mental compound was applied and the seedlings were incubatedfor 4 or 24 h wrapped with yellow long-pass filter film with trans-mittance below 450 nm (Stasinopoulos &Hangarter, 1990) on anorbital shaker (150 rpm). The green fluorescent protein (GFP) sig-nal was observed in roots using a Zeiss LSM510 confocal laser scan-ning microscope (Carl Zeiss, Jena, Germany) with excitation at488 nm using an argon laser and a 920 objective. The quantifica-tion of the fluorescence intensity was performed with image analy-sis software ImageJ (http://rsb.info.nih.gov/ij/) by selecting theroot tip area (meristem, transition and elongation zones included).
Analysis of the inhibitory effect of auxin on endocytosis
The experiments were performed as described in Paciorek et al.(2005). Four to 5-d-old wild-type Arabidopsis seedlings wereincubated in sterile liquid medium supplemented with the com-pounds at 25 lM concentration for 30 min, followed by 90 minof treatment with 25 lM brefeldin A (BFA). The immunodetec-tion of PIN1 and PIN2 proteins was performed according toFriml et al. (2003). The anti-PIN1 antibody (1 : 1000) (Benkov�aet al., 2003), anti-PIN2 antibody (1 : 1000) (Abas et al., 2006)and fluorochrome-conjugated secondary antibody anti-rabbit-Cy3 (1 : 600) (Dianova, Hamburg, Germany) were used. Imag-ing was performed as described previously (Robert et al., 2010).The rate of the inhibitory effect of each compound was measuredby counting the number of BFA bodies in individual cells.
Auxin accumulation assay
The accumulation of radiolabelled auxins was measured asdescribed in Petr�a�sek et al. (2006).
IBA metabolic assay
Two-day-old BY-2 cells were equilibrated in the uptake buffer asdescribed for accumulation assays. Experiments were performedin the uptake buffer and under standard cultivation conditions.Cells were incubated with 20 nM [3H]IBA for a period from 30 sto 48 h in darkness. After incubation, the cells were filtered and200 mg fresh weight cells (after draining the medium off underreduced pressure) were flash frozen in liquid nitrogen and storedat �80°C. The extraction and purification of the auxin metabo-lites from cells and media were performed as described previously(Dobrev et al., 2005). The analysis was monitored by a Ramona2000 flow-through radioactivity detector (Raytest GmbH, Strau-benhardt, Germany) after online mixing with three volumes(1.8 ml min�1) of liquid scintillation cocktail (Flo-Scint III, Per-kin Elmer Life and Analytical Sciences, Shelton, CT, USA), andthe results were normalized on the basis of the total accumulatedradioactivity.
Statistical analysis
Differences between control and experimental variants were anal-ysed using one-way ANOVA. When significant differences weredetected at a 95% level of confidence, the multi-range Tukey’spost-hoc test was applied. All statistical tests were performed usingMicrosoft Excel and GraphPad software (http://graphpad.com).
Results
Selection of the auxin analogues
The selection of compounds was performed on the basis of theirstructural similarity to major native (IAA) and synthetic (NAAand 2,4-D) auxins. As all active auxins are weak organic acids andcontain aromatic or heterocyclic ring(s), we have selected various
New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist1036

derivatives of diverse rings bearing one carboxyl group on a short(carbon–carbon or oxygen–carbon) side-chain. Thus, carboxyderivatives of benzene (BA), indole (IAAs, IPA, IBA, ILA), naph-thalene (NAAs) and phenol (PAA and phenoxyacetic acids) wereused (Supporting Information Fig. S1). Compounds containingonly corresponding heterocyclic and aromatic rings, that is,indole and naphthalene, respectively, and an auxin conjugate(IAAla) were chosen as controls.
Effect of auxin analogues on Arabidopsis primary rootgrowth and lateral root formation
The auxin-like physiological competence of selected compoundswas analysed in Arabidopsis thaliana seedlings using auxin assaysbased on the inhibition of primary root elongation and theinduction of lateral root formation.
IAA and most of its halogenated derivatives inhibited primaryroot elongation efficiently at concentrations between 100 and500 nM. The most active compound tested was 6-Cl-IAA, whichexhibited maximum inhibition at a concentration as low as10 nM (Fig. 1a). Non-halogenated indolic acids (IPA and IBA)required a much higher concentration (10 lM) for maximumeffect. Other indole-based compounds had only slight effects onroot elongation (Fig. 1b).
The application of NAA caused maximum inhibition ofprimary root elongation at 500 nM, whereas its positionalanalogue, 2-NAA, as well as the weak native auxin PAA,inhibited root elongation completely at a concentration oneorder of magnitude higher (Fig. 1c,d). By contrast, the dichlo-rophenoxy derivatives 2,4-D and 2,4-DP were very active, withmaximum inhibitory effects at 100 nM, but 2,4,5-T, anotherstructural analogue of this class, was comparatively less effective(Fig. 1d).
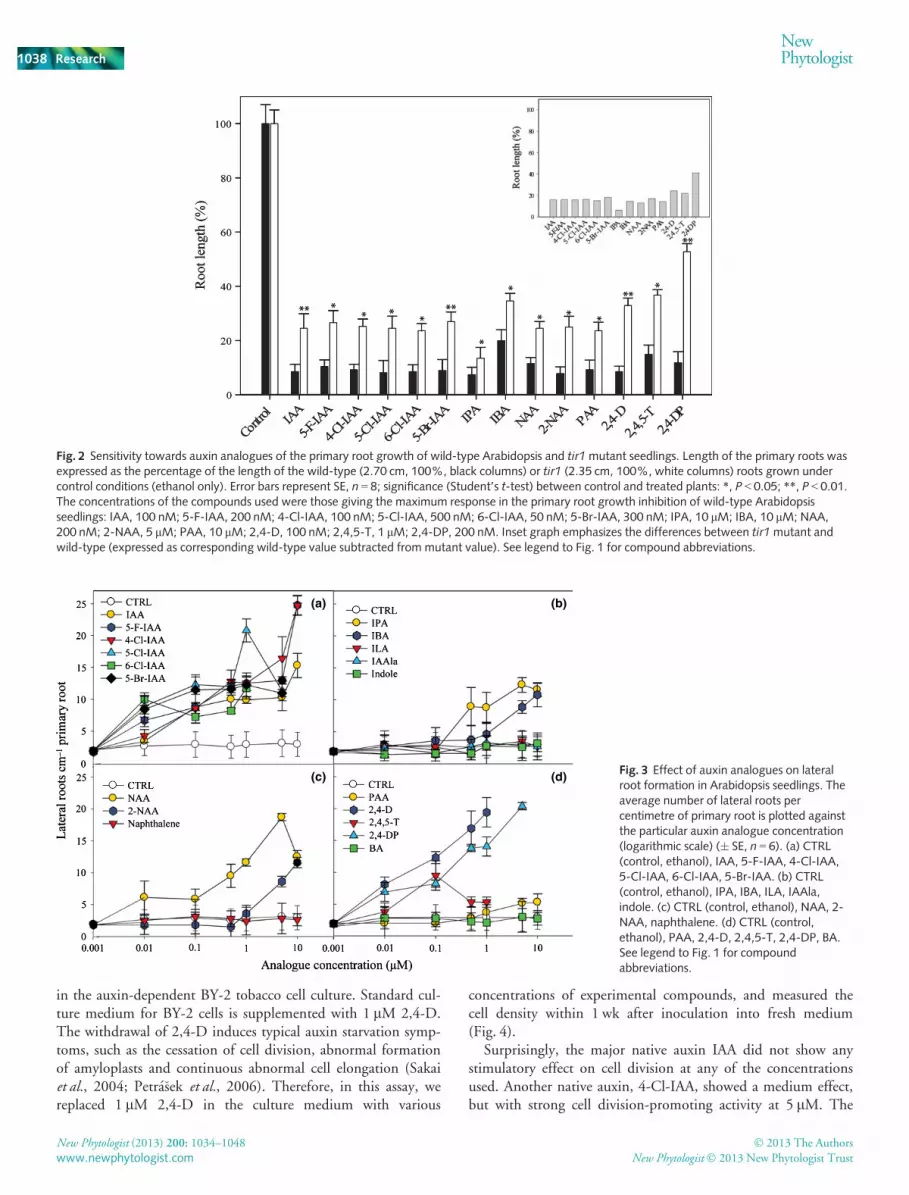
As TIR1 seems to be the auxin receptor that is important formany processes in plant development, we also analysed the sensi-tivity of tir1 knockout mutant seedlings to these compounds(Fig. 2). Most of the tested compounds showed more or less simi-lar effects on the primary root elongation of both wild-type andtir1 seedlings. However, treatments with 2,4-D and its structuralanalogues, that is, 2,4,5-T and 2,4-DP, resulted in less inhibitionin tir1 relative to the wild-type. By contrast, IPA showed thehighest inhibition of primary root elongation in tir1 (Fig. 2,inset), and it appears to be the least dependent on TIR1 toexpress its auxin-like activity.
In lateral root initiation (Fig. 3), 6-Cl-IAA was the compoundwith the greatest activity (Fig. 3a). 5-Cl-IAA showed its highesteffect at 1 lM; however, above 1 lM concentration its action wasinhibitory. Similarly, NAA showed an increasing effect up to5 lM and above this concentration its effect was inhibitory(Fig. 3c). IAA, 4-Cl-IAA and 5-F-IAA increased lateral root num-ber with an increase in concentration of the tested compounds upto 10 lM, the last two compounds promoting more lateral rootsthan IAA. 2,4-D and 2,4-DP also showed high activity (Fig. 3d),but other compounds showed either very weak or no activity inthis assay.
Arabidopsis root growth assays showed that, apart from IAAitself, the most potent auxin-like compounds included most ofthe IAA derivatives halogenated in positions 4, 5 and 6 of theindole ring, as well as 2,4-D and its structural analogues, andNAA.
Effects of auxin analogues on cell division in suspension-grown tobacco BY-2 cells
Compounds showing very obvious effects on root phenotype inArabidopsis were tested for their cell division-stimulating activity
(a) (b)
(c) (d)
Fig. 1 Effect of auxin analogues on thegrowth of the primary root in Arabidopsisseedlings. Length of the root (cm) is plottedagainst the particular auxin analogueconcentration (logarithmic scale) (� SE,n = 6). (a) CTRL (control, ethanol),IAA (indole-3-acetic acid), 5-F-IAA(5-fluoroindole-3-acetic acid), 4-Cl-IAA(4-chloroindole-3-acetic acid), 5-Cl-IAA (5-chloroindole-3-acetic acid), 6-Cl-IAA (6-chloroindole-3-acetic acid), 5-Br-IAA(5-bromoindole-3-acetic acid). (b) CTRL(control, ethanol), IPA (indole-3-propionicacid), IBA (indole-3-butyric acid), ILA (indole-3-lactic acid), IAAla (indole-3-acetyl alanine),indole. (c) CTRL (control, ethanol), NAA(naphthalene-1-acetic acid), 2-NAA(naphthalene-2-acetic acid), naphthalene.(d) CTRL (control, ethanol), PAA(phenylacetic acid), 2,4-D (2,4-dichlorophenoxyacetic acid), 2,4,5-T (2,4,5-trichlorophenoxyacetic acid), 2,4-DP (2-(2,4-dichlorophenoxy) propionic acid), BA(benzoic acid).
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 1034–1048
www.newphytologist.com
NewPhytologist Research 1037
in the auxin-dependent BY-2 tobacco cell culture. Standard cul-ture medium for BY-2 cells is supplemented with 1 lM 2,4-D.The withdrawal of 2,4-D induces typical auxin starvation symp-toms, such as the cessation of cell division, abnormal formationof amyloplasts and continuous abnormal cell elongation (Sakaiet al., 2004; Petr�a�sek et al., 2006). Therefore, in this assay, wereplaced 1 lM 2,4-D in the culture medium with various
concentrations of experimental compounds, and measured thecell density within 1 wk after inoculation into fresh medium(Fig. 4).
Surprisingly, the major native auxin IAA did not show anystimulatory effect on cell division at any of the concentrationsused. Another native auxin, 4-Cl-IAA, showed a medium effect,but with strong cell division-promoting activity at 5 lM. The
Fig. 2 Sensitivity towards auxin analogues of the primary root growth of wild-type Arabidopsis and tir1mutant seedlings. Length of the primary roots wasexpressed as the percentage of the length of the wild-type (2.70 cm, 100%, black columns) or tir1 (2.35 cm, 100%, white columns) roots grown undercontrol conditions (ethanol only). Error bars represent SE, n = 8; significance (Student’s t-test) between control and treated plants: *, P < 0.05; **, P < 0.01.The concentrations of the compounds used were those giving the maximum response in the primary root growth inhibition of wild-type Arabidopsisseedlings: IAA, 100 nM; 5-F-IAA, 200 nM; 4-Cl-IAA, 100 nM; 5-Cl-IAA, 500 nM; 6-Cl-IAA, 50 nM; 5-Br-IAA, 300 nM; IPA, 10 lM; IBA, 10 lM; NAA,200 nM; 2-NAA, 5 lM; PAA, 10 lM; 2,4-D, 100 nM; 2,4,5-T, 1 lM; 2,4-DP, 200 nM. Inset graph emphasizes the differences between tir1mutant andwild-type (expressed as corresponding wild-type value subtracted from mutant value). See legend to Fig. 1 for compound abbreviations.
(a) (b)
(c) (d) Fig. 3 Effect of auxin analogues on lateralroot formation in Arabidopsis seedlings. Theaverage number of lateral roots percentimetre of primary root is plotted againstthe particular auxin analogue concentration(logarithmic scale) (� SE, n = 6). (a) CTRL(control, ethanol), IAA, 5-F-IAA, 4-Cl-IAA,5-Cl-IAA, 6-Cl-IAA, 5-Br-IAA. (b) CTRL(control, ethanol), IPA, IBA, ILA, IAAla,indole. (c) CTRL (control, ethanol), NAA, 2-NAA, naphthalene. (d) CTRL (control,ethanol), PAA, 2,4-D, 2,4,5-T, 2,4-DP, BA.See legend to Fig. 1 for compoundabbreviations.
New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist1038

application of 5-Cl-IAA and 5-F-IAA also stimulated cell divisionwith increasing concentrations, and both compounds were highlyactive at 5 lM. NAA was mildly active, whereas its structuralcounterpart, 2-NAA, exhibited no cell division-promoting activ-ity at any of the concentrations tested. As expected, 2,4-D wasthe most active compound, but only at concentrations up to3 lM; higher concentrations had an inhibitory effect. Interest-ingly, 2,4,5-T was highly active only at 0.5 lM and all other con-centrations appeared to be inhibitory. Strikingly, IBA was veryactive at 5 lM (Fig. 4). As IBA was much more active in thisassay than IAA itself, and as it has been suggested to act as anIAA precursor (Strader et al., 2010), we tracked the metabolicfate of radiolabelled IBA in BY-2 cells, tracing free radiolabelledIAA at different time points from 30 s to 48 h after labelled IBAapplication. Free labelled IAA was not detected at any of the timepoints measured (Fig. S2), suggesting that, under our experimen-tal conditions, at least at the beginning of the subculture interval,IBA stimulates cell division independently of IAA.
Altogether, this assay confirmed 2,4-D as a preferential auxin-like compound for the standard growth of BY-2 cells and indi-cated that other 2,4-D-like compounds (namely 2,4,5-T) are ableto promote cell division efficiently at low concentration. It alsopointed to an IAA-independent action of IBA in BY-2 cells.
Active transport of auxin analogues across PM insuspension-grown cells
For tobacco cells, auxin influx and efflux carrier affinity has beenattributed mostly to the major synthetic auxins, either 2,4-D orNAA, respectively, whereas the native auxin IAA is a good
substrate for both classes of carriers in cells of tobacco cv XanthiXHFD8 (Delbarre et al., 1996) or BY-2 and VBI-0 (Pacioreket al., 2005). This distinct substrate specificity of auxin carriersfacilitates the quantitative analysis of the active influx and effluxof different compounds during accumulation assays in tobaccocells: using a simple displacement of [3H]2,4-D and [3H]NAA,respectively, by non-labelled auxin analogues, the relative affinityof a particular type of carrier for such analogues can be measured.For comparison, we also performed competitive auxin accumula-tion assays on suspension-grown Arabidopsis cells.
Of all the compounds tested, at the influx level, IAA maximallydecreased the accumulation of [3H]2,4-D (Figs 5a, S3a). Intobacco cells, this decrease was even higher than that resulting fromthe application of the auxin influx carrier inhibitor 2-naphthoxyacetic acid (2-NOA, La�nkov�a et al., 2010) or unlabelled 2,4-D.Halogenated derivatives of IAA were also highly competitive tolabelled 2,4-D in both tobacco (Fig. 5a) and Arabidopsis (Fig. S4a)cells. NAA did not compete for auxin influx carriers but, notably,its structural isomer 2-NAA decreased the accumulation of labelled2,4-D significantly in both tobacco (Fig. 5a) and Arabidopsis(Fig. S4a) cells. This finding is consistent with its status as an auxininflux carrier ‘inhibitor’ in tobacco cv Xanthi XHFD8 cells(Delbarre et al., 1996). Surprisingly, IBA increased slightly theaccumulation of labelled 2,4-D in tobacco cells, but did not showany effect in Arabidopsis cells (Figs 5a, S4a). Representative shapesof accumulation curves for all experimental compounds are shownin Figs S3(a) and S5(a) for BY-2 and Arabidopsis, respectively.
IAA and all of its halogenated derivatives competed efficientlywith labelled NAA for active auxin efflux (thus resulting inincreased [3H]NAA accumulation in cells), but the major native
Fig. 4 Effects of auxin analogues on celldivision in suspension-grown tobacco BY-2cells. Cells were grown in auxin-free (CTRL,control) standard cultivation mediumsupplemented with auxin analogues atconcentrations of 0.5, 1.0, 3.0 and 5.0 lM(as indicated in the panels). Each pointrepresents the mean value of eightmeasurements. Error bars were always< 0.2% of the mean and were omitted in thegraph to avoid its ‘illegibility’. See legend toFig. 1 for compound abbreviations.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 1034–1048
www.newphytologist.com
NewPhytologist Research 1039

auxin IAA showed the least activity. Of all the compounds testedin tobacco cells, 5-Cl-IAA competed the most strongly, followedby 6-Cl-IAA and 5-Br-IAA (Fig. 5b); however, in Arabidopsiscells, 5-Br-IAA was the most effective competitor (Fig. S4b).Non-labelled NAA appeared to be less competitive than someother compounds (such as halogenated IAAs), and 2-NAA wasonly a slightly less active inhibitor than NAA itself in bothtobacco and Arabidopsis cells. A widely used chemical inhibitorof active auxin efflux, 1-naphthylphthalamic acid (NPA), wasused as a positive control; it showed the maximum inhibition ofauxin efflux, being relatively more effective in Arabidopsis thanin BY-2 cells. Representative shapes of the accumulation curvesfor all compounds tested are shown in Figs S3(b) and S5(b) forBY-2 and Arabidopsis, respectively. These findings reveal distinct
specificities of auxin influx and efflux carriers towards some syn-thetic auxins.
The ability of auxin analogues to induce DR5-driven geneexpression
To address the possible activity of the auxin analogues in elic-iting an auxin response, we tested their ability to induce theexpression of DR5rev::GFP in Arabidopsis roots. Generally,IAA and all of its halogenated derivatives induced DR5rev::GFP expression efficiently, and IAA itself and 6-Cl-IAA werethe most active. Similarly, 2,4-D and 2,4-DP showed highlevels of activity. NAA induced DR5rev::GFP to a slightlylesser extent and other compounds did not show statistically
(a)
(b)
Fig. 5 Active transport of auxin and itsanalogues across the plasma membrane intobacco BY-2 cells. (a) Accumulation ofradiolabelled 2,4-D, as a marker for the activeinflux of auxin into cells, in competition withauxin analogues, measured 20min after theaddition of [3H]2,4-D (2 nM) together withthe tested compound (10 lM). (b)Accumulation of radiolabelled NAA, as amarker of the active auxin efflux from cells, incompetition with auxin analogues, measured20min after the addition of [3H]NAA (2 nM)together with the tested compound (10 lM).Data are expressed as a percentage of theaccumulation of labelled 2,4-D or NAA attime 20min after their addition to the cellstogether with the solvent (ethanol) only, in(a) and (b), respectively. 2-Naphthoxy aceticacid (2-NOA) and 1-naphthylphthalamic acid(NPA) (10 lM each) were used as positivecontrols for active auxin influx (a) and efflux(b), respectively. Error bars represent SE,n = 8. See legend to Fig. 1 for compoundabbreviations.
New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist1040

significant effects (Fig. 6a,b). To test the possibility that someof the inactive compounds may only be active at higher con-centrations and/or only after longer treatment, the same assaywas repeated using higher concentrations (5 lM) of all theweakly active and inactive compounds and for a longer timeof treatment (24 h). Under these conditions, 2-NAA induceda considerable effect and 2,4,5-T exhibited a very high activ-ity (Fig. S6).
Taken together, these results show that those compounds thataffected the Arabidopsis root phenotype were also active in induc-ing DR5rev::GFP expression; however, at higher concentrations
and after longer treatment, certain other compounds also inducedDR5rev::GFP expression to a considerable extent.
The involvement of auxin analogues in the inhibition ofendocytosis of PIN auxin efflux carriers
BFA is widely used to inhibit vesicle trafficking from endosomalcompartments to the PM in plant tissues, and treatment withBFA leads to the deposition of some PM proteins in so-calledBFA compartments inside cells (Geldner et al., 2001). Thus,BFA can be used to visualize proteins internalized by endocytosis.
(a)
(b)
Fig. 6 Activity of auxin analogues in thestimulation of DR5-driven gene expression.Four-day-oldDR5rev::GFPArabidopsisseedlings were treated with the specifiedcompound and single confocal scansthrough the root tip were taken after 4 h.(a) Representative image from control(mediumwith the corresponding volume ofsolvent only) and each compound (1 lM).Bar, 500 lm. Note that the active auxin andauxin analogues induced green fluorescenceprotein (GFP) fluorescence along all this partof the root, in contrast with control andinactive compounds, where the fluorescencewas restricted only to the very tip. (b) Graphrepresenting total GFP fluorescence intensityin each experimental variant. Error barsrepresent SE, n = 10. Significant differencebetween control and experimental variantsas determined by one-way ANOVA:**, P < 0.01. See legend to Fig. 1 forcompound abbreviations.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 1034–1048
www.newphytologist.com
NewPhytologist Research 1041

We followed the inhibitory effect of auxins and auxin-like com-pounds on the BFA-induced intracellular deposition of PIN1and PIN2 in Arabidopsis root stele and cortex/epidermis regions,
respectively. The remarkable inhibitory effect of some auxins(namely 2,4-D and NAA) on endocytosis has been reported pre-viously (Paciorek et al., 2005).
Control BFA IAABFA
4-Cl-IAABFA
5-Cl-IAABFA
6-Cl-IAABFA
5-Br-IAABFA
IPABFA
IBABFA
ILABFA
IAAlaBFA
NAABFA
2-NAABFA
NaphthaleneBFA
PAABFA
2,4-DBFA
2,4,5-TBFA
2,4-DPBFA
BABFA
5-F-IAABFA
IndoleBFA
Cont
rol
BFA
IAA
+ BF
A5-
F-IA
A +
BFA
4-Cl
-IAA
+ BF
A5-
Cl-IA
A +
BFA
6-Cl
-IAA
+ BF
A5-
Br-IA
A +
BFA
IPA
+ BF
AIB
A +
BFA
ILA
+ BF
AIA
Ala +
BFA
Indo
le +
BFA
NAA
+ BF
A2-
NAA
+ BF
A
Naph
thal
ene +
BFA
PAA
+ BF
A2,4
-D +
BFA
2,4,5-
T +
BFA
2,4-D
PBA
BFA
bod
ies p
er c
ell
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
**
** ** ****
**
** **
** **
****
****
*
(a)
(b)
Fig. 7 Activity of auxin analogues in the inhibition of endocytosis of PIN-FORMED (PIN) proteins. (a) Representative image from control (mediumwith thecorresponding volume of solvent only) and each compound (25 lM). PIN1 and PIN2 proteins were visualized using anti-PIN1 and anti-PIN2 antibodies.Inhibitory effect of auxin analogues on endocytosis was assessed by observing the level of PIN proteins in brefeldin A (BFA) compartments (arrowheads). Bar,10 lm. (b) Graph representing quantification of BFA compartments per cell in each experimental variant. Error bars, + SE, at least 120 cells from six differentplants for each variant were measured. Significant difference between treatments with BFA alone and BFA plus experimental compound as determined byone-way ANOVA: *, P < 0.05; **, P < 0.01. Note that most of the physiologically active auxin analogues showed reduced or no BFA compartments, exceptfor 5-F-IAA. See legend to Fig. 1 for compound abbreviations.
New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist1042

As shown in Fig. 7, 5-Br-IAA was the most active compoundin inhibiting the endocytosis of PINs. Halogenated derivatives ofIAA, as well as NAA, 2-NAA, 2,4-D, 2,4,5-T and 2,4-DP, inhib-ited PIN1 and PIN2 internalization with high efficiency, and cer-tain other compounds, including IAA itself, also exhibitedactivity (Fig. 7b). However, there was one exception among com-pounds active in both the inhibition of endocytosis and in otherassays, namely 5-F-IAA, which was entirely ineffective here(Fig. 7a,b). These results showed that, with the exception of 5-F-IAA, which induces DR5rev::GFP expression, but does notinhibit endocytosis (cf. Robert et al., 2010), auxins andauxin analogues active in DR5rev::GFP induction also inhibitendocytosis.
The observed inhibitory effect of 2-NAA on endocytosis doesnot agree with previous reports, in which experiments underslightly different conditions suggested that 2-NAA is not effectivecompared with NAA (Paciorek et al., 2005). Therefore, we com-pared the effects of 2-NAA and NAA at different concentrations.Our conditions with 25 lM BFA treatments are generally moresensitive than those used in previous studies. Furthermore, weestimated quantitatively the number of BFA bodies, which fur-ther increased the precision of our measurements. Using thesetechniques, we determined that the inhibitory effect of 2-NAA isindeed weaker than that of NAA, but that, at a concentrationhigher than 5 lM, both compounds are effective (Fig. S7).
Discussion
Physiological effects of auxins and their analogues
The introduction of different halogen atoms into variouspositions on the indole ring of IAA had a positive effect on theauxin-like activity in various bio-assays, for example, in the peacurvature test (Porter & Thimann, 1965), and, in particular,substitution by Cl at the 4 position of the indole ring to give 4-Cl-IAA had a strong stimulatory effect on oat and pea growthrelative to IAA itself (B€ottger et al., 1978; Katekar & Geisler,1983). In the wheat coleoptile straight growth bio-assay forauxins, 4-Cl-IAA, 5-Cl-IAA and 6-Cl-IAA were considerablymore active than IAA itself, their individual activities correspond-ing to their ability to displace labelled IAA bound to the PM-enriched microsomal fraction (Za�z�ımalova & Kuta�cek, 1985). Areport on pea fruit growth demonstrated the activity of 4-Cl-IAAand the inactivity of IAA itself (Reinecke et al., 1995).
Similarly, in the present study, the observed effect ofhalogenation on auxin-like activity was different in differentbioassays. In the primary root elongation inhibition assay inArabidopsis, 6-Cl-IAA was more active than IAA, whereas 5-Cl-IAA and 5-Br-IAA showed much lower activity. In lateral root for-mation, 5-Cl-IAA was much more active than any other haloge-nated derivative and IAA itself. A negative effect of halogenationappeared in 2,4,5-T, in which the addition of a further Cl atom to2,4-D at position 5 of the benzene ring reduced its auxin-likeactivity considerably. In contrast with this, the addition of amethyl group to the side-chain had little effect on activity in bothprimary root and lateral root assays, as seen in the case of 2,4-DP.
These findings show that halogenation at different ring positionsresults in distinct activities in various physiological processes.They also highlight the fine differences in the specificity patternsof these processes towards compounds which are not native and socould not have created selection pressure during evolution.
Some compounds showed activity only in some of the assaysperformed. Thus, IAA and NAA, which were very potent in rootassays, were not the most active auxins for the stimulation of BY-2cell division. Here, 2,4-D and related compounds were the mostactive, probably reflecting differential mechanisms for NAA and2,4-D action in BY-2 cells (Campanoni & Nick, 2005). Next, 2-NAA, a positional analogue of NAA, was quite effective in bothprimary root growth inhibition and the promotion of lateral rootsat higher concentrations. This observation is very interesting,because this compound was either considered as a very weak auxin(Katekar, 1979) or as an inactive analogue of the potent syntheticauxin NAA (Kepinski & Leyser, 2005). The compound corre-sponding to the basic core of the IAAmolecule – indole –inhibitedslightly primary root elongation at high concentrations, possiblyas a result of its likely role as a precursor for the tryptophan-inde-pendent IAA biosynthetic pathway (Woodward & Bartel, 2005).
As reported previously (Hayashi et al., 2008), lengthening ofthe side-chain of an auxin molecule reduces its auxin-like activity,and may explain why IBA and IPA are much weaker auxins thanIAA. However, and importantly, IBA was effective in the stimula-tion division of BY-2 cells, whereas IAA showed almost no activ-ity. There are currently two different views on the auxin-likeactivity of IBA: one notion considers IBA to be a precursor whichis converted to IAA by b-oxidation (Zolman et al., 2008; Strader& Bartel, 2011); the other suggests that IBA acts independentlyof IAA (Ludwig-M€uller, 2000). A recent study using stable iso-tope labelling of IAA and IBA showed that a considerableamount of IBA is converted to IAA in Arabidopsis hypocotyls(Liu et al., 2012). However, interestingly, the presence of endoge-nous IBA in Arabidopsis thaliana has been challenged recently(Nov�ak et al., 2012). Our study did not reveal any IAA convertedfrom [3H]IBA in tobacco cells; this could be either because themetabolic machinery in Arabidopsis and tobacco BY-2 cells dif-fers considerably in terms of conversion of IBA to IAA, orbecause the generated IAA is very rapidly (within a few seconds)converted to IAA metabolites.
Comparison of the structure–activity relationships for rootelongation, lateral root formation (both in Arabidopsis seedlings)and cell division (tobacco BY-2 cell line) highlights differences inthe structural requirements of these auxin-related physiologicalprocesses, thus making the differential phenotypic outcome ofthe same compound a very important aspect of auxin biology.
Intercellular auxin transport
In view of the results described above, it is not surprising that thesubstrate specificity of both auxin influx and efflux carriers differsbetween BY-2 and Arabidopsis cells. An interesting observation inseveral assays performed was that the activity of the major nativeauxin IAA was much lower than that of some of its synthetic andeven native analogues (typically 4-Cl-IAA). This could be
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 1034–1048
www.newphytologist.com
NewPhytologist Research 1043

(a) (d)
(b) (e)
(c) (f)
(g)
Fig. 8 Overview of effectiveness of different auxins in different assays. The auxin activities of all the tested compounds normalized in relation to therelevant controls are presented at a selected concentration for all assays performed. (a) Inhibition of primary root growth in Arabidopsis. Concentration ofcompounds 500 nM, normalized to IAA (100%). (b) Stimulation of lateral root formation in Arabidopsis. Concentration of compounds 500 nM, normalizedto IAA (100%). (c) Stimulation of cell division in tobacco BY-2 cells. Concentration of compounds 1 lM, normalized to 2,4-D (100%). (d) Competition foractive auxin influx. Concentration of compounds 10 lM, normalized to IAA (100%). (e) Competition for active auxin efflux. Concentration of compounds10 lM, normalized to NAA (100%). (f) Stimulation of DR5rev::GFP expression. Concentration of compounds 1 lM, normalized to IAA (100%). (g)Inhibition of endocytosis of PIN1 and PIN2 proteins. Concentration of compounds 25 lM, normalized to NAA (100%). See legend to Fig. 1 for compoundabbreviations.
New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist1044

explained by the fast metabolic turnover of IAA. With respect tothe fact that IAA is the major endogenous auxin, a well-organizedmetabolic machinery can be expected to be present, with preferen-tial affinity towards IAA. Fast metabolism of NAA and IAA hasbeen reported in tobacco cells (Nicotiana tabacum L., cv XanthiXHFD8 and cv BY-2; Delbarre et al., 1994 and Ho�sek et al.,2012, respectively). As these metabolic changes proceed insidecells, they should not affect the auxin influx assay in which IAA iscoming from outside (in its non-metabolized form); indeed, IAAshows a high degree of competition with the influx carrier ‘marker’2,4-D. In the auxin efflux assay, cold NAA competed with [3H]NAAmuch less than did some of the halogenated IAAs, suggestingthat endogenous auxin efflux carriers in BY-2 cells possess higheraffinity towards some halogenated IAAs relative to NAA. Carrier-mediated but AUX1- and PIN-independent transport of IBA hasbeen reported in Arabidopsis (Rashotte et al., 2003; Poupart et al.,2005; Yang et al., 2006). An interesting observation in this studywas that IBA consistently stimulated an increase (of c. 10%) in[3H]2,4-D accumulation in BY-2 cells. This may reflect reportsthat PDR9 (a member of the ABCG/PDR subfamily of ABC trans-porters) transports 2,4-D out of cells (Ito & Gray, 2006) and thatIBA also utilizes the same system for its efflux (Strader et al., 2008);our data may therefore reflect the competition of cold IBA for thePDR-like-driven export of [3H]2,4-D from cells, thus increasingits accumulation. However, in Arabidopsis cells, we did not observeany increase in [3H]2,4-D intracellular accumulation by IBA. Thismight be explained either by the different relative affinity of effluxcarriers towards IBA and 2,4-D in tobacco and Arabidopsis cellsand/or by the fact that the Arabidopsis cell suspension used herewas derived from stem explants (May & Leaver, 1993), whereasPDR9 genes are widely expressed only in roots (van den Brule &Smart, 2002; R�u�zi�cka et al., 2010). A parallel IBA transportmachinery has recently been proposed to play a role in auxinhomeostasis by actively transporting IBA across the PM as a storageform of IAA (Strader & Bartel, 2009, 2011; R�u�zi�cka et al., 2010).
Auxin signalling
Basically, auxin signalling pathways result in either the regulationof gene expression (TIR1-related, Calderon-Villalobos et al.,2010) or in effects which do not involve directly transcription ortranslation (mediated by ABP1, Badescu & Napier, 2006; Robertet al., 2010). In this study, the activation by auxin analogues ofDR5rev::GFP, reflecting the transcriptional control of auxin sig-nalling pathways, is in good agreement with Arabidopsis root phe-notyping. This is further supported by visible induction of theDR5-GFP signal by IAAla and IBA only after prolonged treat-ment and at higher concentrations, suggesting that this signalcould originate from the IAA released (Rampey et al., 2004) orconverted (Woodward & Bartel, 2005) from them. IBA itself doesnot seem to be a good substrate for the TIR1-mediated pathway,probably because of its longer side-chain (Hayashi et al., 2008).
The resistance of the tir1 mutant to primary root elongationinhibition by auxins was relatively low and may be caused by thefunctional redundancy between TIR1 and other members of theAFB family of auxin receptors, as reported previously (Parry et al.,
2009). Compared with the wild-type, the mutant was notablyresistant only to 2,4-D and its structural analogues, namely 2,4-DP and 2,4,5-T. The effect of 2,4,5-T was rather unexpectedbecause of its weak ability to induce DR5rev::GFP expression.Therefore, the auxin-like activity of 2,4,5-T could be attributedmainly to the interaction with the TIR1 co-receptor itself, and itsinteraction with other members of the AFB family may be verylow. A similar effect has been reported previously for a mutant ofthe TIR1 homologue AFB5, which is resistant to the picolinate-type auxin picloram, but sensitive to 2,4-D and IAA (Walsh et al.,2006). Moreover, the activity of 2,4,5-T might also reflect finetuning of the recently identified TIR1/AFB-Aux/IAA co-receptormachinery (Calderon Villalobos et al., 2012). As functionalredundancy obviously compromises the accuracy of specificitystudies performed in vivo, other methods, such as protein crystal-lography of individual proteins, as performed for TIR1 (Tanet al., 2007), can provide much better information about struc-ture–activity relationships for particular auxin (co-)receptors.
The inhibition of endocytosis was investigated to obtain aninsight into the ‘non-transcriptional’ auxin signalling pathway(s).It revealed the non-canonical behaviour of 5-F-IAA, which wasnot active in inhibiting endocytosis, even though it was very effec-tive in inducing a DR5-GFP signal (and also effective in certainother assays). A more detailed and focused study on the inhibitionof endocytosis of PM proteins has been reported by Robert et al.(2010). Interestingly, the inactivity of 5-F-IAA in the inhibitionof the endocytosis of PINs may also contribute to its lower activity(relative to other halogenated IAAs) in competition for activeauxin efflux: when endocytosis is not inhibited, there are fewerPIN efflux carriers on the PM for which to compete.
Altogether, this study shows that the auxin-like potential is avery complex property of a compound, determined by its chemi-cal and metabolic stability, ability to distribute in a tissue in apolar manner and its efficacy to activate various auxin signallingpathways. In Fig. 8, the auxin activities of selected compoundsare presented at selected concentrations and normalized in rela-tion to relevant controls. This figure shows that, except for thepresence of both aromatic (or heterocyclic) ring(s) and a carboxylgroup, no general criteria can be set for the specificity patterns ofdifferent auxin-related processes. The non-canonical behaviour ofsome synthetic auxin analogues is worth stressing because of theirpotential to be used as tools in further studies dissecting themolecular mechanism of auxin action. Thus, IPA and 2,4,5-Tmay serve to distinguish between TIR1 and other AFB protein-related processes, and 5-F-IAA can help to discriminate betweentranscriptional and non-transcriptional (ABP1-dependent) path-ways of auxin signalling. Last, but not least, this study has dem-onstrated that structure–activity relationships determinedprecisely at the level of a particular protein (receptor, carrier) maynot correspond fully to the final auxin-like physiological activityof a particular compound.
Acknowledgements
The authors thank Dr Christian Luschnig (University of NaturalResources and Life Sciences (BOKU), Vienna, Austria) for the
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 1034–1048
www.newphytologist.com
NewPhytologist Research 1045

anti-PIN2 antibody, Professor Mark Estelle (University of Cali-fornia, San Diego, CA, USA) for tir1-1 mutant seeds and, lastbut not least, to Dr David Morris for critical reading of the man-uscript. We also thank Mark�eta Pa�rezov�a and Jana St�yblov�a forexcellent technical assistance. This work was supported by theGrant Agency of the Czech Republic (P305/11/0797 to E.Z. and13-40637S to J.F.), the Central European Institute of Technol-ogy project CZ.1.05/1.1.00/02.0068 from the EuropeanRegional Development Fund and by a European ResearchCouncil starting independent research grant ERC-2011-StG-20101109-PSDP (to J.F.).
References
Abas L, Benjamins R, Malenica N, Paciorek T, Wi�sniewska J, Moulinier-Anzola
JC, Sieberer T, Friml J, Luschnig C. 2006. Intracellular trafficking and
proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root
gravitropism. Nature Cell Biology 8: 249–256.Badescu GO, Napier RM. 2006. Receptors for auxin: will it all end in TIRs?
Trends in Plant Science 11: 217–223.Benkov�a E, Michniewicz M, Sauer M, Teichmann T, Seifertov�a D, J€urgens G,
Friml J. 2003. Local, efflux-dependent auxin gradients as a common module
for plant organ formation. Cell 15: 591–602.Bennett MJ, Marchant A, Green HG, May ST, War SP, Millner PA, Walker
AR, Schulz B, Feldmann KA. 1996. Arabidopsis AUX1 gene: a permease-like
regulator of root gravitropism. Science 273: 948–950.B€ottger M, Engvild CK, Soll H. 1978. Growth of avena coleoptiles and pH drop
of protoplast suspensions induced by chlorinated indoleacetic acids. Planta140: 89–92.
van den Brule S, Smart CC. 2002. The plant PDR family of ABC transporters.
Planta 216: 95–106.Calderon Villalobos LI, Lee S, De Oliveira C, Ivetac A, Brandt W, Armitage L,
Sheard LB, Tan X, Parry G, Mao HB et al. 2012. A combinatorial TIR1/
AFB-Aux/IAA co-receptor system for differential sensing of auxin. NatureChemical Biology 8: 477–485.
Calderon-Villalobos LI, Tan X, Zheng N, Estelle M. 2010. Auxin perception –structural insights. Cold Spring Harbor Perspectives in Biology 2: a005546.
Campanoni P, Nick P. 2005. Auxin-dependent cell division and cell elongation.
1-Naphthaleneacetic acid and 2,4-dichlorophenoxyaceic acid activate different
pathways. Plant Physiology 137: 939–948.Chandler JW. 2009. Local auxin production: a small contribution to a big field.
BioEssays 31: 60–70.Christian M, Hannah WB, L€uthen H, Jones AM. 2008. Identification of
auxins by a chemical genomic approach. Journal of Experimental Botany 59:2757–2767.
Darwin C, Darwin F. 1980. The power of movement in plants. New York, NY,
USA: D. Appleton and Co.
Delbarre A, Muller P, Imhoff V, Guern J. 1996. Comparison of mechanisms
controlling uptake and accumulation of 2,4-dichlorophenoxyacetic acid,
naphthalene-1-acetic acid, and indole-3-acetic acid in suspension-cultured
tobacco cells. Planta 198: 532–541.Delbarre A, Muller P, Imhoff V, Morgat JL, Barbier-Brygoo H. 1994. Uptake,
accumulation and metabolism of auxins in tobacco leaf protoplasts. Planta 195:159–167.
Depta H, Rubery PH. 1984. A comparative study of carrier participation in the
transport of 2,3,5-triiodobenzoic acid, indole-3-acetic acid and
2,4-dichlorophenoxyacetic acid by Cucurbita pepo L. hypocotyls segments.
Journal of Plant Physiology 115: 371–387.Depuydt S, Hardtke CS. 2011.Hormone signalling crosstalk in plant growth
regulation. Current Biology 21: R365–R373.Dharmasiri N, Dharmasiri S, Estelle M. 2005. The F-box protein TIR1 is an
auxin receptor. Nature 435: 441–445.Dobrev PI, Havl�ı�cek L, V�agner M, Malbeck J, Kam�ınek M. 2005. Purification
and determination of plant hormones auxin and abscisic acid using solid phase
extraction and two-dimensional high performance liquid chromatography.
Journal of Chromatography A 1075: 159–166.Ferro N, Gallegos A, Bultinck P, Jacobsen HJ, Carbo-Dorca R, Reinard T.
2006. Coulomb and overlap self-similarities: a comparative selectivity analysis
of structure–function relationship for auxin-like molecules. Journal of ChemicalInformation and Modeling 46: 1751–1762.
Friml J, Vieten A, Sauer M, Weijers D, Schwarz H, Hamann T, Offringa R,
Jurgens G. 2003. Efflux-dependent auxin gradients establish the apical–basalaxis of Arabidopsis. Nature 426: 147–153.
Geldner N, Friml J, Stierhof YD, J€urgens G, Palme K. 2001. Auxin
transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 413:425–428.
Grossmann K. 2010. Auxin herbicides: current status of mechanism and mode of
action. Pest Management Science 66: 113–120.Grunewald W, Friml J. 2010. The march of the PINs: developmental plasticity
by dynamic polar targeting in plant cells. EMBO Journal 29: 2700–2714.Hayashi K, Tan X, Zheng N, Hatate T, Kimura Y, Kepinski S, Nozaki H. 2008.
Small-molecule agonists and antagonists of F-box protein–substrateinteractions in auxin perception and signaling. Proceedings of the NationalAcademy of Sciences, USA 105: 5632–5637.
Hertel R, Evan ML, Leopold AC, Sell HM. 1969. The specificity of the auxin
transport system. Planta 85: 238–249.Hertel R, Thomson KS, Russo EA. 1972. In-vitro auxin binding to particulate
cell fractions from corn coleoptiles. Planta 107: 325–340.Ho�sek P, Kube�s M, La�nkov�a M, Dobrev PI, Kl�ıma P, Kohoutov�a M, Petr�a�sek J,Hoyerov�a K, Ji�rina M, Za�z�ımalov�a E. 2012. Auxin transport on cellular level:
new insights supported by mathematical-modelling. Journal of ExperimentalBotany 63: 3815–3828.
Ito H, Gray WM. 2006. A gain-of-function mutation in the Arabidopsispleiotropic drug resistance transporter PDR9 confers resistance to auxinic
herbicides. Plant Physiology 142: 63–74.Katekar GF. 1979. Auxins: on the nature of the receptor site and molecular
requirements of auxin activity. Phytochemistry 18: 223–233.Katekar GF, Geisler AE. 1983. Structure–activity differences betweenindoleacetic acid auxins on pea and wheat. Phytochemistry 22: 27–31.
Kepinski S, Leyser O. 2005. The Arabidopsis F-box protein TIR1 is an auxin
receptor. Nature 435: 446–451.Koepfli JB, Thimann KV, Went FW. 1938. Phytohormones: structure and
physiological activity. Ι. Journal of Biological Chemistry 122: 763–780.K€ogl F, Haagen-Smit AJ, Erxleben H. 1934. €Uber die neues auxin (heteroauxin)
aus hain. XI Mitteilung. Zeitschrift f€ur Physiologische Chemie 228: 90–103.K�re�cek P, Sk�upa P, Libus J, Naramoto S, Tejos R, Friml J, Za�z�ımalov�a E. 2009.
The PIN-FORMED (PIN) protein family of auxin transporters. GenomeBiology 10: 249.
La�nkov�a M, Smith RS, Pe�sek B, Kube�s M, Za�z�ımalov�a E, Petr�a�sek J, Hoyerov�a
K. 2010. Auxin influx inhibitors 1-NOA, 2-NOA and CHPAA interfere
with membrane dynamics in tobacco cells. Journal of Experimental Botany 61:3589–3698.
Leyser O. 2011. Auxin, self-organisation, and the colonial nature of plants.
Current Biology 21: R331–R337.Liu X, Barkawi L, Gardner G, Cohen JD. 2012. Transport of indole-3-butyric
acid and indole-3-acetic acid in Arabidopsis thaliana hypocotyls using stableisotope labelling. Plant Physiology 158: 1988–2000.
Ljung K. 2013. Auxin metabolism and homeostasis during plant development.
Development 140: 943–950.Ljung K, Hull AK, Kowalczyk M, Marchant A,Celenza J, Cohen JD, Sandberg
G. 2002. Biosynthesis, conjugation, catabolism and homeostasis of
indole-3-acetic acid in Arabidopsis thaliana. Plant Molecular Biology 49:249–272.
L€obler M, Kl€ambt D. 1985. Auxin-binding proteins from coleoptiles membranes
of corn (Zea mays L). I. Purification by immunological methods and
characterization. Journal of Biological Chemistry 260: 9848–9853.Loper MT, Spanswick RM. 1991. Auxin transport in suspension-cultured
soybean root cells: II. Anion effects on carrier-mediated uptake. Plant Physiology96: 192–197.
Ludwig-M€uller J. 2000. Indole-3-butyric acid in plant growth and development.
Plant Growth Regulation 32: 219–230.
New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist1046

Martin HV, Pilet PE. 1986. Saturable uptake of indol-3yl-acetic acid by maize
roots. Plant Physiology 81: 889–895.May MJ, Leaver CJ. 1993.Oxidative stimulation of glutathione synthesis in
Arabidopsis thaliana suspension cultures. Plant Physiology 103: 621–627.M€uller F, Appleby AP. 2010.Weed control, 2. Individual herbicides. In: Elvers
B, ed. Ullmann’s Encyclopedia of Industrial Chemistry. New York, NY, USA:
John Wiley & Sons, Inc., Wiley Online Library, doi: 10.1002/14356007.
o28_o01.
Nagata T, Nemoto Y,Hasezawa S. 1992.Tobacco BY-2 cell line as the “HeLa” cell
in the cell biology of higher plants. International Review of Cytology 132: 1–30.Novak O, Henykova E, Sairanen I, Kowalczyk M, Pospısil T, Ljung K. 2012.
Tissue-specific profiling of the Arabidopsis thaliana auxin metabolome. PlantJournal 72: 523–536.
Paciorek T, Za�z�ımalov�a E, Ruthardt N, Petr�a�sek J, Stierhof YD, Kleine-Vehn J,
Morris DA, Emans N, J€urgens G, Geldner N et al. 2005. Auxin inhibits
endocytosis and promotes its own efflux from cells. Nature 435: 1251–1256.Parry G, Calderon-Villalobos LI, Prigge M, Peret B, Dharmasiri S, Itoh H,
Lechner E, Gray WM, Bennett M, Estelle M. 2009. Complex regulation of
the TIR1/AFB family of auxin receptors. Proceedings of the National Academy ofSciences, USA 106: 22540–22545.
Parry G, Delbarre A, Marchant A, Swarup R, Napier R, Perrot-Rechenmann C,
Bennett MJ. 2001. Novel auxin transport inhibitors phenocopy the auxin
influx carrier mutation aux1. Plant Journal 25: 399–406.Petr�a�sek J, El�ckner M, Morris DA, Za�z�ımalov�a E. 2002. Auxin efflux carrier
activity and auxin accumulation regulate cell division and polarity in tobacco
cells. Planta 216: 302–308.Petr�a�sek J, Friml J. 2009. Auxin transport routes in plant development.
Development 36: 2678–2688.Petr�a�sek J, Mravec J, Bouchard R, Blakeslee JJ, Abas M, Seifertov�a D,
Wisniewska J, Tadele Z, Kube�s M, �Covanov�a M et al. 2006. PINproteins perform a rate-limiting function in cellular auxin efflux. Science 312:914–918.
Porter WL, Thimann KV. 1965.Molecular requirements for auxin action – I.Halogenated indoles and indoleacetic acid. Phytochemistry 4: 229–243.
Poupart J, Rashotte AM, Muday GK, Waddell CS. 2005. The rib1 mutant of
Arabidopsis has alterations in indole-3-butyric acid transport, hypocotyl
elongation, and root architecture. Plant Physiology 39: 1460–1470.Rampey RA, LeClere S, Kowalczyk M, Ljung K, Sandberg G, Bartel B. 2004.
A family of auxin-conjugate hydrolases that contributes to free
indole-3-acetic acid levels during Arabidopsis germination. Plant Physiology35: 978–988.
Rashotte AM, Poupart J, Waddell CS, Muday GK. 2003. Transport of the two
natural auxins, indole-3-butyric acid and indole-3-acetic acid, in Arabidopsis.Plant Physiology 133: 761–772.
Reinecke DM, Ozga JA, Magnus V. 1995. Effect of halogenated substitution of
indole-3-acetic acid on biological activity of pea fruit. Phytochemistry 40: 1361–1366.
Robert S, Kleine-Vehn J, Barbez E, Sauer M, Paciorek T, Baster P, Vanneste S,
Zhang J, Simon S, �Covanov�a M et al. 2010. ABP1 mediates auxin inhibition
of clathrin-dependent endocytosis in plants. Cell 143: 111–121.Rosquete MR, Barbez E, Kleine-Vehn J. 2012. Cellular auxin homeostasis:
gatekeeping is housekeeping.Molecular Plant 5: 772–786.Rubery PH. 1977. The specificity of carrier-mediated auxin transport by
suspension-cultured crown gall cells. Planta 135: 275–283.Ruegger M, Dewey E, Gray WM, Hobbie L, Turner J, Estelle M. 1998. The
TIR1 protein of Arabidopsis functions in auxin response and is related to
human SKP2 and yeast grr1p. Genes & Development 12: 198–207.R�u�zi�cka K, Strader LC, Bailly A, Yang H, Blakeslee J, Łangowski L, Nejedl�a E,Fujita H, Ito H, Sy�ono K et al. 2010. Arabidopsis PIS1 encodes the ABCG37
transporter of auxinic compounds including the auxin precursor
indole-3-butyric acid. Proceedings of the National Academy of Sciences, USA 107:
10749–10753.Sabater M, Rubery PH. 1987. Auxin carriers in Cucurbita vesicles III. Specificity,with particular references to 1-naphthylacetic acid. Planta 171: 514–518.
Sakai A, Miyazawa Y, Kuroiwa T. 2004. Studies on dynamic changes
of organelles using tobacco BY-2 as the model plant cell line. In:
Nagata T, Hasezawa S, Inz�e D, eds. Biotechnology in agriculture and forestry
vol. 53, Tobacco BY-2 cells. Berlin, Heidelberg, Germany: Springer,
192–216.Stasinopoulos TC, Hangarter RP. 1990. Preventing photochemistry in culture
media by long-pass light filters alters growth of cultured tissues. Plant Physiology93: 1365–1369.
Strader LC, Bartel B. 2009. The Arabidopsis PLEIOTROPIC DRUG
RESISTANCE8/ABCG36 ATP binding cassette transporter modulates
sensitivity to the auxin precursor indole-3-butyric acid. Plant Cell 21: 1992–2007.
Strader LC, Bartel B. 2011. Transport and metabolism of the endogenous auxin
precursor indole-3-butyric acid.Molecular Plant 4: 477–486.Strader LC, Culler AH, Cohen JD, Bartel B. 2010. Conversion of endogenous
indole-3-butyric acid to indole-3-acetic acid drives cell expansion in
Arabidopsis seedlings. Plant Physiology 153: 1577–1586.Strader LC, Monroe-Augustus M, Rogers KC, Lin GL, Bartel B. 2008.
Arabidopsis iba response5 suppressors separate responses to various hormones.
Genetics 180: 2019–2031.Swarup K, Benkov�a E, Swarup R, Casimiro I, P�eret B, Yang Y, Parry G, Nielsen
E, De Smet I, Vanneste S et al. 2008. The auxin influx carrier LAX3 promotes
lateral root emergence. Nature Cell Biology 10: 946–954.Tan X, Calderon-Villalobos LI, Sharon M, Zheng C, Robinson CV, Estelle M,
Zheng N. 2007.Mechanism of auxin perception by the TIR1 ubiquitin ligase.
Nature 446: 640–645.Ulmasov T, Murfett J, Hagen G, Guilfoyle TJ. 1997. Aux/IAA proteins repress
expression of reporter genes containing natural and highly active synthetic
auxin response elements. Plant Cell 9: 1963–1971.Walsh TA, Neal R, Merlo AO, Honma M, Hicks GR, Wolff K, Matsumura M,
Davies JP. 2006.Mutations in an Auxin Receptor Homolog AFB5 and in
SGT1b confer resistance to synthetic picolinate auxins and not to
2,4-dichlorophenoxyacetic acid or indole-3-acetic acid in Arabidopsis. PlantPhysiology 142: 542–552.
Went FW, Thimann KV. 1937. Phytohormones. New York, NY, USA:
Macmillan.
Woodward AW, Bartel B. 2005. Auxin: regulation, action, and interaction.
Annals of Botany 95: 707–735.Yang H, Murphy AS. 2009. Functional expression and characterization of
Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomycespombe. Plant Journal 59: 179–191.
Yang Y, Hammes UZ, Taylor CG, Schachtman DP, Nielsen E. 2006.
High-affinity auxin transport by the AUX1 influx carrier protein. CurrentBiology 16: 1123–1127.
Za�z�ımalova E, Kuta�cek M. 1985. Auxin-binding site in wheat shoots:
interactions between indol-3-ylacetic acid and its halogenated derivatives.
Biologia Plantarum 27: 114–118.Zimmerman PW, Wilcoxon F. 1935. Several chemical growth substances which
cause initiation of root and other responses in plants. Contributions of the BoyceThompson Institute 7: 209–229.
Zolman BK, Martinez N, Millius A, Adham AR, Bartel B. 2008. Identification
and characterization of Arabidopsis indole-3-butyric acid response mutants
defective in novel peroxisomal enzymes. Genetics 180: 237–251.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Chemical structure of auxins and auxin analogues used inthis study.
Fig. S2 Metabolic profile of indole-3-butyric acid (IBA) intobacco BY-2 suspension-grown cells.
Fig. S3 Representative curves depicting the accumulation of aux-ins and auxin analogues in tobacco BY-2 cells.
� 2013 The Authors
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 200: 1034–1048
www.newphytologist.com
NewPhytologist Research 1047

Fig. S4 Active transport of auxin and its analogues across theplasma membrane in Arabidopsis thaliana, ecotype Landsbergerecta cells.
Fig. S5 Representative curves depicting the accumulation of aux-ins and auxin analogues in Arabidopsis thaliana, ecotype Lands-berg erecta cells.
Fig. S6 Activity of auxins and auxin analogues in the stimulationof DR5-driven gene expression at higher concentration and pro-longed treatment.
Fig. S7 Activity of naphthalene-1-acetic acid (NAA) and naph-thalene-2-acetic acid (2-NAA) in the inhibition of endocytosis atdifferent concentrations.
Please note: Wiley Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
New Phytologist is an electronic (online-only) journal owned by the New Phytologist Trust, a not-for-profit organization dedicatedto the promotion of plant science, facilitating projects from symposia to free access for our Tansley reviews.
Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged. We are committed to rapid processing, from online submission through to publication ‘as ready’ via Early View – our average timeto decision is <25 days. There are no page or colour charges and a PDF version will be provided for each article.
The journal is available online at Wiley Online Library. Visit www.newphytologist.com to search the articles and register for tableof contents email alerts.
If you have any questions, do get in touch with Central Office ([email protected]) or, if it is more convenient,our USA Office ([email protected])
For submission instructions, subscription and all the latest information visit www.newphytologist.com
New Phytologist (2013) 200: 1034–1048 � 2013 The Authors
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist1048




















