
Decomposition of 13C-labelled plant material in a European 65-40º latitudinal transect of...
17
Decomposition of 13 C-labelled plant material in a European 65– 408 latitudinal transect of coniferous forest soils: simulation of climate change by translocation of soils Pierre Bottner a, *, Marie-Madeleine Couˆteaux a , Jonathan M. Anderson b , Bjo¨rn Berg c , Georges Bille`s a , Tom Bolger d , Herve´ Casabianca e , Joan Romanya´ f , Pere Rovira f a CEFE-CNRS, 1919 Route de Mende, 34293 Montpellier Cedex 5, France b Department of Biological Sciences, Hatherly Laboratories, University of Exeter, Prince of Wales Road, Exeter EX4 4PS, UK c Department of Forest Soils, Swedish University of Agricultural Sciences, P.O. Box 7001, S-750 07 Uppsala, Sweden d Department of Zoology, University of Dublin, Belfield, Dublin 4, Ireland e Service Central d’Analyse, CNRS, BP 22, 69390 Vernaison, France f Dept Biologia Vegetal, Universitat de Barcelona, 645 Diagonal, 08028 Barcelona, Spain Accepted 27 September 1999 Abstract Standard 13 C-labelled plant material was exposed over 2–3 yr at 8 sites in a north–south climatic gradient of coniferous forest soils, developed on acid and calcareous parent materials in Western Europe. In addition to soils exposed in their sites of origin, replicate units containing labelled material were translocated in a cascade sequence southwards along the transect, to simulate the eects of climate warming on decomposition processes. The current Atlantic climate represented the most favourable soil temperature and moisture conditions for decomposition. Northward this climatic zone, where the soil processes are essentially temperature-limited, the prediction for a temperature increase of 38C estimated a probable increase of C mineralisation by 20– 25% for the boreal zone and 10% for the cool temperate zone. Southward the cool Atlantic climate zone, (the Mediterranean climate), where the processes are seasonally moisture-limited, the predicted increase of temperature by 1–28C little aected the soil organic matter dynamics, because of the higher water deficit. A significant decrease of C mineralisation rates was observed only in the superficial layers recognised in Mediterranean forest soils as ‘xeromoder’ and subject to frequent dry conditions. In the deeper Mediterranean soil organic horizons (the mull humus types), representing the major C storage in this zone, C mineralisation was not aected by a simulated 28C temperature increase. The temperature eect is probably counteracted by a higher water deficit. 7 2000 Elsevier Science Ltd. All rights reserved. Keywords: Decomposition; Carbon; Coniferous forests; 13 C-labelling; Climate change; North–south transect; Translocation; Organic matter; Car- bon mineralisation; Forest soils; Europe; Tracer techniques 1. Introduction One of the key issues in climate change research is the future dynamics of organic carbon in soils which contains about two-thirds of the total organic C in terrestrial systems. Even small changes of the mineralisation rates of these large soil pools could therefore have significant eects on concentrations of atmospheric CO 2 . As yet there are few indications as to whether soils will be net sources of CO 2 , since climate warming increases mineralisation rates, or net sinks for C as a consequence of the CO 2 eect on plant litter production, quality and decomposition rates (Couˆteaux et al., 1991; Cotrufo et al., 1994). A number of dierent approaches have been used to ob- Soil Biology & Biochemistry 32 (2000) 527–543 0038-0717/00/$ - see front matter 7 2000 Elsevier Science Ltd. All rights reserved. PII: S0038-0717(99)00182-0 www.elsevier.com/locate/soilbio * Corresponding author. Fax: +33-4-6741-2138. E-mail address: [email protected] (P. Bottner).
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Decomposition of 13C-labelled plant material in a European 65-40º latitudinal transect of...

Decomposition of 13C-labelled plant material in a European 65±408 latitudinal transect of coniferous forest soils: simulation of
climate change by translocation of soils
Pierre Bottnera,*, Marie-Madeleine Couà teauxa, Jonathan M. Andersonb, BjoÈ rn Bergc,Georges BilleÁ s a, Tom Bolgerd, Herve Casabiancae, Joan Romanya f, Pere Roviraf
aCEFE-CNRS, 1919 Route de Mende, 34293 Montpellier Cedex 5, FrancebDepartment of Biological Sciences, Hatherly Laboratories, University of Exeter, Prince of Wales Road, Exeter EX4 4PS, UK
cDepartment of Forest Soils, Swedish University of Agricultural Sciences, P.O. Box 7001, S-750 07 Uppsala, SwedendDepartment of Zoology, University of Dublin, Bel®eld, Dublin 4, Ireland
eService Central d'Analyse, CNRS, BP 22, 69390 Vernaison, FrancefDept Biologia Vegetal, Universitat de Barcelona, 645 Diagonal, 08028 Barcelona, Spain
Accepted 27 September 1999
Abstract
Standard 13C-labelled plant material was exposed over 2±3 yr at 8 sites in a north±south climatic gradient of coniferous forestsoils, developed on acid and calcareous parent materials in Western Europe. In addition to soils exposed in their sites of origin,
replicate units containing labelled material were translocated in a cascade sequence southwards along the transect, to simulatethe e�ects of climate warming on decomposition processes. The current Atlantic climate represented the most favourable soiltemperature and moisture conditions for decomposition. Northward this climatic zone, where the soil processes are essentiallytemperature-limited, the prediction for a temperature increase of 38C estimated a probable increase of C mineralisation by 20±
25% for the boreal zone and 10% for the cool temperate zone. Southward the cool Atlantic climate zone, (the Mediterraneanclimate), where the processes are seasonally moisture-limited, the predicted increase of temperature by 1±28C little a�ected thesoil organic matter dynamics, because of the higher water de®cit. A signi®cant decrease of C mineralisation rates was observed
only in the super®cial layers recognised in Mediterranean forest soils as `xeromoder' and subject to frequent dry conditions. Inthe deeper Mediterranean soil organic horizons (the mull humus types), representing the major C storage in this zone, Cmineralisation was not a�ected by a simulated 28C temperature increase. The temperature e�ect is probably counteracted by a
higher water de®cit. 7 2000 Elsevier Science Ltd. All rights reserved.
Keywords: Decomposition; Carbon; Coniferous forests; 13C-labelling; Climate change; North±south transect; Translocation; Organic matter; Car-
bon mineralisation; Forest soils; Europe; Tracer techniques
1. Introduction
One of the key issues in climate change researchis the future dynamics of organic carbon in soilswhich contains about two-thirds of the total organicC in terrestrial systems. Even small changes of the
mineralisation rates of these large soil pools could
therefore have signi®cant e�ects on concentrations
of atmospheric CO2. As yet there are few indications
as to whether soils will be net sources of CO2, since
climate warming increases mineralisation rates, or net
sinks for C as a consequence of the CO2 e�ect on
plant litter production, quality and decomposition
rates (Couà teaux et al., 1991; Cotrufo et al., 1994). A
number of di�erent approaches have been used to ob-
Soil Biology & Biochemistry 32 (2000) 527±543
0038-0717/00/$ - see front matter 7 2000 Elsevier Science Ltd. All rights reserved.
PII: S0038-0717(99 )00182 -0
www.elsevier.com/locate/soilbio
* Corresponding author. Fax: +33-4-6741-2138.
E-mail address: [email protected] (P. Bottner).

serve these trends which involve ®eld measurements,
manipulative experiments and simulation models.
The net changes in the balances between C input
to soils and mineralisation rates are generally toosmall to detect by direct measurements, because of
the variability of the ¯uxes and pools. Using indir-
ect evidence, a large current terrestrial CO2 sink inthe northern hemisphere, referred as `missing sink' was
indicated (i) by atmospheric chemistry measurements:
CO2 pressure gradients (Tans et al., 1990), CO213C/12C ratio (Ciais et al., 1995) and O2 concentration
(Keeling et al., 1996) and (ii) by analysis of climatevariability during the last decades (Dai and Fung,
1993).
The e�ect of climate controls over soil Cdynamics at geographical scales have been investi-
gated using a number of di�erent approaches. Soilrespiration data have been extensively employed in
empirically-based statistical models, to predict
annual and global CO2 emission from terrestrial soils(Raich and Schlesinger, 1992) and to de®ne the spatial
and temporal climate controls of soil respiration
(Raich and Potter, 1995). Since a possible imbalance inthe ecosystem C cycle arises from the di�ering re-
sponses of production and decomposition to tempera-
ture change, the mechanistic models are generallybased on coupled production and decomposition sub-
models. A relatively simple model developed by Town-
send et al. (1992) describes the temperature e�ect onnet ecosystem production using a linear function and
an exponential function for response by soil respir-
ation. More complex decomposition models partitionthe organic matter into multiple C and N pools with
speci®c turnover rates (Jenkinson et al., 1991; Schimel
et al., 1994). Production- and decomposition-submo-dels have been linked by the litter quality and plant
detritus chemistry (C-to-N ratio and lignin content)and by the nitrogen cycle (the dynamics of N mineral-
isation controlling N uptake by the plants). The spatial
dimension of the decomposition models necessitatedthe integration of the geographical distribution of soil
types and some essential intrinsic soil properties.
In our study 13C- and 15N-labelled standard plantmaterial was exposed, at eight sites along a north±
south climatic gradient of coniferous forest soils inWestern Europe, which included boreal, Atlantic and
Mediterranean climates. In addition, the soils with
their labelled plant material were translocated fromnorth to south according to a cascade procedure, in
order to simulate a south to north climate shift. The
objective was to investigate the decomposition pro-cesses in the climatic transect, where the current cli-
matic spatial di�erences were used as an analogue for
expected climate change. This publication only pre-sents results for 13C dynamics.
2. Materials and methods
2.1. The sites and humus types
Within each climatic zone, except for boreal, twosites were identi®ed (on acidic and calcareous parentmaterials; Fig. 1 and Table 1). In boreal coniferousforests the surface organic layers are generally of lowpH irrespective of the base status of the underlyingparent material. The complement of sites detailed inTable 1 included representatives of the major humustypes developed under coniferous forest stands onwell-drained soils in Western Europe. The series ofhumus types developed on acid soils included, fromnorth to south: `mor' (Oh horizons at Vindeln andJaÈ draaÊ s), acid `mull' (A1 horizons at Haldon, The zanand Desert) and acid `xeromoder' (Oh at The zan). Thecalcareous soils series comprised acid `mull' at Friston,`calcic mull' at La Clape (where the exchange capacitywas saturated by Ca2+ without the presence of car-bonates), `calcareous mull' at Maials (Ca2+ saturatedexchange capacity with presence of carbonates) and®nally neutral `xeromoder' at la Clape and Maials.
Mor and xeromoder are Oh horizons where the or-ganic layer with a high C content, and high C-to-Nratio (Fig. 2) is developed on the surface of the min-eral soil, resulting from the slow turnover rates of lowquality litters under temperature-limited conditions inthe boreal region (mor) and under moisture limitationin the Mediterranean region (xeromoder). The mullsoils are mainly developed under Atlantic and Mediter-ranean conditions where organic matter decomposingat higher rates is incorporated into the mineral soil, byfaunal activity, forming stabilised organo-mineral com-plexes. On acid soils, the pH (H2O) values of mull are5±6, irrespective of the climatic conditions (Fig. 2). Inthe calcareous soils sequence, the pH values, the Ca2+
saturation and the carbonate concentrations in themull soils increase from north to south, indicating adecreasing capacity of dissolution and leaching of car-bonates from the organic horizon. At The zan, LaClape and Maials, the Oh horizon (xeromoder) over-lies the A1 horizon (mull). This is a common situationunder Mediterranean conditions (Fig. 2).
2.2. 13C and 15N soil labelling and ®eld incubation incylinders
The soil was divided into pedological horizons(Fig. 2) de®ned by the distribution of organic matterin the pro®le. The material from each horizon wassieved (4 mm mesh) and thoroughly homogenised. Thematerial from the horizons selected for labelling wasair-dried in the laboratory before the labelled plantmaterial was added to the soil. The moist materialsfrom the unlabelled horizons were placed in increments
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543528
in plastic cylinders (inside dia 12 cm, length 30 cm)and packed down with a heavy metal ram to reconsti-tute a bulk density close to the ®eld value. Discs of 1mm mesh polyester netting were placed between thehorizons to facilitate sampling. The depth and mass ofthe horizons established for each experimental unit aregiven in Fig. 2.
The labelled plant material was produced by grow-ing wheat (`Florence Aurore', an old spring-wheat cul-tivar with low N requirements) over 4 months, in a
labelling chamber with facilities for maintaining tem-perature, radiation, moisture and CO2 conditions, in anutrient solution with low 15N, P and K concen-trations, plus micro-nutrients, under a 13CO2-labelledatmosphere. In order to obtain material with a high C-to-N ratio, only the stems and leaves were used in theexperiment. The harvested plant material was air-dried, milled into 2±7 mm long ®brous particles andhomogenised.
The labelled plant material was added separately to
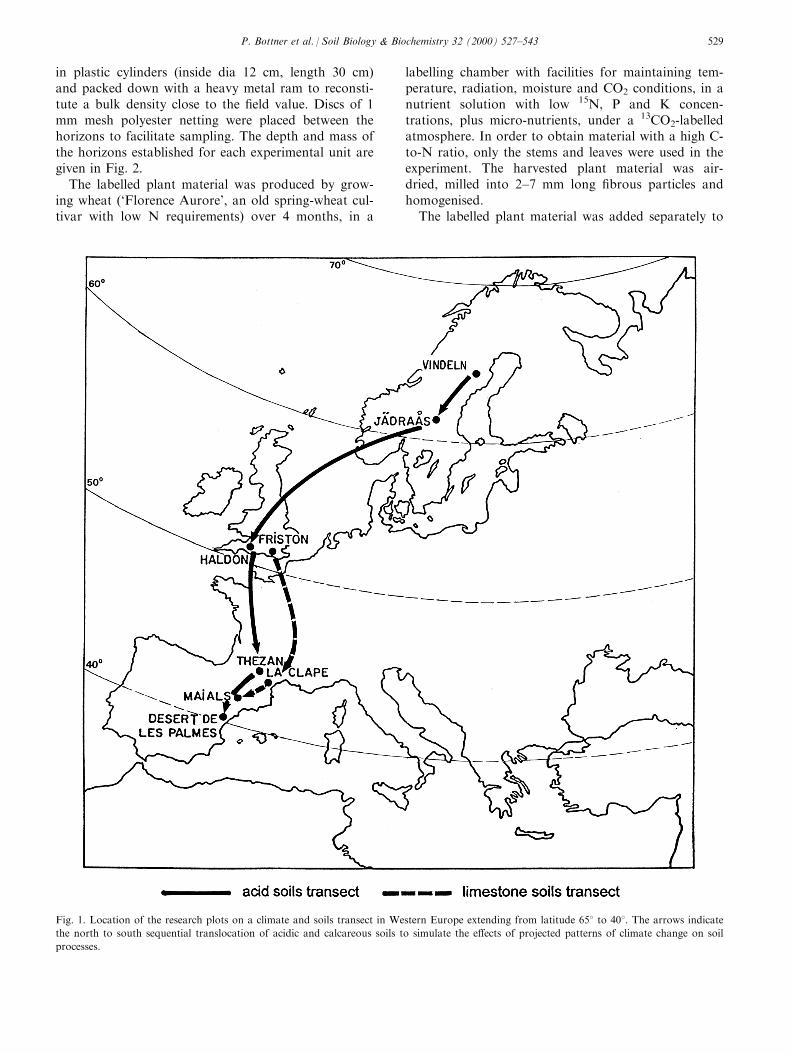
Fig. 1. Location of the research plots on a climate and soils transect in Western Europe extending from latitude 658 to 408. The arrows indicate
the north to south sequential translocation of acidic and calcareous soils to simulate the e�ects of projected patterns of climate change on soil
processes.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 529
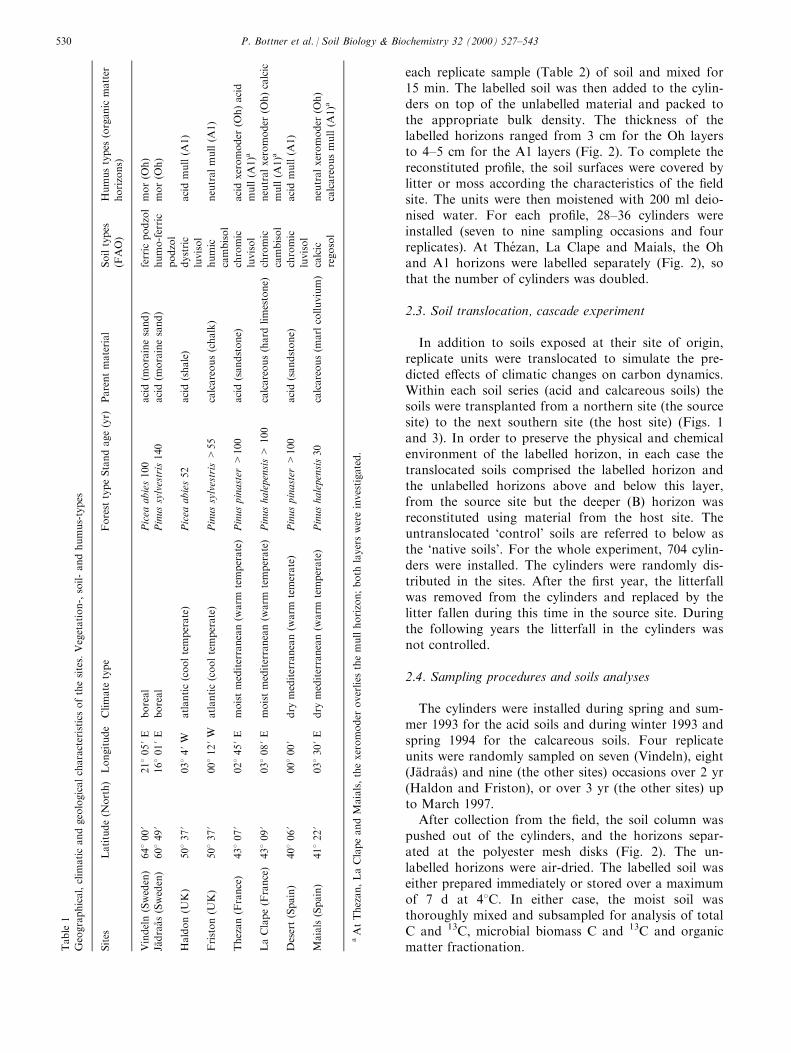
each replicate sample (Table 2) of soil and mixed for15 min. The labelled soil was then added to the cylin-ders on top of the unlabelled material and packed tothe appropriate bulk density. The thickness of thelabelled horizons ranged from 3 cm for the Oh layersto 4±5 cm for the A1 layers (Fig. 2). To complete thereconstituted pro®le, the soil surfaces were covered bylitter or moss according the characteristics of the ®eldsite. The units were then moistened with 200 ml deio-nised water. For each pro®le, 28±36 cylinders wereinstalled (seven to nine sampling occasions and fourreplicates). At The zan, La Clape and Maials, the Ohand A1 horizons were labelled separately (Fig. 2), sothat the number of cylinders was doubled.
2.3. Soil translocation, cascade experiment
In addition to soils exposed at their site of origin,replicate units were translocated to simulate the pre-dicted e�ects of climatic changes on carbon dynamics.Within each soil series (acid and calcareous soils) thesoils were transplanted from a northern site (the sourcesite) to the next southern site (the host site) (Figs. 1and 3). In order to preserve the physical and chemicalenvironment of the labelled horizon, in each case thetranslocated soils comprised the labelled horizon andthe unlabelled horizons above and below this layer,from the source site but the deeper (B) horizon wasreconstituted using material from the host site. Theuntranslocated `control' soils are referred to below asthe `native soils'. For the whole experiment, 704 cylin-ders were installed. The cylinders were randomly dis-tributed in the sites. After the ®rst year, the litterfallwas removed from the cylinders and replaced by thelitter fallen during this time in the source site. Duringthe following years the litterfall in the cylinders wasnot controlled.
2.4. Sampling procedures and soils analyses
The cylinders were installed during spring and sum-mer 1993 for the acid soils and during winter 1993 andspring 1994 for the calcareous soils. Four replicateunits were randomly sampled on seven (Vindeln), eight(JaÈ draaÊ s) and nine (the other sites) occasions over 2 yr(Haldon and Friston), or over 3 yr (the other sites) upto March 1997.
After collection from the ®eld, the soil column waspushed out of the cylinders, and the horizons separ-ated at the polyester mesh disks (Fig. 2). The un-labelled horizons were air-dried. The labelled soil waseither prepared immediately or stored over a maximumof 7 d at 48C. In either case, the moist soil wasthoroughly mixed and subsampled for analysis of totalC and 13C, microbial biomass C and 13C and organicmatter fractionation.T
able
1
Geographical,clim
aticandgeologicalcharacteristics
ofthesites.Vegetation-,soil-andhumus-types
Sites
Latitude(N
orth)
Longitude
Climate
type
Forest
typeStandage(yr)
Parentmaterial
Soiltypes
(FAO)
Humustypes
(organic
matter
horizons)
Vindeln(Sweden)
64800'
21805'E
boreal
Picea
abies100
acid(m
orainesand)
ferric
podzol
mor(O
h)
JaÈdraaÊs(Sweden)
60849'
16801'E
boreal
Pinussylvestris140
acid(m
orainesand)
humo-ferric
podzol
mor(O
h)
Haldon(U
K)
50837'
0384'W
atlantic(cooltemperate)
Picea
abies52
acid(shale)
dystric
luvisol
acidmull(A
1)
Friston(U
K)
50837'
00812'W
atlantic(cooltemperate)
Pinussylvestris>
55
calcareous(chalk)
humic
cambisol
neutralmull(A
1)
Thezan(France)
43807'
02845'E
moistmediterranean(w
arm
temperate)
Pinuspinaster
>100
acid(sandstone)
chromic
luvisol
acidxeromoder
(Oh)acid
mull(A
1)a
LaClape(France)
43809'
03808'E
moistmediterranean(w
arm
temperate)
Pinushalepensis>
100
calcareous(hard
limestone)
chromic
cambisol
neutralxeromoder
(Oh)calcic
mull(A
1)a
Desert(Spain)
40806'
00800'
dry
mediterranean(w
arm
temerate)
Pinuspinaster
>100
acid(sandstone)
chromic
luvisol
acidmull(A
1)
Maials(Spain)
41822'
03830'E
dry
mediterranean(w
arm
temperate)
Pinushalepensis30
calcareous(m
arlcolluvium)
calcic
regosol
neutralxeromoder
(Oh)
calcareousmull(A
1)a
aAtThezan,LaClapeandMaials,thexeromoder
overlies
themullhorizon;both
layerswereinvestigated.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543530
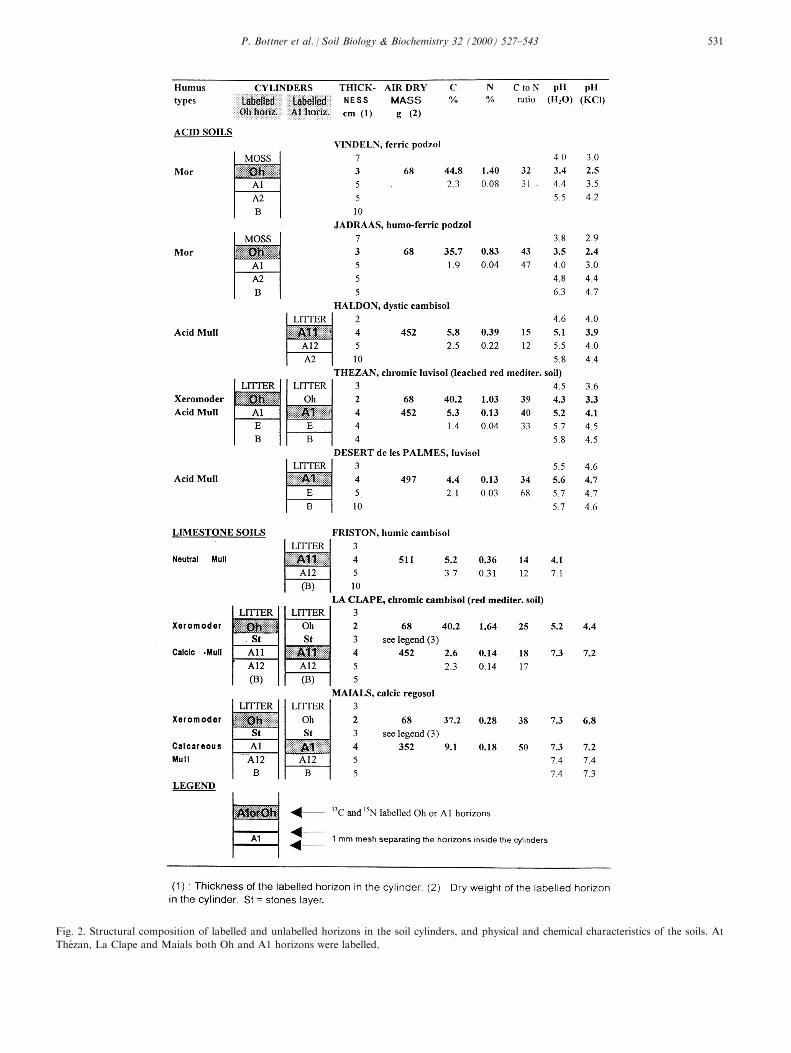
Fig. 2. Structural composition of labelled and unlabelled horizons in the soil cylinders, and physical and chemical characteristics of the soils. At
The zan, La Clape and Maials both Oh and A1 horizons were labelled.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 531
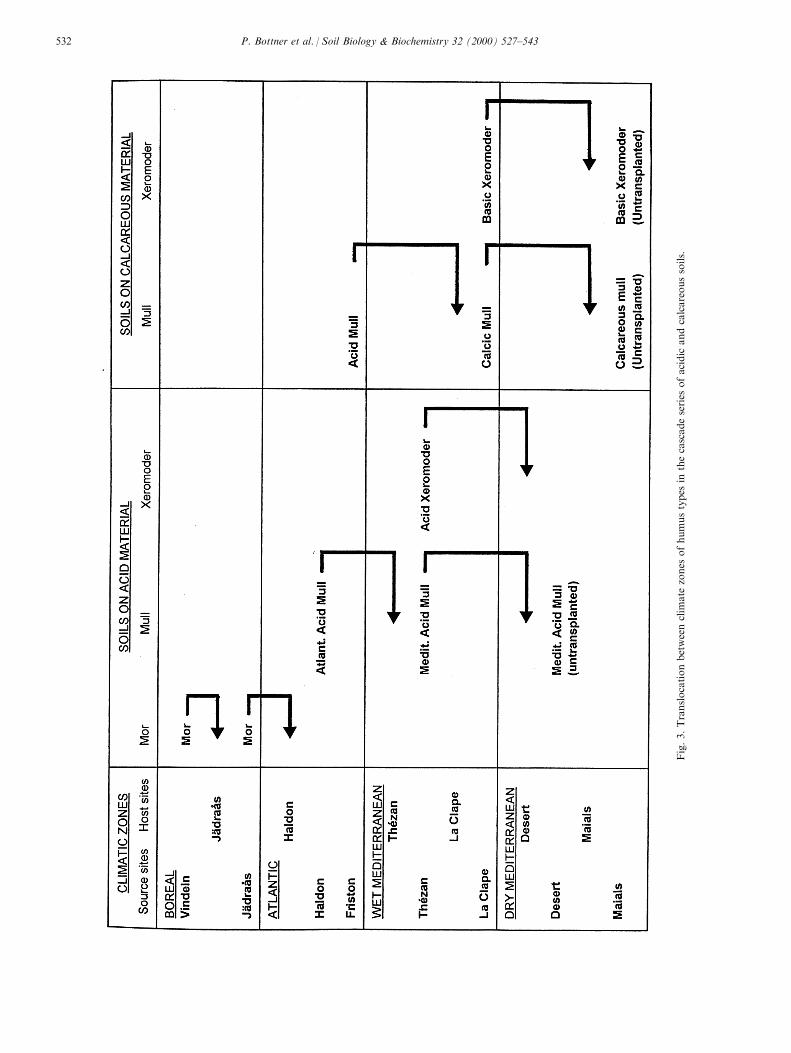
Fig.3.Translocationbetweenclim
ate
zones
ofhumustypes
inthecascadeseries
ofacidic
andcalcareoussoils.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543532

2.5. C, N and mass spectrometry analysis
Since highly labelled plant material was added to
the soils in small amounts, preparation methods
were investigated to determine the sample homogen-
eity required to reduce variation in results of mass
spectrometry. The following procedures were carried
out on the labelled horizons and on the soil ma-
terials above and below this layer.
For the mineral soil samples (A1 horizons), about
60% of the initial mass remained after subsampling
for microbial biomass (and inorganic N). This ma-
terial was air-dried and homogenised. One-third was
stored for organic matter fractionation and the
remaining material was ground in a blender for 5
min. One-third of this ground material was ground
again using a ball mill (Retsch MM2, 10 ml cups,
10 min) and then a third of this material was
ground again to a ®nal ®ne powder using a liquid
N freeze mill (Spex 6700 freezer/mill, 5 min). For
the labelled Oh horizons (mor and xeromoder) the
blender was replaced by a ultracentrifuge mill
(Retsch ZM1).
Total C, and 13C isotopic ratios were measured at
the Service Central d'Analyses of the CNRS, Solaize
(France), using a CN elemental analyser (CNRS)
coupled with a mass spectrometer (Finnigan delta S or
MAT 252).
One analysis was generally carried out for each
®eld replicate, but 10±25 replicates were initially
analysed (time `t0') according to humus types. This
was necessary for two reasons. Firstly, it was di�cult
to obtain homogeneous soil samples because the unde-
composed labelled wheat material was initially resist-
ant to grinding, and secondly, it was necessary to
obtain very accurate initial values for isotope ratios,
since the 13C remaining in soils during the ®eld ex-
posure period was calculated in percent of the initial
material.
The natural 13C isotopic ratio, measured in four
replicates at each site before the installation of the
experiment, varied from 1.081 to 1.083%. The 13C
derived from added plant material was calculated,
using the 13C isotopic enrichments: enrichment
� atom% excess � �measured isotopic ratio ÿnatural isotopic ratio� � 100: In highly labelled exper-
iments this formulation was preferable to d13C -(Boutton, 1991). In Table 2, d13C - varied from
+262 to +56 for the initial soil 13C and from +67 to
+5 for the ®nal soil 13C. In the ®gures the bares rep-
resent the standard deviation for the four replicates.
The comparison of data was performed using the test
of Student ( p value).
3. Results
3.1. Isotopic 13C
The characteristics of the labelled material were:C � 40:220:2%; 13C isotopic ratio � 10:54020:013%;N � 1:2820:02% �15N isotopic ratio � 9:73520:018%�and C-to-N ratio � 31:4: The added N in the labelledplant material, as % of soil native total N, rangedbetween 0.8 and 3.4%. The added C in plant material,expressed as % of soil native organic C, ranged gener-ally from 1.5 to 3.4%. The initial isotopic ratiosmeasured at t0 for the experiment ranged from 1.173to 1.399%. The ®nal isotopic ratios measured on thelast sampling occasion after 2±3 yr ®eld exposure ran-ged from 1.116 to 1.184% (Table 2). These ratios weresigni®cantly di�erent from 1.081±1.083% determinedfor the unlabelled soils sampled before the experiment(P < 0.01). Thus, because of the low N and high 13C-labelling, the plant material could be added to the soilas a very small proportion of the total mass, that iswithout changing the chemical, physical and biologicalproperties of the soil native organic matter and the tra-cer was su�ciently concentrated to be detectedthroughout the experiment.
3.2. Transfers of labelled C
In all cases the 13C isotopic ratios in the layerslocated directly above the labelled horizons were closeto the natural ratios. Hence upwards transfers of thelabelled-C (for instance by fungi) were negligible. The13C measured in the horizon located directly below thelabelled one was also low, ranging from 2 to 7% ofthe initial 13C. The highest 13C leaching occurred inthe Vindeln (boreal) soil of 4±7% of the initial 13C.The lowest values were observed in the acid Mediterra-nean soils with generally less than 3% of the initiallabel. Data for 13C in these adjacent horizons are notpresented separately.
3.3. 13C mineralisation rates
Fig. 4(a and b) shows the percentage 13C remainingin soils (sum of 13C remaining in the initially labelledhorizon plus the 13C recovered in the horizons locateddirectly under and above the initially labelled layer).For technical reasons, the installation of the exper-iment in the donor site for the native soil and in thehost site for the translocated soil was generally notachieved exactly at the same date. The time scale istherefore shown as exposure days for both native andtransplanted soils, rather than from the date on instal-lation. Fig. 4(a and b) illustrates the dynamics of 13Cmineralisation assuming that (i) the leaching oflabelled carbon below the layers analysed was negli-
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 533

gible and (ii) 13C remaining in the pro®le can be bud-geted as the di�erence between initial 13C and 13CO2
lost by respiration. The ®rst assumption is probablyvalid, since the amount of 13C translocated down thepro®le was low. The second assumption presupposesthat there were no 13C-bicarbonates and carbonates ac-cumulating in the soil from the 13CO2 released by thesoil respiration. This will not occur in acid and neutralsoil but could occur in soils containing CaCO3. TheLa Clape soil contained less than 1% carbonates butwas Ca2+ saturated �pH � 7:3). The 13C measured inthe A12 horizon (directly located below the labelledhorizon; Fig. 2) never exceeded 4%, indicating that13C immobilisation was probably negligible. In con-trast the Maials soil contained 40±42% carbonates inall horizons and showed unrealistic high 13C values(15±20%) in the below horizon. In this case 13CO2
may have been immobilised. The di�erentiation of car-bonate-13C from organic-13C is necessary to clarify theCO2-
13C sequestration. The results for this soil aretherefore not presented in this paper.
3.4. Soils from the boreal zone
The relocation of soil from Vindeln to JaÈ draaÊ s, i.e.from a north to a more south Scandinavian climate(Figs. 1 and 3), was accompanied by (i) an increase inmean annual air temperatures of 2.58C (averaged over3 yr), (ii) an increase of 17% of the sum of mean dailytemperatures greater than 08C and (iii) a mean annualprecipitation increase of 23% (Table 3). At both sites,the water de®cit (PET-AET) was low and similar tothe long-term values, except for the second year atJaÈ draaÊ s.
At both Vindeln and JaÈ draaÊ s, the experiment wasinstalled in June 1993. In the native Vindeln soil, thetotal soil C remained relatively constant (Fig. 4a).When transplanted to JaÈ draaÊ s, the total C decreasedprogressively by 20% and 13C remaining in the trans-located soil was 10% lower than the native soil (P <0.05). During the ®rst year of incubation, the rapid in-itial decomposition phase was interrupted during the6±7 winter months by temperatures falling below 08C.Consequently both the native and transplanted soilsshowed a stepwise pattern of 13C depletion. This e�ectwas not evident during the second (1994±1995) andthird (1995±1996) winters.
The translocation of the JaÈ draaÊ s soil from JaÈ draaÊ s(boreal) to Haldon (Atlantic climate) correspondedover the 3 yr to a mean annual air temperatureincrease of 78C. Precipitation increased by a factor oftwo over this period. The summer water de®cit waslow at both sites and of the same order of magnitude(<50 mm; Table 3) and was probably not a limitingfactor for C mineralisation. The JaÈ draaÊ s native soilwas installed in June and the transplanted soil wasT
able
213C-labellingcharacteristics
VindelnOh
JaÈdraaÊsOh
HaldonA1
FristonA1
The zanOh
The zanA1
LaClapeOh
LaClapeA1
DesertA1
MaialsOh
MaialsA1
Labelledplantmaterial
Added
plantmaterialgcylinderÿ1
1.31
1.31
0.94
1.19
1.09
0.82
1.12
0.90
0.92
1.13
1.10
Added
C%
nativesoilC
cylinderÿ1
2.17
2.50
1.78
1.94
1.84
1.47
1.97
3.39
1.84
2.98
1.08
Added
N%
nativesoilN
cylinderÿ1
2.19
3.39
0.81
0.86
2.26
1.87
1.48
1.77
1.96
2.23
1.10
Initialsoil
13C
(attime0)
Measuredisotopic
ratio(%
)1.218
1.275
1.194
1.253
1.200
1.191
1.206
1.399
1.230
1.280
1.173
S.D
.0.024
0.022
0.012
0.009
0.024
0.018
0.013
0.03
0.022
0.014
0.030
Replicates(n)
29
22
35
19
19
32
20
26
19
10
13
Measurednaturalisotopic
ratio�n�
4)
1.081
1.081
1.082
1.082
1.083
1.082
1.082
1.082
1.082
1.083
1.082
Finalsoil
13C
(measuredatthelast
samplingoccasion)
13C
isotopic
ratio
1.128
1.160
1.116
1.139
1.128
1.128
1.142
1.185
1.136
1.184
1.127
S.D
.�n�
4)
0.0054
0.0019
0.0013
0.0039
0.0075
0.0079
0.0056
0.0081
0.0036
0.0051
0.0017
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543534
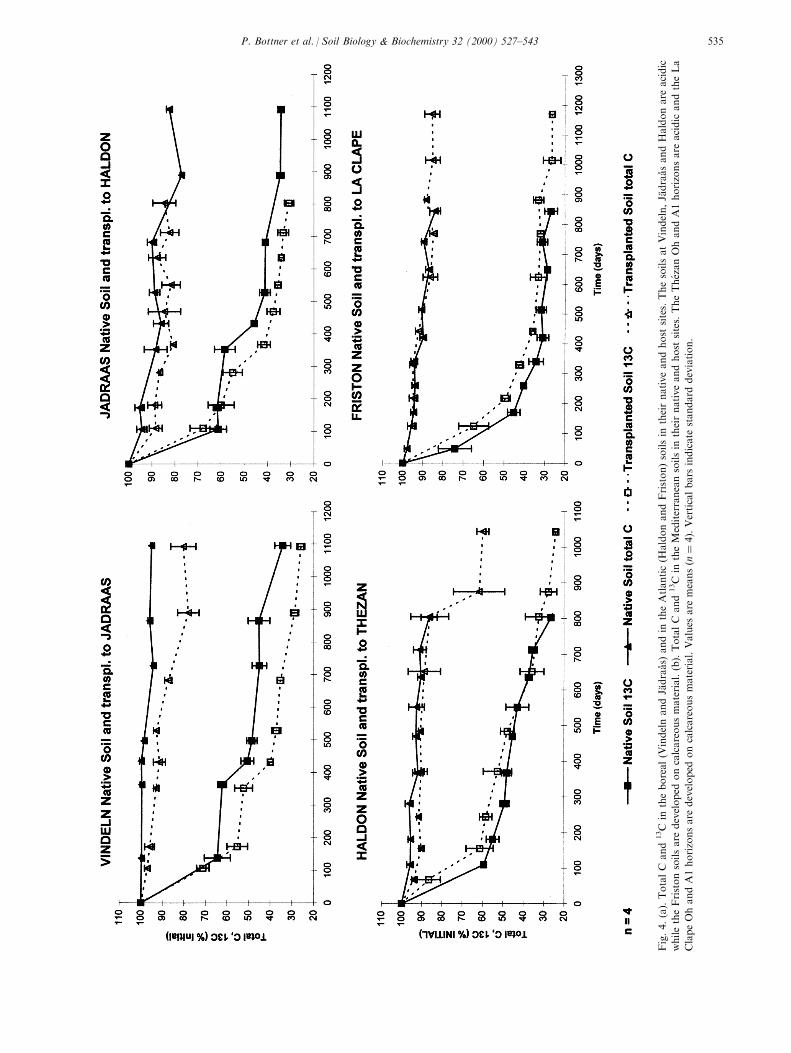
Fig.4.(a).TotalC
and
13C
intheboreal(V
indelnandJaÈdraaÊs)
andin
theAtlantic(H
aldonandFriston)soilsin
theirnativeandhost
sites.ThesoilsatVindeln,JaÈdraaÊsandHaldonare
acidic
whiletheFristonsoilsare
developed
oncalcareousmaterial.(b).TotalC
and
13C
intheMediterraneansoilsin
theirnativeandhost
sites.TheThe zanOhandA1horizonsare
acidic
andtheLa
ClapeOhandA1horizonsare
developed
oncalcareousmaterial.Values
are
means�n�
4).Verticalbars
indicate
standard
deviation.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 535
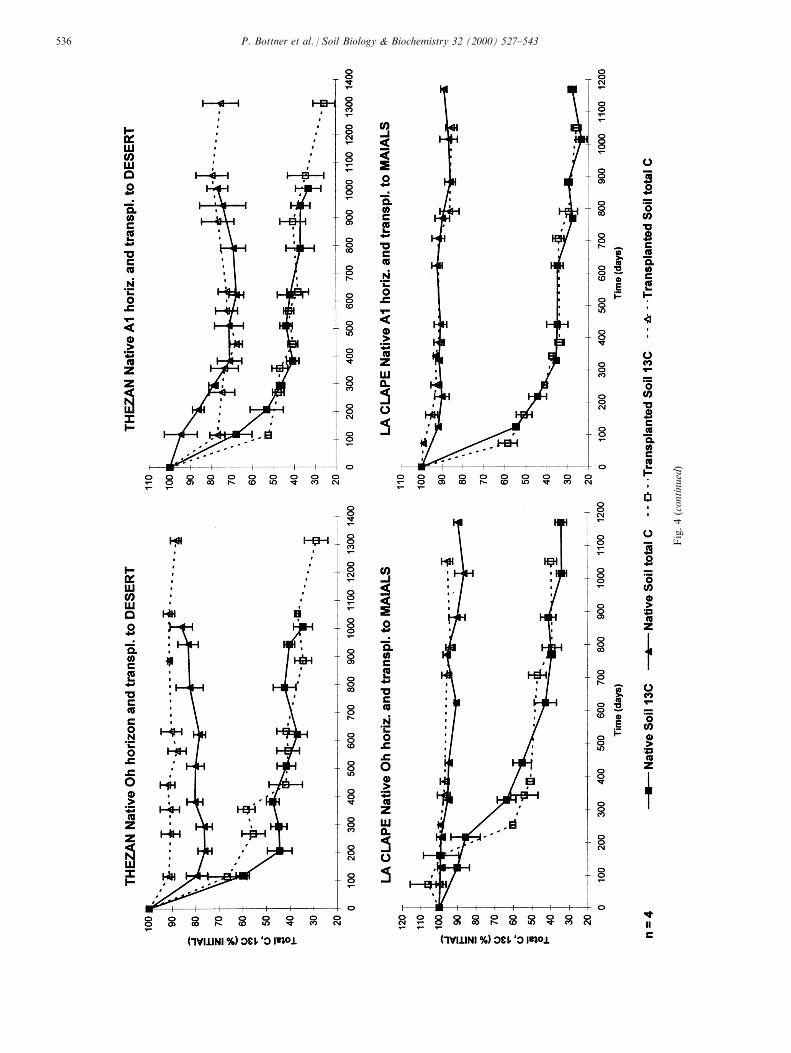
Fig.4(continued)
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543536
installed at Haldon only 3 months later. The stepwisetemperature-linked 13C-curve was also clearly observedfor the native soil during the ®rst winter period, butdisappeared for this soil when transplanted underAtlantic conditions. Both sites showed a similardecrease in total C of about 15%. The translocationincreased the 13C mineralisation by 5±10% (P < 0.05;Fig. 4a).
3.5. Soils from the Atlantic zone
In the acid soil sequence (Figs. 1 and 3), the Haldonsoil (Atlantic climate) was transplanted to The zan (wetMediterranean). The soil was thus exposed to anincrease in annual mean air temperature of 5.48C anda decrease in annual precipitation from 1505 to 731mm throughout the experiment. The water de®cit alsoincreased from 47 mm (with PET-AET > 0 over 3months) at Haldon to 313 mm with 7 months waterde®cit at The zan during spring, summer and autumn(Table 3). The native soil was installed in September1993 and the transplanted soil was installed 2 monthslater. The total C decreased by 15% of the initial totalC for both soils over 3 yr (Fig. 4a). Translocation ofHaldon soil to The zan resulted in a slower rate of 13Cdecrease in the transplanted soil. During the initial andrapid decomposition phase, the amount of 13C in thetranslocated soil was 5±10% higher than in the nativesoil (P<0.05 at samplings 2 and 3). During the slowerdecomposition phase, the mineralisation rates becamecomparable.
When the calcareous Friston soil (Atlantic climate)was transplanted to La Clape (wet Mediterranean)(Figs. 1 and 3) the soil was exposed to an increase inmean annual air temperature of 3.58C and a decreasein precipitation from 750 to 572 mm (Table 3). Thesevalues are comparable to long-term records but bothsites experienced a dry summer during the second yearof the experiment. The water de®cit increased from111 to 281 mm and the dry months increased from 5to 8.5 at La Clape. The third winter was at La Clapeexceptionally wet with 1199 mm precipitation. Never-theless, the moisture e�ect of the second and thirdyear on the 13C mineralisation rates was of little sig-ni®cance since, as shown in Fig. 4a, the pattern of 13Closses was already stabilised after the ®rst year. Thenative Friston soil was installed in March 1994 but thetransplanted soil had already been installed at LaClape since January 1994. The total organic C contentof the Friston soil decreased by 10±15% for bothnative and transplanted soils. The 13C curve of thetranslocated soil was generally higher by 5±10% com-pared to the native soil (0.1> P>0.05).
Hence, moisture de®cit limited 13C mineralisation inthe translocated Haldon and Friston soils. In contrastto the soils maintained under boreal conditions, forT
able
3
Climaticcharacteristics
ofthesites
Sites
Beginningof
experim
ent
Endof
experim
ent
Meanannuala
airtemperature
(8C)
Annualaprecipitation
(mm/year)
Sum
ofdaily
airtemperature
>08(8C)
Waterde®citb
PET-A
ET
(mm)
Number
ofmonths
withPET-A
ET
>0mm
year1
year2
year3
long
term
year1
year2
year3
long
term
year
year2
year3
year1
year2
year3
long
term
year1
year2
year3
long
term
Vindeln
16-06-93
15-06-96
ÿ0.6
2.0
0.3
0.2
707
563
341
604
1458
1712
1693
397
19
14
3
JaÈdraaÊs
19-06-93
15-06-96
2.3
4.4
2.6
3.9
620
804
560
764
1739
2067
2062
14
24
30
33
4
Haldon
22-09-93
04-12-95
9.8
11.0
9.6
10.1
1723
1475
1317
1354
3566
4020
3539
70
24
15
32
3
Friston
04-03-94
25-06-96
11.9
11.9
11.1
1073
428
801
4328
3994
38
184
59
37
5
Thezan
07-07-93
02-04-96
15.0
15.8
15.6
14.4
682
598
912
579
5486
5767
5704
183
392
364
154
510
74
LaClape
12-01-94
27-03-97
15.5
15.4
14.5
14.8
692
453
1199
587
5657
5605
5300
197
366
192
710
6
Desert
30-07-93
06-03-97
16.4
16.6
16.5
14.9
234
401
526
538
5982
6043
6029
557
464
337
232
10
11
11
5
Maials
18-04-94
05-03-97
16.2
16
15.7
15.2
356
383
405
388
5928
5843
5741
459
482
414
810
8
aTheannualdata
are
calculatedfrom
thedatesofthebeginningoftheexperim
ent.
bCalculatedusingtheWATBUG
model.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 537

both native Atlantic climate soils (Haldon and Friston)and the boreal JaÈ draaÊ s soil transplanted to Haldon,the generally favourable seasonal patterns of moistureand temperature resulted in steady and regular rates of13C mineralisation. The variability was particularly lowfor the native Haldon and Friston soils.
3.6. Soils in the Mediterranean zone
In the acid soils series, the xeromoder (Oh) and theacid mull (A1) of The zan were transplanted from wetMediterranean conditions to Desert de les Palmes(Figs. 1 and 3) which had the driest climatic conditionsin the transect. This involved a (3 yr) mean annualprecipitation decrease from 731 to 387 mm, an increasein moisture de®cit (PET-AET) from 313 to 453 mmyrÿ1; the number of dry months (PET-AET > 0)increased from 7 to 10 and the temperature increasedfrom 15.5 to 16.58C (Table 3). Units at The zan wereinstalled in July 1993 and at Desert 3 weeks later. Thecurves for residual organic-C and 13C showed greatervariation compared with results from of the Atlanticsites (Fig. 4b). This was probably the consequence ofthe heterogeneous spatial distribution of the Mediter-ranean vegetation in the sites, creating greater vari-ation in microclimate.
The more arid environment of the transplanted soilsa�ected the pattern of total organic C and 13C lossesfrom the Oh layer (Fig. 4b). During the ®rst year,total organic C was reduced by about 20% for thenative soil and 10% for the transplanted soil and 13Cby 55 and 45%, respectively (P < 0.05 at samplings 2and 3). Over this ®rst year the Desert site was particu-larly dry with a water de®cit for 10 months comparedto 5 months at The zan. Over the remainder of the ex-periment, the curves for residual 13C were not signi®-cantly di�erent (P > 0.1) for the two soil andstabilised at relatively high values. In contrast to theboreal soils where freezing markedly a�ected the 13Cmineralisation curves, the dry summer e�ect of theMediterranean soils was probably masked by the highvariability of the data.
Results for the The zan A1 horizon showed highvariability for the same reasons considered for the Ohmaterial. The curves for total organic C and 13C weresimilar for the native and transplanted soil, except onthe ®rst sampling occasion (Fig. 4b).
When the calcareous La Clape soil was transplantedfrom a wet Mediterranean climate to dry Mediterra-nean conditions at Maials (Figs. 1 and 3), the soilschanged from a mean annual precipitation of 781 to381 mm yrÿ1. The temperature increased from 15.1 to16.08C. Water de®cit increased from 282 (8 drymonths) to 468 mm yrÿ1 (9 dry months). As with theThe zan soil, the Oh and A1 horizons were investigated(Fig. 2). Simulating the natural ®eld pro®le, the Oh
horizon was separated from the underlying A1 horizonby a stone layer (St layer in Fig. 2). The experimentwas installed at La Clape in January 1994 and atMaials in April.
During the ®rst spring and summer, conditions wereextremely dry with 7±8 dry months and at Maialsthere were no decreases in Oh 13C from April (installa-tion) to September. At La Clape in Oh only 10±15%of the initial 13C was mineralised during this period.At Maials maximum rates of 13C mineralisationoccurred during the ®rst wet autumn and winter(1994). During the summer of 1995, mineralisation of13C was again reduced. Thus Maials Oh showed astepwise pattern of 13C losses controlled by seasonalalternate dry and wet periods while at La Clape thedrought e�ects were attenuated and only manifestedduring the ®rst summer period.
The La Clape labelled A1 mull horizon was locatedin the mineral soil under the xeromoder horizon andthe stony layer (Fig. 2). This bu�ered the variability inmoisture conditions at both sites. The dry summerconditions did not a�ect the 13C mineralisation(Fig. 4b). This soil, with basic pH and bu�ered moist-ure conditions, showed high mineralisation rates result-ing in 65% 13C loss during the ®rst year. In the nativeand transplanted soils, the curves for total organic Cand 13C were similar despite the comparatively lowvariability of the data.
3.7. Temperature e�ect on 13C mineralisation andstabilisation
Fig. 5 shows the residual 13C in the native andtransplanted soils in relation to the mean annual airtemperatures of the donor and host sites. The slopesof the lines indicate the temperature e�ect on 13C min-eralisation. The increase in soil temperatures producedby translocation had the largest e�ect (highest negativeslope) on 13C mineralisation between Vindeln andJaÈ draaÊ s, i.e. from north to south boreal conditions.The 13C mineralisation rates were enhanced over the 3yr of decomposition as shown by signi®cant (P <0.001) di�erences in the slopes of V1, V2 and V3. Thetranslocation from boreal to Atlantic conditions alsoproduced an increase in 13C mineralisation, but theslopes are less steep and the temperature e�ectdecreased from the ®rst (J1; P < 0.01) to the last year(J3; P < 0.05). The temperature increase had no sig-ni®cant e�ects when the Haldon soil was translocatedto The zan and the positive slope for H1 indicates atendency �P � 0:13� for 13C mineralisation to decreaseduring the ®rst year. For the Friston soil transplantedto La Clape, the mineralisation of 13C was signi®cantlyreduced during the ®rst (P < 0.01 for F1) and secondyears (P<0.01 for F2). Translocation within the Med-iterranean region had no e�ect on 13C mineralisation
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543538
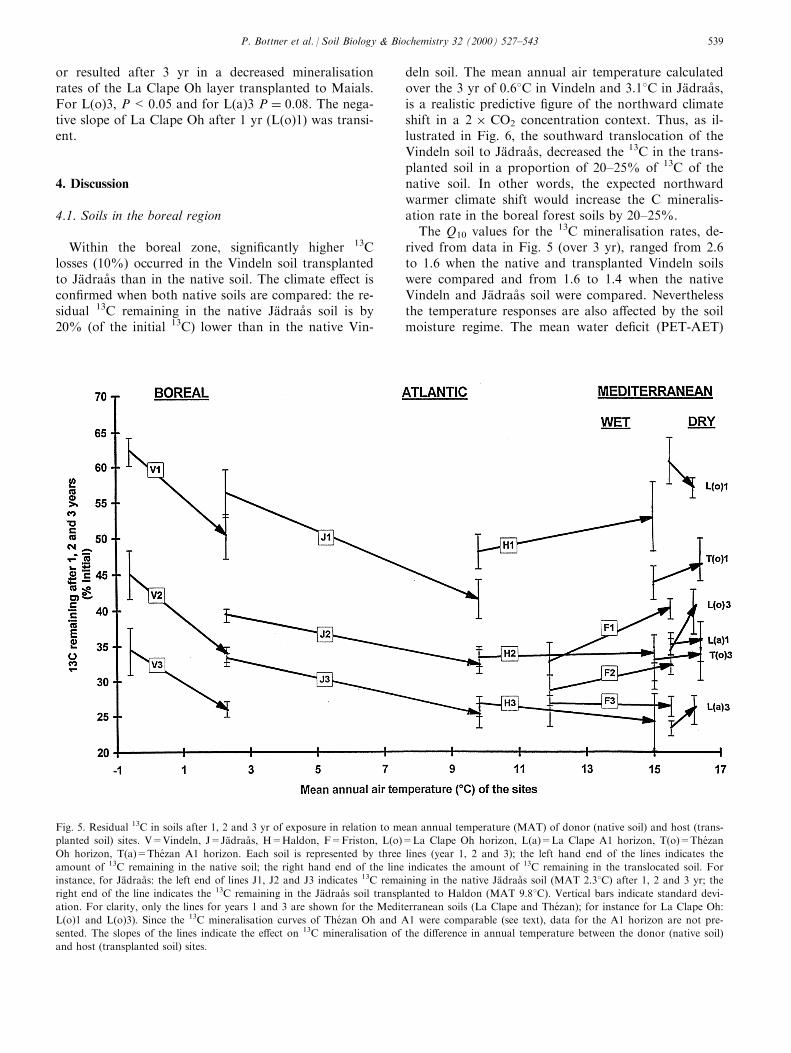
or resulted after 3 yr in a decreased mineralisationrates of the La Clape Oh layer transplanted to Maials.For L(o)3, P<0.05 and for L(a)3 P � 0:08: The nega-tive slope of La Clape Oh after 1 yr (L(o)1) was transi-ent.
4. Discussion
4.1. Soils in the boreal region
Within the boreal zone, signi®cantly higher 13Closses (10%) occurred in the Vindeln soil transplantedto JaÈ draaÊ s than in the native soil. The climate e�ect iscon®rmed when both native soils are compared: the re-sidual 13C remaining in the native JaÈ draaÊ s soil is by20% (of the initial 13C) lower than in the native Vin-
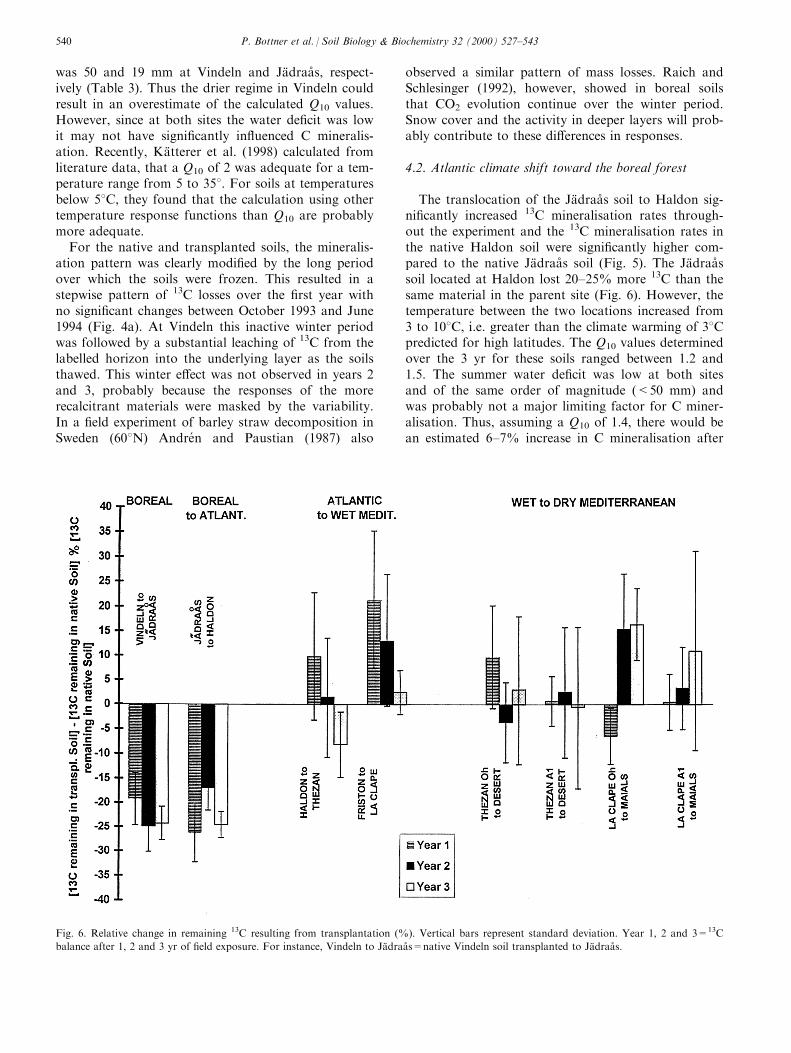
deln soil. The mean annual air temperature calculatedover the 3 yr of 0.68C in Vindeln and 3.18C in JaÈ draaÊ s,is a realistic predictive ®gure of the northward climateshift in a 2 � CO2 concentration context. Thus, as il-lustrated in Fig. 6, the southward translocation of theVindeln soil to JaÈ draaÊ s, decreased the 13C in the trans-planted soil in a proportion of 20±25% of 13C of thenative soil. In other words, the expected northwardwarmer climate shift would increase the C mineralis-ation rate in the boreal forest soils by 20±25%.
The Q10 values for the 13C mineralisation rates, de-rived from data in Fig. 5 (over 3 yr), ranged from 2.6to 1.6 when the native and transplanted Vindeln soilswere compared and from 1.6 to 1.4 when the nativeVindeln and JaÈ draaÊ s soil were compared. Neverthelessthe temperature responses are also a�ected by the soilmoisture regime. The mean water de®cit (PET-AET)
Fig. 5. Residual 13C in soils after 1, 2 and 3 yr of exposure in relation to mean annual temperature (MAT) of donor (native soil) and host (trans-
planted soil) sites. V=Vindeln, J=JaÈ draaÊ s, H=Haldon, F=Friston, L(o)=La Clape Oh horizon, L(a)=La Clape A1 horizon, T(o)=The zan
Oh horizon, T(a)=The zan A1 horizon. Each soil is represented by three lines (year 1, 2 and 3); the left hand end of the lines indicates the
amount of 13C remaining in the native soil; the right hand end of the line indicates the amount of 13C remaining in the translocated soil. For
instance, for JaÈ draaÊ s: the left end of lines J1, J2 and J3 indicates 13C remaining in the native JaÈ draaÊ s soil (MAT 2.38C) after 1, 2 and 3 yr; the
right end of the line indicates the 13C remaining in the JaÈ draaÊ s soil transplanted to Haldon (MAT 9.88C). Vertical bars indicate standard devi-
ation. For clarity, only the lines for years 1 and 3 are shown for the Mediterranean soils (La Clape and The zan); for instance for La Clape Oh:
L(o)1 and L(o)3). Since the 13C mineralisation curves of The zan Oh and A1 were comparable (see text), data for the A1 horizon are not pre-
sented. The slopes of the lines indicate the e�ect on 13C mineralisation of the di�erence in annual temperature between the donor (native soil)
and host (transplanted soil) sites.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 539
was 50 and 19 mm at Vindeln and JaÈ draaÊ s, respect-ively (Table 3). Thus the drier regime in Vindeln couldresult in an overestimate of the calculated Q10 values.However, since at both sites the water de®cit was lowit may not have signi®cantly in¯uenced C mineralis-ation. Recently, KaÈ tterer et al. (1998) calculated fromliterature data, that a Q10 of 2 was adequate for a tem-perature range from 5 to 358. For soils at temperaturesbelow 58C, they found that the calculation using othertemperature response functions than Q10 are probablymore adequate.
For the native and transplanted soils, the mineralis-ation pattern was clearly modi®ed by the long periodover which the soils were frozen. This resulted in astepwise pattern of 13C losses over the ®rst year withno signi®cant changes between October 1993 and June1994 (Fig. 4a). At Vindeln this inactive winter periodwas followed by a substantial leaching of 13C from thelabelled horizon into the underlying layer as the soilsthawed. This winter e�ect was not observed in years 2and 3, probably because the responses of the morerecalcitrant materials were masked by the variability.In a ®eld experiment of barley straw decomposition inSweden (608N) Andre n and Paustian (1987) also
observed a similar pattern of mass losses. Raich andSchlesinger (1992), however, showed in boreal soilsthat CO2 evolution continue over the winter period.Snow cover and the activity in deeper layers will prob-ably contribute to these di�erences in responses.
4.2. Atlantic climate shift toward the boreal forest
The translocation of the JaÈ draaÊ s soil to Haldon sig-ni®cantly increased 13C mineralisation rates through-out the experiment and the 13C mineralisation rates inthe native Haldon soil were signi®cantly higher com-pared to the native JaÈ draaÊ s soil (Fig. 5). The JaÈ draaÊ ssoil located at Haldon lost 20±25% more 13C than thesame material in the parent site (Fig. 6). However, thetemperature between the two locations increased from3 to 108C, i.e. greater than the climate warming of 38Cpredicted for high latitudes. The Q10 values determinedover the 3 yr for these soils ranged between 1.2 and1.5. The summer water de®cit was low at both sitesand of the same order of magnitude (<50 mm) andwas probably not a major limiting factor for C miner-alisation. Thus, assuming a Q10 of 1.4, there would bean estimated 6±7% increase in C mineralisation after
Fig. 6. Relative change in remaining 13C resulting from transplantation (%). Vertical bars represent standard deviation. Year 1, 2 and 3=13C
balance after 1, 2 and 3 yr of ®eld exposure. For instance, Vindeln to JaÈ draaÊ s=native Vindeln soil transplanted to JaÈ draaÊ s.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543540

the ®rst year of the JaÈ draaÊ s soil for a 38C rise in tem-perature and a increase of 4% after 3 y. Thus a 38Cincrease in the JaÈ draaÊ s soil had a substantially smallere�ect than the same temperature increase in the Vin-deln soil, indicating that there were di�erences in thetemperature sensitivity of the organic matter at thetwo latitudes.
4.3. Soils in the warmer temperate regions
The mean temperature increased from 9.8 at Haldonto 158C at The zan and from 11.9 at Friston to 168C atLa Clape but the 13C mineralisation was not a�ectedbecause the temperature e�ect was probably counter-acted by the increasing moisture de®cits down thetransect (Fig. 6). Water de®cit signi®cantly reduced13C mineralisation during the ®rst 2 yr when the Fris-ton soil was translocated to La Clape and also forHaldon during the ®rst year. Thus the expected climateshift of wet Mediterranean climate toward the Atlanticzone would lower the decomposition rates of the tran-sient labile compounds during the initial mineralisationphases. The slow 13C mineralisation phases were notsigni®cantly a�ected, either because the decompositionof the stabilised material was less sensitive to moistureregime (e.g. changes in the composition of the mi-crobial community, inducing functional shifts; Zogg etal., 1997) or the high variability masked the di�erencebetween the treatments. In addition Pinol et al. (1995)observed that ®eld CO2 e�ux from Mediterraneansoils are not entirely explained by moisture and tem-perature conditions.
In western temperate Europe, with a doubling of at-mospheric CO2, a global warming of 1.1±2.48C is pre-dicted with higher frequencies of seasonal and weatherextremes. Summer precipitation is predicted todecrease in southern UK and potential evapotranspira-tion to increase in both winter and summer (CCIRG,1996). Nevertheless, despite the lack of more precisepredictions for moisture regime, the climate conditionsof The zan and La Clape compared to Haldon andFriston probably overestimate the e�ects of predictedclimate shifts. Assuming roughly a linear relationbetween water de®cit and C mineralisation for therange of precipitation between the donor and hostsites, a decrease of about 6±12 % in C mineralisationper 100 mm decline in rainfall is a realistic estimate.More precise predictions would be questionable; themost ®rm result is that the temperature e�ect is clearlycounteracted by the moisture e�ect.
4.4. E�ects of increased temperature and aridity in thewet Mediterranean zone
The translocation of soils by 2±38 latitude, from thewet Mediterranean sites (The zan and La Clape in
southern France) to the dry Mediterranean Spanishhost sites (Desert and Maials), resulted in a mean airtemperature increase over the 3 yr of 1.0 and 0.88C forThe zan to Desert and La Clape to Maials, respectively.The corresponding decrease in precipitation was 344and 400 mm yrÿ1 and the increased water de®cits were140 and 187 mm, respectively. The general circulationmodels predict a temperature increase of 1.3±1.98C inthe Mediterranean with a doubling of atmosphericCO2 with longer periods of consecutive hot periodsduring the dry seasons (Rambal and Ho�, 1998). Anessential characteristic of the Mediterranean soils is thedeep distribution of the soil organic matter within thesoil pro®le as a consequence of both deep rooting sys-tems and soil faunal animal activity: a strategy ofplants and animals to survive during the dry season.Thus the soil organic matter quality (humus types) isstrongly in¯uenced by the physical and chemicalcharacteristics of the mineral soil.
With the exception of the La Clape Oh horizonduring the ®rst year (Lo1 in Fig. 5) the host environ-ment generally reduced 13C mineralisation rates in Ohand A1 as a consequence of moisture limitation over-riding temperature e�ects. However, the di�erencebetween native and transplanted soil was really im-portant and signi®cant only for the La Clape Oh hor-izon (L(o)3, Fig. 5). The increasing moisture limitationsubstantially a�ected only the Oh layers (Fig. 6).
At La Clape, the Oh horizon was separated fromthe underlying A1 horizon by a stone layer of about 5cm thickness. In Mediterranean soils, this frequentlyobserved layer results from the residual accumulationof 2±7 cm dia stones after erosion of soil ®nes, whenthe primary Mediterranean forest was destroyed. Afterthe more recent natural forest reinstalled, the xeromo-der (Oh) developed on the surface of the stone layerand was consequently subject to severe desiccation and¯uctuating moisture regimes. Similar conditions lead-ing to the development of xeromoders are quite preva-lent in Mediterranean forests, especially on calcareoussoils. The columns set up with the La Clape soilsincluded this stone layer (Fig. 2) and probably contrib-uted to the larger residual 13C in the Oh horizon thanin the A1 horizons for both the native and translo-cated systems.
While the Oh material at La Clape has developedon a soil recently renewed by erosion and recolonisedby forest, the The zan Oh is developed on a old stabil-ised `luvisol', representative of large areas in the Medi-terranean region. Here the clays and ®ne particlefractions have been weathered out of the topsoil intothe B horizon during long pedogenesis periods, so thatxeromoder horizon has developed on the surface of animpoverished, very permeable, residual, sandy and dryhorizon (Bottner et al., 1995). Although the humustype (Oh) is similar, the pH is 5.2 for La Clape on cal-
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 541

careous material and 4.3 for The zan on acid sand ma-terial. The 13C mineralisation rates in Oh were similarfor the two soils, emphasising the dominant e�ects ofmoisture on decomposition processes in the top layersin this region.
The 13C mineralisation characteristics of the The zanA1 horizon (not shown in Fig. 5) were similar to thoseof the The zan Oh horizon. The The zan A1 horizonwas also developed in the leached and extremely per-meable top mineral layer below the xeromoder (Fig. 2);so soil processes in this horizon were probably alsomoisture-limited for a signi®cant period of the year.
The La Clape A1 horizon is of special interest as itshowed the highest mineralisation rates with morethan 75% losses of 13C from both the native andtranslocated soils (Fig. 4b). The most likely reasonsfor this are that (i) the water draining rapidly throughthe Oh and stone layers is retained within this clayand loam horizon and moisture losses by evaporationare reduced by the mulching e�ects of the overlyingmaterials and (ii) in this Ca2+. saturated soil the ac-tivity is stimulated by soil neutral pH (7.3). In this A1horizon, the processes are not moisture-limited forextended periods, when desiccation limits decompo-sition of surface organic matter (Oh). The la Clape A1horizon illustrates the high activity of the Mediterra-nean soil deep organic layers, where favourable tem-perature conditions meet bu�ered moisture conditionslasting beyond the rain season.
4.5. The climate shift from Mediterranean to Atlantic: akey situation for transition of soils as carbon source orsink
The concave shape of the curves shown in Fig. 5 in-dicates a number of key features of the e�ects of cli-mate change on these soils. (1) The magnitude of thetemperature e�ect decreased progressively from thenorthern boreal climate at Vindeln to the Atlantic cli-mate at Haldon. Then from Atlantic to Mediterraneanregions the moisture limitation counteracts the e�ectsof higher temperatures. (2) In the more temperature-limited situations (Vindeln to JaÈ draaÊ s), the temperaturee�ect persists over the 3 yr. When the temperaturee�ect becomes less dominant (JaÈ draaÊ s to Haldon) onlythe ®rst year decomposition is a�ected (the active de-composition phase). (3) Similarly, the water de®cita�ects generally only the ®rst year decomposition i.e.H1 and F1 in Fig. 5. The lowering of 13C mineralis-ation persists over the 3 yr only under the driest con-ditions, i.e. La Clape Oh, (L(o)3 in Fig. 5). (4) Theconcave part of the centre of the Figure (the Haldonsite with Haldon native soil and the soil transplantedfrom JaÈ draaÊ s and the native Friston soil) illustratesthat optimum combined temperature and moisture
conditions occur under the current Atlantic climate,and results in the highest 13C mineralisation rates.
The transition between the current Atlantic climateand wet Mediterranean climate (in the more generalHoldridge life zone chart: the transition between cooltemperate and warm temperate) is probably a key situ-ation, with an expected net stimulation of the C miner-alisation controlled by the predicted temperatureincrease to the north and a net slowdown controlledby the predicted water de®cit increase to the south.This is illustrated by Fig. 5: The ordinate of thenorth±south soils sequences follows asymmetrical con-cave shaped curves (year 1: V14 J14H14 F1; year2: V2, 4 J2, 4 H2 4 F2; year 3: V3, 4 J3 4 H3 4F3) indicating from north to south a temperature con-trolled decrease of C mineralisation from Vindeln toHaldon and Friston. Southward, beyond the Atlanticclimate, despite the temperature increase, the C miner-alisation remains constant or tends to be slightly low-ered. The increased water de®cit from wet to dryMediterranean climate, lowered the C mineralisationclearly in particularly unfavourable conditions, illus-trated by the xeromoders. The high and prolonged soilactivity persists under Mediterranean conditions onlyunder particular favourable microclimatic circum-stances, which were illustrated in this paper by thedeep organic and moisture bu�ered layers of the Medi-terranean soils (La Clape A1, i.e. L(a)3 in Fig. 5). Theroot litter derived from of the deeply distributed rootsystem of the Mediterranean vegetation and the or-ganic matter incorporated by animals provides sub-strate for microbial activity.
5. Conclusion
The combination of (i) tracer techniques, (ii) latitu-dinal climatic gradient investigation and (iii) transloca-tion of di�erent humus forms has provided a novelinsight into the potential e�ects of climate change ondecomposition processes and the dynamics of soil Cpools. In the West European 65±408 latitudes, themost favourable temperature and moisture conditionsoccur under the current Atlantic climate. Northwardthis climate zone (the cool temperate and boreal cli-mate), where the soil processes are essentially tempera-ture-limited, the predicted temperature increase of 38Cwill probably enhance C mineralisation. Based on Q10
values derived from the comparison of native andtransplanted soils, the calculation estimates an increaseof the C mineralisation of about 10% for the cool tem-perate zone and of about 20±25% for the boreal zone.This estimation is based on the assumption that inthese northern sites the di�erence in moisture e�ectbetween native and host site is of small importancecompared to the temperature e�ect. Southward the
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543542

Atlantic climate zone, where soil processes are season-ally moisture-limited, increased temperatures are likelyto have little e�ects on soil organic matter dynamics,because of the higher water de®cit. A signi®cant re-duction of C mineralisation rates was shown only inthe surface organic layers, morphologically recognisedin Mediterranean forest soils as `xeromoder' and sub-ject to frequent dry conditions. The amount of Cstored in this thin and patch-wise distributed organicsurface horizon is unknown. In the deeper organiclayers (the mull humus types) which constitute themain soil C pools in Mediterranean soils, microbial ac-tivity is maintained for longer into the dry seasons asa consequence of moisture storage. However a tem-perature increase of 28C did not signi®cantly a�ect Cmineralisation in this layer because the e�ects ofhigher temperatures may have been counteracted byincreased water de®cits.
Acknowledgements
This work was supported by the EnvironmentalResearch and Development Programme of the Com-mission of the European Community (Vamos pro-gramme). We are grateful to V. Couà teaux, P. Jame, F.Picasso and P. Splatt for technical assistance with theproject.
References
Andre n, O., Paustian, K., 1987. Barley straw decomposition in the
®eld: a comparison of models. Ecology 68, 1190±1200.
Bottner, P., Couà teaux, M.M., Vallejo, V.R., 1995. Soil organic mat-
ter in Mediterranean-type ecosystems and global climatic changes:
a case study: the soils of the Mediterranean Basin. In: Moreno,
J.M., Oechel, W.C. (Eds.), Global Changes and Mediterranean-
Type Ecosystems. Springer, New York, pp. 306±325.
Boutton, T.W., 1991. Stable carbon isotopic ratios of natural ma-
terial: Sample preparation and mass spectrometic analysis. In:
Coleman, D.C., Fry, B. (Eds.), Carbon Isotopic Techniques.
Academic Press, New York, pp. 155±171.
CCIRG, 1996. Review of the potential e�ects of climate change in
the United Kindom. Department of Environment, HMSO,
London.
Ciais, P., Tans, P.P., Trollier, M., White, J.W.C., Francey, R.J.,
1995. A large northern hemisphere terrestrial CO2 sink indicated
by the 13C/12C ratio of atmospheric CO2. Science 269, 1098±1102.
Cotrufo, M.F., Ineson, P., Rowland, A.P., 1994. Decomposition of
tree leaf litters grown under elevated CO2: e�ects of litter quality.
Plant and Soil 163, 121±130.
Couà teaux, M.M., Mousseau, M., Ce le rier, M.L., Bottner, P., 1991.
Increased atmospheric CO2 and litter quality: decomposition of
sweet chestnut leaf litter with animal food webs of di�erent com-
plexities. Oikos 61, 54±64.
Dai, A., Fung, I.Y., 1993. Can climate variability contribute to the
``missing'' CO2 sink? Global Biogeochemical Cycles 7, 599±609.
Jenkinson, D.S., Adams, D.E., Wild, A., 1991. Model estimates of
CO2 emissions from soil in response to global warming. Nature
351, 304±306.
KaÈ tterer, T., Reichstein, M., Andre n, O., Lomander, A., 1998.
Temperature dependance of organic matter decomposition: a
critical review using literature data analysed with di�erent
models. Biology and Fertility of Soils 27, 258±262.
Keeling, R.F., Piper, S.C., Heimann, M., 1996. Global and hemi-
spheric CO2 sinks deduced from changes in atmospheric O2 con-
centration. Nature 381, 218±221.
Pinol, J., Alcaniz, J.M., Roda, F., 1995. Carbon dioxide e�ux and
pCO2 in soils of three Quercus ilex montane forests.
Biogeochemistry 30, 191±215.
Raich, J.W., Potter, C.S., 1995. Global patterns of carbon dioxide
emissions from soils. Global Biogeochemical Cycles 9, 23±36.
Raich, J.W., Schlesinger, W.H., 1992. The global carbon dioxide ¯ux
in soil respiration and its relationship to vegetation and climate.
Tellus 44B, 81±99.
Rambal, S., Ho�, C., 1998. Mediterranean ecosystems and ®re: the
threats of global change. In: Moreno, J.M. (Ed.), Large Forest
Fires. Backhuys, Leiden, pp. 187±213.
Schimel, D.S., Braswell, B.H., Holland, E.A., Mckeown, R., Ojima,
D.S., Painter, T.H., Parton, W.J., Townsend, A.R., 1994.
Climatic, edaphic and biotic controls over storage and turnover
of carbon in soils. Global Biogeochemical Cycles 8, 279±293.
Tans, P.P., Fung, I.Y., Takahashi, T., 1990. Observational con-
straints on the global atmospheric CO2 budget. Science 247,
1431±1438.
Townsend, A.R., Vitousek, P.M., Holland, E.A., 1992. Tropical soils
could dominate the short-term carbon cycle feedback to increased
global temperature. Climatic Change 22, 293±303.
Zogg, G.P., Zak, D.R., Ringelberg, D.B., MacDonald, N.W., 1997.
Compositional and functional shifts in microbial communities
due to soil warming. Soil Science Society of America Journal 61,
475±481.
P. Bottner et al. / Soil Biology & Biochemistry 32 (2000) 527±543 543




















