
Decoding a Perceptual Decision Process across Cortex
15
Neuron Article Decoding a Perceptual Decision Process across Cortex Adria ´ n Herna ´ ndez, 1 Vero ´ nica Na ´ cher, 1 Rogelio Luna, 1 Antonio Zainos, 1 Luis Lemus, 1 Manuel Alvarez, 1 Yuriria Va ´ zquez, 1 Liliana Camarillo, 1 and Ranulfo Romo 1, * 1 Instituto de Fisiologı ´a Celular-Neurociencias, Universidad Nacional Auto ´ noma de Me ´ xico, 04510 Me ´ xico, D.F., Mexico *Correspondence: [email protected] DOI 10.1016/j.neuron.2010.03.031 SUMMARY Perceptual decisions arise from the activity of neurons distributed across brain circuits. But, decod- ing the mechanisms behind this cognitive operation across brain circuits has long posed a difficult problem. We recorded the neuronal activity of diverse cortical areas, while monkeys performed a vibrotac- tile discrimination task. We find that the encoding of the stimuli during the stimulus periods, working memory, and comparison periods is widely distrib- uted across cortical areas. Notably, during the comparison and postponed decision report periods the activity of frontal brain circuits encode both the result of the sensory evaluation that corresponds to the monkey’s possible choices and past information on which the decision is based. These results sug- gest that frontal lobe circuits are more engaged in the readout of sensory information from working memory, when it is required to be compared with other sensory inputs, than simply engaged in motor responses during this task. INTRODUCTION In its simplest formulation, a perceptual decision results from the interaction between past and current sensory information. A major problem in this formulation involves understanding how brain circuits represent past and current sensory events and how these representations are linked to perceptual reports (Romo and Salinas, 1999). Previously, we addressed this problem using a vibrotactile discrimination task (Herna ´ n- dez et al., 1997). In this task, trained monkeys compare infor- mation of the first stimulus frequency (f1) temporarily stored in working memory to the current sensory information of the second stimulus frequency (f2) to form a decision, i.e., whether f2 > f1 or f2 < f1, and to immediately report their perceptual evaluation by pressing one of two push buttons. Because this sequence depends on discrimination of highly simplified stimuli, the neuronal activity of diverse cortical areas can be examined during the same behavior (Brody et al., 2003; Chow et al., 2009; Herna ´ ndez et al., 2000, 2002; Jun et al., 2010; Luna et al., 2005; Machens et al., 2005; Romo et al., 1999, 2002, 2003, 2004; Romo and Salinas, 2003; Salinas et al., 2000). The task used in these studies simulates the behavioral condition in which the decision based on a sensory evaluation is immediately reported through a voluntary movement (Herna ´ n- dez et al., 1997). There are, however, behavioral conditions in which a perceptual decision can be postponed for later report. But, in theory, once the subject reaches a decision, this becomes categorical, no matter whether it must be reported immediately or reported later. If postponed, memory circuits may store the categorical decision for later report (de Lafuente and Romo, 2005; Shadlen and Newsome, 1996). However, an alternative could be that the memory circuits store not only the categorical decision, but also the information on which the decision is based (Lemus et al., 2007). This last possibility could be extremely advantageous since it gives flexibility for the deci- sion-making process. In this case, it is possible that the deci- sion is revised or updated as long as there is time for it to be reconsidered. In a variant of the vibrotactile discrimination task, in which monkeys were asked to postpone their decision report, we found that the activity of medial premotor cortex (MPC, presupplemen- tary motor area, and supplementary motor cortex) neurons during this period encodes both the result of the sensory evalu- ation (which corresponds to the monkey’s two possible choices) and past information on which the decision is based (Lemus et al., 2007). These responses could switch back and forth with remarkable flexibility across the postponed decision report period. Moreover, these responses covaried with the animal’s decision report. Thus, the MPC circuits appear critically suited to integrate and reorganize all of the elements associated with decision making in this task. Furthermore, they reflect the flexi- bility that is needed when a perceptual decision must be either immediately reported (Herna ´ ndez et al., 2002) or postponed for later report (Lemus et al., 2007). This result prompted us to further explore whether the neuronal responses recorded during the postponed decision period are a unique property of the MPC circuit (Lemus et al., 2007) or whether similar processes are also present in other cortical areas of the parietal and frontal lobes during this variant of the task. To further investigate this question, we recorded the neuronal activities of diverse cortical areas while trained monkeys reported a postponed decision based on previous sensory evaluation. In this task, monkeys must hold f1 in working memory and must compare it to the current sensory stimulus (f2) and must postpone the decision report until a cue triggers the 300 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Decoding a Perceptual Decision Process across Cortex

Neuron
Article
Decoding a Perceptual DecisionProcess across CortexAdrian Hernandez,1 Veronica Nacher,1 Rogelio Luna,1 Antonio Zainos,1 Luis Lemus,1 Manuel Alvarez,1 Yuriria Vazquez,1
Liliana Camarillo,1 and Ranulfo Romo1,*1Instituto de Fisiologıa Celular-Neurociencias, Universidad Nacional Autonoma de Mexico, 04510 Mexico, D.F., Mexico
*Correspondence: [email protected] 10.1016/j.neuron.2010.03.031
SUMMARY
Perceptual decisions arise from the activity ofneurons distributed across brain circuits. But, decod-ing the mechanisms behind this cognitive operationacross brain circuits has long posed a difficultproblem. We recorded the neuronal activity of diversecortical areas, while monkeys performed a vibrotac-tile discrimination task. We find that the encoding ofthe stimuli during the stimulus periods, workingmemory, and comparison periods is widely distrib-uted across cortical areas. Notably, during thecomparison and postponed decision report periodsthe activity of frontal brain circuits encode both theresult of the sensory evaluation that corresponds tothe monkey’s possible choices and past informationon which the decision is based. These results sug-gest that frontal lobe circuits are more engaged inthe readout of sensory information from workingmemory, when it is required to be compared withother sensory inputs, than simply engaged in motorresponses during this task.
INTRODUCTION
In its simplest formulation, a perceptual decision results from
the interaction between past and current sensory information.
A major problem in this formulation involves understanding
how brain circuits represent past and current sensory events
and how these representations are linked to perceptual
reports (Romo and Salinas, 1999). Previously, we addressed
this problem using a vibrotactile discrimination task (Hernan-
dez et al., 1997). In this task, trained monkeys compare infor-
mation of the first stimulus frequency (f1) temporarily stored in
working memory to the current sensory information of the
second stimulus frequency (f2) to form a decision, i.e., whether
f2 > f1 or f2 < f1, and to immediately report their perceptual
evaluation by pressing one of two push buttons. Because
this sequence depends on discrimination of highly simplified
stimuli, the neuronal activity of diverse cortical areas can be
examined during the same behavior (Brody et al., 2003; Chow
et al., 2009; Hernandez et al., 2000, 2002; Jun et al., 2010;
Luna et al., 2005; Machens et al., 2005; Romo et al., 1999,
300 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
2002, 2003, 2004; Romo and Salinas, 2003; Salinas et al.,
2000).
The task used in these studies simulates the behavioral
condition in which the decision based on a sensory evaluation
is immediately reported through a voluntary movement (Hernan-
dez et al., 1997). There are, however, behavioral conditions in
which a perceptual decision can be postponed for later report.
But, in theory, once the subject reaches a decision, this becomes
categorical, no matter whether it must be reported immediately
or reported later. If postponed, memory circuits may store the
categorical decision for later report (de Lafuente and Romo,
2005; Shadlen and Newsome, 1996). However, an alternative
could be that the memory circuits store not only the categorical
decision, but also the information on which the decision is
based (Lemus et al., 2007). This last possibility could be
extremely advantageous since it gives flexibility for the deci-
sion-making process. In this case, it is possible that the deci-
sion is revised or updated as long as there is time for it to be
reconsidered.
In a variant of the vibrotactile discrimination task, in which
monkeys were asked to postpone their decision report, we found
that the activity of medial premotor cortex (MPC, presupplemen-
tary motor area, and supplementary motor cortex) neurons
during this period encodes both the result of the sensory evalu-
ation (which corresponds to the monkey’s two possible choices)
and past information on which the decision is based (Lemus
et al., 2007). These responses could switch back and forth with
remarkable flexibility across the postponed decision report
period. Moreover, these responses covaried with the animal’s
decision report. Thus, the MPC circuits appear critically suited
to integrate and reorganize all of the elements associated with
decision making in this task. Furthermore, they reflect the flexi-
bility that is needed when a perceptual decision must be either
immediately reported (Hernandez et al., 2002) or postponed for
later report (Lemus et al., 2007).
This result prompted us to further explore whether the
neuronal responses recorded during the postponed decision
period are a unique property of the MPC circuit (Lemus et al.,
2007) or whether similar processes are also present in other
cortical areas of the parietal and frontal lobes during this variant
of the task. To further investigate this question, we recorded the
neuronal activities of diverse cortical areas while trained
monkeys reported a postponed decision based on previous
sensory evaluation. In this task, monkeys must hold f1 in working
memory and must compare it to the current sensory stimulus (f2)
and must postpone the decision report until a cue triggers the

Figure 1. Discrimination Task
(A) Sequence of events during discrimination
trials. The mechanical probe is lowered, indenting
the glabrous skin of one digit of the restrained
hand (pd); the monkey places its free hand on an
immovable key (kd); the probe oscillates vertically,
at the base stimulus frequency (f1); after a fixed
delay (3 s), a second mechanical vibration is deliv-
ered at the comparison frequency (f2); after
another fixed delay (3 s) between the end of f2
and probe up (pu), the monkey releases the key
(ku) and presses either a lateral or a medial push-
button (pb) to indicate whether the comparison
frequency was higher or lower than the base,
respectively.
(B) Stimulus set used during recordings. Each box
indicates a base/comparison frequency stimulus
pair. The number inside the box indicates overall
percentage of correct trials for that (f1, f2) stimulus pair, except when the stimulus pair was identical (22 Hz; we plotted the number of times that animal pressed
the lateral push button).
(C) Psychophysical performance when f1 was maintained fixed at 22 Hz and f2 was variable (red curve), and when f2 was fixed at 22 Hz and f1 was variable (green
curve). D.L. is the discrimination threshold in Hz.
(D) Top view of the monkey brain and the cortical areas recorded during perceptual discrimination (orange spots). Recordings were made in primary somatosen-
sory cortex (S1) and secondary somatosensory cortex (S2) contralateral to the stimulated hand (left hemisphere) and in primary motor cortex (M1) contralateral to
the responding hand/arm (right hemisphere). Recordings were made contralateral and ipsilateral to the stimulated fingertip in prefrontal cortex (PFC), ventral pre-
motor cortex (VPC), medial premotor cortex (MPC), and dorsal premotor cortex (DPC).
Neuron
Decision Making across Cortex
motor report, i.e., whether f2 > f1 or f2 < f1. Clearly, the neuronal
processes associated with the postponed decision report and
the task components that precede it can be analyzed across
diverse cortical areas.
Here we report the extent to which the stimulus identity is
encoded across diverse cortical areas in this task. We found
that the encoding of f1 and f2 through all task periods is widely
distributed across cortical areas. We also found that the activity
of frontal lobe circuits encodes both the result of the sensory
evaluation and past information on which those choices are
based. Notably, the activity of primary motor cortex (M1) showed
processes similar to those observed in the premotor areas
(ventral premotor cortex, VPC; dorsal premotor cortex, DPC;
and MPC) and prefrontal cortex (PFC), both during the compar-
ison and postponed decision report periods. These results
suggest that frontal lobe neurons have the capacity to encode
during the comparison and postponed decision report periods
both the final result of the sensory evaluation and past informa-
tion about it.
Here we also document the nature of the neuronal responses
during the stimuli and their interactions. In addition to the stan-
dard discrimination test, the neuronal activity of all cortical areas
was studied when the stimuli were delivered but monkeys were
not requested to perform the task. Under this condition, most
neurons across the cortical areas no longer encode information
about the stimuli and their interactions during these trials. The
only areas that responded in this case were S1 and S2. This
would suggest that those cortical areas central to S1 that encode
information about the stimuli are more likely associated with
the sensory evaluation, than engaged simply in encoding the
sensory stimulus. We also tested each neuron in a simpler
task, in which trials proceeded exactly as in the vibrotactile
task, but the stimuli were not delivered to the skin and the move-
ments were guided by visual cues. Neurons responded during
movement execution but not during the periods preceding it.
These control tests show that the neuronal responses from all
the cortical areas studied, except for S1, reflect both the active
comparisons between f1 and f2 and the execution of the motor
choice that is specific to the context of the vibrotactile discrimi-
nation task.
RESULTS
Optimal Conditions for Studying PerceptualDiscriminationFour monkeys (Macaca mulatta) were trained to discriminate the
difference in frequency between two consecutive vibrotactile
stimuli, f1 and f2 delivered to one fingertip (Figure 1A). Monkeys
were asked to report discrimination after a fixed delay period of
3 s between the end of f2 and the cue that triggered the motor
report (probe up, pu in Figure 1A). This delay period thus sepa-
rates the comparison between the two stimuli from the motor
response. In this task, monkeys must hold f1 in working memory,
must compare the current sensory input f2 to the memory trace
of f1, and must postpone the decision until the sensory cue
triggers the motor report. Animals were trained to perform the
task up to their psychophysical thresholds (Figures 1B and C).
After training, we recorded the activity of single neurons from
diverse cortical areas while the monkeys performed the task
(Figure 1D). These recordings were made in primary somatosen-
sory cortex (S1), secondary somatosensory cortex (S2), PFC,
VPC, DPC, and MPC contralateral to the stimulated finger and
in PFC, VPC, DPC, MPC, and M1 contralateral to the responding
hand/arm. All neurons were recorded using the stimulus set of
Figure 1B. In these trials, the comparison frequency (f2) can be
judged higher or lower than f1. Thus, the neuronal responses
across trials can be analyzed as functions of f1, f2, f2 � f1, or
as functions of the monkey’s two possible motor choices.
Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc. 301

Neuron
Decision Making across Cortex
Neuronal Responses across Cortical Areas duringPerceptual DiscriminationWe recorded from 2509 neurons (Table 1) that had average firing
rates that were significantly different from their firing rates during
a pre-trial control period (500 ms immediately before probe
down; pd in Figure 1A; p < 0.01, Wilcoxon rank-sum test [Siegel
and Castellan, 1988]). Some of these neurons enhanced or
reduced their firing rates (222/2509, 9%), but their firing rate
did not depend significantly on the applied stimulus frequencies
or the animal’s decision report (Experimental Procedures). Here
we focus exclusively on the 91% of neurons (n = 2287) that were
stimulus dependent (Romo et al., 1999), although not necessarily
during the stimulus presentation periods: stimulus-dependent
modulations could occur also during the working memory,
comparison and postponed decision report periods of the task
(Table 1). This is illustrated for an example neuron recorded in
the MPC contralateral to the stimulated finger (Figure 2A). The
discharge rate of this neuron during the delay period between
f1 and f2 varied as a monotonic function of f1. This neuron dis-
charged more strongly for the lower frequency and decreased
its firing rate steadily for increasing f1. We refer for this type of
response as ‘‘negative monotonic’’ (Romo et al., 1999). Other
neurons had discharge rates that varied in the opposite direc-
tion. We refer to those as ‘‘positive monotonic’’ (Romo et al.,
1999). Thus, for this example neuron the analysis showed that
f1 appeared to be encoded directly with various strengths and
at various periods of time during the delay period between f1
and f2. Neurons with positive or negative modulations were
also recorded in S2 contralateral to the stimulated hand and
bilaterally in PFC, VPC, DPC, and MPC. Neurons from these
areas could respond exclusively to the f1 presentation, or
encode f1 during the delay period between f1 and f2 (Figure 2A)
or during both periods. It is also worth mentioning that the large
majority of S1 neurons encoded f1 almost exclusively in a posi-
tive monotonic fashion during the first stimulus period only, and
that none of the M1 neurons showed any modulation of their
firing rates as a function of f1 during the f1 presentation or during
the delay period between f1 and f2 (Figure 3 and see Figure S1
available online).
As the task progressed, trials for the example neuron of
Figure 2A can be divided into two types: those on which f2 > f1
(black label) and those on which f2 < f1 (gray label). During the
final 200 ms of f2 presentation and early component (1 s) of
the postponed decision report period of our example neuron,
the firing rates were modulated by both f1 and f2. The main
determinant of the firing rate was not, however, the particular
values that f1 or f2 took on any given trial. Instead, it was simply
whether the trial belonged to the f2 > f1 group or f2 < f1 group.
This corresponds to the monkey’s two possible choices, repre-
sented during the comparison and postponed decision report
periods. Not only did f1 modulate the response to f2 in this
neuron, but even more notably, this happened such that by
the end of f2 and during the early component of the postponed
decision period, the responses became mostly correlated
with the monkey’s choice. This type of response that occurred
during the comparison and postponed decision report periods
was observed in area S2 contralateral to the stimulated hand
and bilaterally in PFC, VPC, DPC, and in M1 contralateral to
302 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
the responding arm (Figure 3), but not in S1. In brief, the
stimulus identity of f1 and f2 could be encoded in a positive
(S1, S2, PFC, VPC, DPC, and MPC) or negative (S2, PFC, VPC,
DPC, and MPC) monotonic fashion during the stimulus pre-
sentations. The f2 > f1 and f2 < f1 responses during the f2 and
postponed decision report periods recorded in S2, PFC, VPC,
MPC, and M1 suggest that they result from an interaction
between the two stimuli. This requires, however, understanding
precisely how f1 and f2 are encoded and how these interactions
are computed during the comparison and postponed decision
report periods.
Decoding Perceptual Discrimination across CorticalAreasTo further estimate the representation of f1 and f2 and their
interactions during the task components, we used multivariate
regression analysis. For each neuron, we modeled the firing
rate’s dependency on f1 and f2. In principle, the response during
f2 could be an arbitrary linear function of both f1 and f2: firing
rate(t) = a1(t) f1 + a2(t) f2 + a3(t) (Draper and Smith, 1966; Hernan-
dez et al., 2002; Press et al., 1992; Romo et al., 2002, 2004). In
this formulation, t represents time, and the coefficients a1 and
a2 serve as direct measurements of firing rate dependence on
f1 and f2, respectively. These measurements were calculated
in sliding windows of 200 ms moving in steps of 20 ms. To illus-
trate this analysis, the resulting coefficients a1 and a2 and
their interactions were plotted in panels C and D of Figure 2 for
the example neuron as a function of time. We also plotted the
values of a1 and a2 against each other to compare the responses
at different points during the task (Figure 2C). Three lines are
of particular relevance in these plots: points that fall on the
a1 = 0 axis represent responses that depend on f2 only (red dots
in Figure 4); points that fall on a2 = 0 axis represent responses
that depend on f1 only (green dots in Figure 4), and points that
fall near the a2 = �a1 line represent responses that are a func-
tion of f2� f1 only (black and blue dots in Figure 4). This last con-
sideration is of particular importance since the sign of the differ-
ence between f1 and f2 determines correct task performance.
However, the result of the analysis is not restricted to these three
conditions. For example, in those hypothetical cases when the
modulation imposed by f1 and f2 results in f1 + f2, the point
would fall close to a1 = a2 line. In this case, the memory of
f1 is added to the f2 representation, but this result was rarely
observed. Importantly, the larger area of the plane represents
those responses where the strengths of a1 and a2 are signifi-
cantly different from zero and significantly different from each
other. This plot can also reveal whether one of the two stimulus
frequencies is more strongly represented than the other (ja1jsja2j; a1 s 0; a2 s 0; black dots in panels C and D of Figure 2 and
black dots of Figure 4).
The population analysis of each cortical area shows that most
of the neurons encode f1 in a positive or negative monotonic
manner at various coefficients strengths. This result agrees
with results obtained from a large data base recorded in
monkeys reporting discrimination immediately after f2 (Romo
et al., 2004). As before, most S1 neurons encode f1 during the
stimulus presentation in a positive monotonic fashion whereas
the rest of cortical areas—except for M1 neurons which did

Table 1. Database
Task Component
Area Responsive Tuned f1 delay f1 � f2 f2 delay f2 � pu mt
S1 189 f1 59 + (31.2%)
5 � (2.6%)
f2 61 + (32.2%)
4 � (2.1%)
c
d
S2 426 f1 49 + (11.5%) 28 + (6.5%) 6 + (1.4%) 8 + (1.8%) 4 + (0.9 %)
45 � (10.5%) 21 � (4.9%) 5 � (1.1%) 6 � (1.4%) 3 � (0.7%)
f2 57 + (13.3%) 31 + (7.2%) 6 + (1.4%)
54 � (12.6%) 25 � (5.8 %) 5 � (1.1%)
c 34 (7.9%) 15 (3.5%) 12 (2.8%)
d 21 (4.9%) 26 (6.1%) 23 (5.4%)
VPC 375 f1 48 + (12.8%) 63 + (16.8%) 76 + (20.2%) 31 + (8.2%) 14 + (3.7%)
36 � (9.6%) 45 � (12.0%) 57 � (15.2%) 26 � (6.9%) 12 � (3.2%)
f2 47 + (12.5%) 27 + (7.2%) 15 + (4.0%)
30 � (8.0%) 25 � (6.6%) 7 � (1.8%)
c 62 (16.5%) 11 (2.9%) 5 (1.3%)
d 108 (28.8%) 68 (18.1%) 16 (4.3%)
PFC 358 f1 25 + (6.9%) 49 + (13.7%) 25 + (6.9%) 14 + (3.9%) 5 + (1.4%)
26 � (7.7%) 42 � (11.7%) 21 � (5.8%) 9 � (2.5%) 8 � (2.2%)
f2 35 + (9.7%) 27 + (7.5%) 11 + (3.0%)
21 � (8.0%) 31 � (8.7%) 13 � (3.6%)
c 53 (14.8%) 26 (7.3%) 3 (0.8%)
d 82 (22.9%) 72 (20.1%) 16 (4.5%)
MPC 494 f1 29 + (5.9%) 47 + (9.5%) 33 + (6.7%) 39 + (7.9%) 19 + (3.8%)
24 � (4.9%) 39 � (7.9%) 28 � (5.7%) 32 � (6.5%) 14 � (2.8%)
f2 63 + (12.8%) 67 + (13.6%) 16 + (3.2%)
41 � (8.3%) 59 � (11.9%) 17 � (3.4%)
c 53 (10.7%) 42 (8.5%) 13 (2.6%)
d 83 (16.8%) 74 (15.0%) 45 (9.1%)
DPC 164 f1 10 + (6.0%) 12 + (7.3%) 3 + (1.8%) 9 + (5.5%) 4 + (2.4%)
12 � (7.3%) 8 � (4.9%) 6 � (3.7%) 12 � (7.3%) 5 � (3.0%)
f2 15 + (9.1%) 11 + (6.7%) 9 + (5.5%)
16 � (9.8%) 13 � (7.9%) 10 � (6.1%)
c 9 (5.5%) 3 (1.8%) 3 (1.8%)
d 4 (2.4%) 9 (5.5%) 16 (9.8%)
M1 281 f1 2 + (0.7%) 3 + (1.0%) 3 + (1.0%) 17 + (6.0%) 4 + (1.4%)
3 � (1.0%) 4 � (1.4%) 7 � (2.4%) 24 � (8.5%) 6 � (2.1%)
f2 25 + (8.8%) 49 + (17.4%) 14 + (4.9%)
27 � (9.6%) 52 � (18.5%) 17 � (6.0%)
c 15 (5.3%) 27 (9.6%) 15 (5.3%)
d 17 (6.0%) 35 (12.5%) 29 (10.3%)
Recorded, n = 2509. Responsive, n = 2287 (91%). f1, first stimulus; f2 second stimulus; pu, probe up; mt, movement time; S1, primary somatosensory
cortex; S2, second somatosensory cortex; VPC, ventral premotor cortex; PFC, prefrontal cortex; MPC, medial premotor cortex; DPC, dorsal premotor
cortex; M1, primary motor cortex. Tuned to stimulus frequency with positive (+) or negative (�) slopes; c, tuned to stimuli with differential activity for
f2 > f1 or f2 < f1; d, differential activity to f2 > f1 or f2 < f1.
Neuron
Decision Making across Cortex
Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc. 303

Figure 2. Responses of a MPC Neuron
during the Discrimination Task and Control
Tests
(A) Raster plots of responses during the discrimi-
nation task. This neuron responded with an f1
negative monotonic fashion to the increasing stim-
ulus frequency f1 during the delay period between
f1 and f2 and during the early delay period
between the end of f2 and the beginning of the
decision motor report (pu). Each row of ticks is
a trial, and each tick is an action potential. Trials
were delivered in random order (10 trials per stim-
ulus pair). Labels at left indicate f1:f2 stimulus
pairs. Black indicates f2 > f1; gray indicates f2 < f1.
(B) Firing rate modulation (mean ± SEM) as a func-
tion of f1 or f2.
(C) Resulting coefficient values for f1 (a1, green)
and f2 (a2, red) for panels in (B).
(D) Coefficients values as functions of time. Green
and red traces correspond to a1 and a2, respec-
tively. Filled circles indicate significant values.
Black circles indicate points at which a1 and a2
were significant and of different magnitudes, but
had opposite signs; these are partially differential
(c) responses. Blue circles indicate points at which
a1 and a2 were significant and of similar magni-
tude but had opposite signs; these are fully differ-
ential (d) or categorical responses.
(E) Responses of the same neuron when the same
set of stimuli (A) was delivered to the fingertip, but
discrimination was restricted, just by removing the
key and the interrupt target switches. Thus, in this
condition the animal remained alert—by rewarding
with drops of liquid at different times—but was no
longer using the stimuli to indicate discrimination
with the free hand/arm. Under this test condition,
the neuron does not encode information about
the stimuli.
(F) Choice probability indices as function of time
during the discrimination task. Filled circles are
significant values that deviated from 0.5 (green
for f1 values; black for c values; and blue for
d values of D).
(G) Choice probability index for the same neuron tested in the light instruction task. Under this condition, the choice probability indices were calculated by
comparing the response distributions for lateral versus medial push button presses. Arm movements in this situation were identical to those in the vibrotactile
discrimination task but were cued by visual stimuli.
Neuron
Decision Making across Cortex
not encode f1 at all—do so in dual form: positive or negative
monotonic encoding (green dots in Figure 4). These encodings
are also observed during the delay period between f1 and f2,
except for S1 and M1 neurons, which do not show any encoding
during this period (green dots in Figure 4). During the comparison
period, most S1 neurons encoded f2 in a positive monotonic
fashion (red dots in Figure 4), whereas most of the cortical areas
encoded the current stimulus f2, but also f1 (Figure 4). Again, the
encoding of f1 and f2 could be positive or negative monotonic.
But, in addition, the information of f1 and f2 is combined to
generate a differential response (blue dots in Figure 4). An inter-
esting observation is that some neurons during the comparison
period switched from an f1 encoding to a combination of f2
and f1 (black dots in Figure 4), and then to a differential response
(blue dots in Figure 4). During the postponed decision report,
many neurons from PFC, VPC, MPC, DPC, and M1 encoded
information on which the decision is based and the resulting
304 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
operation, f2 > f1 or f2 < f1 (Figure 4). Again, S1 did not show
any sign of participation in the postponed decision report
(Figure 4).
Dynamics of the Perceptual Discrimination Processacross Cortical AreasFigure 4 shows the number of neurons with significant a1 and
a2 coefficients during the relevant components of the task.
However, it does not tell much about the dynamics of the popu-
lation response of each cortical area across the task compo-
nents. An analysis of the coefficients as functions of time shows
that all cortical areas studied here encoded f1 during the f1
presentation, except M1 (green traces in Figure 5A). The earliest
response began in S1 compared to S2, PFC, VPC, MPC, and
DPC, then S2 responded earlier than PFC, VPC, MPC, and
DPC, and finally PFC and VPC responded earlier than MPC
and DPC (Figure 6; p < 0.01 between response distributions;
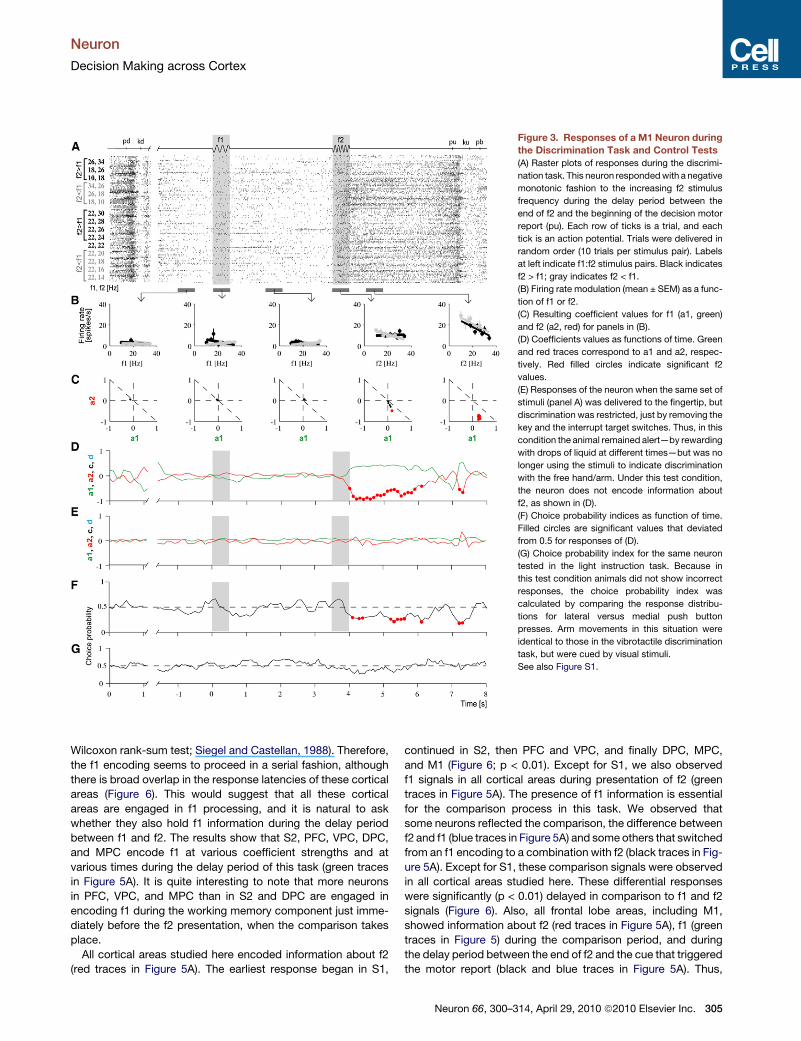
Figure 3. Responses of a M1 Neuron during
the Discrimination Task and Control Tests
(A) Raster plots of responses during the discrimi-
nation task. This neuron responded with a negative
monotonic fashion to the increasing f2 stimulus
frequency during the delay period between the
end of f2 and the beginning of the decision motor
report (pu). Each row of ticks is a trial, and each
tick is an action potential. Trials were delivered in
random order (10 trials per stimulus pair). Labels
at left indicate f1:f2 stimulus pairs. Black indicates
f2 > f1; gray indicates f2 < f1.
(B) Firing rate modulation (mean ± SEM) as a func-
tion of f1 or f2.
(C) Resulting coefficient values for f1 (a1, green)
and f2 (a2, red) for panels in (B).
(D) Coefficients values as functions of time. Green
and red traces correspond to a1 and a2, respec-
tively. Red filled circles indicate significant f2
values.
(E) Responses of the neuron when the same set of
stimuli (panel A) was delivered to the fingertip, but
discrimination was restricted, just by removing the
key and the interrupt target switches. Thus, in this
condition the animal remained alert—by rewarding
with drops of liquid at different times—but was no
longer using the stimuli to indicate discrimination
with the free hand/arm. Under this test condition,
the neuron does not encode information about
f2, as shown in (D).
(F) Choice probability indices as function of time.
Filled circles are significant values that deviated
from 0.5 for responses of (D).
(G) Choice probability index for the same neuron
tested in the light instruction task. Because in
this test condition animals did not show incorrect
responses, the choice probability index was
calculated by comparing the response distribu-
tions for lateral versus medial push button
presses. Arm movements in this situation were
identical to those in the vibrotactile discrimination
task, but were cued by visual stimuli.
See also Figure S1.
Neuron
Decision Making across Cortex
Wilcoxon rank-sum test; Siegel and Castellan, 1988). Therefore,
the f1 encoding seems to proceed in a serial fashion, although
there is broad overlap in the response latencies of these cortical
areas (Figure 6). This would suggest that all these cortical
areas are engaged in f1 processing, and it is natural to ask
whether they also hold f1 information during the delay period
between f1 and f2. The results show that S2, PFC, VPC, DPC,
and MPC encode f1 at various coefficient strengths and at
various times during the delay period of this task (green traces
in Figure 5A). It is quite interesting to note that more neurons
in PFC, VPC, and MPC than in S2 and DPC are engaged in
encoding f1 during the working memory component just imme-
diately before the f2 presentation, when the comparison takes
place.
All cortical areas studied here encoded information about f2
(red traces in Figure 5A). The earliest response began in S1,
continued in S2, then PFC and VPC, and finally DPC, MPC,
and M1 (Figure 6; p < 0.01). Except for S1, we also observed
f1 signals in all cortical areas during presentation of f2 (green
traces in Figure 5A). The presence of f1 information is essential
for the comparison process in this task. We observed that
some neurons reflected the comparison, the difference between
f2 and f1 (blue traces in Figure 5A) and some others that switched
from an f1 encoding to a combination with f2 (black traces in Fig-
ure 5A). Except for S1, these comparison signals were observed
in all cortical areas studied here. These differential responses
were significantly (p < 0.01) delayed in comparison to f1 and f2
signals (Figure 6). Also, all frontal lobe areas, including M1,
showed information about f2 (red traces in Figure 5A), f1 (green
traces in Figure 5) during the comparison period, and during
the delay period between the end of f2 and the cue that triggered
the motor report (black and blue traces in Figure 5A). Thus,
Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc. 305

Figure 4. Population Coefficient Values
across Cortical Areas during the Different
Components of the Discrimination Task
Each point represents one neuron with at least one
coefficient significantly different from zero. We
analyzed five periods: f1 (500 ms), delay between
f1 and f2 (3000 ms), f2 (500 ms), delay between
the end of f2 and pu (3000 ms), and during a period
posterior to pu (1000 ms). For each neuron, we
identified a 200 ms bin with the highest modulation
during each period. n = number of neurons.
Green and red circles correspond respectively
to neurons with significant a1 coefficients only or
a2 coefficients only. Black circles correspond to
neurons with both significant a1 and a2 coeffi-
cients of opposite signs but of significantly
different magnitudes; these are partially differen-
tial responses (c). Blue circles correspond to
neurons with both significant a1 and a2 coeffi-
cients, but of opposite signs and statistically equal
magnitude; these are fully differential or categor-
ical responses encoding f2 � f1 (d). S1, primary
somatosensory cortex; S2, secondary somato-
sensory cortex; VPC, ventral premotor cortex;
PFC, prefrontal cortex; MPC, medial premotor
cortex; DPC, dorsal premotor cortex; M1, primary
motor cortex.
Neuron
Decision Making across Cortex
during the postponed decision report period these cortical
circuits maintain in working memory all the elements associated
with this cognitive operation. However, we noticed that there
were more neurons in MPC, DPC, and M1 than in PFC and
VPC that carried information about f2 during the postponed deci-
sion period (red traces in Figure 5A), indicating that the most
recent information sensory information (f2) is more likely to be
kept in working memory than immediately-preceding sensory
information (f1).
State-Dependent Responses across Cortical AreasCortical population dynamics illustrated in Figure 5A shows that
neurons of diverse cortical areas encode the different task
components. But, to what extent are these neuronal events
associated with the task components and animal’s sensory eval-
uation?
Do these events occur only during the task execution or are
irrespective of the animal’s state? To answer these questions,
in addition to the standard test, many of the neurons that
306 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
encoded information about the stimuli
and motor choice were also tested in
a variant of the task (Experimental Proce-
dures). In this test, the neuronal activity
of these cortical areas was studied
when the stimuli were delivered but
monkeys were not requested to perform
the task. Under this condition, most
neurons across the cortical areas no
longer encoded information about the
stimuli and their interactions during the
task components (Figure 5B). The only
areas that responded in this case were S1 and S2. This would
suggest that those cortical areas central to S1 that encoded
information about the stimuli are more likely associated with
the sensory evaluation, than engaged in encoding the stimuli.
Choice Signals across Cortical AreasResponses during correct trials alone did not allow us to deter-
mine to what extent (f2 � f1)-dependent responses were corre-
lated with the sensory stimuli, or with the monkey’s action
choice. For each neuron of each cortical area we sorted the
responses into correct and errors trials and calculated a choice
probability index as a function of time (Britten et al., 1996; Green
and Swets, 1966; Romo et al., 2002). This quantified for each
stimulus (f1, f2) pair whether neuronal responses during error
trials were different from responses during correct trials (panel
F in Figures 2 and 3). If the responses are exclusively stimulus
dependent, they should show no differences between correct
and errors trials, except when the differences expected at
chance level result of the intrinsic variability of the neural activity.

Figure 5. Cortical Population Dynamics
during the Discrimination Task
(A) Percentage of neurons with significant co-
efficients as a function of time. Green and red
traces correspond to a1 and a2 coefficients,
respectively. Black traces indicate percentage of
neurons with a1 and a2 coefficients of opposite
sign but of different magnitudes. These neurons
combine differential response with a sensory
component. Blue traces indicate percentage of
neurons with coefficients a1 and a2 of opposite
sign but similar magnitude; these produce a differ-
ential signal.
(B) Percentage of neurons that responded during
passive stimulation. All these neurons are part of
the populations studied in (A). S1, primary somato-
sensory cortex; S2, secondary somatosensory
cortex; VPC, ventral premotor cortex; PFC, pre-
frontal cortex; MPC, medial premotor cortex; DPC,
dorsal premotor cortex; M1, primary motor cortex.
n = number of neurons.
Neuron
Decision Making across Cortex
In contrast, if the responses were dependent on the animal’s
choice, then they should vary according to which button the
monkey chose to press (panel F in Figures 2 and 3). For all
cortical areas, we computed a choice probability index sepa-
rately for the neurons that responded as a function of f1 (green
traces in Figure 7A), f2 (red traces in Figure 7A) or that depended
specifically on f2 – f1 (black and blue traces in Figure 7A). The
result of this analysis shows that very few neurons from S1
reflect significant differences between correct and error trials.
In contrast, many neurons from S2, PFC, VPC, DPC, MPC, and
M1 predicted in their activity the animal’s choice. These
responses began during the f1 stimulus presentation for areas
S2, PFC, VPC, DPC, and MPC, and were also present during
the delay period between f1 and f2 for areas PFC, VPC, MPC,
DPC, and M1, just before f2 presentation. During the f2 period,
all these areas except S1 had significant choice probability
indices (Figures 7A and S2). These signals were maintained
during the postponed decision report, during the delay between
the end of f2 and the cue that triggered the motor response
(Figures 7A and S2). These results suggest that neuronal activity
that correlates with error responses are widely distributed across
cortical areas, and that an error could be due to a failure to
encode the sensory inputs and their interactions during the
task components.
In addition to analyzing error trials, we tested each neuron in
a variant of the task, in which trials proceeded exactly as in the
vibrotactile task, but the stimuli were not delivered to the skin
and the movements were guided by visual cues (Experimental
Procedures). Thus, movements were triggered by visual cues.
Neurons responded exactly as in Figure 7A during movement
Neuron 66, 300–3
execution but not during the period
preceding it (Figure 7B). These control
tests show that some of the neuronal
responses from all the cortical areas
studied, except for S1, reflect the active
comparison between f1 and f2 whereas
some other neurons encode the motor choice that is specific
to the context of the vibrotactile discrimination task.
Neuronal Correlates of Bias Behavior across CortexIn the task used here, the decision must result of the difference
between f1 and f2 (choice = f(f2 – f1)). However, the discrimina-
tion thresholds plotted in Figure 1C suggest that subjects could
base their decisions by paying more attention to the weight of f2
than to the weight of f1 (choice = f(w2*f2 – w1*f1)). To further esti-
mate the weights w2 and w1, we used multivariate regression
analysis and plotted the subject’s performance as the percen-
tage of f2 > f1 correct responses for each stimulus pair—we
used only stimulus pairs labeled red and green in Figure 1C. Thus,
the resulting coefficients a1 and a2 of the multivariate regres-
sion analysis correspond to w1 and w2, respectively. If during
the decision subjects gave more weight to f2 than to the weight
of f1, f2 must be reported higher than f1. This is what the anal-
ysis shows on the data of Figure 1C (a1 = 4.3, a2 = 5.6; r =
a1/a2 = 0.76). If this analysis is carried out for each experiment
of the 312 reported here, we obtain a geometric mean of 0.79
(Figure 8A). On the other hand, we know that those neurons
with sensory differential activity (labeled in black in Figures 4
and 5) show coefficients a1 and a2 significant different from
zero and between each other. Therefore, it is natural to ask
whether the activity of these neurons is correlated with the
behavioral bias illustrated in Figure 8A. For example, if we use
the a1 and a2 values of the MPC neurons for those periods of
times that show the condition described above and calculate
the ratio a1/a2, we obtain the histogram of Figure 8B. This plot
shows that the larger percentage of neural responses (77%;
14, April 29, 2010 ª2010 Elsevier Inc. 307

Figure 6. Box Plots Illustrate Response Latency Distributions for f1
(Green), f2 (Red), Comparison (c, Black), and Differential (d, Blue)
across Cortical Areas
These boxes have lines at the lower quartile, median, and upper quartile
values. The whiskers are lines extending from each end of the boxes to
show the extent of the rest of the data. A comparative analysis (Wilcoxon
rank-sum test; Siegel and Castellan, 1988) of the response latencies between
the cortical areas showed that the f1 and f2 began earlier in S1 (p < 0.01) than in
Neuron
Decision Making across Cortex
308 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
data distribution to the left relative to 1 in Figure 8B) corresponds
to a1 < a2 responses. Such proportion of bias can not be ob-
tained by chance (p < 0.001; binomial test [100, 55, and 0.05]).
Interestingly, there is a strong relationship between the direc-
tion and magnitude of the bias behavior. To further establish in
which cortical area this bias is generated, we estimated the
neuronal bias (a1/a2) for each neuron of each cortical area using
a sliding window of 200 ms duration moving in steps of 20 ms
(S2, m = 0.79; VPC, m = 0.82; PFC, m = 0.81; MPC, m = 0.81;
M1, m = 0.86)—beginning at f2 onset and ending at probe up
that triggered the decision motor report—and compared this
value against the behavioral bias (a1/a2) obtained simulta-
neously in the same experiment. The results are shown
in Figure 8C. Except for S1, we found that a large fraction of
these neurons correlated with the behavioral bias. This neural
bias was more evident in the PFC, MPC, DPC, and M1 than in
S2 (Figure 8C). Thus, when one of the two stimulus frequencies
is more strongly represented than the other, it could bias
the psychophysical performance in this task. This interpretation
is consistent with the fact that f2 is more strongly represented
during the comparison and postponed decision periods than f1
(Figure 5).
DISCUSSION
To understand perceptual discrimination, we need to know
where in the brain are the physical relevant variables encoded
and what are their relative contributions to the final percept.
Our study focuses on this problem using highly simplified stimuli,
in which the neuronal responses from diverse cortical areas can
be examined while trained monkeys executed the same task.
Although not sufficiently exhaustive, this study shows how
cortical circuits are associated with perceptual discrimination.
For example, our results show that S1 is essentially sensory
and M1 is not necessarily primarily associated with motor
outputs only. Also, those cortical areas that receive the S1 inputs
combine the sensory representations of S1 with sensory signals
stored in working memory. Notably, these cortical areas encode
at various strengths and times the stimulus parameters of both
past and current sensory information on which the perceptual
decision report is based. Moreover, the sensory, memory and
comparison signals are gradually conveyed to the frontal lobe
S2, PFC, VPC, MPC, DPC, and M1 (f1 was not present in M1). The response
latencies for f1 and f2 in S2 (p < 0.01) began earlier than PFC, VPC, MPC,
DPC, and M1. The response latencies for f1 and f2 began earlier in PFC and
VPC (p < 0.01) than in MPC, DPC and M1. We found no differences in the
response latencies for f1 and f2 between MPC, DPC, and M1 (p > 0.01).
All f1 and f2 response latencies in all these cortical areas began earlier
(p < 0.01) than the comparison (c) and differential responses (d). We found
no statistical differences (p > 0.01) between the comparison and differential
responses across the cortical areas. L, left hemisphere (contralateral to the
stimulated hand); R, right hemisphere (ipsilateral to the stimulated hand).
Recordings in primary somatosensory cortex (S1) and secondary somatosen-
sory cortex (S2) were made contralateral to the stimulated hand (left hemi-
sphere) and in primary motor cortex (M1) contralateral to the responding
hand/arm (right hemisphere). Recordings were made bilaterally in prefrontal
cortex (PFC), ventral premotor cortex (VPC), medial premotor cortex (MPC),
and dorsal premotor cortex (DPC).

Figure 7. Correlation between Neuronal
Responses of Diverse Cortical Areas and
Behavioral Choice
(A) Percentage of neurons that had significant
choice probability indices as a function of time.
Green trace: neurons that encoded information
about f1; red trace: neurons that carried informa-
tion about f2; black trace: partially differential
neurons that carried information about f1 and f2
(c); blue trace: fully differential neurons that carried
information specifically about f2 � f1 only (d). See
also Figure S2.
(B) Percentage of the neurons in (A) that showed
significant choice probability indices during the
visual control task. In this test, animals had to
follow a visual cue to produce the motor choice
response. S1, primary somatosensory cortex;
S2, secondary somatosensory cortex; PFC, pre-
frontal cortex; VPC, ventral premotor cortex;
MPC, medial premotor cortex; DPC, dorsal premo-
tor cortex; M1, primary motor cortex. n = number of
neurons.
Neuron
Decision Making across Cortex
circuits that in turn drive the motor circuits for a movement
execution. Although this suggests a feedforward processing
beginning in S1 and ending in M1, this seems unlikely given feed-
back/recurrent communications between cortical and subcor-
tical areas (Lamme and Roelfsema, 2000). This problem is
currently addressed by recording the simultaneous activity of
neurons distributed across cortical circuits engaged in the task
used here (Hernandez et al., 2008).
This study shows how distinct cortical areas contribute to the
entire sequence of the processing steps that link sensation and
decision making. One could argue that the neuronal events
recorded in frontal lobe circuits during this task reflect other
processes, such as preparation for a future action, particularly
during the postponed decision report. This seems unlikely, how-
ever, because (1) delay responses between f1 and f2 depended
on f1 regardless of subsequent movements; (2) responses
during the postponed decision period often reflected f1 or f2
information; (3) choice probability indices indicated that there
were significant differences between correct versus error
trials—except for S1, variability in the responses of those S2
and frontal neurons associated with encoding the stimuli corre-
lates with the behavioral choice, although less stronger than
for the differential responses; (4) when the same movements
were guided by visual cues the differential activity disappeared,
except for some neurons that maintained their differential activity
during movement execution (Figure 7B); and (5) except for S1, all
these processes are dependent on active stimulus comparisons,
because they disappeared when subjects were not engaged in
solving the task (Figure 5B). We found it surprising that during
the comparison and postponed decision period some M1
neurons encoded information on which the decision is based.
This result could suggest that M1 is engaged in the readout of
sensory information from working memory, when it is required
to be compared with other sensory inputs, than engaged simply
in a motor response in this task. However, considering the
activity observed in other cortical areas notably in PFC, VPC,
MPC, and DPC during the same task, it would seem that this
process involved conjoined activity of these cortical areas, not
only during the postponed decision report, but also during the
task components preceding it. Thus, a comparison of the
strengths (Figure 4), dynamics (Figure 5A), and latencies (Fig-
ure 6) of the f1 and f2 responses and their interactions across
cortical areas is instructive.
Our results show that the strength of the f1 responses during
the stimulus period is stronger in S1 and gradually decreasing
in S2, PFC, VPC, MPC, and DPC (green dots in Figure 4). Also,
more neurons with f1 responses were recorded in S1, S2, PFC
and VPC than in MPC and DPC (green traces in Figure 5). This
suggests that f1 is preferentially encoded in some of the cortical
areas studied during this task. Accurate performance of the task
can be consistent only with a sensory percept elicited during the
f1 period. The lifetime of the percept directly induced by f1 could
not be measured, if it were not kept in working memory. It there-
fore remained possible that the lifetime of a quantitative, induced
percept was confined to the period of stimulation. Our results
show that the induced percept can be quantitatively memorized
as illustrated in Figures 4 and 5. Although the strength of this
signal varies across areas (green dots in Figure 4), all of them
except S1 and M1 store the value of f1 at different strengths
and times during the working memory component of the task
(green dots in Figure 4 and green traces in Figure 5A). These
results are in accord with the proposal that there is a large
cortical network that dynamically stores sensory information
during working memory (Fuster, 1997; Romo et al., 2004).
During the comparison period, f2 is processed similarly by the
same cortical areas and also in M1 (red dots in Figure 4 and red
Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc. 309

Figure 8. Neuronal Correlates of Bias Behavior
(A) Distribution of coefficients a1/a2 ratios (312 experiments in four animals),
obtained from linear regression analysis to the behavioral data. The histogram
shows that coefficient a2 has a stronger weight than coefficient a1.
(B) Bin distribution ratios for coefficients a1/a2 for neurons from medial premo-
tor cortex (MPC) that showed coefficients a1 and a2 significantly different from
zero and from each other. For each MPC neuron, we estimated the ratio
between weights a1/a2 in a sliding window of 200 ms moving in steps of
20 ms, beginning during the onset of the comparison period and ending during
the probe up that triggers the decision report. All these neurons showed the
properties described in B and illustrated in Figures 4 and 5 (black dots and
traces, respectively). This panel shows that coefficient a2 was more often
higher than coefficient a1, and consequently there are more bins to the left
relative to 1.
(C) Distribution of bin ratios for behavioral bias/neuron bias. For each neuron of
each cortical area, the resulting value a1/a2 of each cortical neuron was
compared against the behavioral value a1/a2 obtained simultaneously in the
same experiment. Data from primary somatosensory cortex (S1) are not
shown, since there are no neurons that show the properties described in (B).
DPC, dorsal premotor cortex; PFC, prefrontal cortex; M1, primary motor
Neuron
Decision Making across Cortex
310 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
traces in Figure 5A). Again, accurate performance of the task can
be consistent only with a sensory percept elicited during the f2
period. But, it is during the f2 period that the comparison
between stored (f1) and ongoing sensory information (f2) takes
place. During this period, f2 must not only be present, but f1
too, as shown in Figure 4 (red and green dots) and Figure 5
(red and green traces). The comparison between f1 and f2 is
observed in S2, VPC, PFC, MPC, DPC, and M1, again at various
strengths across these cortical areas (black dots in Figure 4 and
black traces in Figure 5). Some of these comparison signals
evolve into a signal that is consistent with the animal’s motor
choice (blue dots in Figure 4 and blue traces in Figure 5). During
the postponed report period, the activity of all cortical areas
except S1 encodes both the result of the comparison and past
information on which the decision is based (Figures 4 and 5A)
and covaried with the animal’s decision report (Figure 7A).
During the comparison and postponed delay periods, more
neurons in MPC, DPC, and M1 encoded f2 (red traces in Fig-
ure 5A) than information about f1 (green traces in Figure 5A)
and comparison signals (black and blue traces in Figure 5A).
This would suggest that these frontal circuits are more likely to
store recent sensory information (f2) than immediately preceding
sensory information (f1) during this task. Consistent with this
observation is the fact that the lifetime of the percept kept in
working memory seems to impact the decision report, as
observed in Figures 2B and 2C. These results suggest that
frontal lobe circuits do not simply wait for the result of a sensory
evaluation to be communicated but that actively participate in
this process. Although highly speculative, we suggest that main-
taining in working memory the original stimulus information on
which the decision is based could serve to continuously update
the postponed decision report in this task, and that very likely
depends on the conjoined activity of these cortical areas.
Assuming that neurons from distinct cortical circuits coordi-
nate their activities to solve this perceptual discrimination task,
we wonder how these events evolve in time. The comparative
analysis of the response latencies of f1, f2, and comparison
signals could shed some light on this problem. For instance,
compare S1 and S2: their response latencies were significantly
different (p < 0.01), with the f1 and f2 signals beginning earlier
in S1 than in S2 (Figure 6). This type of comparative analysis
also shows that the response latencies of S2 began significantly
earlier (p < 0.01) than in VPC, PFC, MPC, and DPC. This would
suggest that S2 could send information about the stimuli to these
frontal lobe circuits because their response types are quite
similar to S2 (Figures 4 and 5A). The question is whether frontal
lobe circuits receive at the same time S2 inputs or at different
times. An analysis of the response latencies for f1 and f2 showed
that the PFC and VPC respond significantly earlier (p < 0.01) than
DPC, MPC and M1 (f1 was not present in M1), with no significant
differences between PFC and VPC (p > 0.01). This would suggest
that the PFC and VPC receive the S2 inputs and that very likely
cortex; S2, secondary somatosensory cortex; VPC, ventral premotor cortex;
m, geometric mean (vertical line in each histogram). Gray bars in the histograms
show percentage of bins close to 1 (arbitrary range, 1 ± 0.22). A value of 1
means close correspondence between neuronal activity and behavioral
report.

Neuron
Decision Making across Cortex
distribute them to DPC, MPC, and M1. However, further studies
are needed to establish whether the functional connectivity
between these cortical circuits proceeds in such order in the
task used here.
The comparative analysis of the response latencies shows, in
this task, that sensory information processing proceeds in
a serial order. An important observation is that we did not find
significant differences (p > 0.01) in the response latencies
between the comparison and differential signals in the cortical
areas that showed these responses (Figure 6). The response
latencies for the comparison and differential signals were all
significantly (p < 0.01) delayed in comparison to the f1 and f2
signals. These findings suggest that S1 generates first a neural
representation of the stimulus (positive monotonic encoding)
and that then the S2 circuit transforms it into a dual representa-
tion (positive and negative monotonic encoding). The S2 repre-
sentations could be then used by the frontal lobe circuits to
not only encode the sensory information but also for the decision
process (Romo et al., 2003). Also, these findings suggest that
frontal lobe circuits coordinate the sensory, memory and deci-
sion components of the task, but that these processes are first
coordinated in PFC and VPC (Romo et al., 2004). These interpre-
tations, however, need further support by analyzing the neuronal
responses simultaneously recorded across cortical areas during
this task (Hernandez et al., 2008).
A fundamental problem posed by the results obtained in this
study is whether the neuronal signals across the task compo-
nents are associated with decision making or motor planning.
But, is there any difference between making a decision and plan-
ning a motor action that can be also postponed? In this respect,
the study by Gold and Shadlen (2000) is particularly revealing.
They recorded neural activity and injected microstimulation
current in the frontal eye field of monkeys performing a two-
alternative visual motion discrimination task. They found that
this area gradually accumulates evidence for motion in one or
another direction, such that the process of forming a decision
and motor preparation seem to be indistinguishable. The deci-
sion process studied here seems to proceed as it were part of
an encompassing, established plan. In this respect, decision
making could be conceived as the creation of a highly flexible
motor plan that can be delayed (if the action requires a go signal
to be triggered) or rapidly reconfigured (if the motor output is not
specified ahead of time), for example (Hernandez, et al., 2002;
Lemus et al., 2007; Romo et al., 2002, 2004). In fact, the stronger
signals that come closest to encoding the output of a decision
making process in our task have been found in areas involved
in motor actions: PFC, VPC, MPC, DPC, and M1. These results
fit quite well with the interpretation that those frontal lobe circuits
that show preparatory activity during delay periods not only
encode the planning of motor actions but encode also informa-
tion on which the motor action is based (Carpenter et al., 1999;
Hoshi and Tanji, 2004; Mushiake et al., 2006; Ohbayashi et al.,
2003; Shima et al., 2007).
Intuitively, the circuits that generate motor commands should
stand at the other end of the decision making processes
because their output needs to be expressed physically. Consis-
tent with this conjecture is the fact that some neurons from the
frontal lobe circuits respond differentially during the movement
execution in the light instruction task (Figure 7B), but not during
the periods that preceded it. This result is very similar to that
reported by Salinas and Romo (1998) in a categorization task.
Another possibility is that motor planning in this task is main-
tained in other circuits, for example, in the spinal cord (Prut
and Fetz, 1999). In this case weak signals sent from the cortical
lobe circuits could activate the execution of the motor plan in this
task. This conjecture is supported by the fact that the frontal lobe
circuits studied in our task send projections to the motor circuits
of the spinal cord (Dum and Strick, 1991; He et al., 1993).
However, few studies have explored the functional role of frontal
lobe neurons that project to the spinal cord during cognitive
tasks (Kraskov et al., 2009). Thus, further studies are needed
to explore the functional roles of frontal lobe circuit neurons
that send projections to the spinal cord, and whether spinal
motor circuits receive an instruction signal to execute the motor
plan in this task.
Our study is different from other paradigms used to investigate
perceptual discrimination processes (de Lafuente and Romo,
2005; Newsome et al., 1989). They have shown how neurons
from distinct cortical areas vary their discharges as a function
of varying a sensory stimulus and of the behavioral responses
(de Lafuente and Romo, 2006; Gold and Shadlen, 2007). Our
study focused on how neurons from several cortical areas
respond during decision making based on the evaluation of
two stimuli. In our task, decision making arises from the interac-
tion between stored information of f1 and current f2. Thus, the
fundamental mechanism behind decision making based on the
evaluation of one single stimulus or about two stimuli resides
on understanding the contribution of memory: how is it com-
bined with the current sensory input to produce a decision?
Although quite speculative, it is very likely that the working
memory signals associated with the delay period of our task is
a reflection of the stimulus recall triggered by the sensory
inputs. In this vein, it could be possible that the synapses of
the neuronal networks associated with the discrimination task
store the ‘‘task rule’’ (Mongillo et al., 2008), in which case the
scalar analog stimuli trigger synaptic graded responses, which
are then reflected in the neuronal firing rates. This mechanism
could be responsible for encoding the stimuli during the stimulus
periods, working memory and decision periods in cortical areas
central to S1.
In brief, this study shows how the dynamics of distinct cortical
circuits contribute to perceptual discrimination. However, to
reveal the flow of information between these circuits and the
operations described in our task, it would be desirable to simul-
taneously record the neuronal events (Hernandez et al., 2008).
Experiments of this type would reveal how neuronal populations
of distinct brain circuits joint efforts, in real time, to solve percep-
tual discrimination.
EXPERIMENTAL PROCEDURES
Discrimination Task
This study was performed on four male monkeys, Macaca mulatta, 5–7 kg. The
sensory discrimination task used here has already been described (Lemus
et al., 2007 and Figure 1). The monkey sat on a primate chair with its head fixed.
The right hand was restricted through a half-cast and kept in palm-up position.
The left hand operated an immovable key (elbow at �90�) and two push
Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc. 311

Neuron
Decision Making across Cortex
buttons in front of the animal, 25 cm away from the shoulder and at eye level.
The centers of the switches were located 7 and 10.5 cm to the left of the
midsagittal plane. In all trials, the monkey first placed the left hand and later
projected to one of the two switches. Stimuli were delivered to the skin of
the distal segment of one digit of the right, restrained hand, via a computer-
controlled stimulator (2 mm round tip, BME Systems). The initial indentation
was 500 mm (pd in Figure 1A). Vibrotactile stimuli were trains of short mechan-
ical pulses. Each of these pulses consisted of a single-cycle sinusoid lasting
20 ms. Stimulation amplitudes were adjusted to produce equal subjective
intensities (Hernandez et al., 1997; Mountcastle et al., 1990). During trials,
two vibrotactile stimuli (f1 and f2) were delivered consecutively to the glabrous
(hairless) skin of one fingertip, separated by a fixed interstimulus delay period
of 3 s. The monkey was asked to report discrimination of the two stimuli after
a fixed delay period of 3 s between the end of f2 and a cue signal that triggered
the beginning of the motor response (pu in Figure 1A). Discrimination was indi-
cated by pressing one of two pushbuttons with the left hand (lateral push
button for f2 > f1, medial push button for f2 < f1). The animal was rewarded
for correct discriminations with a drop of liquid. Performance was quantified
through psychometric techniques (Figures 1B and C). Monkeys were handled
according to the institutional standards of the National Institutes of Health and
Society for Neuroscience.
Because we were interested in finding cortical activity related to sensory
events, it was crucial to minimize or eliminate modulatory effects arising
from the well known dependence on arm movement direction (Georgopoulos
et al., 1988; Schwartz et al., 1988) or on parameter that covary with it. The
setup was thus arranged to filter out the classic directionally tuned responses.
The distance between target switches was 3.5 cm, and these were 18 cm away
from the immovable key. Thus, the difference between medial and lateral
movements was�11�. On average, the directional cells reported by Schwartz
et al. (1988, their Figure 13) fire at frequencies that range between �5 and
25 spikes/s, corresponding to their antipreferred and preferred directions,
respectively. Therefore, on average, directional cells modulate their firing rates
by�20 spikes/s when movement direction changes 180�. The expected effect
on an 11� change in direction is thus on the order of 1 spike/s. Under these
conditions some activity related to arm motion may be expected, but should
be practically identical for the two arm movements.
Visual Instruction Task
During recordings, we used a simpler control task. Trials in this test began
exactly as described above and in Figure 1A, except that when the probe
touched the skin, one of the target switches was illuminated. The monkey
had to respond by holding the immovable key. Then, after a long delay period
(8–9 s) the light was turned off and the probe was simultaneously lifted off from
the skin and triggered the hand/arm movement. The monkey was rewarded for
pressing the previously illuminated pushbutton. Arm movements in this situa-
tion were identical to those in the vibrotactile discrimination task, but were
cued by visual stimuli.
Passive Stimulation
In this condition, the same set of stimuli (Figure 1B) were delivered to the
fingertip while recording cortical neurons from diverse cortical areas, but
discrimination was restricted, just by removing the key and the interrupt target
switches. Thus, in this condition the animal remained alert—by rewarding with
drops of liquid at different times—but was no longer using the stimuli to indi-
cate discrimination with the free hand/arm.
Recordings
Neuronal recordings were obtained with an array of seven independent micro-
electrodes (2–3 MU; Romo et al., 1999) inserted into each cortical area, contra-
lateral (left hemisphere) or ipsilateral (right hemisphere) to the stimulated hand,
except for S1 in which recordings were contralateral to the stimulated hand
and in M1 (arm region) contralateral to the responding hand/arm. We collected
data using the stimulus set of Figure 1B, usually 10 trials per stimulus pair.
We used well-established electrophysiological and anatomical criteria to
distinguish between cortical areas (Figure 1C; Hernandez et al., 2002; Romo
et al., 1999, 2002, 2004; Salinas et al., 2000; Salinas and Romo, 1998). For
example, in S1 we recorded single neurons with cutaneous receptive fields
312 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
confined to the distal segments of the glabrous skin of fingertips 2, 3 or 4
and had quickly adapting properties. All neurons recorded in S2 had large
cutaneous receptive fields confined to the hand contralateral to the recording
site. All neurons in MPC were recorded in the pre-SMA. Pre-SMA is located
rostral to a line passing from the midline to the posterior edge of the arcuate
sulcus (Matsuzaka et al., 1992). Neuronal activity from VPC was recorded in
area F5 (Rizzolatti et al., 1988). Recordings in PFC were made anterior to the
arcuate sulcus and lateral to the principal sulcus (Romo et al., 1999). All record-
ings in DPC were made in the arm region of F2 (Rizzolatti and Luppino 2001).
This region is in front of M1 (F1), lateral to the central dimple, posterior to F7
and genu of arcuate sulcus (Rizzolatti and Luppino, 2001). Recordings in M1
were confined to the arm region of M1, confined to the anterior bank and crown
of the central sulcus, medial to the level of the posterior genu of the arcuate
sulcus and lateral to the central dimple (Rizzolatti and Luppino 2001). On all
recordings sessions in M1, acceptable penetrations sites were first identified.
The criterion was that, throughout the penetration track (maximum depth of
2000 mm), neurons were found that responded both during the task and to
passive movements of the contralateral arm. The passive responses had to
be related to shoulder and elbow joints; when they were associated with wrist
and finger movements the penetration was discarded. If these conditions were
met, then other neurons with different characteristics but recorded in the same
penetration were also studied and considered in the analysis. Recordings sites
changed from session to session and the locations of the penetrations were
used to construct surface maps of all the penetrations in each cortical area.
This was done by marking the edges of the small chamber (7 mm in diameter)
placed above each cortical area.
Data Analysis
We considered a neuron’s response as task-related if during any of the rele-
vant periods (f1, delay between f1 and f2, f2, delay between f2 and pu, reaction
time [RT] or movement time ([MT]) its mean firing rate was significantly different
from a control period of equal duration (500 ms) but preceding the initial probe
indentation at the beginning of each trial (Wilcoxon test, p < 0.01; Siegel and
Castellan, 1988). By definition, f1 and f2 correspond to the base and compar-
ison periods, respectively. The first delay was divided into consecutive inter-
vals of 500 ms beginning at the end of f1 and up to the beginning of f2. Similar
intervals were used for the second delay between the end of f2 and pu. The RT
was the period from the end of pu to the beginning of the key up (ku; Figure 1A).
The MT was the period from the end of ku to the beginning of the push button
press (pb; Figure 1A).
To estimate the a1, a2, and a3 coefficient values through multivariate regres-
sion analysis (Draper and Smith, 1966; Press et al., 1992), we used the 18 stim-
ulus frequency pairs labeled in green and red in Figure 1B. The correlation
coefficient between these stimulus pairs is zero, indicating that f1 and f2 are
absolutely linearly independent. After finding the best-fit coefficients a1 and
a2, differences between fitted and measured responses to the individual (f1,
f2) stimulus pairs were calculated, resulting in a full 2D covariance matrix of
errors (Press et al., 1992). Coefficients were considered significantly different
from (0, 0) if they were more than two standard deviations away. Neuronal
responses were defined unambiguously as dependent on f1 or f2 if the
coefficients of the planar fit were within two standard deviations of either the
a2 = 0 or the a1 = 0 lines, respectively. Responses were considered dependent
on f2 � f1 if the coefficients were more than 2 standard deviations away from
these two lines and within 2 standard deviations of the a2 = �a1 line
(Figure 2C). Responses not satisfying this criterion were classified as ‘‘mixed.’’
The dynamics of these coefficients was analyzed using a sliding window of
200 ms duration moving in steps of 20 ms.
The beginning of the f1 tuned response latency was estimated for each
neuron by identifying the first three consecutive 20 ms bins after f1 onset, in
which a1 was significantly different from zero and a2 was not significantly
different from zero. We then obtained a post-stimulus-time-histogram (PSTH,
500 ms immediately before f1 onset until the end of f1 using a time resolution
of 1 ms). The PSTH was smoothed using a 3-point moving average filter. We
then obtained the mean and standard deviation from the PSTH corresponding
to the 500 ms immediately before stimulus onset. From this, we identified the
first bin after the stimulus onset with a value higher to the mean and two
standard deviations of the control period (p < 0.01). The time of this bin was

Neuron
Decision Making across Cortex
considered as the response latency. The beginning of the f2 tuned responses
was similarly estimated for each neuron as for the f1 tuned response, but a2
was significantly different from zero and a1 was not significantly different
from zero. The beginning of the comparison response (c; black dots and traces
in Figures 2–5) was estimated as before but, (1) a1 and a2 were both signifi-
cantly different from zero; (2) either a1 or a2 was two standard deviations
away from the a2 = �a1 line.. The beginning of the differential response
(d; blue dots and traces in Figures 2–5) was estimated similarly as for the
comparison responses by identifying the first of three consecutive 20 ms
bins in which the coefficients a1 and a2 were significantly different from zero
and both coefficients were of equal magnitudes, within two standard devia-
tions of the a2 = �a1 line.
The choice probability index was calculated using methods from signal
detection theory (Green and Swets, 1966). This quantity measures the overlap
between two response distributions, in this case between correct and error
responses for each stimulus (f1, f2) pair. We restricted the analysis to those
stimulus (f1, f2) pairs for which the animals had between 30% and 70% of
errors. Notice that a value of 0.5 indicates full overlap, while 1 and 0 indicates
completely separate distributions (Figures 2 and 3). Thus, the choice proba-
bility index quantifies selectivity for one or the other outcome of the discrimi-
nation process. To compute it at different times, we used a sliding window
of 200 ms duration moving in 20 ms steps, beginning 1000 ms immediately
before f1 and ending 1000 ms immediately after the animal reported the
comparison between f2 and f1. To establish the significance of the choice
probability values, the neuronal responses in each time window were shuffled,
such that correct and error trials were randomized, and new choice probability
indices for the shuffled data were generated (permutation test). By comparing
the indices from the shuffled and unshuffled data and repeating the process
1000 times, we estimated the probability of obtaining choice probability values
as large or larger than those observed initially (with the unshuffled data) just by
chance (Figures 7A and S2). In the light instruction task, choice probability
indices were calculated by comparing the response distributions for lateral
versus medial push button presses (Figure 7B).
SUPPLEMENTAL INFORMATION
Supplemental Information includes two figures and can be found with this
article online at doi:10.1016/j.neuron.2010.03.031.
ACKNOWLEDGMENTS
The research of R.R. was partially supported by an International Research
Scholars Award from the Howard Hughes Medical Institute, and grants from
the Direccion del Personal Academico de la Universidad Nacional Autonoma
de Mexico and the Consejo Nacional de Ciencia y Tecnologıa. We thank Carlos
Brody, Andres Ojeda, and Emilio Salinas for invaluable comments.
Accepted: March 24, 2010
Published: April 28, 2010
REFERENCES
Britten, K.H., Newsome, W.T., Shadlen, M.N., Celebrini, S., and Movshon, J.A.
(1996). A relationship between behavioral choice and the visual responses of
neurons in macaque MT. Vis. Neurosci. 13, 87–100.
Brody, C.D., Hernandez, A., Zainos, A., and Romo, R. (2003). Timing and
neural encoding of somatosensory parametric working memory in macaque
prefrontal cortex. Cereb. Cortex 13, 1196–1207.
Carpenter, A.F., Georgopoulos, A.P., and Pellizzer, G. (1999). Motor cortical
encoding of serial order in a context-recall task. Science 283, 1752–1757.
Chow, S.S., Romo, R., and Brody, C.D. (2009). Context-dependent modula-
tion of functional connectivity: S2 to PFC connections in two-stimulus-interval
discrimination task. J. Neurosci. 29, 7238–7245.
de Lafuente, V., and Romo, R. (2005). Neuronal correlates of subjective
sensory experience. Nat. Neurosci. 8, 1698–1703.
de Lafuente, V., and Romo, R. (2006). Neural correlate of subjective sensory
experience gradually builds up across cortical areas. Proc. Natl. Acad. Sci.
USA 103, 14266–14271.
Draper, N., and Smith, H. (1966). Applied Regression Analysis, Second Edition
(New York: John Wiley and Sons).
Dum, R.P., and Strick, P.L. (1991). The origin of corticospinal projections from
the premotor areas in the frontal lobe. J. Neurosci. 11, 667–689.
Fuster, J.M. (1997). Network memory. Trends Neurosci. 20, 451–459.
Georgopoulos, A.P., Kettner, R.E., and Schwartz, A.B. (1988). Primate motor
cortex and free arm movements to visual targets in three-dimensional space.
II. Coding of the direction of movement by a neuronal population. J. Neurosci.
8, 2928–2937.
Gold, J.I., and Shadlen, M.N. (2000). Representation of a perceptual decision
in developing oculomotor commands. Nature 404, 390–394.
Gold, J.I., and Shadlen, M.N. (2007). The neural basis of decision making.
Annu. Rev. Neurosci. 30, 535–574.
Green, D.M., and Swets, J.A. (1966). Signal Detection Theory and Psycho-
physics (New York: John Wiley).
He, S.Q., Dum, R.P., and Strick, P.L. (1993). Topographic organization of
corticospinal projections from the frontal lobe: motor areas on the lateral
surface of the hemisphere. J. Neurosci. 13, 952–980.
Hernandez, A., Salinas, E., Garcıa, R., and Romo, R. (1997). Discrimination in
the sense of flutter: new psychophysical measurements in monkeys. J. Neuro-
sci. 17, 6391–6400.
Hernandez, A., Zainos, A., and Romo, R. (2000). Neuronal correlates of
sensory discrimination in the somatosensory cortex. Proc. Natl. Acad. Sci.
USA 97, 6191–6196.
Hernandez, A., Zainos, A., and Romo, R. (2002). Temporal evolution of a deci-
sion-making process in medial premotor cortex. Neuron 33, 959–972.
Hernandez, A., Nacher, V., Luna, R., Alvarez, M., Zainos, A., Cordero, S.,
Camarillo, L., Vazquez, Y., Lemus, L., and Romo, R. (2008). Procedure for
recording the simultaneous activity of single neurons distributed across
cortical areas during sensory discrimination. Proc. Natl. Acad. Sci. USA 105,
16785–16790.
Hoshi, E., and Tanji, J. (2004). Differential roles of neuronal activity in the
supplementary and presupplementary motor areas: from information retrieval
to motor planning and execution. J. Neurophysiol. 92, 3482–3499.
Jun, J.K., Miller, P., Hernandez, A., Zainos, A., Lemus, L., Brody, C.D., and
Romo, R. (2010). Heterogenous population coding of a short-term memory
and decision task. J. Neurosci. 30, 916–929.
Kraskov, A., Dancause, N., Quallo, M.M., Shepherd, S., and Lemon, R.N.
(2009). Corticospinal neurons in macaque ventral premotor cortex with mirror
properties: a potential mechanism for action suppression? Neuron 64,
922–930.
Lamme, V.A., and Roelfsema, P.R. (2000). The distinct modes of vision offered
by feedforward and recurrent processing. Trends Neurosci. 23, 571–579.
Lemus, L., Hernandez, A., Luna, R., Zainos, A., Nacher, V., and Romo, R.
(2007). Neural correlates of a postponed decision report. Proc. Natl. Acad.
Sci. USA 104, 17174–17179.
Luna, R., Hernandez, A., Brody, C.D., and Romo, R. (2005). Neural codes for
perceptual discrimination in primary somatosensory cortex. Nat. Neurosci.
8, 1210–1219.
Machens, C.K., Romo, R., and Brody, C.D. (2005). Flexible control of
mutual inhibition: a neural model of two-interval discrimination. Science 307,
1121–1124.
Matsuzaka, Y., Aizawa, H., and Tanji, J. (1992). A motor area rostral to the
supplementary motor area (presupplementary motor area) in the monkey:
neuronal activity during a learned motor task. J. Neurophysiol. 68, 653–662.
Mongillo, G., Barak, O., and Tsodyks, M. (2008). Synaptic theory of working
memory. Science 319, 1543–1546.
Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc. 313

Neuron
Decision Making across Cortex
Mountcastle, V.B., Steinmetz, M.A., and Romo, R. (1990). Frequency discrim-
ination in the sense of flutter: psychophysical measurements correlated with
postcentral events in behaving monkeys. J. Neurosci. 10, 3032–3044.
Mushiake, H., Saito, N., Sakamoto, K., Itoyama, Y., and Tanji, J. (2006). Activity
in the lateral prefrontal cortex reflects multiple steps of future events in action
plans. Neuron 50, 631–641.
Newsome, W.T., Britten, K.H., and Movshon, J.A. (1989). Neuronal correlates
of a perceptual decision. Nature 341, 52–54.
Ohbayashi, M., Ohki, K., and Miyashita, Y. (2003). Conversion of working
memory to motor sequence in the monkey premotor cortex. Science 301,
233–236.
Press, N., Teukolsky, S.A., Vettering, W.T., and Fannery, B.P. (1992). Numer-
ical Recipes in C, Second Edition (Cambridge: Cambridge University Press).
Prut, Y., and Fetz, E.E. (1999). Primate spinal interneurons show pre-move-
ment instructed delay activity. Nature 401, 590–594.
Rizzolatti, G., and Luppino, G. (2001). The cortical motor system. Neuron 31,
889–901.
Rizzolatti, G., Camarda, R., Fogassi, L., Gentilucci, M., Luppino, G., and
Matelli, M. (1988). Functional organization of inferior area 6 in the macaque
monkey. II. Area F5 and the control of distal movements. Exp. Brain Res. 71,
491–507.
Romo, R., and Salinas, E. (1999). Sensing and deciding in the somatosensory
system. Curr. Opin. Neurobiol. 9, 487–493.
Romo, R., and Salinas, E. (2003). Flutter discrimination: neural codes, percep-
tion, memory and decision making. Nat. Rev. Neurosci. 4, 203–218.
314 Neuron 66, 300–314, April 29, 2010 ª2010 Elsevier Inc.
Romo, R., Brody, C.D., Hernandez, A., and Lemus, L. (1999). Neuronal
correlates of parametric working memory in the prefrontal cortex. Nature
399, 470–473.
Romo, R., Hernandez, A., Zainos, A., Lemus, L., and Brody, C.D. (2002).
Neuronal correlates of decision-making in secondary somatosensory cortex.
Nat. Neurosci. 5, 1217–1225.
Romo, R., Hernandez, A., Zainos, A., and Salinas, E. (2003). Correlated
neuronal discharges that increase coding efficiency during perceptual discrim-
ination. Neuron 38, 649–657.
Romo, R., Hernandez, A., and Zainos, A. (2004). Neuronal correlates of
a perceptual decision in ventral premotor cortex. Neuron 41, 165–173.
Salinas, E., and Romo, R. (1998). Conversion of sensory signals into motor
commands in primary motor cortex. J. Neurosci. 18, 499–511.
Salinas, E., Hernandez, A., Zainos, A., and Romo, R. (2000). Periodicity and
firing rate as candidate neural codes for the frequency of vibrotactile stimuli.
J. Neurosci. 20, 5503–5515.
Schwartz, A.B., Kettner, R.E., and Georgopoulos, A.P. (1988). Primate motor
cortex and free arm movements to visual targets in three-dimensional space.
I. Relations between single cell discharge and direction of movement.
J. Neurosci. 8, 2913–2927.
Shadlen, M.N., and Newsome, W.T. (1996). Motion perception: seeing and
deciding. Proc. Natl. Acad. Sci. USA 93, 628–633.
Shima, K., Isoda, M., Mushiake, H., and Tanji, J. (2007). Categorization of
behavioural sequences in the prefrontal cortex. Nature 445, 315–318.
Siegel, S., and Castellan, N.J. (1988). Nonparametric Statistics for Behavioral
Sciences (New York: McGraw-Hill).




















