
Danger is worse when it moves: Neural and behavioral indices of enhanced attentional capture by...
6
Neuropsychologia 47 (2009) 364–369 Contents lists available at ScienceDirect Neuropsychologia journal homepage: www.elsevier.com/locate/neuropsychologia Danger is worse when it moves: Neural and behavioral indices of enhanced attentional capture by dynamic threatening stimuli Luis Carretié a,∗ , José A. Hinojosa b , Sara López-Martín a , Jacobo Albert a , Manuel Tapia a , Miguel A. Pozo b a Facultad de Psicología, Universidad Autónoma de Madrid, 28049 Madrid, Spain b Instituto Pluridisciplinar, Universidad Complutense de Madrid, 28040 Madrid, Spain article info Article history: Received 31 March 2008 Received in revised form 26 August 2008 Accepted 8 September 2008 Available online 13 September 2008 Keywords: Emotion Automatic attention Motion ERPs P1 abstract Both dynamic non-emotional stimuli (moving dots or digits) and danger-related static stimuli have pre- viously shown to capture attention. This study explored whether the combination of the two factors (i.e., threatening moving stimuli), frequent in natural situations, enhances attentional capture. To this end, static and moving distractors containing emotionally negative and non-negative information were pre- sented to 30 volunteers while they were engaged in a digit categorization task. Behavioral responses and event-related potentials (ERPs) were analyzed. Behavioral and electrophysiological data were con- vergent: moving negative distractors produced the longest reaction times in the digit categorization task, and elicited the highest amplitudes in the P1 component of the ERPs (peaking at 112 ms), an electrophys- iological signal of attentional capture. These results suggest that motion provides additional salience to threatening stimuli that facilitates attentional capture. © 2008 Elsevier Ltd. All rights reserved. 1. Introduction Several data suggest that moving stimuli are detected more eas- ily and more quickly than static stimuli, and that moving distractors tend to be efficient in capturing attention, disrupting the ongoing task (Franconeri & Simons, 2003, 2005; McLeod, Driver, & Crisp, 1988; Von Mühlenen, Rempel, & Enns, 2005). Thus, behavioral experiments confirm the advantage of motion over other phys- ical features such as luminance or color for capturing attention (Franconeri & Simons, 2003, 2005). In line with this, peripheral moving distractors cause visual cortex activation when subjects are engaged in a primary task concerning centrally located visual stim- uli, this activation being greater than that elicited by motionless distractors (Rees, Frith, & Lavie, 2001). However, the extent to which moving distractors gain access to processing systems appears to be modulated by several factors, since dynamic stimuli are not capable of capturing attention under certain circumstances. For example, static but not dynamic stimuli capture attention within a context of moving stimuli (e.g., Pinto, Olivers, & Theeuwes, 2006). The pro- cessing load involved in the primary task is also a limiting factor: visual cortical areas responsible for processing motion stimulation are less activated by moving distractors as the involvement in the ongoing task increases (Buchel et al., 1998; Rees, 1997; Rees et ∗ Corresponding author. Fax: +34 914975215. E-mail address: [email protected] (L. Carretié). al., 2001). Another important albeit unexplored modulating factor might be the information contained in the moving distractor itself. In the studies mentioned above, moving stimuli typically con- sist of dots or digits. An interesting question worth to be explored is whether the attentional capture facilitated by dynamic stimuli is enhanced when they contain highly relevant information. Several experiments using static stimuli have suggested that emotionally negative events capture attention to a greater extent than non- negative events (e.g., Carretié, Hinojosa, Martín-Loeches, Mercado, & Tapia, 2004; Doallo, Holguin, & Cadaveira, 2006; Pratto & John, 1991; Vuilleumier, Armony, Driver, & Dolan, 2001). But, also in this case, several factors may modulate this pattern. For example, deep involvement in the primary task reduces the capability of negative stimuli to capture attention (Doallo et al., 2006; Pessoa, Kastner, & Ungerleider, 2002; Schwartz et al., 2005). Moreover, low salience negative events fail to attract attention in normal populations (see reviews in Siegle, Ingram, & Matt, 2002; Williams, Mathews, & MacLeod, 1996). It could be expected that motion is also a modulat- ing factor, providing additional salience to negative stimuli for cap- turing attention. In biological terms, this enhanced salience of nega- tive events provided by motion would involve clear adaptive advan- tages. In fact, negative events such as the presence of predators are not well detected when these are still (Pinto, Jankowski, & Sesack, 2003) or when they camouflage their motion (Anderson, 2003). Therefore, the conjunction of danger-related information and motion would maximize attentional capture. To the best of our knowledge, there are as yet no experimental data on the direct 0028-3932/$ – see front matter © 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.neuropsychologia.2008.09.007
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Danger is worse when it moves: Neural and behavioral indices of enhanced attentional capture by...

Da
La
b
a
ARRAA
KEAMEP
1
itt1ei(meudmmosocvao
0d
Neuropsychologia 47 (2009) 364–369
Contents lists available at ScienceDirect
Neuropsychologia
journa l homepage: www.e lsev ier .com/ locate /neuropsychologia
anger is worse when it moves: Neural and behavioral indices of enhancedttentional capture by dynamic threatening stimuli
uis Carretiéa,∗, José A. Hinojosab, Sara López-Martína, Jacobo Alberta, Manuel Tapiaa, Miguel A. Pozob
Facultad de Psicología, Universidad Autónoma de Madrid, 28049 Madrid, SpainInstituto Pluridisciplinar, Universidad Complutense de Madrid, 28040 Madrid, Spain
r t i c l e i n f o
rticle history:eceived 31 March 2008eceived in revised form 26 August 2008ccepted 8 September 2008
a b s t r a c t
Both dynamic non-emotional stimuli (moving dots or digits) and danger-related static stimuli have pre-viously shown to capture attention. This study explored whether the combination of the two factors (i.e.,threatening moving stimuli), frequent in natural situations, enhances attentional capture. To this end,static and moving distractors containing emotionally negative and non-negative information were pre-
vailable online 13 September 2008
eywords:motionutomatic attentionotion
sented to 30 volunteers while they were engaged in a digit categorization task. Behavioral responsesand event-related potentials (ERPs) were analyzed. Behavioral and electrophysiological data were con-vergent: moving negative distractors produced the longest reaction times in the digit categorization task,and elicited the highest amplitudes in the P1 component of the ERPs (peaking at 112 ms), an electrophys-iological signal of attentional capture. These results suggest that motion provides additional salience to
cilita
am
sieenn&1cisUnrMi
RPs1
threatening stimuli that fa
. Introduction
Several data suggest that moving stimuli are detected more eas-ly and more quickly than static stimuli, and that moving distractorsend to be efficient in capturing attention, disrupting the ongoingask (Franconeri & Simons, 2003, 2005; McLeod, Driver, & Crisp,988; Von Mühlenen, Rempel, & Enns, 2005). Thus, behavioralxperiments confirm the advantage of motion over other phys-cal features such as luminance or color for capturing attentionFranconeri & Simons, 2003, 2005). In line with this, peripheral
oving distractors cause visual cortex activation when subjects arengaged in a primary task concerning centrally located visual stim-li, this activation being greater than that elicited by motionlessistractors (Rees, Frith, & Lavie, 2001). However, the extent to whichoving distractors gain access to processing systems appears to beodulated by several factors, since dynamic stimuli are not capable
f capturing attention under certain circumstances. For example,tatic but not dynamic stimuli capture attention within a contextf moving stimuli (e.g., Pinto, Olivers, & Theeuwes, 2006). The pro-
essing load involved in the primary task is also a limiting factor:isual cortical areas responsible for processing motion stimulationre less activated by moving distractors as the involvement in thengoing task increases (Buchel et al., 1998; Rees, 1997; Rees et∗ Corresponding author. Fax: +34 914975215.E-mail address: [email protected] (L. Carretié).
tttn2
mk
028-3932/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.oi:10.1016/j.neuropsychologia.2008.09.007
tes attentional capture.© 2008 Elsevier Ltd. All rights reserved.
l., 2001). Another important albeit unexplored modulating factoright be the information contained in the moving distractor itself.In the studies mentioned above, moving stimuli typically con-
ist of dots or digits. An interesting question worth to be exploreds whether the attentional capture facilitated by dynamic stimuli isnhanced when they contain highly relevant information. Severalxperiments using static stimuli have suggested that emotionallyegative events capture attention to a greater extent than non-egative events (e.g., Carretié, Hinojosa, Martín-Loeches, Mercado,Tapia, 2004; Doallo, Holguin, & Cadaveira, 2006; Pratto & John,
991; Vuilleumier, Armony, Driver, & Dolan, 2001). But, also in thisase, several factors may modulate this pattern. For example, deepnvolvement in the primary task reduces the capability of negativetimuli to capture attention (Doallo et al., 2006; Pessoa, Kastner, &ngerleider, 2002; Schwartz et al., 2005). Moreover, low salienceegative events fail to attract attention in normal populations (seeeviews in Siegle, Ingram, & Matt, 2002; Williams, Mathews, &acLeod, 1996). It could be expected that motion is also a modulat-
ng factor, providing additional salience to negative stimuli for cap-uring attention. In biological terms, this enhanced salience of nega-ive events provided by motion would involve clear adaptive advan-ages. In fact, negative events such as the presence of predators are
ot well detected when these are still (Pinto, Jankowski, & Sesack,003) or when they camouflage their motion (Anderson, 2003).Therefore, the conjunction of danger-related information andotion would maximize attentional capture. To the best of our
nowledge, there are as yet no experimental data on the direct

sychologia 47 (2009) 364–369 365
eetiswscictaoropoppmHy
2
2
aTatuooad
2
stmnpfnpes6ipmc
saricedotdtactp
Fig. 1. Examples of static or S stimuli (top) and moving or M stimuli (bottom) belong-ifSh
2
Iderdm
l(ofitiwema(s(
2
2
paaq
L. Carretié et al. / Neurop
xploration of this hypothesis through the use of biologically rel-vant dynamic stimuli. In the present study we aim to comparehe effect of static and moving peripheral distractors contain-ng negative and non-negative information. In order to equalizeemantic/categorical influences across conditions, all distractorsill consist of insects. Insects are natural stimuli that may be more
uitable for the exploration of our hypothesis, framed in biologi-al terms, than ‘culture-mediated’ threatening stimuli. Moreover,nsects can actually either move or remain static in natural cir-umstances, and may be displayed at close to realistic sizes byypical stimulation instruments. The primary, cognitive task willsk from subjects to categorize two digits appearing at the centerf the screen. Performance in this task will be measured througheaction times and number of errors in order to analyze the levelf interference caused by distractors. Additionally, event-relatedotentials (ERPs) will be recorded in order to facilitate the detectionf rapid neural changes underlying motion–emotion interactionrocesses. Several studies report that P1, an early sensory com-onent of ERPs peaking around 100 ms, constitutes an index ofobilization of automatic attentional resources (see a review inopfinger & Mangun, 2001; see also Carretié et al., 2004), so anal-ses will focus on this component.
. Materials and methods
.1. Subjects
Thirty students (26 women) from the Universidad Autónoma de Madrid, withn age range of 20–40 years (mean = 23.89, S.D. = 3.91), took part in this experiment.hey participated voluntarily and reported normal or corrected-to-normal visualcuity. All subjects provided their informed consent, and received course credit forheir participation. These subjects were selected from a wider sample of 115 vol-nteers as those who reported fear of both spiders and cockroaches: they scoredver 6 on a 0 (nil fear) to 10 (maximum fear) scale of fears of 18 different animals,bjects and situations. On the other hand, they all scored under 4 for ladybirdsnd butterflies (see next section). Selected subjects reported to be free of affectiveisorders.
.2. Stimuli and procedure
As illustrated in Fig. 1, each presentation included a set of visual elements,ome of them appearing in the center of the screen (relevant for the primaryask) and others peripherally (distractors). Peripheral elements were of four types:
oving negative (M−), static negative (S− moving non-negative (M0) and staticon-negative (S0). Negative stimuli contained a spider (50%) or a cockroach (50%) aseripheral elements (i.e., frightening or aversive for the subjects, who had reported
ear of spiders and cockroaches, as explained in the Subjects section), and non-egative contained butterflies (50%) or ladybirds (50%). Average height of theseeripheral elements, in visual angle, was 0.26◦ . In the case of M stimuli, these periph-ral elements moved in a rectilinear path starting from the bottom border of thecreen and going towards the center of the screen. Length of this vertical path was.6◦ . Peripheral elements appeared and remained static in the middle of that path
n S stimuli. Both S and M stimuli lasted 300 ms (M stimuli were composed of sixhotograms that resulted in a 22◦/s speed). Intertrial interval was 2000 ms. An ani-ation reproducing several S and M trials as well as their temporal characteristics
an be seen at http://www.uam.es/carretie/grupo/movingdanger.htm.The primary task concerned the central visual element, which consisted of a
equence of 4 digits (height: 1.59◦). Those relevant for the task were the secondnd the third (i.e., the central digits): participants had to press, “as accurately andapidly as possible”, one key if both the second and the third digits were even orf both were odd (i.e., if they were “concordant”), and a different key if those twoentral digits were of different types (i.e., if they where “discordant”). Within eachmotional and motion condition (M−, M0, S−, and S0), half of the trials were concor-ant and the other half were discordant. The first and fourth digits in the sequence,
r flankers, were always different from the central digits, and were included inhe sequence in order to increase task difficulty and, hence, top-down attentionalemand. The even/odd condition of the flankers was controlled: within each emo-ional and motion category, these flanker digits were of the same even/odd categorys the central digits (1st = 2nd and 4th = 3rd) in half of the trials, and of a differentategory (1st /= 2nd and 4th /= 3rd) in the other half. Total number of presenta-ions was 160 (40 for each of the stimulus categories: M−, M0, S−, and S0). Order ofresentation was random.Kmrpiacia
ng to the same category (emotionally negative). In the M example, photogramsorming the actual stimulus are superimposed. An animation reproducing several
and M trials as well as their temporal characteristics can be downloaded fromttp://www.uam.es/carretie/grupo/movingdanger.htm.
.3. Recording
Electroencephalographic data were recorded using an electrode cap (ElectroCapnternational) with tin electrodes. A total of 60 electrode locations homogeneouslyistributed all over the scalp were used. All active scalp electrodes (58), as well as onelectrode at the left mastoid (M1), were originally referenced to an electrode at theight mastoid (M2). For the entire sample of electrodes, originally M2-referencedata were re-referenced offline using the average of the mastoids (M1 and M2)ethod.
Electrooculographic (EOG) data were recorded from supra- and infra-orbitallyocated electrodes (vertical EOG) as well as from the left versus the right orbital rimhorizontal EOG). Electrode impedances were always kept below 3 k�. A bandpassf 0.1–50 Hz (3 dB points for −6 dB/octave roll-off) was set for the recording ampli-ers. The channels were continuously digitizing data at a sampling rate of 250 Hz forhe entire duration of the recording session. The continuous recording was dividednto 1000 ms epochs for each trial, starting 200 ms before stimulus onset. Trials in
hich subjects pressed the wrong button or to which they did not respond to wereliminated. Moreover, visual inspection was also carried out, epochs with eye move-ents or blinks being eliminated. This artifact and error rejection led to the average
cceptance of 28.93 S− trials (standard error of means, S.E.M.: 0.63), 30.93 M− trialsS.E.M.: 0.67), 27.50 S0 trials (S.E.M.: 0.71), and 27.57 M0 trials (S.E.M.: 0.95), and noignificant differences existed in this respect for the Motion by Emotion interactionwhich was the relevant contrast, as explained later: F(1,29) = 3.94, p > 0.05).
.4. Data analysis
.4.1. Detection and quantification of ERP componentsWith the aim of testing whether P1 component was present in the ERPs, com-
onents explaining most of the ERP variance in the temporal domain were detectednd quantified through covariance-matrix-based temporal principal componentnalysis (tPCA). This strategy has been widely recommended for detection anduantification of ERP components (e.g., Chapman & McCrary, 1995; Coles, Gratton,ramer, & Miller, 1986; Dien, Beal, & Berg, 2005; Donchin & Heffley, 1978). Theain advantage of tPCA over traditional procedures based on visual inspection of
ecordings and on ‘temporal windows of interest’ is that it presents each ERP com-
onent separately and with its ‘clean’ shape, extracting and quantifying it free of thenfluences of adjacent or subjacent components. Indeed, the waveform recorded atsite on the head over a period of several hundreds of milliseconds represents a
omplex superposition of different overlapping electrical potentials. Such record-ngs can stymie visual inspection. In brief, tPCA computes the covariance betweenll ERP time points, which tends to be high between those time points involved in
3 sychologia 47 (2009) 364–369
tTctqbwA
srdtifrforcatsfsb
2
eatotat2RM
3
3
(gaicPiii
2ti(s
(rit(wcfpNE
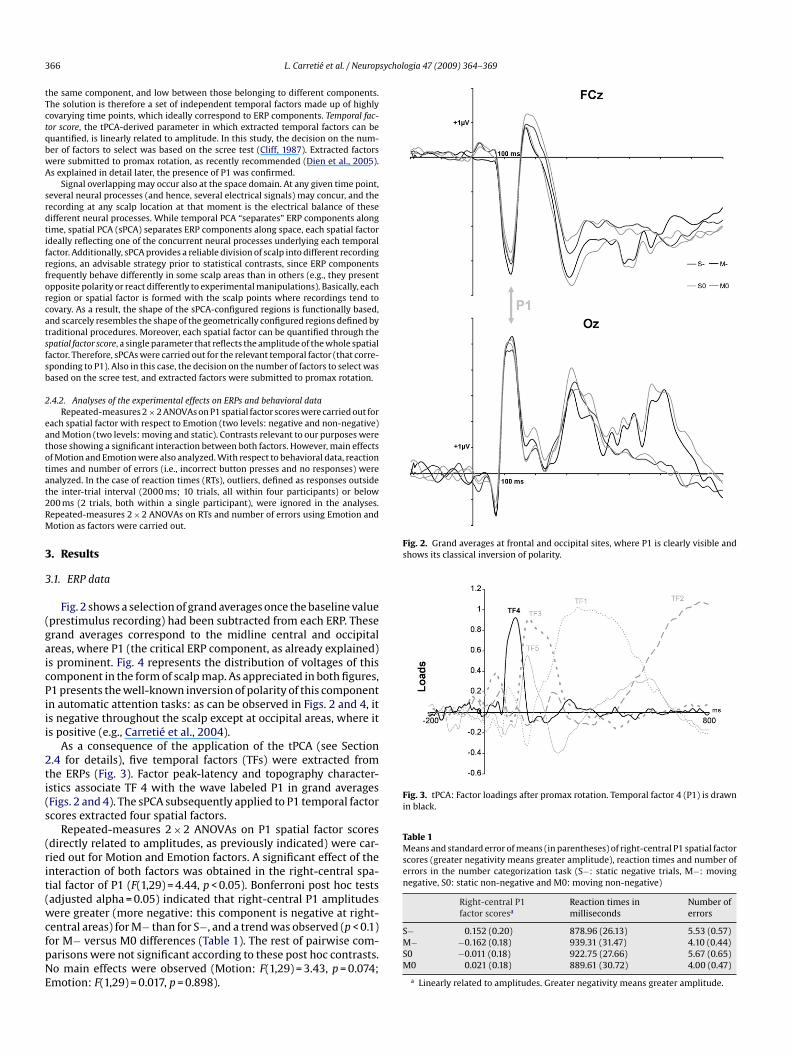
Fig. 2. Grand averages at frontal and occipital sites, where P1 is clearly visible andshows its classical inversion of polarity.
Fig. 3. tPCA: Factor loadings after promax rotation. Temporal factor 4 (P1) is drawnin black.
Table 1Means and standard error of means (in parentheses) of right-central P1 spatial factorscores (greater negativity means greater amplitude), reaction times and number oferrors in the number categorization task (S−: static negative trials, M−: movingnegative, S0: static non-negative and M0: moving non-negative)
Right-central P1factor scoresa
Reaction times inmilliseconds
Number oferrors
66 L. Carretié et al. / Neurop
he same component, and low between those belonging to different components.he solution is therefore a set of independent temporal factors made up of highlyovarying time points, which ideally correspond to ERP components. Temporal fac-or score, the tPCA-derived parameter in which extracted temporal factors can beuantified, is linearly related to amplitude. In this study, the decision on the num-er of factors to select was based on the scree test (Cliff, 1987). Extracted factorsere submitted to promax rotation, as recently recommended (Dien et al., 2005).s explained in detail later, the presence of P1 was confirmed.
Signal overlapping may occur also at the space domain. At any given time point,everal neural processes (and hence, several electrical signals) may concur, and theecording at any scalp location at that moment is the electrical balance of theseifferent neural processes. While temporal PCA “separates” ERP components alongime, spatial PCA (sPCA) separates ERP components along space, each spatial factordeally reflecting one of the concurrent neural processes underlying each temporalactor. Additionally, sPCA provides a reliable division of scalp into different recordingegions, an advisable strategy prior to statistical contrasts, since ERP componentsrequently behave differently in some scalp areas than in others (e.g., they presentpposite polarity or react differently to experimental manipulations). Basically, eachegion or spatial factor is formed with the scalp points where recordings tend toovary. As a result, the shape of the sPCA-configured regions is functionally based,nd scarcely resembles the shape of the geometrically configured regions defined byraditional procedures. Moreover, each spatial factor can be quantified through thepatial factor score, a single parameter that reflects the amplitude of the whole spatialactor. Therefore, sPCAs were carried out for the relevant temporal factor (that corre-ponding to P1). Also in this case, the decision on the number of factors to select wasased on the scree test, and extracted factors were submitted to promax rotation.
.4.2. Analyses of the experimental effects on ERPs and behavioral dataRepeated-measures 2 × 2 ANOVAs on P1 spatial factor scores were carried out for
ach spatial factor with respect to Emotion (two levels: negative and non-negative)nd Motion (two levels: moving and static). Contrasts relevant to our purposes werehose showing a significant interaction between both factors. However, main effectsf Motion and Emotion were also analyzed. With respect to behavioral data, reactionimes and number of errors (i.e., incorrect button presses and no responses) werenalyzed. In the case of reaction times (RTs), outliers, defined as responses outsidehe inter-trial interval (2000 ms; 10 trials, all within four participants) or below00 ms (2 trials, both within a single participant), were ignored in the analyses.epeated-measures 2 × 2 ANOVAs on RTs and number of errors using Emotion andotion as factors were carried out.
. Results
.1. ERP data
Fig. 2 shows a selection of grand averages once the baseline valueprestimulus recording) had been subtracted from each ERP. Theserand averages correspond to the midline central and occipitalreas, where P1 (the critical ERP component, as already explained)s prominent. Fig. 4 represents the distribution of voltages of thisomponent in the form of scalp map. As appreciated in both figures,1 presents the well-known inversion of polarity of this componentn automatic attention tasks: as can be observed in Figs. 2 and 4, its negative throughout the scalp except at occipital areas, where its positive (e.g., Carretié et al., 2004).
As a consequence of the application of the tPCA (see Section.4 for details), five temporal factors (TFs) were extracted fromhe ERPs (Fig. 3). Factor peak-latency and topography character-stics associate TF 4 with the wave labeled P1 in grand averagesFigs. 2 and 4). The sPCA subsequently applied to P1 temporal factorcores extracted four spatial factors.
Repeated-measures 2 × 2 ANOVAs on P1 spatial factor scoresdirectly related to amplitudes, as previously indicated) were car-ied out for Motion and Emotion factors. A significant effect of thenteraction of both factors was obtained in the right-central spa-ial factor of P1 (F(1,29) = 4.44, p < 0.05). Bonferroni post hoc testsadjusted alpha = 0.05) indicated that right-central P1 amplitudesere greater (more negative: this component is negative at right-
entral areas) for M− than for S−, and a trend was observed (p < 0.1)or M− versus M0 differences (Table 1). The rest of pairwise com-arisons were not significant according to these post hoc contrasts.o main effects were observed (Motion: F(1,29) = 3.43, p = 0.074;motion: F(1,29) = 0.017, p = 0.898).
S− 0.152 (0.20) 878.96 (26.13) 5.53 (0.57)M− −0.162 (0.18) 939.31 (31.47) 4.10 (0.44)S0 −0.011 (0.18) 922.75 (27.66) 5.67 (0.65)M0 0.021 (0.18) 889.61 (30.72) 4.00 (0.47)
a Linearly related to amplitudes. Greater negativity means greater amplitude.
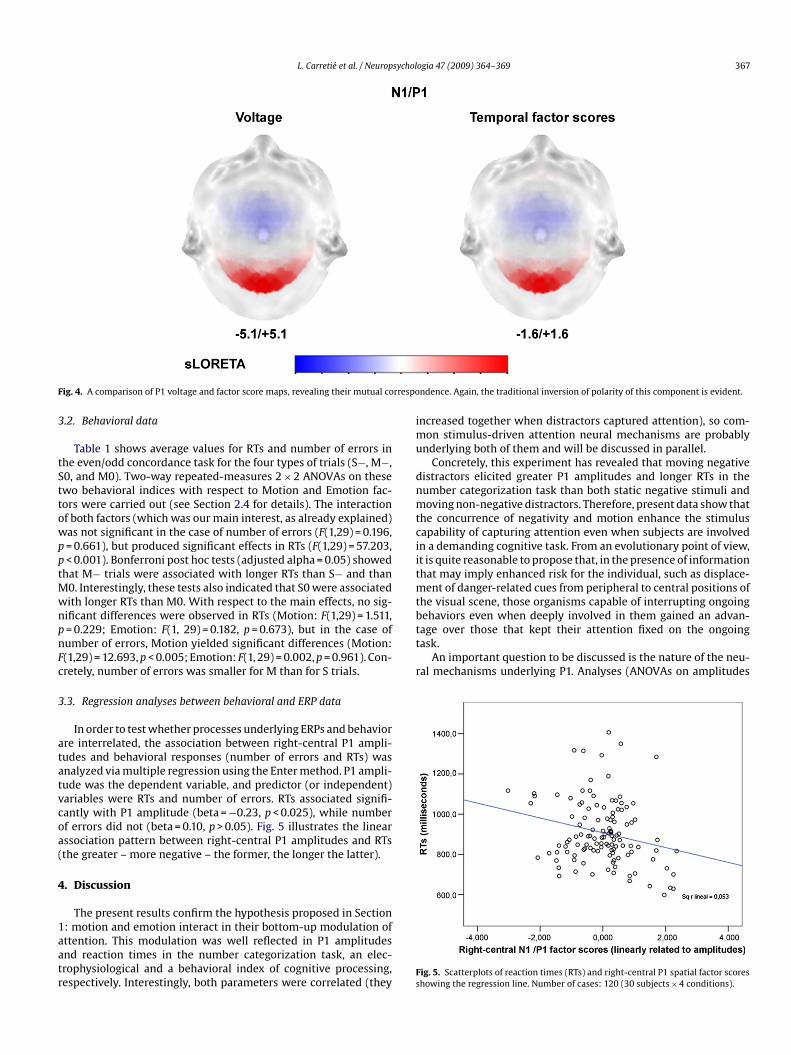
L. Carretié et al. / Neuropsychologia 47 (2009) 364–369 367
F rrespo
3
tSttowpptMwnpnFc
3
atatvcoa(
4
1aatr
imu
dnmtciitmtbehaviors even when deeply involved in them gained an advan-tage over those that kept their attention fixed on the ongoingtask.
An important question to be discussed is the nature of the neu-ral mechanisms underlying P1. Analyses (ANOVAs on amplitudes
ig. 4. A comparison of P1 voltage and factor score maps, revealing their mutual co
.2. Behavioral data
Table 1 shows average values for RTs and number of errors inhe even/odd concordance task for the four types of trials (S−, M−,0, and M0). Two-way repeated-measures 2 × 2 ANOVAs on thesewo behavioral indices with respect to Motion and Emotion fac-ors were carried out (see Section 2.4 for details). The interactionf both factors (which was our main interest, as already explained)as not significant in the case of number of errors (F(1,29) = 0.196,= 0.661), but produced significant effects in RTs (F(1,29) = 57.203,< 0.001). Bonferroni post hoc tests (adjusted alpha = 0.05) showed
hat M− trials were associated with longer RTs than S− and than0. Interestingly, these tests also indicated that S0 were associatedith longer RTs than M0. With respect to the main effects, no sig-ificant differences were observed in RTs (Motion: F(1,29) = 1.511,= 0.229; Emotion: F(1, 29) = 0.182, p = 0.673), but in the case ofumber of errors, Motion yielded significant differences (Motion:(1,29) = 12.693, p < 0.005; Emotion: F(1, 29) = 0.002, p = 0.961). Con-retely, number of errors was smaller for M than for S trials.
.3. Regression analyses between behavioral and ERP data
In order to test whether processes underlying ERPs and behaviorre interrelated, the association between right-central P1 ampli-udes and behavioral responses (number of errors and RTs) wasnalyzed via multiple regression using the Enter method. P1 ampli-ude was the dependent variable, and predictor (or independent)ariables were RTs and number of errors. RTs associated signifi-antly with P1 amplitude (beta = −0.23, p < 0.025), while numberf errors did not (beta = 0.10, p > 0.05). Fig. 5 illustrates the linearssociation pattern between right-central P1 amplitudes and RTsthe greater – more negative – the former, the longer the latter).
. Discussion
The present results confirm the hypothesis proposed in Section
: motion and emotion interact in their bottom-up modulation ofttention. This modulation was well reflected in P1 amplitudesnd reaction times in the number categorization task, an elec-rophysiological and a behavioral index of cognitive processing,espectively. Interestingly, both parameters were correlated (theyFs
ndence. Again, the traditional inversion of polarity of this component is evident.
ncreased together when distractors captured attention), so com-on stimulus-driven attention neural mechanisms are probably
nderlying both of them and will be discussed in parallel.Concretely, this experiment has revealed that moving negative
istractors elicited greater P1 amplitudes and longer RTs in theumber categorization task than both static negative stimuli andoving non-negative distractors. Therefore, present data show that
he concurrence of negativity and motion enhance the stimulusapability of capturing attention even when subjects are involvedn a demanding cognitive task. From an evolutionary point of view,t is quite reasonable to propose that, in the presence of informationhat may imply enhanced risk for the individual, such as displace-
ent of danger-related cues from peripheral to central positions ofhe visual scene, those organisms capable of interrupting ongoing
ig. 5. Scatterplots of reaction times (RTs) and right-central P1 spatial factor scoreshowing the regression line. Number of cases: 120 (30 subjects × 4 conditions).

3 sychol
aetprieteatitw
tviepsartavAlmcvsrDusipD2cip
taTagdidiidedtco1
rar
p“iisshowesrcni
isetiecer
A
0I
R
AA
B
C
C
C
C
C
C
D
D
D
D
68 L. Carretié et al. / Neurop
nd correlation with RTs) suggest that this component reflectsnhanced mobilization of visual processing resources in trials con-aining efficient distractors (moving spiders and cockroaches, in theresent experiment). As explained in Section 1, P1 has already beenelated to visual attentional capture processes in previous stud-es (see a review in Hopfinger & Mangun, 2001; see also Carretiét al., 2004), and its amplitude increases as this type of atten-ion increases. However, an open question is whether P1 reflectsnhanced attention towards efficient distractors whenever theyppear or whether it reflects increased mobilization of resourcesowards the ongoing task (digit categorization, in this experiment)n order to ‘compensate’ the disruptive effect of efficient distrac-ors. Further research is needed to respond this relevant question,hich may be pertinent in other studies on attentional capture.
Whatever the precise mechanism underlying P1 is, it appearso be clear that it reflects attentional processes. However, a pre-ious, “preattentive” (Öhman, Flykt, & Esteves, 2001) mechanisms necessary to evaluate whether the distractor is potentially rel-vant (e.g., harmful or dangerous) and deserves attention. Thisreattentive mechanism appears no to be reflected in ERPs, but thehort latency of ‘attentional’ P1 (it peaks at 112 ms) provides somedditional clues on its nature. Obviously, it should involve emotion-elated structures of the brain, and these structures should be ableo increase the activity of visual cortex when cues signaling dangerppear in the visual scene. How is the nervous system able to pro-ide motion information to emotion-related structures so rapidly?key element in the answer to this question is the magnocellu-
ar visual pathway. As compared to the parvocellular system, theagnocellular pathway has fewer neurons – which therefore each
over wider areas of the visual scene – and it rapidly distributesisual information (which is, however, of low-quality—e.g., highpatial frequencies are lacking) to subcortical emotion-related, fast-esponding structures such as the amygdala (Vuilleumier, Armony,river, & Dolan, 2003). Interestingly, the amygdala is able to mod-late visual cortex activity (e.g., Adolphs, 2004). Previous studiesuggest that the magnocellular pathway is more involved in thenitial, rapid processing of emotional stimuli than the parvocellularathway (Carretié, Hinojosa, López-Martín, & Tapia, 2007; Pourtois,an, Grandjean, Sander, & Vuilleumier, 2005; Vuilleumier et al.,003). The magnocellular pathway is deeply involved in the pro-essing of motion (e.g., Chapman, Hoag, & Giaschi, 2004); in fact,t provides the major input to cortical areas responsible for motionrocessing (DeYoe & Van Essen, 1988; Maunsell & Newsome, 1987).
Two additional findings are worth to be discussed. On one hand,his study has shown that, separately, motion and emotional neg-tivity may not produce significant effects in bottom-up attention.hus, dynamic peripheral distractors (negative and non-negatives a whole) did not elicit longer RTs, higher number of errors, norreater P1 amplitudes, than static distractors. Similarly, negativeistractors (dynamic and static as a whole) did not show these
ndices of attentional capture to a greater extent than non-negativeistractors. As explained in Section 1, both motion and negativ-
ty do not capture attention under certain circumstances, and thiss apparently the case in the present study. Interestingly, dynamicistractors (regardless of their emotional content) yielded fewerrrors (but not shorter RTs nor greater P1 amplitudes) than staticistractors. A possible explanation is that motion of irrelevant itemsowards the relevant, central items may have served as a ‘movingue’ that signaled the target. The fact that cues facilitate detectionf targets is very well known in attention research (Posner & Cohen,
984).On the other hand, non-negative static trials elicited longereaction times (but not greater number of errors nor greater P1mplitudes) than non-negative moving trials in the digit catego-ization task. Two ideas provide a possible explanation to this
F
F
ogia 47 (2009) 364–369
henomenon. First, “non-negative” does not necessarily meanneutral”. Butterflies, for example, are assessed as emotionally pos-tive by many individuals (Lang, Bradley, & Cuthbert, 2005). If its the case in the current sample, the fact that positive emotionaltimuli have been reported to be efficient capturers of attentionhould be taken into account (Rothermund, 2001), though theyave been much less studied than negative at this respect. Sec-nd, while threats probably increase their danger or harmful valuehen they move, motion in non-negative cues (clouds, leaves, birds,
tc.) is not a characteristic conveying dramatic information or con-equences. In truth, static pleasant stimuli would be more easilyecognized than dynamic pleasant stimuli, so they would be moreapable of capturing attention. This tentative explanation on RTs toon-negative trials is worth to be further explored through exper-
ments designed ad hoc to test this issue.In sum, the main conclusion that derives from electrophysiolog-
cal and behavioral results suggest that motion provides additionalalience to threatening stimuli that facilitates attentional captureven when the individual is engaged in a demanding task. An addi-ional idea that can be extracted from this study concerns thenterest of reproducing natural conditions as far as possible in thexperimental research of emotion. Particularly, motion appears toonstitute an important ingredient in many affective events forliciting full and intense psychological reactions, such as attention-elated processes.
cknowledgements
This work was supported by the grants SEJ2005-08461-C02-1/02/PSIC and PSI2008-03688 from the Ministerio de Ciencia ennovación of Spain.
eferences
dolphs, R. (2004). Emotional vision. Nature Neuroscience, 7, 1167–1168.nderson, A. J. (2003). Humans deceived by predatory stealth strategy camouflaging
motion. Proceedings: Biological Sciences, 270, 18–20.uchel, C., Josephs, O., Rees, G., Turner, R., Frith, C. D., & Friston, K. J. (1998). The
functional anatomy of attention to visual motion. A functional MRI study. Brain,121, 1281–1294.
arretié, L., Hinojosa, J. A., López-Martín, S., & Tapia, M. (2007). An electrophysiolog-ical study on the interaction between emotional content and spatial frequencyof visual stimuli. Neuropsychologia, 45, 1187–1195.
arretié, L., Hinojosa, J. A., Martín-Loeches, M., Mercado, F., & Tapia, M. (2004). Auto-matic attention to emotional stimuli: Neural correlates. Human Brain Mapping,22, 290–299.
hapman, C., Hoag, R., & Giaschi, D. (2004). The effect of disrupting the humanmagnocellular pathway on global motion perception. Vision Research, 44, 2551–2557.
hapman, R. M., & McCrary, J. W. (1995). EP component identification and measure-ment by principal components analysis. Brain and Cognition, 27, 288–310.
liff, N. (1987). Analyzing multivariate data. San Diego, Calif: Harcourt BraceJovanovich.
oles, M. G. H., Gratton, G., Kramer, A. F., & Miller, G. A. (1986). Principles of signalacquisition and analysis. In M. G. H. Coles, E. Donchin, & S. W. Porges (Eds.),Psychophysiology: Systems, processes and applications (pp. 183–221). Amsterdam:Elsevier.
eYoe, E. A., & Van Essen, D. C. (1988). Concurrent processing streams in monkeyvisual cortex. Trends in Neurosciences, 11, 219–226.
ien, J., Beal, D. J., & Berg, P. (2005). Optimizing principal components analysis ofevent-related potentials: Matrix type, factor loading weighting, extraction, androtations. Clinical Neurophysiology, 116, 1808–1825.
oallo, S., Holguin, S. R., & Cadaveira, F. (2006). Attentional load affects automaticemotional processing: Evidence from event-related potentials. Neuroreport, 17,1797–1801.
onchin, E., & Heffley, E. F. (1978). Multivariate analysis of event-related potentialdata: A tutorial review. In D. Otto (Ed.), Multidisciplinary perspectives in eventrelated brain potential research (pp. 555–572). Washington, DC: USA: Govern-
ment Printing Office.ranconeri, S. L., & Simons, D. J. (2003). Moving and looming stimuli capture atten-tion. Perception & Psychophysics, 65, 999–1010.
ranconeri, S. L., & Simons, D. J. (2005). The dynamic events that capture visualattention: A reply to abrams and christ (2005). Perception & Psychophysics, 67,962–966.

sychol
H
L
M
M
Ö
P
P
P
P
P
P
R
R
R
S
S
V
V
Neuron, 30, 829–841.
L. Carretié et al. / Neurop
opfinger, J. B., & Mangun, G. R. (2001). Electrophysiological studies of reflexiveattention. In C. L. Folk & B. S. Gibson (Eds.), Attraction, distraction and action:Multiple perspectives on attentional capture (pp. 3–26). Amsterdam: Elsevier.
ang, P. J., Bradley, M. M., & Cuthbert, B. N. (2005). International affective picturesystem (IAPS): Affective ratings of pictures and instruction manual. Gainesville, FL:University of Florida.
aunsell, J. H., & Newsome, W. T. (1987). Visual processing in monkey extrastriatecortex. Annual Review of Neuroscience, 10, 363–401.
cLeod, P., Driver, J., & Crisp, J. (1988). Visual search for a conjunction of movementand form is parallel. Nature, 332, 154–155.
hman, A., Flykt, A., & Esteves, F. (2001). Emotion drives attention: Detecting thesnake in the grass. Journal of Experimental Psychology General, 130, 466–478.
essoa, L., Kastner, S., & Ungerleider, L. G. (2002). Attentional control of the process-ing of neutral and emotional stimuli. Cognitive Brain Research, 15, 31–45.
into, A., Jankowski, M., & Sesack, S. R. (2003). Projections from the paraventricularnucleus of the thalamus to the rat prefrontal cortex and nucleus accumbensshell: Ultrastructural characteristics and spatial relationships with dopamineafferents. The Journal of Comparative Neurology, 459, 142–155.
into, Y., Olivers, C. N., & Theeuwes, J. (2006). When is search for a static targetamong dynamic distractors efficient? Journal of Experimental Psychology. HumanPerception and Performance, 32, 59–72.
osner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma &
D. Bouwhuis (Eds.), Attention and performance (pp. 531–556). Hillsdale, NJ: Erl-baum.ourtois, G., Dan, E. S., Grandjean, D., Sander, D., & Vuilleumier, P. (2005). Enhancedextrastriate visual response to bandpass spatial frequency filtered fearful faces:Time course and topographic evoked-potentials mapping. Human Brain Map-ping, 26, 65–79.
V
W
ogia 47 (2009) 364–369 369
ratto, F., & John, O. P. (1991). Automatic vigilance: The attention-grabbing powerof negative social information. Journal of Personality and Social Psychology, 61,380–391.
ees, G. (1997). Modulating irrelevant motion perception by varying attentional loadin an unrelated task. Science, 278, 1616–1619.
ees, G., Frith, C. D., & Lavie, N. (2001). Perception of irrelevant visual motion duringperformance of an auditory task. Neuropsychologia, 39, 937–949.
othermund, K. (2001). Automatic attention to stimuli signalling chances and dan-gers: Moderating effects of positive and negative goal and action contexts.Cognition & Emotion, 15, 231–248.
chwartz, S., Vuilleumier, P., Hutton, C., Maravita, A., Dolan, R. J., & Driver, J. (2005).Attentional load and sensory competition in human vision: Modulation of fMRIresponses by load at fixation during task-irrelevant stimulation in the peripheralvisual field. Cerebral Cortex, 15, 770–786.
iegle, G. J., Ingram, R. E., & Matt, G. E. (2002). Affective interference: An explanationfor negative attention biases in dysphoria? Cognitive Therapy and Research, 26,73–87.
on Mühlenen, A., Rempel, M. I., & Enns, J. T. (2005). Unique temporal change is thekey to attentional capture. Psychological Science, 16, 979–986.
uilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J. (2001). Effects of attention andemotion on face processing in the human brain an event-related fMRI study.
uilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J. (2003). Distinct spatial fre-quency sensitivities for processing faces and emotional expressions. NatureNeuroscience, 6, 624.
illiams, J. M. G., Mathews, A., & MacLeod, C. (1996). The emotional stroop task andpsychopathology. Psychological Bulletin, 120, 3–24.




















