d-Glucose releases 5-hydroxytryptamine from human bon cells as a model of enterochromaffin cells
7
GASTROENTEROLOGY2001;121:1400-1406 D-Glucose Releases 5-Hydroxytryptamine From Human BON Cells as a Model of Enterochromaffin Cells MINSOO KIM,* HELEN J. COOKE,t NAJMA H. JAVED,§ HANNAH V. CAREY,qt FIEVOS CHRISTOFI,¶ and HELEN E. RAYBOULD # Departments of *Pharmacology, tNeuroscience, and ¶Anesthesiology, The Ohio State University, Columbus, Ohio; §Department of Physiology and Health Science, Ball State University, Muncie, Indiana; IIDepartment of Comparative Biosciences, University of Wisconsin-Madison, Madison, Wisconsin; and #Department of Anatomy, Physiology, and Cell Biology, University of California, Davis, California Background&Aims: 5-Hydroxytryptamine (5-HT) is re- leased from enterochromaffin cells and activates neural reflex programs regulating motility and secretion. Al- though sugars are reported to release 5-HT in vivo, it is unclear whether they act directly on enterochromaffin cells or indirectly through an intermediary messenger. The aim was to determine if D-glucose is a stimulus for 5-HT release. Methods: Human BON cells, derived from enterochromaffin cells, were treated with D-glucose, ga- lactose, and the nonmetabolizable methyl oL-D-glucopy- ranoside, or with fructose. Results: Reverse-transcription polymerase chain reaction together with Western blot analysis revealed an SGLT-like protein. D-Glucose caused a concentration-dependent increase in 5-HT release, which was mimicked by methyl O~-D-glucopyranoside and galactose but not fructose. D-Glucose-stimulated 5-HT release was significantly reduced by phloridzin. Concen- trations of mannitol below 75 mmol/L were ineffective in releasing 5-HT. Brefeldin A abolished forskolin-stimu- lated 5-HT release without affecting basal or constitutive release. Conclusions: The results show that high concen- trations of metabolizable and nonmetabolizable hex- oses activate signal transduction pathways, leading to release of 5-HT. These findings imply a role for entero- chromaffin cells as "glucose sensors" during ingestion of a meal. I ngestion of a meal slows gastric emptying and secre- tion while stimulating pancreatic and biliary secre- tion. 1 These processes depend on transduction of chemical and mechanical signals from the duodenum, processing and integration in the central nervous system, and adjustments in efferent neural outflow to prevent gastric intake and emptying from overloading of the digestive and absorptive capacity of the intestine. 1 One of the chemical signals during a meal is D-glucose, which can reach highs of 300 mmol/L during digestion of carbohydrates2 Previous stud- ies have shown that intestinal D-glucose inhibits gastric emptying and stimulates pancreatic secretion by activation of extrinsic vagal and spinal afferent pathways containing 5-hydroxytryptamine 3 (5-HT3) receptors. 36 Although glucose is undoubtedly a signaling molecule, it is unclear which cells are the sensors. Intrinsic primary afferents in the enteric plexus and extrinsic primary afferents are excited by D-glucose. 7-9 Because the mucosal epithelium separates the nerve end- ings from the luminal environment, direct exposure of the neurons to nutrients is questionable. This fact raises the concept that an intermediate mediator may be re- leased from cells whose apical membranes are exposed to the luminal milieu containing glucose. In support of this "taste cell" concept are studies showing that glucose infusion into the small intestine releases gastric inhibi- tory polypeptide (GIP), 7,~ cholecystokinin (CCK), 9 and 5-HT3 ° Because release of GIP and CCK was inhibited by phloridzin, it was suggested that sodium-D-glucose co- transporter 1 (SGLT1) might be found in endocrine cells. 11 Given the reported involvement of 5-HT 3 receptors in nutrient-evoked adjustments in gastric emptying, it is possible that 5-HT is also an important paracrine medi- ator of afferent neuronal signals from the intestine to the brain. In this study, we examined the ability of extracellular hexoses to induce release of 5-HT from BON cells, a model for human enterochromaffin ceils. The results suggest that BON cells of enterochromaffin origin ex- press an SGLT-like protein and release 5-HT in response to D-glucose and galactose as well as to the nonmetabo- lizable hexose methyl O~-D-glucopyranoside. Abbreviations used in this paper: AEBSF, 4-(2-aminoethyl)benzene- sulfonylfluoride-HCI;CCK,cholecystokinin;EBBS,Earle balanced buffer solution; GIP, gastric inhibitory polypeptide;GLUT,glucose transporter; 5-HT, 5-hydroxytryptamine; e~MG,(~-D-glucopyranoside; SGLT1, sodium- D-glucosecotransporter1. © 2001 by the American Gastroenterological Association 0016-5085/01/$35.00 doi:10.1053/gast.2001.29567
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of d-Glucose releases 5-hydroxytryptamine from human bon cells as a model of enterochromaffin cells

GASTROENTEROLOGY 2001;121:1400-1406
D-Glucose Releases 5-Hydroxytryptamine From Human BON Cells as a Model of Enterochromaffin Cells
MINSOO K I M , * HELEN J. COOKE, t NAJMA H. JAVED,§ HANNAH V. CAREY,qt FIEVOS CHRISTOFI,¶
and HELEN E. RAYBOULD # Departments of *Pharmacology, tNeuroscience, and ¶Anesthesiology, The Ohio State University, Columbus, Ohio; §Department of Physiology and Health Science, Ball State University, Muncie, Indiana; IIDepartment of Comparative Biosciences, University of Wisconsin-Madison, Madison, Wisconsin; and #Department of Anatomy, Physiology, and Cell Biology, University of California, Davis, California
Background&Aims: 5-Hydroxytryptamine (5-HT) is re- leased from enterochromaffin cells and activates neural reflex programs regulating motility and secretion. Al- though sugars are reported to release 5-HT in vivo, it is unclear whether they act directly on enterochromaffin cells or indirectly through an intermediary messenger. The aim was to determine if D-glucose is a stimulus for 5-HT release. Methods: Human BON cells, derived from enterochromaffin cells, were treated with D-glucose, ga- lactose, and the nonmetabolizable methyl oL-D-glucopy- ranoside, or with fructose. Results: Reverse-transcription polymerase chain reaction together with Western blot analysis revealed an SGLT-like protein. D-Glucose caused a concentration-dependent increase in 5-HT release, which was mimicked by methyl O~-D-glucopyranoside and galactose but not fructose. D-Glucose-stimulated 5-HT release was significantly reduced by phloridzin. Concen- trations of mannitol below 75 mmol /L were ineffective in releasing 5-HT. Brefeldin A abolished forskolin-stimu- lated 5-HT release without affecting basal or constitutive release. Conclusions: The results show that high concen- trations of metabolizable and nonmetabolizable hex- oses activate signal transduction pathways, leading to release of 5-HT. These findings imply a role for entero- chromaffin cells as "glucose sensors" during ingestion of a meal.
I ngestion of a meal slows gastric emptying and secre- tion while stimulating pancreatic and biliary secre-
tion. 1 These processes depend on transduction of chemical
and mechanical signals from the duodenum, processing and integration in the central nervous system, and adjustments
in efferent neural outflow to prevent gastric intake and emptying from overloading of the digestive and absorptive
capacity of the intestine. 1 One of the chemical signals
during a meal is D-glucose, which can reach highs of 300 mmol/L during digestion of carbohydrates2 Previous stud-
ies have shown that intestinal D-glucose inhibits gastric emptying and stimulates pancreatic secretion by activation
of extrinsic vagal and spinal afferent pathways containing
5-hydroxytryptamine 3 (5-HT3) receptors. 36 Although
glucose is undoubtedly a signaling molecule, it is unclear
which cells are the sensors. Intrinsic primary afferents in the enteric plexus and
extrinsic primary afferents are excited by D-glucose. 7-9
Because the mucosal epithelium separates the nerve end-
ings from the luminal environment, direct exposure of
the neurons to nutrients is questionable. This fact raises
the concept that an intermediate mediator may be re-
leased from cells whose apical membranes are exposed to
the luminal milieu containing glucose. In support of this
"taste cell" concept are studies showing that glucose
infusion into the small intestine releases gastric inhibi-
tory polypeptide (GIP), 7,~ cholecystokinin (CCK), 9 and
5-HT3 ° Because release of GIP and CCK was inhibited by
phloridzin, it was suggested that sodium-D-glucose co-
transporter 1 (SGLT1) might be found in endocrine cells. 11
Given the reported involvement of 5-HT 3 receptors in
nutrient-evoked adjustments in gastric emptying, it is
possible that 5-HT is also an important paracrine medi-
ator of afferent neuronal signals from the intestine to the
brain. In this study, we examined the ability of extracellular
hexoses to induce release of 5-HT from BON cells, a
model for human enterochromaffin ceils. The results
suggest that BON cells of enterochromaffin origin ex-
press an SGLT-like protein and release 5-HT in response
to D-glucose and galactose as well as to the nonmetabo-
lizable hexose methyl O~-D-glucopyranoside.
Abbreviations used in this paper: AEBSF, 4-(2-aminoethyl)benzene- sulfonylfluoride-HCI; CCK, cholecystokinin; EBBS, Earle balanced buffer solution; GIP, gastric inhibitory polypeptide; GLUT, glucose transporter; 5-HT, 5-hydroxytryptamine; e~MG, (~-D-glucopyranoside; SGLT1, sodium- D-glucose cotransporter 1.
© 2001 by the American Gastroenterological Association 0016-5085/01/$35.00
doi:10.1053/gast.2001.29567

December 2001 D-GLUCOSE AND 5-HT RELEASE 1401
M e t h o d s
Culture of Human Carcinoid Tumor Cell Line
Carcinoid tumors are endocrine neoplasms derived from enterochromaffin cells. They secrete vasoactive substances and biogenic amines and cause carcinoid syndrome character- ized by flushing, diarrhea, bronchoconstriction, and other symptoms. 12 BON cells were obtained from C. M. Townsend Jr. (University of Texas, Galveston, TX). An immortalized BON cell line was derived from a lymph node metastasis of a carcinoid tumor originating from the pancreas. BON is a noncloned cell line that is a mixed population of large round cells and small ceils with dendritic-like extensions and prop- erties characteristic of foregut and midgut carcinoid tumors. They form monoiayers and are not contact inhibited. 13 They test positive for ePithelial antigens and not for neuronal or glial antigens. 13 The cells have been reported to release pan- creastatin , neurotensin, and 5-HT but not insulin, glucagon, or somat0statin. 13 Carcinoid tumor cells have apical microvilli and a supernuclear Golgi apparatus that suggest they are polarized cells. ~4
Cells (BON0) were subcloned, and clone 7 (BONy) was chosen on the basis of having the highest content of 5-HT released by the calcium ionophore A23187 (10 b~mol/L). Monolayers of BON ceils were maintained in Dulbecco mod- ified Eagle medium-nutr ient mixture F- 12 (DMEM/F 12; 1 : 1), supplemented with 10% fetal calf serum, 100 IU/mL penicil- lin, and 100 b~g/mL streptomycin (Sigma Chemical Co., St. Louis, MO) in a humidified atmosphere of 95% air and 5% CO2 at 37°C. Cells were trypsinized and seeded at a density of 1 × 106 cells per well in 12-well culture plates (Costar, Cambridge, MA). Cells were grown for 48 hours to achieve confluence before experiments. Passage numbers of cells used in the present study were 10-30.
Treatment of Cells for 5-HT Release
Cells were washed once and preincubated with 0.5 mL Earle balanced buffer solution (EBBS; pH 7.3) containing 0.1% bovine serum albumin. After preincubation with EBBS at 37°C for 30 minutes, medium was removed and cells were washed twice and incubated in 0.5 mL of EBBS containing 1 × 10 -5 mol/L alaproclate (5-HT uptake inhibitor) and pargyline (monoamine oxidase inhibitor). After 20 minutes at 37°C, the incubation medium was removed and centrifuged for 30 seconds in a microcentrifuge to remove any nonadherent cells. Supernatants were stored at - 8 0 ° C until assay.
Enzyme Immunoassay to Detect 5-HT
Basal and stimulated 5-HT release were measured by enzyme immunoassay using a kit (Beckman Coulter Inc., Brea, CA). Assays were performed according to the manufacturer's instructions. The absorbance was measured at 405 nm, and 5-HT concentration was determined from a standard curve. The sensitivity of the assay is approximately 0.5 nmol/L (1.8 pg/well).
Reverse-Transcription Polymerase Chain Reaction
Total RNA was prepared from BON cells using a guanidium thiocyanate and phenol procedure. Cells were lysed in 4 mol/L guanidium thiocyanate denaturing solution with 2 mol/L sodium acetate buffer (pH 4) followed by phenol, chlo- roform extraction, and precipitation with isoproterenol. After the pellet was washed, it was resuspended in deionized water. Aliquots of each sample were run on a formaldehyde denatur- ing gel to check for quality of RNA. RNA (2 I~g) was primed with oligo (dT) and reverse transcribed with avian myeloblas- tosis virus reverse transcriptase (Promega, Madison, WI). The resulting complementary DNA (cDNA) was amplified by PCR in the presence of polymerase (Perkin-Elmer Corp., Branch- burg, NJ) and 1 I~g of forward (5 '-TCT TCG ATT ACA TCC AGT CCA-3') .5 and reverse (5 ' -GAT AAT CGT G G G ACA GTT GCT-3') 16 primers. The primers encompassed a 208- nucleotide band from 1365 to 1573 base pairs of the human SGLT1 (GenBank M24847; National Center for Biotechnol- ogy, Bethesda, MD). Each reaction cycle consisted of incuba- tions at 95°C for 1 minute, 60°C for 30 seconds, and 72°C for 1 minute. The amplified DNA bands were separated by elec- trophoresis on a 3% agarose gel. The specific bands were excised, purified, and sequenced.
Antisera
Rabbit anti-SGLT1. The antigenic epitope for SGLT1 antiserum is a peptide corresponding to amino acids 402-420 of the putative extracellular loop of SGLT1 of rabbit small intestine (lot 20070618; Chemicon International, Inc., Te- mecula, CA). It cross-reacts to rat, mouse, human, rabbit, porcine, feline, and canine tissues. The SGLT1 control peptide (amino acids 402-420; Chemicon International, Inc.) was modified by addition of cysteine at the amino terminus for coupling to keyhole l impet hemocyanin.
Rabbit anti-GLUT2. The immunogenic site was a peptide encompassing amino acids 40 -55 corresponding to the exoplasmic loop between helices 1 and 2 of the liver glucose transporter coupled to keyhole limpet hemocyanin (lot 20060426, Chemicon International, Inc.). It recognizes GLUT2 in mouse, rat, and human tissues.
Rabbit anti-GLUT5. The immunogenic site was a peptide specific for the carboxy terminus of human GLUT5 sequence (ELKELPPVTSEQ) conjugated to keyhole limpet hemocyanin (lot 19102127; Chemicon International, Inc.). It reacts only with human GLUT5 and does not bind to GLUTs 1-4.
Western Blotting
BON cells. The frozen BON cells were thawed and scraped from culture plates. After centrifugation (3000g for 5 minutes at 4°C), pellets were resuspended in lysis buffer containing 10 mrnol/L Tris-HCl, pH 8.0, ~% Triton X-100, 60 mmol/L octyl glucose, 150 mmol/L NaC1, 5 mmol/L EDTA, 500 I~mol/L 4-(2-aminoethyl)benzenesulfonylfluoride-
1402 KIM El" AL. GASTROENTEROLOGY Vol. 121, No. 6
HCI (AEBSF), 150 nmol/L aprotinin, t b~mol/L E-64, and 1 b~mol/L leupeptin. After incubation for 30 minutes on ice, the tubes were centrifuged at 10,000g for 3 minutes at 4°C. The pellets were discarded. Protein concentrations of the superna- tants were determined (Bio-Rad, Hercules, CA). Solubilized proteins (10-20 b~g) were separated by 10%-12.5% denatur- ing sodium dodecyl sulfate-polyacrylamide gel and transferred to nitrocellulose membranes.
Human jejunal m u c o s a . Tissue was obtained from clinically obese individuals undergoing Roux-n-Y bypass sur- gery. Protocols were in accordance with The Ohio State Uni- versity Human Subjects Review Board. Tissue was pulverized in liquid nitrogen and lysed in 10 mmol/L HEPES (pH 7.9), 1,5 mmol/L MgCl2, 10 mmol/L KC1, 500 t*mol/L AEBSF, 150 nmol/L aprotinin, 1 btmol/L E-64, and 1 b~mol/L leupep- tin. After 20 homogenizing strokes and another 30-minute incubation on ice, samples were centrifuged at 8000g for 3 minutes at 4°C. The pellets were discarded, and a protein assay was done on the supernatants.
Antibodies used include SGLT1, GLUT2, and GLUT5. For irnmunoneutralization, antibodies were combined with a 5-fold (by weight) excess of blocking peptide and incubated for 2 hours at room temperature. Nitrocellulose membranes were blocked with 5% nonfat milk for 2 hours at 37°C and probed with primary antibodies to SGLTI (1:4000), GLUT2 (1:2000), or GLUT5 (1:1000) for 1 hour at 37°C. Membranes were rinsed with phosphate-buffered saline/Tween 20 (80 mmol/L Na2HPO4 " 7H20, 20 mmol/L KHePO4, 100 mmol/L NaCl, 1% [vol/vol] Tween 20, pH 7.5) and incubated with peroxidase-conjugated anti-rabbit immunoglobulin G at 1:3000 dilution for 30 minutes at room temperature. Immunopositive bands were visualized with the ECL kit (Amersham, Piscat- away, N J).
S t a t i s t i c s . All data were expressed as means -+ SEM. Statistical significance of the data was evaluated by the paired or unpaired Student t test.
Results
Effect of Brefeldin A on Basal (Constitutive) and Forskolin-Evoked 5-HT Release
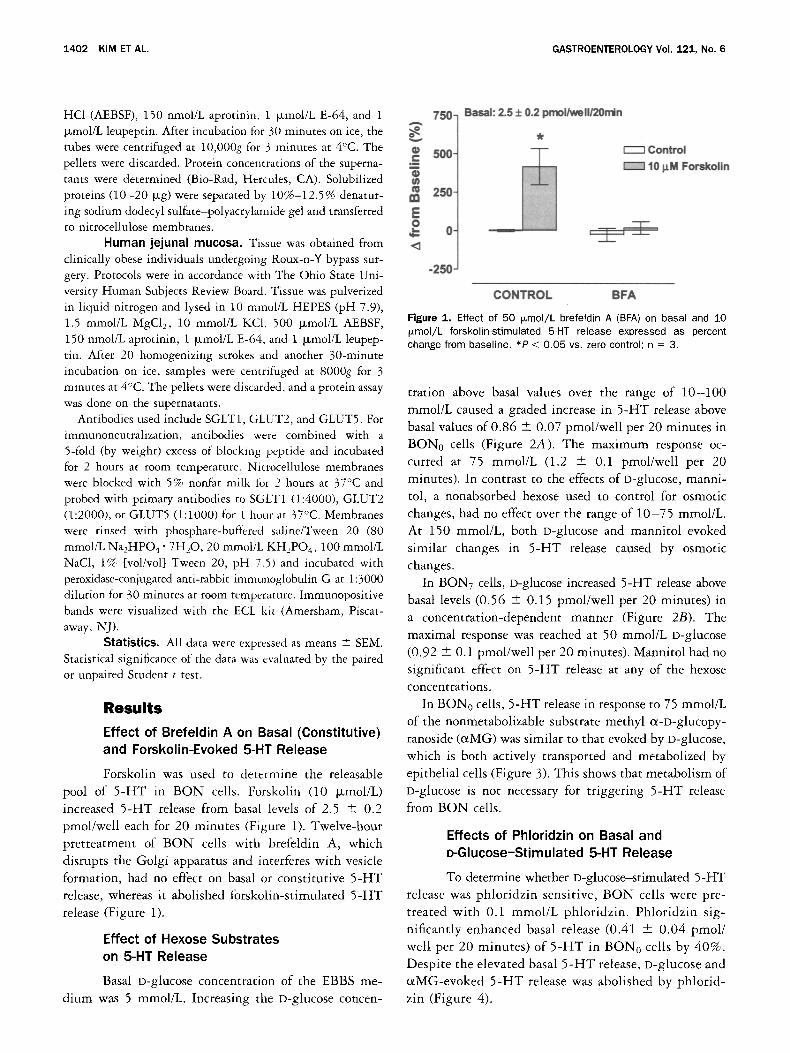
Forskolin was used to determine the releasable pool of 5 -HT in BON cells. Forskolin (10 ~zmol/L)
increased 5 -HT release from basal levels of 2.5 -+ 0.2 pmol/well each for 20 minutes (Figure 1). Twelve-hour pretreatment of B O N cells with brefeldin A, which disrupts the Golgi apparatus and interferes with vesicle
formation, had no effect on basal or constitutive 5 -HT release, whereas it abolished forskolin-stimulated 5 -HT release (Figure 1).
Effect of Hexose Substrates on 5-HT Release
Basal D-glucose concentration of the EBBS me- dium was 5 mmol/L. Increasing the D-glucose concen-
750- g • 500"
Q ~q eu 250-
-250
Basal: 2.5 + 0,2 pmol/well/2Omin
Control 10 pM Forskolin
CONTROL BFA
Figure 1. Effect of 50 Fmol/L brefeldin A (BFA) on basal and 10 Fmol/L forskolin-stimulated 5-HT release expressed as percent change from baseline. *P < 0.05 vs. zero control; n = 3.
tration above basal values over the range of 10-100 mmol/L caused a graded increase in 5 -HT release above
basal values of 0.86 + 0.07 pmol/well per 20 minutes in BON0 cells (Figure 2A). The maximum response oc- curred at 75 mmol/L (1.2 + 0.1 prnol/well per 20 minutes). In contrast to the effects of D-glucose, manni- tol, a nonabsorbed hexose used to control for osmotic changes, had no effect over the range of 10-75 mmol/L.
At 150 mmol/L, both D-glucose and mannitol evoked similar changes in 5 -HT release caused by osmotic changes.
In BONy cells, D-glucose increased 5-HT release above basal levels (0.56 +- 0.15 pmol/well per 20 minutes) in a concentration-dependent manner (Figure 2B). The
maximal response was reached at 50 mmol/L D-glucose (0.92 --+ 0.1 pmol/well per 20 minutes). Mannitol had no significant effect on 5 -HT release at any of the hexose concentrations.
In BON0 cells, 5 -HT release in response to 75 mrnol/L of the nonmetabolizable substrate methyl OL-D-glucopy- ranoside (otMG) was similar to that evoked by D-glucose, which is both actively transported and metabolized by
epithelial cells (Figure 3). This shows that metabolism of D-glucose is not necessary for triggering 5-HT release from BON cells.
Effects of Phloridzin on Basal and D-Glucose-Stimulated 5-HT Release
To determine whether D-glucose-stimulated 5-HT release was phloridzin sensitive, B O N cells were pre-
treated with 0.1 mmol /L phloridzin. Phloridzin sig- nificantly enhanced basal release (0.41 + 0.04 pmol /
well per 20 minutes) of 5 - H T in BON0 cells by 40%. Despite the elevated basal 5 - H T release, D-glucose and otMG-evoked 5 - H T release was abolished by phlorid- zin (Figure 4).

December 2001 D-GLUCOSE AND 5-HT RELEASE 1403
A 150-
A
v 100- @ e- Q w m 5 0 . m
E o
<l 0"
-50
glucose
i - - - - I mannitol
9r
10 25 50 75 150 [mM]
B 150-
A
• 100-
< 0"
-50
glucose
r---I mannltol
-z---E-
~t
10 25 50 75 [mM]
Figure 2. Effect of increasing concentrations of D-glucose and man- nitol on 5-HT release expressed as percent change from baseline. (A) Noncloned BON cells. Basal 5-HT release was 0.86 -- 0.07 pmol/well per 20 minutes; n = 5. (B) Subcloned BON cells. Basal 5-HT release was 0.56 -+ 0.15 pmol/well pe r20 minutes; n = 3 -4 . * P < 0,05 vs. zero control.
Effect of Galactose and Fructose on 5-HT Release
In these experiments, 50 mmol/L galactose, which is a substrate for SGLT1 but not for SGLT2, and 50 mmol/L fructose, which is a substrate for GLUT5 and GLUT2 in epithelial ceils, were used to stimulate 5-HT release. In BON7 ceils, fructose did not stimulate 5-HT release above basal values of 1.2 --- 0.16 pmol/well per 20 minutes, whereas D-glucose and galactose significantly increased release of 5-HT (Figure 5).
Gene and Protein Expression
Reverse-transcription polymerase chain reaction using specific primers based on human SGLT1 cDNA sequence and subsequent gel electrophoresis showed a
125"
A
100- Q c
• - 75- m
o m m 50- m E O 25- 4= <1
O-
-25
i @ T
"k
1 GLU a M G MAN ISO
Figure 3. Comparison of nonmetabolizable (xMG with other stimu- lants of 5-HT release. GLU, glucose; MAN, mannitol; ISO, isoprotere- nol. 5-HT release is expressed as percent change from baseline. BONo cells were treated with 75 mmol/L ~MG, GLU, or MAN, and 0.1 mmol/L ISO was used as a positive control. Baseline 5-HT release was 0.73 _+ 0.06 pmol/well per 20 minutes; n = 7-9. * P < 0.05 vs. zero controls.
208-base pair (bp) band in BON0 cells and in HEK293 cells (Figure 6). Water controls lacked bands. The se- quence of the purified 208-bp band, which encompassed nucleotides 1365-1573, was identical to the comparable sequence of SGLT-1 reported for humans, iv
Western blotting with anti-SGLT1 antibody revealed a prominent protein band of 55-60 kilodaltons (Figure
100- ~ vehicle p h l o r i d z i n
A
75- 4)
• - 50- m
4)
m 25- m
E O 0- 4=
<~ -25-
-50
"k
CON GLU ~ M G
Figure 4. Effect of 0.1 mmol/L phloridzin on 75 mmol/L glucose (GLU)- and 75 mmol/L c~MG-stimulated 5-HT release expressed as percent change from baseline. Basal.5-HT release was 0.41 - 0.04 pmol/well per 20 minutes; n = 4 -11 . *P < 0.05 vehicle vs. phlorid- zin.

1404 KIM ET AL. GASTROENTEROLOGY Vol. 121, No. 6
A
Q ¢:
IBm i
Q W m
E o ,P.
75-
0-
25-
.
-25
"k
io 'A'
GLU GAL FRU Figure 5. Comparison of the effects of 50 mmol/L D-glucose (GLU), 50 mmol/L galactose (GAL), and 50 mmol/L fructose (FRU) on 5-HT release from BON7 cells, expressed as percent change from baseline. Basal 5-HT release was 1.2 _+ 0.3_6 pmoi/well per 20 minutes; n - 4 -10 . *P < 0.05.
7A) in both human jejunal lysates and BON cell lysates. A very faint band was detected in both at 66 kilodaltons.
Immunoneutralization with the blocking peptide pre- vented appearance of the 55-60-ki lodal ton bands but did not eliminate faint bands outside of that size range that were present in some samples.
Western blotting for GLUT2 and GLUT5 indicated the presence of bands of 47 -50 kilodaltons and 45 kilo-
208 bp
Figure 6. cDNA amplified by polymerase chain reaction. Reverse- transcribed cDNA from HEK293 cells was a positive control (HEK). For negative control, water was substituted for the cDNA (H20). The 208-bp polymerase chain reaction product (arrow) from the BON cell was excised, purified, and sequenced.
A WB: SGLT1 Ab SGLT1 Ab +
Blocking Peptide
66 kda - -~ .... :!9~;:~i:a: ::::¢, ,: • ......
45 kda
;oo x-~o,,"
B
GLUT: 45 kda
W ¸
GLUT5 .. ~ ~/
45 kda ~ i ~
Figure 7. Western blotting of SGLT1, GLUT2, and GLUT5. Protein preparations from human jejunum and BON cells were solubilized in sample buffer and separated by 10%-12.5% sodium dodecyl sulfate- polyacrylamide gel electrophoresis, Proteins were transferred to the nitrocellulose membrane. (A) Membrane probing with polyclonal anti- SGLT1 antibody (Ab) indicated the presence of a specific bands of 5 5 - 6 0 kilodaltons in both human jejunum and BON cell preparations. Preabsorption of anti-SGLT1 antibody with immunogenic blocking pep- tide prevented appearance of the band. (B) Membrane probing with polyclonal anti-GLUT2 and anti-GLUT5 antibodies revealed specific bands of 47-50 kilodaltons and 45 kilodaltons, respectively, in hu- man jejunum and BON cells.
daltons, respectively, in human jejunum and BON cells (Figure 7B).
Discussion
The suitability of BON ceils as a model for study- ing 5-HT release has been documented in a previous publication. 1* However, caution must be used in extrap-
olation of the results to enterochromaffin cells in vivo because these cells were derived from a metastasis of a pancreatic carcinoid tumor. Nevertheless, BON cells are currently the only cell line available for study of 5 -HT release, and useful information can be gained that might

December 2001 D-GLUCOSE AND 5-HT RELEASE 1405
have therapeutic potential for treatment of carcinoid
syndrome. Forskolin stimulated 5-HT release and has been used to assess the cyclic adenosine monophosphate- mediated releasable pool of 5-HT in DON cells. Studies with brefeldin A showed an inhibitory effect on forsko-
lin-stimulated 5-HT release without an effect on basal or constitutive release. These disparate effects suggest that different mechanisms may underlie 5-HT release from constitutive and regulated pathways. 19 Our results indi-
cate that D-glucose affects only the phloridzin-sensitive regulated pathway. Although there was a small effect of phloridzin on the constitutive pathway, it was not sig-
nificant. The SGLT family consists of SGLTI, SGLT2, SGLT3,
and possibly other members whose functions are un- known. 2° SGLT1 and SGLT3 are found in intestinal and renal epithelia. 20,21 In the intestine, the predominate form is
SGLT1 based on identification of specific genetic defects that cause glucose-galactose malabsorption. 2° Although
SGLT3 also exists in the intestine, its function is unknown. These transporters vary in their substrate specificity
and affinity. 2°,el For example, SGLT1 is a high-affinity
transporter for D-glucose and galactose. In contrast, SGLT3 is a low-affinity transporter selective for D-glu-
cose in the kidney, although functional studies predict a lower-affinity transporter yet to be cloned.
Our results demonstrate the presence of SGLT-like sequence in human DON cells comparable to a 208- nucleotide domain of the human SGLT1. iv Western
blotting with the putative anti-SGLT1 antibody recog-
nized a predominant protein of 55 -60 kilodaltons in DON cell lysates that appears to be at the lower limit of the range reported for SGLT1. 22 The antiserum was
specific for the 55-60-kilodalton band, which disap- peared in blots incubated with antibody neutralized by prior incubation with the blocking peptide. The 55-60-
kilodalton band was not unique to DON cells; it was also detected in human jejunal lysates.
The functional studies also support the presence of an
SGLT-like protein that may function as a "glucose sen- sor" in DON cells. Like SGLT1 in epithelial cells, D- glucose, galactose, and oLMG, but not fructose, were
substrares for the SGLT-like protein because they in- duced release of 5-HT. This observation could not be explained by osmotic effects at mannitol concentrations
less than 150 mmol/L. Further evidence that SGLT-like protein is necessary
for D-glucose stimulation of 5-HT release from BON
ceils comes from the observation that phloridzin, which blocks SGLT1 in epithelial cells, but not GLUT2 or GLUT5, inhibited 5-HT release stimulated by glucose, e3
The specific mechanism of phloridzin's ability to inhibit glucose release cannot be determined from this study. However, it is possible that interfering with D-glucose
binding and/or translocation prevents 5-HT release. Our results indicate that metabolizable and nonme-
tabolizable hexoses stimulate 5-HT release in DON cells in a concentration-dependent manner. In contrast, 5-HT
is not released by diffusive transport of fructose, a sub- strate for GLUT2 and GLUT5, nor is it mimicked by
iso-osmotic mannitol concentrations below 75 mmol/L. Thus DON cells have chemosensitive properties that suggest they are a model for study of 5-HT release.
The results indicate that the SGLT-like protein in
DON cells has both similarities to and differences from SGLT1. Like intestinal epithelial cells that express SGLT1, DON cells have an SGLT-Iike protein that is involved in chemotransduction in response to metaboliz-
able and nonmetabolizable hexoses. Unlike epithelial cells, detection of extracellular D-glucose by DON cells occurs at high concentrations of glucose. In this regard,
SGLT-like protein in DON cells resembles SGLT32 ° In the ileum, a subset of calbindin-positive primary
afferent neurons containing acetylcholine and substance P are glucoresponsive. 24 These neurons have adenosine
tr iphosphate-gated potassium channels that couple cell metabolism with membrane potential, similar to the
pancreatic beta cell. Chemotransduction in DON cells differs from that in glucosresponsive neurons in the enteric ganglia in 2 ways. First, the D-glucose threshold
(50-75 mmol/L) for evoking 5-HT release in DON cells was 10-15-fold higher than the 5 mmol/L threshold for activation of myenteric neurons. 24 Second, both metab-
olizable and nonmetabolizable hexoses triggered 5-HT release from DON cells, whereas intracellular D-glucose
metabolism was necessary for neural activation. Our results on D-glucose-stimulated 5-HT release
from DON cells are consistent with studies showing that perfusion of the small intestinal lumen with actively
transported hexoses can reflexively inhibit gastric emp- tying by a mechanism independent of their intracellular metabolism. 1 Whether this is mediated by 5-HT release is unknown, but it is likely given that vagal afferents
contain 5-HT 3 receptors on their nerve terminals in close association with the mucosa, and the reflex control of
gastric emptying in this model is blocked by 5-HT 3 receptor antagonists. Like the effect of D-glucose on gastric emptying, carbohydrate-induced pancreatic secre-
tion is dependent on the release of endogenous 5-HT from enterochromaffin cells and activation of 5-HT3 receptors on vagal afferent fibers. 5,6 Therefore, if DON cells are a model of enterochromaffin cells, our results

1406 KIM ET AL. GASTROENTEROLOGY Vol. 121, No. 6
imply that detection of D-glucose and subsequent release of 5-HT may be an important determinant of gastroin- testinal function.
References 1. Raybould HE. Does your gut taste? Sensory transduction in the
gastrointestinal tract. News Physiol Sci 1998;13:275-280. 2. Pappenheimer JR. On the coupling of membrane digestion with
intestinal absorption of sugars and amino acids. Am J Physiol 1993;265:G409-G417.
3. Raybould HE, Bitticaca M, Cann S, Tabrizi Y, Meyer JH, Wong H, Walsh JH, Sternini C. 5HT3 receptors (5HT3R) localized on vagal afferents innervating the stomach and duodenum contribute to the intestinal inhibition of gastric emptying (abstr). Gastroenter- ology 1997;112:A811.
4. Raybould HE, Zitte TT. Inhibition of gastric motility induced by intestinal glucose in awake rats: role of Na(+)-glucose co-trans- porter. Neurogastroenterol Motil 1995;7:9-14.
5. Li Y, Hao Y, Zhu J, Owyang C. Serotonin released from intestinal enterochromaffin cells mediates luminal non-cholecystokinin- stimulated pancreatic secretion in rats. Gastroenterology 2000; 118:1197-1207.
6. Zhu JX, Wu XY, Owyang C, Li Y. Intestinal serotonin acts as a paracrine substance to mediate vagal signal transmission evoked by luminal factors in the rat. J Physiol (Lond) 2001;530: 43•-442.
7. Boden G, Essa N, Owen OE, Reichle FA. Effects of intraduodenal administration of HCl and glucose on circulating immunoreactive, secretin and insulin concentrations. J Clin Invest 1974;53:1185- 1193.
8. Kieffer T J, Buchan AM, Barker H, Brown JC, Pederson RA. Re- lease of gastric inhibitory polypeptide from cultured canine endo- crine cells. Am J Physiol 1994;267:E489-E496.
9. Raybould HE. Nutrient tasting and signaling mechanisms in the gut. I. Sensing of lipid by the intestinal mucosa. Am J Physiol 1999;277:G751-G755.
10. Martin DC, Magnani AD, Kellum JM Jr. Luminal hypertonic solu- tions stimulate concentration-dependent duodenal sereotonin re- lease. Surgery 1989;106:325-331.
11. Buchan AM. Nutrient tasting and signaling mechanisms in the gut. Ill. Endocrine cell recognition of luminal nutrients. Am J Physiol 1999;277:G1103-G1107.
12. Evers BM, Ishizuka J, Townsend CM Jr, Thompson JC. The human carcinoid cell line, BON. A model system for the study of carcinoid tumors. Ann N Y Acad Sci 1994;733:393-406.
13. Parekh D. Ishizuka J, Townsend CM Jr, Haber B, Beauchamp RD, Karp G, Kim SW, Rajaraman S, Greeley G Jr, Thompson JC. Characterization of a human pancreatic carcinoid in vitro: mor-
phology, amine and peptide storage, and secretion. Pancreas 1994;9:83-90.
14. Ahlund L, Nilsson O, Kling-Petersen T, Wigander A, Theodorsson E, Dahlstrom A, Ahiman, H. Serotonin-producing carcinoid tumor cells in long-term culture. Acta Oncologica 1989;28:341-346.
15. Schroppel B, Fischereder M, Wiese P, Segerer S, Huber S, Kret- zler M, Heiss P, Sitter T, Schlondorff D. Expression of glucose transporters in human peritoneal mesothelial cells. Kidney Int 1998;53 :1278-1287.
16. Matosin-Matekalo M, Mesonero JE, Delezay O, Poiree JC, Ilun- dain AA, Brot-Laroche E. Thyroid hormone regulation of the Na+ / glucose cotransporter SGLT1 in Caco-2 cells. Biochem J 1998; 334:633-640.
17. Hediger MA, Turk E, Wright EM. Homology of the human intestinal Na+/glucose and Escherichia coil Na+/prol ine cotransporters. Proc Natl Acad Sci U S A 1989;86 :5748-5752.
18. Kim M, Javed NH, Yu J-G, Christofi F, Cooke HJ. Mechanical stimulation activates G(~q signaling pathways and 5-hydroxytryp- tamine release from human carcinoid BON ceils. J Clin Invest 2001;108:1051-1059.
19. Maroto R, Hamill OP. Brefeldin A block of integrin-dependent mechanosensitive ATP release from Xenopus oocytes reveals a novell mechanism of mechanotransduction. J Biol Chem 2001; 276:23867-23872.
20. Diez-Sampedro A, Eskandari S, Wright EM, Hirayama BA. Na+- to-sugar stoichiometry of SGLT3. Am J Physiol 2001;49:F278- F282.
21. You G, Lee W-S, Barros EJG, Kanai Y, Huo T-L, Khawaja S, Wells RG, Nigam SK, Hediger MA. Molecular characteristics of Na+- coupled glucose transporters in adult and embryonic rat kidney. J Biol Chem 2001;270:29365-27105.
22. Del6zay O, Baghdiguian S, Fantini J. The Development of Na+- dependent glucose transport during differentiation of an intesti- nal epithelial cell clone is regulated by protein kinase C. J Biol Chem 1995;270:12536-12541.
23. Wielert-Badt S, Lin JT, Lorenz M, Fritz S, Kinne RK. Probing the conformation of the sugar transport inhibitor phlorizin by 2D- NMR, molecular dynamics studies, and pharmacophore analysis. J Med Chem 2000;43:1692-1698.
24. Liu M, Seino S, Kirchgessner AL. Identification and characteriza- tion of glucoresponsive neurons in the enteric nervous system. J Neurosci 1999;19:10305-10317.
Received March 31, 2001. Accepted August 16, 2001. Address requests for reprints to: Helen J. Cooke, Ph.D., Department
of Neuroscience, 333 West lOth Avenue, Columbus, Ohio 43210. e-mail: [email protected]; fax: (614) 688-8742.
Supported by the National Institutes of Health grants DK37240 (to H.J.C.), DK57016 (to H.J.C.), DK44179 (to F.C.), and DK41004 (to H.E.R.) and a grant from the Indiana Academy of Science (to N.H.J.)