
Cytotoxic effects in 3T3-L1 mouse and WI-38 human fibroblasts following 72hour and 7day exposures to...
13
Cytotoxic effects in 3T3-L1 mouse and WI-38 human fibroblasts following 72 hour and 7 day exposures to commercial silica nanoparticles Maciej Stępnik a, ⁎, Joanna Arkusz a , Anna Smok-Pieniążek a , Anna Bratek-Skicki b , Anna Salvati b , Iseult Lynch b , Kenneth A. Dawson b , Jolanta Gromadzińska a , Wim H. De Jong c , Konrad Rydzyński a a Nofer Institute of Occupational Medicine, Łódź, Poland b Centre for BioNano Interactions, School of Chemistry and Chemical Biology, University College Dublin, Belfield, Dublin 4, Ireland c National Institute for Public Health and the Environment, Antonie van Leeuwenhoeklaan 9 NL‐3720, Bilthoven, The Netherlands abstract article info Article history: Received 24 February 2012 Revised 27 May 2012 Accepted 4 June 2012 Available online 13 June 2012 Keywords: Cell cycle Cytotoxicity Fibroblasts Nanoparticles Oxidative stress Silica The potential toxic effects in murine (3T3-L1) and human (WI-38) fibroblast cell lines of commercially available silica nanoparticles (NPs), Ludox CL (nominal size 21 nm) and CL-X (nominal size of 30 nm) were investigated with particular attention to the effect over long exposure times (the tests were run after 72 h exposure up to 7 days). These two formulations differed in physico-chemical properties and showed different stabilities in the cell culture medium used for the experiments. Ludox CL silica NPs were found to be cytotoxic only at the higher concentrations to the WI-38 cells (WST-1 and LDH assays) but not to the 3T3-L1 cells, whereas the Ludox CL-X silica NPs, which were less stable over the 72 h exposure, were cytotoxic to both cell lines in both assays. In the clonogenic assay both silica NPs induced a concentration dependent decrease in the surviving fraction of 3T3-L1 cells, with the Ludox CL-X silica NPs being more cytotoxic. Cell cycle anal- ysis showed a trend indicating alterations in both cell lines at different phases with both silica NPs tested. Buthionine sulfoximine (γ-glutamylcysteine synthetase inhibitor) combined with Ludox CL-X was found to induce a strong decrease in 3T3-L1 cell viability which was not observed for the WI-38 cell line. This study clearly indicates that longer exposure studies may give important insights on the impact of nanomaterials on cells. However, and especially when investigating nanoparticle effects after such long exposure, it is fun- damental to include a detailed physico-chemical characterization of the nanoparticles and their dispersions over the time scale of the experiment, in order to be able to interpret eventual impacts on cells. © 2012 Elsevier Inc. All rights reserved. Introduction Silica is commonly found in the earth's crust and is widely produced for many industrial purposes (Merget et al., 2002), therefore human exposure to its different physical forms is inevitable. There are two basic forms of silica which fundamentally differ in their biological activity, yet have the same molecular formula: crystalline and non- crystalline (amorphous) silica. Occupational exposure to crystalline silica was associated with development of pulmonary silicosis, increased risk of chronic bronchitis, chronic obstructive pulmonary disease, and lung cancer (IARC, 1997; Merget et al., 2002). In contrast, occupational exposure to amorphous silica is generally not associated with persistent pathological changes in man (McLaughlin et al., 1997; Merget et al., 2002) or in experimental animals (Arts et al., 2007; Johnston et al., 2000). Moreover, there is inadequate evidence in humans for the carcinogenicity of amorphous silica (IARC, 1997). Although the biological effects of exposure to silica at micro-scale size are relatively well known, its effects at nano-scale size are still unclear. Assuming the existence of differential toxicity between materials at nano- and microscale, e.g. as demonstrated for TiO 2 (Oberdörster, 2000) or polystyrene NPs (Brown et al., 2001), it is likely that nano-sized silica can have different biological effects from silica at the micro-scale size. The issue becomes more important considering the increasing use of silica NPs in consumer products and therapeutics. Silica dye-doped or functionalized NPs have found appli- cations in bioanalysis and bioimaging (Kumar et al., 2008; Liong et al., 2008). The usefulness of silica nanoparticles was also demonstrated for labeling human cancer cells in vitro, as carriers for DNA transfection and drug delivery, coatings for contrast agents, and in other applica- tions (Fukumori et al., 2007; Santra and Dutta, 2007). Ultrafine amorphous silica (diameter 14 nm with no surface charac- teristics given; from Sigma-Aldrich, St. Louis, MO, USA) induced a severe but reversible pulmonary inflammation in mice after intratracheal instil- lation (Cho et al., 2007). In addition, ultrafine silica particles had greater Toxicology and Applied Pharmacology 263 (2012) 89–101 ⁎ Corresponding author at: Department of Toxicology and Carcinogenesis, Nofer Institute of Occupational Medicine, 8 Sw. Teresy St 91‐348 Łódź, Poland. Fax: +48 42 6314 610. E-mail address: [email protected] (M. Stępnik). 0041-008X/$ – see front matter © 2012 Elsevier Inc. All rights reserved. doi:10.1016/j.taap.2012.06.002 Contents lists available at SciVerse ScienceDirect Toxicology and Applied Pharmacology journal homepage: www.elsevier.com/locate/ytaap
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Cytotoxic effects in 3T3-L1 mouse and WI-38 human fibroblasts following 72hour and 7day exposures to...

Toxicology and Applied Pharmacology 263 (2012) 89–101
Contents lists available at SciVerse ScienceDirect
Toxicology and Applied Pharmacology
j ourna l homepage: www.e lsev ie r .com/ locate /ytaap
Cytotoxic effects in 3T3-L1 mouse and WI-38 human fibroblasts following 72 hourand 7 day exposures to commercial silica nanoparticles
Maciej Stępnik a,⁎, Joanna Arkusz a, Anna Smok-Pieniążek a, Anna Bratek-Skicki b, Anna Salvati b,Iseult Lynch b, Kenneth A. Dawson b, Jolanta Gromadzińska a, Wim H. De Jong c, Konrad Rydzyński aa Nofer Institute of Occupational Medicine, Łódź, Polandb Centre for BioNano Interactions, School of Chemistry and Chemical Biology, University College Dublin, Belfield, Dublin 4, Irelandc National Institute for Public Health and the Environment, Antonie van Leeuwenhoeklaan 9 NL‐3720, Bilthoven, The Netherlands
⁎ Corresponding author at: Department of ToxicoloInstitute of Occupational Medicine, 8 Sw. Teresy St 91‐36314 610.
E-mail address: [email protected] (M. Stępnik).
0041-008X/$ – see front matter © 2012 Elsevier Inc. Alldoi:10.1016/j.taap.2012.06.002
a b s t r a c t
a r t i c l e i n f oArticle history:Received 24 February 2012Revised 27 May 2012Accepted 4 June 2012Available online 13 June 2012
Keywords:Cell cycleCytotoxicityFibroblastsNanoparticlesOxidative stressSilica
The potential toxic effects in murine (3T3-L1) and human (WI-38) fibroblast cell lines of commerciallyavailable silica nanoparticles (NPs), Ludox CL (nominal size 21 nm) and CL-X (nominal size of 30 nm) wereinvestigated with particular attention to the effect over long exposure times (the tests were run after 72 hexposure up to 7 days). These two formulations differed in physico-chemical properties and showed differentstabilities in the cell culture medium used for the experiments. Ludox CL silica NPs were found to be cytotoxiconly at the higher concentrations to the WI-38 cells (WST-1 and LDH assays) but not to the 3T3-L1 cells,whereas the Ludox CL-X silica NPs, which were less stable over the 72 h exposure, were cytotoxic to bothcell lines in both assays. In the clonogenic assay both silica NPs induced a concentration dependent decreasein the surviving fraction of 3T3-L1 cells, with the Ludox CL-X silica NPs being more cytotoxic. Cell cycle anal-ysis showed a trend indicating alterations in both cell lines at different phases with both silica NPs tested.Buthionine sulfoximine (γ-glutamylcysteine synthetase inhibitor) combined with Ludox CL-X was found toinduce a strong decrease in 3T3-L1 cell viability which was not observed for the WI-38 cell line. This studyclearly indicates that longer exposure studies may give important insights on the impact of nanomaterialson cells. However, and especially when investigating nanoparticle effects after such long exposure, it is fun-damental to include a detailed physico-chemical characterization of the nanoparticles and their dispersionsover the time scale of the experiment, in order to be able to interpret eventual impacts on cells.
© 2012 Elsevier Inc. All rights reserved.
Introduction
Silica is commonly found in the earth's crust and is widely producedfor many industrial purposes (Merget et al., 2002), therefore humanexposure to its different physical forms is inevitable. There are twobasic forms of silica which fundamentally differ in their biologicalactivity, yet have the same molecular formula: crystalline and non-crystalline (amorphous) silica. Occupational exposure to crystallinesilica was associated with development of pulmonary silicosis,increased risk of chronic bronchitis, chronic obstructive pulmonarydisease, and lung cancer (IARC, 1997; Merget et al., 2002). In contrast,occupational exposure to amorphous silica is generally not associatedwith persistent pathological changes in man (McLaughlin et al., 1997;Merget et al., 2002) or in experimental animals (Arts et al., 2007;
gy and Carcinogenesis, Nofer48 Łódź, Poland. Fax: +48 42
rights reserved.
Johnston et al., 2000). Moreover, there is inadequate evidence inhumans for the carcinogenicity of amorphous silica (IARC, 1997).
Although the biological effects of exposure to silica at micro-scalesize are relatively well known, its effects at nano-scale size are stillunclear. Assuming the existence of differential toxicity betweenmaterials at nano- and microscale, e.g. as demonstrated for TiO2
(Oberdörster, 2000) or polystyrene NPs (Brown et al., 2001), it islikely that nano-sized silica can have different biological effects fromsilica at the micro-scale size. The issue becomes more importantconsidering the increasing use of silica NPs in consumer products andtherapeutics. Silica dye-doped or functionalized NPs have found appli-cations in bioanalysis and bioimaging (Kumar et al., 2008; Liong et al.,2008). The usefulness of silica nanoparticles was also demonstrated forlabeling human cancer cells in vitro, as carriers for DNA transfectionand drug delivery, coatings for contrast agents, and in other applica-tions (Fukumori et al., 2007; Santra and Dutta, 2007).
Ultrafine amorphous silica (diameter 14 nm with no surface charac-teristics given; from Sigma-Aldrich, St. Louis, MO, USA) induced a severebut reversible pulmonary inflammation in mice after intratracheal instil-lation (Cho et al., 2007). In addition, ultrafine silica particles had greater

90 M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
ability to induce lung inflammation and tissue damage compared to finesilica particles, suggesting that silica NPs may indeed pose a humanhealth hazard (Kaewamatawong et al., 2005). At the cellular level, amor-phous silica NPs were found to induce cytotoxicity in a range of differentcell types. Lin et al. (2006) reported cytotoxic effects of 15 and 46 nmamorphous silica (hydrodynamic diameters: 590±104 nm and 617±107 nm, respectively; from Degussa Co., Parsippany, NJ, USA) to A549human lung cancer cells after 48 and 72 h of exposure to concentrationsof tens ofmicrograms permilliliter. This effectwas highly correlatedwiththe induction of oxidative stress in the cells. In the study by Lison et al.(2008), cytotoxicity of Ströber amorphous silica NPs after 24 h of expo-sure (TEMand hydrodynamic diameters of 29.3±4.4 and 35 nm, respec-tively) at concentrations below 200 μg/ml was shown in A549 cells, J774mouse monocyte–macrophage and EAHY926 human endothelium cells.The effect of Ströber silica nano- and ultrafine particles (size range of16.4–335 nm) on EAHY926 cells after 24 h of exposure to high concen-trations evidently depended on their size, the smaller particles having alower IC50 value compared to the larger particles (Napierska et al.,2009). Human fibroblasts with longer doubling times were shown tobe more susceptible to damage after 48 h exposure to silica NPs (meandiameters by SEM for silica prepared from silicate: 21.58±4.36 nmwith hydrodynamic size of 188.3±11.5 nm; and prepared from TEOS:80.21±14.43 nm with hydrodynamic size of 236.3±6.85 nm) thanhuman tumor cells with shorter doubling times (Chang et al., 2007).The data provided by Nan et al. (2008) indicated a growth inhibitory ef-fect of silica nanotubes on MDA-MB-231 breast cancer and primaryhuman umbilical vein endothelial cells. The effect of unmodified nan-otubes was clearly visible at concentrations as low as 5 μg/ml and wasmore significant for HUVEC cells. Thus, it can be concluded that undersome conditions, some silica NPs do express toxicity for cell lines invitro (Ahmad et al., 2012; Drescher et al., 2011; Eom and Choi, 2011;Gehrke et al., 2012; Lu et al., 2011; Nabeshi et al., 2011a; Napierskaet al., 2010; Rabolli et al., 2011). There are however, reports, includingfrom our consortium (Barnes et al., 2008), where silica NPs from arange of sources were found not to be cytotoxic (MTT, WST-1, LDH,propidium iodide tests) or genotoxic (Comet assay), following short ex-posures (up to 24 h). All the data indicate that a deeper analysis of thephysico-chemical characteristics of the various NPs used, the exposuredoses, the cell types used and so on are important in order to draw anygeneral conclusions regarding the in vitro toxicity of silica NPs.
In this study we investigated two different types of silica NPs, onepositively charged with an alumina coating and one negatively chargedwith no coating, although stabilized with ethylene glycol. The two silicananoparticle formulations studied had an identical composition but adifferent surface charge/coating and dispersing agent. The Ludox CL sil-ica NPs had an alumina coating and were dispersed in a chloridestabilized solution inducing a positive surface charge on the silica NPs(in water, although this became negative when the particles where dis-persed in DMEMwith 10% fetal bovine serum), whereas the Ludox CL-Xsilica NPs were present in a sodium stabilized solution resulting in anegative surface charge of the silica NPs (in water) and also had 7% eth-ylene glycol in the solution. Considering reports indicating potential in-terference of test materials with assays (Wörle-Knirsch et al., 2006) weapplied two different standard cytotoxicity assays, the 72 h tetrazoliumreduction (WST-1) assay and the LDH release test, which measure cel-lularmetabolismandmembrane permeability, respectively. These cyto-toxicity tests were further supported with cell cycle analysis. To detectmore subtle changes induced by the NPs prolonged exposure timeswere used. Such a strategy requires more careful standardization oftesting parameters (e.g. adjusting initial cell density or determinationof nanoparticle stability over the entire experiment). The effects oflong-term exposure of the two fibroblast cell lines to the silica NPswere investigated using a clonogenic survival assay and the 7-daycytotoxicity test. As oxidative stress induction is suggested as animportant mechanism for nanoparticle cytotoxicity, the involvementof reactive oxygen species in the observed effects was indirectly
assessed by inhibition of glutathione synthesis in the cells. In order toinvestigate whether any of the observed effects were due to the NPsthemselves or a result of some residual impurities from the synthesisor stabilization of theNPs, both Ludox CL and CL-XNPswere extensivelydialyzed against Milli‐Q water, and the dialyzed NPs were tested inparallel with the as-purchased NPs. Thus, both dialyzed and nondialyzed nanoparticle samples were investigated to exclude possibletoxic effects from production residues/contaminants in the silica nano-particle preparations.
The study was performed using mouse embryonic 3T3-L1 fibro-blasts as they are readily available, undergo contact inhibition, aremore closely representative of a physiologic model than e.g. cancercell lines, and they represent one of the most commonly non-epithelial cell lines used in short- and long-term nanotoxicologicalin vitro studies on cytotoxicity, biocompatibility, or mechanisms ofcellular uptake of nanoparticles (Napierska et al., 2010). Moreover,fibroblasts are known to be actively involved in excess collagendeposition, ultimately resulting in interstitial fibrosis and formationof silicotic nodules, a hallmark of silicosis, thus represent a goodmodel for the impact of silica nanoparticles in particular. To comparethe results, a similar in respect to origin and behavior, normal humanembryonic lung fibroblast cell line WI-38 was chosen.
Materials and methods
Chemicals and reagents. L-Buthionine-sulfoximine (BSO, #B2515),propidium iodide (PI #81845), N-acetyl-L-cysteine (#A9165), and5,5′-dithio-bis(2-nitrobenzoic acid) (DTNB) were obtained fromSigma-Aldrich Co. Cytotoxicity Detection Kit (LDH, #11 644 793 001)and Cell Proliferation Reagent (WST-1, #11 644 807 001) werepurchased from Roche. RNase A was obtained from Fermentas(#EN0531). The composition of Hanks' Balanced Salt Solution (HBSS)was the following: 0.4 g/l KCl, 0.06 g/l KH2PO4, 8 g/l NaCl, 0.35 g/lNaHCO3, 0.04788 g/l Na2HPO4 (pH 7.2–7.4). Stock solution of BSO(10 mM) was prepared in deionized H2O, filter-sterilized (0.2 μm) andstored at 4 °C until used.
Particle source and their characteristics. Two amorphous silica NPs:LUDOX® CL colloidal silica 30 wt.% in H2O (alumina coated chloridestabilizing counterion) (#420883) and LUDOX® CL-X colloidalsilica 45 wt.% suspension in H2O (sodium-stabilizing counterion)(#420891) were purchased from Sigma-Aldrich Co. Thus, the parti-cles were with a common size but with varied surface characteristics.Before use the particles were investigated for possible bacterialcontamination.
Particle size, pH and zeta potential weremeasured for particles afterdialysis. DLS and zeta potential measurements were performed on aphoton correlation spectrophotometer Malvern 3000HS Zetasizer. Forthese measurements, particle stocks were diluted in filtered Milli‐Qwater to final concentrations of 10 mg/ml. Stability of CL and CL-X silicaNPs was studied in water at the concentration of 40 μg/ml and inDMEM supplemented with 10% FBS at 37 °C immediately after prepa-ration (0 h) and after 24 or 72 h of incubation. The concentrationrange in DMEM was from 40 to 800 μg/ml. Zeta potential of NPssuspended in DMEM/ 10% FBS was measured after 72 h of incubationat 37 °C.
Preparation of silica dilutions. Stock solutions of silica NPs (30%) weredialyzed extensively against Milli-Q water, and then diluted using0.2 μm filtered deionized water to 10 mg/ml. Then, a range of testconcentrations for each nanoparticle was prepared in culture medi-um, and was applied to the cells immediately (within 10 min). Thefinal concentration of water in each test sample was 10%. For compar-ison, the undialyzed stock was also diluted directly with the filteredMilli-Q water, and then culture media.

91M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
Cell culture. Murine fibroblasts 3T3-L1 (#CL-173) were purchased fromAmerican Type Culture Collection and human fibroblasts WI-38(#90020107) were purchased from the European Collection of CellCultures (ECACC). Both cell linesweremaintained in Dulbecco'smodifiedEagle's medium (DMEM) (Sigma #D5671) supplementedwith 10% heat-inactivated Fetal Bovine Serum (Gibco #10106-169, lot#41F3271F),25 mM Hepes, 4 mM L-glutamine, 100 U/ml penicillin, and 100 μg/mlstreptomycin (Sigma-Aldrich Co.). The cells were screened for Mycoplas-ma sp. infection using indicator cell line 3T6 cells (ATCC#CCL-96) andMycoTech Kit (Gibco BRL).
WST-1 reduction assay. The cells seeded onto 96-well microplates ona prior day (1.5×103 cells/well for 3T3-L1 and 4×103 cells/well forWI-38 in 100 μl) were incubated in the absence or presence of testsamples for 72 h without removal of the medium. For 3T3-L1 andWI-38 fibroblasts an extended cytotoxicity protocol was also used,with 7 days of exposure to dialyzed Ludox silica samples. In this pro-tocol the cells were plated onto 96-well microplates at a density of 50cells/well (3T3-L1) or 500 cells/well (WI-38) and allowed to adherefor at least 16 h before treatment with silica at selected concentra-tions. The cells were treated with silica for 7 days without mediumrenewal. After exposure, to all wells 10 μl WST-1 reagent was addedfor 1 h.
The optical density of solubilized formazan product was deter-mined using a spectrophotometer (with a 450 nm and 620 nm filteras a reference). The results were expressed as the percent of cellsurvival (OD of exposed vs. OD of control non-exposed cells). Beforeexperiments possible interference of the NPs with the viability assaywas assessed. To this end, the range of silica NPs dispersions overthe same concentration range as used in the cytotoxicity assay wereincubated in culture medium without cells for 72 h and absorbancemeasurements at the same wavelengths used for WST-1 reductionassay were performed. Also, a standard protocol for WST-1 reductionwas applied to check if the NPs had an effect on the reduction poten-tial per se. The measurements did not show any significant reducingpotential of either type of silica NPs on the WST-1 reagent (data in-cluded in Supplementary information, §S7). Only a minimal increasein absorbance was observed in the case of CL-X silica at the highestconcentrations, and therefore all results were corrected accordingly.
Lactate dehydrogenase measurement. The assay was performed ac-cording to the manufacturer's protocol. Briefly, cells were plated onto96-well microplates (1.5×103 cells/well for 3T3-L1 and 4×103 cells/well for WI-38 seeded on a prior day) and incubated in the absence orpresence of test samples for 72 h. For testing the released LDH activity,10 μl of supernatantswas diluted 10 times inHBSS to reduce undesirableeffect of FBS and phenol red dye present in the culture medium. Next,100 μl reaction mixture (freshly prepared) was added to each well andincubated for up to 30 min (protected from the light). Afterwards, 50 μl1 N HCl was added to stop the reaction and the absorbance wasmeasured at 450 nm (with 620 nm filter as a reference). Results wereexpressed as the percent of LDH activitymeasured in control (non-treat-ed) samples. To determine themaximum LDH release, cells were treatedwith 1% Triton-X100 for 60 min before running the assay. Thereafter, forsuch prepared lysates the same protocol as for supernatants was used.Similar toWST-1 test, we performedmeasurements to check for possibleinterference of the NPs with the LDH reagent as well as their potential toinhibit LDH activity. The measurements did not show any significant in-fluence of silica NPs (data presented in Supplementary information, §S8).
Cytotoxicity of dialyzed Ludox silica NPs on 3T3-L1 mouse fibroblasts inclonogenic survival assay. 3T3-L1 cells were seeded onto 60-mmplates at the density of 100 cells/plate in 3 ml. On the next day thecells were exposed to dialyzed Ludox samples at selected concentra-tions and incubated for 72 h. On the 4th day supernatants wereexchanged for fresh cDMEM medium without Ludox silica and the
cells were incubated for an additional 4 days. After the exposure theplates were washed with PBS, the colonies were fixed with 2% forma-lin, stained with 0.5% crystal violet and the number of survivors (i.e.,colonies containing >50 cells) was scored. Plating Efficiency (PE) wascalculated as the ratio of the number of colonies to the number ofcells seeded:
PE ¼ Number of colonies formed=Number of cells seededð Þ � 100%:
Surviving fraction (SF) calculated as the number of colonies thatarose after treatment of cells was expressed in terms of PE:
SF ¼ Number of colonies formed after treatment=Number of cells seededð Þ�PE:
Cell cycle analysis. To assess cell cycle changes of 3T3-L1 and WI-38 fi-broblasts exposed to dialyzed Ludox silica NPs a standard propidium io-dide staining protocol was applied. After exposure for 24 or 48 h thecells were trypsinized, washed twice with ice-cold PBS, incubated for15 min in PBS on ice and fixed with 70% ethanol at −20 °C overnight.Then, the cells were spun, washed in PBS, resuspended in PBS con-taining 50 μg/ml propidium iodide and 10 μg/ml RNAse A and subse-quently analyzed with an FACS CantoII flow cytometer (BectonDickinson). Cycle profiles of ten thousands collected cells were deter-mined using ModFit LT 3.0 software (Verity Software House, Inc.).
Treatment with BSO. To modulate potential silica-induced oxidativestress in exposed cells glutathione level in cells was reduced by a spe-cific inhibitor of glutathione synthesis (BSO). 3T3-L1 and WI-38 cellsplated onto 96-well microplates on a previous day were co-incubatedwith dialyzed silica samples and BSO for 72 h. After the incubation pe-riod WST-1 reduction assay was performed as described above.
Glutathione (GSH) measurements. The cells were incubated in 6-wellmicroplates with BSO at the concentration of 10 μM (for 3T3-L1cells) or at 2.5 μM (forWI-38 cells) for 24 or 72 h. After the incubationperiod, they were processed for GSH measurements. The spectropho-tometric method for reduced GSH determination is based on oxida-tion of GSH by the sulfhydryl reagent 5,5′-dithio-bis(2-nitrobenzoicacid) (DTNB) to form the yellow derivative 5′-thio-2-nitrobenzoic acid,measurable at 415 nm on amicroplate reader (Rahman et al., 2007). Be-fore colorimetric reaction cells were lysed and deproteinized using 5%HClO4. Reduced glutathione (5 nmol/ml) was used to generate standardcurve. Protein concentration in cell lysates was determined using Brad-ford method with bovine serum albumin as standard (Bradford, 1976).
Statistical analysis. All results are presented as a mean±SD from thenumber of independent experiments shown. All samples wereassayed in triplicates. Bartlett's test of homogeneity of variance wasused to determine if the results had equivalent variances at thepb0.05 level. If the variances were not significantly different, the re-sults were compared using a standard one-way analysis of variance(ANOVA). When the F-test from ANOVA was significant, the Dun-nett's test was used to compare means from the control and exposedsamples.
Results
Dispersion measurements
The physico-chemical characterization of the NP dispersions inMilli‐Q water at 40 μg/ml is shown in Table 1. The size of thedispersed NPs in solution is slightly different from the nominalmanufacturer's stated size, likely as a result of the difference betweenthe hydrodynamic diameter and the dry diameter. The DLS size
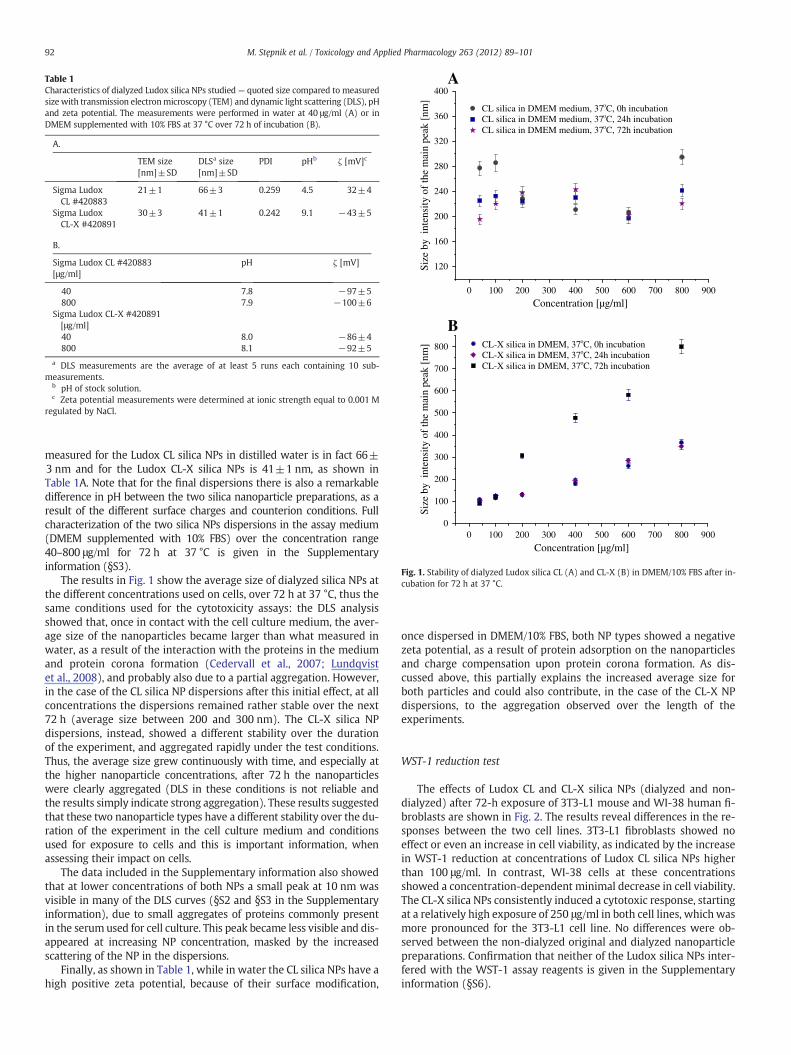
Table 1Characteristics of dialyzed Ludox silica NPs studied— quoted size compared to measuredsize with transmission electronmicroscopy (TEM) and dynamic light scattering (DLS), pHand zeta potential. The measurements were performed in water at 40 μg/ml (A) or inDMEM supplemented with 10% FBS at 37 °C over 72 h of incubation (B).
A.
TEM size[nm]±SD
DLSa size[nm]±SD
PDI pHb ζ [mV]c
Sigma LudoxCL #420883
21±1 66±3 0.259 4.5 32±4
Sigma LudoxCL-X #420891
30±3 41±1 0.242 9.1 −43±5
B.
Sigma Ludox CL #420883[μg/ml]
pH ζ [mV]
40 7.8 −97±5800 7.9 −100±6
Sigma Ludox CL-X #420891[μg/ml]40 8.0 −86±4800 8.1 −92±5
a DLS measurements are the average of at least 5 runs each containing 10 sub-measurements.
b pH of stock solution.c Zeta potential measurements were determined at ionic strength equal to 0.001 M
regulated by NaCl.
120
160
200
240
280
320
360
400A
B
Size
by
inte
nsity
of
the
mai
n pe
ak [
nm]
0 100 200 300 400 500 600 700 800 900
0 100 200 300 400 500 600 700 800 9000
100
200
300
400
500
600
700
800
Size
by
inte
nsity
of
the
mai
n pe
ak [
nm]
CL silica in DMEM medium, 37oC, 0h incubation CL silica in DMEM medium, 37oC, 24h incubation CL silica in DMEM medium, 37oC, 72h incubation
CL-X silica in DMEM, 37oC, 0h incubation CL-X silica in DMEM, 37oC, 24h incubation CL-X silica in DMEM, 37oC, 72h incubation
Concentration [µg/ml]
Concentration [µg/ml]
Fig. 1. Stability of dialyzed Ludox silica CL (A) and CL-X (B) in DMEM/10% FBS after in-cubation for 72 h at 37 °C.
92 M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
measured for the Ludox CL silica NPs in distilled water is in fact 66±3 nm and for the Ludox CL-X silica NPs is 41±1 nm, as shown inTable 1A. Note that for the final dispersions there is also a remarkabledifference in pH between the two silica nanoparticle preparations, as aresult of the different surface charges and counterion conditions. Fullcharacterization of the two silica NPs dispersions in the assay medium(DMEM supplemented with 10% FBS) over the concentration range40–800 μg/ml for 72 h at 37 °C is given in the Supplementaryinformation (§S3).
The results in Fig. 1 show the average size of dialyzed silica NPs atthe different concentrations used on cells, over 72 h at 37 °C, thus thesame conditions used for the cytotoxicity assays: the DLS analysisshowed that, once in contact with the cell culture medium, the aver-age size of the nanoparticles became larger than what measured inwater, as a result of the interaction with the proteins in the mediumand protein corona formation (Cedervall et al., 2007; Lundqvistet al., 2008), and probably also due to a partial aggregation. However,in the case of the CL silica NP dispersions after this initial effect, at allconcentrations the dispersions remained rather stable over the next72 h (average size between 200 and 300 nm). The CL-X silica NPdispersions, instead, showed a different stability over the durationof the experiment, and aggregated rapidly under the test conditions.Thus, the average size grew continuously with time, and especially atthe higher nanoparticle concentrations, after 72 h the nanoparticleswere clearly aggregated (DLS in these conditions is not reliable andthe results simply indicate strong aggregation). These results suggestedthat these two nanoparticle types have a different stability over the du-ration of the experiment in the cell culture medium and conditionsused for exposure to cells and this is important information, whenassessing their impact on cells.
The data included in the Supplementary information also showedthat at lower concentrations of both NPs a small peak at 10 nm wasvisible in many of the DLS curves (§S2 and §S3 in the Supplementaryinformation), due to small aggregates of proteins commonly presentin the serum used for cell culture. This peak became less visible and dis-appeared at increasing NP concentration, masked by the increasedscattering of the NP in the dispersions.
Finally, as shown in Table 1, while in water the CL silica NPs have ahigh positive zeta potential, because of their surface modification,
once dispersed in DMEM/10% FBS, both NP types showed a negativezeta potential, as a result of protein adsorption on the nanoparticlesand charge compensation upon protein corona formation. As dis-cussed above, this partially explains the increased average size forboth particles and could also contribute, in the case of the CL-X NPdispersions, to the aggregation observed over the length of theexperiments.
WST-1 reduction test
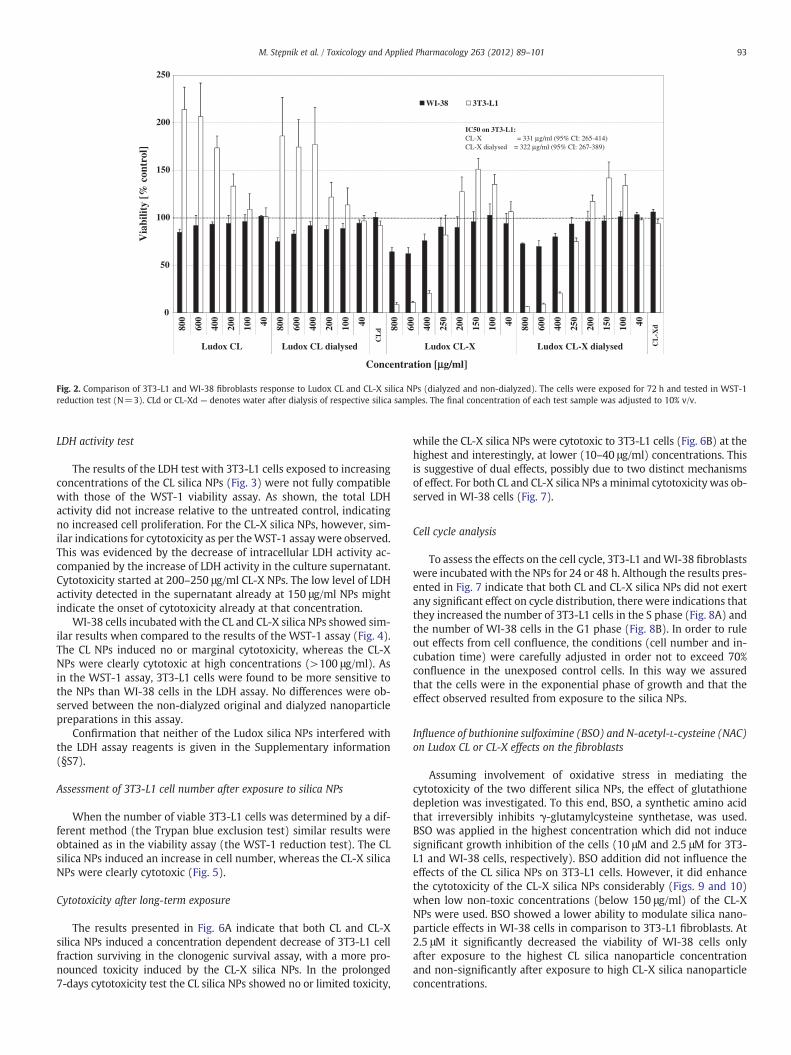
The effects of Ludox CL and CL-X silica NPs (dialyzed and non-dialyzed) after 72-h exposure of 3T3-L1 mouse and WI-38 human fi-broblasts are shown in Fig. 2. The results reveal differences in the re-sponses between the two cell lines. 3T3-L1 fibroblasts showed noeffect or even an increase in cell viability, as indicated by the increasein WST-1 reduction at concentrations of Ludox CL silica NPs higherthan 100 μg/ml. In contrast, WI-38 cells at these concentrationsshowed a concentration-dependent minimal decrease in cell viability.The CL-X silica NPs consistently induced a cytotoxic response, startingat a relatively high exposure of 250 μg/ml in both cell lines, which wasmore pronounced for the 3T3-L1 cell line. No differences were ob-served between the non-dialyzed original and dialyzed nanoparticlepreparations. Confirmation that neither of the Ludox silica NPs inter-fered with the WST-1 assay reagents is given in the Supplementaryinformation (§S6).
0
50
100
150
200
250
800
600
400
200
100 40 800
600
400
200
100 40 800
600
400
250
200
150
100 40 800
600
400
250
200
150
100 40
Ludox CL Ludox CL dialysed Ludox CL-X Ludox CL-X dialysed
Via
bilit
y [%
con
trol
]
Concentration [µµg/ml]
WI-38 3T3-L1
IC50 on 3T3-L1:CL-X = 331 µg/ml (95% CI: 265-414) CL-X dialysed = 322 µg/ml (95% CI: 267-389)
CL
d
CL
-Xd
Fig. 2. Comparison of 3T3-L1 and WI-38 fibroblasts response to Ludox CL and CL-X silica NPs (dialyzed and non-dialyzed). The cells were exposed for 72 h and tested in WST-1reduction test (N=3). CLd or CL-Xd — denotes water after dialysis of respective silica samples. The final concentration of each test sample was adjusted to 10% v/v.
93M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
LDH activity test
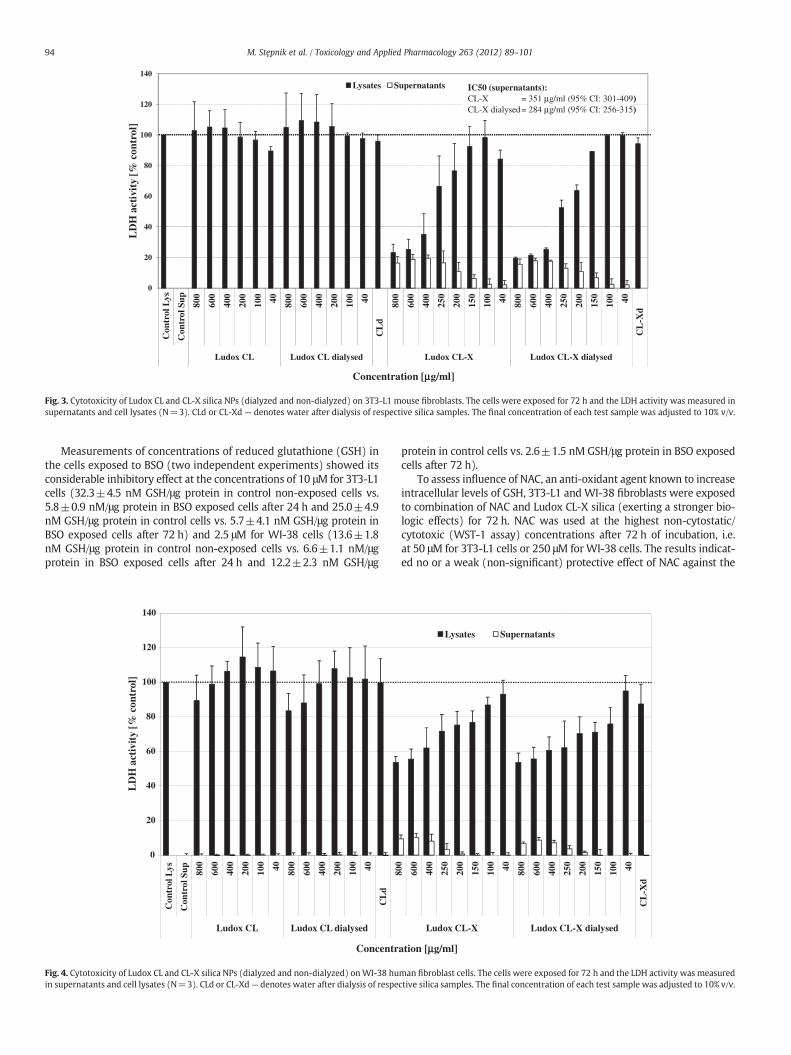
The results of the LDH test with 3T3-L1 cells exposed to increasingconcentrations of the CL silica NPs (Fig. 3) were not fully compatiblewith those of the WST-1 viability assay. As shown, the total LDHactivity did not increase relative to the untreated control, indicatingno increased cell proliferation. For the CL-X silica NPs, however, sim-ilar indications for cytotoxicity as per theWST-1 assay were observed.This was evidenced by the decrease of intracellular LDH activity ac-companied by the increase of LDH activity in the culture supernatant.Cytotoxicity started at 200–250 μg/ml CL-X NPs. The low level of LDHactivity detected in the supernatant already at 150 μg/ml NPs mightindicate the onset of cytotoxicity already at that concentration.
WI-38 cells incubated with the CL and CL-X silica NPs showed sim-ilar results when compared to the results of the WST-1 assay (Fig. 4).The CL NPs induced no or marginal cytotoxicity, whereas the CL-XNPs were clearly cytotoxic at high concentrations (>100 μg/ml). Asin the WST-1 assay, 3T3-L1 cells were found to be more sensitive tothe NPs than WI-38 cells in the LDH assay. No differences were ob-served between the non-dialyzed original and dialyzed nanoparticlepreparations in this assay.
Confirmation that neither of the Ludox silica NPs interfered withthe LDH assay reagents is given in the Supplementary information(§S7).
Assessment of 3T3-L1 cell number after exposure to silica NPs
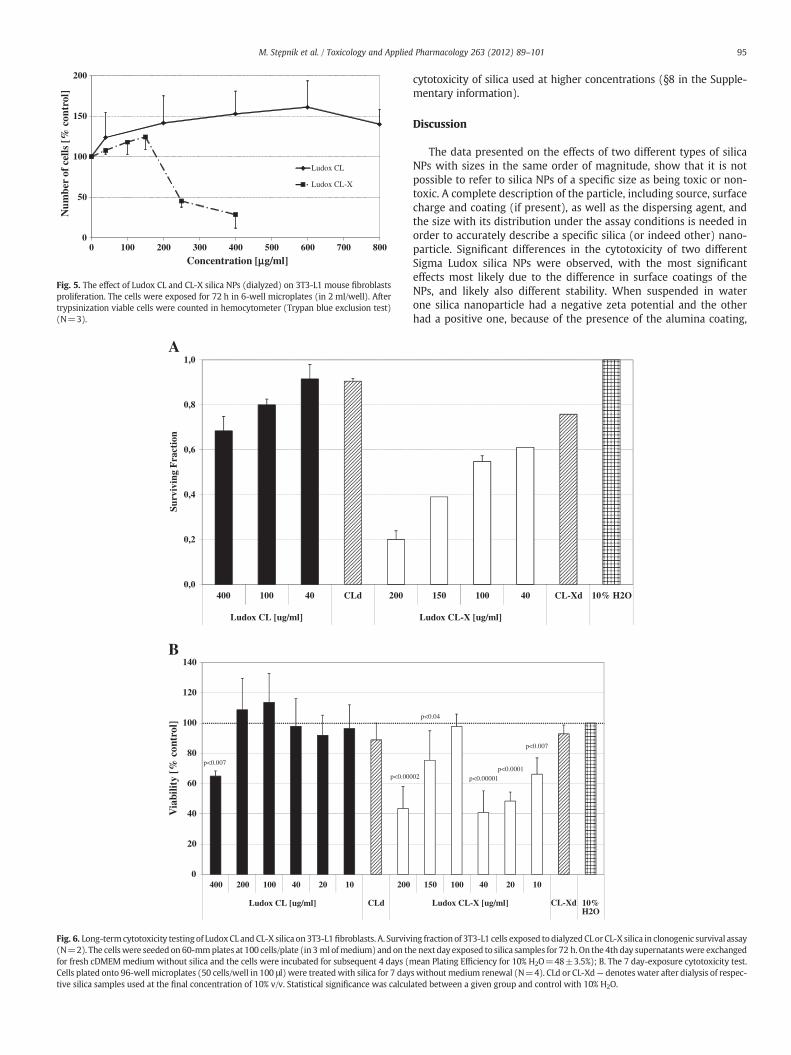
When the number of viable 3T3-L1 cells was determined by a dif-ferent method (the Trypan blue exclusion test) similar results wereobtained as in the viability assay (the WST-1 reduction test). The CLsilica NPs induced an increase in cell number, whereas the CL-X silicaNPs were clearly cytotoxic (Fig. 5).
Cytotoxicity after long-term exposure
The results presented in Fig. 6A indicate that both CL and CL-Xsilica NPs induced a concentration dependent decrease of 3T3-L1 cellfraction surviving in the clonogenic survival assay, with a more pro-nounced toxicity induced by the CL-X silica NPs. In the prolonged7-days cytotoxicity test the CL silica NPs showed no or limited toxicity,
while the CL-X silica NPs were cytotoxic to 3T3-L1 cells (Fig. 6B) at thehighest and interestingly, at lower (10–40 μg/ml) concentrations. Thisis suggestive of dual effects, possibly due to two distinct mechanismsof effect. For both CL and CL-X silica NPs a minimal cytotoxicity was ob-served in WI-38 cells (Fig. 7).
Cell cycle analysis
To assess the effects on the cell cycle, 3T3-L1 andWI-38 fibroblastswere incubated with the NPs for 24 or 48 h. Although the results pres-ented in Fig. 7 indicate that both CL and CL-X silica NPs did not exertany significant effect on cycle distribution, there were indications thatthey increased the number of 3T3-L1 cells in the S phase (Fig. 8A) andthe number of WI-38 cells in the G1 phase (Fig. 8B). In order to ruleout effects from cell confluence, the conditions (cell number and in-cubation time) were carefully adjusted in order not to exceed 70%confluence in the unexposed control cells. In this way we assuredthat the cells were in the exponential phase of growth and that theeffect observed resulted from exposure to the silica NPs.
Influence of buthionine sulfoximine (BSO) and N-acetyl-L-cysteine (NAC)on Ludox CL or CL-X effects on the fibroblasts
Assuming involvement of oxidative stress in mediating thecytotoxicity of the two different silica NPs, the effect of glutathionedepletion was investigated. To this end, BSO, a synthetic amino acidthat irreversibly inhibits γ-glutamylcysteine synthetase, was used.BSO was applied in the highest concentration which did not inducesignificant growth inhibition of the cells (10 μM and 2.5 μM for 3T3-L1 and WI-38 cells, respectively). BSO addition did not influence theeffects of the CL silica NPs on 3T3-L1 cells. However, it did enhancethe cytotoxicity of the CL-X silica NPs considerably (Figs. 9 and 10)when low non-toxic concentrations (below 150 μg/ml) of the CL-XNPs were used. BSO showed a lower ability to modulate silica nano-particle effects in WI-38 cells in comparison to 3T3-L1 fibroblasts. At2.5 μM it significantly decreased the viability of WI-38 cells onlyafter exposure to the highest CL silica nanoparticle concentrationand non-significantly after exposure to high CL-X silica nanoparticleconcentrations.
0
20
40
60
80
100
120
140
Con
trol
Lys
Con
trol
Sup 800
600
400
200
100 40 800
600
400
200
100 40 800
600
400
250
200
150
100 40 800
600
400
250
200
150
100 40
Ludox CL Ludox CL dialysed Ludox CL-X Ludox CL-X dialysed
LD
H a
ctiv
ity
[% c
ontr
ol]
Concentration [µµg/ml]
Lysates Supernatants IC50 (supernatants):CL-X = 351 µg/ml (95% CI: 301-409)CL-X dialysed= 284 µg/ml (95% CI: 256-315)
IC50 (supernatants):CL-X = 351 µg/ml (95% CI: 301-409)CL-X dialysed= 284 µg/ml (95% CI: 256-315)
CL
-Xd
CL
d
Fig. 3. Cytotoxicity of Ludox CL and CL-X silica NPs (dialyzed and non-dialyzed) on 3T3-L1 mouse fibroblasts. The cells were exposed for 72 h and the LDH activity was measured insupernatants and cell lysates (N=3). CLd or CL-Xd — denotes water after dialysis of respective silica samples. The final concentration of each test sample was adjusted to 10% v/v.
94 M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
Measurements of concentrations of reduced glutathione (GSH) inthe cells exposed to BSO (two independent experiments) showed itsconsiderable inhibitory effect at the concentrations of 10 μM for 3T3-L1cells (32.3±4.5 nM GSH/μg protein in control non-exposed cells vs.5.8±0.9 nM/μg protein in BSO exposed cells after 24 h and 25.0±4.9nM GSH/μg protein in control cells vs. 5.7±4.1 nM GSH/μg protein inBSO exposed cells after 72 h) and 2.5 μM for WI-38 cells (13.6±1.8nM GSH/μg protein in control non-exposed cells vs. 6.6±1.1 nM/μgprotein in BSO exposed cells after 24 h and 12.2±2.3 nM GSH/μg
0
20
40
60
80
100
120
140
Con
trol
Lys
Con
trol
Sup 80
0
600
400
200
100 40 800
600
400
200
100 40
Ludox CL Ludox CL dialysed
LD
H a
ctiv
ity
[% c
ontr
ol]
Concentr
CL
d
Fig. 4. Cytotoxicity of Ludox CL and CL-X silica NPs (dialyzed and non-dialyzed) onWI-38 huin supernatants and cell lysates (N=3). CLd or CL-Xd— denotes water after dialysis of respe
protein in control cells vs. 2.6±1.5 nM GSH/μg protein in BSO exposedcells after 72 h).
To assess influence of NAC, an anti-oxidant agent known to increaseintracellular levels of GSH, 3T3-L1 and WI-38 fibroblasts were exposedto combination of NAC and Ludox CL-X silica (exerting a stronger bio-logic effects) for 72 h. NAC was used at the highest non-cytostatic/cytotoxic (WST-1 assay) concentrations after 72 h of incubation, i.e.at 50 μM for 3T3-L1 cells or 250 μM forWI-38 cells. The results indicat-ed no or a weak (non-significant) protective effect of NAC against the
800
600
400
250
200
150
100 40 800
600
400
250
200
150
100 40
Ludox CL-X Ludox CL-X dialysed
ation [µµg/ml]
Lysates SupernatantsC
L-X
d
man fibroblast cells. The cells were exposed for 72 h and the LDH activity was measuredctive silica samples. The final concentration of each test sample was adjusted to 10% v/v.
0
50
100
100 200 300 400 500 600 700 800
150
200
0
Num
ber
of c
ells
[%
con
trol
]
Concentration [µµg/ml]
Ludox CL
Ludox CL-X
Fig. 5. The effect of Ludox CL and CL-X silica NPs (dialyzed) on 3T3-L1 mouse fibroblastsproliferation. The cells were exposed for 72 h in 6-well microplates (in 2 ml/well). Aftertrypsinization viable cells were counted in hemocytometer (Trypan blue exclusion test)(N=3).
0,0
0,2
0,4
0,6
0,8
1,0
400 100 40 CLd 200
Ludox CL [ug/ml]
Surv
ivin
g F
ract
ion
0
20
40
60
80
100
120
140B
400 200 100 40 20 10 200
A
Ludox CL [ug/ml] CLd
Via
bilit
y [%
con
trol
]
p<0.007
p<0.000
Fig. 6. Long-termcytotoxicity testing of LudoxCL and CL-X silica on3T3-L1fibroblasts. A. Surviv(N=2). The cellswere seededon 60-mmplates at 100 cells/plate (in 3 ml ofmedium) and on thfor fresh cDMEMmedium without silica and the cells were incubated for subsequent 4 days (mCells plated onto 96-well microplates (50 cells/well in 100 μl) were treatedwith silica for 7 daytive silica samples used at the final concentration of 10% v/v. Statistical significance was calcul
95M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
cytotoxicity of silica used at higher concentrations (§8 in the Supple-mentary information).
Discussion
The data presented on the effects of two different types of silicaNPs with sizes in the same order of magnitude, show that it is notpossible to refer to silica NPs of a specific size as being toxic or non-toxic. A complete description of the particle, including source, surfacecharge and coating (if present), as well as the dispersing agent, andthe size with its distribution under the assay conditions is needed inorder to accurately describe a specific silica (or indeed other) nano-particle. Significant differences in the cytotoxicity of two differentSigma Ludox silica NPs were observed, with the most significanteffects most likely due to the difference in surface coatings of theNPs, and likely also different stability. When suspended in waterone silica nanoparticle had a negative zeta potential and the otherhad a positive one, because of the presence of the alumina coating,
150 100 40 CL-Xd 10% H2O
Ludox CL-X [ug/ml]
150 100 40 20 10
Ludox CL-X [ug/ml] CL-Xd 10%H2O
02
p<0.04
p<0.00001
p<0.007
p<0.0001
ing fraction of 3T3-L1 cells exposed to dialyzedCLor CL-X silica in clonogenic survival assaye next day exposed to silica samples for 72 h.On the 4th day supernatantswere exchangedean Plating Efficiency for 10% H2O=48±3.5%); B. The 7 day-exposure cytotoxicity test.
s without medium renewal (N=4). CLd or CL-Xd— denotes water after dialysis of respec-ated between a given group and control with 10% H2O.
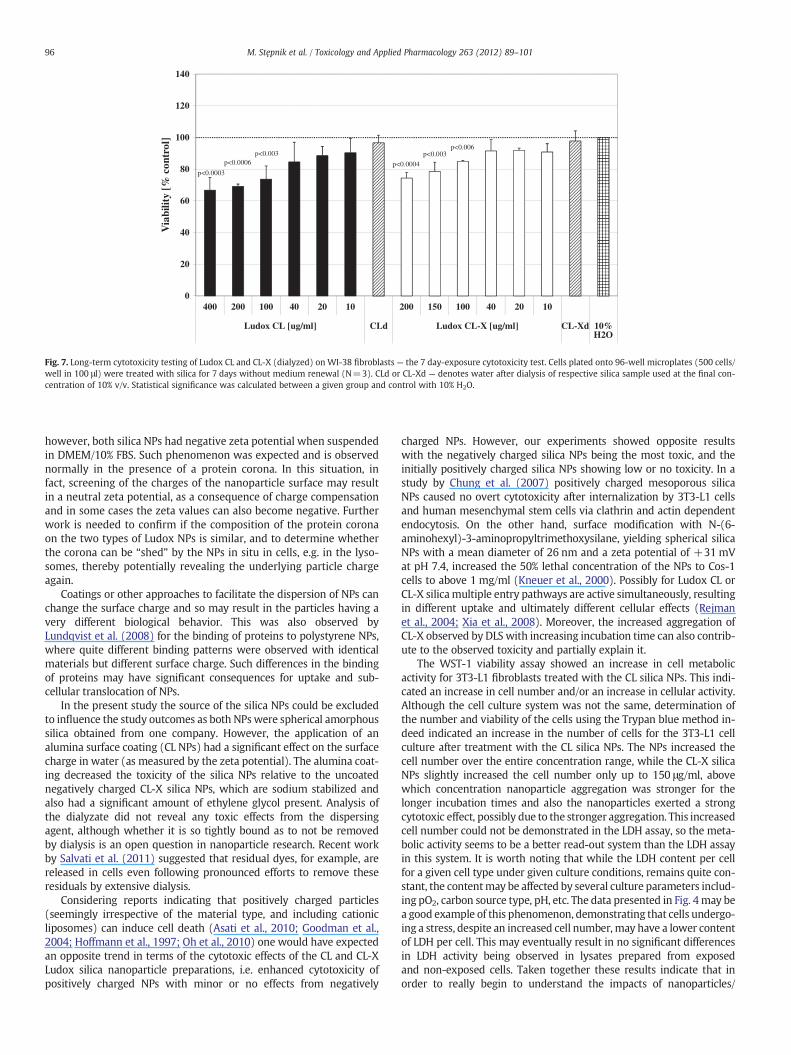
0
20
40
60
80
100
120
140
400 200 100 200 150 100 40 20 1040 20 10
Ludox CL [ug/ml] CLd Ludox CL-X [ug/ml] CL-Xd 10%H2O
Via
bilit
y [%
con
trol
]p<0.0003
p<0.0006p<0.003
p<0.0004
p<0.003p<0.006
Fig. 7. Long-term cytotoxicity testing of Ludox CL and CL-X (dialyzed) on WI-38 fibroblasts — the 7 day-exposure cytotoxicity test. Cells plated onto 96-well microplates (500 cells/well in 100 μl) were treated with silica for 7 days without medium renewal (N=3). CLd or CL-Xd — denotes water after dialysis of respective silica sample used at the final con-centration of 10% v/v. Statistical significance was calculated between a given group and control with 10% H2O.
96 M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
however, both silica NPs had negative zeta potential when suspendedin DMEM/10% FBS. Such phenomenon was expected and is observednormally in the presence of a protein corona. In this situation, infact, screening of the charges of the nanoparticle surface may resultin a neutral zeta potential, as a consequence of charge compensationand in some cases the zeta values can also become negative. Furtherwork is needed to confirm if the composition of the protein coronaon the two types of Ludox NPs is similar, and to determine whetherthe corona can be “shed” by the NPs in situ in cells, e.g. in the lyso-somes, thereby potentially revealing the underlying particle chargeagain.
Coatings or other approaches to facilitate the dispersion of NPs canchange the surface charge and so may result in the particles having avery different biological behavior. This was also observed byLundqvist et al. (2008) for the binding of proteins to polystyrene NPs,where quite different binding patterns were observed with identicalmaterials but different surface charge. Such differences in the bindingof proteins may have significant consequences for uptake and sub-cellular translocation of NPs.
In the present study the source of the silica NPs could be excludedto influence the study outcomes as both NPswere spherical amorphoussilica obtained from one company. However, the application of analumina surface coating (CL NPs) had a significant effect on the surfacecharge in water (as measured by the zeta potential). The alumina coat-ing decreased the toxicity of the silica NPs relative to the uncoatednegatively charged CL-X silica NPs, which are sodium stabilized andalso had a significant amount of ethylene glycol present. Analysis ofthe dialyzate did not reveal any toxic effects from the dispersingagent, although whether it is so tightly bound as to not be removedby dialysis is an open question in nanoparticle research. Recent workby Salvati et al. (2011) suggested that residual dyes, for example, arereleased in cells even following pronounced efforts to remove theseresiduals by extensive dialysis.
Considering reports indicating that positively charged particles(seemingly irrespective of the material type, and including cationicliposomes) can induce cell death (Asati et al., 2010; Goodman et al.,2004; Hoffmann et al., 1997; Oh et al., 2010) one would have expectedan opposite trend in terms of the cytotoxic effects of the CL and CL-XLudox silica nanoparticle preparations, i.e. enhanced cytotoxicity ofpositively charged NPs with minor or no effects from negatively
charged NPs. However, our experiments showed opposite resultswith the negatively charged silica NPs being the most toxic, and theinitially positively charged silica NPs showing low or no toxicity. In astudy by Chung et al. (2007) positively charged mesoporous silicaNPs caused no overt cytotoxicity after internalization by 3T3-L1 cellsand human mesenchymal stem cells via clathrin and actin dependentendocytosis. On the other hand, surface modification with N-(6-aminohexyl)-3-aminopropyltrimethoxysilane, yielding spherical silicaNPs with a mean diameter of 26 nm and a zeta potential of +31 mVat pH 7.4, increased the 50% lethal concentration of the NPs to Cos-1cells to above 1 mg/ml (Kneuer et al., 2000). Possibly for Ludox CL orCL-X silicamultiple entry pathways are active simultaneously, resultingin different uptake and ultimately different cellular effects (Rejmanet al., 2004; Xia et al., 2008). Moreover, the increased aggregation ofCL-X observed by DLS with increasing incubation time can also contrib-ute to the observed toxicity and partially explain it.
The WST-1 viability assay showed an increase in cell metabolicactivity for 3T3-L1 fibroblasts treated with the CL silica NPs. This indi-cated an increase in cell number and/or an increase in cellular activity.Although the cell culture system was not the same, determination ofthe number and viability of the cells using the Trypan blue method in-deed indicated an increase in the number of cells for the 3T3-L1 cellculture after treatment with the CL silica NPs. The NPs increased thecell number over the entire concentration range, while the CL-X silicaNPs slightly increased the cell number only up to 150 μg/ml, abovewhich concentration nanoparticle aggregation was stronger for thelonger incubation times and also the nanoparticles exerted a strongcytotoxic effect, possibly due to the stronger aggregation. This increasedcell number could not be demonstrated in the LDH assay, so the meta-bolic activity seems to be a better read-out system than the LDH assayin this system. It is worth noting that while the LDH content per cellfor a given cell type under given culture conditions, remains quite con-stant, the contentmay be affected by several culture parameters includ-ing pO2, carbon source type, pH, etc. The data presented in Fig. 4may bea good example of this phenomenon, demonstrating that cells undergo-ing a stress, despite an increased cell number, may have a lower contentof LDH per cell. This may eventually result in no significant differencesin LDH activity being observed in lysates prepared from exposedand non-exposed cells. Taken together these results indicate that inorder to really begin to understand the impacts of nanoparticles/
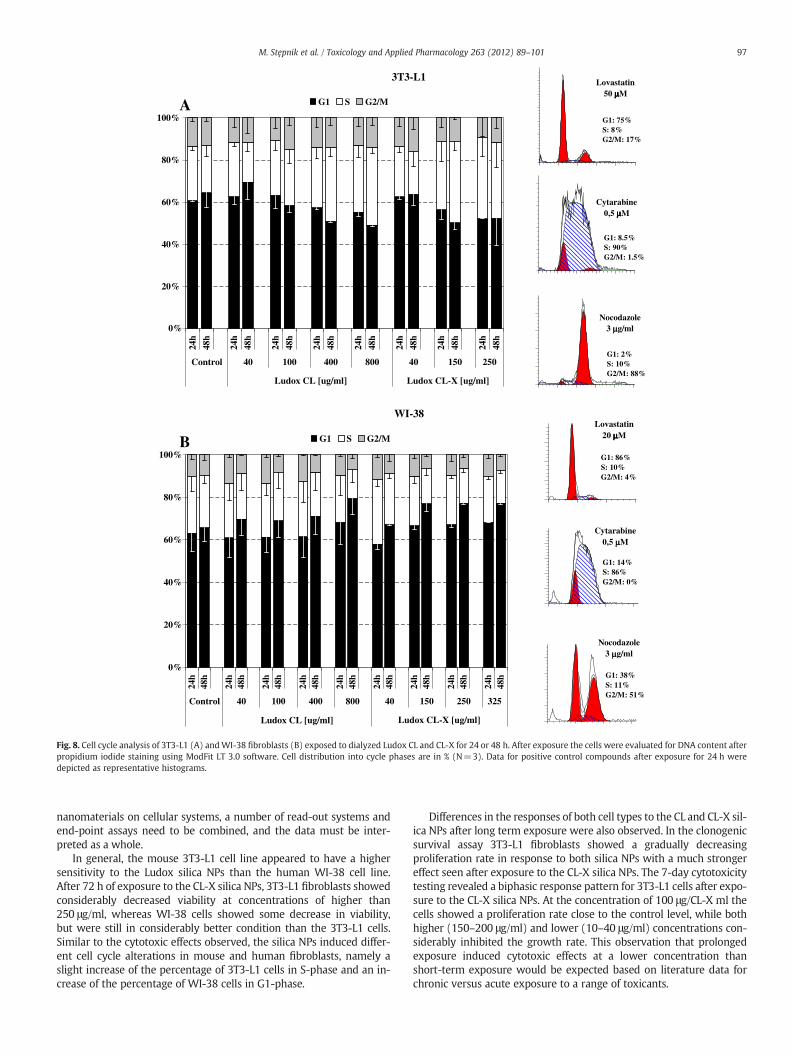
0%
20%
40%
60%
80%
100%
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
Control 40 100 400 800 40 150 250
Ludox CL [ug/ml] Ludox CL-X [ug/ml]
Ludox CL [ug/ml] Ludox CL-X [ug/ml]
G1 S G2/M
3T3-L1
Cytarabine0,5 µµµµM
Lovastatin50 µµµµM
G1: 8.5%S: 90%G2/M: 1.5%
G1: 75%S: 8%G2/M: 17%
G1: 2%S: 10%G2/M: 88%
Nocodazole3 µµg/ml
0%
20%
40%
60%
80%
100%B
A
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
24h
48h
Control 40 100 400 800 40 150 250 325
G1 S G2/M
WI-38
Cytarabine0,5 µµM
Lovastatin20 µµµµM
G1: 14%S: 86%G2/M: 0%
G1: 86%S: 10%G2/M: 4%
G1: 38%S: 11%G2/M: 51%
Nocodazole3 µµg/ml
Fig. 8. Cell cycle analysis of 3T3-L1 (A) andWI-38 fibroblasts (B) exposed to dialyzed Ludox CL and CL-X for 24 or 48 h. After exposure the cells were evaluated for DNA content afterpropidium iodide staining using ModFit LT 3.0 software. Cell distribution into cycle phases are in % (N=3). Data for positive control compounds after exposure for 24 h weredepicted as representative histograms.
97M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
nanomaterials on cellular systems, a number of read-out systems andend-point assays need to be combined, and the data must be inter-preted as a whole.
In general, the mouse 3T3-L1 cell line appeared to have a highersensitivity to the Ludox silica NPs than the human WI-38 cell line.After 72 h of exposure to the CL-X silica NPs, 3T3-L1 fibroblasts showedconsiderably decreased viability at concentrations of higher than250 μg/ml, whereas WI-38 cells showed some decrease in viability,but were still in considerably better condition than the 3T3-L1 cells.Similar to the cytotoxic effects observed, the silica NPs induced differ-ent cell cycle alterations in mouse and human fibroblasts, namely aslight increase of the percentage of 3T3-L1 cells in S-phase and an in-crease of the percentage of WI-38 cells in G1-phase.
Differences in the responses of both cell types to the CL and CL-X sil-ica NPs after long term exposure were also observed. In the clonogenicsurvival assay 3T3-L1 fibroblasts showed a gradually decreasingproliferation rate in response to both silica NPs with a much strongereffect seen after exposure to the CL-X silica NPs. The 7-day cytotoxicitytesting revealed a biphasic response pattern for 3T3-L1 cells after expo-sure to the CL-X silica NPs. At the concentration of 100 μg/CL-X ml thecells showed a proliferation rate close to the control level, while bothhigher (150–200 μg/ml) and lower (10–40 μg/ml) concentrations con-siderably inhibited the growth rate. This observation that prolongedexposure induced cytotoxic effects at a lower concentration thanshort-term exposure would be expected based on literature data forchronic versus acute exposure to a range of toxicants.
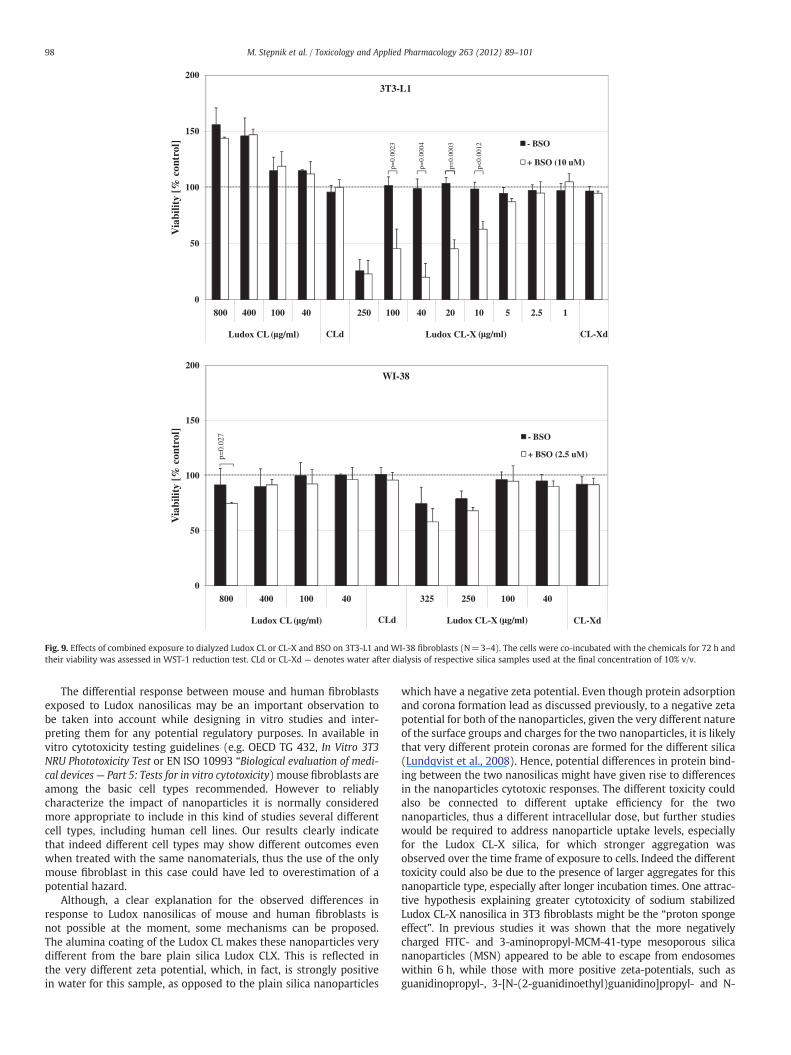
0
50
100
150
200
800 400 100 40
800 400 100 40 325 250 100 40
250 100 40 20 10 2.55 1
Ludox CL-X CLd CL-Xd
Via
bilit
y [%
con
trol
] - BSO
+ BSO (10 uM)
3T3-L1
p=0.
0023
p=0.
0004
p=0.
0003
p<0.
0012
0
50
100
150
200
Via
bilit
y [%
con
trol
]
- BSO
+ BSO (2.5 uM)
WI-38
p=0.
027
( g/ml)Ludox CL ( g/ml)
Ludox CL-X CLd CL-Xd( g/ml)Ludox CL ( g/ml)
Fig. 9. Effects of combined exposure to dialyzed Ludox CL or CL-X and BSO on 3T3-L1 and WI-38 fibroblasts (N=3–4). The cells were co-incubated with the chemicals for 72 h andtheir viability was assessed in WST-1 reduction test. CLd or CL-Xd — denotes water after dialysis of respective silica samples used at the final concentration of 10% v/v.
98 M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
The differential response between mouse and human fibroblastsexposed to Ludox nanosilicas may be an important observation tobe taken into account while designing in vitro studies and inter-preting them for any potential regulatory purposes. In available invitro cytotoxicity testing guidelines (e.g. OECD TG 432, In Vitro 3T3NRU Phototoxicity Test or EN ISO 10993 “Biological evaluation of medi-cal devices— Part 5: Tests for in vitro cytotoxicity) mouse fibroblasts areamong the basic cell types recommended. However to reliablycharacterize the impact of nanoparticles it is normally consideredmore appropriate to include in this kind of studies several differentcell types, including human cell lines. Our results clearly indicatethat indeed different cell types may show different outcomes evenwhen treated with the same nanomaterials, thus the use of the onlymouse fibroblast in this case could have led to overestimation of apotential hazard.
Although, a clear explanation for the observed differences inresponse to Ludox nanosilicas of mouse and human fibroblasts isnot possible at the moment, some mechanisms can be proposed.The alumina coating of the Ludox CL makes these nanoparticles verydifferent from the bare plain silica Ludox CLX. This is reflected inthe very different zeta potential, which, in fact, is strongly positivein water for this sample, as opposed to the plain silica nanoparticles
which have a negative zeta potential. Even though protein adsorptionand corona formation lead as discussed previously, to a negative zetapotential for both of the nanoparticles, given the very different natureof the surface groups and charges for the two nanoparticles, it is likelythat very different protein coronas are formed for the different silica(Lundqvist et al., 2008). Hence, potential differences in protein bind-ing between the two nanosilicas might have given rise to differencesin the nanoparticles cytotoxic responses. The different toxicity couldalso be connected to different uptake efficiency for the twonanoparticles, thus a different intracellular dose, but further studieswould be required to address nanoparticle uptake levels, especiallyfor the Ludox CL-X silica, for which stronger aggregation wasobserved over the time frame of exposure to cells. Indeed the differenttoxicity could also be due to the presence of larger aggregates for thisnanoparticle type, especially after longer incubation times. One attrac-tive hypothesis explaining greater cytotoxicity of sodium stabilizedLudox CL-X nanosilica in 3T3 fibroblasts might be the “proton spongeeffect”. In previous studies it was shown that the more negativelycharged FITC- and 3-aminopropyl-MCM-41-type mesoporous silicananoparticles (MSN) appeared to be able to escape from endosomeswithin 6 h, while those with more positive zeta-potentials, such asguanidinopropyl-, 3-[N-(2-guanidinoethyl)guanidino]propyl- and N-
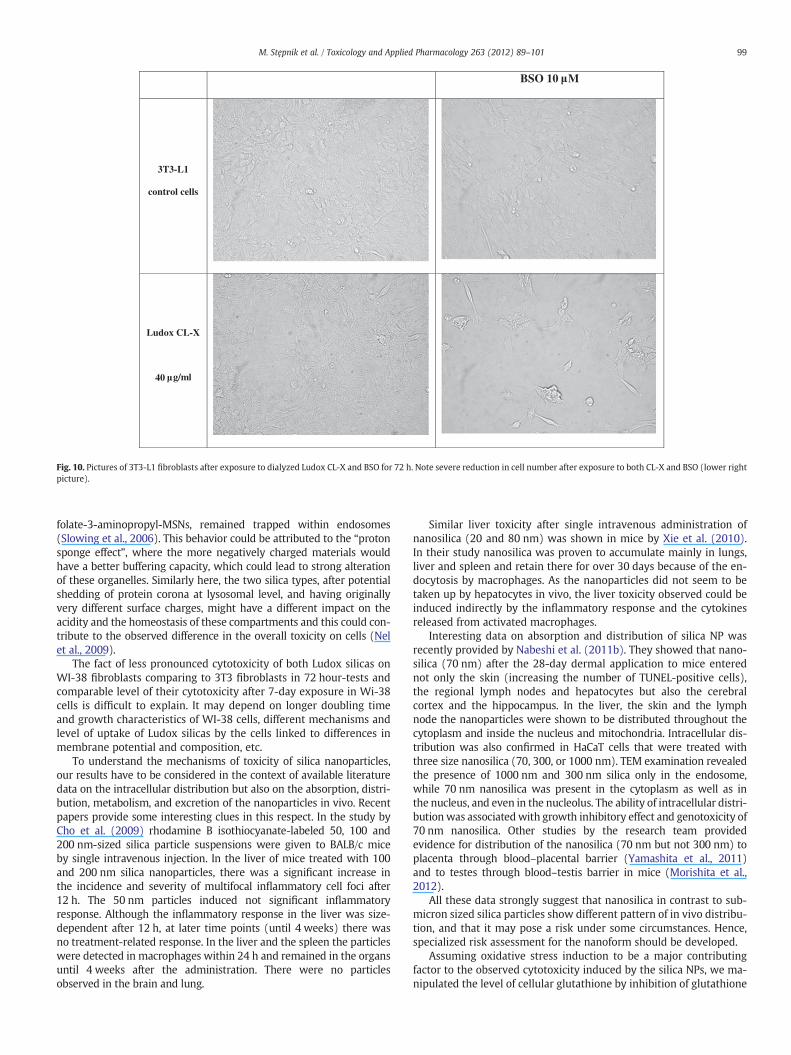
BSO 10 M
3T3-L1
control cells
Ludox CL-X
40 g/ml
Fig. 10. Pictures of 3T3-L1 fibroblasts after exposure to dialyzed Ludox CL-X and BSO for 72 h. Note severe reduction in cell number after exposure to both CL-X and BSO (lower rightpicture).
99M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
folate-3-aminopropyl-MSNs, remained trapped within endosomes(Slowing et al., 2006). This behavior could be attributed to the “protonsponge effect”, where the more negatively charged materials wouldhave a better buffering capacity, which could lead to strong alterationof these organelles. Similarly here, the two silica types, after potentialshedding of protein corona at lysosomal level, and having originallyvery different surface charges, might have a different impact on theacidity and the homeostasis of these compartments and this could con-tribute to the observed difference in the overall toxicity on cells (Nelet al., 2009).
The fact of less pronounced cytotoxicity of both Ludox silicas onWI-38 fibroblasts comparing to 3T3 fibroblasts in 72 hour-tests andcomparable level of their cytotoxicity after 7-day exposure in Wi-38cells is difficult to explain. It may depend on longer doubling timeand growth characteristics of WI-38 cells, different mechanisms andlevel of uptake of Ludox silicas by the cells linked to differences inmembrane potential and composition, etc.
To understand the mechanisms of toxicity of silica nanoparticles,our results have to be considered in the context of available literaturedata on the intracellular distribution but also on the absorption, distri-bution, metabolism, and excretion of the nanoparticles in vivo. Recentpapers provide some interesting clues in this respect. In the study byCho et al. (2009) rhodamine B isothiocyanate-labeled 50, 100 and200 nm-sized silica particle suspensions were given to BALB/c miceby single intravenous injection. In the liver of mice treated with 100and 200 nm silica nanoparticles, there was a significant increase inthe incidence and severity of multifocal inflammatory cell foci after12 h. The 50 nm particles induced not significant inflammatoryresponse. Although the inflammatory response in the liver was size-dependent after 12 h, at later time points (until 4 weeks) there wasno treatment-related response. In the liver and the spleen the particleswere detected in macrophages within 24 h and remained in the organsuntil 4 weeks after the administration. There were no particlesobserved in the brain and lung.
Similar liver toxicity after single intravenous administration ofnanosilica (20 and 80 nm) was shown in mice by Xie et al. (2010).In their study nanosilica was proven to accumulate mainly in lungs,liver and spleen and retain there for over 30 days because of the en-docytosis by macrophages. As the nanoparticles did not seem to betaken up by hepatocytes in vivo, the liver toxicity observed could beinduced indirectly by the inflammatory response and the cytokinesreleased from activated macrophages.
Interesting data on absorption and distribution of silica NP wasrecently provided by Nabeshi et al. (2011b). They showed that nano-silica (70 nm) after the 28-day dermal application to mice enterednot only the skin (increasing the number of TUNEL-positive cells),the regional lymph nodes and hepatocytes but also the cerebralcortex and the hippocampus. In the liver, the skin and the lymphnode the nanoparticles were shown to be distributed throughout thecytoplasm and inside the nucleus and mitochondria. Intracellular dis-tribution was also confirmed in HaCaT cells that were treated withthree size nanosilica (70, 300, or 1000 nm). TEM examination revealedthe presence of 1000 nm and 300 nm silica only in the endosome,while 70 nm nanosilica was present in the cytoplasm as well as inthe nucleus, and even in the nucleolus. The ability of intracellular distri-bution was associatedwith growth inhibitory effect and genotoxicity of70 nm nanosilica. Other studies by the research team providedevidence for distribution of the nanosilica (70 nm but not 300 nm) toplacenta through blood–placental barrier (Yamashita et al., 2011)and to testes through blood–testis barrier in mice (Morishita et al.,2012).
All these data strongly suggest that nanosilica in contrast to sub-micron sized silica particles show different pattern of in vivo distribu-tion, and that it may pose a risk under some circumstances. Hence,specialized risk assessment for the nanoform should be developed.
Assuming oxidative stress induction to be a major contributingfactor to the observed cytotoxicity induced by the silica NPs, we ma-nipulated the level of cellular glutathione by inhibition of glutathione

100 M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
synthetase via BSO. It was assumed that if silica NPs are able togenerate reactive oxygen species causing the observed cytotoxiceffects, the combined exposure to silica NPs and BSO would increasesuch effects. Interestingly in 3T3-L1 fibroblasts such combined expo-sure resulted in induction of considerable cytotoxicity already at theCL-X concentrations of 10 μg/ml. However, no significant protectiveeffect of N-acetyl-L-cysteine on the cytotoxic effect of Ludox CL-Xwas observed.
Oxidative stress has often been reported as toxic mechanism ofnanosilica-induced cytotoxicity in various cell types, including myo-cardial cells (Ye et al., 2010a), HepG2 human hepatic cells (Lu et al.,2011; Ye et al., 2010b), HEK293 human embryonic kidney cells(Wang et al., 2009), MCF-7 human breast cancer cells (Shi et al.,2012), or HaCaT human keratinocytes (Gong et al., 2012). However,the molecular mechanisms of nanosilica-induced ROS generationand the oxidative stress responding signal transduction pathwaysremain poorly understood. Recently some new concepts trying tocorrelate physico-chemical properties of nanosilica with its biologicaleffects suggested that full hydroxylation of nanosilica surface, withconsequent stable coverage by water molecules, may lead toreduced/inhibited toxic behavior (Gazzano et al., 2012). It wasshown that pyrogenic nanosilicas (Aerosil50, 40 nm and Aerosil300,7 nm) were remarkably more active than the precipitated one(FK320, 15 nm) as to cytotoxicity, ROS production, lipid peroxidation,nitric oxide synthesis and production of tumor necrosis factor-α,when compared both per mass and per unit surface. Silanols densitywas much higher on the precipitated nanosilica comparing with theothers.
There are several mechanisms of cytotoxicity proposed so far fornanosilica which link ROS induction with activation of signal trans-duction pathways ultimately leading to cell death. In BEAS-2B cellssilica nanoparticles exert toxicity via oxidative stress, which leads tothe induction of heme oxyganase-1 via the nuclear factor-E2-relatedfactor-2 (Nrf-2), as well as the extracellular regulated mitogen-activated protein kinase (ERK-MAP) signaling pathway (Eom andChoi, 2009). In HaCaT cells exposed to 15-nm, 30-nm silica andmicro-sized silica particles down-regulation of peroxiredoxins Prx1and Prx6 as well as thioredoxin can be observed (Yang et al., 2010).Moreover, the molecular chaperone proteins, like cyclophilin andpeptidyl prolyl isomerase (PPIA), were also found to be altered. Theresearch by Ye et al. (2010b) has shown that 21 nm silica colloidsinduced apoptosis in L-02 human hepatic cells, which was correlatedwith ROS-mediated oxidative stress, the activation of p53 and up-regulation of Bax/Bcl-2 ratio. Mitochondria, which are the mainsources of cellular ROS and the basis of the ROS metabolism may bedirectly or indirectly affected by nanosilica leading to mitochondrialmembrane potential collapse and ultimately apoptosis. The followingsilica nanoparticles-mediated signaling pathway for apoptosis relatedevents was proposed by Liu and Sun (2010): exposure to silicananoparticles/ROS production/(JNK/c-Jun phosphorylation)/P53 acti-vation/Bax upregulation/Bcl-2 downregulation/loss of mitochondrialmembrane potential/caspase-3 activation/apoptosis. Such scenariowas further confirmed by Ahmad et al. (2012) and by Sun et al.(2011) in silica nanoparticles treated HepG2 cells.
Conclusions
Although, more and more reports on cytotoxic effects of silica NPsin vitro are published (Chang et al., 2007; Lin et al., 2006; Lison et al.,2008; Napierska et al., 2009), the mechanisms of the toxic effectshave not been clarified so far. In this study several aspects of cellular re-sponses to amorphous silica NPs were studied. The results show that:
— although the CL and CL-X silica NPs show opposite charges whenmeasured in water, they both show negative zeta potential ofcomparable value when measured in DMEM supplemented with
10% FBS as observed often after protein adsorption, as a conse-quence of charge compensation,
— the CL-X silica NPs stabilized with ethylene glycol aggregate at thehigher concentrations over the 72 h exposure and are more cyto-toxic to both human and mouse fibroblasts than the CL silica NPswhich have an alumina coating, probably in connection to theobserved aggregation,
— it is not possible to refer to silica NPs of a specific size being toxicor non-toxic, but instead a complete description of the particle, itssurface coating and/or dispersing agents and size distribution inthe assay medium is needed to accurately describe a specificnanoparticle,
— murine 3T3-L1 fibroblasts are more sensitive to the cytotoxiceffects of the CL-X silica NPs than human WI-38 fibroblasts,
— differences in the responses of both cell types to the CL and CL-Xsilica NPs after long term exposure were also observed using theclonogenic survival assay,
— although the differences are not clear at the moment the resultsindicate the need for further studies, as eventually they mayhave practical usefulness in development of better risk assessmentprocedures,
— induction of oxidative stress may be an important contributingfactor in mediating the cytotoxic effects of CL-X silica NPs in3T3-L1 cells,
— using only a single cytotoxicity assay may provide inaccurate re-sults, for a variety of reasons, and thus in order to draw reliableconclusions regarding the impact of NPs on biological entities, apanel of tests measuring different endpoints is required,
— in interpretation of cytotoxicity results additional control testshave to carried out in order to rule out possible interference ofNPs with the detection systems being utilized (e.g. non-specificreaction of NPs with WST-1 or LDH reagents, stability of LDH re-leased from cells after prolonged incubation in culture medium,potential inhibition of its activity by NPs, etc.).
Conflict of interest
The authors report no conflict of interests. The authors are entirelyresponsible for the content and writing of the manuscript.
Acknowledgments
This work was supported by EU project within the Sixth Frame-work Program, Acronym: NanoInteract (NMP4-CT-2006-033231).
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.taap.2012.06.002.
References
Ahmad, J., Ahamed, M., Akhtar, M.J., Alrokayan, S.A., Siddiqui, M.A., Musarrat, J., Al-Khedhairy, A.A., 2012. Apoptosis induction by silica nanoparticles mediatedthrough reactive oxygen species in human liver cell line HepG2. Toxicol. Appl.Pharmacol. 259, 160–168.
Arts, J.H.E., Muijser, H., Duistermaat, E., Junker, K., Kuper, C.F., 2007. Five-day inhalationtoxicity study of three types of synthetic amorphous silicas in Wistar rats andpost-exposure evaluations for up to 3 months. Food Chem. Toxicol. 45, 1856–1867.
Asati, A., Santra, S., Kaittanis, C., Perez, J.M., 2010. Surface-charge-dependent celllocalization and cytotoxicity of cerium oxide nanoparticles. ACS Nano 4,5321–5331.
Barnes, C.A., Elsaesser, A., Arkusz, J., Smok, A., Palus, J., Leśniak, A., Salvati, A., Hanrahan,J.P., Jong, W.H., Dziubałtowska, E., Stępnik, M., Rydzyński, K., McKerr, G., Lynch, I.,Dawson, K.A., Howard, C.V., 2008. Reproducible comet assay of amorphous silicananoparticles detects no genotoxicity. Nano Lett. 8, 3069–3074.
Bradford, M.M., 1976. A rapid and sensitivemethod for the quantitation of microgram quan-tities of protein utilizing the principle of protein–dyebinding. Anal. Biochem. 7, 248–254.
Brown, D.M., Wilson, M.R., MacNee, W., Stone, V., Donaldson, K., 2001. Size-dependentproinflammatory effects of ultrafine polystyrene particles: a role for surface area

101M. Stępnik et al. / Toxicology and Applied Pharmacology 263 (2012) 89–101
and oxidative stress in the enhanced activity of ultrafines. Toxicol. Appl.Pharmacol. 175, 191–199.
Cedervall, T., Lynch, I., Lindman, S., Berggård, T., Thulin, E., Nilsson, H., Dawson, K.A.,Linse, S., 2007. Understanding the nanoparticle-protein corona using methods toquantify exchange rates and affinities of proteins for nanoparticles. Proc. Natl.Acad. Sci. U. S. A. 104 (7), 2050–2055.
Chang, J.S., Chang, K.L., Hwang, D.F., Kong, Z.L., 2007. In vitro cytotoxicitiy of silicananoparticles at high concentrations strongly depends on the metabolic activitytype of the cell line. Environ. Sci. Technol. 41, 2064–2068.
Cho, W.S., Choi, M., Han, B.S., Cho, M., Oh, J., Park, K., Kim, S.J., Kim, S.H., Jeong, J., 2007.Inflammatory mediators induced by intratracheal instillation of ultrafine amor-phous silica particles. Toxicol. Lett. 175, 24–33.
Cho, M., Cho, W.S., Choi, M., Kim, S.J., Han, B.S., Kim, S.H., Kim, H.O., Sheen, Y.Y., Jeong, J.,2009. The impact of size on tissue distribution and elimination by single intrave-nous injection of silica nanoparticles. Toxicol. Lett. 189 (3), 177–183.
Chung, T.H., Wu, S.H., Yao, M., Lu, C.W., Lin, Y.S., Hung, Y., Mou, C.Y., Chen, Y.C., Huang,D.M., 2007. The effect of surface charge on the uptake and biological function ofmesoporous silica nanoparticles in 3T3-L1 cells and human mesenchymal stemcells. Biomaterials 28, 2959–2966.
Drescher, D., Orts-Gil, G., Laube, G., Natte, K., Veh, R.W., Österle, W., Kneipp, J., 2011.Toxicity of amorphous silica nanoparticles on eukaryotic cell model is determinedby particle agglomeration and serum protein adsorption effects. Anal. Bioanal.Chem. 400 (5), 1367–1373.
Eom, H.J., Choi, J., 2009. Oxidative stress of silica nanoparticles in human bronchialepithelial cell, Beas-2B. Toxicol. In Vitro 23, 1326–1332.
Eom, H.J., Choi, J., 2011. SiO(2) nanoparticles induced cytotoxicity by oxidative stress inhuman bronchial epithelial cell, Beas-2B. Environ. Health Toxicol. 26, e2011013(Epub 2011 Sep 8).
Fukumori, Y., Takada, K., Takeuchi, H., 2007. Nanoporous and nanosize materials fordrug delivery systems. In: Kumar, C. (Ed.), Nanomaterials for Medical Diagnosisand Therapy. Wiley-VCH Verlag GmbH&Co. KGaA, Weinheim, pp. 255–309.
Gazzano, E., Ghiazza, M., Polimeni, M., Bolis, V., Fenoglio, I., Attanasio, A., Mazzucco, G.,Fubini, B., Ghigo, D., 2012. Physico-chemical determinants in the cellular responsesto nanostructured amorphous silicas. Toxicol. Sci. (Apr 5. [Electronic publicationahead of print]. PMID: 22491428.).
Gehrke, H., Frühmesser, A., Pelka, J., Esselen, M., Hecht, L.L., Blank, H., Schuchmann,H.P., Gerthsen, D., Marquardt, C., Diabaté, S., Weiss, C., Marko, D., 2012. In vitrotoxicity of amorphous silica nanoparticles in human colon carcinoma cells.Nanotoxicology (Jan 20. [Electronic publication ahead of print]).
Gong, C., Tao, G., Yang, L., Liu, J., He, H., Zhuang, Z., 2012. The role of reactive oxygenspecies in silicon dioxide nanoparticle-induced cytotoxicity and DNA damage inHaCaT cells. Mol. Biol. Rep. 39, 4915–4925.
Goodman, C.,McCusker, C., Yilmaz, T., Rotello, V., 2004. Toxicity of goldnanoparticles func-tionalized with cationic and anionic side chains. Bioconjug. Chem. 15, 897–900.
Hoffmann, F., Cinatl, J., Kabickova, H., Kreuter, J., Stieneker, F., 1997. Preparation,characterization and cytotoxicity of methylmethacrylate copolymer nanoparticleswith a permanent positive surface charge. Int. J. Pharm. 157, 189–198.
IARC, 1997. Silica, some silicates, coal dust and para-aramid fibrils. IARC Monographson the Evaluation of Carcinogenic Risks to Humans, 68, pp. 41–242.
Johnston, C.J., Driscoll, K.E., Finkelstein, J.N., Baggs, R., O'Reilly, M.A., Carter, J., Gelein, R.,Oberdorster, G., 2000. Pulmonary chemokine and mutagenic responses in rats aftersubchronic inhalation of amorphous and crystalline silica. Toxicol. Sci. 56, 405–413.
Kaewamatawong, T., Kawamura, N., Okajima, M., Sawada, M., Morita, T., Shimada, A.,2005. Acute pulmonary toxicity caused by exposure to colloidal silica: particlesize dependent pathological changes in mice. Toxicol. Pathol. 33, 743–749.
Kneuer, C., Sameti, M., Bakowsky, U., Schiestel, T., Schirra, H., Schmidt, H., Lehr, C.M.,2000. A nonviral DNA delivery system based on surface modified silica-nanoparticles can efficiently transfect cells in vitro. Bioconjug. Chem. 11, 926–932.
Kumar, R., Roy, I., Ohulchanskyy, T.Y., Goswami, L.N., Bonoiu, A.C., Bergey, E.J.,Tramposch, K.M., Maitra, A., Prasad, P.N., 2008. Covalently dye-linked, surface-controlled, and bioconjugated organically modified silica nanoparticles as targetedprobes for optical imaging. ACS Nano 2, 449–456.
Lin, W., Huang, Y.W., Zhou, X.D., Ma, Y., 2006. In vitro toxicity of silica nanoparticles inhuman lung cancer cells. Toxicol. Appl. Pharmacol. 217, 252–259.
Liong, M., Lu, J., Kovochich, M., Xia, T., Ruehm, S.G., Nel, A.E., Tamanoi, F., Zink, J.I., 2008.Multifunctional inorganic nanoparticles for imaging, targeting, and drug delivery.ACS Nano 2, 889–896.
Lison, D., Thomassen, L.C., Rabolli, V., Gonzalez, L., Napierska, D., Seo, J.W., Kirsch-Volders, M., Hoet, P., Kirschhock, C.E., Martens, J.A., 2008. Nominal and effectivedosimetry of silica nanoparticles in cytotoxicity assays. Toxicol. Sci. 104, 155–162.
Liu, X., Sun, J., 2010. Endothelial cells dysfunction induced by silica nanoparticlesthrough oxidative stress via JNK/P53 and NF-κB pathways. Biomaterials 31,8198–8209.
Lu, X., Qian, J., Zhou, H., Gan, Q., Tang, W., Lu, J., Yuan, Y., Liu, C., 2011. In vitro cytotoxicityand induction of apoptosis by silica nanoparticles in human HepG2 hepatoma cells.Int. J. Nanomedicine 6, 1889–1901.
Lundqvist, M., Stigler, J., Elia, G., Lynch, I., Cedervall, T., Dawson, K.A., 2008. Nanoparticlesize and surface properties determine the protein corona with possible implicationsfor biological impacts. Proc. Natl. Acad. Sci. U. S. A. 105, 14265–14270.
McLaughlin, J.K., Chow, W.H., Levy, L.S., 1997. Amorphous silicas: a review of healtheffects from inhalation exposure with particular reference to cancer. J. Toxicol.Environ. Health 50, 553–566.
Merget, R., Bauer, T., Kupper, H.U., Philippou, S., Bauer, H.D., Breitstadt, R., Bruening, T.,2002. Health hazards due to the inhalation of amorphous silica. Arch. Toxicol. 75,625–634.
Morishita, Y., Yoshioka, Y., Satoh, H., Nojiri, N., Nagano, K., Abe, Y., Kamada, H., Tsunoda,S., Nabeshi, H., Yoshikawa, T., Tsutsumi, Y., 2012. Distribution and histologic effectsof intravenously administered amorphous nanosilica particles in the testes of mice.Biochem. Biophys. Res. Commun. 420 (2), 297–301.
Nabeshi, H., Yoshikawa, T., Matsuyama, K., Nakazato, Y., Tochigi, S., Kondoh, S., Hirai, T.,Akase, T., Nagano, K., Abe, Y., Yoshioka, Y., Kamada, H., Itoh, N., Tsunoda, S.,Tsutsumi, Y., 2011a. Amorphous nanosilica induce endocytosis-dependent ROSgeneration and DNA damage in human keratinocytes. Part. Fibre Toxicol. 8, 1–10.
Nabeshi, H., Yoshikawa, T., Matsuyama, K., Nakazato, Y., Matsuo, K., Arimori, A., Isobe,M., Tochigi, S., Kondoh, S., Hirai, T., Akase, T., Yamashita, T., Yamashita, K.,Yoshida, T., Nagano, K., Abe, Y., Yoshioka, Y., Kamada, H., Imazawa, T., Itoh, N.,Nakagawa, S., Mayumi, T., Tsunoda, S., Tsutsumi, Y., 2011b. Systemic distribution,nuclear entry and cytotoxicity of amorphous nanosilica following topical applica-tion. Biomaterials 32, 2713–2724.
Nan, A., Bai, X., Son, S.J., Lee, S.B., Ghandehari, H., 2008. Cellular uptake and cytotoxicityof silica nanotubes. Nano Lett. 8, 2150–2154.
Napierska, D., Thomassen, L.C., Rabolli, V., Lison, D., Gonzalez, L., Kirsch-Volders, M.,Martens, J.A., Hoet, P.H., 2009. Size-dependent cytotoxicity of monodisperse silicananoparticles in human endothelial cells. Small 5, 846–853.
Napierska, D., Thomassen, L.C., Lison, D., Martens, J.A., Hoet, P.H., 2010. The nanosilicahazard: another variable entity. Part. Fibre Toxicol. 7 (1), 39–71.
Nel, A.E., Lutz, M., Velegol, D., Xia, T., Hoek, E.M.V., Somasundaran, P., Klaessig, F.,Castranova, V., Thompson, M., 2009. Understanding biophysicochemical interactionsat the nano–bio interface. Nat. Mater. 8, 543–557.
Oberdörster, G., 2000. Toxicology of ultrafine particles: in vivo studies. Trans. R. Soc.Lond. A 358, 2719–2740.
Oh, W.K., Kim, S., Choi, M., Kim, C., Jeong, Y.S., Cho, B.R., Hahn, J.S., Jang, J., 2010. Cellularuptake, cytotoxicity, and innate immune response of silica–titania hollownanoparticles based on size and surface functionality. ACS Nano 4, 5301–5313.
Rabolli, V., Thomassen, L.C., Uwambayinema, F., Martens, J.A., Lison, D., 2011. The cyto-toxic activity of amorphous silica nanoparticles is mainly influenced by surfacearea and not by aggregation. Toxicol. Lett. 206 (2), 197–203.
Rahman, I., Kode, A., Saibal, K., 2007. Assay for quantitative determination of glutathioneand glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 1,3159–3165.
Rejman, J., Oberle, V., Zuhorn, I.S., Hoekstra, D., 2004. Size-dependent internalization ofparticles via the pathways of clathrin- and caveolae-mediated endocytosis.Biochem. J. 377, 159–169.
Salvati, A., Aberg, C., Dos Santos, T., Varela, J., Pinto, P., Lynch, I., Dawson, K.A., 2011. Exper-imental and theoretical comparison of intracellular import of polymeric nanoparticlesand small molecules: toward models of uptake kinetics. Nanomedicine 7, 818–826.
Santra, S., Dutta, D., 2007. Nanoparticles for optical imagining of cancer. In: Kumar, C.(Ed.), Nanotechnologies for the Life Sciences. Wiley-VCH Verlag GmbH&Co.KGaA, Weinheim, pp. 44–85.
Shi, J., Karlsson, H.L., Johansson, K., Gogvadze, V., Xiao, L., Li, J., Burks, T., Garcia-Bennett, A., Uheida, A., Muhammed, M., Mathur, S., Morgenstern, R., Kagan, V.E.,Fadeel, B., 2012. Microsomal glutathione transferase 1 protects against toxicityinduced by silica nanoparticles but not by zinc oxide nanoparticles. ACS Nano 6,1925–1938.
Slowing, I., Trewyn, B.G., Lin, W.S.Y., 2006. Effect of surface functionalization of MCM-41-type mesoporous silica nanoparticles on the endocytosis by human cancercells. J. Am. Chem. Soc. 128, 14792–14793.
Sun, L., Li, Y., Liu, X., Jin, M., Zhang, L., Du, N., Guo, C., Huang, P., Sun, Z., 2011. Cytotox-icity and mitochondrial damage caused by silica nanoparticles. Toxicol. In Vitro 25,1619–1629.
Wang, F., Gao, F., Lan, M., Yuan, H., Huang, Y., Liu, J., 2009. Oxidative stress contributesto silica nanoparticle-induced cytotoxicity in human embryonic kidney cells.Toxicol. In Vitro 23, 808–815.
Wörle-Knirsch, J.M., Pulskamp, K., Krug, H.F., 2006. Oops they did it again! Carbonnanotubes hoax scientists in viability assays. Nano Lett. 6, 1261–1268.
Xia, T., Kovochich, M., Liong, M., Zink, J.I., Nel, A.E., 2008. Cationic polystyrenenanosphere toxicity depends on cell-specific endocytic and mitochondrial injurypathways. ACS Nano 2, 85–96.
Xie, G., Sun, J., Zhong, G., Shi, L., Zhang, D., 2010. Biodistribution and toxicity of intra-venously administered silica nanoparticles in mice. Arch. Toxicol. 84 (3),183–190.
Yamashita, K., Yoshioka, Y., Higashisaka, K., Mimura, K., Morishita, Y., Nozaki, M., Yoshida,T., Ogura, T., Nabeshi, H., Nagano, K., Abe, Y., Kamada, H., Monobe, Y., Imazawa, T.,Aoshima, H., Shishido, K., Kawai, Y., Mayumi, T., Tsunoda, S., Itoh, N., Yoshikawa, T.,Yanagihara, I., Saito, S., Tsutsumi, Y., 2011. Silica and titanium dioxide nanoparticlescause pregnancy complications in mice. Nat. Nanotechnol. 6, 321–328.
Yang, X., Liu, J., He, H., Zhou, L., Gong, C., Wang, X., Yang, L., Yuan, J., Huang, H., He, L.,Zhang, B., Zhuang, Z., 2010. SiO2 nanoparticles induce cytotoxicity and proteinexpression alteration in HaCaT cells. Part. Fibre Toxicol. 7, 1–11.
Ye, Y., Liu, J., Chen, M., Sun, L., Lan, M., 2010a. In vitro toxicity of silica nanoparticles inmyocardial cells. Environ. Toxicol. Pharmacol. 29, 131–137.
Ye, Y., Liu, J., Xu, J., Sun, L., Chen, M., Lan, M., 2010b. Nano-SiO2 induces apoptosis viaactivation of p53 and Bax mediated by oxidative stress in human hepatic cellline. Toxicol. In Vitro 24, 751–758.




















