
Myofibroblast activation in colorectal cancer lymph node metastases
Upload
independentCategory
view
0download
0
www.elsevier.com/locate/vetimm
Veterinary Immunology and Immunopathology 114 (2006) 135–148
Cytokine and antibody subclass responses in the intestinal lymph
of sheep during repeated experimental infections with the
nematode parasite Trichostrongylus colubriformis
Anton Pernthaner *, Sally-Ann Cole, Lilian Morrison, Richard Green,Richard J. Shaw, Wayne R. Hein
AgResearch Limited, Wallaceville Animal Research Centre, PO Box 40063, Ward Street, Upper Hutt, New Zealand
Received 22 November 2005; received in revised form 30 June 2006; accepted 3 August 2006
Abstract
The expression of interleukin (IL)-4, IL-5, IL-10, IL-13, TNF-a and IFN-g genes, and parasite-specific IgM, IgG1, IgG2, IgA
and total IgE levels, were monitored daily in intestinal lymph of sheep infected repeatedly with the nematode parasite
Trichostrongylus colubriformis. Host genotype had a significant influence on IL-13 gene activity, with resistant-line (R) sheep
consistently expressing higher levels of mRNA than susceptible-line (S) sheep. Mean gene expression of IL-13, IL-4 and IFN-g did
not differ significantly between the first and second nematode challenge. Field-primed R and S as well as field-primed R and naı̈ve S
sheep had lower mean gene expression of IL-5 and IL-10, respectively, during the second when compared to primary challenge.
Genes for IL-13 and IL-5 were transiently and strongly up-regulated after nematode infection, particularly in animals with previous
exposure to nematodes. Genes for TNF-a and IFN-g were also transiently up-regulated, but to a lesser extent and more typically
after primary challenge. Naı̈ve sheep of both genotypes produced relatively little antibody response after primary challenge. A
second nematode challenge resulted in large increases in the lymphatic levels of all antibody sub-classes which were significant for
adult antigen-specific IgA and larval antigen-specific IgG1. In naı̈ve S line sheep, the larval-specific IgA and IgG2 response
appeared delayed when compared to the R line animals. Field-primed R and S line sheep had relatively high lymphatic IgG1 levels
prior to experimental infection and these did not change significantly afterwards. These results demonstrate that during nematode
infections, the intestinal micro-environment of sheep is transiently skewed towards Th2 cytokine dominance, although IFN-g gene
expression continues. This response is accompanied by increases of nematode-specific IgG1, IgA, IgG2 and IgM, as well as of total
IgE in lymph plasma.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Intestinal immune response; Afferent lymph; Efferent lymph; Cytokines; Antibody subclass; Nematode infection; Sheep
Abbreviations: AIL, afferent intestinal lymph; AILC, afferent
intestinal lymph cells; EIL, efferent intestinal lymph; EILC, efferent
intestinal lymph cells; FEC, faecal egg count; DC, dendritic cells
* Corresponding author. Tel.: +64 4 529 0577; fax: +64 4 529 0380.
E-mail address: [email protected]
(A. Pernthaner).
0165-2427/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.vetimm.2006.08.004
1. Introduction
The acquisition of immunity to gastro-intestinal
nematodes has seemingly paradoxical features. While
on the one hand, the immune mechanisms acting against
different parasites are uniquely adapted to specific life-
cycle stages (Meeusen et al., 2005), on the other hand
the host immune system appears to mount a stereotypic

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148136
Th2 response which activates a set of generic effector
mechanisms that protects against most intestinal
nematode parasites (Finkelman et al., 2004). The
inability to understand better the complex mix of
regulatory mechanisms which underlie immunity and
susceptibility to gastro-intestinal nematodes accounts,
at least partly, for the relative lack of success in
developing nematode vaccines.
Immune responses against gastro-intestinal nema-
todes are mounted in what is arguably the most
complex immunological micro-environment in the
body. The intestinal immune system mediates crucial
and often opposing immunological functions such as
protective responses to intestinal pathogens and
tolerance to harmless commensal bacteria and dietary
proteins in order to maintain homeostasis of the gut
(Mowat, 2003). To achieve these outcomes, the gut
immune system contains several structural and
functional specialisations for the uptake and proces-
sing of antigens and the dissemination of immune
responses. Achieving a better understanding of the
events unfolding within this regional microenviron-
ment during nematode infections will be critical to
the development of new vaccines. Research into the
role of the lymphatic system recently gained wide
spread attention as crucial functions of the normal
immune system and its functions in human diseases
are being recognized (Skobe et al., 2001; Swartz and
Skobe, 2001). A major cell population trafficking in
lymph is highly mobile dendritic cells (DC) which
form sentinels in the periphery, and their migrating
through afferent lymphatic vessels is crucial for the
execution of their functions (Randolph et al., 2005).
In large domestic animal species cannulation of the
afferent and efferent lymph ducts of a lymph node has
greatly facilitated the study of the immune system
functions, both under physiological conditions and
during immune responses to various pathogens. Many
of these studies utilize the lymphatic system draining
the skin of sheep (Gohin, 1997). The lymphatic
system draining from the wall of the small intestine,
which is an important site of gut-associated lymphoid
tissue (Lowden and Heath, 1993), has been less
explored.
Recently, we reported methods for long-term
collection of afferent and efferent lymph draining the
small intestine of sheep and showed that this approach
could be used to study the regional immune response to
diverse pathogens such as Mycobacterium avium
subsp. paratuberculosis or the nematode parasite
Trichostrongylus colubriformis (Hein et al., 2004).
We subsequently used this approach to compare the
nature of the regional response during a single
experimental infection with T. colubriformis in sheep
with different levels of genetically determined resis-
tance. When compared to susceptible-line (S) sheep,
resistant-line (R) sheep expressed higher levels of some
Th2 cytokines in immune cells migrating in intestinal
lymph (Pernthaner et al., 2005a), and developed
enhanced total- and nematode-specific IgE responses
in their intestinal lymph plasma (Pernthaner et al.,
2005b).
In the work reported here, we extended these
studies in two ways. Firstly, regional immune
responses were monitored in animals during two,
three, or four successive nematode infection cycles,
each of 21-day duration. This approach enabled the
detection of immunological events that require
repeated infections, as does the development of
protective immunity. Secondly, in addition to mon-
itoring the expression levels of the six cytokine genes
measured during primary infection in the earlier study,
we also measured nematode-specific IgA, IgG1, IgG2
and IgM in intestinal lymph at daily intervals
throughout the infection. The combination of cytokine
gene expression and antibody sub-class levels in
intestinal lymph provides a significantly more detailed
description of regional immune response parameters
during nematode infection than has been achieved
previously.
2. Material and methods
2.1. Surgical procedures
All animal experiments and surgical procedures
were approved by the Animal Ethics Committee of the
Wallaceville Animal Research Centre. Efferent lym-
phatic vessels draining from lymph nodes in the region
of the upper or mid jejunum were cannulated using a
vinyl cannula. A two-stage surgical procedure was used
to gain access to afferent lymph draining from the small
intestine as described previously (Pernthaner et al.,
2002). Briefly, in the first stage, mesenteric lymph nodes
which drain from the jejunum were removed, and in a
second stage operation, a reconstituted ‘pseudo-
afferent’ lymphatic vessel was located and cannulated
enabling continuous sampling of lymph. Methods for
long-term collection and characterization of afferent
and efferent lymph draining from the small intestine of
sheep were reported in detail elsewhere and are repeated
here only in brief to enable understanding of the overall
procedures and outline of the experiment (Hein et al.,
2004).

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148 137
2.2. Animals and experimental design
The aim of this study was to detect differences in the
cytokine gene expression and antibody subclass
responses in afferent and efferent intestinal lymph
between nematode resistant (R) and susceptible (S) line
sheep during two experimental infections with T.
colubriformis. Cytokine genes that are important in
mediating nematode disease or nematode immunity and
that are indicative for either a Th1 (IFN-g) or a Th2 type
response (IL-4, IL-5, IL-13), or are regulative (IL-10) or
predominantly pro-inflammatory (TNF-a) were
assayed.
Sheep were derived from the Wallaceville R (n = 6)
or nematode S (n = 5) sheep lines which differ at least
36-fold in their autumn faecal egg counts under natural
challenge on pasture (Bisset et al., 2001; Morris et al.,
2000). A total of 11 male sheep, aged between one and
two years was included in the study. Animals were
challenged orally on day 0 with 50,000 infective larvae
of T. colubriformis and the infection was terminated at
day 21 by drenching with oxfendazole (5 mg/kg body
weight; Systamex1, COOPERS). Sheep were then re-
challenged 7 days after drenching. All sheep had a
patent cannula for more than 20 days after the second T.
colubriformis infection. In two separate field-primed R-
line sheep, the cannulae remained patent throughout
three or four 21-day cycles of challenge infection
respectively, allowing extended observations in these
animals.
Animals were either nematode naı̈ve (pen-raised;
Rn, n = 3; Sn, n = 2) or had experienced previous
natural polygeneric nematode challenge (field-primed;
Rp, n = 3; Sp, n = 3). These animals were fitted with
either afferent (n = 5) or efferent (n = 6) lymphatic
cannulae.
2.3. Parasitology
Infective T. colubriformis larvae were cultured under
standard procedures from eggs obtained from faeces of
a mono-specifically infected sheep. Faecal egg counts
were performed at weekly intervals throughout the
duration of the experiment using the modified
McMaster method (Whitlock, 1948).
2.4. Sampling
Five to 15 ml of fresh lymph was collected daily over
a period of 30–60 min into sample tubes under aseptic
conditions to allow sampling of cells and lymph plasma.
Cells were separated from lymph by centrifugation,
washed once in PBS and re-suspended in RNAlater
(Ambion) to preserve the integrity of mRNA. Cell and
lymph samples were stored at�20 8C until subjected to
either RNA extraction and reverse transcription or
antibody subclass analysis.
2.5. RNA extraction and reverse transcription
RNA extraction and reverse transcription was
performed as described previously (Pernthaner et al.,
2005a). Briefly, RNAlater was removed prior to RNA
extraction by centrifugation and total RNA isolated
from cells using TRI REAGENT1LS (Molecular
Research Center, Inc.) based protocol according to
the manufacturer’s instructions. Total RNA yield was
calculated based on the absorption at 260/280 nm. First-
strand cDNA synthesis was performed using Super-
ScriptTMII RNAse H- reverse transcriptase (InVitrogen)
and poly(A) oligo(dT)12–18 primer to reverse tran-
scribe up to 5 mg of total RNA according to the
manufacturer’s protocol, and adjusted to an initial
concentration of 20 ng/ml RNA. cDNA was stored at
�20 8C until used for real time PCR.
2.6. Primer design, real time PCR and
quantification of gene expression
For each target gene a primer pair was selected using
the Primer Express software (Applied Biosystems) for
use in a SYBR1Green real time PCR assay, which were
described previously (Hein et al., 2004).
Cytokine gene expression was detected by real time
PCR using a GeneAmpTM 5700 sequence detection
system in combination with sequence detector software
(Applied Biosystems, Foster City, CA, USA) as
described previously (Pernthaner et al., 2005a). Briefly,
reactions were assembled in duplicates in optical 96-
well reaction plates using SYBR1Green PCR master
mix (Applied Biosystems). For each sample a melting
curve was generated after completion of amplification,
and analysed in comparison to the positive and negative
controls. Mean threshold cycle (CT) values of duplicate
samples were used for analysis.
The comparative CT method was employed for
relative quantification where the amount of target is
normalized to an endogenous reference (housekeeping
gene) (User Bulletin #2: ABI PRISM 7700 Sequence
Detection System; Applied Biosystems). The CT
indicates the fractional cycle number at which the
amount of amplified target reaches a fixed threshold,
and DCT represents the difference in threshold cycles
for the target and housekeeping gene.

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148138
Fig. 1. FEC in individual naı̈ve (n) and field-primed (p) R and S line
sheep during two cycles of experimental nematode infection. Animals
were infected on days 0 and 28 with 50,000 L3 T. colubriformis and
drenched 21 days after each challenge. FEC were determined 21 days
after each infection. FEC in Rp sheep were significantly (P < 0.05)
lower than in Rn, Sp and Sn sheep after the first challenge infection.
Both Rn and Rp sheep had significantly (P < 0.01) lower FEC than Sn
and Sp sheep after the second challenge infection. Numbers in
parentheses indicate the challenge infection.
2.7. Enzyme immunoassay (EIA) to detect specific
antibody subclass responses in lymph
Levels of T. colubriformis specific IgA, IgM, IgG1 and
IgG2 antibody subclasses in lymph were determined
using a non-competitive antibody sandwich immunoas-
say (EIA) as described previously (Douch et al., 1994).
Briefly, microtitre plates were coated either with T.
colubriformis excretory secretory products (TcESL3)
obtained from infective larvae or with somatic antigen
obtained from adult worms (TcAd) and incubated with
diluted lymph or serum samples. For IgG1 the samples
were diluted 1:400, for IgA, IgG2 and IgM 1:10.
Nematode specific antibody subclasses were then
detected using subclass-specific anti-ovine mAb and a
peroxidase based detection system. Lymphatic IgE levels
were detected using a non-competitive antibody sand-
wich EIA as described previously (Pernthaner et al.,
2005b; Shaw et al., 1997).
2.8. Nematode larvae and antigen preparation
Infective larvae (L3) of T. colubriformis were
obtained from cultures of faeces taken from a mono-
specifically infected sheep. T. colubriformis excretory
secretory product was obtained from exsheathed
infective larvae as described previously (Douch et al.,
1994). Somatic antigen was prepared by homogenizing
adult worms under liquid nitrogen. Soluble protein was
extracted in 5 mM Tris buffer pH 7.6 and stored at
�70 8C until used for assay.
2.9. Statistical analysis
Cytokine gene expression data were analysed on DCT
values using procedures for pulse analysis and REML
analysis of group means (Patterson and Thompson, 1971)
as described previously (Pernthaner et al., 2005a) with
the exception that subgroups were pooled when
appropriate. Pulse analysis for individual cytokine gene
expression profiles was done using the variables control
chart method, which is used for tracking processes over
time and is able to detect special cases not fitting into the
baseline variation, as described previously (Pernthaner
et al., 2005a). Data for antibody subclasses were analysed
using the same procedures as for cytokine gene
expression. FEC data were analysed as an unbalanced
two-way ANOVA after log(x + 100) transformation. The
non-parametric Kruskal–Wallis test was used to compare
time points of when antibody levels started to rise above
background between naı̈ve R and S line sheep. P < 0.05
was considered to be significant.
3. Results
3.1. Parasitology
FEC determined 3 weeks after the first challenge
infection showed that the Rp animals had significantly
(P < 0.05) lower FEC than the Rn, Sp or Sn sheep. At
this stage FEC of the Rn tended to be lower than for the
Sn sheep but this difference did not achieve signifi-
cance. The difference in FEC between R and S sheep
diverged further after the second challenge and both the
Rn and Rp groups showed significantly (P < 0.01)
lower FEC than the corresponding S groups (Fig. 1). As
expected FEC determined at weeks 1 and 2 after each
challenge infection were always 0.
3.2. Comparison of primary and secondary
nematode challenged cytokine responses in
intestinal lymph cells
3.2.1. IL-13 gene expression in lymph cells during
primary and secondary challenge infection of naı̈ve
and field-primed R and S sheep
The genotype of the animal but not the type of lymph
had a significant influence on the IL-13 gene expression.
Therefore, data for AILC and EILC were combined
for analysis. Nematode naı̈ve (n = 3) and field-primed
R sheep (n = 3) expressed significantly higher mean
levels of the IL-13 gene than S line sheep during two
experimental infections with T. colubriformis

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148 139
Fig. 2. Cytokine gene expression in AILC and EILC of individual naı̈ve (n) and field-primed (p) R and S sheep. Animals were infected on days 0 and
28 with 50,000 L3 T. colubriformis and drenched 21 days after each challenge. FEC determined at days 21 and 49 revealed large differences in egg
production between R and S sheep (for details see Fig. 1). IL-13 was significantly (P < 0.01) higher expressed in the R than S sheep. The time
required for the nematode development (L3, third stage larvae; L4, fourth stage larvae; EAd, early adult nematodes) is indicated. Results are
presented as DCT values, which is the difference in threshold cycles for the target and housekeeping gene. Therefore, low DCT values represent high
gene expression and high DCT values represent low gene expression. (a) Within naı̈ve R and S sheep the mean IL-13 gene expression did not differ
significantly between the first and second challenge infection. Large but transient increases occurred in the R sheep 8–14 days after primary and 2–8
days after secondary challenge. (b) For field-primed R and S sheep large but transient increases in the IL-13 gene expression occurred 1–7 days after
primary and secondary challenge. Only R sheep had an additional period of IL-13 gene up-regulation 15–21 days after primary challenge.
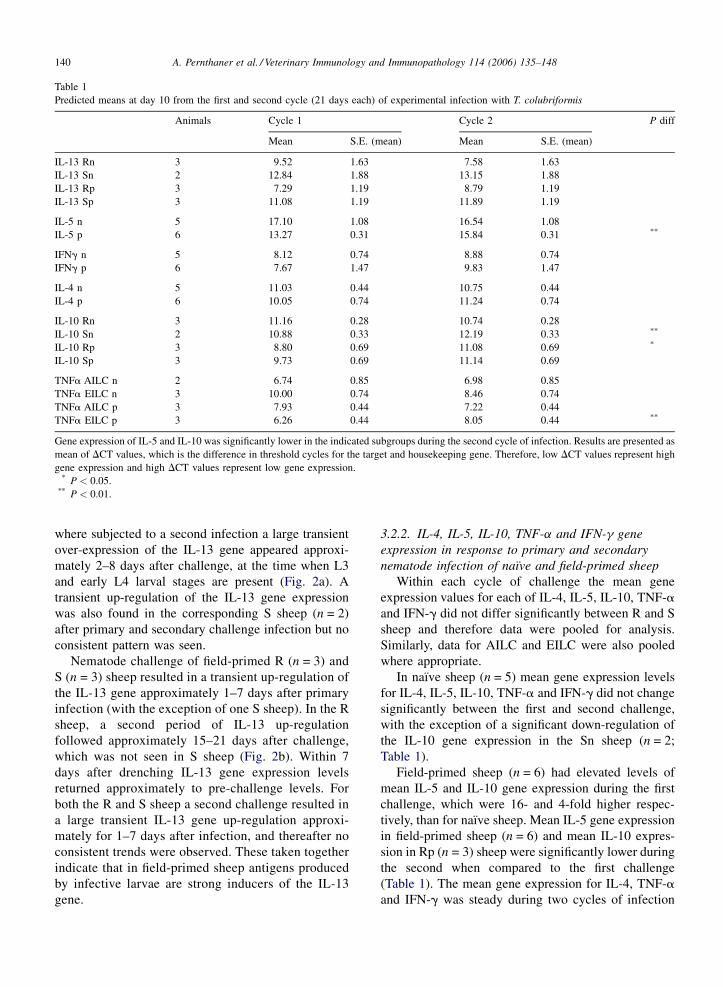
(P < 0.01; data from individual naı̈ve and field-primed
R and S sheep are shown in Fig. 2a and b). Naı̈ve R
sheep were able to respond to primary experimental
infection with a trend for IL-13 up-regulation, which
continued during the secondary infection, but differ-
ences of mean expression between cycle one and two
were not statistically significant (Table 1). The mean IL-
13 gene expression in the naı̈ve S sheep (n = 2) during
two challenge infections was low resulting in an
increase from an 11-fold to a 32-fold difference from
the first to the second challenge between R and S sheep.
In contrast, the 14-fold difference in the mean IL-13
gene expression seen between field-primed R (n = 3)
and S sheep (n = 3) during the first challenge declined to
a six-fold difference during the second challenge due to
a tendency for down-regulation in the R animals. In two
field-primed R sheep the mean expression of the IL-13
gene remained elevated throughout a third and fourth
challenge (data not shown).
When different developmental stages of T. colu-
briformis were taken into account, a large transient
over-expression of the IL-13 gene was consistently seen
in naı̈ve R animals (n = 3) approximately 8–14 days
post primary challenge, at the time when mostly L4
larval stages are present (Fig. 2a). In these sheep,
termination of the infection at day 21 resulted in a
decline of the IL-13 gene expression to approximately
pre-challenge levels within 7 days. When naı̈ve R sheep
A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148140
Table 1
Predicted means at day 10 from the first and second cycle (21 days each) of experimental infection with T. colubriformis
Animals Cycle 1 Cycle 2 P diff
Mean S.E. (mean) Mean S.E. (mean)
IL-13 Rn 3 9.52 1.63 7.58 1.63
IL-13 Sn 2 12.84 1.88 13.15 1.88
IL-13 Rp 3 7.29 1.19 8.79 1.19
IL-13 Sp 3 11.08 1.19 11.89 1.19
IL-5 n 5 17.10 1.08 16.54 1.08
IL-5 p 6 13.27 0.31 15.84 0.31 **
IFNg n 5 8.12 0.74 8.88 0.74
IFNg p 6 7.67 1.47 9.83 1.47
IL-4 n 5 11.03 0.44 10.75 0.44
IL-4 p 6 10.05 0.74 11.24 0.74
IL-10 Rn 3 11.16 0.28 10.74 0.28
IL-10 Sn 2 10.88 0.33 12.19 0.33 **
IL-10 Rp 3 8.80 0.69 11.08 0.69 *
IL-10 Sp 3 9.73 0.69 11.14 0.69
TNFa AILC n 2 6.74 0.85 6.98 0.85
TNFa EILC n 3 10.00 0.74 8.46 0.74
TNFa AILC p 3 7.93 0.44 7.22 0.44
TNFa EILC p 3 6.26 0.44 8.05 0.44 **
Gene expression of IL-5 and IL-10 was significantly lower in the indicated subgroups during the second cycle of infection. Results are presented as
mean of DCT values, which is the difference in threshold cycles for the target and housekeeping gene. Therefore, low DCT values represent high
gene expression and high DCT values represent low gene expression.* P < 0.05.
** P < 0.01.
where subjected to a second infection a large transient
over-expression of the IL-13 gene appeared approxi-
mately 2–8 days after challenge, at the time when L3
and early L4 larval stages are present (Fig. 2a). A
transient up-regulation of the IL-13 gene expression
was also found in the corresponding S sheep (n = 2)
after primary and secondary challenge infection but no
consistent pattern was seen.
Nematode challenge of field-primed R (n = 3) and
S (n = 3) sheep resulted in a transient up-regulation of
the IL-13 gene approximately 1–7 days after primary
infection (with the exception of one S sheep). In the R
sheep, a second period of IL-13 up-regulation
followed approximately 15–21 days after challenge,
which was not seen in S sheep (Fig. 2b). Within 7
days after drenching IL-13 gene expression levels
returned approximately to pre-challenge levels. For
both the R and S sheep a second challenge resulted in
a large transient IL-13 gene up-regulation approxi-
mately for 1–7 days after infection, and thereafter no
consistent trends were observed. These taken together
indicate that in field-primed sheep antigens produced
by infective larvae are strong inducers of the IL-13
gene.
3.2.2. IL-4, IL-5, IL-10, TNF-a and IFN-g gene
expression in response to primary and secondary
nematode infection of naı̈ve and field-primed sheep
Within each cycle of challenge the mean gene
expression values for each of IL-4, IL-5, IL-10, TNF-a
and IFN-g did not differ significantly between R and S
sheep and therefore data were pooled for analysis.
Similarly, data for AILC and EILC were also pooled
where appropriate.
In naı̈ve sheep (n = 5) mean gene expression levels
for IL-4, IL-5, IL-10, TNF-a and IFN-g did not change
significantly between the first and second challenge,
with the exception of a significant down-regulation of
the IL-10 gene expression in the Sn sheep (n = 2;
Table 1).
Field-primed sheep (n = 6) had elevated levels of
mean IL-5 and IL-10 gene expression during the first
challenge, which were 16- and 4-fold higher respec-
tively, than for naı̈ve sheep. Mean IL-5 gene expression
in field-primed sheep (n = 6) and mean IL-10 expres-
sion in Rp (n = 3) sheep were significantly lower during
the second when compared to the first challenge
(Table 1). The mean gene expression for IL-4, TNF-a
and IFN-g was steady during two cycles of infection

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148 141
with the exception of the down-regulation of TNF-a in
EILC of field-primed sheep (n = 3; P < 0.01; Table 1).
The gene expression of IL-5, TNF-a and IFN-g was
modulated in both AILC and EILC in response to
primary and secondary challenge infection. Large
transient increases in IL-5 gene expression levels were
seen in individual naı̈ve sheep during the first and
second challenge (data for individual R and S sheep are
given in Fig. 3a). Consistent trends were only seen for
IL-5 in R (n = 3) sheep which showed a transient
increase approximately 7–10 days after primary
challenge and 1–9 days after secondary challenge,
though base line expression levels differed substantially
between animals (Fig. 3a). Large transient increases in
Fig. 3. IL-5 gene expression in AILC and EILC of individual naı̈ve (n) or fie
with 50,000 L3 T. colubriformis and drenched 21 days after each challenge
production between R and S sheep (for details see Fig. 1). The time required
larvae; EAd, early adult nematodes) is indicated. Results are presented as DC
housekeeping gene. Therefore, low DCT values represent high gene expressi
gene expression did not differ significantly between the first and second chall
occurred in particular 7–10 days after the first and 1–9 days after the second
higher during the first than second challenge infection of field-primed (p) R an
after each challenge infection.
IL-5 gene expression were also seen in field-primed R
and S sheep (with the exception of an R animal during
the second challenge) in particular approximately 1–7
days after the first and second challenge (data for
individual R and S sheep are given in Fig. 3b). In field-
primed animals the main peaks for IL-5 gene expression
appear to coincide with the major peaks in IL-13
expression approximately 3–7 days after challenge.
The expression of the TNF-a gene was also
transiently up-regulated in particular after primary
challenge but no consistent trends within sheep lines
were seen (data not shown). Large but transient
increases in the IFN-g gene expression occurred in
naı̈ve and field-primed R and S sheep in particular
ld-primed (p) R and S sheep. Animals were infected on days 0 and 28
. FEC determined at days 21 and 49 revealed large differences in egg
for the nematode development (L3, third stage larvae; L4, fourth stage
T values, which is the difference in threshold cycles for the target and
on and high DCT values represent low gene expression. (a) Mean IL-5
enge infection of naı̈ve (n) R and S sheep. Large but transient increases
challenge infection. (b) Mean IL-5 gene expression was significantly
d S sheep. Large but transient increases occurred in particular 1–7 days

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148142
during the first challenge but animal-to-animal varia-
tions were large and no consistent trends within line
animals were seen (data not shown). In contrast,
changes in the IL-4 and IL-10 gene expression within
cycles of challenge infections were comparatively small
(data not shown).
3.3. Antibody responses in afferent and efferent
intestinal lymph
3.3.1. Lymphatic antibody subclass response of
naı̈ve R and S line sheep
No significant genotype related difference for the
lymphatic IgG1, IgM and IgE antibody subclass
responses was found in naı̈ve and field-primed animals
during two challenge infections. Therefore, data from R
and S animals were pooled for analysis. The only
genotype related differences were significantly higher
adult antigen-specific IgA and larval antigen-specific
IgG2 levels in the Rn when compared to the Sn sheep
(data not shown). Rn (n = 3) but not Sn sheep (n = 2)
had a significant increase in larval antigen-specific IgG2
levels between the first and second challenge infection
(Table 2).
Table 2
Predicted means of antibody subclass levels at day 10 from the first and secon
Animals Cycle 1
Mean S.E. (m
L3IgA n 5 �2.190 0.164
L3IgA p 6 �0.707 0.351
AdIgA n 5 �2.052 0.160
AdIgA p 6 �1.126 0.332
L3IgG1 n 5 �2.025 0.135
L3IgG1 p 6 �0.056 0.376
AdIgG1 n 5 �2.079 0.407
AdIgG1 p 6 �1.724 0.251
L3IgG2 Rn 3 �2.047 0.117
L3IgG2 Sn 2 �1.814 0.135
L3IgG2 Rp 3 �1.367 0.313
L3IgG2 Sp 3 �2.076 0.313
AdIgG2 n 5 �2.453 0.183
AdIgG2 p 6 �1.970 0.259
L3IgM n 5 �1.775 0.217
L3IgM p 6 �1.492 0.140
AdIgM n 5 �2.474 0.202
AdIgM p 6 �1.859 0.118
IgE n 5 0.879 0.358
IgE p 6 2.384 0.133
Levels of adult antigen-specific IgA (AdIgA), larval antigen-specific Ig
significantly from the first to the second cycle of infection in naı̈ve (n) s
which did not increase further during challenges. Data for IgA, IgG1, IgG2 a
IgE are mean values of U/ml.
Overall, antibody subclass responses of naı̈ve R and
S line sheep to primary experimental infection were
small, but for some animals adult antigen-specific IgA
and IgG1 and total IgE was detected, particularly at the
later stage of the challenge (data for IgA and IgG1 in
AIL and EIL from individual animals are given in
Fig. 4a and c, data for IgE not shown). A second
nematode challenge resulted in an increase in the
lymphatic IgA, IgG1, IgG2 and IgE production (data for
larval and adult antigen-specific IgA and IgG1 in AIL
and EIL from individual animals are given in Fig. 4a–d;
data for IgG2 and IgE not shown). This increase
achieved statistical significance for the adult antigen-
specific IgA (P < 0.01) and larval antigen-specific IgG1
(P < 0.05) responses (Table 2). Little larval but no adult
antigen-specific IgM response was seen during primary
infection which did not change during the secondary
challenge of naı̈ve sheep (Table 2). R line sheep (n = 3)
tended to respond earlier than S line sheep (n = 2) with a
TcES IgA and IgG2 response, although these differ-
ences did not achieve significance ( p = 0.08) due to the
small group size.
These findings show that naı̈ve R and S line sheep
were able to respond at the late stage of a primary
d cycle (21 days each) of experimental infection with T. colubriformis
Cycle 2 P diff
ean) Mean S.E. (mean)
�1.770 0.164
�0.653 0.351
�1.310 0.160 **
�0.958 0.332
�1.542 0.135 *
�0.023 0.376
�1.438 0.407
�1.712 0.251
�1.455 0.117 **
�1.923 0.135
�1.433 0.313
�1.995 0.313
�2.194 0.183
�1.890 0.259
�1.819 0.217
�1.558 0.140
�2.472 0.202
�1.849 0.118
1.539 0.358
2.654 0.133
G1 (L3IgG1) and larval antigen-specific IgG2 (L3IgG2) increased
heep. Field-primed (p) sheep had existing high levels of antibodies
nd IgM represent mean values of natural log transformed OD; data for

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148 143
Fig. 4. IgA and IgG1 subclass responses in AILC and EILC of individual naı̈ve (n) R and S sheep. Animals were infected on days 0 and 28 with
50,000 L3 T. colubriformis and drenched 21 days after each challenge. FEC determined at days 21 and 49 revealed large differences in egg
production between the R and S sheep after the second challenge infection (for details see Fig. 1). The time required for the nematode development
(L3, third stage larvae; L4, fourth stage larvae; EAd, early adult nematodes) is indicated. (a) Adult antigen-specific IgA levels during the first and
second nematode challenge of individual naı̈ve sheep (n = 5). Mean adult antigen-specific IgA levels increased significantly from the first to the
second nematode challenge (P < 0.01). (b) Larval (L3) antigen-specific IgA levels during the first and second nematode challenge of individual
naı̈ve sheep (n = 5). (c) Adult antigen-specific IgG1 levels during the first and second nematode challenge of individual naı̈ve sheep (n = 5). (d)
Larval (L3) antigen-specific IgG1 levels during the first and second nematode challenge of individual naı̈ve sheep (n = 5). Mean L3 antigen-specific
IgG1 levels increased significantly from the first to the second nematode challenge (P < 0.01).
experimental nematode infection with secretion of adult
antigen-specific lymphatic IgG1. With some exceptions
these animals were also able to respond to the second
nematode challenge with a nematode specific IgA,
IgG1, IgG2, and total IgE production. The antigen-
specific IgA and IgG2 response appeared to be
enhanced in R when compared to S line sheep.
3.3.2. Lymphatic antibody subclass response of
field-primed R and S line sheep
Field-primed R (n = 3) and S line sheep (n = 3) had
established larval and adult antigen-specific lymphatic
IgA, IgG1, IgG2, IgM and total IgE levels prior to
challenge, and mean values did not differ significantly
between the first and second challenge infection
(Table 2). IgG1, IgG2 and IgM levels remained
approximately steady throughout two challenge infec-
tions (data not shown). In two field-primed R sheep
these already elevated antibody subclass levels did not
change significantly during a third or fourth challenge
infection (data not shown).
In contrast to IgG1, IgG2 and IgM responses, large
transient increases in lymphatic IgA and total IgE levels
were seen after primary and secondary experimental T.
colubriformis infection in both line sheep (data for EIL
and AIL from individual R and S sheep are shown in
Fig. 5a–c). These transient increases appeared to be
biphasic with the first peak appearing approximately 2–
13 days and the second 16–20 days after the first
challenge. When developmental stages are considered

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148144
Fig. 5. IgA and IgE subclass responses in AILC and EILC of individual field-primed (p) R and S sheep. Animals were infected on days 0 and 28 with
50,000 L3 T. colubriformis and drenched 21 days after each challenge. FEC determined at days 21 and 49 revealed large differences in egg
production between the R and S sheep after each challenge infection (for details see Fig. 1). The time required for the nematode development (L3,
third stage larvae; L4, fourth stage larvae; EAd, early adult nematodes) is indicated. (a) Adult antigen-specific IgA levels during the first and second
nematode challenge of individual field-primed sheep (n = 6). (b) Larval (L3) antigen-specific IgA levels during the first and second nematode
challenge of individual field-primed sheep (n = 6). (c) Total IgE levels during the first and second nematode challenge of individual field-primed
sheep (n = 6).

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148 145
the first increase coincides with L3 and early L4 stages,
and the later peak with adult nematode parasites.
Transient increases were also seen during the second
challenge, again approximately 2–14 days after
challenge, but thereafter no consistent trends were
observed. Although these increases tended to be higher
in R than S line sheep, no significant difference was
detected due to the large animal-to-animal variation.
These findings demonstrate that field-primed R and S
line sheep were able to respond in a similar manner to
two experimental nematode infections in addition to
total IgE with antigen-specific IgG1, IgA, IgG2 and
IgM production and the response was directed against
both larval excretory/secretory and adult somatic
nematode antigens.
4. Discussion
Nematode challenge of the host is met by a range of
humoral and cellular responses but immunity is usually
Th2 dependent. Recent evidence suggests that the Th2
pathway is not a ‘default pathway’ but one that is
actively instructed by mechanisms that are only
beginning to be understood. Areas of intensive
investigation include studies on the role of antigen
presenting cells in Th2 response development, the
inhibitory function of IL-10, regulatory T-cells and
decoy receptors on chronic Th2-mediated inflamma-
tion, and the role of chitinases in mediating Th2 disease
which accelerated our understanding of the contribution
of these factors to Th2 immunity (Ramalingam et al.,
2005).
Recently, we described and validated reliable
methods for long-term collection of ovine afferent
and efferent intestinal lymph using experimental
infection with T. colubriformis as a model for Th2-
type and natural infection with Mycobacterium avium
spp. paratuberculosis as Th1-type responses (Hein
et al., 2004). In consecutive studies on Th2-type
responses we used the same lymphatic cannulation
procedures in the Wallaceville R and S line sheep while
they were subjected to a single experimental infection
with T. colubriformis. This approach allowed us to
monitor, without any need for further in vitro
stimulation, the in vivo expression levels of several
cytokine genes in cells migrating in lymph directly from
those sites where anti-nematode immune responses are
generated without compromising the integrity of the
immune system. We found that R line sheep were able
to respond to a single experimental nematode infection
in the presence of IFN-g with a significant up-regulation
of the genes for IL-13, IL-5 and TNF-a when compared
to S-line sheep (Pernthaner et al., 2005a). IgE levels
were also found to be higher in intestinal lymph from R
line sheep than S line sheep (Pernthaner et al., 2005b).
This study extends the previous findings and
demonstrates that cannulation of intestinal afferent
and efferent lymphatic vessels can also effectively be
used to investigate over periods of two or more
challenge infections in vivo intestinal nematode
immune responses. We were able to characterize
cytokine gene expression profiles and antibody subclass
responses during this prolonged period of time which
enabled us to compare primary with secondary
challenge responses.
The animals used in this study exhibited the expected
phenotypic differences (Bisset et al., 2001; Morris et al.,
2000) and overall, FEC were lower in the R than S
groups. The immune response to the second challenge
of Rn and to both challenge infections of Rp sheep was
fully protective, but S sheep remained susceptible to
nematode infection throughout the course of the
experiment. FEC were also lower in Rn than Sn sheep
following primary infection, although this difference
did not achieve statistical significance.
The genotype related difference in the mean IL-13
gene expression, which was observed previously during
primary infection (Pernthaner et al., 2005a), was
maintained during the second challenge of both naı̈ve
and field-primed sheep. In the case of naı̈ve animals, the
mean IL-13 gene expression increased in R, but not S
sheep between the first and second challenge, although
this increase in the R sheep did not achieve statistical
significance. The only genotype related change between
the first and second challenge was seen in the mean IL-
10 gene expression with a significant down-regulation
in the Sn and Rp animals during the second challenge.
In humans, enhanced secretion of IL-10 by CD14+
monocytes is associated with reduced chlamydia
antigen-specific lymphocyte proliferative and IFN-g
responses (Ohman et al., 2006). However, in this study
the down-regulation in IL-10 did not coincide with an
increase in the IFN-g response nor did it significantly
enhance the expression of the Th2 effector cytokines IL-
5 and IL-13 (Maizels, 2005). The reasons for the
genotype dependent IL-10 effects remain unknown and
require further study.
Compartment related differences in mean gene
expression between the first and second challenge
infection were generally small and only the down-
regulation of TNF-a in EILC of field-primed animals
was significant. Maybe some of the key functions of
TNF-a such as promoting activation of innate cells and
release of pro-inflammatory cytokines or influencing

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148146
the transition from innate to adaptive immunity
(Rahman and McFadden, 2006), are not required to
the same extent in animals previously exposed to
natural nematode infections. Interestingly, this down-
regulation is only seen in cells migrating in efferent
lymph, a cell population consisting almost exclusively
of lymphocytes. In contrast, afferent lymph cells, which
contain other cell subsets with the potential of TNF-a
expression like antigen presenting cells and natural
killer cells, retained the elevated TNF-a gene expres-
sion throughout two cycles of challenge. This indicates
that TNF-a mediated activation of antigen presenting
cells and natural killer cells, and differentiation of T and
B cells, remains steady at a mucosal level during
nematode challenge.
The only other significant difference in mean gene
expression levels between the first and second challenge
was the down-regulation of the IL-5 gene during the
second challenge of field-primed sheep. These animals
had elevated mean IL-5 gene expression levels during
the first challenge, which induced IgE production and
may have mediated nematode immunity via induction
of eosinophilia and mastocytosis. These functions of IL-
5 are well established in both sheep (Bao et al., 1996;
Gill et al., 2000) and rodents (Zhou et al., 1996) and
maintenance of these functions, once initiated, may not
require a permanent IL-5 stimulus.
Interestingly, in all experimental groups, mean IL-4
and IFN-g gene expression levels remained constant
over two challenge infections indicating that in sheep,
the transcription of IL-4 and IFN-g genes is not
modulated to the same extent as in laboratory animals
following nematode exposure (Deschoolmeester and
Else, 2002; Finkelman et al., 1997).
Although the overall changes in mean gene
expression levels between the first and second challenge
were comparatively small, large transient increases of
IL-13, IL-5, TNF-a and IFN-g over shorter periods of
time were present in many animals. When we attempted
to relate these temporary changes with the biology of
the nematode infection, stage-specific induction, in
particular of IL-13 and IL-5, became obvious. These
stage-specific transient increases, however, were not
entirely consistent between cycles of challenge infec-
tion, genotype or pre-challenge status. For the naı̈ve R
but not S sheep the induction of the IL-13 gene in
response to primary infection appears to be induced by
L4 larvae. When these R sheep were re-challenged, a
transient up-regulation appeared earlier in the course of
the infection indicating induction by L3 larvae.
Transient IL-13 up-regulation generally of lower
magnitude occurred also in the naı̈ve S sheep but these
peaks could not be related to any developmental stages.
In contrast, when field-primed R and S sheep were
subjected to a primary challenge, antigens produced by
L3 larvae appeared to induce an initial up-regulation of
the IL-13 gene, but a second peak at a later stage of the
infection was only seen in R sheep, when L4 and adult
nematodes would have been present. When field-
primed sheep were re-challenged, S sheep showed a
large transient up-regulation of the IL-13 gene at the
early stage of the infection.
Our data further show that in addition to IL-13, gene
expression of IL-5, IFN-g and TNF-a was clearly
modulated during two nematode challenge infections.
Nematode challenge of naı̈ve R sheep resulted in a
transient up-regulation of the IL-5 gene. Although
small animal numbers and a large animal-to-animal
variation might have masked some of the underlying
trends, a consistent up-regulation of the IL-5 gene was
induced at a time when L4 larval stages were present.
On the other hand, each of the two challenge infections
in the majority of field-primed R and S sheep resulted in
a transient up-regulation of the IL-5 gene expression
levels which tended to coincide with peaks in IL-13
gene expression. The major peaks for IL-13 and IL-5
gene expression appeared approximately 3–10 days
after challenge, which was consistent for field-primed
R and S sheep during both challenge infections and for
naı̈ve R sheep during the first challenge. These taken
together provide evidence that nematode antigens
expressed at specific developmental stages are power-
ful inducers of IL-13 and IL-5 gene transcription.
Furthermore, antigens produced by infective T.
colubriformis larvae are considered to be effective
inducers of Th2-type cytokines which is well known for
filarial infections (Osborne and Devaney, 1998) or
glycans derived from several nematode species (Tawill
et al., 2004). The identification of such T. colubriformis
nematode antigens, especially of the IL-13 inducing
antigens, is considered as having great potential in the
discovery of vaccine candidates, as the critical role of
IL-13 in mediating nematode immunity is generally
well recognized (Finkelman et al., 2004; Grencis,
1997; Grencis, 2001; McKenzie et al., 1998; Wynn,
2003). We found previously that IL-5 was over-
expressed in AILC of R when compared to S sheep
(Pernthaner et al., 2005a), but mice are able to develop
and maintain immunity to nematode infection in the
absence of IL-5 (Dent et al., 1999; Fallon et al., 2002;
Hokibara et al., 1997). This makes the discovery of
predominately IL-5 inducing nematode antigens a less
attractive option for identifying potential vaccine
candidates.

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148 147
Primary challenge of naı̈ve and field-primed sheep
resulted in repeated transient up-regulation of the genes
for TNF-a and IFN-g in most animals, which could not
be consistently related to any developmental stages.
This transient over-expression was less prominent and
somehow controlled during the second challenge. As
seen previously for primary challenge (Pernthaner et al.,
2005a), field-primed R line sheep were able to maintain
a protective immune response during the second
challenge infection in the presence of and despite
transient up-regulation of the IFN-g gene.
To our knowledge this is the first extensive report on
lymphatic antibody subclass titres in response to
repeated nematode infections. However, as mentioned
before, the group size of subgroups was small and
possible effects of lymphatic compartment or genotype
might not have been detected. Nonetheless, our data
show some interesting findings. Mean antibody titres for
antigen-specific IgG1, IgG2, IgA and total IgE
increased from the first to the second challenge of
naı̈ve sheep and this increase was statistically sig-
nificant for adult antigen-specific IgA and for larval
antigen-specific IgG1. Increases in other antigen-
specific subclass responses might not have been
identified as such as some naı̈ve animals were able to
respond to primary infection with a specific antibody
production. The only genotype related difference
detected was the significantly higher increase in larval
antigen-specific IgG2 response between the first and the
second challenge of Rn but not Sn sheep. Field-primed
sheep had previously experienced natural nematode
challenge and had existing lymphatic antibody titres
prior to challenge. As expected, these animals were able
to respond to nematode challenge with an early up-
regulation of lymphatic antibody subclass titres but
mean values did not increase significantly following a
second challenge. The pattern of the overall lymphatic
antibody responses was similar to those observed
previously in serum (Douch et al., 1994) with IgG1 and
IgG2 tending to continuously increase over time until
reaching a plateau.
In contrast to IgG1 and IgG2, the increase in antigen-
specific IgA and total IgE following primary and
secondary challenge of field-primed sheep appeared to
be biphasic, with a first peak coinciding with the
presence of infective and L4 larvae, and a second peak
by which time adult stages would have developed. This
is a strong indication of the stage-specific induction of
IgA and IgE, especially when the short serum half-life
of IgA and IgE is taken into consideration (Klobasa and
Werhahn, 1989; Curtis and Bourne, 1973; Vieira and
Rajewsky, 1988).
Although compromised by the small group size, our
data also provide evidence that R line sheep are able to
respond earlier than S line animals with a larval antigen-
specific IgA and IgG2 response. The response to adult
antigens was similar for both lines. Evidently, in R line
sheep, antigens produced by L3 and L4 larval
developmental stages serve as effective inducers of
multi-valent anti-nematode immune responses marked
by the expression of Th2-type cytokines and IgA and
IgG2. It remains to be elucidated if S line sheep respond
with a dominating IL-13 and IL-5 response in
conjunction with a larval antigen-specific IgA and
IgG2 production similar to that one seen in R line sheep
once protective nematode immunity is developed.
Taken together these results demonstrate that during
repeated nematode infections, the intestinal micro-
environment of sheep is transiently skewed towards Th2
cytokine dominance, although IFN-g gene expression
continues. The up-regulation of Th2 cytokines appears
to be biphasic with induction by antigens predominantly
produced by early larval developmental stages and, with
some restrictions, by adult nematodes. This response is
accompanied by increases of nematode-specific IgG1,
IgA, IgG2 and IgM, as well as of total IgE in lymph
plasma.
Acknowledgements
This work was supported by the Foundation for
Research, Science & Technology. We thank Roger
Littlejohn for advice about the statistical analyses used.
References
Bao, S., McClure, S.J., Emery, D.L., Husband, A.J., 1996. Interleukin-
5 Mrna expressed by eosinophils and gamma/delta T cells in
parasite-immune sheep. Eur. J. Immunol. 26, 552–556.
Bisset, S.A., Morris, C.A., McEwan, J.C., Vlassoff, A., 2001. Breed-
ing sheep in New Zealand that are less reliant on anthelmintics to
maintain health and productivity. N. Z. Vet. J. 49, 236–246.
Curtis, J., Bourne, F.J., 1973. Half-lives of immunoglobulins IgG, IgA
and IgM in the serum of new-born pigs. Immunology 24, 147–155.
Dent, L.A., Daly, C.M., Mayrhofer, G., Zimmerman, T., Hallett, A.,
Bignold, L.P., Creaney, J., Parsons, J.C., 1999. Interleukin-5
transgenic mice show enhanced resistance to primary infections
with nippostrongylus brasiliensis but not primary infections with
Toxocara canis. Infect Immun. 67, 989–993.
Deschoolmeester, M.L., Else, K.J., 2002. Cytokine and chemokine
responses underlying acute and chronic Trichuris muris infection.
Int. Rev. Immunol. 21, 439–467.
Douch, P.G.C., Green, R.S., Risdon, P.L., 1994. Antibody responses of
sheep to challenge with Trichostrongylus Colubriformis and the
effect of dexamethasone treatment. Int. J. Parasitol. 24, 921–928.
Fallon, P.G., Jolin, H.E., Smith, P., Emson, C.L., Townsend, M.J.,
Fallon, R., McKenzie, A.N., 2002. Il-4 induces characteristic Th2

A. Pernthaner et al. / Veterinary Immunology and Immunopathology 114 (2006) 135–148148
responses even in the combined absence of Il-5, Il-9, and Il-13.
Immunity 17, 7–17.
Finkelman, F.D., Shea-Donohue, T., Goldhill, J., Sullivan, C.A.,
Morris, S.C., Madden, K.B., Gause, W.C., Urban Jr., J.F., 1997.
Cytokine regulation of host defense against parasitic gastrointest-
inal nematodes: lessons from studies with rodent models. Annu.
Rev. Immunol. 15, 505–533.
Finkelman, F.D., Shea-Donohue, T., Morris, S.C., Gildea, L., Strait,
R., Madden, K.B., Schopf, L., Urban Jr., J.F., 2004. Interleukin-4-
and interleukin-13-mediated host protection against intestinal
nematode parasites. Immunol. Rev. 201, 139–155.
Gill, H.S., Altmann, K., Cross, M.L., Husband, A.J., 2000. Induction of
T helper 1- and T helper 2-type immune responses during hae-
monchus contortus infection in sheep. Immunology 99, 458–463.
Gohin, I., 1997. The lymphatic system and its functioning in sheep.
Vet. Res. 28, 417–438.
Grencis, R.K., 1997. Th2-mediated host protective immunity to
intestinal nematode infections. Philos. Trans. R. Soc. Lond. B.
Biol. Sci. 352, 1377–1384.
Grencis, R.K., 2001. Cytokine regulation of resistance and suscept-
ibility to intestinal nematode infection—from host to parasite. Vet.
Parasitol. 100, 45–50.
Hein, W.R., Barber, T., Cole, S.A., Morrison, L., Pernthaner, A., 2004.
Long-term collection and characterization of afferent lymph from
the ovine small intestine. J. Immunol. Meth. 293, 153–168.
Klobasa, F., Werhahn, E., 1989. Variations in the concentrations of the
immunoglobulins IgG1, IgG2, IgM and IgA in sheep. 2. Changes
in the blood of lambs of different breeds and crossbreeds during
the course of the rearing period. Berl. Munch. Tierarztl.
Wochenschr. 102, 331–337.
Hokibara, S., Takamoto, M., Tominaga, A., Takatsu, K., Sugane, K.,
1997. Marked eosinophilia in interleukin-5 transgenic mice fails to
prevent Trichinella spiralis infection. J. Parasitol. 83, 1186–1189.
Lowden, S., Heath, T., 1993. Lymphatic drainage from the distal small
intestine in sheep. J. Anat. 183 (Pt 1), 13–20.
Maizels, R.M., 2005. Infections and allergy - helminths, hygiene and
host immune regulation. Curr. Opin. Immunol. 17, 656–661.
McKenzie, G.J., Bancroft, A., Grencis, R.K., McKenzie, A.N., 1998.
A distinct role for interleukin-13 in Th2-cell-mediated immune
responses. Curr. Biol. 8, 339–342.
Meeusen, E.N., Balic, A., Bowles, V., 2005. Cells, cytokines and other
molecules associated with rejection of gastrointestinal nematode
parasites. Vet. Immunol. Immunopathol..
Morris, C.A., Vlassoff, A., Bisset, S.A., Baker, R.L., Watson, T.G.,
West, C.J., Wheeler, M., 2000. Continued selection of romney
sheep for resistance or susceptibility to nematode infection:
estimates of direct and correlated responses. Anim. Sci. 70, 17–27.
Mowat, A.M., 2003. Anatomical basis of tolerance and immunity to
intestinal antigens. Nat. Rev. Immunol. 3, 331–341.
Ohman, H., Tiitinen, A., Halttunen, M., Birkelund, S., Christiansen,
G., Koskela, P., Lehtinen, M., Paavonen, J., Surcel, H.M., 2006.
IL-10 polymorphism and cell-mediated immune response to
Chlamydia trachomatis. Genes Immun..
Osborne, J., Devaney, E., 1998. The L3 of Brugia induces a Th2-
polarized response following activation of an IL-4-producing
CD4-CD8- alphabeta T cell population. Int Immunol. 10,
1583–1590.
Patterson, H.D., Thompson, R., 1971. Recovery of inter-block
information when block sizes are unequal. Biometrika 58,
545–554.
Pernthaner, A., Cole, S.A., Gatehouse, T., Hein, W.R., 2002. Pheno-
typic diversity of antigen-presenting cells in ovine-afferent intest-
inal lymph. Arch. Med. Res. 33, 405–412.
Pernthaner, A., Cole, S.A., Morrison, L., Hein, W.R., 2005a. Increased
expression of interleukin-5 (Il-5), Il-13, and tumor necrosis factor
alpha genes in intestinal lymph cells of sheep selected for
enhanced resistance to nematodes during infection with Trichos-
trongylus colubriformis. Infect. Immun. 73, 2175–2183.
Pernthaner, A., Shaw, R.J., McNeill, M.M., Morrison, L., Hein, W.R.,
2005b. Total and nematode-specific Ige responses in intestinal
lymph of genetically resistant and susceptible sheep during infec-
tion with Trichostrongylus colubriformis. Vet. Immunol. Immu-
nopathol. 104, 69–80.
Rahman, M.M., McFadden, G., 2006. Modulation of tumor necrosis
factor by microbial pathogens. PLoS Pathog. 2, e4.
Ramalingam, T.R., Reiman, R.M., Wynn, T.A., 2005. Exploiting
worm and allergy models to understand Th2 cytokine biology.
Curr. Opin. Allergy Clin. Immunol. 5, 392–398.
Randolph, G.J., Angeli, V., Swartz, M.A., 2005. Dendritic-cell traf-
ficking to lymph nodes through lymphatic vessels. Nat. Rev.
Immunol. 5, 617–628.
Shaw, R.J., McNeill, M.M., Gatehouse, T.K., Douch, P.G., 1997.
Quantification of total sheep Ige concentration using anti-ovine
Ige monoclonal antibodies in an enzyme immunoassay. Vet.
Immunol. Immunopathol. 57, 253–265.
Skobe, M., Hawighorst, T., Jackson, D.G., Prevo, R., Janes, L.,
Velasco, P., Riccardi, L., Alitalo, K., Claffey, K., Detmar, M.,
2001. Induction of tumor lymphangiogenesis by Vegf-C promotes
breast cancer metastasis. Nat. Med. 7, 192–198.
Swartz, M.A., Skobe, M., 2001. Lymphatic function, lymphangiogen-
esis, and cancer metastasis. Microsc. Res. Technol. 55, 92–99.
Tawill, S., Le Goff, L., Ali, F., Blaxter, M., Allen, J.E., 2004. Both
free-living and parasitic nematodes induce a characteristic Th2
response that is dependent on the presence of intact glycans.
Infect. Immun. 72, 398–407.
Vieira, P., Rajewsky, K., 1988. The half-lives of serum immunoglo-
bulins in adult mice. Eur. J. Immunol. 18, 313–316.
Whitlock, H.V., 1948. Some modifications of the Mcmaster Helminth
egg-counting techniques and apparatus. J. Council Sci. Ind. Res.
21, 177–180.
Wynn, T.A., 2003. IL-13 effector functions. Annu. Rev. Immunol. 21,
425–456.
Zhou, Y., Bao, S., Rothwell, T.L., Husband, A.J., 1996. Differential
expression of interleukin-5 Mrna+ cells and eosinophils in nip-
postrongylus brasiliensis infection in resistant and susceptible
strains of mice. Eur. J. Immunol. 26, 2133–2139.
Copyright © 2022 FDOKUMEN




















