
CURRENT INVASIONS OF ASIAN CYCLOPID SPECIES (COPEPODA: CYCLOPIDAE) IN CRIMEA, WITH TAXONOMICAL AND...
22
Proof INTRODUCTION Crimea harbors a high landscape and biological diversity resulting from a complex geological history (Muratov et al. 1984, Apostolov et al. 1999). The Crimean fauna is a mixture of different faunistic components and belongs to two ecoregions of the Pale- arctic ecozone: northern Crimea is part of the Euro- Siberian region, while southern Crimea belongs to the Mediterranean Basin (Polishchuk 1998, Apostolov et al. 1999). As an example of the complex origin of the fauna, 44% of the mammal species are European, 32% are Mediterranean, and 22% are Central Asian (Apos- tolov et al. 1999). Cyclopidae are one of the largest and most diverse groups of copepods in inland waters (Monchenko CURRENT INVASIONS OF ASIAN CYCLOPID SPECIES (COPEPODA: CYCLOPIDAE) IN CRIMEA, WITH TAXONOMICAL AND ZOOGEOGRAPHICAL REMARKS ON THE HYPERSALINE AND FRESHWATER FAUNA ELENA ANUFRIIEVA 1, * , MARIA HOŁYŃSKA 2 and NICKOLAI SHADRIN 1 1 Institute of Biology of the Southern Seas, 2, Nakhimov ave., Sevastopol, 99011, Ukraine 2 Museum and Institute of Zoology, Wilcza 64, 00-679 Warsaw, Poland * Corresponding author: e-mail: [email protected] Abstract.— The Crimean Peninsula holds a large number of hypersaline water bodies. Our studies focused on these poorly investigated habitats, and included few brackish – and freshwater ponds. Seventeen species were identified, of which only 4(5) were collected from hypersaline waters sometimes with extremely high salinities (Acanthocyclops sp. copepodid, 210 ppt; Eucyclops sp. copepodid, 150 ppt; Diacyclops bisetosus and Cyclops furcifer, 140–150 ppt). We also report on the occurrence of three alien thermophilic species (Eucyclops roseus Ishida, 1997, Mesocyclops isabellae Dussart et Fernando, 1988, and Mesocyclops pehpeiensis Hu, 1943) from the brackish and fresh waters of Crimea. Morphological descriptions, illustrations of the diagnostic characters and comments on relevant taxonomic issues are supplemented with discussion of the putative ways of dispersal of the alien copepods to Crimea. We provisionally reinstate Eucyclops roseus, regarded by others as a subspecies of E. agiloides (G. O. Sars, 1909), and retain the name Acanthocyclops trajani Mirabdullayev et Defaye, 2002 which was recently synonymized with A. americanus (Marsh, 1893) here considered a nomen dubium. Species accumulation curves based on our and literature data showed that significantly larger sampling efforts could yield a total of 6–8 species in the hypersaline waters and 47–50 species in all types of continental waters of Crimea. Key words.— Inland waters, species richness estimation, alien microcrustaceans, morphology. ANNALES ZOOLOGICI (Warszawa), 2014, 64(1): xxx-xxx PL ISSN 0003-4541 © Fundacja Natura optima dux doi: ???
Transcript of CURRENT INVASIONS OF ASIAN CYCLOPID SPECIES (COPEPODA: CYCLOPIDAE) IN CRIMEA, WITH TAXONOMICAL AND...

Proof
INTRODUCTION
Crimea harbors a high landscape and biologicaldiversity resulting from a complex geological history(Muratov et al. 1984, Apostolov et al. 1999). TheCrimean fauna is a mixture of different faunistic components and belongs to two ecoregions of the Pale-arctic ecozone: northern Crimea is part of the Euro-
Siberian region, while southern Crimea belongs to theMediterranean Basin (Polishchuk 1998, Apostolov etal. 1999). As an example of the complex origin of thefauna, 44% of the mammal species are European, 32%are Mediterranean, and 22% are Central Asian (Apos-tolov et al. 1999).
Cyclopidae are one of the largest and most diversegroups of copepods in inland waters (Monchenko
CURRENT INVASIONS OF ASIAN CYCLOPID SPECIES(COPEPODA: CYCLOPIDAE) IN CRIMEA, WITH
TAXONOMICAL AND ZOOGEOGRAPHICAL REMARKS ONTHE HYPERSALINE AND FRESHWATER FAUNA
ELENA ANUFRIIEVA1, *, MARIA HOŁYŃSKA2
and NICKOLAI SHADRIN1
1Institute of Biology of the Southern Seas, 2, Nakhimov ave., Sevastopol, 99011,Ukraine
2Museum and Institute of Zoology, Wilcza 64, 00-679 Warsaw, Poland*Corresponding author: e-mail: [email protected]
Abstract.— The Crimean Peninsula holds a large number of hypersaline water bodies. Ourstudies focused on these poorly investigated habitats, and included few brackish – andfreshwater ponds. Seventeen species were identified, of which only 4(5) were collected fromhypersaline waters sometimes with extremely high salinities (Acanthocyclops sp.copepodid, 210 ppt; Eucyclops sp. copepodid, 150 ppt; Diacyclops bisetosus and Cyclopsfurcifer, 140–150 ppt). We also report on the occurrence of three alien thermophilic species(Eucyclops roseus Ishida, 1997, Mesocyclops isabellae Dussart et Fernando, 1988, andMesocyclops pehpeiensis Hu, 1943) from the brackish and fresh waters of Crimea.Morphological descriptions, illustrations of the diagnostic characters and comments onrelevant taxonomic issues are supplemented with discussion of the putative ways ofdispersal of the alien copepods to Crimea. We provisionally reinstate Eucyclops roseus,regarded by others as a subspecies of E. agiloides (G. O. Sars, 1909), and retain the nameAcanthocyclops trajani Mirabdullayev et Defaye, 2002 which was recently synonymizedwith A. americanus (Marsh, 1893) here considered a nomen dubium. Speciesaccumulation curves based on our and literature data showed that significantly largersampling efforts could yield a total of 6–8 species in the hypersaline waters and 47–50species in all types of continental waters of Crimea.
�
Key words.— Inland waters, species richness estimation, alien microcrustaceans, morphology.
AA NN NN AA LL EE SS ZZ OO OO LL OO GG II CC II ((WWaarrsszzaawwaa)),, 22001144,, 6644((11)):: xxxxxx--xxxxxx
PL ISSN 0003-4541 © Fundacja Natura optima duxdoi: ???

2003). The earliest surveys on the Crimean cyclopoidswere provided by Sovinsky (1891), Lebedinsky (1904)and Velichkevich (1931). More comprehensive over-views of the Crimean copepod fauna were published byTseeb (1947, 1961) and Ulomsky (1955). By the time V.I. Monchenko started his studies in Crimea, 25 cyclopidspecies had been known from the region. In a mono-graph of the Cyclopoida of the Ponto-Caspian basinMonchenko (2003) listed 34 taxa that he had identifiedhimself from 103 samples from the Crimea, yet failed tofind 6 other taxa previously recorded in the peninsula.Thence, according to this census (Monchenko 2003) ~ 40 cyclopid species occur in the Crimea. This speciesrichness is highlighted when compared with that oflarger areas such as Poland which is more than eleventimes as large as the Crimea (51 cyclopid species)(Hołyńska 2008) and Italy (its area is similar toPoland) with a very rich subterranean fauna (101species) (Stoch 2007). Crimea is the largest (nearly26.5 thousands km2) peninsula in the Black Sea. North-ern Crimea is a steppe plain (19 thousands km2), whilstthe southern part (7 thousands km2) is highland withthree mountain ranges (the highest peak: Roman-Kosh,1545 m a.s.l.). The climate varies from subtropical inthe South Crimean Coast to semiarid in North, Eastand West Crimea. The summer maximum temperaturecan reach more than +40°C, and winter minimum to -10°C (average: -5 to +1°C). The total precipita-tion is 360–390 mm in the coastal lowland (Sudak, Sevastopol), and reaches 1500 mm in the mountains.
Natural freshwater bodies are relatively scarce inCrimea – they are represented by small springs andadjacent pools, small mountain and steppe rivers/streams, cave waters, etc. Artificial water bodies how-ever are present in a wide variety, such as reservoirscreated by dams, the North Crimean Canal which is oneof the largest canal systems in Europe, ponds, decora-tive pools, rice fields, and other small water bodies(Oliferov and Timchenko 2005). The hypersaline waterbodies constitute a very characteristic and peculiarhabitat type in the region: fifty relatively large lakesand numerous small (from several to hundreds meterslong) hypersaline water bodies (Shadrin 2009) areknown in the peninsula. The Crimean hypersalinelakes are divided into two types: of marine origin (tha-lassohaline); and of continental origin (athalassoha-line–sulfate; the athalassohaline lakes formed in thecalderas of ancient mud volcanoes). Also, salinizationprocess has recently started in some artificial ponds(Shadrin et al. 2012). Despite the long history of cope-pod studies in Ukraine (Sovinsky 1891), very littleresearch has been done on the hypersaline lakes and,as a consequence, their fauna is still poorly under-stood (Apostolov et al. 1999). There are no data onCyclopidae either; the only exceptions are records of“unidentified Cyclopoida„ (Belmonte et al. 2012) and
Acanthocyclops americanus (Marsh, 1893) (Anufri-ieva and Shadrin 2012) from few hypersaline lakes.Comprehensive faunistic surveys of the hypersalinewaters in Crimea has begun very recently (Anufriievaand Shadrin 2012, Belmonte et al. 2012).
Given the current poor state of knowledge of thehypersaline fauna in Crimea, we started a long-termbiodiversity project in 2000, and from 2012 we focusedon the Cyclopidae. Our first (partial) results, supple-mented with data on some freshwater ponds, are pre-sented herein.
MATERIAL AND METHODS
A total of 117 samples were collected from 39 waterbodies in 2012–2013; 100 samples were collected fromhypersaline water bodies, 11 from brackish waters,and 6 from fresh waters. Fig. 1 shows a map of all sam-pling sites. Only 17 samples contained specimens ofCyclopidae, the characteristics of those water bodiesare given in Table 1. Most of the investigated waterbodies are very shallow, therefore water was collectedby a 5 liter bucket. Up to 50–100 L of water were fil-tered through plankton net with mesh size of 110 µm,the resulting samples were preserved in the field witha 4% buffered formalin solution. Salinity, temperature,and pH were measured during each sampling. Speci-mens were dissected in glycerin under an Olympus SZ-ST stereo microscope. Identifications and measure-ments were made using an Olympus BX50 compoundmicroscope with Nomarski (differential interferencecontrast) optics. Drawings were prepared with the aidof a drawing tube attached to the compound micro-scope. The morphometric measurements were takenfollowing Koźmiński (1936). Dissected specimens weremounted in glycerin and the semi-permanent slideswere sealed with nail polish. In a few cases we couldnot identify the species, because only juvenile speci-mens were collected in the samples. Parameters ofregression equations (accumulation curves) and corre-lation coefficients were calculated in Excel; the confi-dence level of the correlation coefficients was deter-mined from the table of Müller et al. (1979). Randompermutations of data were made on-line (http://www.webcalculator.co.uk/statistics/rpermute3.htm) to cal-culate accumulation curves. The Chao 2 index, a non-parametric species richness estimator is calculated asSChao2 = Sobs + F1
2/2F2, where: SChao2 and Sobs are theexpected and observed species richness, respectively;F1 and F2 are the number of singletons (number ofspecies occurring in single sample only) and double-tons (number of species occurring in two samples).
In the species diagnoses the setae of the caudalrami are denoted by Roman numerals, following the scheme applied by Huys and Boxshall (1991).
2 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN

Reference slides are deposited in the:IBSS – Institute of Biology of the Southern Seas, Sevas-
topol, Ukraine;MIZ – Museum and Institute of Zoology PAS, Warsaw,
Poland.Morphological abbreviations:
P1–P4 – swimming legs 1–4, exp – exopodite, enp – endopodite.
RESULTS
Cyclopidae were present in 6%, 100% and 100% ofthe samples collected in the hypersaline, brackish (to30 ppt) and freshwater habitats, respectively. A total of17 taxa were identified on species level and three taxawere identified only at the genus level, 14 of which werecollected from freshwater, 8 from brackish habitats,and 4(5) from hypersaline water bodies (Table 2).Some of these taxa withstand surprisingly high salini-ties: Acanthocyclops sp. (copepodid, 210 ppt), Eucy-clops sp. (copepodid, 150 ppt), Diacyclops bisetosus(Rehberg, 1880) and Cyclops furcifer Claus, 1857
(140–150 ppt). Diacyclops bisetosus was the most fre-quent species in the hypersaline samples.
We identified two species of Mesocyclops, whichhave never (M. isabellae Dussart et Fernando, 1988)or very rarely (M. pehpeiensis Hu, 1943), been report-ed from Europe. Another interesting finding was theoccurrence of Eucyclops roseus Ishida, 1997 in twolocalities (oligohaline/fresh waters). The only Euro-pean record of this species is from northern Germany(Ishida 1997, Alekseev and Defaye 2011). Acanthocy-clops trajani Mirabdullayev et Defaye, 2002 is alsoreported in Ukraine for the first time, though previousrecords of A. americanus (Monchenko 2003, Anufriie-va and Shadrin 2012) very likely refer to the formerspecies. We also collected Thermocyclops oithono-ides (G. O. Sars, 1863) in a spring pool (freshwater).The species is common in other parts of Ukraine, but ithas never been recorded in Crimea yet (Monchenko2003). Thus, the total number of copepod species so faridentified in Crimea is 44 (or 43) (see also the taxonom-ical notes on E. roseus and A. trajani).
We provide short diagnoses and taxonomic notes on those taxa that are new to Europe or Ukraine,and/or show previously unreported morphologicalpeculiarities.
CYCLOPIDAE IN CRIMEA 3
Figure 1. Map showing all the sampling sites in Crimea (numbers 1–17 indicate sites with cyclopids. Numbers on the map correspond to the numbers of the water bodies in Table 1). Stars are sampling sites without cyclopoids.
4 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN
No.
of
Geo
grap
hica
lTy
pe o
fTo
tal r
ange
of
fact
ors
Sam
plin
g in
form
atio
nw
ater
Wat
er b
ody,
nam
eco
ordi
nate
sha
bita
tS
, pp
tT,
°C
pHD
ate
S,
ppt
T, °
CpH
body 1
Ach
i45
°9'N
35
°25'
E1
65–3
700–
336.
7–8.
315
.04.
1321
0–21
221
8.0
2A
ygul
skoy
e45
°59'
N34
°35'
E2
150–
270
0–30
6.0–
7.3
12.0
4.13
150
17–
3S
him
akha
nsko
ye45
°10'
N36
°25'
E1
23–4
500–
496.
4–9.
114
.04.
1314
016
8.6
4C
hers
ones
sus
Lake
44°5
9'N
33°3
9'E
223
–290
0–37
7.1–
9.3
23.0
5.13
2322
–5
Pon
d ne
ar W
cor
ner
of L
ake
Tobe
chik
45°1
1'N
36°1
8'E
118
–145
0–31
7.4–
7.9
13.0
4.13
40–4
220
–6
Akt
ashs
koye
45°2
2'N
35°4
9'E
214
–360
0–31
6.7–
8.4
15.0
4.13
36–3
814
8.4
7K
oyas
hsko
ye (
SE
par
t)45
°02'
N35
°12'
E2
30–3
900–
316.
0–8.
714
.04.
1330
–32
157.
98
Yan
ishs
koye
45°0
7'N
36°2
4'E
221
–166
0–36
6.9–
8.5
13.0
4.13
2122
–9
Pon
d ne
ar v
. C
hela
ydin
ovo
45°1
3'N
36°2
2'E
39–
240–
327.
0–7.
613
.04.
1317
16–
10P
ond
befo
re V
ladi
slav
ovka
45°0
9'N
35°2
3'E
310
–20
0–25
8.6
15.0
4.13
12–1
417
8.6
11K
uchu
k-A
djig
ol45
°06'
N35
°27'
E4
0–80
0–25
7.0–
9.5
15.0
4.13
515
8.6
12K
uchu
k-A
djig
ol45
°06'
N35
°27'
E4
0–80
0–25
7.0–
9.5
8.08
.12
030
7.4
13P
ond
No.
1,
near
Yal
ta44
°32'
N35
°27'
E5
0–
–15
.05.
130
20–
14P
ond
No.
2,
near
Yal
ta44
°32'
N35
°34'
E6
0–
–15
.05.
130
21–
15P
ond
No.
3,
near
Yal
ta44
°32'
N35
°34'
E7
0–
–15
.05.
130
21–
16P
ond
No.
4,
near
Yal
ta44
°32'
N35
°34'
E5
0–
–15
.05.
130
20–
17P
ond
No.
5,
near
Yal
ta44
°31'
N35
°35'
E5
0–
–16
.05.
130
21–
18P
ool,
near
Sev
asto
pol
44°3
5'N
33°2
4'E
80
––
23.0
5.13
021
–
Tab
le 1
. Geo
grap
hic
coor
dina
tes
and
phys
icoc
hem
ical
feat
ures
of t
he w
ater
bod
ies
in w
hich
Cyc
lopi
dae
wer
e pr
esen
t. T
ype
of h
abit
at: 1
– a
thal
asso
halin
e –
sulfa
te h
yper
salin
e la
ke,
2 –
thal
asso
halin
e hy
pers
alin
e la
ke, 3
– b
rack
ish
pond
, 4 –
form
erly
a h
yper
salin
e la
ke, 5
– fr
eshw
ater
pon
d, 6
– s
prin
g po
ol, 7
– p
ond-
wet
land
, 8 –
dec
orat
ive
fres
hwat
er p
ool

Num
ber
of w
ater
bod
y in
Tab
. 1
Spe
cies
12
34
56
78
910
1112
1314
1516
1718
Aca
ntho
cycl
ops
sp.
++
A.
traj
aniM
irabd
ulla
yev
et D
efay
e, 2
002
++
A.
cf.
traj
aniM
irabd
ulla
yev
& D
efay
e, 2
002
+A
. ve
rnal
is(F
isch
er,
1853
)+
Cyc
lops
sp.
+C
. fu
rcife
rC
laus
, 18
57+
+C
. st
renu
usF
isch
er,
1851
++
C.
vici
nus
Ulja
nin,
187
5+
Dia
cycl
ops
sp.
++
D.
bise
tosu
s(R
ehbe
rg,
1880
)+
++
++
+D
. bi
cusp
idat
us(C
laus
, 18
57)
++
Euc
yclo
pssp
.+
+E
. ro
seus
Ishi
da,
1997
++
E.
cf.
sper
atus
(Lill
jebo
rg,
1901
)+
Mac
rocy
clop
s al
bidu
s(J
urin
e, 1
820)
+M
esoc
yclo
ps is
abel
lae
Dus
sart
et
Fer
nand
o, 1
988
+M
. pe
hpei
ensi
sH
u, 1
943
+M
egac
yclo
ps v
iridi
s(J
urin
e, 1
820)
++
Mic
rocy
clop
s ru
bellu
s(L
illje
borg
, 19
01)
+P
arac
yclo
ps f
imbr
iatu
s(F
isch
er,
1853
)+
The
rmoc
yclo
ps c
rass
us(F
isch
er,
1853
)+
Th.
oith
onoi
des
(G.
O.
Sar
s, 1
863)
+C
yclo
poid
a*+
Tota
l num
ber
of s
peci
es p
er la
ke1
31
11
11
22
23
41
51
41
3
CYCLOPIDAE IN CRIMEA 5
Tab
le 2
. Lis
t of
Cyc
lopi
dae
spec
ies
foun
d in
diff
eren
t w
ater
bod
ies
of C
rim
ea
*– U
nide
ntifi
ed la
rvae
(C
II, C
III)
of C
yclo
pida
e

TAXONOMIC ACCOUNTS
Family Cyclopidae Rafinesque, 1815
Subfamily Eucyclopinae Kiefer, 1927
EEuuccyyccllooppss rroosseeuuss Ishida, 1997(Figs 2A–F, 3A–E)
Eucyclops roseus Ishida, 1997: 350–354, Figs 1–3; Ishida, 1998: 24;Chang, 2009: 394–397, 619–620, Figs 208–209; Eucyclops agilo-ides roseus: Alekseev and Defaye, 2011: 61–64, 67–69.
? Eucyclops agiloides: Monchenko, 2003: 79–80, 83, 87.
MMaatteerriiaall eexxaammiinneedd. Ukraine, Crimea: LakeKuchuk-Ajigol, leg. E. Anufriieva, 15 April 2013, twofemales (IBSS); Sevastopol, decorative pool, leg. E.Anufriieva, 23 May 2013, one female (IBSS).
OOtthheerr mmaatteerriiaall. Iraq, South Iraq: Al Salal marsh,20 km NW Basra, 30°40’N 47°36’E, lake shore, leg. S. D.Salman and M. Hołyńska, 24 March 2012, three females(MIZ); Shatt al Arab river, Basra, 30°31’N 47°49’E, leg.S. D. Salman and M. Hołyńska, 21 March 2012, onefemale and one male (MIZ).
DDiiaaggnnoossiiss. Total body length without caudal setae:1103 µm; prosome/urosome: 1.58; cephalothorax length/width: 1.24; genital double-somite length/width: 0.89.
Lateral margins of pediger 5 with hairs (Fig. 2A).Seminal receptacle typical of genus, lateral arms ofanterior part slightly shorter than those of posteriorpart (Fig. 2A). Hyaline frill on posterior edge of genitaldouble-somite irregularly serrated. Ventral surface ofgenital double-somite with 2 posterolateral sensilla.Anal somite (Fig. 2B) bearing row of small spinules onposterior margin, and 2 sensilla on dorsal surface. Pro-ctodeum (hindgut) with single row of long hairs on bothsides. Anal operculum distinct, convex (Fig. 2B). Cau-dal rami slightly divergent without hairs on inner mar-gin, 5.0 times as long as wide. Longitudinal row of spin-ules (“serra”) present along most of outer edge of eachramus. Posterolateral (III) caudal seta with row of shortspinules on outer margin and longer setules on innermargin; spinules present at insertion of seta. Relativelengths of terminal caudal setae from terminal accesso-ry (VI) to posterolateral (III): 1.15; 7.08; 4.69; 1.00.
Antennule 12-segmented, last 3 segments withnearly smooth hyaline membrane (Fig. 2C). Antennaenp2 with 9 setae. Frontal surface of antennal coxoba-sis (Fig. 2D) ornamented with: two groups of long hair-like spinules in distal half of segment (next distal mar-gin and proximal to medial seta), three oblique andparallel rows of spinules in proximal part; and 2 groupsof shorter spinules on lateral margin. Caudal surface ofantennal coxobasis (Fig. 2E) bearing: 5 strong spinulesnext distal margin; 6 strong spinules more proxim-ally at height of exopodite seta; 2 longitudinal rows of
spinules of similar size near lateral margin; transverserow of spinules near implantation of medial setae, andfew spinules proximal to insertion of medial setae; tinyspinules proximally to longitudinal rows, and nearmedial margin.
Maxillulary palp (Fig. 2F) two-segmented. Proxi-mal article bearing 3 medial setae and 1 lateral seta(remnant of exopodite); anterior surface with spinulesin distinct oval pattern. Distal segment, representingendopodite, armed with 3 setae.
Spine formula of exp3 of P1-P4, 3–4–4–3. Medialseta on P1 basipodite with short setules at whole length(Fig. 3A). Caudal surface of P2–P4 couplers (Figs3B–D) with transverse rows of hairs; pilosity mostdense on P4 coupler, hairs continuous on distal margin.P4 coxopodite bearing rich ornamentation of spinules,typical of genus. Coxopodal seta with heteronomoussetulation: long setules in proximal half and shortsetules in distal half on inner (medial) margin; setulesonly present in distal half on outer (lateral) margin. P4enp3 2.3 times as long as wide. Inner apical spine of P4enp3 1.6 times longer than outer apical spine, and 1.2times as long as segment (Fig. 3E); inner (medial) andouter (lateral) setae not reaching tip of longer apicalspine. Leg 5 (Fig. 2A) one-segmented, bearing medialspine and apical and lateral setae. Medial spine dis-tinctly longer than segment, small spinules present atbase of spine.
Notes on taxonomy
Eucyclops roseus was originally described fromthe Okinawa Island (Ryukyus), Japan (Ishida 1997),yet Ishida (1997, 1998) himself recorded this speciesfrom several distant places, such as the more northernJapanese islands (Honshu and Hokkaido), the RussianFar East (Primorskiy), northern Germany, and LakeVictoria and Lake Naivasha in Kenya. Ishida (1997)also mentioned that E. roseus might be present in Chi-na, and some of the records of E. speratus (Lilljeborg,1901) (e.g. Tai and Chen 1979) might in fact refer to E. roseus. This species is the most frequent and abun-dant cyclopid in South Korean inland waters (Chang2009). El-Shabrawy (2013) reported it from the RiverNile, Lower Egypt, and from ponds and irrigationcanals in the neighborhood of Cairo. The species alsooccurs in Sudan (G. Idris and M. Hołyńska unpublishedresults) and the Mesopotamian marshes (H. H. Mo-hammed and M. Hołyńska unpublished results).
In a review of the Eucyclops serrulatus group,Alekseev and Defaye (2011) pointed to the close rela-tionships of E. roseus and E. agiloides (G. O. Sars,1909), and considered E. roseus as a subspecies of E. agiloides. The authors mentioned (identificationkey) one character [two (E. agiloides roseus) vs. one
6 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN

CYCLOPIDAE IN CRIMEA 7
Figure 2. Eucyclops roseus Ishida, 1997 from decorative pool, Sevastopol, Ukraine, female: (A) pediger 5 and genital double-somite, ventral view;(B) anal somite and caudal rami, dorsal view; (C) last antennulary segment; (D) antennal coxobasis, frontal view; (E) antennal coxobasis, caudal
view; (F) maxillulary palp. Scale bars = 50 µm.

8 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN
Figure 3. Eucyclops roseus Ishida, 1997 from decorative pool, Sevastopol, Ukraine, female: (A) leg 1 coupler, coxopodite and basipodite(coxopodal seta broken), frontal view; (B) leg 2 coupler, coxopodite and basipodite, caudal view; (C) leg 3 coupler, coxopodite and basipodite,
caudal view; (D) leg 4 coupler, coxopodite and basipodite, caudal view; (E) leg 4, third segment of endopodite. Scale bars = 50 µm.

(E. agiloides s. str.) distal group of long spinules onthe frontal surface of the antennal coxobasis], by whichthe two forms can be distinguished from each other,but they did not specify if any difference in the geo-graphic distribution between the nominotypical sub-species and ’E. agiloides roseus‘ existed. Alekseevand Defaye also provided a short diagnosis and somedrawings (mouthparts were not illustrated) of E. agi-loides s. str., based on specimen(s?) from LakeMalawi. In the original description (Sars 1909) thediagnosis and illustration of Eucyclops agiloideswere based on a female from Lake Victoria (VictoriaNyanza) [the type locality is not Lake Malawi (LakeNyasa), as it was stated by Alekseev and Defaye(2011)], and in the same work Sars also mentioned twoother females, apparently the same species found inthe southern and southwestern part of Lake Tanganyi-ka. Sars’ (1909) description and drawings naturally donot report any microcharacter, yet some features (slen-der habitus; terminal accessory (VI) caudal seta ~ 1.6times as long as posterolateral (III) caudal seta; termi-nal segment of antennule relatively short, ca. five timesas long as wide; and the inner apical spine of P4 enp3shorter than segment) might indicate that E. agiloidessensu G. O. Sars is not conspecific with the LakeMalawi specimen shown by Alekseev and Defaye(2011). The females from Crimea differ from bothforms: two distal groups of long spinules are present onthe frontal surface of antennal coxobasis (one group ofspinules is present in E. agiloides agiloides sensuAlekseev and Defaye); the terminal accessory and pos-terolateral caudal setae are subequal, the terminal seg-ment of antennule is ~ 8 times longer than wide, andthe medial apical spine on P4 enp3 is 1.2 times longerthan segment (terminal accessory caudal seta distinct-ly longer, terminal segment of antennule relativelyshort, and medial apical spine of P4 enp3 shorter thansegment in E. agiloides agiloides sensu G. O. Sars).We do not deny a supposedly close relationshipbetween E. agiloides G. O. Sars and E. roseus Ishida,yet as long as the morphology of E. agiloides is unsuf-ficiently known, it is better to keep the name E. roseusrather than to sink it in the poorly-defined E. agi-loides. The latter species or a form named as E. cf.agiloides have been recorded from East- North- andWest Africa, several countries in Asia [Israel, Turkey,Iran, India, China and Indonesia (Sumatra, Java)](Kiefer 1933, Dussart 1981, Dussart and Defaye 2006,Alekseev and Defaye 2011) and also from the LesserCaucasus and Talysh region (SE Azerbaijan) (Mon-chenko 2003). Some of those records (cf. notes on thedistribution of E. roseus at the beginning of this sec-tion) could refer to E. roseus (unless E. roseus turnsto be a younger synonym of E. agiloides). Interesting-ly, Monchenko (2003) found E. agiloides in a springnear Mountain Chatyr-Dag (South Crimea, Crimean
Mountains), whose relation to E. roseus needs to beclarified.
Finally, we would like to comment a feature, thesetulation pattern of P4 coxopodal seta, which was given high diagnostic value in the Eucyclops serrula-tus group (Alekseev et al. 2006, Alekseev and Defaye2011). The seta can be homonomously (holotype of E. roseus – Fig. 2l in Ishida 1997), or heteronomouslysetulose (E. roseus in Crimea, see Fig. 3D). Also, thelack of setulation (“gap”) on the outer margin at leastin proximal half of the seta is used as a diagnosticcharacter in some species (e.g. E. serrulatus s. str.).This state is also present in some specimens of E. roseus (Crimea, see Fig. 3D; South Korea – Fig.209H in Chang 2009). We observed intra-populationvariation of this feature (setules can be absent or present on the outer margin in the proximal half of P4coxopodal seta) in specimens from South Iraq (Al Salalmarsh). The character state varied even in one speci-men (left and right seta had different setulation), whichindicate that setulation of P4 coxopodite seta some-times is intraspecifically variable character and shouldbe given less weight in the identification of E. roseus.
HHaabbiittaattss. Lakes, rivers, streams, springs, wells,ponds, puddles, swamps, bogs, salt marshes and estu-aries, from fresh to brackish waters (Chang 2012).
DDiissttrriibbuuttiioonn. Japan, Russian Far East (Primor-skiy), South Korea, China (?), Iraq, Egypt, Sudan,Kenya, Germany, [introduced?], Ukraine (Crimea)[range extension?].
Subfamily Cyclopinae Rafinesque, 1815
AAccaanntthhooccyyccllooppss ttrraajjaannii Mirabdullayev et Defaye, 2002
(Fig. 4A–G)
Acanthocyclops trajani Mirabdullayev et Defaye, 2002: 14–18, Figs24–44; Mirabdullayev and Defaye, 2004: 34–36.
Acanthocyclops americanus: Monchenko, 1974: 235–246; Miracle etal., 2013: 875–878, Figs 3–6.
MMaatteerriiaall eexxaammiinneedd. Ukraine, Crimea: LakeKuchuk-Ajigol, leg. E. Anufriieva, 8 August 2012, onefemale (IBSS); Sevastopol, decorative pool, leg. E.Anufriieva, 23 May 2013, two females (IBSS); Pondnear v. Vladislavovka, leg. E. Anufriieva, 15 April 2013,2 CV copepodids (IBSS).
DDiiaaggnnoossiiss. Total body length without caudal setae:1033 µm; prosome/urosome: 1.55; cephalothoraxlength/width: 1.17; genital double-somite length/width:1.20.
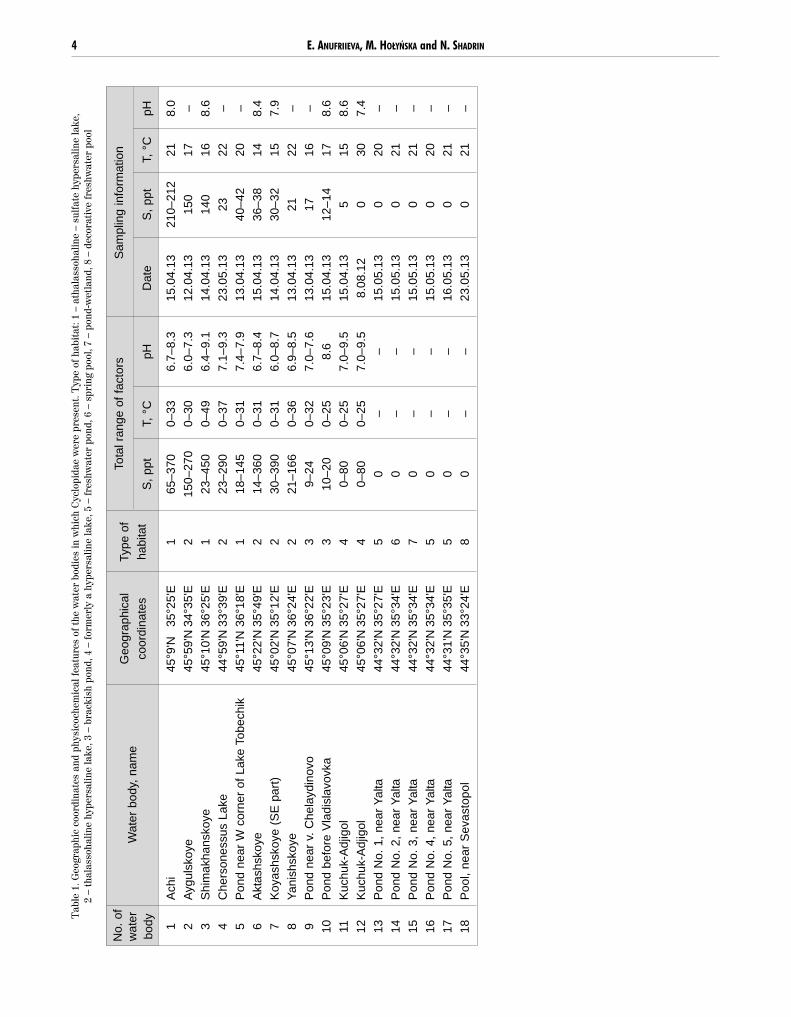
Pediger 5 and genital double-somite naked, integu-mental perforation pattern on dorsal surface as in Fig.4A. Genital double-somite broadly rounded in anteriorpart (Fig. 4A). Anal somite with continuous row of
CYCLOPIDAE IN CRIMEA 9

small spinules on posterior margin and with two dorsalhair-sensilla anteriorly to operculum (Fig. 4B). Longi-tudinal row of hairs on both sides of proctodeum.
Caudal rami parallel and naked, 4.3 times as long aswide. Spinules present at implantation of posterolater-al (III) caudal seta. Caudal setae with homonomoussetulation. Terminal accessory (VI) seta 1.9 times aslong as posterolateral (III) seta. Dorsal (VII) caudalseta 1.2 times as long as posterolateral (III) seta.
Antennule 17-segmented. Antenna enp2 with 9 se-tae. Antennal coxobasis adorned with few robust spin-ules near lateral margin on frontal surface (Fig. 4C),with long basal spinules next lateral margin, and twogroups of robust spinules on caudal surface (Fig. 4D).Maxilla (Fig. 4E) with 2-segmented endopodite, withtwo and three setae, respectively. Robust seta in frontof claw-like attenuation of basal endite, bearing in mid-dle few spinules on both edges (Fig. 4E).
P1-P4 rami 3-segmented, spine formula 3–4–4–4. P4coupler (Fig. 4F) with one row of spinules, P1-P3 cou-plers without ornamentation on caudal surface. Caudalsurface of P4 coxopodite with: intermittent row of spin-ules near distal margin; one row near proximal marginin middle; two oblique rows of longer and thinner spin-ules near, and one sensillum next to lateral margin(Fig. 4F). Leg 4 basipodite with apical hairs. P4 enp3(Fig. 4G) ~ 2.3 times as long as wide. Inner (medial)apical spine slightly longer than outer (lateral) spineand about as long as segment. Outer seta transformedinto spine and inserted at distance of 0.64 length meas-ured from proximal margin of segment. Medial setaealso modified, bearing few long setules proximally andshort spinules in distal half (Fig. 4G). P6 composed ofmedial seta and 2 short spines (Fig. 4A).
Notes on the synonymy of AA.. aammeerriiccaannuuss and AA.. ttrraajjaannii
The taxonomy of A. trajani has been closely con-nected to the debates about the identity of A. ameri-canus (Marsh, 1893); therefore, we provide a shortoverview of the history of research on the latterspecies. Marsh (1893) described an Acanthocyclopsunder the name Cyclops americanus from the littoralof three lakes (Green, Little Green, and Rush) and stag-nant pools in the vicinity of Ripon, Wisconsin State inthe United States. The original diagnosis and drawingsshow a medium-sized (1.2 mm) Acanthocyclops withgenital double-somite seemingly rounded laterally,short (about 3x longer than wide) caudal rami, termi-nal accessory (VI) and posterolateral (III) (“the firstand fourth terminal setae”) caudal setae of “nearlyequal” length, outer apical spine on the third endopo-dal segment of P4 slightly longer than inner one anddistinctly shorter (~ 0.6x) than segment. No type(s)
were designated in the original paper, and the deposi-tory of the Wisconsin material is unknown. The name’americanus’ afterwards appeared in various combina-tions: Marsh (1910) later referred to it as Cyclopsviridis americanus; Gurney (1933) reported on theoccurrence of Cyclops vernalis americanus inBritain; Yeatman (1944) synonymized Marsh’s specieswith Cyclops (Acanthocyclops) vernalis Fischer,1853; Dussart (1969), Monchenko (1974, 2003) andAlekseev et al. (2002) considered A. americanus as a good species and found it in several places in Europe.In a revision of the robustus-vernalis group, Kiefer(1976) examined Marsh’s legacy (wet material fromKansas and Alaska, and drawings labelled by Marsh as’Cyclops americanus’) deposited in the NationalMuseum of Natural History in Washington. Kiefer iden-tified both A. robustus and A. vernalis in this materi-al, which prompted him to synonymize A. americanuspartly with A. robustus and partly with A. vernalis.Kiefer (1976: 104) also made a note of Marsh’s originalillustration of the genital double-somite, suggestingthat the rather indefinite shape of the segment in thedrawing might have been caused by the compression ofthe body by a cover slide. Later works, among others a recent Acanthocyclops monograph by Einsle (1996)followed Kiefer’s classification. Mirabdullayev andDefaye (2002, 2004) focused on the A. robustus-likeforms (genital double-somite widely rounded laterally)made further and significant contributions to the tax-onomy of the robustus-complex. They redescribed thefemale from Lake Mjosa in Norway (the only availablematerial identified by G. O. Sars himself – Kiefer, 1976:96) and the male of A. robustus s. str. from Lapplandin Sweden, and described two new species, A. trajaniand A. einslei, both widely distributed in the Holarctic.These authors re-checked the samples originally iden-tified by Marsh and found four (!) species [A. vernalis,A. brevispinosus (Herrick, 1894) A. einslei and A. trajani] in the vials labeled as ’Cyclops ameri-canus‘. They also pointed out that the morphology ofP4 in Marsh’s illustration corresponds to the one of A. vernalis, while the drawing of the genital-doublesomite (of the robustus-type) should be given lessweight (see Kiefer’s comments above). Recently, Miracle et al. (2013) did major rearrangements in the’robustus-group‘: i) they redescribed A. robustus,based on recently collected material from Maridals-vann (presumed to be the type locality) near Oslo, andsynonym-ized A. einslei with A. robustus; ii) designat-ed the neotype of A. americanus from a materialrecently collected in a pond in Madison (~ 90 km fromthe collecting sites of the original description), and synonym-ized A. trajani with A. americanus. In ouropinion both decisions were wrong, because theseauthors overlooked existing morphological differencesbetween A. robustus sensu G. O. Sars and A. einslei
10 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN
CYCLOPIDAE IN CRIMEA 11
Figure 4. Acanthocyclops trajani Mirabdullayev et Defaye, 2002 from Lake Kuchuk-Ajigol, Crimea, Ukraine, female: (A) pediger 5 and genitaldouble-somite, dorsal view; (B) anal somite and caudal rami, dorsal view; (C) antennal coxobasis, frontal; (D) antennal coxobasis, caudal view; (E) maxillary basis and endopodite, caudal view; (F) leg 4 coupler, coxopodite and basipodite, caudal view; (G) leg 4 endopodite 3, with
heteronomous setulation of medial setae (figured separately) . Scale bars = 50 µm.

Mirabdullayev et Defaye and A. americanus sensuMarsh and A. trajani Mirabdullayev et Defaye. Thepresence/ absence of the spinule group on the antennalcoxobasis next to the exopodal seta is not the onlycharacter by which A. robustus differs from A. einslei.The posterolateral part of pediger 5 is strongly “acute-ly” produced in A. robustus, while this structure isweakly produced in A. einslei (see illustrations inMirabdullayev and Defaye 2002, 2004); the terminalaccessory (VI) caudal seta is short, 1.2–1.7 times aslong as the posterolateral (III) seta in A. robustus, yet1.5–2.0 times as long in A. einslei; the lateral seta onP4 enp3 is inserted at distance of 0.6–0.7 segmentlength in A. robustus, yet 0.7–0.8 segment length in A. einslei. As to the male morphology, the P6 has a medial spine that is shorter than the middle seta andabout half as long as the lateral seta in A. robustus(differing from A. trajani as well, where the medialspine is slightly longer than the middle seta), while themedial spine much longer than middle seta, and almostas long as the lateral seta in A. einslei. We think thatspecimens from Lake Maridalsvann, used by Miracle etal. (2013) as the “reference point” were conspecificwith A. einslei but not with A. robustus. We also thinkthat the designation of the neotype of A. americanus,made by Miracle et al. (2013), did not fulfill at least twoconditions of a valid neotype designation:
“the author’s reasons for believing the name-bearing type specimen(s) (i.e. holotype, or lectotype, orall syntypes or prior neotype) to be lost or destroyed,and the steps that had been taken to trace it or them”– Miracle et al. (2013) did not mention whether theytried to find and examine the depository of the originalWisconsin material which Marsh’s description wasbased on.
”evidence that the neotype is consistent with whatis known of the former name-bearing type from theoriginal description and from other sources” – Theshort apical spines on the third endopodal segment ofP4 in the original drawing (the longer apical spine ~ 0.6 times as long as segment in the Marsh drawing,yet ~ 0.9 times as long in the neotype), length propor-tion of the outer and inner apical spines (outer spine isslightly longer in the original drawing, while it is short-er in the neotype), as well as similar length of the ter-minal accessory (VI) and posterolateral (III) caudalsetae mentioned in the original diagnosis (seta VI 2.1times as long as seta III in the neotype) indicate that A. americanus sensu Marsh is not conspecific withthe neotype designated by Miracle et al. (2013) for theformer taxon. The “neotype” very likely is conspecificwith A. trajani Mirabdullayev & Defaye.
For the reasons discussed above, we decided not toaccept the classification proposed by Miracle et al.(2013), and keep A. trajani as valid name, and consid-er A. americanus (Marsh, 1893) as nomen dubium.
HHaabbiittaattss. Acanthocyclops trajani occurs in dif-ferent kinds of water bodies including lakes, ponds,estuaries, alkali-marsh, and rice fields. Salinity range:0–80 ppt, temperature range: 10–30°C.
DDiissttrriibbuuttiioonn. Europe (including also Scandinavia),Russia (including Siberia), North Africa (Algeria,Tunisia, Egypt), Middle East (Iraq – unpublished dataof H. H. Mohammed and M. Hołyńska), West Asia(Iran), Central Asia (Uzbekistan, Kazakhstan), andNorth America [Canada, U.S.A, Mexico(?) Mirabdul-layev and Defaye 2002, Miracle et al. 2013]. The previ-ous records of A. americanus in Ukraine (Monchenko2003, Anufriieva and Shadrin 2012) likely refer to A. trajani.
DDiiaaccyyccllooppss bbiisseettoossuuss (Rehberg, 1880)(Fig. 5A–I)
Cyclops bisetosus Rehberg, 1880: 543–544; Gurney, 1933: 227–231;Diacyclops bisetosus: Dussart, 1969: 149–152, Fig. 69; Mon-chenko, 1974: 284–290.
MMaatteerriiaall eexxaammiinneedd. Ukraine, Crimea: Lake Shi-makhanskoye, leg. E. Anufriieva, 14 April 2013, onefemale and one CV copepodid (IBSS); Lake Yanish-skoye, leg. E. Anufriieva, 13 April 2013, three femalesand one CIV copepodid (IBSS); Lake Aygulskoe, leg. E.Anufriieva, 12 April, 2013, one male (IBSS); pond nearv. Chelaydinovo, leg. E. Anufriieva, 13 April 2013, threefemales and two CV copepodids (IBSS); Tobechik, pondnear W corner of Lake Tobechik, leg. E. Anufriieva, 13April, 2013, two females and two males (IBSS); LakeKoyashskoye, leg. E. Anufriieva, 14 April 2013, onefemale (IBSS).
Species identification was based on P5 morphology(Fig. 5A), segmentation of the antennule (17-segment-ed) and swimming legs (P1–P4 three-segmented),length proportion of the apical spines on the thirdendopodal segment of P4 (inner spine longer than out-er one) (Fig. 5B), and length proportion of the terminalaccessory (VI) and posterolateral (III) caudal setae(subequal, or seta VI slightly shorter).
While other characters were stable, we found a sur-prisingly great variation in the shape of the seminalreceptacle (Fig. 5C–I). The posterior part most often isa large sac. In the anterior part two horn-like process-es were formed in almost every specimen, the exten-sion of the which (and the volume of the anterior part)varied between the specimens from different lakes, andeven within specimens from the same sample (e.g.Chelaydinovo, pond) (Fig. 5D, F, I). Notes on the pres-ence of the anterolateral processes can be found inevery monograph of the European cyclopid fauna, yetwe failed to find any illustration of the variation of thisfeature. We speculate that the observed variationmight show the different stages of the formation
12 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN

CYCLOPIDAE IN CRIMEA 13
Figure 5. Diacyclops bisetosus (Rehberg, 1880), female: (A) pediger 5, Lake Shimakhanskoye; (B) leg 4 endopodite 3, Lake Shimakhanskoye;Seminal receptacle: (C) Lake Shimakhanskoye; (D, F) I pond near v. Chelaydinovo; (E) Lake Yanishskoye; (G) Lake Koyashskoye; (H) Tobechik
(pond near west corner of the lake). Scale bars = 50 µm.

process of the anterolateral horns (i.e. the femalesreached different stages of the process) rather thanreflecting individual-specific difference.
HHaabbiittaattss. Diacyclops bisetosus often occurs insmall ephemeral waterbodies, where it survives the dryperiod in a resting stage (likely both as copepodid Vand adult stage – Dussart 1969). Not rare in brackishand underground waters (cave, spring). Gurney (1933)found it in a tree-hole in England. The species has widetemperature (0–26°C), salinity (0–150 ppt), and pH(4.0–9.6) tolerance.
DDiissttrriibbuuttiioonn. There are numerous records in thePalearctic region, but it was also recorded from Cana-da (Edmonton), Cuba, Chile, Australia (incl. Tasma-nia), and New Zealand (Gurney 1933, Andrew et al.1989, Ustao™lu 2004, Dussart and Defaye 2006).
MMeessooccyyccllooppss ppeehhppeeiieennssiiss Hu, 1943(Fig. 6A–I)
Mesocyclops Leuckarti pehpeiensis Hu, 1943: 124–126, Fig. C;Mesocyclops pehpeiensis: Dussart and Fernando, 1988: 249–250,Figs 24–27; Guo, 2000: 34–40, Figs 2–4; Hołyńska et al., 2003:138–141, Fig. 57.
Mesocyclops ruttneri Kiefer, 1981: 178–180, Fig. 14; Reid, 1993:177–183, Figs 3–5.
MMaatteerriiaall eexxaammiinneedd. Ukraine, Crimea, LakeKuchuk-Ajigol, leg, E. Anufriieva, 8 August 2012, onefemale (IBSS).
The species has been described several times,therefore our diagnosis (based on the specimen fromLake Kuchuk-Ajigol) only mentions the most importantspecies distinguishing characters.
Body length without caudal setae 1210 µm; pro-some/urosome: 1.7; cephalothorax length/width: 1.2.
Pediger 5 (Fig. 6A) without lateral hairs. Genitaldouble-somite 1.2 times as long as wide, no hairs. Seminal receptacle (Fig. 6B) with wide lateral arms,transverse duct-like structures meet at acute angle (V-shape) next to horseshoe-shaped copulatory pore,copulatory duct long and slightly curved right. Cau-dal rami (Fig. 6C) 3.6 times as long as wide, with-out medial hairs. Spinules present at implantation of anterolateral (II) and posterolateral (III) caudalsetae. Terminal caudal setae with homonomous setula-tion. Relative lengths of terminal caudal setae from ter-minal accessory (VI) to posterolateral (III): 2.6; 5.8;4.13; 1.0.
Antennule 17-segmented, hyaline membrane on lastsegment with one large notch. Antenna enp2 with 7setae. Antennal coxobasis with long longitudinal row ofspinules (24) on the frontal surface, no spinules next toinsertion of exopodal seta (Fig. 6D). The spinule pat-tern on caudal surface of antennal coxobasis (Fig. 6E)composed of: oblique row of tiny spinules near medial
edge of segment; row near implantation of medialsetae; few spinules near distal margin; patch at medio-proximal angle; longitudinal row (11) near lateral mar-gin, and oblique row (8) more proximally.
P1 (Figs 6F–H) basipodite without medial spine.Medial expansion of basipodite pilose in P1–P3 yetnaked in P4. P4 coupler bearing two large (longer thanwide) acute protuberances (Fig. 6I). P4 enp3 3.0 timesas long as wide; inner apical spine 1.2 times as long asouter one; outer margin of inner apical spine withmany teeth. P5 (Fig. 6A) as typical of the genus; apicalseta 1.3 times as long as medial spiniform seta.
HHaabbiittaattss. Eurytopic species, found also in ricefield, green-house water tank, artificial water contain-ers, and wells (Reid 1993, Hołyńska et al. 2003,Suárez-Morales et al. 2005).
Geographical distribution: it is native in East-,South-, and Central Asia (Kazakhstan Uzbekistan),and probably introduced in U.S.A (District of Columbia,Mississippi and Louisiana), Cuba (Habana Province)and Mexico (Chiapas, Pacific coast) (Reid 1993, Mirab-dullayev 1996, Guo 2000, Suárez-Morales et al. 2005,Dussart and Defaye 2006). In Europe it was only known(under the name Mesocyclops ruttneri Kiefer, 1981,a younger synonym of M. pehpeiensis) from warmwater basin in the botanical garden in Vienna. TheCrimean record is the first finding of the species in a natural water body in Europe.
MMeessooccyyccllooppss iissaabbeellllaaee Dussart et Fernando, 1988(Fig. 7A–J)
Mesocyclops isabellae Dussart et Fernando, 1988: 246–248, Figs12–17; Hołyńska et al., 2003: 174–176, Fig. 74.
MMaatteerriiaall eexxaammiinneedd. Ukraine, Crimea, pond nearvillage Chelaydinovo, leg. E. Anufriieva, 13 April 2013,one female (IBSS).
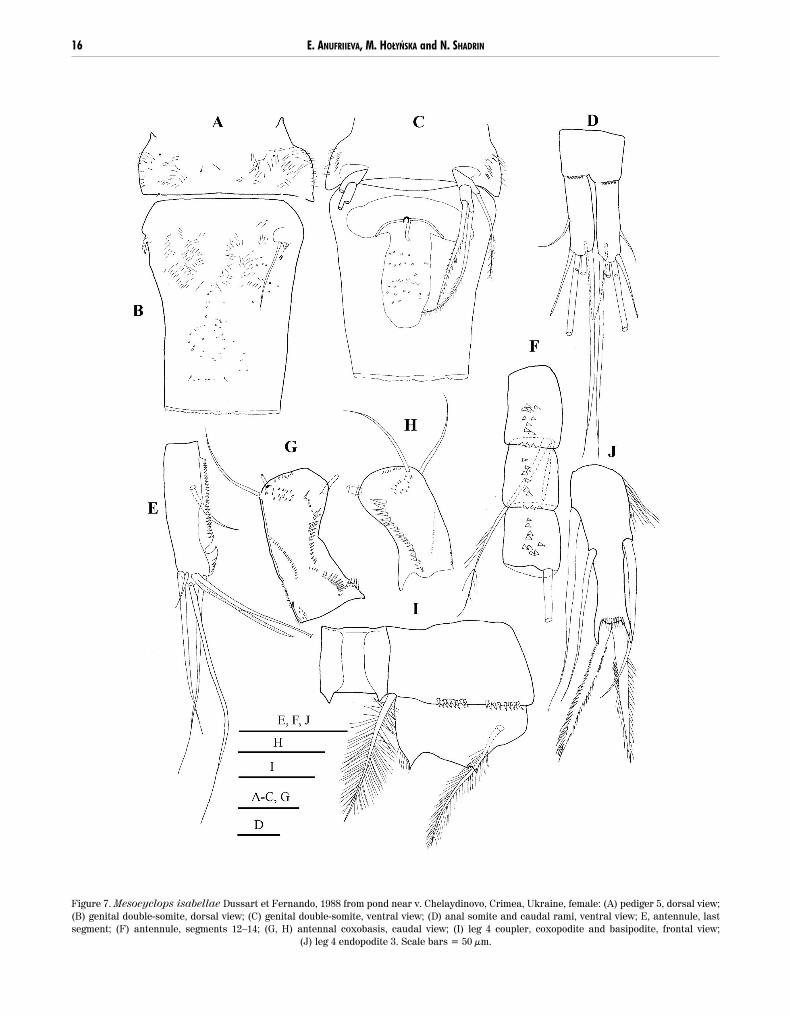
DDiiaaggnnoossiiss. Pediger 5 dorsally and laterally pilose(Fig. 7A). Genital double-somite with shallow integu-mental pits ventrally, and dorsally pilose in anteriorhalf (Fig. 7B). Seminal receptacle with wide lateralarms, transverse duct-like structures meet at straightangle next to copulatory pore, copulatory duct short(Fig. 7C). Posterior margin of anal somite with spinuleson ventral surface only (Fig. 7D). Caudal rami (Fig. 7D)3.5 times as long as wide, without hairs on inner mar-gin. No spinules at implantation of anterolateral (II)and posterotlateral (III) caudal setae. Length propor-tion of terminal caudal setae from terminal accessory(VI) to posterolateral (III): 3.0; broken seta; 4.2; 1.0.Dorsal (VII) caudal seta 1.1 times as long as posterolat-eral (III) seta.
Antennule 17-segmented; serrate hyaline mem-brane of terminal segment, extending beyond insertion
14 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN

CYCLOPIDAE IN CRIMEA 15
Figure 6. Mesocyclops pehpeiensis Hu, 1943 from Lake Kuchuk-Ajigol, Crimea, Ukraine, female: (A) leg 5; (B) seminal receptacle; (C) anal somiteand caudal rami, ventral view; (D) antennal coxobasis, frontal; (E) antennal coxobasis, caudal view; (F) leg 1 coxopodite and basipodite, frontal
view; (G) leg 1 exopodite; (H) leg 1 endopodite; (I) leg 4 coupler, frontal view. Scale bars = 50 µm.
16 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN
Figure 7. Mesocyclops isabellae Dussart et Fernando, 1988 from pond near v. Chelaydinovo, Crimea, Ukraine, female: (A) pediger 5, dorsal view;(B) genital double-somite, dorsal view; (C) genital double-somite, ventral view; (D) anal somite and caudal rami, ventral view; E, antennule, lastsegment; (F) antennule, segments 12–14; (G, H) antennal coxobasis, caudal view; (I) leg 4 coupler, coxopodite and basipodite, frontal view;
(J) leg 4 endopodite 3. Scale bars = 50 µm.

of medial seta of segment, with one large subapicalnotch (Fig. 7E). Spinules present on antennular seg-ments 1, 4–5, and 7–14 (Fig. 7F). Antenna enp2 with 7 setae. Spinule ornamentation on caudal surface ofantennal coxobasis is similar to that in M. pehpeien-sis but: spinules (19, 21) in longitudinal row near later-al margin distally conspicuously increasing in size(Figs 7G, H); and additional group of tiny spinules pres-ent between longitudinal and proximal oblique rows(Fig. 7G).
P1 basipodite lacking medial spine. P4 couplernaked on frontal and caudal surfaces (Fig. 7I showsfrontal surface), distal margin bearing two small andacute protuberances. Medial expansion of P4 basi-podite bearing apical hairs and transverse row of hairson caudal surface, near proximal margin. P4 enp3 (Fig.7J) 2.9 times as long as wide, medial and lateral apicalspines subequal, lateral edge of medial (inner) spinewith many fine spinules. Leg 5 (Fig. 7C) typical ofgenus; apical seta 1.3 times as long, and lateral seta 0.9times as long as spiniform medial seta.
NNootteess. Mesocyclops leuckarti (Claus, 1857), a common species in European freshwaters, can be distinguished from both Mesocyclops species hererecorded by the wide, short and sinuously curved cop-ulatory duct, and the spinular ornamentation of theantennule (spinules present on segments 1, 4–5, 7–10,and 12–13).
HHaabbiittaattss. Lakes, reservoirs, temporary pools,marshes. Temperature tolerance: 10 to 39.5°C, Salinitytolerance: 7 to 17 ppt (Mohammed et al. 2008). Mo-hammed et al. (2011) observed diapause in copepodidIV and V instars found at a depth of 10–20 cm duringwinter (12–14°C) in southern Iraq. This species cannotreproduce if the temperature drops below 10°C (Mo-hammed et al. 2008).
DDiissttrriibbuuttiioonn. South (Sri Lanka, India, Nepal,Bangladesh) and West Asia (Iraq) (Hołyńska et al.2003, Mohammed et al. 2008), Ukraine (Crimea)[range extension?].
DISCUSSION
Alien cyclopid species in Crimea and Europe
We found three cyclopid species in Crimea (i.e.:Mesocyclops pehpeiensis, M. isabellae, and Eucy-clops roseus) which had not yet been reported fromUkraine. The findings of Acanthocyclops trajani inoligohaline- and fresh waters in East and South Crimeavery likely are not new records, because this samespecies could be reported as ’Acanthocyclops ameri-canus’ from different areas of Ukraine (Monchenko1974, 2003) including Crimea (Anufriieva and Shadrin2012).
It is also possible that Monchenko’s report (2003)on the occurrence of Eucyclops agiloides in Chatyr-Dag (Crimean Mts.) in fact refers to the species that weidentified as E. roseus from the East and SouthCrimean coast. Considering that both the fresh-(Monchenko 2003) and saline/hypersaline waters (ourdata) have already been sampled in Crimea ratherextensively, we suppose that E. roseus is not very com-mon in the peninsula. The only other European recordof E. roseus is from northern Germany (a stream inOldenburg) (Ishida 1997). In a review of the world dis-tribution of the E. serrulatus-group, which E. roseusbelongs to, Alekseev and Defaye (2011) did not mentionany other European localities for E. agiloides roseus(they considered E. roseus as a subspecies of E. agi-loides). They also stated that E. agiloides is a tropi-cal/subtropical species, and E. agiloides roseus wasintroduced to northern Germany by human agency. Asto Crimea, we speculate that E. roseus reached thepeninsula via Caucasia from Western Asia (Iran) in a gradual process (range extension) rather than as a result of human-mediated introduction because thereis no aquaculture activities in the studied water bodies.Monchenko’s report (2003) on the occurrence of ’E. agiloides‘ in the Lesser Caucasus and Talyshregion (SE Azerbaijan) might support a southeasterndispersal route.
There are no records of the genus Mesocyclops inthe earlier fauna studies of Crimea. This predominant-ly (sub)tropical genus is also absent in the Iberianpeninsula and in southern Italy and Greece (Van deVelde 1984). The first report for Crimea of the occur-rence of M. leuckarti Claus, 1857, one of the common-est species in the temperate/cold temperate waters ofEurasia (incl. large part of Ukraine), was that from theSimferopol Reservoir in 1955–1960 (Tseeb 1961).Mesocyclops leuckarti was probably the first aliencyclopid species recorded in Crimea; it could have beenaccidentally introduced into the Simferopol Reser-voir, or transported by the North Crimean Canal(Tseeb 1961, Monchenko 2003). In this survey we foundtwo tropical/subtropical species (M. isabellae and M. pehpeiensis) which are alien species not only inCrimea but also in Europe. A range extension cannotbe excluded in these taxa either, yet the closest knownoccurrences of these species are from fairly distantregions: northern Karakalpakstan in Uzbekistan (ca. 1800 km) in M. pehpeiensis, and southern Iraq(ca. 1900 km) in M. isabellae. In the sites where M. pehpeiensis (recently desalinated fresh lake) and M. isabellae (a brackish pond) live, there is nohuman agency that could lead to introductions of alien species. Long-distance transportation by birdsseems to be more plausible explanation for the ap-pearance of M. isabellae and M. pehpeiensis inCrimea.
CYCLOPIDAE IN CRIMEA 17

Crimea lies in the crossroads of important birdmigration routes between the Western and EasternPalearctic, Africa and Asia (Shadrina et al. 2002,Maclean et al. 2009, Khomenko and Shadrin 2009),which might facilitate a connection of the crustaceanfauna of Crimea to Western and Central Asia, andSiberia. The case of the anostracan crustacean,Artemia urmiana Günther, 1899 (Khomenko andShadrin 2009, Shadrin et al. 2012) is an example forsuch distribution pattern. Artemia urmiana occurs inLake Urmia (Iran), in some hypersaline lakes inCrimea and in few soda lakes in Altai (Siberia); allthese sites lay along the same migratory route of sever-al migrant waterfowl species. There are at least 3 birdspecies (Tadorna tadorna (Linneaus, 1758), Tringatotanus (Linneaus, 1758) and Recurvirostra avoset-ta (Linneaus, 1758)) which might transport the brineshrimp cysts, as well as resting stages of other aquaticanimals between Western (Iran) and Central Asia andCrimea (Khomenko and Shadrin 2009). The presence ofresting stage that might survive such a long travel hasbeen demonstrated in M. isabellae (CIV and CVstages) (Mohammed et al. 2011). We are not aware ofany study proving the occurrence of resting stage in M. pehpeiensis, yet this Oriental species has beenfound in several localities (ricefields, urban ponds) inNorth and Central America, outside its native distribu-tional range, which hints that some dormant stageresistant to lack of free water might exist. Interesting-ly, in the western hemisphere also birds were presum-ably transporting two Neotropical Mesocyclopsspecies [M. longisetus (Thiébaud, 1912) and M. vene-zolanus Dussart, 1987] to arctic Canada (Yukon Terri-tory) (Reid and Reed 1994).
Eucyclops roseus, M. isabellae and M. pehpeien-sis are thermophilic species, therefore occurrence ofthese cyclopids might be an indication of the effect ofthe climate warming on the copepod fauna of Ukraine(and Europe), although we need to collect more data toverify this hypothesis in a wider time scale.
Not only the biological properties of the potentialcolonizers determine the success in a new environ-ment, but also the ’immunity‘ of the invaded ecosystem;destabilized or new ecosystems in the shallow waterbodies have very low ecological resilience (Shadrin2000, Gomoiu et al. 2002). It should be noted that in ourcase all alien species were found in newly formedwater bodies where stable communities did not exist.The pond in the quarry of the former Kamysh-Burunore mine near the village of Chelyadinovo, where M. isabellae was found, was naturally filled with rain-water in the last 10–12 years. Lake Kuchuk-Adjigol,investigated by us since 2003, was a hypersaline lakebefore 2012 and it now hosts M. pehpeiensis, E. ro-seus and A. trajani. A dramatic desalination occurredin 2012 due to leakage from a freshwater reservoir
filled from the North Crimean Canal. All the hyperha-line fauna previously existing there disappeared,which created empty niches for the new incomers. Theother site of E. roseus is a decorative artificial poolnear Kazachija Bay.
Mesocyclops pehpeiensis and M. isabellae areboth predator forms which naturally influence the den-sity and species composition of their preys (cladocer-ans, rotifers, and dipteran larvae – Dieng et al. 2003,Nagata and Hanazato 2006, Mohammed et al. 2008,Sarma et al. 2013). These alien predator cyclopids maychange the zooplankton composition in their new habi-tats, further destabilizing ecosystems and openingdoors for new incomers. We need to mention thatalthough the problem of biological invasions is becom-ing a rapidly-growing research area, there is a largegap in the studies of the invasions of cyclopids inEurope and worldwide. In a recent review of the inva-sions of exotic species in Europe there is no informa-tion pertaining to Cyclopoida (Leppäkoski et al. 2002,Nentwig 2009). In the Global Invasive Species Database(http://www.issg.org/database) we have not found anydata on invasive cyclopoids either. This gap should befilled in the near future.
Species richness of Crimean Cyclopidae
Obviously, not all species of Cyclopidae occurring inCrimea have been recorded yet, so the question is towhat degree the cyclopid fauna of the peninsula hasbeen explored? There is a fairly tight correlationbetween the number of samples analyzed and the num-ber of observed species (Pesenko 1982, Shen et al.2003, Brucet et al. 2009).
Various types of functions are used to describe therelationship between the sampling effort (the numberof identified specimens, or the number of samples) andthe observed species richness; the power and logarith-mic functions are the most used. Relying upon our ownand Monchenko’s (2003, Table 28) data, we found thatthe species accumulation curve for the whole fauna(including both fresh and saline/hypersaline waters)(Fig. 8A) is best described by the following logarithmicfunction (R = 0.976, p <0.0001):
Y = 12.09 ln(X) – 26.08 (1)
where Y – the number of observed species, ln(X) –logarithm of the number of analyzed samples.
This function corresponds to one of the many possi-ble orderings of 220 samples. We analyzed 10 moreaccumulation curves with 10 different orderings of thesamples. The species richness for 500 samples, apply-ing different orderings of the samples, was estimatedas 47–50 species for all habitats of Crimea. Taking 44
18 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN

species (including the three alien taxa here reported)as a current estimate of the diversity of Cyclopidae inCrimea (both fresh- and saline waters), the known fauna is nearly 90% of the expected species richness.That indicates that the cyclopids of Crimea are wellexplored in general, but this may not necessarily holdtrue if the hypersaline fauna alone is considered. Infact, when similar analyses were carried out only forthe samples from hypersaline waters, the species accu-mulation curve (Fig. 8B) was described by a logarith-mic function (R = 0.976, p <0.0005):
Y = 2.503 ln(X) – 7.683 (2)
Eleven different random permutations of the sam-ples yielded 6–8 species, if very large sample (500)would be analyzed. This implies that we only found halfor two thirds of the cyclopid species living in hyper-saline habitats.
How much can we trust these calculations? Weshould remember that the species accumulation curvemethod provides realistic estimate of the species rich-ness, when species are distributed uniformly or ran-domly in space and time (Pesenko 1982), which israther rare in nature, and cannot be expected in sucha heterogeneous region as Crimea either. Therefore,the values given above are only rough estimates of theexpected diversity.
CYCLOPIDAE IN CRIMEA 19
Figure 8. Species accumulation curves: (A) fresh and saline/ hypersaline water bodies pooled; (B) hypersaline waters only.
There is a non-parametric approach to estimateasymptotic species richness, Chao2, which is based onthe frequencies of rare species in the original samplingdata (Chao et al. 2009). The method is robust to non-independence arising from small-scale spatial aggrega-tion of species occurrences. But if we have more than60–80 samples, the different calculation methods givevery close results (Gotelli and Colwell 2011). The Chao2estimate for the expected species richness in hyper-saline waters (2 species are singleton and 1 species isdoubleton) is 6, which is exactly the lower bound of theestimates provided by the species accumulation curvemethod.
Both values of the observed and expected speciesrichness indicate that only few species of Cyclopidaecould adapt to hypersaline environment. Our cyclopidfindings agree with previous observations on thesetaxa, in fact, Diacyclops bisetosus and Cyclops fur-cifer have already been recorded from waterbodieswith salinity about 100 ppt in Germany and Spain,respectively (Zernov 1949, Hammer 1986). Acanthocy-clops americanus (syn. A. trajani) has been report-ed from high-salinity (80 ppt) waters in Crimea (Anufri-ieva and Shadrin 2012) and Eucyclops serrulatusFischer, 1851 was found in waters with 55 ppt (Brucetet al. 2009).
CONCLUSIONS
Cyclopidae were present only in few (6%) of thesamples collected from the hypersaline waters of Cri-mea. All the taxa recorded (Eucyclops sp., Cyclopsfurcifer, Acanthocyclops sp. and Diacyclops bise-tosus) have been previously reported from hypersalinewaters. Diacyclops bisetosus was the most frequentspecies in this habitat.
The observed species richness for Crimea is half totwo thirds of the expected (asymptotic) species rich-ness (6–8) in the hypersaline waters, while this valuereaches 90% if the hypersaline and freshwater samplesare pooled.
Our fauna survey also included few brackish andfresh water bodies, some of which were formerly hyper-saline systems. Three alien, Asian/Afro-Asian ther-mophilic species, Eucyclops roseus, Mesocyclopsisabellae and M. pehpeiensis were found in destabi-lized or new ecosystems. Long-term observations couldbe useful to assess if the new colonizers can form persistent populations in Crimea.
ACKNOWLEDGEMENTS
The authors are very grateful to the Distribut-ed European School of Taxonomy (DEST) EC for the

partial financial support for E. Anufriieva to partici-pate in the training programme in Crustacean system-atics with specialisation in Cyclopidae (Copepoda) atthe Museum and Institute of Zoology, Warsaw, Poland. We are grateful to all those colleagues who assisted inthe fieldwork, especially to Mr. O. Yu. Eryomin, for hishelp in the organization and conducting of all expedi-tions. All advices given by our colleagues are kindlyappre-ciated here. Special thanks from the first authorto Museum and Institute of Zoology (Warsaw) for the hospitality during her research visit. We also thank twoanonymous reviewers who helped us to improve ourmanuscript.
REFERENCES
Alekseev, V., Fefilova, E. and H. J. Dumont. 2002. Some note-worthy free-living copepods from surface freshwater inBelgium. Belgian Journal of Zoology, 132: 133–139.
Alekseev, V., Dumont, H. J., Pensaert, J., Baribwegure, D. andJ. R. Van Fleteren. 2006. A redescription of Eucyclopsserrulatus (Fischer, 1851) (Crustacea: Copepoda:Cyclopoida) and some related taxa, with a phylogeny ofthe E. serrulatus-group. Zoologica Scripta, 35: 123–147.
Alekseev, V. and D. Defaye. 2011. Taxonomic differentiationand world geographical distribution of the Eucyclops ser-rulatus group (Copepoda, Cyclopidae, Eucyclopinae), pp.41–72. In: D. Defaye, E. Suárez-Morales and J. C. von Vau-pel Klein (eds.). Studies on Freshwater Copepoda: A Vol-ume in Honour of Bernard Dussart. Crustaceana Mono-graphs 16, Brill, Netherlands.
Andrew, T. E., Cabrera, S. and V. Montecino. 1989. Diurnalchanges in zooplankton respiration rates and the phyto-plankton activity in two Chilean lakes. Hydrobiologia, 175:121–135.
Anufriieva, E. V. and N. V. Shadrin. 2012. Crustacean diversi-ty in hypersaline Chersonessus Lake (Crimea). Optimiza-tion and Protection of Ecosystems, 7: 55–61. (In Russian).
Apostolov, L. G. (ed.) 1999. [Biological and landscape diversi-ty of the Crimea: problems and perspectives]. Sonat, Sim-feropol: 180 pp. (In Russian).
Belmonte, G., Moscatello, S., Batogova, E. A., Pavlovskaya, T.,Shadrin, N. V. and L. F. Litvinchuk. 2012. Fauna of hyper-saline lakes of the Crimea (Ukraine). Thalassia Salentina,34: 11–24.
Brucet, S., Boix, D., Gascon, S., Sala, J., Quintana, X. D.,Badosa, A., Søndergaard, M., Lauridsen, T. L. and E. Jep-pesen. 2009. Species richness of crustacean zooplanktonand trophic structure of brackish lagoons in contrastingclimate zones: north temperate Denmark and Mediter-ranean Catalonia (Spain). Ecography, 32: 692–702.
Chang, C. Y. 2009. Illustrated Encyclopedia of Fauna & Floraof Korea. Vol. 42. Inland-water Copepoda. JeonghaengsaPubl. Co., Seoul, 687 pp. (In Korean).
Chang, C. Y. 2012. Discovery of Halicyclops continentalis(Cyclopidae, Halicyclopinae) from estuaries and saltmarshes on the west coast of South Korea. Animal Sys-tematics, Evolution and Diversity, 28: 12–19.
Chao, A., Colwell, R. K., Lin, C. W. and N. J. Gotelli. 2009.
Sufficient sampling for asymptotic minimum species rich-ness estimators. Ecology, 90: 1125–1133.
Dieng, H., Boots, M., Tuno, N., Tsuda, Y. and M. Takagi. 2003. Life history effects of prey choice by copepods: implications for biocontrol of vector mosquitoes. Jour-nal of the American Mosquito Control Association, 19:67–73.
Dussart, B. H. 1969. Les Copépodes des eaux continentalesd’Europe occidentale II: Cyclopoïdes et Biologie. N. Bou-bée & Cie, Paris, 289 pp.
Dussart, B. H. 1981. Contribution à l’étude des copépodes d’Iran. Crustaceana, 41: 162–166.
Dussart, B. H. and C. H. Fernando. 1988. Sur quelques Meso-cyclops (Crustacea, Copepoda). Hydrobiologia, 157:241–264.
Dussart, B. H. and D. Defaye. 2006. World directory of Crus-tacea Copepoda of inland waters II – Cyclopiformes. Back-huys Publishers, Leiden, 354 pp.
Einsle, U. 1996. Copepoda: Cyclopoida Genera Cyclops,Megacyclops, Acanthocyclops. Guides to the identifica-tion of the microinvertebrates of the continental waters of the world. Vol. 10, SPB Academic Publishing, Amsterdam.83 pp.
El-Shabrawy, G. 2013. Report on the training programme inCrustacean systematics with specialization in Cyclopidae(Copepoda), 2012. Monoculus Copepod Newsletter, 64:5–7.
Gomoiu, M. T., Alexandrov, B., Shadrin, N. and Y. Zaitsev.2002. The Black Sea – a recipient, donor and transit area for alien species, pp. 341–350. In: E. Leppäkoski, S. Gollasch and S. Olenin, (eds.), Invasive Aquatic Speciesof Europe, Distribution, Impacts and Management.Springer.
Gotelli, N. J. and R. K. Colwell. 2011. Estimating species rich-ness, pp. 39–54. In: A. E. Magurran and B. J. McGill (eds.),Biological diversity: frontiers in measurement and assess-ment. Vol. 12. Oxford University Press, Oxford.
Guo, X. 2000. A redescription of Mesocyclops pehpeiensisHu, 1943 and notes on Mesocyclops ruttneri Kiefer, 1981(Copepoda, Cyclopidae). Hydrobiologia, 418: 33–43.
Gurney, R. 1933. British freshwater Copepoda, 3. The RaySociety, London, 384 pp.
Hammer, U. T. 1986. Saline lake ecosystems of the world.Monographiae Biologicae 59. Dr. W. Junk Publishers, Dor-drecht, Netherlands, 616 pp.
Hołyńska, M. 2008. Cyclopoida, pp. 301–311. In: W. Bogdano-wicz, E. Chudzicka, I. Pilipiuk I. and E. Skibińska (eds.),Fauna Polski – charakterystyka i wykaz gatunków [Fau-na of Poland: Checklist and characterization of thespecies]. Muzeum i Instytut Zoologii PAN, Warsaw.
Hołyńska , M., Reid, J. W. and H. Ueda. 2003. Genus Mesocy-clops Sars, 1914, pp. 12–213. In: H. Ueda and J.W. Reid(eds.). Copepoda: Cyclopoida. Genera Mesocyclops andThermocyclops. Vol. 20, in Dumont H. J. F. (ed.) Guides tothe Identification of the Microinvertebrates of the Conti-nental Waters of the World. Backhuys, Leiden.
Hu, Y. T. 1943. Notes on fresh-water copepods from Pehpei,Szechwan. Sinensia, 14: 115–128.
Huys, R. and G. A. Boxshall. 1991. Copepod evolution. Vol. 159.The Ray Society, London, 468 pp.
Ishida, T. 1997. Eucyclops roseus, a new Eurasian copepod
20 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN

and the E. serrulatus-speratus problem in Japan. Japan-ese Journal of Limnology, 58: 349–358.
Ishida, T. 1998. Eucyclops biwensis, a new cyclopoid cope-pod (Crustacea) from Lake Biwa, Japan, with records of a morphologically identical population of the new speciesand Eucyclops roseus from Lake Victoria, Kenya. Bul-letin of the Biogeographical Society of Japan, 53: 23–27.
Khomenko, S. V. and N. V. Shadrin. 2009. Iranian endemicArtemia urmiana in hypersaline Lake Koyashskoe(Crimea, Ukraine): a preliminary discussion of introduc-tion by birds. Branta. Transaction of Azov-Black Seaornithological station, 12: 81–91. (In Russian).
Kiefer, F. 1933. Die freilebenden Copepoden der Binnenge-wässer von Insulinde. Archiv für Hydrobiologie supple-ment, 12: 519–621.
Kiefer, F. 1976. Revision der robustus-vernalis-Gruppe derGattung Acanthocyclops Kiefer (Crustacea, Copepoda)(Mit eingehender Beurteilung des „Cyclops americanusMarsh, 1892”). Beiträge zur naturkundlichen Forschungin Südwestdeutschland, 35: 95–110.
Kiefer, F. 1981. Beitrag zur Kenntnis von Morphologie, Tax-onomie und geographischer Verbreitung von Mesocy-clops leuckarti auctorum. Archiv für Hydrobiologie sup-plement, 62: 148–190.
Koźmiński, Z. 1936. Morphometrische und ökologische Unter-suchungen an Cyclopiden der strenuus-Gruppe. Interna-tionale Revue der gesamten Hydrobiologie und Hydrogra-phie, 33: 161–240.
Lebedinsky, J. 1904. [On fauna of the Crimean caves II].Zapiski Novorosiyskogo obschestva estestvoispytatelei,25: 75–88. (In Russian).
Leppäkoski, E., Gollasch, S. and S. Olenin (eds.). 2002. Inva-sive aquatic species of Europe. Distribution, Impacts andManagement. Kluwer Academic Publishers, Dordrecht,Boston, London, 592 pp.
Maclean, I. M. D., Rehfisch, M. M., Delany, S. and R. A. Robin-son. 2009. The effects of climate change on migratorywaterbirds within the African-Eurasian flyway. ResearchReport No. 486. British Trust for Ornithology, London, 99pp.
Marsh, C. D. 1893. On the Cyclopidae and Calanidae of CentralWisconsin. Transactions of the Wisconsin Academy of Sci-ences, Arts, and Letters, 9: 189–234.
Marsh, C. D. 1910. A revision of the North American species ofCyclops. Transactions of the Wisconsin Academy of Sci-ences, Arts, and Letters, 16: 1067–1134.
Mirabdullayev, I. M. 1996. The genus Mesocyclops (Crustacea:Copepoda) in Uzbekistan (Central Asia). InternationaleRevue der gesamten Hydrobiologie und Hydrographie, 84:93–100.
Mirabdullayev, I. M. and D. Defaye. 2002. On the taxonomy ofthe Acanthocyclops robustus species complex (Copepo-da, Cyclopidae) 1. Acanthocyclops robustus (G. O. Sars,1863) and Acanthocyclops trajani n. sp. Selevinia, 1-4:7–20.
Mirabdullayev, I. M. and D. Defaye. 2004. On the taxonomy ofthe Acanthocyclops robustus species-complex (Copepo-da, Cyclopidae): Acanthocyclops brevispinosus and A. einslei sp. n. Vestnik Zoologii, 38: 27–37.
Miracle, M. R., Alekseev, V., Monchenko, V., Sentandreu, V.and E. Vicente. 2013. Molecular-genetic-based contribu-
tion to the taxonomy of the Acanthocyclops robustusgroup. Journal of Natural History, 47: 863–888.
Mohammed, H. H., Salman, S. D. and A. A. M. Abdullah. 2008.Some aspects of the biology of two copepods: Apocyclopsdengizicus and Mesocyclops isabellae from a pool inGarmat – Ali, Basrah, Iraq. Turkish Journal of Fisheriesand Aquatic Sciences, 8: 239–247.
Mohammed, H. H., Salman, S. D. and A. A. M. Abdullah. 2011.Does predation affect the life style of freshwater Copepo-da, pp. 18–33. In: V. K. Gupta, A. K. Verma and G. D. Singh(eds.) Perspectives in Animal Ecology and Reproduction,8. Daya Publishing House, Delhi.
Monchenko, V. I. 1974. [Cyclopidae Fauna of the Ukraine].Fauna Ukrainii, 27. Naukova Dumka, Kyiv, 450 pp. (InUkrainian).
Monchenko, V. I. 2003. Free-living cyclopoid copepods of Ponto-Caspian basin. Naukova Dumka, Kyiv, 350 pp. (InRussian).
Muratov, M. V., Arkhipov, I. V. and Y. A. Uspenskaya. 1984.Structural evolution of the Crimean Mountains and com-parison with the western Caucasus and the easternBalkan ranges. International Geology Review, 26:1259–1266.
Müller, P. H., Neuman, P. and R. Storm. 1979. Tafeln der mathematischen Statistik. VEB Fachbuchverlag, Leipzig,272 pp.
Nagata, T. and T. Hanazato. 2006. Different predation impactsof two cyclopoid species on a small-sized zooplanktoncommunity: an experimental analysis with mesocosms.Hydrobiologia, 556: 233–242.
Nentwig, W. 2009. Handbook of alien species in Europe.Springer, 399 pp.
Oliferov, A. N. and Z. V. Timchenko. 2005. [Rivers and lakes inthe Crimea]. Dolia, Simferopol, 216 pp. (In Russian).
Pesenko, J. A. 1982. [Principles and methods of quantitativeanalysis in faunal studies]. Nauka, Moscow, 278 pp. (InRussian).
Polishchuk, V. V. 1998. Biogeography of continental waterbodies of Ukraine and neighboring territories. 1. HoloceneComplexes. Hydrobiological Journal, 34: 10–35.
Rehberg, H. 1880. Beitrag zur Kenntnis der freilebenden Süss-wasser-Copepoden. Abhandlungen des Naturwissen-schaftlichen Vereins zu Bremen, 6: 533–553 + Pl. VI.
Reid, J. W. 1993. New records and redescriptions of Americanspecies of Mesocyclops and of Diacyclops bernardi(Petkovski, 1986) (Copepoda: Cyclopoida). Bijdragen totde Dierkunde, 63: 173–191.
Reid, J. W. and E. B. Reed. 1994. First records of two Neotrop-ical species of Mesocyclops (Copepoda) from Yukon Ter-ritory: Cases of passive dispersal? Arctic, 47: 80–87.
Sarma, S. S., Jiménez-Contreras, J., Fernández, R., Nandini,S. and G. García-García. 2013. Functional responses andfeeding rates of Mesocyclops pehpeiensis Hu (Copepo-da) fed different diets (rotifers, cladocerans, alga andcyanobacteria). Journal of Natural History, 47: 841–852.
Sars, G. O. 1909. Zoological results of the third Tanganyikaexpedition, conducted by Dr. W. A. Cunnington, F.Z.S.,1904–1905. Report on the Copepoda. Proceedings of theZoological Society of London, 31–77.
Shadrin, N. V. 2000. The species introducers in the Black Seaand in the Sea of Azov: The causes and the consequences,
CYCLOPIDAE IN CRIMEA 21

22 E. ANUFRIIEVA, M. HOŁYŃSKA and N. SHADRIN
pp. 76–90. In: G. G. Matishov (ed.), Species Introducers in the European Seas in Russia. KSC RAS, Apatity. (InRussian).
Shadrin, N. V. 2009. The Crimean hypersaline lakes: towardsdevelopment of scientific basis of integrated sustainablemanagement. In: Proceedings of 13th World Lake Confer-ence, Wuhan, China, 1–5 November, 2009: http://www.ilec.or.jp/eg/wlc/wlc13/wlc13papers1.html; http://wldb.ilec.or.jp/data/ilec/WLC13_Papers/S12/s12-1.pdf
Shadrin, N., Anufriieva, E. and E. Galagovets. 2012. Distribu-tion and historical biogeography of Artemia Leach, 1819(Crustacea: Anostraca) in Ukraine. International Journalof Artemia Biology, 2: 30–42.
Shadrina, S. N., Solovjov, A. I., Shadrin, N. V. and N. V. Shik.2002. [Birds wintering in Sevastopol Bay as a resource forsustainable development of tourism], pp. 178–185. In: N. V.Shadrin (ed.) Problems of sustainable development ofcoastal cities and towns. Aquavita Publisher, Sevastopol.(In Russian).
Shen, T. J., Chao, A. and C. F. Lin. 2003. Predicting the num-ber of new species in further taxonomic sampling. Ecolo-gy, 84: 798–804.
Sovinsky, V. K. 1891. [Materials on the fauna of crustaceans ofsouth-western region]. Zapiski Kievskogo obschestvaesyestvoispytatelei, 11: 107–177. (In Russian).
Stoch, F. 2007 [2006]. Crustacea Copepoda Cyclopoida, pp.93–97. In: S. Ruffo and F. Stoch (eds.) Checklist and distri-bution of the Italian fauna. Memorie del Museo Civico diStoria Naturale di Verona, 2 serie, Sezopne Sceinze dellaVita 17, with data on CD-ROM.
Suárez- Morales, E., Gutiérrez-Aguirre, M. A., Torres, J. L.and F. Hernández. 2005. The Asian Mesocyclops peh-peiensis Hu, 1943 (Crustacea, Copepoda, Cyclopidae) in
Southeast Mexico with comments on the distribution ofthe species. Zoosystema, 27: 245–256.
Tai, A. and G. Chen. 1979. Cyclopoida, pp. 310–420. In: C-JShen et al. (eds.) Fauna Sinica, Crustacea, FreshwaterCopepoda. Science Press Peking.
Tseeb, J. J. 1947. [Zoogeographical essay and history of theCrimean hydrofauna]. Uchenye zapiski Orlovskogo gos-pedinstituta, seriaa estestvennyhk i khimicheskikh nauk,2: 67–112. (In Russian).
Tseeb, J. J. 1961. [Hydrobiological characterization of theCrimean water bodies], pp. 230–242. In: Small water bodies of plain regions of the USSR and their use. VGBO,Chisinau. (In Russian).
Ulomsky, S. N. 1955. [Plankton of inland water bodies in theCrimea and its biomass]. Trudy Karadagskoy biostantsii,13: 131–162. (In Russian).
Ustao™lu, R. M. 2004. A check-list for zooplankton of Turkishinland waters. E.U. Journal of Fisheries & Aquatic Sci-ences, 21: 191–199.
Van de Velde, I. 1984. Revision of the African species of thegenus Mesocyclops Sars, 1914 (Copepoda: Cyclopidae).Hydrobiologia, 109: 3–66.
Velichkevich, A. I. 1931. [On ecology of the malaria mosquitoesspreading on the southern coast of the Crimea]. Parasito-logichesky sbornik Zoologicheskogo muzeya AkademiiNauk, 2: 273–314. (In Russian).
Yeatman, H. C. 1944. American cyclopoid copepods of theviridis-vernalis group (including a description ofCyclops carolinianus n. sp.). The American MidlandNaturalist Journal, 32: 1–90.
Zernov, S.A. 1949. [General Hydrobiology]. Akademia NaukUSSR, Moscow, 587 pp. (In Russian).
Received: mm xx, 201xAccepted: mm xx, 201x




















