
Critical centrifugal forces induce adhesion rupture or structural reorganization in cultured cells
12
Cell Motility and the Cytoskeleton 33:276-287 (1996) Critical Centrifugal Forces Induce Adhesion Rupture or Structural Reorganization in Cultured Cells Olivier Thoumine, Albrecht Ott, and Daniel Louvard Section de Recherche, lnstitut Curie, Paris Cedex, France Cultured epithelial cells were exposed to accelerations ranging from 9,000 to 70,OOOg for time periods of 5, 15, or 60 min, by centrifugation in a direction tangential to their plastic substrate. Three regimes describe the cellular response: (1) Cell morphology and density remain unaltered at forces below a threshold of about lop7 N; (2) Between this critical force and a second threshold of about 1.5 lop7 N, the number of adherent cells decreases exponentially with time and acceleration, with no alteration of cell morphology. This behavior can be modeled by a constant probability of detaching and by an exponential distribution of cell-to-substrate adhesive forces; (3) Past the second threshold, cells that are still adherent exhibit elongated morphologies, the degree of elongation increasing linearly with the force. The fact that cells lose their vinculin-rich focal contacts past the first threshold and that cells cultured on gelatin-coated plastic show an increased resistance to detachment suggests a rupture of cell-to-substrate adhe- sions upon centrifugation. Immunofluorescent labeling of cells for actin and tu- bulin shows a reorganization of the cytoskeleton upon centrifugation, and treat- ment of cells with the drugs cytochalasin D and nocodazole demonstrates that cytoskeletal elements are actively involved in the structural deformation of cells past the second acceleration threshold, microtubules and microfilaments playing antagonistic roles. 0 1996 Wiley-Liss, Inc. Key words: centrifugation, epithelial, cell deformation, cytoskeletal organization, tangential accelera- tion, adhesive force INTRODUCTION Cellular morphogenesis is essential to embryonic development and pathological situations such as cancer metastasis and would healing. Cell shape is determined by several types of forces acting on the cell structure [Penman, 19951. These forces include osmotic [Al- brecht-Buehler and Bushnell, 19821 and hydrostatic [Be- reiter-Hahn, 19871 pressures acting on the membranes, prestress integrated by the cytoskeleton [Sims et al., 1992; Ingber, 19931, adhesive forces conveyed between the cell structure and the external environment through cell-cell and cell-matrix transmembrane mechano-chem- ical connections such as focal contacts [Luna and Hitt, 1992; Wang et al., 19931, and other external physical forces (e.g. gravity, shearing stresses in the circulation). Changes in one or more of these factors alter the overall 0 1996 Wiley-Liss, Inc. force balance and may affect cellular morphology and structural organization. Experiments using environments of reduced or in- creased gravity have shown that gravitational forces could influence the oriented growth of plants [Sack, 19911, cytoplasmic streaming in plant cells [Wayne et al., 19921, animal embryogenesis [Suda et al., 19941, adult physiology [Kirsch et al., 19841, as well as mam- malian cell proliferation [Cogoli et a]., 19841. In such biological systems, gravitational fields alter the force distribution by acting differentially on components of different mass densities. Specific connections between Received November 23, 1995; accepted November 30, 1995 Address reprint requests to Olivier Thoumine, Ph.D., Section de Re- cherche, Institut Curie, 75231 Paris Cedex 05, France.
Transcript of Critical centrifugal forces induce adhesion rupture or structural reorganization in cultured cells

Cell Motility and the Cytoskeleton 33:276-287 (1996)
Critical Centrifugal Forces Induce Adhesion Rupture or Structural Reorganization in
Cultured Cells
Olivier Thoumine, Albrecht Ott, and Daniel Louvard
Section de Recherche, lnstitut Curie, Paris Cedex, France
Cultured epithelial cells were exposed to accelerations ranging from 9,000 to 70,OOOg for time periods of 5 , 15, or 60 min, by centrifugation in a direction tangential to their plastic substrate. Three regimes describe the cellular response: (1) Cell morphology and density remain unaltered at forces below a threshold of about lop7 N; (2) Between this critical force and a second threshold of about 1.5 lop7 N, the number of adherent cells decreases exponentially with time and acceleration, with no alteration of cell morphology. This behavior can be modeled by a constant probability of detaching and by an exponential distribution of cell-to-substrate adhesive forces; (3) Past the second threshold, cells that are still adherent exhibit elongated morphologies, the degree of elongation increasing linearly with the force. The fact that cells lose their vinculin-rich focal contacts past the first threshold and that cells cultured on gelatin-coated plastic show an increased resistance to detachment suggests a rupture of cell-to-substrate adhe- sions upon centrifugation. Immunofluorescent labeling of cells for actin and tu- bulin shows a reorganization of the cytoskeleton upon centrifugation, and treat- ment of cells with the drugs cytochalasin D and nocodazole demonstrates that cytoskeletal elements are actively involved in the structural deformation of cells past the second acceleration threshold, microtubules and microfilaments playing antagonistic roles. 0 1996 Wiley-Liss, Inc.
Key words: centrifugation, epithelial, cell deformation, cytoskeletal organization, tangential accelera- tion, adhesive force
INTRODUCTION
Cellular morphogenesis is essential to embryonic development and pathological situations such as cancer metastasis and would healing. Cell shape is determined by several types of forces acting on the cell structure [Penman, 19951. These forces include osmotic [Al- brecht-Buehler and Bushnell, 19821 and hydrostatic [Be- reiter-Hahn, 19871 pressures acting on the membranes, prestress integrated by the cytoskeleton [Sims et al., 1992; Ingber, 19931, adhesive forces conveyed between the cell structure and the external environment through cell-cell and cell-matrix transmembrane mechano-chem- ical connections such as focal contacts [Luna and Hitt, 1992; Wang et al., 19931, and other external physical forces (e.g. gravity, shearing stresses in the circulation). Changes in one or more of these factors alter the overall
0 1996 Wiley-Liss, Inc.
force balance and may affect cellular morphology and structural organization.
Experiments using environments of reduced or in- creased gravity have shown that gravitational forces could influence the oriented growth of plants [Sack, 19911, cytoplasmic streaming in plant cells [Wayne et al., 19921, animal embryogenesis [Suda et al., 19941, adult physiology [Kirsch et al., 19841, as well as mam- malian cell proliferation [Cogoli et a]., 19841. In such biological systems, gravitational fields alter the force distribution by acting differentially on components of different mass densities. Specific connections between
Received November 23, 1995; accepted November 30, 1995
Address reprint requests to Olivier Thoumine, Ph.D., Section de Re- cherche, Institut Curie, 75231 Paris Cedex 05, France.

Tangential Centrifugation of Cells 277
trated cell suspensions. Cell volume (V,) was approxi- mated by V, = 4.rrr3/3 where r is the radius of trypsinized cells: r = 6.7 + 0.6 pm (n = 40 cells). The mass density of cells was found to be p, = 1.29 r 0.06 g/cm' (4 experiments, each using about 2.5 lo7 cells). This value was used to calculate the effective force (f) from the acceleration (a), using: f = (p, - pl). V,.a, where p1 is the mass density of the culture medium (p, = 1.01 g/cm3).
Perturbation of Cell Adhesion or Cytoskeleton To test whether the strength of cell-to-substrate ad-
hesion could affect the response of cells to the centrifugal field, cells were cultured onto plastic slides which had previously been coated for 2 h at 37°C with 0.2% gelatin in phosphate-buffered saline (PBS).
To assess whether the integrity of the cytoskeleton contributed to the cellular response to the centrifugal force, cells were incubated with MEM containing 1 pg/ml cytochalasin D or 5 pg/ml nocodazole for 10 min at 37"C, before centrifugation.
Cellular Uptake of Glass Microspheres Glass microspheres (diameter 1.5 pm, specific
gravity 2.45 g/ml) were coated by suspension in gelatin (a 0.2% solution in PBS, at 37"C), followed by two cycles of sedimentation and washing in PBS at 4°C. Cells were incubated for 2 h at 37°C with these glass beads suspended in culture medium at a concentration of 1 mg/ml (about 1.9 lo* beaddml). The beads bound to the cells and were phagocytosed. Microspheres present in cells were counted using a 100 X / I .3 n.a. phase con- trast objective, after placing the slide carrying the cells upside down onto a coverslip: each cell had taken up an average of 40 +- 25 glass beads (74 cells examined).
Centrifugation Cells were inserted into centrifuge tubes filled with
MEM containing 20 mM Hepes, without NaHCO,, and supplemented with 10% fetal calf serum. Tubes were placed in a swinging-bucket rotor and centrifuged at ac- celerations ranging from 9,000 to 70,OOOg for time pe- riods of 5 , 15, or 60 min in a Beckman (Palo Alto, CA) ultracentrifuge at 37°C. After centrifugation, cells were observed and stained for vinculin, tubulin, or actin, or immersed in supplemented MEM containing NaHCO, and placed back into the incubator to allow for recovery.
Cell Morphometry and Statistical Analysis Three to five 15 mm-long areas were selected along
the slide. In each area, the acceleration was within an interval whose limits are given by a mean value +- 7-12%, depending on the rotational speed. Cells were filmed using a Zeiss (Germany) microscope equipped
such components and the cytoskeleton [Buchen et al., 1993; Braun and Sievers, 19931 or the extracellular ma- trix [Wayne et al., 19921 may be important in gravity sensing by transmitting the forces generated by gravita- tional fields, although the precise mechanisms of force transduction remain unclear.
Past studies have shown that cell adhesion [Lotz et al., 19891 and nuclear position [Krokhina and Nadezh- dina, 19911 were sensitive to centrifugation, but no di- rect effect of acceleration on cell morphology has been reported yet. Here, in order to assess whether gravita- tional fields could directly influence mammalian cell morphology and dictate pattern formation in their cy- toskeleton, epithelial cells were exposed to high centrif- ugal accelerations tangential to their substrate, corre- sponding to effective forces in the range of previously reported cell-associated stresses. The experiments show the existence of critical forces for both cell detachment and cell elongation, which quantitatively characterize the adhesive and morphologic behavior of the cells, respec- tively.
MATERIALS AND METHODS Materials
Cell culture reagents were obtained from Biologi- cal Industries (Israel). Rhodamine-phalloidin was from Molecular Probes (Eugene, OR). Cytochalasin D, no- codazole, monoclonal antibodies to vinculin and to p-tu- bulin, FITC-conjugated anti-mouse IgG, and all other chemical reagents were purchased from Sigma (St. Louis, MO). Uniform glass microspheres were obtained from Duke Scientific Corp. (Palo Alto, CA).
Cell Culture Immortal human mammary epithelial cells
HBLlOO [Caron de Fromentel et al., 1985; Lebeau et al., 19911, passage 59, were generously provided by the lab- oratory of Gerard Goubin (Section de Recherche, Institut Curie). Cells were grown on tissue culture plastic dishes and fed with minimum essential medium (MEM) supple- mented with 50 IU/ml penicillin, 0.05 mg/ml streptomy- cin, 2 mM L-glutamine, and 10% fetal calf serum (FCS). Cells were grown in a humidified incubator, provided with an atmosphere of 95% air-5% CO, at 37°C. One day before centrifugation, cells were plated at a 1:6 surface ratio onto rectangular plastic slides (2 X 13 X 70 mm) with field-off corners, cut from a tissue culture flask with a hot scalpel. Cells were used at a subconfluent level since attempts to centrifuge confluent monolayers led to cell detachment by sheet.
Measurement of Cell Mass Density Cell weight was determined by weighing a 10-ml
calibrated Gay-Lussac pycnometer filled with concen-

278 Thoumine et al.
with phase contrast optics, a CCD camera, and a video recording system. Cell counts were made directly from the video monitor. The longest segment which could be traced between two points situated on the edges of a given cell was taken as the cell length. Widths of cells were taken halfway on this segment. Measurements were made directly on the video monitor for 70-320 cells in each of the areas mentioned.
Errors associated with cell length and width mea- surements were around 0.5 pm, and the error on cell number was about 3%. Due to heterogeneity in the cel- lular response to the centrifugal field, standard errors associated with morphometric measurements were large (i.e., typically 50 to 100%). Errors associated with cell density determinations were also high since subconfluent cells tended to form clusters. Hence, limits of the 95% confidence intervals calculated from a t-distribution were added to the mean values in the graphs. Tukey's paired comparison analysis [Pearson and Hartley, 19701 was performed to ascertain that when these intervals did not intersect, two means were indeed significantly different from each other.
Fluorescent Stainings for Vinculin, Tubulin, and Actin
Vinculin labeling was performed essentially as de- scribed by Turner et al. [1989]. For staining of microtu- bules, cultures were extracted for 10 rnin in 5% acetic acid:95% methanol (v/v) at -2O"C, rehydrated in PBS, incubated for 2 h with monoclonal antibodies to (3-tubu- lin, rinsed in PBS, incubated for 1 h with FITC-conju- gated goat anti-mouse IgG, and finally rinsed in PBS. For filamentous (F-) actin labeling, cells were fixed for 15 rnin with 3.7% formaldehdye in PBS and permeabi- lized for 10 min with 0.2% Triton X-100 in PBS. Cells were incubated for 2 h with rhodamine-phalloidin in PBS, then rinsed for 1 h. Incubations and rinses were carried out at room temperature. Glass coverslips were mounted onto the slides previously covered with a 1:l glycerol-PBS solution. Fluorescent labelings were visu- alized using a 100 X /1.4 n.a. Plan-Apochromat objective with a Zeiss inverted microscope equipped with epiflu- orescence optics.
RESULTS Adhesive and Morphologic Behavior of Epithelial Cells in a Tangential Centrifugal Field
Under normal gravity condition, HBLlOO cells tend to grow as clusters and exhibit rather polygonal morphologies (Fig. 1A). Cells exposed to accelerations lower or equal to 37,OOOg (& 3,500g) for 15 min showed only a slight degree of detachment and retained a poly- gonal-like morphology (Fig. 1B). In cultures exposed for
15 min to accelerations of 51,OOOg (k 3,500g) and 64,000 (+ 3,50Og), cell density was strongly reduced, while the majority of still adherent cells were elongated in the direction of the centrifugal field (Fig. 1C and D). At high accelerations, cell bodies often pointed towards the bottom of the slide, and cellular material formed long tails oriented in the direction of the force (Fig. ID).
Cells which were exposed to these large accelera- tions were still alive after centrifugation, for they re- mained adherent to the slides for several days and con- tinued to proliferate. Cells which had adopted elongated morphologies recovered a normal polygonal-like mor- phology in a matter of several hours (not shown): after overnight incubation at lg, most of the elongated cells had relaxed.
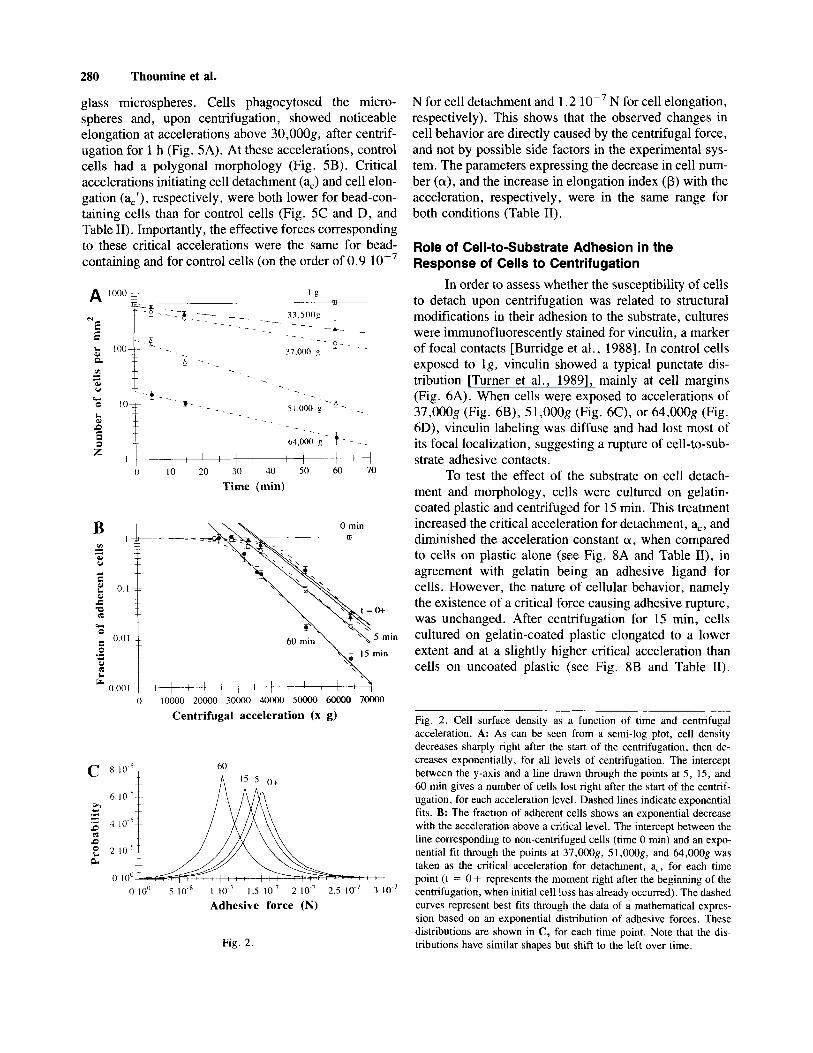
Decrease of Cell Density Over Time Cell surface density decreased sharply right after
the start of the centrifugation (Fig. 2A). After this initial loss, it decreased in an exponential fashion with time. Assuming the probability of a cell to detach at times t > 0 as being constant, we can write:
-FN/6T = N(t) or N(t) = N,+(a) exp[- V T ] ,
where FN is the fraction of cells detaching during a time interval Ft, N(t) is the number of adherent cells at time t, No + (a) is the number of cells after the initial loss at the considered acceleration, and T is the characteristic time leading to a cell surface density reduced by a factor of e. Exponential fits through the data gave values for initial cell loss and characteristic time T. The fraction of adher- ent cells at t = 0 + decreased with acceleration (Table I, Fig. 2B), while T was relatively independent of acceler- ation (Table I, Fig. 2A), confirming that the probability of detaching is constant.
Decrease of Cell Density With Respect to Centrifugal Acceleration
Expressing data as a function of acceleration on a semi-log plot (Fig. 2B) reveals that cell density remains fairly constant at accelerations below a threshold, a,, and exhibits an exponential decrease with acceleration above this critical value. Thus we can write:
@ (a) = exp [-(a - a,)/a] for a 2 a,,
where is the fraction of adherent cells, a is the cen- trifugal acceleration, and a is a characteristic accelera- tion, accelerations of a, + a reducing cell density by a factor of e.
If one associates to each cell an adhesive force (f) which can evolve with time and vary from cell to cell, and further assumes that the population of these adhesive

Tangential Centrifugation of Cells 279
Fig. 1. Morphology of HBLlOO cells exposed to tangential centrifu- gal forces. Subconfluent cultures of HBLlOO under normal gravity condition (A) or exposed to accelerations of 37,OOOg (B), 51,OOOg (C), or 64,OOOg (D) for 15 min, were photographed using phase- contrast microscopy. Cells exposed to 37,OOOg do not exhibit major
structural alteration, as compared with non-centrifuged cells: they re- tain a rather polygonal morphology. Cells exposed to accelerations of 51,OOOg and 64,OOOg show a dramatic elongation in the direction of the field (from top to bottom). Bar = 50 pm.
forces follows a given statistical distribution, the fraction of cells able to resist an applied force f,, P (f>f,), is given by the area under the distribution curve [Truskey and Pirone, 19901. For an exponential distribution cen- tered on fA, with a characteristic slope of y: P(f) = O.Syexp[(-lf - fAI)/y]. Based on such a model, one can derive a second expression for the fraction of adherent cells upon centrifugation, where a and aA are the accel- erations corresponding to the centrifugal force f, and the adhesive force fA, respectively:
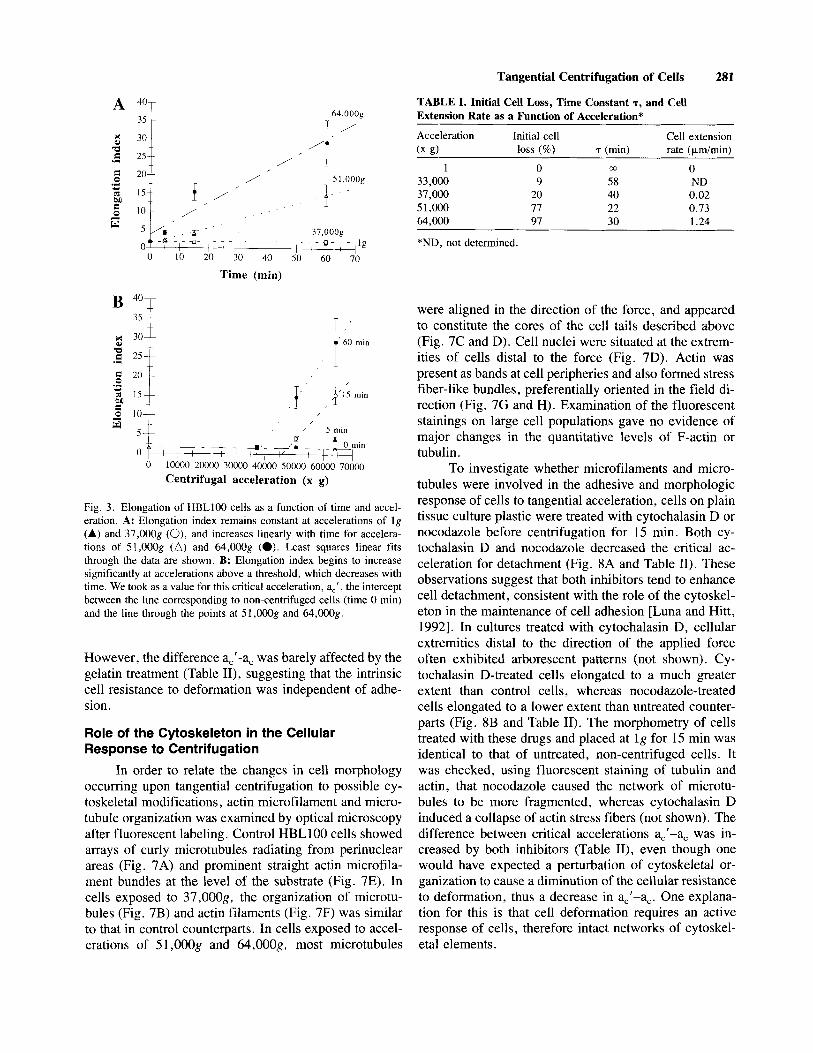
stant around 1.5 at accelerations below 37,00Og, and increased linearly with time for accelerations of 5 1,OOOg and 64,OOOg (Fig. 3A). Cell extension rates, given as the slopes of linear fits through the length data, increased with the acceleration (Table I).
Elongation index (E) was found to increase in a relatively linear fashion with the acceleration in excess of a second threshold value, a=’:
E(a) = E(0) + (a-a,’)+ for a > a,’,
@ (a) = 0.5 exp[(a - aA)/y] for a < aA where a is the acceleration and p is the inverse of the slope of a linear fit (Fig. 3B). The critical acceleration and CD (a) = 1 - 0.5 exp [(a - a*)/?] for a > aA.
We determined values of fA and y for which we obtained the best visual fit with the experimental data (Fig. 2B). The agreement is very good, suggesting that adhesive forces in the HBL 100 cell population indeed follow exponential distributions, which are presented as a function of time in Figure 2C. Based on the interpre- tation, the threshold acceleration a, roughly corresponds to fA - y. The mean adhesive force of the population decreases linearly with time, as fA (t) = fA(0) - Et, with fA(0) = 43,OOOg and E = 222g min-’ (r = 0.98). Ranging from 6,500g to 7,5OOg, the parameter y is close
a,’ decreased with time whereas p remained constant within the observed range (Table 11).
Thus cell behavior in a tangential centrifugal field can essentially be described by two parameters: a critical acceleration for detachment (a,) which roughly corre- sponds to the mean cell-to-substrate adhesive force, and another critical acceleration (a,’), the difference between the two representing the additional resistance to defor- mation provided by the cell structure itself, which is independent of time (Table 11). Three regimes can be distinguished depending on time and acceleration, which are described in Figure 4.
to (Y and almost time-independent.
Elongation of Cells at Accelerations Exceeding Critical Levels
An elongation index was computed as the ratio of cell length divided by width. This index remained con-
Influence of Ingested Heavy Particles on the Detachment and Elongation of Cells
To assess whether the presence of internalized dense bodies could affect the cellular response to the centrifugal field, HBLlOO cells were incubated with
280 Thournine et al.
glass microspheres. Cells phagocytosed the micro- spheres and, upon centrifugation, showed noticeable elongation at accelerations above 30,00Og, after centrif- ugation for 1 h (Fig. 5A). At these accelerations, control cells had a polygonal morphology (Fig. 5B). Critical accelerations initiating cell detachment (a,) and cell elon- gation (a,'), respectively, were both lower for bead-con- taining cells than for control cells (Fig. 5C and D, and Table 11). Importantly, the effective forces corresponding to these critical accelerations were the same for bead- containing and for control cells (on the order of 0.9 lo-'
CI
0.01 .; E
L4 0.001 0 10000 20000 30000 4OOO0 5 0 0 Mwx)o 7oooO
Centrifugal acceleration (x g)
Adhesive force (N)
N for cell detachment and 1.2 N for cell elongation, respectively). This shows that the observed changes in cell behavior are directly caused by the centrifugal force, and not by possible side factors in the experimental sys- tem. The parameters expressing the decrease in cell num- ber (a), and the increase in elongation index (p) with the acceleration, respectively, were in the same range for both conditions (Table 11).
Role of Cell-to-Substrate Adhesion in the Response of Cells to Centrifugation
In order to assess whether the susceptibility of cells to detach upon centrifugation was related to structural modifications in their adhesion to the substrate, cultures were immunofluorescently stained for vinculin, a marker of focal contacts [Burridge et al., 19881. In control cells exposed to lg, vinculin showed a typical punctate dis- tribution [Turner et al., 19891, mainly at cell margins (Fig. 6A). When cells were exposed to accelerations of 37,OOOg (Fig. 6B), 51,OOOg (Fig. 6C), or 64,OOOg (Fig. 6D), vinculin labeling was diffuse and had lost most of its focal localization, suggesting a rupture of cell-to-sub- strate adhesive contacts.
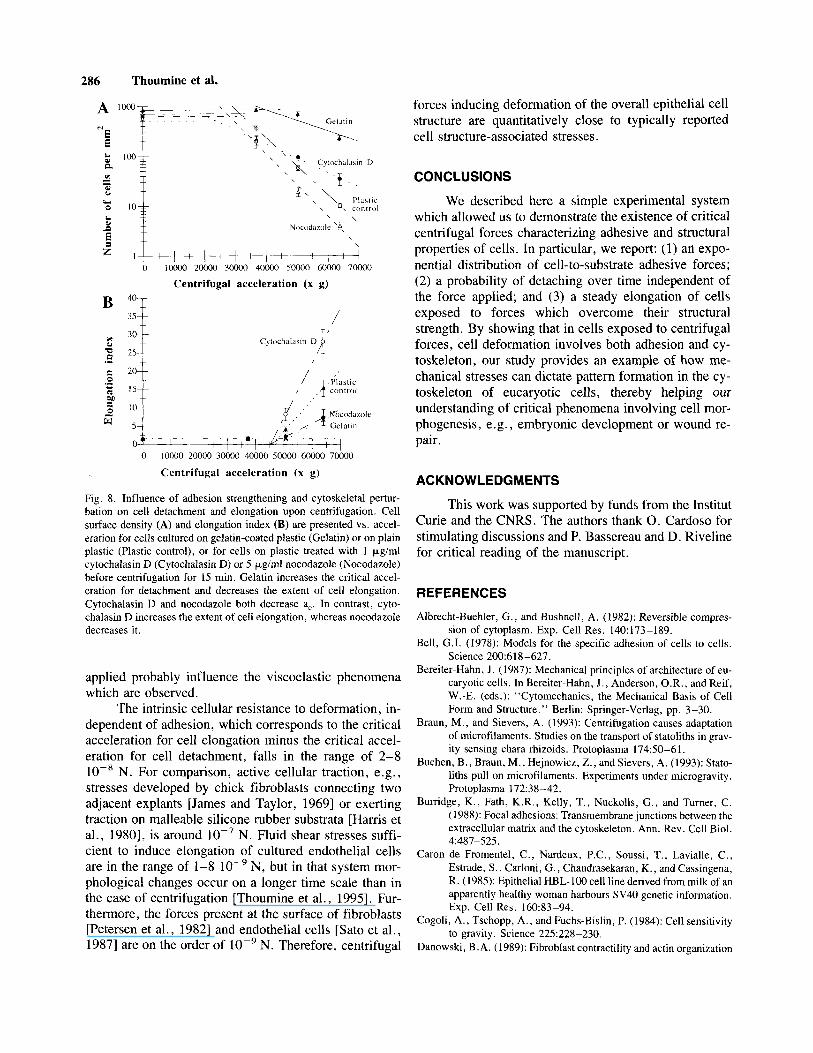
To test the effect of the substrate on cell detach- ment and morphology, cells were cultured on gelatin- coated plastic and centrifuged for 15 min. This treatment increased the critical acceleration for detachment, a,, and diminished the acceleration constant a, when compared to cells on plastic alone (see Fig. 8A and Table II), in agreement with gelatin being an adhesive ligand for cells. However, the nature of cellular behavior, namely the existence of a critical force causing adhesive rupture, was unchanged. After centrifugation for 15 min, cells cultured on gelatin-coated plastic elongated to a lower extent and at a slightly higher critical acceleration than cells on uncoated plastic (see Fig. 8B and Table 11).
Fig. 2. Cell surface density as a function of time and centrifugal acceleration. A: As can be seen from a semi-log plot, cell density decreases sharply right after the start of the centrifugation, then de- creases exponentially, for all levels of centrifugation. The intercept between the y-axis and a line drawn through the points at 5 , 15, and 60 min gives a number of cells lost right after the start of the centrif- ugation, for each acceleration level. Dashed lines indicate exponential fits. B: The fraction of adherent cells shows an exponential decrease with the acceleration above a critical level. The intercept between the line corresponding to non-centrifuged cells (time 0 min) and an expo- nential fit through the points at 37,OOOg, 51,00Og, and 64,OOOg was taken as the critical acceleration for detachment, a,, for each time point (t = 0 + represents the moment right after the beginning of the centrifugation, when initial cell loss has already occurred). The dashed curves represent best fits through the data of a mathematical expres- sion based on an exponential distribution of adhesive forces. These distributions are shown in C, for each time point. Note that the dis- tributions have similar shapes but shift to the left over time. Fig. 2
Tangential Centrifugation of Cells 281
TABLE I. Initial Cell Loss, Time Constant T, and Cell Extension Rate as a Function of Acceleration*
Acceleration Initial cell Cell extension (x g) loss (%) r (min) rate (Fdmin)
33,000 9 58 ND 37,000 20 40 0.02 5 1,000 77 22 0.73 64.000 97 30 1 24
1 0 m 0
A 401 1 35
64,OOOg T /
C 0
m M C 0
w
.I *
-
/I, ’ / ,
1 51’000g
/
i / ’ I 0 / ’
Time (min)
I,,’ 60 rnin
0 10000 20000 30000 40000 50000 60000 70000 Centrifugal acceleration (x g)
Fig. 3. Elongation of HBLl00 cells as a function of time and accel- eration. A: Elongation index remains constant at accelerations of Ig (A) and 37,000g (O), and increases linearly with time for accelera- tions of 51,000g (A) and 64,000g (0). Least squares linear fits through the data are shown. B: Elongation index begins to increase significantly at accelerations above a threshold, which decreases with time. We took as a value for this critical acceleration, ac’, the intercept between the line corresponding to non-centrifuged cells (time 0 min) and the line through the points at 51,OOOg and 64,000g.
However, the difference a,’-a, was barely affected by the gelatin treatment (Table II), suggesting that the intrinsic cell resistance to deformation was independent of adhe- sion.
Role of the Cytoskeleton in the Cellular Response to Centrifugation
In order to relate the changes in cell morphology occumng upon tangential centrifugation to possible cy- toskeletal modifications , actin microfilament and micro- tubule organization was examined by optical microscopy after fluorescent labeling. Control HBLlOO cells showed arrays of curly microtubules radiating from perinuclear areas (Fig. 7A) and prominent straight actin microfila- ment bundles at the level of the substrate (Fig. 7E). In cells exposed to 37 ,OOOg, the organization of microtu- bules (Fig. 7B) and actin filaments (Fig. 7F) was similar to that in control counterparts. In cells exposed to accel- erations of 5 1,OOOg and 64,00Og, most microtubules
*ND, not determined.
were aligned in the direction of the force, and appeared to constitute the cores of the cell tails described above (Fig. 7C and D). Cell nuclei were situated at the extrem- ities of cells distal to the force (Fig. 7D). Actin was present as bands at cell peripheries and also formed stress fiber-like bundles, preferentially oriented in the field di- rection (Fig. 7G and H). Examination of the fluorescent stainings on large cell populations gave no evidence of major changes in the quantitative levels of F-actin or tubulin .
To investigate whether microfilaments and micro- tubules were involved in the adhesive and morphologic response of cells to tangential acceleration, cells on plain tissue culture plastic were treated with cytochalasin D or nocodazole before centrifugation for 15 min. Both cy- tochalasin D and nocodazole decreased the critical ac- celeration for detachment (Fig. 8A and Table 11). These observations suggest that both inhibitors tend to enhance cell detachment, consistent with the role of the cytoskel- eton in the maintenance of cell adhesion [Luna and Hitt, 19921. In cultures treated with cytochalasin D, cellular extremities distal to the direction of the applied force often exhibited arborescent patterns (not shown). Cy- tochalasin D-treated cells elongated to a much greater extent than control cells, whereas nocodazole-treated cells elongated to a lower extent than untreated counter- parts (Fig. SB and Table 11). The morphometry of cells treated with these drugs and placed at l g for 15 min was identical to that of untreated, non-centrifuged cells. It was checked, using fluorescent staining of tubulin and actin, that nocodazole caused the network of microtu- bules to be more fragmented, whereas cytochalasin D induced a collapse of actin stress fibers (not shown). The difference between critical accelerations a,’-a, was in- creased by both inhibitors (Table II), even though one would have expected a perturbation of cytoskeletal or- ganization to cause a diminution of the cellular resistance to deformation, thus a decrease in a,’-a,. One explana- tion for this is that cell deformation requires an active response of cells, therefore intact networks of cytoskel- eta1 elements.

282 Thoumine et al.
TABLE 11. Critical Accelerations for Cell Detachment (a,) and Cell Elongation (a:), Acceleration Dependence of the Decrease in Cell Density (a), and Acceleration Dependence of the Increase in Cell Elongation (p), as a Function of Time and Cell Treatment*
Time (min)
5 15 60 15 15 15 15 60 60
Treatment a,
None 36 None 31 None 26 Gelatin 40 Plastic control 33 Cytochalasin D 24 Nocodazole 29 Glass beads 20 Control 27
a,' >51"
47 38 50 47 46 46 21 34
a,'-a,
> 15" 16 12 10 14 22 17 7 I
(Y R 7.9 NDa 7.9 1.2 6.5 0.9
20 2.6 7.5 1.2
16 0.7 7.0 2.6 5.7 1.0 5.0 1.3
*Values are to be multiplied by lo3 g. "A regression between accelerations of 51,OOOg and 64,OOOg was not realistic since cells were not elongated after only 5 min of centrifu- gation.
t . . . . . . . . . . . . . . . . . . . \ . . . .
0 , ' ~ ~ I i I ! - I ~ I ~ ~: I 0 10000 20000 30000 40000 50000 60000 70000
Centrifugal acceleration (x g)
Fig. 4. Summary diagram of cell behavior in a tangential centrifugal field. The two curves represent the critical accelerations for cell de- tachment (a,) and for cell elongation (ac'), respectively, plotted vs. time. They separate three domains: (1) in the left region, acceleration is too low to produce either cell detachment or elongation; (2) in the middle region, cells detach in an exponential fashion with time at accelerations above the critical level a,, but accelerations are still insufficient to cause cell deformation; and (3) on the right, cells con- tinue to detach, while still adherent cells adopt elongated morpholo- gies, the degree of elongation increasing with the acceleration above the critical level ac'.
DISCUSSION
Adherent epithelial cells exposed to high centrifu- gal accelerations directed in the plane of their substrate exhibited two types of responses: either they detached or they exhibited a dramatic shape reorganization, namely an elongation in the direction of the field. Detachment was insignificant until the force exceeded a critical level in the range of 0.8-1.4 N, depending on time of exposure and on cell treatment. Cells that were still ad- herent showed significant elongation when the force was greater than a second threshold of 1.2-1.7 lop7 N.
Thus, the intrinsic resistance to deformation of the cell structure corresponded to forces around 2-8 lo-' N.
Gravitational Forces Are Conveyed From Dense Cellular Elements to Adhesion Sites
Due to heterogeneity in the density of the various cellular components, force distribution within a cell ex- posed to these centrifugal accelerations was not uniform. In particular, the cell nucleus having the highest density among cellular components, it seems likely that cell nu- clei were effectively pulled in the direction of the cen- trifugal acceleration. This explains why in elongated cells, nuclei were almost always found at the cell ex- tremity pointing in the direction of the force applied. Such a view is consistent with the observation that in embryonic pig kidney epithelial cells, nuclei are dis- placed when cells are exposed to tangential centrifugal fields of 20,OOOg [Krokhina and Nadezhdina, 1991 1. The value of the centrifugal acceleration used in the study mentioned above was, however, too low to induce any significant morphological change in cells, other than a movement of their nuclei.
When cells are exposed to tangential acceleration, forces are being transmitted throughout the cell structure from these heaviest elements of the cell on which the force is the strongest (e.g., the nucleus) to adhesion sites, where the centrifugal effects are resisted by a tight connection to the substrate [Luna and Hitt, 19921, with- out which cells would detach. Such mechanical loads may be conveyed from the nuclear matrix [Penman, 199.51 to focal contacts [Burridge et al., 19881 directly through intermediate filaments [Quantara and Jones, 1991 J , but also indirectly through the integrated micro- tubular and microfilament cytoskeleton [Pollard et a]., 1984; Ingber, 19931, with which the vimentin network can interact [Hollenbeck et al., 19891. Indeed, adhesion forces appear to be involved in resisting the centrifugal force applied for at least three reasons: (1) cells plated on gelatin-coated plastic, thereby being more adherent, show less detachment than cells on uncoated plastic; (2) human dermal fibroblasts, which have a larger contact area with their substrate than epithelial cells, start de- taching at higher centrifugal accelerations (data not shown); and (3) accelerations sufficient to initiate cell detachment induce a disruption of vinculin-rich focal contacts in HBLlOO cells. It should be pointed out that intercellular adhesions also contribute in resisting the force applied since cells grouped in clusters were usually less elongated than isolated cells.
Cell Elongation Upon Centrifugation Involves a Remodeling of the Cytoskeleton
Cell elongation not involves a decrease in adhesive strength, reflected by a loss of focal contacts, but also a

Tangential Centrifugation of Cells 283
L Q,
n
$ 2
loo+
c
0
Glass beads \ i
't Pn, '9 ' \ \
' \
. \
I I I I I 1 I , \ I \ ,
10000 20000 30000 40000 SO000 60000
Centrifugal acceleration (x g)
Fig. 5. Response of HBLlOO cells to tangential centrifugation after uptake of glass microspheres. Cells were incubated with gelatin- coated glass microspheres, then centrifuged for 1 h at accelerations ranging from 24,OOOg to 37,OOOg. A: Bead-containing cells exposed to accelerations of 34,OOOg and photographed using phase contrast microscopy. Upon centrifugation, beads often form aggregates at the extremities of cells (arrows) situated distally with respect to the force
structural reorganization of the cytoskeleton. Indeed in elongated cells, both microtubules and actin stress fibers exhibited a preferential orientation in the direction of the centrifugal field. The fact that blocking microtubule as- sembly with nocodazole, which results in an overall de- polymerization of existing microtubules in the time range considered, inhibited cell elongation in response to cen- trifugal forces in excess of a threshold indicates that in- tact microtubules are essential to the elongation process, e.g., by stabilizing the cell structure in an elongated shape. The formation of patterns of axially aligned mi- crotubules may involve either a spatial reorganization of existing filaments or polymerization of new filaments, or both. We know that externally applied tension is able to stimulate microtubule formation and elongation of cellu- lar processes, as is the case of neurons [Zheng et al., 19931. Interestingly, the rate of elongation of HBLlOO cells exposed to centrifugal forces in excess of the threshold, which is on the order of 1 p d h , roughly
E 0
m DO E 0
.I *
5
t / T , '
0 10000 20000 30000 40000 SO000 60000
Centrifugal acceleration (x g)
applied (from left to right). B: Control cells incubated with bead-free medium and exposed to 34,OOOg. C: Cell density shows an exponen- tial decrease with the acceleration above a critical level both for con- trol cells (O), and for bead-containing cells (0). D: Elongation index increases linearly with acceleration above a threshold in both condi- tions. Note that critical accelerations for both cell detachment and cell elongation are decreased by the uptake of beads. Bar = 50 km.
corresponds to the rate of neurite extension in response to towing tensions in the range of 0.1-0.5 lo-* N [Zheng et al., 19911, and also to the growth velocity of micro- tubules spontaneously assembling in vitro [Fygenson et al., 19941. These comparisons suggest that de novo mi- crotubule polymerization may be involved in cell elon- gation upon centrifugation.
On the other hand, cells treated with cytochalasin D were more elongated than untreated controls, suggest- ing that actin filaments tend to resist the action of the centrifugal force, as they resist the effects that gravita- tional fields exert on statoliths in chara rhizoids [Buchen et al., 1993; Braun and Sievers, 19931. These antago- nistic effects of nocodazole and cytochalasin D on the process of cell elongation are consistent with the idea that microtubules and microfilaments counterbalance each other's mechanical actions in the cytoskeleton. Mechanical competition between the two interacting cy- toskeletal networks [Pollard et al., 19841 has been dem-

284 Thoumine et al.
Fig. 6. Rupture of adhesion sites in HBLlOO cells exposed to tangen- tial centrifugal forces. Cells exposed to l g (A), 37,OOOg (B) 51,0001: (C), or 64,OOOg (D) for 15 min were immunofluorescently labeled for vinculin. The staining reveals the uniform presence of vinculin in cell bodies, except for nuclear areas. In control conditions (lg), vinculin is
also present at focal contacts where it forms bright streaks, mainly localized at cell margins (A). Exposure of cells to centrifugal accel- erations sufficient to initiate cell detachment (B) or induce cell elon- gation (C,D), causes a loss in the focal distribution of vinculin. Force direction is from top to bottom. Bar = 20 km.
onstrated in several cell types [Vasiliev, 1987; Danow- ski, 19891 including neurons [Dennerll et al., 19881 and retinoreceptors [Madreperla and Adler, 19891, and has led to the hypothesis that cells may behave as tensionally integrated structures [Ingber, 19931, in which the micro- filament system would generate contractile forces re- sisted by the microtubular network thereby being under compression [Danowski, 19891.
Critical Forces Characterize the Adhesive and Morphologic Behavior of Cells
We demonstrated a rapid cell loss at centrifugal accelerations above a threshold. This existence of a crit- ical force for cell detachment, indicating a rapid and simultaneous rupture of specific proteic bonds between- cells and their extracellular matrix, is in agreement with

Tangential Centrifugation of Cells 285
Fig. 7 . Cytoskeletal reorganization in HBLlOO cells exposed to tan- gential centrifugal forces. Cells under normal gravity condition (A, E) exposed to 37,OOOg (B, F), 51,OOOg (C, G), or 64,OOOg (D, H) for 15 min, were labeled for P-tubulin (A,B,C,D) or F-Actin (E,F,G,H), and examined by fluorescence microscopy. Cells exposed to accelerations of 37,OOOg do not exhibit major cytoskeletal alteration, as compared
with non-centrifuged cells. Microtubules form wavy filaments radiat- ing from the nuclei, whereas F-actin is organized as peripheral bands and randomly oriented straight bundles. In cells exposed to accelera- tions of 51,OOOg and 64,00Og, both microtubules and actin cables are oriented in the direction of the field (from top to bottom). Bar = 20 pm.
the theoretical predictions made by Bell [ 19781. The ex- ponential decrease of cell number with acceleration above this threshold agrees well with a sharp exponential distribution of cell-to-substrate adhesive forces, whose shape is unaffected by time. In contrast, the mean adhe- sive force decreases with time, indicating that prolonged exposure of cells to centrifugation fragilizes cell-to-sub- strate adhesions. Studies using fibroblasts exposed to shear stress [Truskey and Pirone, 19901 have reported smoother distributions of adhesive stresses (e.g., log- normal), showing that the nature or magnitude of the applied force is important in the adhesive response of cells. When examined over time, cell surface density shows an initial drop, whose amplitude increases with the force applied, then decreases exponentially with time. This is explained by a probability of detaching
independent of acceleration, suggesting that for cells which remain adherent the force per bond stays constant.
Conversion of the critical accelerations (expressed in g) to units of force indicate that the critical centrifugal forces which initiate cell detachment are in the range of 0.8-1.4 lo-' N. Given a projected cell area of about 600 km2, the mean adhesive stress of the HBLlOO cell pop- ulation is around 200 N/m2. This value is one to two orders of magnitude larger than the stresses reported by other investigators for fibroblasts exposed to shear stress (3-46 N/m2) [Truskey and Pirone, 19901 or glial cells centrifuged in a direction perpendicular to the substrate (0.03-1 lo-' N) [Lotz et al., 19891. This discrepancy reflects differences in cells, substrate coatings, times af- ter cell plating, and techniques of measurement. Also, the magnitude of the force and the rate at which it is
286 Thoumine et al.
forces inducing deformation of the overall epithelial cell structure are quantitatively close to typically reported cell structure-associated stresses.
5 e
0 loo00 Z o o 0 0 3oooO 40600 50000 60bO0 7oooO
Centrifugal acceleration (x g)
/ E 30$ 2s
.- A’ Cytrichalasin D
c 20+ .- 0 Y OJ M
0 - w 0 10000 20000 30000 40000 50000 60000 70000
Centrifugal acceleration (x g)
Fig. 8. Influence of adhesion strengthening and cytoskeletal pertur- bation on cell detachment and elongation upon centrifugation. Cell surface density (A) and elongation index (B) are presented vs. accel- eration for cells cultured on gelatin-coated plastic (Gelatin) or on plain plastic (Plastic control), or for cells on plastic treated with 1 pg/ml cytochalasin D (Cytochalasin D) or 5 p,g/ml nocodazole (Nocodazole) before centrifugation for 15 min. Gelatin increases the critical accel- eration for detachment and decreases the extent of cell elongation. Cytochalasin D and nocodazole both decrease a,. In contrast, cyto- chalasin D increases the extent of cell elongation, whereas nocodazole decreases it.
applied probably influence the viscoelastic phenomena which are observed.
The intrinsic cellular resistance to deformation, in- dependent of adhesion, which corresponds to the critical acceleration for cell elongation minus the critical accel- eration for cell detachment, falls in the range of 2-8 lop8 N. For comparison, active cellular traction, e.g., stresses developed by chick fibroblasts connecting two adjacent explants [James and Taylor, 19691 or exerting traction on malleable silicone rubber substrata [Harris et al., 19801, is around lop7 N. Fluid shear stresses suffi- cient to induce elongation of cultured endothelial cells are in the range of 1-8 N, but in that system mor- phological changes occur on a longer time scale than in the case of centrifugation [Thoumine et al., 19951. Fur- thermore, the forces present at the surface of fibroblasts [Petersen et al., 19821 and endothelial cells [Sato et al., 19871 are on the order of lop9 N. Therefore, centrifugal
CONCLUSIONS
We described here a simple experimental system which allowed us to demonstrate the existence of critical centrifugal forces characterizing adhesive and structural properties of cells. In particular, we report: (1) an expo- nential distribution of cell-to-substrate adhesive forces; (2) a probability of detaching over time independent of the force applied; and ( 3 ) a steady elongation of cells exposed to forces which overcome their structural strength. By showing that in cells exposed to centrifugal forces, cell deformation involves both adhesion and cy- toskeleton, our study provides an example of how me- chanical stresses can dictate pattern formation in the cy- toskeleton of eucaryotic cells, thereby helping our understanding of critical phenomena involving cell mor- phogenesis, e.g., embryonic development or wound re- pair.
ACKNOWLEDGMENTS
This work was supported by funds from the Institut Curie and the CNRS. The authors thank 0. Cardoso for stimulating discussions and P. Bassereau and D. Riveline for critical reading of the manuscript.
REFERENCES
Albrecht-Buehler, G., and Bushnell, A. (1982): Reversible compres- sion of cytoplasm. Exp. Cell Res. 140:173-189.
Bell, G.I. (1978): Models for the specific adhesion of cells to cells. Science 200:618-627.
Bereiter-Hahn, J . (1987): Mechanical principles of architecture of eu- caryotic cells. In Bereiter-Hahn, J . , Anderson, O.R., and Reif, W.-E. (eds.): “Cytomechanics, the Mechanical Basis of Cell Form and Structure.” Berlin: Springer-Verlag, pp. 3-30.
Braun, M., and Sievers, A. (1993): Centrifugation causes adaptation of microfilaments. Studies on the transport of statoliths in grav- ity sensing chara rhizoids. Protoplasma 174:50-61.
Buchen, B., Braun, M., Hejnowicz, Z., and Sievers, A. (1993): Stato- liths pull on microfilaments. Experiments under microgravity . Protoplasma 172:38-42.
Burridge, K., Fath, K.R., Kelly, T., Nuckolls, G . , and Turner, C. (1 988): Focal adhesions: Transmembrane junctions between the extracellular matrix and the cytoskeleton. Ann. Rev. Cell Biol.
Caron de Fromentel, C., Nardeux, P.C., Soussi, T., Lavialle, C., Estrade, S . , Carloni, G., Chandrasekaran, K., and Cassingena, R. (1985): Epithelial HBL-100 cell line derived from milk of an apparently healthy woman harbours SV40 genetic information. Exp. Cell Res. 160:83-94.
Cogoli, A., Tschopp, A. , and Fuchs-Bislin, P. (1984): Cell sensitivity to gravity. Science 225:228-230.
Danowski, B .A. (1989): Fibroblast contractility and actin organization
41487-525.

Tangential Centrifugation of Cells 287
Petersen, N.O., McConnaughey, W.B., and Elson, E.L. (1982): De- pendence of locally measured cellular deformability on position on the cell, temperature, and cytochalasin B. Proc. Natl. Acad. Sci. U.S.A. 79:5327-5331.
Pollard, T.D., Selden, S.C., and Maupin, P. (1984): Interaction of actin filaments with microtubules. J. Cell Biol. 99:33~-37s.
Quantara, V., and Jones, J.C.R. (1991): The internal affairs of an integrin. Trends Cell Biol. 1:2-4.
Sack, F. (1991): Plant gravity sensing. Int. Rev. Cytol. 127:193-252. Sato, M., Levesque, M.J., and Nerem, R.M. (1987): An application
of the micropipette technique to the measurement of the me- chanical properties of cultured bovine aortic endothelial cells. J . Biomech. Eng. 109:27-34.
Sirns, J.R., Karp, S., and Ingber, D.E. (1992): Altering the cellular mechanical force balance results in integrated changes in cell, cytoskeletal and nuclear shape. J. Cell Sci. 103: 1215-1222.
Suda, T., Abe, E., Shinki, T. , Katagari, T., Yamaguchi, A,, Yokose, S. , Yoshiki, S . , Horikawa, H., Cohen, G.W., Yasugi, S . , and Naito, M. (1994): The role of gravity in chick embryogenesis. FEBS Lett. 340:34-38.
Thoumine, O., Ziegler, T., Girard, P.R., and Nerem, R.M. (1995): Elongation of confluent endothelial cells in culture: The im- portance of fields of force in the associated alterations of their cytoskeletal structure. Exp. Cell Res. 219:427-441.
Truskey, G.A., and Pirone, J.S. (1990): Effect of fluid shear stress upon cell adhesion to fibronectin-treated surfaces. J. Biomed. Mat. Res. 24:1333-1353.
Turner, C.E., Pavalko, F.M., and Bumdge, K. (1989): The role of phosphorylation and limited proteolytic cleavage of talin and vinculin in the disruption of focal adhesion integrity. J. Biol. Chern. 264:11938-11944.
Vasiliev, J.M. (1987): Actin cortex and microtubular system in mor- phogenesis: Cooperation and competition. J . Cell Sci. (Suppl) 8:l-18.
Wang, N., Butler, J.P., and Ingber, D.E. (1993): Mechanotransduc- tion across the cell surface and through the cytoskeleton. Sci- ence 260: 1 124- 1 127.
Wayne, R., Staves, M.P., and Leopold, A.C. (1992): The contribu- tion of the extracellular matrix to gravisensing in characean cells. J. Cell Sci. 101:611-623.
Zheng, J., Lamoureux, P., Santiago, V., Dennerll, T. , Buxbaum, R.E., and Heidemann, S.R. (1991): Tensile regulation of ax- onal elongation and initiation. J. Neurosci. 11:1117-1125.
Zheng, J., Buxbaum, R.E., and Heidemann, S.R. (1993): Investiga- tion of microtubule assembly and organization accompanying tension-induced neurite initiation. J. Cell Sci. 104: 1239-1250.
are stimulated by microtubule inhibitors. J. Cell Sci. 93:255- 266.
Dennerll, T.J., Joshi, H.C., Steel, V.L., Buxbaum, R.E., and Heide- mann, S.R. (1988): Tension and compression in the cytoskel- eton of PC-12 neurites 11: Quantitative measurements. J. Cell Biol. 107:655-674.
Fygenson, D.K., Braun, E., and Libchaber, A. (1994): Phase diagram of microtubules. Phys. Rev. E. 50:1579-1588.
Hams, A.K., Wild, P., and Stopak, D. (1980): Silicone rubber sub- strata: A new wrinkle in the study of cell locomotion. Science
Hollenbeck, P. J., Bershadsky, A.D., Pletjushkina, O.Y., Tint, I .S. , and Vasiliev, J.M. (1989): Intermediate filament collapse is an ATP-dependent and actin-dependent process. J. Cell Sci. 92:
Ingber, D.E. (1993): Cellular tensegrity: Defining new rules of bio- logical design that govern the cytoskeleton. J. Cell Sci. 104:
James, D.W., and Taylor, J.F. (1969): The stress developed by sheets of chick fibroblasts in vitro. Exp. Cell Res. 54107-1 10.
Kirsch, K.A., Rocker, L. , Gauer, O.H., Krause, R., Leach, C., Wicke, H.J., and Landry, R. (1984): Venous pressure in man during weightlessness. Science 225:218-219.
Krokhina, T.V., and Nadezhdina, E.S. (1991): The role of cytoskel- eta1 elements in the process of the return of cellular nuclei displaced by centrifugation to the center of the cells. Tsi- tologiya (Russian) 33:28-35.
Lebeau, J., Le Chalony, C., Prosperi, M.-T., and Goubin, G . (1991): Constitutive overexpression of a 89 kDa heat shock protein gene in the HBLlOO human mammary cell line converted to a tumorigenic phenotype by the EJ/T24 Harvey-ras oncogene. Oncogene 6:1125-1132.
Lotz, M.M., Burdsal, C.A., Erickson, H.P., and McClay, D.R. (1989): Cell adhesion to fibronectin and tenascin: Quantitative measurements of initial binding and subsequent strengthening response. J. Cell Biol. 109:1795-1805.
Luna, E.J., and Hitt, A.L. (19920: Cytoskeleton-plasma membrane interactions. Science 258:955 -964.
Madreperla, S.A., and Adler, R. (1989): Opposing microtubule- and actin-dependent forces in the development and maintenance of structural polarity in retinal photoreceptors. Dev. Biol. 131:
Pearson, E.S., and Hartley, H.O. (1970): “Biometrika Tables for Statisticians. ” Cambridge: Cambridge University Press, pp.
Penman, S. (1995): Rethinking cell structure. Proc. Natl. Acad. Sci.
208~177-179.
621-631.
613 -627.
149-1 60.
142-145.
U.S.A. 92~5251-5257.




















