
Coupled Temporal Memories in Parkinson's Disease: A Dopamine-Related Dysfunction
16
Coupled Temporal Memories in Parkinson’ s Disease: A Dopamine-Related Dysfunction Chara Malapani Hôpital de la Salpêtrière and Columbia University Brian Rakitin University of Oregon R. Levy Hôpital de la Salpêtrière Warren H. Meck Duke University Bernard Deweer and Bruno Dubois Hôpital de la Salpêtrière John Gibbon Columbia University and N.Y.S. Psychiatric Institute Abstract Dysfunction of the basal ganglia and the brain nuclei inter- connected with them leads to disturbances of movement and cognition, including disordered timing of movement and per- ceptual timing de cits. Patients with Parkinson’s disease (PD) were studied in temporal reproduction tasks. We examined PD patients when brain dopamine (DA) transmission was impaired (OFF state) and when DA transmission was reestablished, at the time of maximal clinical bene t following administration of levodopa + apomorphine (ON state). Patients reproduced target times of 8 and 21 sec trained in blocked trials with the peak interval procedure, which were veridical in the ON state, com- parable to normative performance by healthy young and aged controls (Experiment 1). In the OFF state, temporal reproduc- tion was impaired in both accuracy and precision (variance). The 8-sec signal was reproduced as longer and the 21-sec signal was reproduced as shorter than they actually were (Experi- ment 1). This “migration” effect was dependent upon training of two different durations. When PD patients were trained on 21 sec only (Experiment 2), they showed a reproduction error in the long direction, opposite to the error produced under the dual training condition of Experiment 1. The results are dis- cussed as a mutual attraction between temporal processing systems, in memory and clock stages, when dopaminergic regu- lation in the striatum is dysfunctional. INTRODUCTION Patients with Parkinson’s disease (PD) experience dif culties when producing temporal components of movement and in programming and synchronizing mo- tor responses. These problems are re ected in increased reaction time and movement time (Bloxham, Dick, & Moore, 1987; Evarts, Teravainen, & Calne, 1981), pro- longed interonset latencies when performing sequential or simultaneous movements (Benecke, Rothwell, Dick, Day, & Marsden, 1986; Roy, Saint-Cyr, Taylor, & Lang, 1993), impaired ability to maintain a xed rhythm in tapping tasks (Nakamura, Nagasaki, & Narabayashi, 1978; O’Boyle, Freeman, & Cody, 1996; Stelmach & Worring- © 1998 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 10:3, pp. 316–331 ham, 1988; Wing, Keele, & Margolin, 1984), and increased speech production time (Lieberman et al., 1992; Volk- mann, Hefter, Lange, & Freund, 1992). Parameters such as reaction time, interonset latency, rhythm maintenance, and temporal organization of speech all require accurate timing, and dysfunctions are well correlated with the clinical phenomenon of bradykinesia in PD (Benecke et al., 1986; Hallett & Khoshbin, 1980; Nakamura et al., 1978; Volkmann et al., 1992). There is evidence that the timing de cits of patients with PD may not be limited to the motor domain. Nonmotor de cits include im- paired temporal discrimination of pairs of stimuli in the somaesthetic, visual, and auditory modalities (Artieda, Pastor, Lacruz, & Obeso, 1992), absence of interlimb
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Coupled Temporal Memories in Parkinson's Disease: A Dopamine-Related Dysfunction

Coupled Temporal Memories in ParkinsonrsquosDisease A Dopamine-Related Dysfunction
Chara MalapaniHocircpital de la Salpecirctriegravere and Columbia University
Brian RakitinUniversity of Oregon
R LevyHocircpital de la Salpecirctriegravere
Warren H MeckDuke University
Bernard Deweer and Bruno DuboisHocircpital de la Salpecirctriegravere
John GibbonColumbia University and NYS Psychiatric Institute
Abstract
Dysfunction of the basal ganglia and the brain nuclei inter-connected with them leads to disturbances of movement andcognition including disordered timing of movement and per-ceptual timing decits Patients with Parkinsonrsquos disease (PD)were studied in temporal reproduction tasks We examined PDpatients when brain dopamine (DA) transmission was impaired(OFF state) and when DA transmission was reestablished at thetime of maximal clinical benet following administration oflevodopa + apomorphine (ON state) Patients reproduced targettimes of 8 and 21 sec trained in blocked trials with the peakinterval procedure which were veridical in the ON state com-parable to normative performance by healthy young and aged
controls (Experiment 1) In the OFF state temporal reproduc-tion was impaired in both accuracy and precision (variance)The 8-sec signal was reproduced as longer and the 21-sec signalwas reproduced as shorter than they actually were (Experi-ment 1) This ldquomigrationrdquo effect was dependent upon trainingof two different durations When PD patients were trained on21 sec only (Experiment 2) they showed a reproduction errorin the long direction opposite to the error produced under thedual training condition of Experiment 1 The results are dis-cussed as a mutual attraction between temporal processingsystems in memory and clock stages when dopaminergic regu-lation in the striatum is dysfunctional
INTRODUCTION
Patients with Parkinsonrsquos disease (PD) experiencedifculties when producing temporal components ofmovement and in programming and synchronizing mo-tor responses These problems are reected in increasedreaction time and movement time (Bloxham Dick ampMoore 1987 Evarts Teravainen amp Calne 1981) pro-longed interonset latencies when performing sequentialor simultaneous movements (Benecke Rothwell DickDay amp Marsden 1986 Roy Saint-Cyr Taylor amp Lang1993) impaired ability to maintain a xed rhythm intapping tasks (Nakamura Nagasaki amp Narabayashi 1978OrsquoBoyle Freeman amp Cody 1996 Stelmach amp Worring-
copy 1998 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 103 pp 316ndash331
ham 1988 Wing Keele amp Margolin 1984) and increasedspeech production time (Lieberman et al 1992 Volk-mann Hefter Lange amp Freund 1992) Parameters such asreaction time interonset latency rhythm maintenanceand temporal organization of speech all require accuratetiming and dysfunctions are well correlated with theclinical phenomenon of bradykinesia in PD (Beneckeet al 1986 Hallett amp Khoshbin 1980 Nakamura et al1978 Volkmann et al 1992) There is evidence that thetiming decits of patients with PD may not be limitedto the motor domain Nonmotor decits include im-paired temporal discrimination of pairs of stimuli in thesomaesthetic visual and auditory modalities (ArtiedaPastor Lacruz amp Obeso 1992) absence of interlimb
synchronization in processing of bimodal simultaneousstimuli (Malapani Pillon Dubois amp Agid 1994) and im-paired time estimation (Pastor Artieda Jahanshahi ampObeso 1992)
Although all of these studies report ldquotiming-relatedrdquodecits in PD most of the experimental tasks used werenot specically designed to adress the issue of whetheran internal timekeeper is dysfunctional in PD Recentpsychophysical research suggests that several motor orperceptual tasks that require accurate timing have accessto the same internal timing mechanism because norma-tive performance varies in similar ways across tasks (Ivryamp Hazeltine 1995) A common timekeeping mechanismmay be therefore used in both production and percep-tion tasks that involve time-related decisions in the milli-second range with humans (Ivry amp Hazeltine 1995 Ivryamp Keele 1989 Keele Pokorny Corcos amp Ivry 1985) andin the seconds-to-minutes range with humans and ani-mals (Church 1984 Meck Church amp Olton 1984 Raki-tin et al 1998) Thus timing may be an independentprocess employed whenever its specic computation isneeded In fact accurate timing is required in manybehaviors across species For example the skilled move-ments of musicians athletes and animal predators re-quire precise timing of activity across different groupsof muscles and anticipation of time intervals into theseconds range
Neuropsychological research provides a complemen-tary way to address the issue of whether a commontiming mechanism is used in both production and per-ception and to investigate possible locations of the hy-pothesized internal timekeepers within neural brainsystems This approach asks whether different patientgroups will show either similarities or dissociations inperformance on specic timing tasks as a function of thelocation of their neurological lesion Patients with le-sions in the cerebellum are impaired in both motor andperceptual tasks requiring accurate timing in short (mil-liseconds) time ranges (Ivry amp Keele 1989 Ivry Keele ampDiener 1988) as well as in long (seconds) time ranges(Nichelli Alway amp Grafman 1996) Ivry (1996) suggestedthat the same timing mechanism operates in both motorand perceptual tasks for the short ranges and may in-volve pathways that pass through the cerebellum Perfor-mance in the same perceptual and production timingtasks was not found to be impaired in PD patients (Ivryamp Keele 1989) leading to the hypothesis that timingmechanisms do not involve the basal ganglia in humansat least in the millisecond time ranges (Ivry 1996) How-ever contrary ndings recently obtained in patients withdiseases originating in the basal ganglia have shown thatboth time estimation and motor tasks requiring accuratetiming are impaired in PD patients in short as well aslong ranges (OrsquoBoyle et al 1996 Pastor et al 1992) andin patients with Huntingtonrsquos disease (Freeman 1996)
Pastor et al (1992) showed that PD patients OFF theirlevodopa medication underestimated time intervals in
the seconds range Following administration of levodopaa signicant improvement in time estimation was ob-served supporting the hypothesis that dopamine playsa role in the modulation of internal timekeeping conso-nant with a variety of work with animals in the secondsrange under dopaminergic agonists and antagonists (seeMeck 1996 for review) Impaired timing of fast repeti-tive movements in PD patients OFF their medication iscommonly found (OrsquoBoyle et al 1966 Wing et al 1984)contrary to Ivry and Keelersquos data (1989) reporting nodecit in temporal judgment or temporal reproductiontasks in short ranges However it is possible that a purelymotor origin may explain observed decits in even per-ceptual time estimation tasks if subjects use vocal orsubvocal counting to estimate time intervals as in Pastoret alrsquos study (1992) So the question of whether timeestimation per se is impaired in PD remains to be an-swered
The present study assessed interval timing compe-tence in the seconds range in patients with PD The rstaims of the reported research were to test if temporalprocessing follows normative rules with aged subjectsand to see how it may break down in PD In the rstexperiment we examined temporal reproduction ofmore than one duration Subjects received blocked train-ing on one duration at a time but more than one weretrained in the same session A second experiment inwhich only one duration was trained controlled forbiases and interactions that may occur when more thanone duration has been learned We evaluated the role ofstriatal dopaminergic activity in temporal processing byexamining PD patients when brain DA transmission wasimpaired (OFF state) and when DA transmission wasreestablished at the time of maximal clinical benetfollowing administration of levodopa + apomorphine(ON state) We used a between-subject design that com-pared both ON and OFF states of patients to aged subjects(Experiment 1) and a within-subject design that com-pared PD patients in the OFF state to their own perfor-mance in the ON state (Experiments 1 and 2) Theexperimental questions were (1) Is timing competencein PD patients under the treated (ON) state comparableto normative performance (2) Is timing in the samepatients when OFF medication different either in preci-sion or accuracy (3) Are differences seen between tim-ing one and more than one temporal duration
We used the peak interval (PI) timing procedure origi-nally developed to study interval timing in animals(Catania 1970 Roberts 1981) and to isolate modularcomponents of temporal processing associated with dis-tinct brain area lesions (Meck 1983 1986 1998a 1998bMeck Church amp Olton 1984 Olton Wenk Church ampMeck 1987) Recent work (Rakitin et al 1988) has suc-cessfully used this technique to study normative tempo-ral reproduction of remembered time values in humansin the seconds-to-minutes range
This task has received extensive theoretical and em-
Malapani et al 317
pirical study Normative reproduction in animals andhumans shows a peak in responding at the target timeand variability around this peak Both accuracy (peaklocation) and precision (variability around the peak) willbe studied here in the context of the Scalar ExpectancyTheory (SET) model of temporal information processing(Figure 1)
The timing system is assumed to be comprised ofthree separable components (Figure 1a) a clock systemthat monitors the (subjective) passage of time a memorysystem that records the subjective representations of thetarget times and a decision system that compares cur-rent time to remembered time to generate appropriateresponding Both animal and human data show that vari-ability increases dramatically with increases in the targetor base duration being timed in the PI task and thisincrease follows the ldquoscalar rulerdquo (Figure 1b) A largebody of literature has established this ldquoscalar propertyrdquoa strong form of Weberrsquos Law (Allan amp Gibbon 1991Gibbon 1977 Gibbon Church amp Meck 1984) In itssimplest form the scalar property holds that errors ofestimation are strictly proportional to the target time soentire distributions of estimates superpose when scaledin proportions of the target duration This means in turnthat the standard deviation (not the variance) is propor-tional to the mean The constant of proportionality the
coefcient of variation s m (SIQRmedian) is the funda-mental sensitivity index of the interval time sense com-parable to the Weber fraction of temporal discrimination(Allen amp Gibbon 1991 Getty 1975 Ivry amp Hazeltine1995) The scalar property implies that multiplicativevariance mechanisms are responsible for errors of esti-mation at different target durations (Gibbon 1992 Gib-bon et al 1984)
Scalar variability may be induced in either of the threecomponentsmdashclock memory or decisionmdashin the tem-poral processing system Clock variance is illustrated inFigure 1b Subjective time (on the ordinate) accumulateswith real time (on the abscissa) at an average rate (heavydiagonal) However within and between trials the clockrate may vary (dashed diagonals) so that the recordedtarget times in memory show the scalar property (distri-butions on the ordinate) Scalar variance may also resultin a similar way from variability in the rate of the encod-ing and decoding process for comparison with currenttime And the comparison itself may also generate scalarvariability via a ratio of current to remembered timeexceeding a variable threshold for initiating and termi-nating responding (Gibbon 1992)
Increases in the level of variability in patient popula-tions are frequently cited in short-interval (eg tapping)tasks as indices of dysfunctional temporal processing
Figure 1 (a) Information processing schematic of the Scalar Expectancy Theory model The clock system is assumed to involve a pacemakerthat is gated into an integrator to monitor the ow of subjective time When feedback is given or on xed-time training trials when the targetduration is indicated by a change in the signal the subjective time is encoded (dashed arrow) and stored in memory On later peak interval tri-als these memory values are decoded (dashed arrow) and sampled for comparison with the current time value in the clockintegrator systemThe comparison is made by a ratio When the current subjective time is sufciently close to the remembered time (ratio close to 10) respond-ing is generated (b) Clock System Variance Subjective time is shown growing linearly with real time Variation in the rate of the subjective inte-gration (dashed diagonals) generates subjective time distributions for a short (S) and a long (L) time (on the ordinate) that are scale transformsof each other (the scalar property)
318 Journal of Cognitive Neuroscience Volume 10 Number 3
When variability is shown to be increased relative tonormative performance of control groups an importantobservation is whether or not this increased variabilityremains scalar in the target times being estimated (Gib-bon Malapani Dale amp Gallistel 1997) If so the temporalinformation processing system may have increased mul-tiplicative noise in either the clock the memory or thecomparison mechanism but is otherwise operating nor-mally If however variability is shown to violate thescalar property it is likely that the violation occurs in thememory representation because increased variability inthe clock system should remain scalar at different targettimes (see Figure 1b) and increased threshold variabilityin the comparison process should similarly remain scalar(Gibbon 1992)
Accuracy according to the SET model of temporalinformation processing is expected to be near veridicalas long as training and testing are conducted under thesame conditions Animal data with the PI procedureshow that a clock speed difference does not produce anaccuracy change (Meck 1996) unless it is introducedafter training in an alternative drug condition Animalsunder dopaminergic manipulation originally trainedwithout a dopamine agonist show a dramatic rapid shifttoward underestimation when training is continued un-der the agonist manipulation However peak estimationgradually returns to veridical accuracy as new subjectivetimes overlay previously learned memories for durationwithout the drug Thus a faster (or slower) clock mightgenerate a larger (or smaller) representation of subjec-tive time but as long as the training and testing condi-tions remain the same this same larger (or smaller)representation is accessed on testing trials and shouldbe reproduced at the appropriate veridical real targettime Similarly bias in the decision process that mightlead to systematic under- or overestimation should becorrected during training when subjects are given feed-back on this task Conversely a memory effect may beinferred when an accuracy distortion is relatively perma-nent despite intermittent corrective feedback during testsessions (Meck 1996) Thus only a permanent accuracydistortion implicates a dysfunctional memory repre-sentation induced either on storage or retrieval
In summary then accuracy and nonscalar varianceeffects in patient populations are most likely the resultof dysfunctional memory encoding or decoding where-as additional scalar noise may be present in patient popu-lations at any stage of the temporal processing system
EXPERIMENT 1
In Experiment 1 aged (n = 12 mean age 72 years)subjects with no neurological disease and PD patients(n = 10 mean age 55 years) were trained to remembertime intervals indicated by a visual signal appearing ona computer screen for a standard duration On a follow-ing test block subjects were required to reproduce the
same time intervals by pressing the space bar on thekeyboard Training and testing sessions for each durationwere blocked (20 training and 60 test trials) Aged sub-jects were tested with three different durations (8 12and 21 sec) and their performance was contrasted withthe performance of 10 young (mean age 25 years) col-lege students (previously reported by Rakitin et al1998) PD patients were tested with two durations (8and 21 sec) in both ON and OFF levodopa states Theperformance of PD patients in the OFF state was con-trasted with their own performance in the ON stage andperformances in both states were contrasted with theperformance of the aged group for these durations
Results
1 Young versus Aged Controls
Distributions of responding for the three intervals areshown in Figure 2 Rakitin et alrsquos (1998) young group isshown in the upper panel our aged group is in themiddle panel and the distributions for both groups nor-malized for time relative to the median time and propor-tions of peak responding are in the bottom panel Youngsubjectsrsquo peak estimates are very close to veridical withincreasing variability with increasing target durationsAged subjectsrsquo estimates are veridical for the 8-sec dura-tion close to veridical for the 12-sec duration and showa small underestimation (leftward shift) for the 21-secduration Normalized distributions in the bottom panelplotted in time relative to the median time show super-position across durations for both groups but with some-what larger variance at all values for the aged subjects
Means (standard deviation) of accuracy variability andthe coefcient of variation in both young and agedgroups are summarized in Table 1 The following effectswere found to be statistically reliable Accuracy Allsources of variance were signicant (Age F = 77 p lt005 Duration F = 9932 p lt 005 Age acute Duration F =41 p lt 005) Posthoc comparisons showed that theinteraction effect was due to a signicant leftward shiftof the aged group for the 21-sec duration (p lt 005)Variability A signicant effect of Duration (F = 808 p lt005) and an effect of Age that ldquoapproachedrdquo signicance(F = 37 p = 006) but no interaction effect was foundfor semi-interquartile range (SIQR) No signicant corre-lations were found between SIQRs and the neuropsy-chological scores Coefcient of Variation A signicanteffect of Age (F = 73 p lt 005) was found on SIQRme-dian No Duration or interaction effects were found Thusaged subjects have somewhat greater variability thanyoung ones which nevertheless remains scalar in theirtarget time estimates
2 ON versus OFF States of PD Patients
Distributions of responding for both ON and OFF condi-tions at both signal durations are shown in Figure 3 The
Malapani et al 319
Figure 2 Relative frequencydistributions for the three tar-get durations (8 12 and 21sec) showing accuracy andvariability of estimation inyoung college students (whiteupper panel) and in aged nor-mal controls (black middlepanel) The three functions ofthe two groups when plottedin time relative to the mediantime and normalized as pro-portions of the maximum rela-tive frequency superpose(lower panel)
Table 1 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation in Young and Aged Subjects inExperiment 1
8-sec duration 12-sec duration 21-sec duration
Young Aged Young Aged Young Aged
Median 805 (4) 805 (6) 123 (8) 117 (7) 214 (13) 197 (17)
SIQR 35 (2) 55 (3) 66 (3) 75 (3) 102 (4) 121 (9)
SIQRmedian 043 (02) 068 (04) 053 (02) 067 (03) 047 (02) 061 (05)
320 Journal of Cognitive Neuroscience Volume 10 Number 3
accuracy of temporal reproduction in PD patients whenthey are ON levodopa + apomorphine medication isveridical (open symbols upper panel) They producepeaks in responding that are very close to the targets forboth durations (mean peak times of 78 and 211 sec)The same subjects show a dramatic distortion in accu-racy for both durations when they are OFF their medica-tion (lled symbols upper panel) they overestimate theshort signal and underestimate the long one The variabil-ity of reproduction for 8 sec is also different betweenON and OFF states as can be seen in the different spread
of the curves obtained for the 8-sec target Howevervariability is roughly equal for the 21-sec target in boththe ON and OFF states because those curves are equallybroad The scalar property shown in the lower panelemerges in the ON state when responses are normalizedas a proportion of median time and scaled to maximumresponding The ON distributions show superposition Inthe OFF state however a violation of superposition isobserved The 8- and 21-sec OFF distributions do notsuperpose and they are broader in relative time than theON distributions especially for the 8-sec condition
Figure 3 Relative frequencydistributions for the two tar-get durations (8 and 21 sec)showing accuracy and variabil-ity of estimation in PD pa-tients ON (white) and OFF
(black) their levodopa medica-tion (upper panel) The twofunctions of the same subjectswhen plotted in time relativeto the median time and nor-malized as proportions of themaximum relative frequencysuperpose in the ON state butnot in the OFF state (lowerpanel)
Malapani et al 321
Means of accuracy variability and coefcient of vari-ation in the ON and OFF states of PD patients are summa-rized in Table 2 Statistical analysis showed the followingeffects Accuracy A signicant effect of Drug (F = 104p lt 005) and Duration (F = 5578 p lt 005) on themedians was found The interaction between these twosources of variance was also signicant (F = 516 p lt005) Further paired comparisons showed a signicantdifference between the medians obtained in the ON andOFF states for both 8 sec (F = 607 p lt 005) and 21-secdurations (F = 592 p lt 005) No correlations werefound between accuracy and neuropsychological or mo-tor scores in the ON state Correlations at a signicantlevel were found in the OFF state between the akinesiascore and (1) the rightward shift in accuracy for the8-sec duration (beta = 01 p lt 005) (2) the leftwardshift in accuracy for the 21-sec duration (beta = 005p lt 005) Variability A signicant effect of Duration (F= 731 p lt 005) and Drug (F = 212 p lt 005) wasfound for SIQR The interaction between the two factorswas not signicant (F = 23 p = 01) Further pairedcomparisons showed a signicant difference betweenthe SIQR obtained in the ON and OFF states for 8 sec (F= 10 16 p lt 005) but not for 21 sec (F = 099 p = 03)No correlations were found between SIQR and motor orneuropsychological scores in either drug state Coef-cient of variation A signicant effect of both Durationand Drug was found on SIQRmedian The interactionbetween the two factors was not signicant In order totest the scalar property further paired comparisons con-trasted Duration within Drug condition Coefcients ofvariation for the two durations were not signicantlydifferent in the ON condition (F = 24 p = 01) Howeverwe found a signicant difference between durations inthe OFF condition (F = 109 p lt 005)
3 Aged Controls versus ON amp OFF PD Patients
Statistical analysis between the three groups (Aged PDON PD OFF) showed the following effects Accuracy Nosignicant effect of Group on the median for the 8-sectarget was found Post hoc analysis showed a signicantdifference between the median obtained between PD ON
and PD OFF groups (p lt 005) but no signicant differ-ence between PD ON and Aged or between PD OFF and
Aged A signicant effect of Group on the median for the21-sec target was found (F = 17 9 p lt 005) Post hoccomparisons showed signicant differences between PDON and PD OFF (p lt 005) PD ON and Aged (p lt 005)and PD OFF and Aged (p lt 005) Variability A signicanteffect of Group (F = 84 p lt 005) was found on SIQRfor the 8-sec target Further post hoc comparisonsshowed a signicant difference between ON and OFF
groups (p lt 005) and between PD OFF and Aged (p lt005) but no signicant difference between PD ON andAged For the 21-sec target no signicant effect of Groupwas found and post hoc analysis showed no signicantdifference between any pair of the three groups Coef-cient of variation A signicant effect of Group wasfound on SIQRmedian for the 8-sec target (F = 67 p lt005) Further post hoc comparisons showed a sig-nicant difference between PD OFF and PD ON as wellas between PD OFF and Aged (p lt 005) However nosignicant difference was found between PD ON andAged For the 21-sec target no signicant effect of Groupwas found on SIQRmedian and the post hoc analysisshowed only a signicant effect between PD ON and PDOFF and no signicant differences between either PD ON
and Aged or PD OFF and Aged
Discussion
The comparisons between young and aged controlsshow that the scalar property for temporal informationprocessing holds with age The good superposition seenin Figure 2 is reected in the large effect of duration onSIQR and the lack of an effect of duration on SIQRme-dian However aged people showed an overall increasedvariability compared to young college students and aslight underestimation for the longer duration target Theformer effect along with the failure to nd a differencein the coefcient of variation (the scalar property) sug-gests that temporal processing is more variable in agedpeople but still conforms to Weberrsquos Law Thus as de-scribed earlier this increased variance could arise eitherfrom clock variability memory storage or retrieval vari-ability or criterion-decision variability
The small underestimation seen at 21 sec (and a stillsmaller underestimation at 12 sec) is probably due to adysfunction in mnemonic processing because accuracy
Table 2 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation for PD Patients ON and OFF Medicationin Experiment 1
8-sec duration 21-sec duration
PD ON PD OFF PD ON PD OFF
Median 76 (6) 89 (206) 211 (8) 173 (15)
SIQR 45 (2) 82 (5) 115 (5) 125 (4)
SIQRmedian 059 (03) 092 (03) 054 (02) 072 (03)
322 Journal of Cognitive Neuroscience Volume 10 Number 3
effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

synchronization in processing of bimodal simultaneousstimuli (Malapani Pillon Dubois amp Agid 1994) and im-paired time estimation (Pastor Artieda Jahanshahi ampObeso 1992)
Although all of these studies report ldquotiming-relatedrdquodecits in PD most of the experimental tasks used werenot specically designed to adress the issue of whetheran internal timekeeper is dysfunctional in PD Recentpsychophysical research suggests that several motor orperceptual tasks that require accurate timing have accessto the same internal timing mechanism because norma-tive performance varies in similar ways across tasks (Ivryamp Hazeltine 1995) A common timekeeping mechanismmay be therefore used in both production and percep-tion tasks that involve time-related decisions in the milli-second range with humans (Ivry amp Hazeltine 1995 Ivryamp Keele 1989 Keele Pokorny Corcos amp Ivry 1985) andin the seconds-to-minutes range with humans and ani-mals (Church 1984 Meck Church amp Olton 1984 Raki-tin et al 1998) Thus timing may be an independentprocess employed whenever its specic computation isneeded In fact accurate timing is required in manybehaviors across species For example the skilled move-ments of musicians athletes and animal predators re-quire precise timing of activity across different groupsof muscles and anticipation of time intervals into theseconds range
Neuropsychological research provides a complemen-tary way to address the issue of whether a commontiming mechanism is used in both production and per-ception and to investigate possible locations of the hy-pothesized internal timekeepers within neural brainsystems This approach asks whether different patientgroups will show either similarities or dissociations inperformance on specic timing tasks as a function of thelocation of their neurological lesion Patients with le-sions in the cerebellum are impaired in both motor andperceptual tasks requiring accurate timing in short (mil-liseconds) time ranges (Ivry amp Keele 1989 Ivry Keele ampDiener 1988) as well as in long (seconds) time ranges(Nichelli Alway amp Grafman 1996) Ivry (1996) suggestedthat the same timing mechanism operates in both motorand perceptual tasks for the short ranges and may in-volve pathways that pass through the cerebellum Perfor-mance in the same perceptual and production timingtasks was not found to be impaired in PD patients (Ivryamp Keele 1989) leading to the hypothesis that timingmechanisms do not involve the basal ganglia in humansat least in the millisecond time ranges (Ivry 1996) How-ever contrary ndings recently obtained in patients withdiseases originating in the basal ganglia have shown thatboth time estimation and motor tasks requiring accuratetiming are impaired in PD patients in short as well aslong ranges (OrsquoBoyle et al 1996 Pastor et al 1992) andin patients with Huntingtonrsquos disease (Freeman 1996)
Pastor et al (1992) showed that PD patients OFF theirlevodopa medication underestimated time intervals in
the seconds range Following administration of levodopaa signicant improvement in time estimation was ob-served supporting the hypothesis that dopamine playsa role in the modulation of internal timekeeping conso-nant with a variety of work with animals in the secondsrange under dopaminergic agonists and antagonists (seeMeck 1996 for review) Impaired timing of fast repeti-tive movements in PD patients OFF their medication iscommonly found (OrsquoBoyle et al 1966 Wing et al 1984)contrary to Ivry and Keelersquos data (1989) reporting nodecit in temporal judgment or temporal reproductiontasks in short ranges However it is possible that a purelymotor origin may explain observed decits in even per-ceptual time estimation tasks if subjects use vocal orsubvocal counting to estimate time intervals as in Pastoret alrsquos study (1992) So the question of whether timeestimation per se is impaired in PD remains to be an-swered
The present study assessed interval timing compe-tence in the seconds range in patients with PD The rstaims of the reported research were to test if temporalprocessing follows normative rules with aged subjectsand to see how it may break down in PD In the rstexperiment we examined temporal reproduction ofmore than one duration Subjects received blocked train-ing on one duration at a time but more than one weretrained in the same session A second experiment inwhich only one duration was trained controlled forbiases and interactions that may occur when more thanone duration has been learned We evaluated the role ofstriatal dopaminergic activity in temporal processing byexamining PD patients when brain DA transmission wasimpaired (OFF state) and when DA transmission wasreestablished at the time of maximal clinical benetfollowing administration of levodopa + apomorphine(ON state) We used a between-subject design that com-pared both ON and OFF states of patients to aged subjects(Experiment 1) and a within-subject design that com-pared PD patients in the OFF state to their own perfor-mance in the ON state (Experiments 1 and 2) Theexperimental questions were (1) Is timing competencein PD patients under the treated (ON) state comparableto normative performance (2) Is timing in the samepatients when OFF medication different either in preci-sion or accuracy (3) Are differences seen between tim-ing one and more than one temporal duration
We used the peak interval (PI) timing procedure origi-nally developed to study interval timing in animals(Catania 1970 Roberts 1981) and to isolate modularcomponents of temporal processing associated with dis-tinct brain area lesions (Meck 1983 1986 1998a 1998bMeck Church amp Olton 1984 Olton Wenk Church ampMeck 1987) Recent work (Rakitin et al 1988) has suc-cessfully used this technique to study normative tempo-ral reproduction of remembered time values in humansin the seconds-to-minutes range
This task has received extensive theoretical and em-
Malapani et al 317
pirical study Normative reproduction in animals andhumans shows a peak in responding at the target timeand variability around this peak Both accuracy (peaklocation) and precision (variability around the peak) willbe studied here in the context of the Scalar ExpectancyTheory (SET) model of temporal information processing(Figure 1)
The timing system is assumed to be comprised ofthree separable components (Figure 1a) a clock systemthat monitors the (subjective) passage of time a memorysystem that records the subjective representations of thetarget times and a decision system that compares cur-rent time to remembered time to generate appropriateresponding Both animal and human data show that vari-ability increases dramatically with increases in the targetor base duration being timed in the PI task and thisincrease follows the ldquoscalar rulerdquo (Figure 1b) A largebody of literature has established this ldquoscalar propertyrdquoa strong form of Weberrsquos Law (Allan amp Gibbon 1991Gibbon 1977 Gibbon Church amp Meck 1984) In itssimplest form the scalar property holds that errors ofestimation are strictly proportional to the target time soentire distributions of estimates superpose when scaledin proportions of the target duration This means in turnthat the standard deviation (not the variance) is propor-tional to the mean The constant of proportionality the
coefcient of variation s m (SIQRmedian) is the funda-mental sensitivity index of the interval time sense com-parable to the Weber fraction of temporal discrimination(Allen amp Gibbon 1991 Getty 1975 Ivry amp Hazeltine1995) The scalar property implies that multiplicativevariance mechanisms are responsible for errors of esti-mation at different target durations (Gibbon 1992 Gib-bon et al 1984)
Scalar variability may be induced in either of the threecomponentsmdashclock memory or decisionmdashin the tem-poral processing system Clock variance is illustrated inFigure 1b Subjective time (on the ordinate) accumulateswith real time (on the abscissa) at an average rate (heavydiagonal) However within and between trials the clockrate may vary (dashed diagonals) so that the recordedtarget times in memory show the scalar property (distri-butions on the ordinate) Scalar variance may also resultin a similar way from variability in the rate of the encod-ing and decoding process for comparison with currenttime And the comparison itself may also generate scalarvariability via a ratio of current to remembered timeexceeding a variable threshold for initiating and termi-nating responding (Gibbon 1992)
Increases in the level of variability in patient popula-tions are frequently cited in short-interval (eg tapping)tasks as indices of dysfunctional temporal processing
Figure 1 (a) Information processing schematic of the Scalar Expectancy Theory model The clock system is assumed to involve a pacemakerthat is gated into an integrator to monitor the ow of subjective time When feedback is given or on xed-time training trials when the targetduration is indicated by a change in the signal the subjective time is encoded (dashed arrow) and stored in memory On later peak interval tri-als these memory values are decoded (dashed arrow) and sampled for comparison with the current time value in the clockintegrator systemThe comparison is made by a ratio When the current subjective time is sufciently close to the remembered time (ratio close to 10) respond-ing is generated (b) Clock System Variance Subjective time is shown growing linearly with real time Variation in the rate of the subjective inte-gration (dashed diagonals) generates subjective time distributions for a short (S) and a long (L) time (on the ordinate) that are scale transformsof each other (the scalar property)
318 Journal of Cognitive Neuroscience Volume 10 Number 3
When variability is shown to be increased relative tonormative performance of control groups an importantobservation is whether or not this increased variabilityremains scalar in the target times being estimated (Gib-bon Malapani Dale amp Gallistel 1997) If so the temporalinformation processing system may have increased mul-tiplicative noise in either the clock the memory or thecomparison mechanism but is otherwise operating nor-mally If however variability is shown to violate thescalar property it is likely that the violation occurs in thememory representation because increased variability inthe clock system should remain scalar at different targettimes (see Figure 1b) and increased threshold variabilityin the comparison process should similarly remain scalar(Gibbon 1992)
Accuracy according to the SET model of temporalinformation processing is expected to be near veridicalas long as training and testing are conducted under thesame conditions Animal data with the PI procedureshow that a clock speed difference does not produce anaccuracy change (Meck 1996) unless it is introducedafter training in an alternative drug condition Animalsunder dopaminergic manipulation originally trainedwithout a dopamine agonist show a dramatic rapid shifttoward underestimation when training is continued un-der the agonist manipulation However peak estimationgradually returns to veridical accuracy as new subjectivetimes overlay previously learned memories for durationwithout the drug Thus a faster (or slower) clock mightgenerate a larger (or smaller) representation of subjec-tive time but as long as the training and testing condi-tions remain the same this same larger (or smaller)representation is accessed on testing trials and shouldbe reproduced at the appropriate veridical real targettime Similarly bias in the decision process that mightlead to systematic under- or overestimation should becorrected during training when subjects are given feed-back on this task Conversely a memory effect may beinferred when an accuracy distortion is relatively perma-nent despite intermittent corrective feedback during testsessions (Meck 1996) Thus only a permanent accuracydistortion implicates a dysfunctional memory repre-sentation induced either on storage or retrieval
In summary then accuracy and nonscalar varianceeffects in patient populations are most likely the resultof dysfunctional memory encoding or decoding where-as additional scalar noise may be present in patient popu-lations at any stage of the temporal processing system
EXPERIMENT 1
In Experiment 1 aged (n = 12 mean age 72 years)subjects with no neurological disease and PD patients(n = 10 mean age 55 years) were trained to remembertime intervals indicated by a visual signal appearing ona computer screen for a standard duration On a follow-ing test block subjects were required to reproduce the
same time intervals by pressing the space bar on thekeyboard Training and testing sessions for each durationwere blocked (20 training and 60 test trials) Aged sub-jects were tested with three different durations (8 12and 21 sec) and their performance was contrasted withthe performance of 10 young (mean age 25 years) col-lege students (previously reported by Rakitin et al1998) PD patients were tested with two durations (8and 21 sec) in both ON and OFF levodopa states Theperformance of PD patients in the OFF state was con-trasted with their own performance in the ON stage andperformances in both states were contrasted with theperformance of the aged group for these durations
Results
1 Young versus Aged Controls
Distributions of responding for the three intervals areshown in Figure 2 Rakitin et alrsquos (1998) young group isshown in the upper panel our aged group is in themiddle panel and the distributions for both groups nor-malized for time relative to the median time and propor-tions of peak responding are in the bottom panel Youngsubjectsrsquo peak estimates are very close to veridical withincreasing variability with increasing target durationsAged subjectsrsquo estimates are veridical for the 8-sec dura-tion close to veridical for the 12-sec duration and showa small underestimation (leftward shift) for the 21-secduration Normalized distributions in the bottom panelplotted in time relative to the median time show super-position across durations for both groups but with some-what larger variance at all values for the aged subjects
Means (standard deviation) of accuracy variability andthe coefcient of variation in both young and agedgroups are summarized in Table 1 The following effectswere found to be statistically reliable Accuracy Allsources of variance were signicant (Age F = 77 p lt005 Duration F = 9932 p lt 005 Age acute Duration F =41 p lt 005) Posthoc comparisons showed that theinteraction effect was due to a signicant leftward shiftof the aged group for the 21-sec duration (p lt 005)Variability A signicant effect of Duration (F = 808 p lt005) and an effect of Age that ldquoapproachedrdquo signicance(F = 37 p = 006) but no interaction effect was foundfor semi-interquartile range (SIQR) No signicant corre-lations were found between SIQRs and the neuropsy-chological scores Coefcient of Variation A signicanteffect of Age (F = 73 p lt 005) was found on SIQRme-dian No Duration or interaction effects were found Thusaged subjects have somewhat greater variability thanyoung ones which nevertheless remains scalar in theirtarget time estimates
2 ON versus OFF States of PD Patients
Distributions of responding for both ON and OFF condi-tions at both signal durations are shown in Figure 3 The
Malapani et al 319
Figure 2 Relative frequencydistributions for the three tar-get durations (8 12 and 21sec) showing accuracy andvariability of estimation inyoung college students (whiteupper panel) and in aged nor-mal controls (black middlepanel) The three functions ofthe two groups when plottedin time relative to the mediantime and normalized as pro-portions of the maximum rela-tive frequency superpose(lower panel)
Table 1 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation in Young and Aged Subjects inExperiment 1
8-sec duration 12-sec duration 21-sec duration
Young Aged Young Aged Young Aged
Median 805 (4) 805 (6) 123 (8) 117 (7) 214 (13) 197 (17)
SIQR 35 (2) 55 (3) 66 (3) 75 (3) 102 (4) 121 (9)
SIQRmedian 043 (02) 068 (04) 053 (02) 067 (03) 047 (02) 061 (05)
320 Journal of Cognitive Neuroscience Volume 10 Number 3
accuracy of temporal reproduction in PD patients whenthey are ON levodopa + apomorphine medication isveridical (open symbols upper panel) They producepeaks in responding that are very close to the targets forboth durations (mean peak times of 78 and 211 sec)The same subjects show a dramatic distortion in accu-racy for both durations when they are OFF their medica-tion (lled symbols upper panel) they overestimate theshort signal and underestimate the long one The variabil-ity of reproduction for 8 sec is also different betweenON and OFF states as can be seen in the different spread
of the curves obtained for the 8-sec target Howevervariability is roughly equal for the 21-sec target in boththe ON and OFF states because those curves are equallybroad The scalar property shown in the lower panelemerges in the ON state when responses are normalizedas a proportion of median time and scaled to maximumresponding The ON distributions show superposition Inthe OFF state however a violation of superposition isobserved The 8- and 21-sec OFF distributions do notsuperpose and they are broader in relative time than theON distributions especially for the 8-sec condition
Figure 3 Relative frequencydistributions for the two tar-get durations (8 and 21 sec)showing accuracy and variabil-ity of estimation in PD pa-tients ON (white) and OFF
(black) their levodopa medica-tion (upper panel) The twofunctions of the same subjectswhen plotted in time relativeto the median time and nor-malized as proportions of themaximum relative frequencysuperpose in the ON state butnot in the OFF state (lowerpanel)
Malapani et al 321
Means of accuracy variability and coefcient of vari-ation in the ON and OFF states of PD patients are summa-rized in Table 2 Statistical analysis showed the followingeffects Accuracy A signicant effect of Drug (F = 104p lt 005) and Duration (F = 5578 p lt 005) on themedians was found The interaction between these twosources of variance was also signicant (F = 516 p lt005) Further paired comparisons showed a signicantdifference between the medians obtained in the ON andOFF states for both 8 sec (F = 607 p lt 005) and 21-secdurations (F = 592 p lt 005) No correlations werefound between accuracy and neuropsychological or mo-tor scores in the ON state Correlations at a signicantlevel were found in the OFF state between the akinesiascore and (1) the rightward shift in accuracy for the8-sec duration (beta = 01 p lt 005) (2) the leftwardshift in accuracy for the 21-sec duration (beta = 005p lt 005) Variability A signicant effect of Duration (F= 731 p lt 005) and Drug (F = 212 p lt 005) wasfound for SIQR The interaction between the two factorswas not signicant (F = 23 p = 01) Further pairedcomparisons showed a signicant difference betweenthe SIQR obtained in the ON and OFF states for 8 sec (F= 10 16 p lt 005) but not for 21 sec (F = 099 p = 03)No correlations were found between SIQR and motor orneuropsychological scores in either drug state Coef-cient of variation A signicant effect of both Durationand Drug was found on SIQRmedian The interactionbetween the two factors was not signicant In order totest the scalar property further paired comparisons con-trasted Duration within Drug condition Coefcients ofvariation for the two durations were not signicantlydifferent in the ON condition (F = 24 p = 01) Howeverwe found a signicant difference between durations inthe OFF condition (F = 109 p lt 005)
3 Aged Controls versus ON amp OFF PD Patients
Statistical analysis between the three groups (Aged PDON PD OFF) showed the following effects Accuracy Nosignicant effect of Group on the median for the 8-sectarget was found Post hoc analysis showed a signicantdifference between the median obtained between PD ON
and PD OFF groups (p lt 005) but no signicant differ-ence between PD ON and Aged or between PD OFF and
Aged A signicant effect of Group on the median for the21-sec target was found (F = 17 9 p lt 005) Post hoccomparisons showed signicant differences between PDON and PD OFF (p lt 005) PD ON and Aged (p lt 005)and PD OFF and Aged (p lt 005) Variability A signicanteffect of Group (F = 84 p lt 005) was found on SIQRfor the 8-sec target Further post hoc comparisonsshowed a signicant difference between ON and OFF
groups (p lt 005) and between PD OFF and Aged (p lt005) but no signicant difference between PD ON andAged For the 21-sec target no signicant effect of Groupwas found and post hoc analysis showed no signicantdifference between any pair of the three groups Coef-cient of variation A signicant effect of Group wasfound on SIQRmedian for the 8-sec target (F = 67 p lt005) Further post hoc comparisons showed a sig-nicant difference between PD OFF and PD ON as wellas between PD OFF and Aged (p lt 005) However nosignicant difference was found between PD ON andAged For the 21-sec target no signicant effect of Groupwas found on SIQRmedian and the post hoc analysisshowed only a signicant effect between PD ON and PDOFF and no signicant differences between either PD ON
and Aged or PD OFF and Aged
Discussion
The comparisons between young and aged controlsshow that the scalar property for temporal informationprocessing holds with age The good superposition seenin Figure 2 is reected in the large effect of duration onSIQR and the lack of an effect of duration on SIQRme-dian However aged people showed an overall increasedvariability compared to young college students and aslight underestimation for the longer duration target Theformer effect along with the failure to nd a differencein the coefcient of variation (the scalar property) sug-gests that temporal processing is more variable in agedpeople but still conforms to Weberrsquos Law Thus as de-scribed earlier this increased variance could arise eitherfrom clock variability memory storage or retrieval vari-ability or criterion-decision variability
The small underestimation seen at 21 sec (and a stillsmaller underestimation at 12 sec) is probably due to adysfunction in mnemonic processing because accuracy
Table 2 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation for PD Patients ON and OFF Medicationin Experiment 1
8-sec duration 21-sec duration
PD ON PD OFF PD ON PD OFF
Median 76 (6) 89 (206) 211 (8) 173 (15)
SIQR 45 (2) 82 (5) 115 (5) 125 (4)
SIQRmedian 059 (03) 092 (03) 054 (02) 072 (03)
322 Journal of Cognitive Neuroscience Volume 10 Number 3
effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

pirical study Normative reproduction in animals andhumans shows a peak in responding at the target timeand variability around this peak Both accuracy (peaklocation) and precision (variability around the peak) willbe studied here in the context of the Scalar ExpectancyTheory (SET) model of temporal information processing(Figure 1)
The timing system is assumed to be comprised ofthree separable components (Figure 1a) a clock systemthat monitors the (subjective) passage of time a memorysystem that records the subjective representations of thetarget times and a decision system that compares cur-rent time to remembered time to generate appropriateresponding Both animal and human data show that vari-ability increases dramatically with increases in the targetor base duration being timed in the PI task and thisincrease follows the ldquoscalar rulerdquo (Figure 1b) A largebody of literature has established this ldquoscalar propertyrdquoa strong form of Weberrsquos Law (Allan amp Gibbon 1991Gibbon 1977 Gibbon Church amp Meck 1984) In itssimplest form the scalar property holds that errors ofestimation are strictly proportional to the target time soentire distributions of estimates superpose when scaledin proportions of the target duration This means in turnthat the standard deviation (not the variance) is propor-tional to the mean The constant of proportionality the
coefcient of variation s m (SIQRmedian) is the funda-mental sensitivity index of the interval time sense com-parable to the Weber fraction of temporal discrimination(Allen amp Gibbon 1991 Getty 1975 Ivry amp Hazeltine1995) The scalar property implies that multiplicativevariance mechanisms are responsible for errors of esti-mation at different target durations (Gibbon 1992 Gib-bon et al 1984)
Scalar variability may be induced in either of the threecomponentsmdashclock memory or decisionmdashin the tem-poral processing system Clock variance is illustrated inFigure 1b Subjective time (on the ordinate) accumulateswith real time (on the abscissa) at an average rate (heavydiagonal) However within and between trials the clockrate may vary (dashed diagonals) so that the recordedtarget times in memory show the scalar property (distri-butions on the ordinate) Scalar variance may also resultin a similar way from variability in the rate of the encod-ing and decoding process for comparison with currenttime And the comparison itself may also generate scalarvariability via a ratio of current to remembered timeexceeding a variable threshold for initiating and termi-nating responding (Gibbon 1992)
Increases in the level of variability in patient popula-tions are frequently cited in short-interval (eg tapping)tasks as indices of dysfunctional temporal processing
Figure 1 (a) Information processing schematic of the Scalar Expectancy Theory model The clock system is assumed to involve a pacemakerthat is gated into an integrator to monitor the ow of subjective time When feedback is given or on xed-time training trials when the targetduration is indicated by a change in the signal the subjective time is encoded (dashed arrow) and stored in memory On later peak interval tri-als these memory values are decoded (dashed arrow) and sampled for comparison with the current time value in the clockintegrator systemThe comparison is made by a ratio When the current subjective time is sufciently close to the remembered time (ratio close to 10) respond-ing is generated (b) Clock System Variance Subjective time is shown growing linearly with real time Variation in the rate of the subjective inte-gration (dashed diagonals) generates subjective time distributions for a short (S) and a long (L) time (on the ordinate) that are scale transformsof each other (the scalar property)
318 Journal of Cognitive Neuroscience Volume 10 Number 3
When variability is shown to be increased relative tonormative performance of control groups an importantobservation is whether or not this increased variabilityremains scalar in the target times being estimated (Gib-bon Malapani Dale amp Gallistel 1997) If so the temporalinformation processing system may have increased mul-tiplicative noise in either the clock the memory or thecomparison mechanism but is otherwise operating nor-mally If however variability is shown to violate thescalar property it is likely that the violation occurs in thememory representation because increased variability inthe clock system should remain scalar at different targettimes (see Figure 1b) and increased threshold variabilityin the comparison process should similarly remain scalar(Gibbon 1992)
Accuracy according to the SET model of temporalinformation processing is expected to be near veridicalas long as training and testing are conducted under thesame conditions Animal data with the PI procedureshow that a clock speed difference does not produce anaccuracy change (Meck 1996) unless it is introducedafter training in an alternative drug condition Animalsunder dopaminergic manipulation originally trainedwithout a dopamine agonist show a dramatic rapid shifttoward underestimation when training is continued un-der the agonist manipulation However peak estimationgradually returns to veridical accuracy as new subjectivetimes overlay previously learned memories for durationwithout the drug Thus a faster (or slower) clock mightgenerate a larger (or smaller) representation of subjec-tive time but as long as the training and testing condi-tions remain the same this same larger (or smaller)representation is accessed on testing trials and shouldbe reproduced at the appropriate veridical real targettime Similarly bias in the decision process that mightlead to systematic under- or overestimation should becorrected during training when subjects are given feed-back on this task Conversely a memory effect may beinferred when an accuracy distortion is relatively perma-nent despite intermittent corrective feedback during testsessions (Meck 1996) Thus only a permanent accuracydistortion implicates a dysfunctional memory repre-sentation induced either on storage or retrieval
In summary then accuracy and nonscalar varianceeffects in patient populations are most likely the resultof dysfunctional memory encoding or decoding where-as additional scalar noise may be present in patient popu-lations at any stage of the temporal processing system
EXPERIMENT 1
In Experiment 1 aged (n = 12 mean age 72 years)subjects with no neurological disease and PD patients(n = 10 mean age 55 years) were trained to remembertime intervals indicated by a visual signal appearing ona computer screen for a standard duration On a follow-ing test block subjects were required to reproduce the
same time intervals by pressing the space bar on thekeyboard Training and testing sessions for each durationwere blocked (20 training and 60 test trials) Aged sub-jects were tested with three different durations (8 12and 21 sec) and their performance was contrasted withthe performance of 10 young (mean age 25 years) col-lege students (previously reported by Rakitin et al1998) PD patients were tested with two durations (8and 21 sec) in both ON and OFF levodopa states Theperformance of PD patients in the OFF state was con-trasted with their own performance in the ON stage andperformances in both states were contrasted with theperformance of the aged group for these durations
Results
1 Young versus Aged Controls
Distributions of responding for the three intervals areshown in Figure 2 Rakitin et alrsquos (1998) young group isshown in the upper panel our aged group is in themiddle panel and the distributions for both groups nor-malized for time relative to the median time and propor-tions of peak responding are in the bottom panel Youngsubjectsrsquo peak estimates are very close to veridical withincreasing variability with increasing target durationsAged subjectsrsquo estimates are veridical for the 8-sec dura-tion close to veridical for the 12-sec duration and showa small underestimation (leftward shift) for the 21-secduration Normalized distributions in the bottom panelplotted in time relative to the median time show super-position across durations for both groups but with some-what larger variance at all values for the aged subjects
Means (standard deviation) of accuracy variability andthe coefcient of variation in both young and agedgroups are summarized in Table 1 The following effectswere found to be statistically reliable Accuracy Allsources of variance were signicant (Age F = 77 p lt005 Duration F = 9932 p lt 005 Age acute Duration F =41 p lt 005) Posthoc comparisons showed that theinteraction effect was due to a signicant leftward shiftof the aged group for the 21-sec duration (p lt 005)Variability A signicant effect of Duration (F = 808 p lt005) and an effect of Age that ldquoapproachedrdquo signicance(F = 37 p = 006) but no interaction effect was foundfor semi-interquartile range (SIQR) No signicant corre-lations were found between SIQRs and the neuropsy-chological scores Coefcient of Variation A signicanteffect of Age (F = 73 p lt 005) was found on SIQRme-dian No Duration or interaction effects were found Thusaged subjects have somewhat greater variability thanyoung ones which nevertheless remains scalar in theirtarget time estimates
2 ON versus OFF States of PD Patients
Distributions of responding for both ON and OFF condi-tions at both signal durations are shown in Figure 3 The
Malapani et al 319
Figure 2 Relative frequencydistributions for the three tar-get durations (8 12 and 21sec) showing accuracy andvariability of estimation inyoung college students (whiteupper panel) and in aged nor-mal controls (black middlepanel) The three functions ofthe two groups when plottedin time relative to the mediantime and normalized as pro-portions of the maximum rela-tive frequency superpose(lower panel)
Table 1 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation in Young and Aged Subjects inExperiment 1
8-sec duration 12-sec duration 21-sec duration
Young Aged Young Aged Young Aged
Median 805 (4) 805 (6) 123 (8) 117 (7) 214 (13) 197 (17)
SIQR 35 (2) 55 (3) 66 (3) 75 (3) 102 (4) 121 (9)
SIQRmedian 043 (02) 068 (04) 053 (02) 067 (03) 047 (02) 061 (05)
320 Journal of Cognitive Neuroscience Volume 10 Number 3
accuracy of temporal reproduction in PD patients whenthey are ON levodopa + apomorphine medication isveridical (open symbols upper panel) They producepeaks in responding that are very close to the targets forboth durations (mean peak times of 78 and 211 sec)The same subjects show a dramatic distortion in accu-racy for both durations when they are OFF their medica-tion (lled symbols upper panel) they overestimate theshort signal and underestimate the long one The variabil-ity of reproduction for 8 sec is also different betweenON and OFF states as can be seen in the different spread
of the curves obtained for the 8-sec target Howevervariability is roughly equal for the 21-sec target in boththe ON and OFF states because those curves are equallybroad The scalar property shown in the lower panelemerges in the ON state when responses are normalizedas a proportion of median time and scaled to maximumresponding The ON distributions show superposition Inthe OFF state however a violation of superposition isobserved The 8- and 21-sec OFF distributions do notsuperpose and they are broader in relative time than theON distributions especially for the 8-sec condition
Figure 3 Relative frequencydistributions for the two tar-get durations (8 and 21 sec)showing accuracy and variabil-ity of estimation in PD pa-tients ON (white) and OFF
(black) their levodopa medica-tion (upper panel) The twofunctions of the same subjectswhen plotted in time relativeto the median time and nor-malized as proportions of themaximum relative frequencysuperpose in the ON state butnot in the OFF state (lowerpanel)
Malapani et al 321
Means of accuracy variability and coefcient of vari-ation in the ON and OFF states of PD patients are summa-rized in Table 2 Statistical analysis showed the followingeffects Accuracy A signicant effect of Drug (F = 104p lt 005) and Duration (F = 5578 p lt 005) on themedians was found The interaction between these twosources of variance was also signicant (F = 516 p lt005) Further paired comparisons showed a signicantdifference between the medians obtained in the ON andOFF states for both 8 sec (F = 607 p lt 005) and 21-secdurations (F = 592 p lt 005) No correlations werefound between accuracy and neuropsychological or mo-tor scores in the ON state Correlations at a signicantlevel were found in the OFF state between the akinesiascore and (1) the rightward shift in accuracy for the8-sec duration (beta = 01 p lt 005) (2) the leftwardshift in accuracy for the 21-sec duration (beta = 005p lt 005) Variability A signicant effect of Duration (F= 731 p lt 005) and Drug (F = 212 p lt 005) wasfound for SIQR The interaction between the two factorswas not signicant (F = 23 p = 01) Further pairedcomparisons showed a signicant difference betweenthe SIQR obtained in the ON and OFF states for 8 sec (F= 10 16 p lt 005) but not for 21 sec (F = 099 p = 03)No correlations were found between SIQR and motor orneuropsychological scores in either drug state Coef-cient of variation A signicant effect of both Durationand Drug was found on SIQRmedian The interactionbetween the two factors was not signicant In order totest the scalar property further paired comparisons con-trasted Duration within Drug condition Coefcients ofvariation for the two durations were not signicantlydifferent in the ON condition (F = 24 p = 01) Howeverwe found a signicant difference between durations inthe OFF condition (F = 109 p lt 005)
3 Aged Controls versus ON amp OFF PD Patients
Statistical analysis between the three groups (Aged PDON PD OFF) showed the following effects Accuracy Nosignicant effect of Group on the median for the 8-sectarget was found Post hoc analysis showed a signicantdifference between the median obtained between PD ON
and PD OFF groups (p lt 005) but no signicant differ-ence between PD ON and Aged or between PD OFF and
Aged A signicant effect of Group on the median for the21-sec target was found (F = 17 9 p lt 005) Post hoccomparisons showed signicant differences between PDON and PD OFF (p lt 005) PD ON and Aged (p lt 005)and PD OFF and Aged (p lt 005) Variability A signicanteffect of Group (F = 84 p lt 005) was found on SIQRfor the 8-sec target Further post hoc comparisonsshowed a signicant difference between ON and OFF
groups (p lt 005) and between PD OFF and Aged (p lt005) but no signicant difference between PD ON andAged For the 21-sec target no signicant effect of Groupwas found and post hoc analysis showed no signicantdifference between any pair of the three groups Coef-cient of variation A signicant effect of Group wasfound on SIQRmedian for the 8-sec target (F = 67 p lt005) Further post hoc comparisons showed a sig-nicant difference between PD OFF and PD ON as wellas between PD OFF and Aged (p lt 005) However nosignicant difference was found between PD ON andAged For the 21-sec target no signicant effect of Groupwas found on SIQRmedian and the post hoc analysisshowed only a signicant effect between PD ON and PDOFF and no signicant differences between either PD ON
and Aged or PD OFF and Aged
Discussion
The comparisons between young and aged controlsshow that the scalar property for temporal informationprocessing holds with age The good superposition seenin Figure 2 is reected in the large effect of duration onSIQR and the lack of an effect of duration on SIQRme-dian However aged people showed an overall increasedvariability compared to young college students and aslight underestimation for the longer duration target Theformer effect along with the failure to nd a differencein the coefcient of variation (the scalar property) sug-gests that temporal processing is more variable in agedpeople but still conforms to Weberrsquos Law Thus as de-scribed earlier this increased variance could arise eitherfrom clock variability memory storage or retrieval vari-ability or criterion-decision variability
The small underestimation seen at 21 sec (and a stillsmaller underestimation at 12 sec) is probably due to adysfunction in mnemonic processing because accuracy
Table 2 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation for PD Patients ON and OFF Medicationin Experiment 1
8-sec duration 21-sec duration
PD ON PD OFF PD ON PD OFF
Median 76 (6) 89 (206) 211 (8) 173 (15)
SIQR 45 (2) 82 (5) 115 (5) 125 (4)
SIQRmedian 059 (03) 092 (03) 054 (02) 072 (03)
322 Journal of Cognitive Neuroscience Volume 10 Number 3
effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

When variability is shown to be increased relative tonormative performance of control groups an importantobservation is whether or not this increased variabilityremains scalar in the target times being estimated (Gib-bon Malapani Dale amp Gallistel 1997) If so the temporalinformation processing system may have increased mul-tiplicative noise in either the clock the memory or thecomparison mechanism but is otherwise operating nor-mally If however variability is shown to violate thescalar property it is likely that the violation occurs in thememory representation because increased variability inthe clock system should remain scalar at different targettimes (see Figure 1b) and increased threshold variabilityin the comparison process should similarly remain scalar(Gibbon 1992)
Accuracy according to the SET model of temporalinformation processing is expected to be near veridicalas long as training and testing are conducted under thesame conditions Animal data with the PI procedureshow that a clock speed difference does not produce anaccuracy change (Meck 1996) unless it is introducedafter training in an alternative drug condition Animalsunder dopaminergic manipulation originally trainedwithout a dopamine agonist show a dramatic rapid shifttoward underestimation when training is continued un-der the agonist manipulation However peak estimationgradually returns to veridical accuracy as new subjectivetimes overlay previously learned memories for durationwithout the drug Thus a faster (or slower) clock mightgenerate a larger (or smaller) representation of subjec-tive time but as long as the training and testing condi-tions remain the same this same larger (or smaller)representation is accessed on testing trials and shouldbe reproduced at the appropriate veridical real targettime Similarly bias in the decision process that mightlead to systematic under- or overestimation should becorrected during training when subjects are given feed-back on this task Conversely a memory effect may beinferred when an accuracy distortion is relatively perma-nent despite intermittent corrective feedback during testsessions (Meck 1996) Thus only a permanent accuracydistortion implicates a dysfunctional memory repre-sentation induced either on storage or retrieval
In summary then accuracy and nonscalar varianceeffects in patient populations are most likely the resultof dysfunctional memory encoding or decoding where-as additional scalar noise may be present in patient popu-lations at any stage of the temporal processing system
EXPERIMENT 1
In Experiment 1 aged (n = 12 mean age 72 years)subjects with no neurological disease and PD patients(n = 10 mean age 55 years) were trained to remembertime intervals indicated by a visual signal appearing ona computer screen for a standard duration On a follow-ing test block subjects were required to reproduce the
same time intervals by pressing the space bar on thekeyboard Training and testing sessions for each durationwere blocked (20 training and 60 test trials) Aged sub-jects were tested with three different durations (8 12and 21 sec) and their performance was contrasted withthe performance of 10 young (mean age 25 years) col-lege students (previously reported by Rakitin et al1998) PD patients were tested with two durations (8and 21 sec) in both ON and OFF levodopa states Theperformance of PD patients in the OFF state was con-trasted with their own performance in the ON stage andperformances in both states were contrasted with theperformance of the aged group for these durations
Results
1 Young versus Aged Controls
Distributions of responding for the three intervals areshown in Figure 2 Rakitin et alrsquos (1998) young group isshown in the upper panel our aged group is in themiddle panel and the distributions for both groups nor-malized for time relative to the median time and propor-tions of peak responding are in the bottom panel Youngsubjectsrsquo peak estimates are very close to veridical withincreasing variability with increasing target durationsAged subjectsrsquo estimates are veridical for the 8-sec dura-tion close to veridical for the 12-sec duration and showa small underestimation (leftward shift) for the 21-secduration Normalized distributions in the bottom panelplotted in time relative to the median time show super-position across durations for both groups but with some-what larger variance at all values for the aged subjects
Means (standard deviation) of accuracy variability andthe coefcient of variation in both young and agedgroups are summarized in Table 1 The following effectswere found to be statistically reliable Accuracy Allsources of variance were signicant (Age F = 77 p lt005 Duration F = 9932 p lt 005 Age acute Duration F =41 p lt 005) Posthoc comparisons showed that theinteraction effect was due to a signicant leftward shiftof the aged group for the 21-sec duration (p lt 005)Variability A signicant effect of Duration (F = 808 p lt005) and an effect of Age that ldquoapproachedrdquo signicance(F = 37 p = 006) but no interaction effect was foundfor semi-interquartile range (SIQR) No signicant corre-lations were found between SIQRs and the neuropsy-chological scores Coefcient of Variation A signicanteffect of Age (F = 73 p lt 005) was found on SIQRme-dian No Duration or interaction effects were found Thusaged subjects have somewhat greater variability thanyoung ones which nevertheless remains scalar in theirtarget time estimates
2 ON versus OFF States of PD Patients
Distributions of responding for both ON and OFF condi-tions at both signal durations are shown in Figure 3 The
Malapani et al 319
Figure 2 Relative frequencydistributions for the three tar-get durations (8 12 and 21sec) showing accuracy andvariability of estimation inyoung college students (whiteupper panel) and in aged nor-mal controls (black middlepanel) The three functions ofthe two groups when plottedin time relative to the mediantime and normalized as pro-portions of the maximum rela-tive frequency superpose(lower panel)
Table 1 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation in Young and Aged Subjects inExperiment 1
8-sec duration 12-sec duration 21-sec duration
Young Aged Young Aged Young Aged
Median 805 (4) 805 (6) 123 (8) 117 (7) 214 (13) 197 (17)
SIQR 35 (2) 55 (3) 66 (3) 75 (3) 102 (4) 121 (9)
SIQRmedian 043 (02) 068 (04) 053 (02) 067 (03) 047 (02) 061 (05)
320 Journal of Cognitive Neuroscience Volume 10 Number 3
accuracy of temporal reproduction in PD patients whenthey are ON levodopa + apomorphine medication isveridical (open symbols upper panel) They producepeaks in responding that are very close to the targets forboth durations (mean peak times of 78 and 211 sec)The same subjects show a dramatic distortion in accu-racy for both durations when they are OFF their medica-tion (lled symbols upper panel) they overestimate theshort signal and underestimate the long one The variabil-ity of reproduction for 8 sec is also different betweenON and OFF states as can be seen in the different spread
of the curves obtained for the 8-sec target Howevervariability is roughly equal for the 21-sec target in boththe ON and OFF states because those curves are equallybroad The scalar property shown in the lower panelemerges in the ON state when responses are normalizedas a proportion of median time and scaled to maximumresponding The ON distributions show superposition Inthe OFF state however a violation of superposition isobserved The 8- and 21-sec OFF distributions do notsuperpose and they are broader in relative time than theON distributions especially for the 8-sec condition
Figure 3 Relative frequencydistributions for the two tar-get durations (8 and 21 sec)showing accuracy and variabil-ity of estimation in PD pa-tients ON (white) and OFF
(black) their levodopa medica-tion (upper panel) The twofunctions of the same subjectswhen plotted in time relativeto the median time and nor-malized as proportions of themaximum relative frequencysuperpose in the ON state butnot in the OFF state (lowerpanel)
Malapani et al 321
Means of accuracy variability and coefcient of vari-ation in the ON and OFF states of PD patients are summa-rized in Table 2 Statistical analysis showed the followingeffects Accuracy A signicant effect of Drug (F = 104p lt 005) and Duration (F = 5578 p lt 005) on themedians was found The interaction between these twosources of variance was also signicant (F = 516 p lt005) Further paired comparisons showed a signicantdifference between the medians obtained in the ON andOFF states for both 8 sec (F = 607 p lt 005) and 21-secdurations (F = 592 p lt 005) No correlations werefound between accuracy and neuropsychological or mo-tor scores in the ON state Correlations at a signicantlevel were found in the OFF state between the akinesiascore and (1) the rightward shift in accuracy for the8-sec duration (beta = 01 p lt 005) (2) the leftwardshift in accuracy for the 21-sec duration (beta = 005p lt 005) Variability A signicant effect of Duration (F= 731 p lt 005) and Drug (F = 212 p lt 005) wasfound for SIQR The interaction between the two factorswas not signicant (F = 23 p = 01) Further pairedcomparisons showed a signicant difference betweenthe SIQR obtained in the ON and OFF states for 8 sec (F= 10 16 p lt 005) but not for 21 sec (F = 099 p = 03)No correlations were found between SIQR and motor orneuropsychological scores in either drug state Coef-cient of variation A signicant effect of both Durationand Drug was found on SIQRmedian The interactionbetween the two factors was not signicant In order totest the scalar property further paired comparisons con-trasted Duration within Drug condition Coefcients ofvariation for the two durations were not signicantlydifferent in the ON condition (F = 24 p = 01) Howeverwe found a signicant difference between durations inthe OFF condition (F = 109 p lt 005)
3 Aged Controls versus ON amp OFF PD Patients
Statistical analysis between the three groups (Aged PDON PD OFF) showed the following effects Accuracy Nosignicant effect of Group on the median for the 8-sectarget was found Post hoc analysis showed a signicantdifference between the median obtained between PD ON
and PD OFF groups (p lt 005) but no signicant differ-ence between PD ON and Aged or between PD OFF and
Aged A signicant effect of Group on the median for the21-sec target was found (F = 17 9 p lt 005) Post hoccomparisons showed signicant differences between PDON and PD OFF (p lt 005) PD ON and Aged (p lt 005)and PD OFF and Aged (p lt 005) Variability A signicanteffect of Group (F = 84 p lt 005) was found on SIQRfor the 8-sec target Further post hoc comparisonsshowed a signicant difference between ON and OFF
groups (p lt 005) and between PD OFF and Aged (p lt005) but no signicant difference between PD ON andAged For the 21-sec target no signicant effect of Groupwas found and post hoc analysis showed no signicantdifference between any pair of the three groups Coef-cient of variation A signicant effect of Group wasfound on SIQRmedian for the 8-sec target (F = 67 p lt005) Further post hoc comparisons showed a sig-nicant difference between PD OFF and PD ON as wellas between PD OFF and Aged (p lt 005) However nosignicant difference was found between PD ON andAged For the 21-sec target no signicant effect of Groupwas found on SIQRmedian and the post hoc analysisshowed only a signicant effect between PD ON and PDOFF and no signicant differences between either PD ON
and Aged or PD OFF and Aged
Discussion
The comparisons between young and aged controlsshow that the scalar property for temporal informationprocessing holds with age The good superposition seenin Figure 2 is reected in the large effect of duration onSIQR and the lack of an effect of duration on SIQRme-dian However aged people showed an overall increasedvariability compared to young college students and aslight underestimation for the longer duration target Theformer effect along with the failure to nd a differencein the coefcient of variation (the scalar property) sug-gests that temporal processing is more variable in agedpeople but still conforms to Weberrsquos Law Thus as de-scribed earlier this increased variance could arise eitherfrom clock variability memory storage or retrieval vari-ability or criterion-decision variability
The small underestimation seen at 21 sec (and a stillsmaller underestimation at 12 sec) is probably due to adysfunction in mnemonic processing because accuracy
Table 2 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation for PD Patients ON and OFF Medicationin Experiment 1
8-sec duration 21-sec duration
PD ON PD OFF PD ON PD OFF
Median 76 (6) 89 (206) 211 (8) 173 (15)
SIQR 45 (2) 82 (5) 115 (5) 125 (4)
SIQRmedian 059 (03) 092 (03) 054 (02) 072 (03)
322 Journal of Cognitive Neuroscience Volume 10 Number 3
effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

Figure 2 Relative frequencydistributions for the three tar-get durations (8 12 and 21sec) showing accuracy andvariability of estimation inyoung college students (whiteupper panel) and in aged nor-mal controls (black middlepanel) The three functions ofthe two groups when plottedin time relative to the mediantime and normalized as pro-portions of the maximum rela-tive frequency superpose(lower panel)
Table 1 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation in Young and Aged Subjects inExperiment 1
8-sec duration 12-sec duration 21-sec duration
Young Aged Young Aged Young Aged
Median 805 (4) 805 (6) 123 (8) 117 (7) 214 (13) 197 (17)
SIQR 35 (2) 55 (3) 66 (3) 75 (3) 102 (4) 121 (9)
SIQRmedian 043 (02) 068 (04) 053 (02) 067 (03) 047 (02) 061 (05)
320 Journal of Cognitive Neuroscience Volume 10 Number 3
accuracy of temporal reproduction in PD patients whenthey are ON levodopa + apomorphine medication isveridical (open symbols upper panel) They producepeaks in responding that are very close to the targets forboth durations (mean peak times of 78 and 211 sec)The same subjects show a dramatic distortion in accu-racy for both durations when they are OFF their medica-tion (lled symbols upper panel) they overestimate theshort signal and underestimate the long one The variabil-ity of reproduction for 8 sec is also different betweenON and OFF states as can be seen in the different spread
of the curves obtained for the 8-sec target Howevervariability is roughly equal for the 21-sec target in boththe ON and OFF states because those curves are equallybroad The scalar property shown in the lower panelemerges in the ON state when responses are normalizedas a proportion of median time and scaled to maximumresponding The ON distributions show superposition Inthe OFF state however a violation of superposition isobserved The 8- and 21-sec OFF distributions do notsuperpose and they are broader in relative time than theON distributions especially for the 8-sec condition
Figure 3 Relative frequencydistributions for the two tar-get durations (8 and 21 sec)showing accuracy and variabil-ity of estimation in PD pa-tients ON (white) and OFF
(black) their levodopa medica-tion (upper panel) The twofunctions of the same subjectswhen plotted in time relativeto the median time and nor-malized as proportions of themaximum relative frequencysuperpose in the ON state butnot in the OFF state (lowerpanel)
Malapani et al 321
Means of accuracy variability and coefcient of vari-ation in the ON and OFF states of PD patients are summa-rized in Table 2 Statistical analysis showed the followingeffects Accuracy A signicant effect of Drug (F = 104p lt 005) and Duration (F = 5578 p lt 005) on themedians was found The interaction between these twosources of variance was also signicant (F = 516 p lt005) Further paired comparisons showed a signicantdifference between the medians obtained in the ON andOFF states for both 8 sec (F = 607 p lt 005) and 21-secdurations (F = 592 p lt 005) No correlations werefound between accuracy and neuropsychological or mo-tor scores in the ON state Correlations at a signicantlevel were found in the OFF state between the akinesiascore and (1) the rightward shift in accuracy for the8-sec duration (beta = 01 p lt 005) (2) the leftwardshift in accuracy for the 21-sec duration (beta = 005p lt 005) Variability A signicant effect of Duration (F= 731 p lt 005) and Drug (F = 212 p lt 005) wasfound for SIQR The interaction between the two factorswas not signicant (F = 23 p = 01) Further pairedcomparisons showed a signicant difference betweenthe SIQR obtained in the ON and OFF states for 8 sec (F= 10 16 p lt 005) but not for 21 sec (F = 099 p = 03)No correlations were found between SIQR and motor orneuropsychological scores in either drug state Coef-cient of variation A signicant effect of both Durationand Drug was found on SIQRmedian The interactionbetween the two factors was not signicant In order totest the scalar property further paired comparisons con-trasted Duration within Drug condition Coefcients ofvariation for the two durations were not signicantlydifferent in the ON condition (F = 24 p = 01) Howeverwe found a signicant difference between durations inthe OFF condition (F = 109 p lt 005)
3 Aged Controls versus ON amp OFF PD Patients
Statistical analysis between the three groups (Aged PDON PD OFF) showed the following effects Accuracy Nosignicant effect of Group on the median for the 8-sectarget was found Post hoc analysis showed a signicantdifference between the median obtained between PD ON
and PD OFF groups (p lt 005) but no signicant differ-ence between PD ON and Aged or between PD OFF and
Aged A signicant effect of Group on the median for the21-sec target was found (F = 17 9 p lt 005) Post hoccomparisons showed signicant differences between PDON and PD OFF (p lt 005) PD ON and Aged (p lt 005)and PD OFF and Aged (p lt 005) Variability A signicanteffect of Group (F = 84 p lt 005) was found on SIQRfor the 8-sec target Further post hoc comparisonsshowed a signicant difference between ON and OFF
groups (p lt 005) and between PD OFF and Aged (p lt005) but no signicant difference between PD ON andAged For the 21-sec target no signicant effect of Groupwas found and post hoc analysis showed no signicantdifference between any pair of the three groups Coef-cient of variation A signicant effect of Group wasfound on SIQRmedian for the 8-sec target (F = 67 p lt005) Further post hoc comparisons showed a sig-nicant difference between PD OFF and PD ON as wellas between PD OFF and Aged (p lt 005) However nosignicant difference was found between PD ON andAged For the 21-sec target no signicant effect of Groupwas found on SIQRmedian and the post hoc analysisshowed only a signicant effect between PD ON and PDOFF and no signicant differences between either PD ON
and Aged or PD OFF and Aged
Discussion
The comparisons between young and aged controlsshow that the scalar property for temporal informationprocessing holds with age The good superposition seenin Figure 2 is reected in the large effect of duration onSIQR and the lack of an effect of duration on SIQRme-dian However aged people showed an overall increasedvariability compared to young college students and aslight underestimation for the longer duration target Theformer effect along with the failure to nd a differencein the coefcient of variation (the scalar property) sug-gests that temporal processing is more variable in agedpeople but still conforms to Weberrsquos Law Thus as de-scribed earlier this increased variance could arise eitherfrom clock variability memory storage or retrieval vari-ability or criterion-decision variability
The small underestimation seen at 21 sec (and a stillsmaller underestimation at 12 sec) is probably due to adysfunction in mnemonic processing because accuracy
Table 2 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation for PD Patients ON and OFF Medicationin Experiment 1
8-sec duration 21-sec duration
PD ON PD OFF PD ON PD OFF
Median 76 (6) 89 (206) 211 (8) 173 (15)
SIQR 45 (2) 82 (5) 115 (5) 125 (4)
SIQRmedian 059 (03) 092 (03) 054 (02) 072 (03)
322 Journal of Cognitive Neuroscience Volume 10 Number 3
effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

accuracy of temporal reproduction in PD patients whenthey are ON levodopa + apomorphine medication isveridical (open symbols upper panel) They producepeaks in responding that are very close to the targets forboth durations (mean peak times of 78 and 211 sec)The same subjects show a dramatic distortion in accu-racy for both durations when they are OFF their medica-tion (lled symbols upper panel) they overestimate theshort signal and underestimate the long one The variabil-ity of reproduction for 8 sec is also different betweenON and OFF states as can be seen in the different spread
of the curves obtained for the 8-sec target Howevervariability is roughly equal for the 21-sec target in boththe ON and OFF states because those curves are equallybroad The scalar property shown in the lower panelemerges in the ON state when responses are normalizedas a proportion of median time and scaled to maximumresponding The ON distributions show superposition Inthe OFF state however a violation of superposition isobserved The 8- and 21-sec OFF distributions do notsuperpose and they are broader in relative time than theON distributions especially for the 8-sec condition
Figure 3 Relative frequencydistributions for the two tar-get durations (8 and 21 sec)showing accuracy and variabil-ity of estimation in PD pa-tients ON (white) and OFF
(black) their levodopa medica-tion (upper panel) The twofunctions of the same subjectswhen plotted in time relativeto the median time and nor-malized as proportions of themaximum relative frequencysuperpose in the ON state butnot in the OFF state (lowerpanel)
Malapani et al 321
Means of accuracy variability and coefcient of vari-ation in the ON and OFF states of PD patients are summa-rized in Table 2 Statistical analysis showed the followingeffects Accuracy A signicant effect of Drug (F = 104p lt 005) and Duration (F = 5578 p lt 005) on themedians was found The interaction between these twosources of variance was also signicant (F = 516 p lt005) Further paired comparisons showed a signicantdifference between the medians obtained in the ON andOFF states for both 8 sec (F = 607 p lt 005) and 21-secdurations (F = 592 p lt 005) No correlations werefound between accuracy and neuropsychological or mo-tor scores in the ON state Correlations at a signicantlevel were found in the OFF state between the akinesiascore and (1) the rightward shift in accuracy for the8-sec duration (beta = 01 p lt 005) (2) the leftwardshift in accuracy for the 21-sec duration (beta = 005p lt 005) Variability A signicant effect of Duration (F= 731 p lt 005) and Drug (F = 212 p lt 005) wasfound for SIQR The interaction between the two factorswas not signicant (F = 23 p = 01) Further pairedcomparisons showed a signicant difference betweenthe SIQR obtained in the ON and OFF states for 8 sec (F= 10 16 p lt 005) but not for 21 sec (F = 099 p = 03)No correlations were found between SIQR and motor orneuropsychological scores in either drug state Coef-cient of variation A signicant effect of both Durationand Drug was found on SIQRmedian The interactionbetween the two factors was not signicant In order totest the scalar property further paired comparisons con-trasted Duration within Drug condition Coefcients ofvariation for the two durations were not signicantlydifferent in the ON condition (F = 24 p = 01) Howeverwe found a signicant difference between durations inthe OFF condition (F = 109 p lt 005)
3 Aged Controls versus ON amp OFF PD Patients
Statistical analysis between the three groups (Aged PDON PD OFF) showed the following effects Accuracy Nosignicant effect of Group on the median for the 8-sectarget was found Post hoc analysis showed a signicantdifference between the median obtained between PD ON
and PD OFF groups (p lt 005) but no signicant differ-ence between PD ON and Aged or between PD OFF and
Aged A signicant effect of Group on the median for the21-sec target was found (F = 17 9 p lt 005) Post hoccomparisons showed signicant differences between PDON and PD OFF (p lt 005) PD ON and Aged (p lt 005)and PD OFF and Aged (p lt 005) Variability A signicanteffect of Group (F = 84 p lt 005) was found on SIQRfor the 8-sec target Further post hoc comparisonsshowed a signicant difference between ON and OFF
groups (p lt 005) and between PD OFF and Aged (p lt005) but no signicant difference between PD ON andAged For the 21-sec target no signicant effect of Groupwas found and post hoc analysis showed no signicantdifference between any pair of the three groups Coef-cient of variation A signicant effect of Group wasfound on SIQRmedian for the 8-sec target (F = 67 p lt005) Further post hoc comparisons showed a sig-nicant difference between PD OFF and PD ON as wellas between PD OFF and Aged (p lt 005) However nosignicant difference was found between PD ON andAged For the 21-sec target no signicant effect of Groupwas found on SIQRmedian and the post hoc analysisshowed only a signicant effect between PD ON and PDOFF and no signicant differences between either PD ON
and Aged or PD OFF and Aged
Discussion
The comparisons between young and aged controlsshow that the scalar property for temporal informationprocessing holds with age The good superposition seenin Figure 2 is reected in the large effect of duration onSIQR and the lack of an effect of duration on SIQRme-dian However aged people showed an overall increasedvariability compared to young college students and aslight underestimation for the longer duration target Theformer effect along with the failure to nd a differencein the coefcient of variation (the scalar property) sug-gests that temporal processing is more variable in agedpeople but still conforms to Weberrsquos Law Thus as de-scribed earlier this increased variance could arise eitherfrom clock variability memory storage or retrieval vari-ability or criterion-decision variability
The small underestimation seen at 21 sec (and a stillsmaller underestimation at 12 sec) is probably due to adysfunction in mnemonic processing because accuracy
Table 2 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation for PD Patients ON and OFF Medicationin Experiment 1
8-sec duration 21-sec duration
PD ON PD OFF PD ON PD OFF
Median 76 (6) 89 (206) 211 (8) 173 (15)
SIQR 45 (2) 82 (5) 115 (5) 125 (4)
SIQRmedian 059 (03) 092 (03) 054 (02) 072 (03)
322 Journal of Cognitive Neuroscience Volume 10 Number 3
effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

Means of accuracy variability and coefcient of vari-ation in the ON and OFF states of PD patients are summa-rized in Table 2 Statistical analysis showed the followingeffects Accuracy A signicant effect of Drug (F = 104p lt 005) and Duration (F = 5578 p lt 005) on themedians was found The interaction between these twosources of variance was also signicant (F = 516 p lt005) Further paired comparisons showed a signicantdifference between the medians obtained in the ON andOFF states for both 8 sec (F = 607 p lt 005) and 21-secdurations (F = 592 p lt 005) No correlations werefound between accuracy and neuropsychological or mo-tor scores in the ON state Correlations at a signicantlevel were found in the OFF state between the akinesiascore and (1) the rightward shift in accuracy for the8-sec duration (beta = 01 p lt 005) (2) the leftwardshift in accuracy for the 21-sec duration (beta = 005p lt 005) Variability A signicant effect of Duration (F= 731 p lt 005) and Drug (F = 212 p lt 005) wasfound for SIQR The interaction between the two factorswas not signicant (F = 23 p = 01) Further pairedcomparisons showed a signicant difference betweenthe SIQR obtained in the ON and OFF states for 8 sec (F= 10 16 p lt 005) but not for 21 sec (F = 099 p = 03)No correlations were found between SIQR and motor orneuropsychological scores in either drug state Coef-cient of variation A signicant effect of both Durationand Drug was found on SIQRmedian The interactionbetween the two factors was not signicant In order totest the scalar property further paired comparisons con-trasted Duration within Drug condition Coefcients ofvariation for the two durations were not signicantlydifferent in the ON condition (F = 24 p = 01) Howeverwe found a signicant difference between durations inthe OFF condition (F = 109 p lt 005)
3 Aged Controls versus ON amp OFF PD Patients
Statistical analysis between the three groups (Aged PDON PD OFF) showed the following effects Accuracy Nosignicant effect of Group on the median for the 8-sectarget was found Post hoc analysis showed a signicantdifference between the median obtained between PD ON
and PD OFF groups (p lt 005) but no signicant differ-ence between PD ON and Aged or between PD OFF and
Aged A signicant effect of Group on the median for the21-sec target was found (F = 17 9 p lt 005) Post hoccomparisons showed signicant differences between PDON and PD OFF (p lt 005) PD ON and Aged (p lt 005)and PD OFF and Aged (p lt 005) Variability A signicanteffect of Group (F = 84 p lt 005) was found on SIQRfor the 8-sec target Further post hoc comparisonsshowed a signicant difference between ON and OFF
groups (p lt 005) and between PD OFF and Aged (p lt005) but no signicant difference between PD ON andAged For the 21-sec target no signicant effect of Groupwas found and post hoc analysis showed no signicantdifference between any pair of the three groups Coef-cient of variation A signicant effect of Group wasfound on SIQRmedian for the 8-sec target (F = 67 p lt005) Further post hoc comparisons showed a sig-nicant difference between PD OFF and PD ON as wellas between PD OFF and Aged (p lt 005) However nosignicant difference was found between PD ON andAged For the 21-sec target no signicant effect of Groupwas found on SIQRmedian and the post hoc analysisshowed only a signicant effect between PD ON and PDOFF and no signicant differences between either PD ON
and Aged or PD OFF and Aged
Discussion
The comparisons between young and aged controlsshow that the scalar property for temporal informationprocessing holds with age The good superposition seenin Figure 2 is reected in the large effect of duration onSIQR and the lack of an effect of duration on SIQRme-dian However aged people showed an overall increasedvariability compared to young college students and aslight underestimation for the longer duration target Theformer effect along with the failure to nd a differencein the coefcient of variation (the scalar property) sug-gests that temporal processing is more variable in agedpeople but still conforms to Weberrsquos Law Thus as de-scribed earlier this increased variance could arise eitherfrom clock variability memory storage or retrieval vari-ability or criterion-decision variability
The small underestimation seen at 21 sec (and a stillsmaller underestimation at 12 sec) is probably due to adysfunction in mnemonic processing because accuracy
Table 2 Mean (Standard Deviation) of Accuracy Variability amp Coefcient of Variation for PD Patients ON and OFF Medicationin Experiment 1
8-sec duration 21-sec duration
PD ON PD OFF PD ON PD OFF
Median 76 (6) 89 (206) 211 (8) 173 (15)
SIQR 45 (2) 82 (5) 115 (5) 125 (4)
SIQRmedian 059 (03) 092 (03) 054 (02) 072 (03)
322 Journal of Cognitive Neuroscience Volume 10 Number 3
effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

effects are not expected from dysfunctions in the clocksystem However to the extent that aged people showsome loss of dopaminergic enervation in the striatum(Agid Javoy-Agid amp Ruberg 1987) and given the bodyof literature on clock effects induced by dopaminergicderegulation (see Meck 1996 for review) a clock dys-function may be also present that is small enough to notforce recalibration of the remembered subjective dura-tion with feedback during training and testing This ques-tion is readdressed below when large accuracydistortions in the performance of PD patients in the OFF
state rule out clock effectsThe results show that the estimation of time is normal
in PD patients in the ON medication state Their peaks inresponding are close to veridical for both durations likethe peaks produced by young normal control subjectsMoreover the temporal variability in the ON state in-creases proportionate to the target duration By conse-quence the scalar property holds in the ON state asshown by the absence of a difference between thenormalized functions for the two durations (Figure 3)These results show that temporal information process-ing is regulated by normative processes when thedopaminergic regulation in the brain is reestablished asit is when PD patients are treated with the levodopa
The same patients OFF their medication show dramati-cally impaired accuracy for both target durations Theyoverestimate the short signal and underestimate the longone compared to their own performance in the ON stateThis unexpected ldquomigrationrdquo effect the rightward shiftfor 8 sec and the leftward shift for 21 sec was correlatedwith the akinesia scores in the OFF state A positive anda negative correlation were found between the akinesiascore and the rightward and leftward shift respectivelysuggesting that dopamine depletion in the striatum pro-duces the migration effect in temporal accuracy A pri-mary conclusion from these results is that accurateestimation of time intervals depends upon dopaminergicregulation in the striatum
Where do these distortions lie We propose that themigration of the peaks toward each other reects amemory dysfunction such that the two targets appearmore alikemdashare ldquocoupledrdquo By coupling we mean a mu-tual attraction between the two time values when theyare laid down in memory or retrieved and compared toa current clock reading The migration effect was consis-tently seen over a considerable number of trials in theOFF state Even though subjects were receiving feedbackthey were uniformly surprised that they were ldquooff thetargetrdquo Corrective adjustment was evidently not possiblefor these subjects This is evidence that the migrationeffect is a memory storageretrieval problem rather thana clock problem A memory effect may be inferred whenan accuracy distortion is relatively permanent despiteintermittent corrective feedback during test sessions Asdescribed earlier a clock speed difference is not ex-pected to produce an accuracy change unless it is intro-
duced after training in an alternative drug condition Theclassic clock pattern distortion produced in animals un-der dopaminergic manipulation (Meck 1996) holds thatsubjects originally trained without a dopamine agonistshow a dramatic rapid shift toward underestimationwhen training is continued under the agonist manipula-tion However peak estimation gradually returns toveridical accuracy as new subjective times overlay pre-viously learned memories for duration without the drugThis in turn is followed by a rebound effect in theopposite direction when the drug is removed In ourstudy we analyzed rst halfsecond half performance inthe OFF condition and found no difference in accuracyearly and late in training This would be expected if theeffect was a memory problem such that even underfeedback subjects are unable to recalibrate because theirmemories continue to be laid down or are retrievedldquoincorrectlyrdquo The mismatch between trained values andremembered values persists at least through the blockedtraining (60 trials) for each duration
The migration however is demonstrably not a mixingof retrieval from two different memories Such mixingor switching decit is well documented in PD (Brownamp Marsden 1991 Malapani Pillon et al 1994) even onlevodopa medication (Owen et al 1993) Mixing shouldinduce relative frequency distributions of temporal esti-mates with two peaks one at each criterion time insteadof one peak shifted to the right or to the left as seenhere A careful examination of individual as well as groupfunctions showed no instances of a minor peak at theother criterion time Rather individual as well as groupfunctions show unimodal peaks at a distorted that ismigrated temporal target
In addition to the coupling seen in accuracy whensubjects are OFF the drug there is increased variabilityas well that does not follow the scalar rule either be-tween ON and OFF states or importantly within the OFF
state In the 8-sec condition if the subjectsrsquo dysfunctionin the OFF state was one of accuracy only while thetiming mechanism operated ldquonormallyrdquo as in the ON statebut aimed at a distorted target the OFF state functionwould superpose with the ON state distribution whichit does not do Even controlling for the distorted increasein reproduced time the coefcient of variation for 8 secOFF is larger than that found in the ON condition Thusadded variability is introduced in the 8-sec OFF conditionSimilarly added variability is introduced in the 21-seccondition in the OFF state compared to the ON statebecause the coefcients of variation are signicantlydifferent between the two states Importantly this in-creased variability is not scalar with the distorted accu-racy values OFF the drug The coefcient of variation for8 sec is signicantly greater than that for 21 sec Thusthe lack of dopamine regulation adds nonscalar variabil-ity in timing just as it adds variance in many systems inPD which is well documented in other clinical phenom-ena (Hallett 1993)
Malapani et al 323
In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

In conclusion dopaminergic deregulation in thestriatum produces both accuracy and variance effects ininterval timing when two different time values havebeen learned
EXPERIMENT 2
The second experiment was designed to eliminate cou-pling by studying a condition in which only one durationmust be remembered and retrieved If the distortionsseen in Experiment 1 are a result of prior training on asecond duration that somehow forces migration andnonscalar variance with only one duration in memorywe should see no such distortions It will therefore beof special interest to look for a higher coefcient ofvariation OFF the drug in the single duration conditionWe chose the 21-sec duration to study in Experiment 2because the left shift for 21 sec in Experiment 1 was insome sense more surprising than the right shift seen atthe short duration Slowness of movement and general-ized slowing in PD suggests that perhaps slowed storageor retrieval might produce an overestimation of timeintervals As in Experiment 1 a new group of PD patientsin Experiment 2 were trained and tested both ON andOFF medication
Results
The results obtained in Experiment 2 are shown con-trasted with the results obtained in Experiment 1 for the21-sec duration in Figure 4 This allows a direct compari-son of responding when 21 sec is the only durationtrained versus 21 sec as one of two durations trainedThe relative frequency distributions (upper panel) showthat in both Experiments 1 and 2 subjects were fairlyaccurate at timing the 21-sec duration in the ON condi-tion A shift in accuracy occurs in the OFF state of PDpatients in both experiments compared to their ownperformance in the ON state However the OFF statefunctions are slightly shifted to the right in Experiment2 whereas they are dramatically shifted to the left inExperiment 1 Importantly the 21-sec functions obtainedin the ON and the OFF state for the patients in Experi-ment 2 superpose when plotted in relative time andnormalized as proportions of the maximum (lowerpanel)
Mean median SIQR and SIQRmedian in both ON andOFF states obtained in Experiment 2 are shown in Table3 Statistical analysis of the results showed a signicant(right shift) effect of drug on the median (F = 69 p lt005) with no signicant effect on SIQR (F = 001 p =09) and no effect on SIQRmedian (F = 01 p = 07)
Discussion
Experiment 2 in which only one duration was repro-duced was designed to control for biases and interac-
tions that may occur when more than one duration hasbeen learned Results showed a small but reliable over-estimation of the 21-sec duration in PD patients OFF
medication when it is the only duration to be remem-bered and reproduced In contrast the same duration isunderestimated in Experiment 1 when it is associatedwith a memory for a shorter duration This is evidencethat the underestimation of the long signal obtained inExperiment 1 was due to the dual training requirementIndeed Experiment 2 suggests this left migration mayhave masked a right shift that would have been obtainedin PD patients OFF the drug if the second short intervalhad not been trained as well Again here as in Experi-ment 1 an analysis of rst halfsecond half performancein the OFF condition showed no difference in accuracyearly and late in training Thus as in Experiment 1 theclassic ldquoclock patternrdquo obtained in animals after adminis-tration of dopaminergic antagonists (Meck 1996) also isnot seen in this experiment Rather the relatively perma-nent although small right shift is maintained under con-tinued feedback consistent with a memory distortionThe nonscalar variance evident in Experiment 1 whentwo durations were trained is not paralleled by an in-crease in coefcient variation OFF the drug in Experi-ment 2 Rather variance OFF the drug is scalar withvariance ON the drug as indexed by no signicant changein SIQRmedian Together with the right shift in accuracyit strongly suggests that the temporal processing distor-tions in Experiment 1 were the result of the dual mem-ory feature for both the migration and the addedvariance seen there
GENERAL DISCUSSION
The experiments reported here have both behavioraland functional anatomical implications
Behavioral Implications
The rst conclusion that can be drawn from the presentdata is that normal clock functioning reproduction ofaccurate time estimation exhibiting the scalar propertyis obtained whenever dopamine regulation is normalYoung subjects to a lesser extent aged subjects and PDpatients ON levodopa supplementation treatment allshowed accurate time estimation Our interest centerson what components of the timing process are impairedwhen dysfunctional temporal reproduction is found Twodiffering dysfunctions are observed in accuracy and vari-ability of temporal reproduction in PD patients OFF medi-cation
When patients are required by the task to remembertwo different intervals memory for an earlier learnedinterval appears to affect production of a later learnedinterval by causing a migration of criterion times towardeach other The preponderance of evidence suggests thatthis effect is a memory rather than a clock or decision
324 Journal of Cognitive Neuroscience Volume 10 Number 3
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331
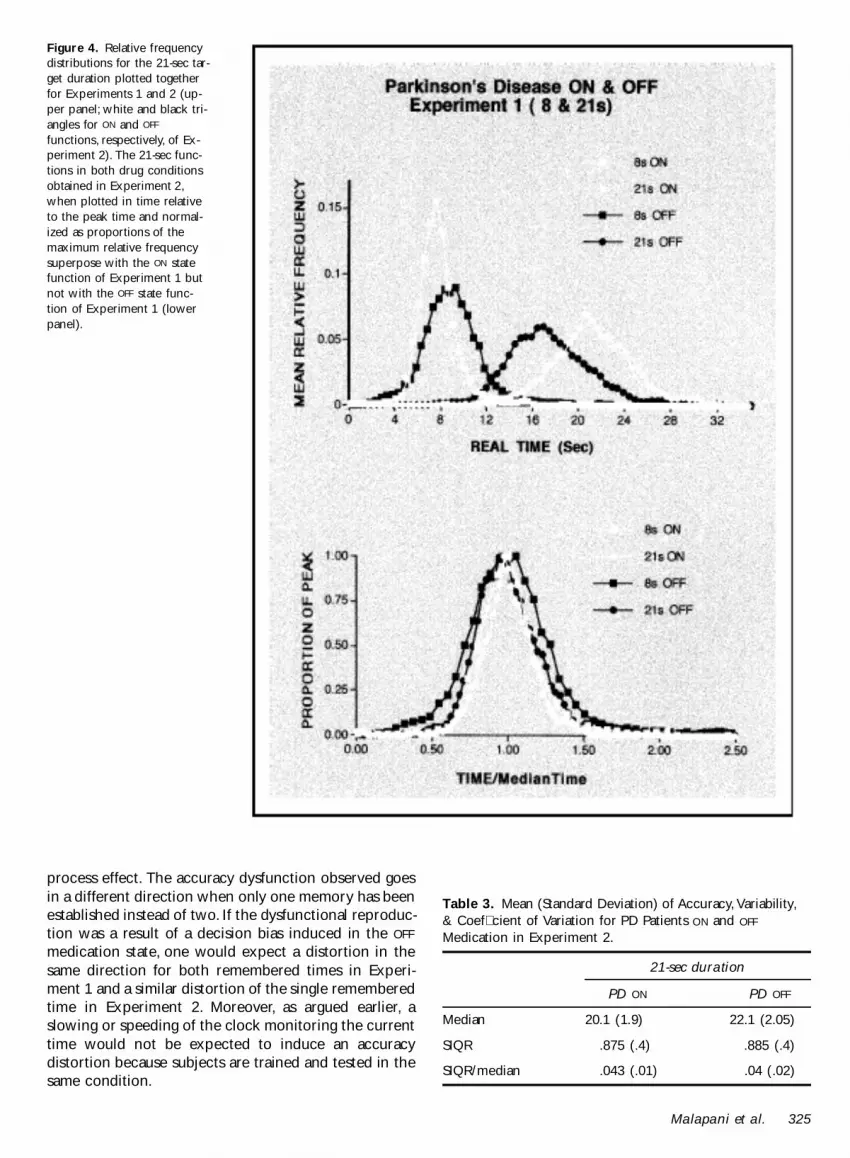
process effect The accuracy dysfunction observed goesin a different direction when only one memory has beenestablished instead of two If the dysfunctional reproduc-tion was a result of a decision bias induced in the OFF
medication state one would expect a distortion in thesame direction for both remembered times in Experi-ment 1 and a similar distortion of the single rememberedtime in Experiment 2 Moreover as argued earlier aslowing or speeding of the clock monitoring the currenttime would not be expected to induce an accuracydistortion because subjects are trained and tested in thesame condition
Figure 4 Relative frequencydistributions for the 21-sec tar-get duration plotted togetherfor Experiments 1 and 2 (up-per panel white and black tri-angles for ON and OFF
functions respectively of Ex-periment 2) The 21-sec func-tions in both drug conditionsobtained in Experiment 2when plotted in time relativeto the peak time and normal-ized as proportions of themaximum relative frequencysuperpose with the ON statefunction of Experiment 1 butnot with the OFF state func-tion of Experiment 1 (lowerpanel)
Table 3 Mean (Standard Deviation) of Accuracy Variabilityamp Coefcient of Variation for PD Patients ON and OFF
Medication in Experiment 2
21-sec duration
PD ON PD OFF
Median 201 (19) 221 (205)
SIQR 875 (4) 885 (4)
SIQRmedian 043 (01) 04 (02)
Malapani et al 325
On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

On these grounds we would ascribe the migration tomnemonic processing There is also internal evidencesupporting a memory rather than a clock or decisiondysfunction here In Experiment 2 subjects showed nor-mal scalar variance for the 21-sec target time even whena small distortion in the reproduced target time waspresent When subjects estimated 21 sec ON medicationthey reproduced nearly veridical estimates When OFF
medication they showed an increase in their estimatebut this increase was associated with scalar varianceThis suggests that variability in the clock process under-lying reproductions of both the veridical and distortedtimes had not changed In Experiment 1 we found twodifferent kinds of distortion when OFF the drug First thetarget times themselves migrated but second variancearound these new (incorrect) target values did not showthe scalar property Importantly this is the case for the8-sec duration in which the distortion was in the samedirection as the 21-sec-only condition in Experiment 2where variance increases according to the scalar ruleThus the greater SIQR for 8 sec OFF in Experiment 1was not due solely to the scalar increase expected onthe basis of a new longer target time Rather addi-tional variance was induced by the dual training condi-tion
There are then two effects induced by dopaminergicderegulation One of these is distortion in the target timeto be reproduced which goes in opposite directionswhen one versus two memories are present The secondis an additional source of nonscalar variability which alsois present only when two memories are resident inlong-term store When only one memory must be re-membered and reproduced this source of variability isnot present Rather the scalar interval timer operatesnormally but on a distorted (in this case overestimated)value as in the 21-sec-only condition in Experiment 2
We hypothesize that either the encode or the decodesystem or both are responsible for distortions in thetimed values to be reproduced Although the memorysystem appears to be implicated in these accuracy andvariance effects clock distortions may be present as wellwhich are not observable in these conditions becausetraining and testing occurs under the same conditionsAdditional study aimed at dissociating these two kindsof effects when trained single- or dual-target values aretested in the alternative drug state without additionalreinforced training is currently in process in our labora-tories
Functional Anatomical Implications
Our experiments indicate that temporal informationprocessing in humans depends on dopaminergic regula-tion in the striatum Previous work with animals hasimplicated dopaminergic transmission systems in thestriatum as being required for any timing performance
at all 6-OHDA lesions in both the substantial nigra andthe striatum dramatically impair interval timing assessedwith the PI procedure in rats (Meck 1998a) whereasspatial discrimination is spared (Robbins Giardini JonesReading amp Sahakian 1990) Importantly interval timingis corrected after levodopa administration in animalswith lesions in the substantial nigra but no such correc-tion occurs when animals are lesioned in the striatum(Meck 1998a) This suggests that dopaminergic modula-tion of striatal circuits is necessary in order for timingfunctions to be processed through the striatum
The dopamine-containing nigrostriatal tract degener-ates in Parkinsonrsquos disease (Agid et al 1987) and theresulting dysfunction of striatal circuits leads to distur-bances of movement and cognition including the disor-dered timing of movement clinically known asbradykinesia andor akinesia (Hallett 1993 Marsden1992 Marsden amp Obeso 1994 Saint-Cyr Taylor amp Nichol-son 1995) The results obtained in this study suggest thatmore specic timing processes in particular storage andretrieval of the memory for time also involve the striatalcircuits Moreover the decit in time reproduction foundin the OFF condition is correlated with the akinesia scoresuggesting that the same dysfunctional timing mecha-nisms may underlie disordered timing of complex move-ments in PD
Recent neurophysiological evidence obtained in be-having animals (Graybiel Aosaki Flaherty amp Kimura1994) suggests that difculties in timing complex move-ments and even cognitive and psychomotor behaviorscould result from reduced temporal coordination of ac-tivity in the modular distributed circuits within thestriatum Coordination of the input-output circuitrythrough the striatum seems to rely on the synchronizedpause of activity in striatal interneurons (TANs) that arespatially distributed throughout the ldquomotorrdquo and ldquocogni-tiverdquo striatum temporally coordinated and predictive ofreward These neurons require tonic dopaminergic inputin order to express the synchronized pause because afterunilateral dopaminergic lesions induced by MPTP injec-tions in primates responses are lost on the side of theinfusion and moreover systemic injections of apomor-phine reinstate the responses (Aosaki Graybiel ampKimura 1994)
A plausible explanation for the results obtained in thisstudy is that the depletion of dopamine in PD OFF pa-tients and the resulting reduced temporal coordinationof activity between striatal modules and striato-corticalcircuits impairs the storage and retrieval processes oftemporal memory under the dual-task condition as wellas possibly impairing clock functioning Whether a dis-tinct role for temporal memory may be attributed to thedopamine-containing nigrostriatal tract or the striatal in-terneurons cannot be answered yet Our current pro-gram of work assessing patients with focal anddegenerative lesions in different parts of the striatal cir-
326 Journal of Cognitive Neuroscience Volume 10 Number 3
cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

cuitry (preliminary results reported by Malapani Rakitinet al 1994) should provide more evidence on this ques-tion
METHODS
General Procedure
The general procedure is diagrammed in Figure 5 Inboth experiments subjects were required to rememberand reproduce time intervals of a standard duration inblocked sessions with one duration presented in eachsession block A block of trials on a given durationconsisted of (1) the training phase of 10 xed time (FT)trials of the appropriate signal duration to allow subjectsto learn the duration to be tested later followed by 10peak interval (PI) trials (described below) with intertrial(ITI) feedback to allow subjects to become familiar withthe response requirements of the task (2) the testingphase of 60 trials including 75 PI trials and 25 FTtrials presented randomly Two-thirds of PI trials werefollowed by ITI feedback For one-third of these trials nofeedback occurred and the subject was instructed tosimply continue to the next trial Sessions generallylasted 15 to 2 h
During FT trials a blue rectangle appeared on thecomputer screen and subjects were instructed to attendto the period of time that the rectangle remained blueAt the end of this interval the rectangle changed colorfrom blue to magenta On PI trials the same blue rectan-gle appeared on the screen signaling the onset of the
interval to be timed but instead of changing color at thecriterion time it remained on the screen for a durationof up to 3 times the criterion time
Subjects were instructed to try to estimate when theythought the criterion time had elapsed by pressing thespace bar on the keyboard (indicated by inverted blacktriangles in the left panel of Figure 5) They were told tomake multiple guesses and that they should begin re-sponding before they thought the criterion time hadelapsed and continue at a high rate until they felt thecriterion time had passed after which they could termi-nate the trial and continue to the next by pressing theEnter key To prevent counting during this phase digitswere imposed over the target stimulus in a randomizedmanner and subjects were instructed to read this distrac-tor aloud (cf Rakitin et al 1998) Feedback was pre-sented during the ITI in the form of a histogram (Figure5 right panel) showing the distribution of the responsesthey made on the previous trial plotted on a relative timescale (tenths of the target interval) so that subjectsobtained no information about the absolute value of thecriterion duration They received two kinds of informa-tion from this gure First the spread of the histogramrevealed how stringent their response criterion wasSecond the location of the peak of the histogram indi-cated accuracy If most of their responding was clusteredto the left of the criterion time (ldquoTrdquo on the abscissa) theywere responding too early if their responses clusteredto the right of ldquoTrdquo this indicated that they were respond-ing too late
Figure 5 Diagram of the peak interval (PI) procedure A blue rectangle appears on the screen at the beginning of a trial (top row) On xedtime (FT trials) the rectangle turns magenta at the criterion time either 8 12 or 21 sec On PI trials (second and third rows) the blue rectan-gle stays on the screen and subjects respond (triangles) just in advance and just after their estimate of the criterion time On ITI feedbacktrials a histogram appears on the screen indicating their proportion of responses in time around T (inset at right)
Malapani et al 327
Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

Data Acquisition and Analysis
In all three experiments data were collected in 05-sectime bins so as to permit assessment of the scalar prop-erty and to measure accuracy and variability of timereproduction Peak functions for individual subjectrsquosblocks were created by collapsing responding from alltrials into a single frequency distribution Group distri-butions were obtained as follows Each individualrsquos timeestimate distribution was pooled within conditions andthen the median of the pooled distribution was alignedwith the group mean median for that condition andaveraged across subjects This method for representingthe spread of individual subjectrsquos time estimate distribu-tion controls for accuracy variation across subjects Thatis the SIQRs from the group distribution are a fair rep-resentation of the mean SIQR from individuals
Experiment 1
Subjects
Twelve aged control subjects with no neurological dis-ease or intellectual impairment and ten patients withidiopathic PD and no clinical evidence of dementia par-ticipated in the study Tables 4 and 5 show the generalcharacteristics and the neuropsychological scores of theaged group and the PD group respectively Results areexpressed as means (standard deviation)
The diagnosis of PD was based on the existence of anakineto-rigid syndrome with or without resting tremorand the absence of neuroleptic treatment focal signson clinical examination andor CT-NMR scans or symp-toms suggesting Progressive Supranuclear Palsy or Multi-ple System Atrophy Patients with a Mini Mental StateExamination score below 27 and a Montgomery andAsberg Depression Rating score greater than 18 wereexcluded None of the patients had undergone thalamo-tomy or were taking anticholinergic drugs Patients wereassessed twice and were divided into two groups accord-ing to their dopamine supplementation treatment at thetime of the study allowing a direct evaluation of theinuence of dopa transmission on their performance ON
PD patients (n = 10) with severe uctuations underlevodopa treatment [Hoehn and Yahr stages = II (n = 3)III (n = 4) IV (n = 3)] were assessed 90 min after acuteadministration of a supraliminar dose of levodopa-carbi-dopa (250 mg) and just after an injection of 15 ml ofapomorphine (when the effect of levodopa was maxi-mal) OFF PD patients (n = 10) were assessed whendopamine supplementation treatment had been with-drawn for at least 18 h (when the parkinsonian disabilitywas maximal)
Design
Stimuli and trial types used were those described in thegeneral experimental procedure Aged subjects were
tested for two sessions on three durations (8 12 and 21sec) The design of the study counterbalanced the orderof testing as follows (8-12-21) (8-21-12) (12-8-21) (12-21-8) (21-8-12) (21-12-8) Subjects received a differentorder of intervals in each of their two sessions PDsubjects had two sessions per day on each drug condi-tion for four consecutive days During each sessionreproduction of two durations (8 and 21 sec) was as-sessed in blocked minisessions The design of the study(Table 6) was counterbalanced for the drug conditionand the training order for the duration Half of thepatients were tested in an OFF -ON-ON-OFF design and theother half in a ON-OFF-OFF -ON design During each session
Table 4 Characteristics of Aged Subjects
n 12
Age (years) 722 (48)
Education (years) 117 (22)
Sex (MF) 84
WRIQ 1067 (173)
DSPANfor 55 (1)
DSPANbac 48 (14)
CVLT15T 545 (84)
Table 5 General Characteristics Motor amp Neuro-psychological Evaluation for PD Patients ON and OFF
Medication in Experiment 1
n 10
SEX (mf) 91
Age (years) 55 (8)
Education (years) 98 (24)
Disease duration (years) 9 (8)
MMS 291 (12)
MADRS 95 (74)
WCST (cr) 54 (29)
ON OFF
UPDRS 20 (10) 477 (145)
Tremor 03 (06) 34 (32)
Akinesia 93 (56) 206 (73)
Rigidity 50 (31) 106 (41)
Levodopa dose 250 mg
Apomorphine dose 4 ml
328 Journal of Cognitive Neuroscience Volume 10 Number 3
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331
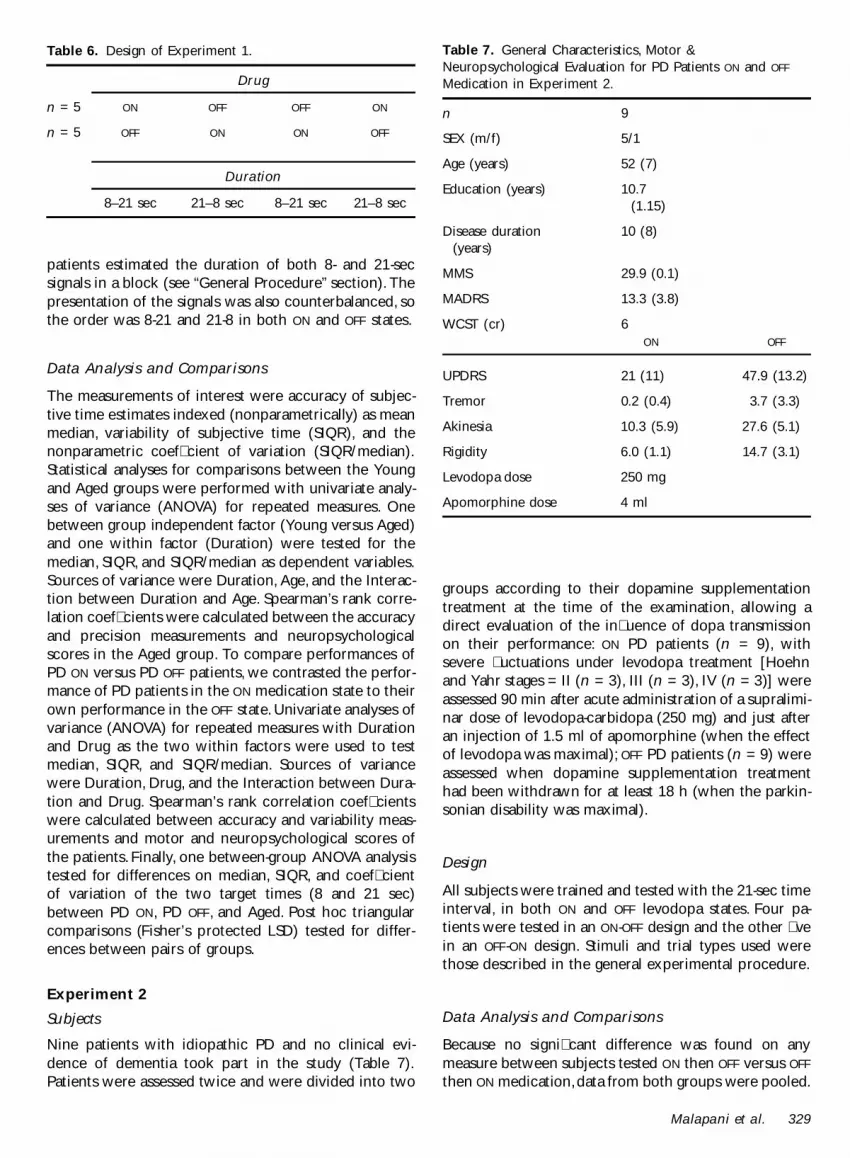
patients estimated the duration of both 8- and 21-secsignals in a block (see ldquoGeneral Procedurerdquo section) Thepresentation of the signals was also counterbalanced sothe order was 8-21 and 21-8 in both ON and OFF states
Data Analysis and Comparisons
The measurements of interest were accuracy of subjec-tive time estimates indexed (nonparametrically) as meanmedian variability of subjective time (SIQR) and thenonparametric coefcient of variation (SIQRmedian)Statistical analyses for comparisons between the Youngand Aged groups were performed with univariate analy-ses of variance (ANOVA) for repeated measures Onebetween group independent factor (Young versus Aged)and one within factor (Duration) were tested for themedian SIQR and SIQRmedian as dependent variablesSources of variance were Duration Age and the Interac-tion between Duration and Age Spearmanrsquos rank corre-lation coefcients were calculated between the accuracyand precision measurements and neuropsychologicalscores in the Aged group To compare performances ofPD ON versus PD OFF patients we contrasted the perfor-mance of PD patients in the ON medication state to theirown performance in the OFF state Univariate analyses ofvariance (ANOVA) for repeated measures with Durationand Drug as the two within factors were used to testmedian SIQR and SIQRmedian Sources of variancewere Duration Drug and the Interaction between Dura-tion and Drug Spearmanrsquos rank correlation coefcientswere calculated between accuracy and variability meas-urements and motor and neuropsychological scores ofthe patients Finally one between-group ANOVA analysistested for differences on median SIQR and coefcientof variation of the two target times (8 and 21 sec)between PD ON PD OFF and Aged Post hoc triangularcomparisons (Fisherrsquos protected LSD) tested for differ-ences between pairs of groups
Experiment 2
Subjects
Nine patients with idiopathic PD and no clinical evi-dence of dementia took part in the study (Table 7)Patients were assessed twice and were divided into two
groups according to their dopamine supplementationtreatment at the time of the examination allowing adirect evaluation of the inuence of dopa transmissionon their performance ON PD patients (n = 9) withsevere uctuations under levodopa treatment [Hoehnand Yahr stages = II (n = 3) III (n = 3) IV (n = 3)] wereassessed 90 min after acute administration of a supralimi-nar dose of levodopa-carbidopa (250 mg) and just afteran injection of 15 ml of apomorphine (when the effectof levodopa was maximal) OFF PD patients (n = 9) wereassessed when dopamine supplementation treatmenthad been withdrawn for at least 18 h (when the parkin-sonian disability was maximal)
Design
All subjects were trained and tested with the 21-sec timeinterval in both ON and OFF levodopa states Four pa-tients were tested in an ON-OFF design and the other vein an OFF -ON design Stimuli and trial types used werethose described in the general experimental procedure
Data Analysis and Comparisons
Because no signicant difference was found on anymeasure between subjects tested ON then OFF versus OFF
then ON medication data from both groups were pooled
Table 6 Design of Experiment 1
Drug
n = 5 ON OFF OFF ON
n = 5 OFF ON ON OFF
Duration
8ndash21 sec 21ndash8 sec 8ndash21 sec 21ndash8 sec
Table 7 General Characteristics Motor ampNeuropsychological Evaluation for PD Patients ON and OFF
Medication in Experiment 2
n 9
SEX (mf) 51
Age (years) 52 (7)
Education (years) 107(115)
Disease duration(years)
10 (8)
MMS 299 (01)
MADRS 133 (38)
WCST (cr) 6ON OFF
UPDRS 21 (11) 479 (132)
Tremor 02 (04) 37 (33)
Akinesia 103 (59) 276 (51)
Rigidity 60 (11) 147 (31)
Levodopa dose 250 mg
Apomorphine dose 4 ml
Malapani et al 329
Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

Data were analyzed similarly to Experiment 1 We usedpaired t-test comparisons to contrast the performance ofPD patients in the ON state on median SIQR andSIQRmedian of reproduced time measures with theirown performance in the OFF state
Acknowledgments
This research was supported by a HFSPO short-term grant toChara Malapani and NIH Grant MH54793 We thank the staff ofthe clinic Federation de Neurologie and Inserm U289 for assis-tance with this study
Reprint requests should be sent to Chara Malapani Dept ofPsychology Schermerhorn Hall Columbia University New YorkNY 10027 or via e-mail charapsychcolumbiaedu
REFERENCES
Agid Y Javoy-Agid F amp Ruberg M (1987) Biochemistry ofneurotransmitters in Parkinsonrsquos disease In C D Marsdenamp S Fahn (Eds) Movement disorders vol 2 (pp 166ndash230) London Butterworth
Allan L amp Gibbon J (1991) Human bisection at the geomet-ric mean Learning and Motivation 22 39ndash58
Aosaki T Graybiel A M amp Kimura M (1994) Changes in re-sponsiveness of striatal neurons to sensory stimuli havealso been shown after systematic treatment with MPTP Sci-ence 265 412
Artieda J Pastor M A Lacruz F amp Obeso J A (1992) Tem-poral discrimination is abnormal in Parkinsonrsquos diseaseBrain 115 199ndash210
Benecke R Rothwell J C Dick J P R Day B L amp Mars-den C D (1986) Performance of simultaneous move-ments in patients with Parkinsonrsquos disease Brain 109739ndash757
Bloxham C A Dick D J amp Moore M (1987) Reactiontimes and attention in Parkinsonrsquos disease Journal of Neu-rology Neurosurgery amp Psychiatry 50 1178ndash1183
Brown R G amp Marsden C D (1991) Dual task perfor-mance and processing resources in normal subjects andpatients with Parkinsonrsquos disease Brain 114 215ndash231
Catania A C (1970) Reinforcement schedules and psycho-physical judgments A study of some temporal propertiesof behavior In W N Schoenfeld (Ed) The theory of rein-forcement schedules (pp 1ndash42) New York Appleton-Cen-tury-Crofts
Church R M (1984) Properties of the internal clock InJ Gibbon amp L Allan (Eds) Timing and time perception(pp 566ndash582) New York New York Academy of Sciences
Evarts EV Teravainen H amp Calne D B (1981) Reactiontime in Parkinsonrsquos disease Brain 104 167ndash186
Freeman J S (1996) Abnormalities of motor timing in Hunt-ingtonrsquos disease Parkinsonism amp Related Disorders 281ndash93
Getty D J (1975) Discrimination of short temporal intervalsA comparison of two models Perception and Psycho-physics 18 1ndash8
Gibbon J (1977) Scalar expectancy theory and Weberrsquos lawin animal timing Psychological Review 84 279ndash325
Gibbon J (1992) Ubiquity of scalar timing with a Poissonclock Journal of Mathematical Psychology 36 283ndash293
Gibbon J Church R M amp Meck W H (1984) Scalar timingin memory In J Gibbon amp L G Allan (Eds) Timing and
time perception (pp 52ndash77) New York New York Acad-emy of Sciences
Gibbon J Malapani C Dale C L amp Gallistel C R (1997)Toward a neurobiology of temporal cognition Advancesand challenges Current Opinion in Neurobiology 7 170ndash184
Graybiel A M Aosaki T Flaherty A W amp Kimura M (1994)The basal ganglia and adaptive motor control Science265 1826ndash1831
Hallett M (1993) Physiology of basal ganglia disorders Anoverview Canadian Journal of Neurological Sciences20 177ndash183
Hallett M amp Khoshbin S (1980) A physiological mecha-nism of bradykinesia Brain 103 301ndash314
Keele S Pokorny R Corcos D amp Ivry R B (1985) Do per-ception and motor production share common timingmechanisms A correlational analysis Acta Psychologica60 173ndash191
Ivry R B (1996) The representation of temporal informationin perception and motor control Current Opinion in Neu-robiology 6 851ndash857
Ivry R B amp Hazeltine R E (1995) Perception and produc-tion of temporal intervals across a range of durations Evi-dence for a common timing mechanism Journal ofExperimental Psychology Human Perception and Perfor-mance 21 3ndash18
Ivry R B amp Keele S W (1989) Timing functions of the cere-bellum Journal of Cognitive Neuroscience 1 137ndash152
Ivry R B Keele S W amp Diener H C (1988) Dissociationof the lateral and medial cerebellum in movement timingand movement execution Experimental Brain Research73 167ndash180
Lieberman P Kako E Friedman J Tajchman G FeldmanL S amp Jiminez E B (1992) Speech production syntaxcomprehension and cognitive decits in Parkinsonrsquos dis-ease Brain amp Language 43 169ndash89
Malapani C Pillon B Dubois B amp Agid Y (1994) Impairedsimultaneous cognitive task performance in Parkinsonrsquos dis-ease A dopamine related dysfunction Neurology 44 319ndash326
Malapani C Rakitin B Deweer B Meck W Gibbon J Pil-lon B Defontaines B Dubois B amp Agid Y (1994) Segre-gation of timing processes within the basal gangliaInternational Neuropsychological Society Abstracts An-gers June 22ndash25 1994
Marsden C D (1992) Parkinsonrsquos disease PostgraduateMedical Journal 68 538ndash543
Marsden C D amp Obeso J A (1994) The functions of the ba-sal ganglia and the paradox of stereotaxic surgery in Park-insonrsquos disease Brain 118 877ndash897
Meck W H (1983) Selective adjustment of the speed of theinternal clock and memory storage processes Journal ofExperimental Psychology Animal Behavior Processes 9171ndash201
Meck W H (1986) Afnity of the dopamine D2 receptor pre-dicts neuroleptic potency in decreasing the speed of theinternal clock Pharmacology Biochemistry and Behav-ior 25 1185ndash1189
Meck W H (1996) Neuropharmacology of timing and timeperception Cognitive Brain Research 3 227ndash242
Meck W H (1998a) Neuroanatomical localization of an inter-nal clock A functional link between mesocorticalmesolombic and nigrostriatal dopaminergic systems Be-havioral Brain Research (in press)
Meck W H (1998b) Frontal cortex or nucleus basalis magno-cellularis lesions but not hippocampal or medial septalarea lesions occasion the loss of control of the speed ofan internal clock Experimental Brain Research (in press)
330 Journal of Cognitive Neuroscience Volume 10 Number 3
Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331

Meck W H Church R M amp Olton D S (1984) Hippocam-pus time and memory Behavioral Neuroscience 98 3ndash22
Nakamura R Nagasaki H amp Narabayashi H (1978) Distur-bances of rhythm formation in patients with Parkinsonrsquosdisease Part 1 Characteristics of tapping response to theperiod signals Perceptual and Motor Skills 46 63ndash75
Nichelli P Alway D amp Grafman J (1996) Perceptual timingin cerebellar degeneration Neuropsychologia 34 863ndash872
OrsquoBoyle D J Freeman J S amp Cody F W J (1996) The accu-racy and precision of timing of self-paced repetition move-ments in subjects with Parkinsonrsquos disease Brain 11951ndash70
Olton D S Wenk G L Church R M amp Meck W H (1987)Attention and the frontal cortex as examined by simultane-ous temporal processing Neuropsychologia 26 307ndash318
Owen A M Roberts A C Hodges J R Summers B APolkey C E amp Robbins T W (1993) Contrasting mecha-nisms of impaired attentional set-shifting in patients withfrontal lobe damage or Parkinsonrsquos disease Brain 1161159ndash1175
Pastor M A Artieda J Jahanshahi M amp Obeso J A (1992)Time estimation and reproduction is abnormal in Parkin-sonrsquos disease Brain 115 211ndash225
Rakitin B C Gibbon J Penney T B Malapani C HintonS C amp Meck W H (1998) Scalar expectancy theory and
peak-interval timing in humans Journal of ExperimentalPsychology Animal Behavior Processes 24 1ndash19
Robbins T W Giardini V Jones G H Reading P ampSahakian B J (1990) Effects of dopamine depletion fromthe caudate-putamen and nucleus accumbens septi on theacquisition and performance of a conditional discrimina-tion task Behavioral Brain Research 38 243ndash261
Roberts S (1981) Isolation of an internal clock Journal ofExperimental Psychology Animal Behavior Processes 7242ndash268
Roy E A Saint-Cyr J Taylor A amp Lang A (1993) Movementsequencing disorders in Parkinsonrsquos disease InternationalJournal of Neuroscience 73 183ndash194
Saint-Cyr J A Taylor A E amp Nicholson K (1995) Behaviorand the basal ganglia Advances in Neurology 65 1ndash28
Stelmach G E amp Worringham C J (1988) The control ofbimanual aiming movements in Parkinsonrsquos disease Jour-nal of Neurology Neurosurgery amp Psychiatry 51 223ndash231
Volkmann J Hefter H Lange H W amp Freund H J (1992)Impairment of temporal organization of speech in basalganglia diseases Brain amp Language 43 386ndash399
Wing M A Keele S amp Margolin L D (1984) Motor disor-der and the timing of repetitive movements In J Gibbonamp L Allan (Eds) Timing and time perception (pp 183ndash192) New York New York Academy of Sciences
Malapani et al 331




















