
Cortisol response and immune-related effects of Atlantic salmon ( Salmo salar Linnaeus) subjected to...
12
This article was published in an Elsevier journal. The attached copy is furnished to the author for non-commercial research and education use, including for instruction at the author’s institution, sharing with colleagues and providing to institution administration. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
wigan-leigh -
Category
Documents
-
view
0 -
download
0
Transcript of Cortisol response and immune-related effects of Atlantic salmon ( Salmo salar Linnaeus) subjected to...

This article was published in an Elsevier journal. The attached copyis furnished to the author for non-commercial research and
education use, including for instruction at the author’s institution,sharing with colleagues and providing to institution administration.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Cortisol response and immune-related effects ofAtlantic salmon (Salmo salar Linnaeus) subjected toshort- and long-term stress
Mark D. Fast 1, Sho Hosoya 2, Stewart C. Johnson, Luis O.B. Afonso*
National Research Council of Canada, Institute for Marine Biosciences 1411 Oxford Street, Halifax,Nova Scotia, B3H 2Z1, Canada
Received 17 August 2007; revised 24 October 2007; accepted 27 October 2007Available online 5 November 2007
KEYWORDSAcute stress;Cortisol;Free cortisol;Handling;Head kidney;Interleukin-1b;Long-term stress;Macrophage
Abstract It is generally considered that stress causes decreased immune function in fish.In this study we examined in Atlantic salmon (Salmo salar Linnaeus) the effects of both short-(a single 15 s out of water) and long-term (4 weeks of daily handling 15 s out of water) stress onplasma cortisol (free and total) and glucose levels, expression of interleukin-1b (IL-1b) and sur-vival of head kidney (HK) macrophages under culture with Aeromonas salmonicida. In theshort-term study, samples were collected prior to the application of the stressor, and at 1,3, 6, 12 and 24 h post stress. Free and total plasma cortisol levels and the percentage of freecortisol increased significantly in the stressed group at 1 and 3 h post stress. Plasma glucoselevels were significantly higher than those of control fish at 1, 3 and 6 h post stress. Constitu-tive expression of IL-1b in macrophages isolated from head kidneys in stressed fish was signif-icantly higher at 1 and 3 h post stress. However, lipopolysaccharide (LPS) stimulated expressionof IL-1b in HK macrophages, exhibited significantly higher fold increases in unstressed fish com-pared to stressed fish. In the long-term study, with the exception of an increase in plasma glu-cose levels at 1 week, there were no significant differences in stress parameters betweengroups. There was a significantly higher constitutive IL-1b expression in macrophages isolatedfrom stressed fish over the first 2 weeks. At weeks 1, 2 and 3 the magnitude of IL-1b response ofisolated HK macrophages to LPS stimulation was reduced in >90% of the stressed fish. At4 weeks there was no significant difference in inducible IL-1b expression between the groups.Macrophages isolated from stressed fish also showed significantly decreased survival when ex-posed to A. salmonicida. This study shows a clear pattern from repeated handling stress,whereby effects on immune cells begin with increased constitutive expression of IL-1b, fol-lowed by decreased stimulation of leucocytes by extracellular antigen, and finally decreased
* Corresponding author. Tel.: þ1 902 868 2180; fax: þ1 902 868 2015.E-mail address: [email protected] (L.O.B. Afonso).
1 Permanent address: School of Marine and Atmospheric Sciences, Stony Brook University, Stony Brook, NY, USA.2 Present address: Graduate School of Agricultural and Life Sciences, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-Ku, Tokyo, 113-8657,
Japan.
1050-4648/$ - see front matter Crown Copyright ª 2007 Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.fsi.2007.10.009
ava i lab le a t www.sc iencedi rec t .com
j ourna l homepage : www.e lsev ie r . com/ loca te / f s i
Fish & Shellfish Immunology (2008) 24, 194e204

Author's personal copy
leukocyte survival when exposed to A. salmonicida. The implications of these changes in theimmune system will be discussed with respect to the use of classical indicators of stress to pre-dict possible effects on the immune system of fish.Crown Copyright ª 2007 Published by Elsevier Ltd. All rights reserved.
Introduction
Similar to other vertebrates, fish elicit an organized set ofresponses, at different levels of organization, to deal withstressors. One of the primary responses is mediated by thehypothalamusepituitaryeinterrenal (HPI) axis, and is char-acterized by increased circulating levels of the glucocorti-coid hormone cortisol [1]. This initial response is thought tobe adaptive [2] and in the short term should provide alert-ness and energy for the animal to deal with the stressor andmaintain homeostasis [2,3].
Glucocorticoid hormones such as cortisol have also beenshown to be immunosuppressive [4,5]. Anti-inflammatoryaction and immunosupression by glucocorticoids is thoughtto occur through their inhibition of transcription factor ac-tivity (i.e. activator protein-1 and nuclear factor kB). Ele-vated levels of glucocorticoids thereby suppress humoralfactors involved in the inflammatory response, inhibit leu-kocyte trafficking to inflammatory sites, and overall reducecirculating leucocytes and lymphocytes [6e10].
Previous work in brown trout (Salmo trutta Linnaeus) hasshown that chronic elevation of plasma cortisol, throughhormone implantation, results in dose-dependent increasesin mortality due to common fungal and bacterial diseases[11]. Furthermore, stress and/or cortisol-induced increasesin pathogen susceptibility have been shown in numerousother hosts to a variety of pathogens [4,12e17]. However,most of these studies addressing the correlation betweencortisol levels and immunosupression and increased suscep-tibility to diseases are based upon exogenous administra-tion of cortisol. This could rather be a pharmacologicalthan a physiological effect. After synthesis and release,steroid hormones such as cortisol are transported in theblood by specific carrier proteins that regulate the avail-ability of the hormone to the tissues [18]. Usually, a variableamount of the steroid hormone will circulate free in theblood (not bound to the carrier protein) and this fractionhas been suggested to be the biologically active componentavailable to interact with the cell [18]. It is possible thatthe administration of exogenous cortisol increases thelevels of free cortisol, which could be mediating the effectsassociated with this hormone.
Recent studies have demonstrated a bi-directional com-munication between the neuroendocrine and immune sys-tems mediated by hormones and cytokines, respectively. Ithas been shown that the HPI-axis can affect [19] or can beaffected [20] by cytokines. Interleukin-1b (IL-1b) is an im-portant regulating cytokine that mediates several responsesafter an organism has been injured or infected [21].
The goals of this study were to investigate the effects ofshort- and long-term stress on the physiological stressresponse of Atlantic salmon and its subsequent effects onmacrophage function and survival. The physiological stress
response was studied by measuring total and free plasmacortisol and plasma glucose levels. Macrophage functionwas examined by studying the expression of IL-1b instimulated and unstimulated cells. Macrophage survival inthe presence of A. salmonicida was also assessed.
Materials and methods
Fish maintenance
Atlantic salmon were reared at the IMB-NRC Marine Re-search Station (Ketch Harbour, NS, Canada). JuvenileAtlantic salmon were initially held in two 500 l flow-through(20e25 l min�1) holding tanks supplied with seawater (tem-perature 10 � 1 �C; oxygen saturation >90%), and fed twicedaily (8:00 and 15:00 h) at a rate of 1.5% of average bodymass with a commercial Atlantic salmon diet (EWOS, Van-couver, BC, Canada). Three weeks prior to experimenta-tion, fish (mass 144.4 � 20 g, length 25.7 � 2 cm) weretransferred to sixteen 120 l experimental tanks (20 fish/tank) and allowed to acclimate to the experimental system.During the acclimation period the water flow, temperatureand oxygen saturation were, respectively: 8.0 l min�1,10 � 1 �C, and 90%. Daily photoperiod was 10 h light, 14 hdark.
Treatments and protocols
Short-term stressThere were two experimental groups: control and stressed(four replicate tanks in each). Following the 3-weekacclimation period, fish in the stress group were subjectedto a single handling stress (15 s out of the water). Tissuesamples (2 per replicate tank) were collected prior to stress(baseline levels in both groups) and at 1, 3, 6, 12 and 24 hpost-stress.
Long-term stressThere were two experimental groups: control and stressed(four replicate tanks in each). Following the 3-weekacclimation period, fish in the stress group were subjectedto a daily handling stress (15 s out of the water) for4 weeks. This procedure was carried out either at 8:00 AMor 3:00 PM, to prevent the fish from anticipating thestressor. Feedings were usually at 8:00 AM and 3:00 PM,but the timing of one of the feedings was adjusted sothat it occurred 1 h after handling. During this study, fish(2 per replicate tank) were sampled prior to initiation ofthe stress protocol (0 week) and 1, 2, 3 and 4 weeks afterit began. Samples were taken just prior to the handlingstress of the day, and were collected at least 18 h afterthe last stress was applied.
Cortisol response and immune-related effects of Atlantic salmon 195

Author's personal copy
Blood and tissue sampling
Two fish from each tank were quickly captured, andimmediately placed in a bucket containing a lethal dose(400 g l�1) of tricaine methanesulfonate (Syndel Laborato-ries Ltd., Vancouver, BC, Canada). Blood samples, collectedfrom the caudal vasculature using a 5 ml heparinized sy-ringe (23 gauge needle), were obtained immediately afterthe fish were anesthetized. The blood was transferred toa 4 ml Vacutainer tube (Becton, Dickinson and Company,Franklin Lakes, NJ, USA) containing 60 IU of heparin, andstored on ice prior to centrifugation. Plasma was separatedby centrifugation (3000 x g for 15 min) at 4 �C, removed andaliquoted into 1.5 ml microcentrifuge tubes. Samples werestored at �80 �C until subsequent analysis. Mass and lengthwere recorded from each fish. Head kidneys were also sam-pled from each fish as described below.
Head kidney macrophage isolation
A macrophage-enriched culture was obtained by isolatingadherent head kidney leukocytes from the anteriorkidney following previously described methods [22,23].Anterior kidneys were removed aseptically from the fishand placed immediately into 5 ml L-15 medium supple-mented with 2% fetal bovine serum (FBS), 100 unitsml�1 penicillin/streptomycin (P/S) and 10 units ml�1 hep-arin. Tissues were then stored on ice (1e2 h) until furtherprocessing. Tissues were dissociated by repeated passagethrough a 3 ml syringe and fragments allowed to settlefor 10 min prior to removal of suspended cells. Cell sus-pensions were pelleted (500 � g for 10 min at 4 �C) andwashed twice prior to layering on Percoll (Sigma, Missas-sauga, ON, Canada) gradients (34/51%). Cells were centri-fuged at 400 � g for 20 min at 4 �C and the macrophageenriched fraction collected at the 34/51% interface. Cellswere then re-suspended in 10 ml L-15/2% FBS, pelletedby centrifugation at 500 � g for 10 min at 4 �C, washedwith 10 ml of L-15/2% FBS and then resuspended in L-15/5% FBS with 100 units ml�1 P/S. Viable cells werecounted using the Trypan blue exclusion method andcell density adjusted to 1 � 107 cells ml�1 in L-15/0.1%FBS. Cells from each fish were then individually platedat 100 ml per well on 96 well plates and incubated at18 �C for 2 h. After 2 h, medium and non-adherent cellswere removed and an equal volume of L-15/5% FBSadded. Cells were maintained for 1.5 days at 18 �C ina humidified chamber, prior to manipulation. All chemi-cals used for cell culture were obtained from Invitrogen(Burlington, ON, Canada) unless otherwise stated.
Separation of free plasma cortisol
Free and protein-bound cortisol were separated by ultra-filtration using commercial columns (Centrifree�- MPS mi-cropartition devices, Millipore Corp., Billerica, MA, USA)following the method described in Hosoya et al. [24]. Sam-ples were thawed on ice and 200 ml of plasma was loadedonto each column. Columns were centrifuged at 2000 � gfor 30 min at 10 �C. Free plasma cortisol was determinedfrom the filtrate using the method described below.
Determination of plasma glucose, and free andtotal plasma cortisol
Plasma glucose levels were measured in duplicate usinga modified Trinder [25] enzymatic assay available in kitform (Diagnostic Chemicals Ltd. Oxford, CT, USA.) Plateswere read at 505 nm on a Molecular Devices VERSAmax mi-croplate reader (Molecular Devices Corporation, Sunnyvale,CA, USA), and intra- and inter-assay coefficients of varia-tion were always less than 5%.
Free and total plasma cortisol levels were determinedusing an enzyme-linked immunosorbent assay (ELISA) kit(Neogen Corp., Lexington, KY, USA), and the MolecularDevice VERSAmax microplate reader at an absorbance of650 nm. Parallelism to the standard curve was confirmedusing serially diluted plasma sample. Intra- and inter-assaycoefficients of variation (based on 4 samples) did not ex-ceed 10%.
Aeromonas salmonicida cultures
Aeromonas salmonicida strain A449 was cultured on trypticsoy agar (TSA) from glycerol stocks. Broth cultures wereprepared from single colonies in TSB and cultured withagitation at 17 �C. The OD was determined at 600 nm andadjusted to 0.1. The bacteria were then pelleted andresuspended in an equal volume of PBS. Bacteria werefurther diluted for the cytotoxicity assay at 1:200,1:2.0 � 10�4, 1:2.0 � 10�6 and 1:2.0 � 10�8 (i.e. 1 � 105,1 � 103, 10, <1 bacteria per well, respectively) at week0. For weeks 2e4, 1 � 105 bacteria were used for the cyto-toxicity assay.
Cytotoxicity assay
Isolated macrophages were incubated with Aeromonas sal-monicida (A449) to determine relative cytotoxicity to cellsfrom stressed and unstressed fish. The cytotoxicity assayfollowed manufacturer’s instructions (Roche, Basel, Swit-zerland). After 1.5 days, plates were washed (2 � 200 ml)with Dulbecco’s PBS (DPBS) (with Caþþ and Mgþþ) (Invitro-gen). To the positive control wells, 100 ml of DPBS þ 100 mlof 2% sterile Triton X-100 (Sigma) were added. To the neg-ative controls, 200 ml of DPBS was added. This was alsodone to blank wells containing no cells. Finally, dilutionsof Aeromonas salmonicida were added (10 ml) þ 190 mlDPBS to treatment wells.
Plates were incubated for 6 h at 18 �C. Following incuba-tion, plates were centrifuged (250 � g) for 5 min and super-natant (100 ml) transferred to a new microtiter plate.Reaction mixture, including catalyst and dye, was addedto each well (100 ml) and the plates were incubated in thedark at RT for 30 min. The release of lactate dehydrogenasewas measured from damaged cells by colorimetric assay.Plates were read at a wavelength combination of 490 nmand 650 nm and the optical density (OD) used to calculatepercentage cytotoxicity:
ðsample OD� negative controlÞðpositive control ODÞ � 100 ð1Þ
196 M.D. Fast et al.

Author's personal copy
Positive and negative controls, as well as all treatments(Aeromonas salmonicida dilutions) were carried out in trip-licate on multiple animals.
Cell stimulation and RNA isolation for gene studies
After 1.5 days medium was removed and 100 ml of freshL-15/5% with 0 or 5 mg ml�1 Aeromonas salmonicida lipo-polysaccharide (LPS) were added. Stimulation of cells wascarried out for 4 h at 18 �C, at which time the mediumwas removed and TRIzol reagent added (1 ml, Invitrogen).Cells were mixed with solution using a pipette and after5 min incubation at room temperature (22 �C), chloroform(200 ml, Sigma) was added to the homogenates and mixedvigorously. Samples underwent a further 3 min incubationat RT prior to centrifugation at 12,000 � g for 15 min at4 �C. Equal volumes of the aqueous phase and molecularbiological grade isopropyl alcohol were mixed (Sigma),incubated for 10 min (RT) and the RNA was pelleted at12,000 � g (4 �C for 5 min). The pellet was washed with1000 ml of 75% ethanol, air dried for 30 min and dissolved in50 ml molecular biology grade water (Sigma). Samples wereincubated at 55 �C for 10 min prior to storage at �80 �C.Addition of LPS had no effects on cell viability, morphologyor adherence properties in this study (data not shown).
Total RNA was measured using the NanoDrop-1000Spectrophotometer (v3.2.1) and samples stored at �80 �Cuntil cDNA generation. For reverse transcription, 1.5 mg oftotal RNA from each sample were dissolved in molecularbiological grade water. Two-step reverse transcription-real-time PCR was carried out using the Superscript III qRT-PCR kit with SYBR green (Invitrogen) on an iCycler iQ�Real-Time detection system (BioRad, Missassauga, ON, Can-ada). Following first strand synthesis, samples were storedat �20 �C until use in real-time PCR assays. All sampleswere first tested for gDNA contamination through PCR ofRT-enzyme negative controls. Any samples showing amplifi-cation were DNase treated and re-confirmed to be absent ofcontaminating gDNA prior to use in expression studies.
Manufacturer’s instructions were followed with thefollowing exceptions. The qPCR Mastermix was made upto 47.5 ml (including 1.25 ml of each primer) and 2.5 ml ofcDNA reaction added for each sample and standard. The to-tal volume of 50 ml was then divided into separate wells forduplication of reactions.
Real-time PCR primers were designed based on pre-viously published Atlantic salmon sequences. PCR productsof elongation factor (EF)-1 A (forward-; reverse-), IL-1b(forward-; reverse-) were cloned into a TA-cloning vector(pCR 4-TOPO, Invitrogen) and sequenced to confirm se-quence of amplified products. Plasmid vectors were iso-lated and used as standards for real-time studies [22]. Thesequences of the oligonucleotide primers used in real-timePCR are described in Fast et al. [22,26].
The PCR profile for both products was as follows: aninitial 3 min denaturation step at 95 �C, followed by 40 cy-cles of denaturation (30 s at 95 �C), annealing (30 s at 53 �C)and extension (30 s at 72 �C). The sensitivity of reactionsand amplification of contaminant products, such as primerdimers indiscriminately detected by SYBR green, were eval-uated by amplifying 10 fold dilutions of the clones (1e10�6 ng) and duplicate samples as well as by performing
a blank without cDNA with each run. The relationship be-tween the threshold cycle (Ct) and the log(RNA) was linear(�3.35 < slope < �3.25) for all reactions.
Data analysis
All data are represented as means � SEM, with mean valuescomprised of values from each of the 4 replicate tanks: 2fish per tank for plasma glucose and cortisol (8 fish pertreatment at each sampling time). Data were submitted toa two-way (main effects group and sampling time) analysisof variance (ANOVA), and the StudenteNewmaneKeuls testwas applied to discern differences among means for thosetreatments that had statistically significant differences(p < 0.05) (SigmaStat 3.0, SPSS Inc., Chicago, IL, USA). When-ever necessary the data was log-transformed to achieveeither a normal distribution or equal variance. Spearmanrank-order correlations were used to determine the relation-ship between plasma glucose, plasma cortisol and IL-1bexpression. Elongation factor-1a was chosen as thehousekeeping gene for this study based on recent findingsin Atlantic salmon [27,28]. The statistical significance ofgene expression differences was assessed using one-wayanalysis of variance (p < 0.05).
Results
Elongation factor-1a validation
Elongation factor-1a was chosen as the house-keeping forthis study and showed no significant changes in its expres-sion in enriched-macrophage culture, regardless of LPS-stimulation of cells or stress application to animals. Thiswas confirmed across both the short- and long-term studies.The mean copy number of EF-1a during the short-termstudy was 1.69 � 107 � 1.69 � 106/1.5 mg of TotalRNA,1.14 � 107 � 4.36 � 106/1.5 mg of TotalRNA, 1.64 � 107 �1.68 � 106/1.5 mg of TotalRNA, 1.86 � 107 � 3.91 � 106/1.5 mg of TotalRNA, for 0, 1, 3 and 24 h, respectively. Themean copy number of EF-1a during the long-term studywas 1.20 � 107 � 3.42 � 106/1.5 mg of TotalRNA, 9.22 �107 � 3.09 � 106/1.5 mg of TotalRNA, 0.52 � 107 � 1.39 �106/1.5 mg of TotalRNA, 1.17 � 107 � 3.30 � 106/1.5 mg ofTotalRNA, 1.37 � 107 � 2.02 � 106/1.5 mg of TotalRNA forweeks 0, 1, 2, 3 and 4, respectively.
Short-term stress (15 s handling stress)
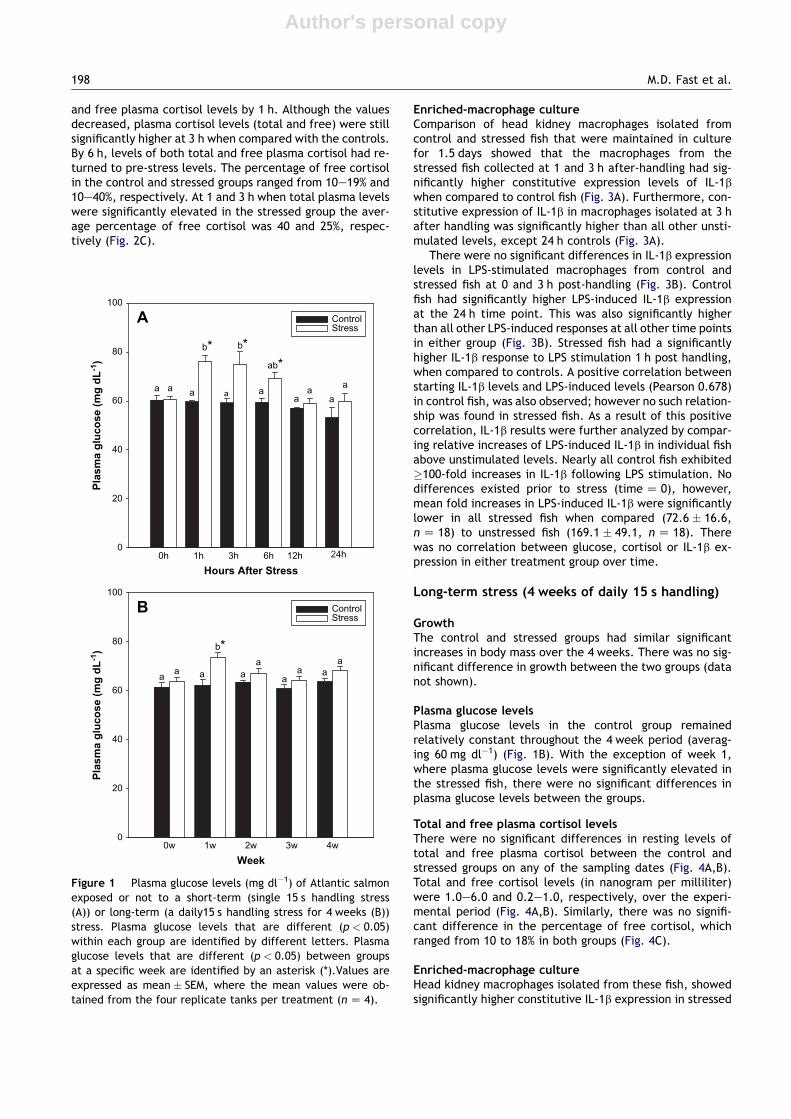
Plasma glucose levelsPlasma glucose levels in the control group remained steady(approximately 60 mg dl�1) at all sampling times (Fig. 1A). Forfish subjected to handling stress, plasma glucose levels in-creased significantly at 1 h after stress, and remained elevatedup to 3 h. By 12 h plasma glucose levels in the stressed fish haddeclined to the same level as that of the controls (Fig. 1A).
Total and free plasma cortisol levels and percentage offree cortisolTotal and free plasma cortisol levels in the control groupremained low at all sampling times (Fig. 2 A,B). Fish sub-jected to handling stress had significantly elevated total
Cortisol response and immune-related effects of Atlantic salmon 197
Author's personal copy
and free plasma cortisol levels by 1 h. Although the valuesdecreased, plasma cortisol levels (total and free) were stillsignificantly higher at 3 h when compared with the controls.By 6 h, levels of both total and free plasma cortisol had re-turned to pre-stress levels. The percentage of free cortisolin the control and stressed groups ranged from 10e19% and10e40%, respectively. At 1 and 3 h when total plasma levelswere significantly elevated in the stressed group the aver-age percentage of free cortisol was 40 and 25%, respec-tively (Fig. 2C).
Enriched-macrophage cultureComparison of head kidney macrophages isolated fromcontrol and stressed fish that were maintained in culturefor 1.5 days showed that the macrophages from thestressed fish collected at 1 and 3 h after-handling had sig-nificantly higher constitutive expression levels of IL-1bwhen compared to control fish (Fig. 3A). Furthermore, con-stitutive expression of IL-1b in macrophages isolated at 3 hafter handling was significantly higher than all other unsti-mulated levels, except 24 h controls (Fig. 3A).
There were no significant differences in IL-1b expressionlevels in LPS-stimulated macrophages from control andstressed fish at 0 and 3 h post-handling (Fig. 3B). Controlfish had significantly higher LPS-induced IL-1b expressionat the 24 h time point. This was also significantly higherthan all other LPS-induced responses at all other time pointsin either group (Fig. 3B). Stressed fish had a significantlyhigher IL-1b response to LPS stimulation 1 h post handling,when compared to controls. A positive correlation betweenstarting IL-1b levels and LPS-induced levels (Pearson 0.678)in control fish, was also observed; however no such relation-ship was found in stressed fish. As a result of this positivecorrelation, IL-1b results were further analyzed by compar-ing relative increases of LPS-induced IL-1b in individual fishabove unstimulated levels. Nearly all control fish exhibited�100-fold increases in IL-1b following LPS stimulation. Nodifferences existed prior to stress (time Z 0), however,mean fold increases in LPS-induced IL-1b were significantlylower in all stressed fish when compared (72.6 � 16.6,n Z 18) to unstressed fish (169.1 � 49.1, n Z 18). Therewas no correlation between glucose, cortisol or IL-1b ex-pression in either treatment group over time.
Long-term stress (4 weeks of daily 15 s handling)
GrowthThe control and stressed groups had similar significantincreases in body mass over the 4 weeks. There was no sig-nificant difference in growth between the two groups (datanot shown).
Plasma glucose levelsPlasma glucose levels in the control group remainedrelatively constant throughout the 4 week period (averag-ing 60 mg dl�1) (Fig. 1B). With the exception of week 1,where plasma glucose levels were significantly elevated inthe stressed fish, there were no significant differences inplasma glucose levels between the groups.
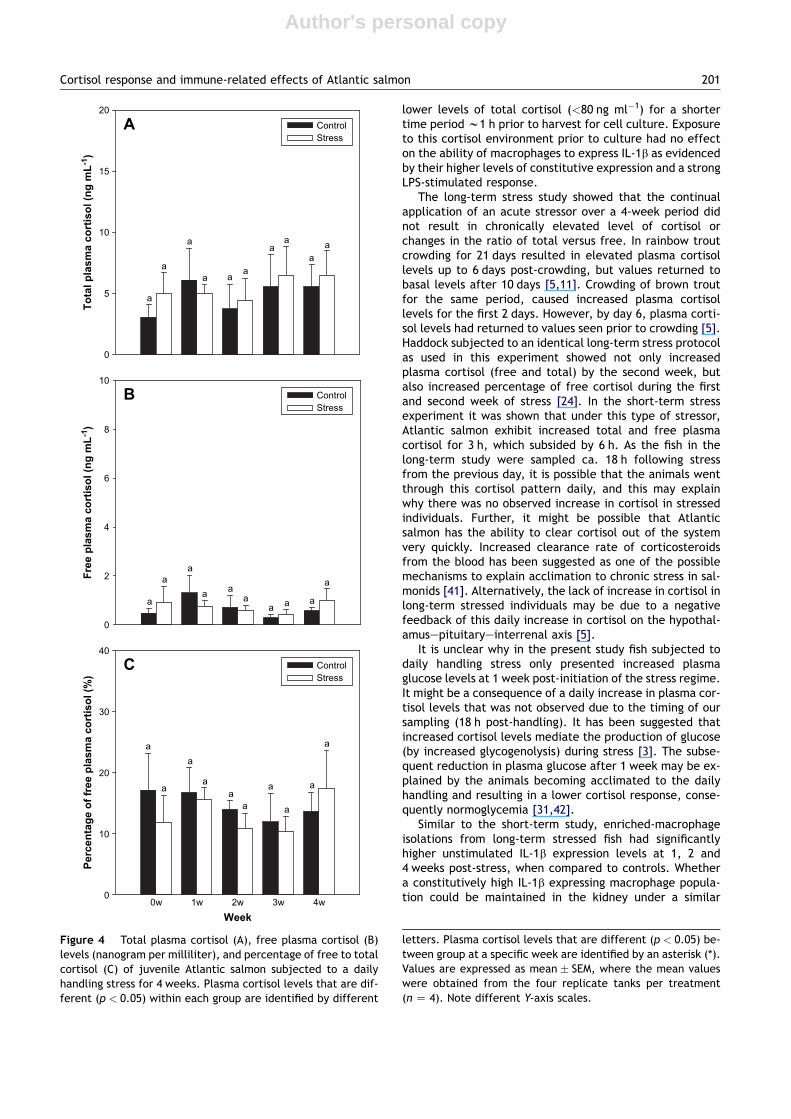
Total and free plasma cortisol levelsThere were no significant differences in resting levels oftotal and free plasma cortisol between the control andstressed groups on any of the sampling dates (Fig. 4A,B).Total and free cortisol levels (in nanogram per milliliter)were 1.0e6.0 and 0.2e1.0, respectively, over the experi-mental period (Fig. 4A,B). Similarly, there was no signifi-cant difference in the percentage of free cortisol, whichranged from 10 to 18% in both groups (Fig. 4C).
Enriched-macrophage cultureHead kidney macrophages isolated from these fish, showedsignificantly higher constitutive IL-1b expression in stressed
Hours After Stress
0h 1h 3h 6h 12h
Plasm
a g
lu
co
se (m
g d
L-1)
0
20
40
60
80
100
Week
0w 1w 2w 3w 4w
Plasm
a g
lu
co
se (m
g d
L-1)
0
20
40
60
80
100
b*
aa
aa
b*
a
b*ab*
a aa a a aa a
a a a aa
A
B
24h
Control Stress
Control Stress
Figure 1 Plasma glucose levels (mg dl�1) of Atlantic salmonexposed or not to a short-term (single 15 s handling stress(A)) or long-term (a daily15 s handling stress for 4 weeks (B))stress. Plasma glucose levels that are different (p < 0.05)within each group are identified by different letters. Plasmaglucose levels that are different (p < 0.05) between groupsat a specific week are identified by an asterisk (*).Values areexpressed as mean � SEM, where the mean values were ob-tained from the four replicate tanks per treatment (n Z 4).
198 M.D. Fast et al.

Author's personal copy
fish when compared to control fish at weeks 1, 2 and 4 post-initiation of the long-term stress regime (Fig. 5A). Macro-phages isolated from control fish at weeks 2 and 3 showedsignificantly higher LPS-induced IL-1b expression whencompared to stressed fish (Fig. 5B). Macrophages isolatedfrom stressed fish at 2 weeks showed the lowest fold-in-crease in IL-1b expression following LPS treatment, in ei-ther study (Fig. 5). There was no correlation between IL-1b starting levels and LPS response levels, or cortisol levelsin either group. Individual responses to LPS stimulationwere often on the order of 10-fold higher following LPSstimulation in both control and stressed fish (11/12 fish ineach group) at 0 and 4 weeks. However, at weeks 1, 2,and 3 post-initiation of the long-term stress regime, only1/18 stressed fish exhibited an LPS-induced response �10-fold higher than resting levels (week 2). Conversely, overthe same period 13/18 control fish had �10-fold higherLPS responses (3, 5 and 5 individuals, respectively). Thesefold changes in both the control and stressed fish were anorder of magnitude lower than those observed from theshort-term stress study.
Head kidney macrophages isolated from fish prior to thestress experiment (time Z 0) were used to determine anoptimal concentration of Aeromonas salmonicida for use incytotoxicity assays. It was determined that addition of1 � 105 A. salmonicida cells resulted in ca. 20% mortalityof host leucocytes within 6 h, and this concentration of A.salmonicida was used in all subsequent trails. Prior to3 weeks post-initiation of the long-term stress regime therewas no significant difference in survival of macrophages be-tween the stressed and control group in the absence ofA. salmonicida. However at 3 and 4 weeks, macrophages iso-lated from stressed individuals had significantly higher ratesof cell death in culture when compared to those obtainedfrom control fish (Fig. 6A). There was no significant differ-ence in the survival of macrophages obtained from stressedand control fish following exposure to A. salmonicida at 0, 1,2 and 3 weeks. Macrophages obtained from stressed individ-uals at week 4 exhibited significantly higher rates of celldeath when incubated with A. salmonicida when comparedto those obtained from control fish (Fig. 6B).
Discussion
In this study, we investigated the effects of short- and long-term stress on physiological and immunological responsesof Atlantic salmon. We showed for the first time the levelsof both free and total cortisol in a salmonid subjected toshort- and long-term stress. Further, we demonstrated thatshort- and long-term stress affected the expression of theinflammatory gene interleukin 1b (IL-1b).
No significant effect of stress was observed on overallgrowth of Atlantic salmon as long-term stressed fish hadsimilar mass to control at the end of the study. The effects
To
tal p
lasm
a co
rtiso
l (n
g m
L-1)
0
20
40
60
80
100
Free p
lasm
a co
rtiso
l (n
g m
L-1)
0
20
40
60
80
100
Hours after stress
0h 1h 3h 6h 12h 24h
Percen
tag
e o
f free p
lasm
a co
rtiso
l (%
)
0
20
40
60
80
100
aa a a aa
a aaa
c*
b*
aa a a a a a aaa
a*
b*
a aa ab ab ab
a a
b*
a
ab*
a
A
C
B
ControlStress
ControlStress
ControlStress
Figure 2 Total plasma cortisol (A), free plasma cortisol (B)levels (ng ml�1), and percentage of free to total cortisol (C)of Atlantic salmon subjected to a single 15 s handling stress.Plasma cortisol levels that are different (p < 0.05) withineach group are identified by different letters. Plasma cortisol
levels that are different (p < 0.05) between groups at a specificsampling time are identified by an asterisk (*). Values are ex-pressed as mean � SEM, where the mean values were obtainedfrom the four replicate tanks per treatment (n Z 4). Note dif-ferent Y-axis scales.
Cortisol response and immune-related effects of Atlantic salmon 199

Author's personal copy
of chronic stress on growth in fish are a controversial issue[29] and the results have been shown to vary not only withthe stressor and its duration, but also with the species (be-tween and within species) (rainbow trout, Oncorhynchusmykiss Walbaum [30,31]; haddock, Melanogrammus aegle-finus Linnaeus [24]). Our findings are similar to the resultsof Barton et al. [31], who reported no change in the growthrate of rainbow trout exposed daily to physical stressors forup to 10 weeks.
The percentage of free cortisol for unstressed Atlanticsalmon ranged from 10 to 20%, and it was within the rangeobserved for other salmonid [32,33] and non salmonid fishes[24]. The percentage of free cortisol for unstressed mam-mals range from 6e14% [48].Short-term stress resulted inincreased total and free plasma cortisol levels at 1 and3 h after stress. By 1 h, 40% of total cortisol was presentin the free form. Mammals usually show a 2% increase in
the percentage of free cortisol after acute stress [34,35].In contrast, a recent study in fish [24] demonstrated thathaddock did not increase the percentage of free cortisol af-ter an acute stress. Thus, these studies suggest there arespecies specific differences in the transport of cortisolwithin the blood. The reason for this difference betweenspecies is unknown. In higher vertebrates steroid hormonetransport in the blood is mediated by specific proteins[36] and the cortisol-binding globulin (CBG) and albuminare the main carriers of the stress hormone cortisol. Infish, proteins involved in plasma cortisol regulation remainto be identified. It is possible that different species of fishmay have different types or levels of carrier proteins intheir plasma or that there may be differences in the affin-ity/binding capacity of carrier proteins between species.
Previous work has shown that acute stress increases thenumbers of leucocytes in the head kidney of juvenile cohosalmon (Oncorhynchus kisutch Walbaum) within 24 h of thestress [6]. These authors also suggest that this redistribu-tion of leucocytes is not solely driven by cortisol. The tim-ing and magnitude of the cortisol response in our study andthe increased constitutive expression of IL-1b, suggest thatthe cortisol response occurred first and may be either di-rectly or indirectly responsible for the increased levels ofexpression in head kidney macrophages at 1 and 3 h post-handling. Common carp (Cyprinus carpio Linnaeus) sub-jected to restraint for 24 h showed significantly increasedplasma cortisol levels and concomitant elevation in IL-1bgene expression in the head kidney, pituitary, and nucleuspreopticus [20,37]. We demonstrated that head kidneymacrophage enriched cell cultures isolated from fish fol-lowing a single acute stress have higher constitutive IL-1bexpression at 1.5 days post- handling when compared tomacrophages from unhandled controls. As high cortisollevels have been removed from these cells for >36 h, theeffects of acute administration should have subsided. Thissuggests that we may be measuring differences in thepopulations of adherent cells taken from fish at 1 and 3 hpost-handling. Previous work has shown that cortisol cannegatively affect adherence in cultured human A549 lungtumor cells [38]. Since cortisol levels were highest at 1and 3 h (same time as increased IL-1b expression in unsti-mulated cells), it suggests that the higher expressingIL-1b cells are more resistant to cortisol affects on adher-ence, and therefore make up a greater proportion of thecells in the kidney at these times. It is also possible thatremoval of the HPI-axis and the bi-directional feedbackon the leucocytes in culture may result in the maintenanceof this higher basal level of expression [20,39].
Subsequent stimulation with LPS resulted in no differ-ence in IL-1b expression between the stressed and non-stressed groups. This occurred despite the differences inconstitutive IL-1b expression between these groups. Inmacrophages isolated from control fish constitutive IL-1bexpression was positively correlated to LPS-stimulated IL-1b expression levels. This was not observed in macrophagesobtained from stressed animals. Zou et al. [40] treated cul-tured rainbow trout leucocytes with cortisol (100 ng ml�1
for 2 h) and then stimulated the cells with LPS while theywere still in the cortisol environment. This resulted in a de-crease (ca. 30%) in IL-1b expression. In our study the headkidney macrophages would have been exposed in vivo to
IL
-1 E
xp
ressio
n (U
nstim
ulated
)
0.00
0.01
0.02
0.03
0.04
0.05
Hours after stress
0h 1h 3h 24h
IL
-1 E
xp
ressio
n (L
PS
)
0
2
4
6
a
*
*
*
*
A
B ControlStress
ControlStress
Figure 3 Mean expression of (�SEM) interleukin-1b relative toEF-1A in control and short-term stressed Atlantic salmon headkidney macrophages, stimulated with (B) or without lipopolysac-charide (A). The asterisk (*) signifies a significant difference(p < 0.05, one-way ANOVA, n Z 6). Note different Y-axis scales.
200 M.D. Fast et al.
Author's personal copy
lower levels of total cortisol (<80 ng ml�1) for a shortertime period w1 h prior to harvest for cell culture. Exposureto this cortisol environment prior to culture had no effecton the ability of macrophages to express IL-1b as evidencedby their higher levels of constitutive expression and a strongLPS-stimulated response.
The long-term stress study showed that the continualapplication of an acute stressor over a 4-week period didnot result in chronically elevated level of cortisol orchanges in the ratio of total versus free. In rainbow troutcrowding for 21 days resulted in elevated plasma cortisollevels up to 6 days post-crowding, but values returned tobasal levels after 10 days [5,11]. Crowding of brown troutfor the same period, caused increased plasma cortisollevels for the first 2 days. However, by day 6, plasma corti-sol levels had returned to values seen prior to crowding [5].Haddock subjected to an identical long-term stress protocolas used in this experiment showed not only increasedplasma cortisol (free and total) by the second week, butalso increased percentage of free cortisol during the firstand second week of stress [24]. In the short-term stressexperiment it was shown that under this type of stressor,Atlantic salmon exhibit increased total and free plasmacortisol for 3 h, which subsided by 6 h. As the fish in thelong-term study were sampled ca. 18 h following stressfrom the previous day, it is possible that the animals wentthrough this cortisol pattern daily, and this may explainwhy there was no observed increase in cortisol in stressedindividuals. Further, it might be possible that Atlanticsalmon has the ability to clear cortisol out of the systemvery quickly. Increased clearance rate of corticosteroidsfrom the blood has been suggested as one of the possiblemechanisms to explain acclimation to chronic stress in sal-monids [41]. Alternatively, the lack of increase in cortisol inlong-term stressed individuals may be due to a negativefeedback of this daily increase in cortisol on the hypothal-amusepituitaryeinterrenal axis [5].
It is unclear why in the present study fish subjected todaily handling stress only presented increased plasmaglucose levels at 1 week post-initiation of the stress regime.It might be a consequence of a daily increase in plasma cor-tisol levels that was not observed due to the timing of oursampling (18 h post-handling). It has been suggested thatincreased cortisol levels mediate the production of glucose(by increased glycogenolysis) during stress [3]. The subse-quent reduction in plasma glucose after 1 week may be ex-plained by the animals becoming acclimated to the dailyhandling and resulting in a lower cortisol response, conse-quently normoglycemia [31,42].
Similar to the short-term study, enriched-macrophageisolations from long-term stressed fish had significantlyhigher unstimulated IL-1b expression levels at 1, 2 and4 weeks post-stress, when compared to controls. Whethera constitutively high IL-1b expressing macrophage popula-tion could be maintained in the kidney under a similar
To
tal p
lasm
a co
rtiso
l (n
g m
L-1)
Free p
lasm
a co
rtiso
l (n
g m
L-1)
Week
0w 1w 2w 3w 4w
Percen
tag
e o
f free p
lasm
a co
rtiso
l (%
)
0
5
10
15
20
A
B
C
ControlStress
ControlStress
ControlStress
0
2
4
6
8
10
0
10
20
30
40
a
a
a
a
aa
a
aa
a
a
a
a
a a
aa
a
aa
a
aa
aa
a a
a
aa
Figure 4 Total plasma cortisol (A), free plasma cortisol (B)levels (nanogram per milliliter), and percentage of free to totalcortisol (C) of juvenile Atlantic salmon subjected to a dailyhandling stress for 4 weeks. Plasma cortisol levels that are dif-ferent (p < 0.05) within each group are identified by different
letters. Plasma cortisol levels that are different (p < 0.05) be-tween group at a specific week are identified by an asterisk (*).Values are expressed as mean � SEM, where the mean valueswere obtained from the four replicate tanks per treatment(n Z 4). Note different Y-axis scales.
Cortisol response and immune-related effects of Atlantic salmon 201

Author's personal copy
cortisol mechanism as suggested for plasma glucose isunknown. However, previous work, in Atlantic salmonundergoing a Lepeophtheirus salmonis infection has shownthe same results. Fast et al. [22] showed no changesin plasma cortisol at 2 and 3 weeks post-infection (pi),whereas increased constitutive expression of IL-1b (12 days)and a decreased constitutive expression of IL-1b (21 days),were observed in macrophage-enriched isolations from thekidney, when compared to controls. Along those same lines,the macrophage-enriched isolations after repeated stress,also exhibited an initial significant increase (at 1 week)and decrease (2 and 3 weeks) of LPS-stimulated IL-1b ex-pression, when compared to controls. Furthermore at2 weeks there was no significant LPS-stimulation of IL-1bexpression in macrophages from stressed fish. The Fastet al. [22] data also supports this, whereby 12 dp L. salmonis
infection, again no IL-1b stimulation via LPS was observed inmacrophages from infected fish, but by 21 days, LPS-induced IL-1b expression was significant. Fast et al. [22]also showed an increase in unstimulated expression anda reduced LPS response in two other immune related genes(major histocompatability class I and cyclooxygenase-2) inthese cells over the same time period.
Basal expression of IL-1b in the long-term study, wasnearly 10-fold higher in fish sampled at time 0, and weeks 1,3 and 4, when compared to the short-term study. Notably,even the fish sampled at the 24 h time point, in the short-term study, showed 2e3 fold higher constitutive expressionwhen compared to the other short-term time points. Thelong-term experiment was conducted shortly after the
IL
-1 E
xp
ressio
n (U
nstim
ulated
)
0.0
0.1
0.2
0.3
0.4
0.5IL
-1 E
xp
ressio
n (L
PS
)
0
2
4
6
* *
*
A
B
0w 1w 2w 3w 4wWeek
* * *
ControlStress
ControlStress
Figure 5 Mean expression of (�SEM) interleukin-1b relativeto EF-1A in control and long-term stressed Atlantic salmonhead kidney macrophages, stimulated without (A) or with lipo-polysaccharide (B). The asterisk (*) signifies a significantdifference (p < 0.05, one-way ANOVA, n Z 6). Note differentY-axis scales.
Co
ntro
l C
yto
to
xicity
0
1
2
3
4
5
Week
0w 1w 2w 3w 4w
Cyto
to
xicity to
A
449
0
10
20
30
40
50
a
**
*
*
A
B
ControlStress
ControlStress
Figure 6 Mean percent cell population death (�SEM) in con-trol and long-term stressed Atlantic salmon head kidney macro-phages, cultured in the absence (A) or presence (B) ofAeromonas salmonicida (isolate A449). Cells were incubatedfor 6 h after which time cell death was determined by lactatedehydrogenase activity released from damaged cells. The as-terisk (*) signifies a significant difference (p < 0.05, one-wayANOVA, n Z 6).
202 M.D. Fast et al.

Author's personal copy
acute study possibly suggesting that monocyte populations(i.e. low and high IL-1b expressing cells) of Atlantic salmon,may undergo cyclical 2e3 week patterns of tissue residenceand replacement, which can be regulated/altered by stress.
The reduced responses of macrophages to extracellularantigen were followed by increased cell death both asa result of incubation with and without A. salmonicida.Again, although no changes in cortisol were measuredover the course of the long-term study, these results aresimilar to other work on the effect of repeated stress orcortisol administration on phagocytic capacity (IL-1b stimu-lates phagocytosis) and disease resistance [7].
There are several studies addressing the effects of stresson the immune function in fish (for review see [43,44]), andmost of them concentrate on the effects of the classical in-dicator of the neuroendocrine response: the glucocorticoidhormone cortisol. A large amount of the evidence forcortisol affecting the immune function in fish comes fromin vivo and in vitro studies where exogenous cortisol hasbeen administered either through diet or implants placedwithin the body cavity to mimic the effects of stress[31,44,45]. Under these conditions levels of cortisol mayincrease to values not only above what would be expectedin a normal physiological response to stress but also forextended periods of time [31,45]. Furthermore, it is notknown whether the cortisol that is present following exog-enous administration is in the bound or free form, whichhas important consequences with respect to its biologicaleffects [18]. Thus, this brings up the question whethercortisol administration is an adequate model for mimickingthe effects of stress in fish. We have demonstrated that theapplication of a handling stress daily for a period of 4 weeksdid not result in chronic elevated levels of plasma cortisolor have any effect on the ratio of free to bound forms.This is the first study in Atlantic salmon examining thelevels of free plasma cortisol and the ratio between freeand total cortisol during short and long-term handlingstress. Rainbow trout subjected to daily handling stressfor 10 weeks did not show chronic elevation of total plasmacortisol levels at 0, 4, and 8 weeks of treatment [31]. Otherstressors such as crowding can result in the chronic eleva-tion of total plasma cortisol levels [5,11]. Haddock exposedto the same long-term stress and sampling regime showedelevated levels of total and free plasma cortisol [24]. Takentogether these data show that it is necessary to understandthe dynamics, magnitude and make-up (free vs. bound) ofthe cortisol response for each stressor and each speciesof fish to be studied.
The long-term stress model used in this study may also bemore appropriate to investigate compensation after expo-sure to a continuous stressor. Prolonged exposure toa stressor can lead to allostasis, which is the ability of thebody to return to physiological levels seen prior to stress[46,47]. But even though animals chronically stressed seemto compensate physiologically to the stressor, their abilityto perform important functions at the whole organism levelmay be affected [47]. By using the long-term stress modelwe showed that the indicators of the whole organism re-sponse to stress (plasma cortisol and glucose levels) returnedto normal values along the treatment; however, the gene ex-pression and death of host cells indicate that the immunefunction may have been impaired. Thus, suggesting that
the long-term handling stress may have imposed an allostaticload [46] that may be compromising the immune function.
In summary, this study shows that short-term but notlong-term stress in Atlantic salmon results in elevation offree and total plasma cortisol levels and in the percentage offree cortisol. It further shows a clear pattern from acute andrepeated handling stress, whereby effects on immune cellscan initially be stimulatory, such as an increased constitutiveexpression of the inflammatory gene IL-1b, followed bybeing inhibitory over the longer-term with decreased stim-ulation of leucocytes by LPS and finally decreased leukocytesurvival in the presence of A. salmonicida. Further studieson the mechanism by which highly IL-1b expressing cellsare maintained in the kidney and their immune functionare needed, as are investigations into the effects that differ-ent methods of cortisol administration have on free andbound cortisol levels. These data provide further evidencethat in salmonids responses at the neuroendocrine systemcan affect the activity of the immune system.
Acknowledgements
This project was funded by the Genomics and HealthInitiative (GHI-3), National Research Council Canada. Weappreciate all the support from Ron Melanson, JohnMahtab, Laura Garrison, Larry Staples and Corey Coldwellfrom IMB’s Marine Research Station.
References
[1] Iwama GK, Afonso LOB, Vijayan MM. Stress in fishes. In:Evans DH, Claiborne JB, editors. The physiology of fishes.Boca Raton: CRC Press; 2005. p. 320e42.
[2] Schreck CB. Stress and compensation in teleostean fishes: re-sponse to social and physical factors. In: Pickering AD, editor.Stress and fish. London: Academic Press; 1981. p. 295e321.
[3] Mommsen TP, Vijayan MM, Moon TW. Cortisol in teleost: dy-namics, mechanism of action, and metabolic regulation. RevFish Biol 1999;9:211e68.
[4] Pickering AD, Pottinger TG. Cortisol can increase the suscepti-bility of brown trout, Salmo trutta L., to disease without re-ducing the white blood cell count. J Fish Biol 1985;27:611e9.
[5] Pickering AD, Pottinger TG. Crowding causes prolonged leuco-penia in salmonid fish, despite interrenal acclimation. J FishBiol 1987;30:701e12.
[6] Maule AG, Shreck CB. Changes in numbers of leukocytes in im-mune organs of juvenile coho salmon after acute stress or cor-tisol treatment. J Aquat Anim Health 1990;2:298e304.
[7] Ainsworth AJ, Dexiang C, Waterstrat PR. Changes in peripheralblood leucocyte percentages and function of neutrophils instressed channel catfish. J Aquat Anim Health 1991;3:41e7.
[8] Cato AC, Wade E. Molecular mechanisms of anti-inflammatoryaction of glucocorticoids. BioEssays 1996;18:371e8.
[9] De Bosscher K, Vanden Berghe W, Haegeman G. Mechanisms ofanti-inflammatory action and of immunosuppression by gluco-corticoids: negative interference of activated glucocorticoidreceptor with transcription factors. J Neuroimmunol 2000;109:16e22.
[10] Engelsma MY, Hougee S, Nap D, Hofenk M, Rombout JHWM,van Muiswinkel WB, et al. Multiple acute temperature stressaffects leucocyte populations and antibody responses in com-mon carp, Cyprinus carpio L. Fish Shellfish Immunol 2003;15:397e410.
Cortisol response and immune-related effects of Atlantic salmon 203

Author's personal copy
[11] Pickering AD, Pottinger TG. Stress responses and disease resis-tance in salmonid fish: effects of chronic elevation of plasmacortisol. Fish Physiol Biochem 1989;58:253e8.
[12] Pickering AD, Duston J. Administration of cortisol to browntrout, Salmo trutta L., and its effects on the susceptibilityto Saprolegenia infection and furunculosis. J Fish Biol 1983;23:163e75.
[13] Maule AG, Schreck CB, Kaattari SL. Changes in the immunesystem of coho salmon (Oncorhynchus kisutch) during theparr-to-smolt transformation and after cortisol implantation.Can J Fish Aquat Sci 1987;44:161e6.
[14] Woo PTK, Leatherland JF, Lee MS. Cryptobia salmositica: cor-tisol increases the susceptibility of Salmo gairdneri Richard-son to experimental cryptobiosis. J Fish Dis 1987;10:75e83.
[15] Wiik R, Andersen K, Uglenes I, Egidius E. Cortisol-induced in-crease in susceptibility of Atlantic salmon, Salmo salar, to Vib-rio salmonicida, together with effects on the blood cellpattern. Aquaculture 1989;83:201e15.
[16] Johnson SC, Albright LJ. Effects of cortisol implants on thesusceptibility and the histopathology of the responses of naıvecoho salmon, Oncorhynchus kisutch, to experimental infec-tion with Lepeophtheirus salmonis (Copepoda: Caligidae).Dis Aquat Org 1992;14:195e205.
[17] Saeij JPJ, Verburg-van Kemenade BML, van Muiswinkel WB,Wiegertjes GF. Daily handling stress reduces resistance ofcarp to Trypanoplasma borreli: in vitro modulatory effectsof cortisol on leukocyte function and apoptosis. Dev Comp Im-munol 2003;27:233e45.
[18] Siiteri PK, Murai JT, Hammond GL, Nisker JA, Raymoure WJ,Kuhn RW. The serum transport of steroid hormones. RecentProg Horm Res 1982;38:457e510.
[19] Engelsma MY, Stet RJM, Saeij JP, Verburg-van Kemenade BML.Differential expression and haplotypic variation of two inter-leukin-1b genes in the common carp (Cyprinus carpio L.). Cy-tokine 2003;22:21e32.
[20] MetzJR,HuisingMO, LeonK,Verburg-vanKemenade BML, FlikG.Central and peripheral interleukin-1b and interleukin-1 receptorI expression and their role in the acute stress response of com-mon carp, Cyprinus carpio L. J Endocrinol 2006;191:25e35.
[21] Engelsma MY, Huising MO, van Muiswinkel WB, Flik G, Kwang J,Savelkoul HFJ, et al. Neuroendocrine-immune interactions infish: a role for interleukin-1. Vet Immunol Immunophathol2002;87:467e79.
[22] Fast MD, Muise DM, Easy RE, Ross NW, Johnson SC. The effectsof Lepeophtheirus salmonis infections on the stress responseand immunological status of Atlantic salmon (Salmo salar).Fish Shellfish Immunol 2006;21:228e41.
[23] Ottinger CA, Johnson SC, Ewart KV, Brown LL, Ross NW. En-hancement of anti-Aeromonas salmonicida activity in Atlanticsalmon (Salmo salar) macrophages by a mannose-binding lec-tin. Comp Biochem Physiol 1999;123C:53e9.
[24] Hosoya S, Johnson SC, Iwama GK, Gamperl AK, Afonso LOB.Changes in free and total plasma cortisol levels in juvenilehaddock (Melanogrammus aeglefinus) exposed to long-termhandling stress. Comp Biochem Physiol 2007;146A:78e86.
[25] Trinder P. Determination of glucose in blood using glucose ox-idase with an alternative oxygen acceptor. Ann Clin Biochem1969;6:24e7.
[26] Fast MD, Johnson SC, Jones SRM. Differential expression of thepro-inflammatory cytokines IL-1b-1, TNFa-1 and IL-8 in vacci-nated pink (Oncorhynchus gorbuscha) and chum (Oncorhynchusketa) salmon juveniles. Fish Shellfish Immunol 2007;22:403e7.
[27] Moore LJ, Somamoto T, Lie KK, Dijkstra JM, Hordvik I. Charac-terisation of salmon and trout CD8a and CD8b. Mol Immunol2005;42:1225e34.
[28] Olsvik PA, Lie KK, Jordal AEO, Nilsen TO, Hordvik I. Evaluationof potential reference genes in real-time RT-PCR studies of At-lantic salmon. BMC Mol Biol 2005;6:21e9.
[29] Van Weerd JH, Komen J. The effects of chronic stress ongrowth in fish: a critical appraisal. Comp Biochem Physiol A1998;120:107e12.
[30] Peters G, Schwarzer R. Changes in hemopoietic tissue of rain-bow trout under influence of stress. Dis Aquat Org 1985;1:1e10.
[31] Barton BA, Schreck CB, Barton LD. Effects of chronic cortisoladministration and daily acute stress on growth, physiologicalconditions, and stress responses in juvenile rainbow trout. DisAquat Org 1987;2:173e85.
[32] Barry TP, Unwin MJ, Malison JA, Quinn TP. Free and total cor-tisol levels in semelparous and iteroparous chinook salmon.J Fish Biol 2001;59:1673e6.
[33] Caldwell CA, Kattesh HG, Strange RJ. Distribution of cortisolamong its free and protein-bound fractions in rainbow trout(Oncorhynchus mykiss): evidence of control by sexual matura-tion. Comp Biochem Physiol 1991;99A:593e5.
[34] Fleshner M, Deak T, Spencer RL, Laudenslager ML, Watkins LR,Maier SF. A long term increase in basal levels of corticosteroneand a decrease in corticosteroid-binding globulin after acutestressor exposure. Endocrinology 1995;136:5336e42.
[35] Hamrahian AH, Oseni TS, Arafah BM. Measurements of serumfree cortisol in critically ill patients. N Engl J Med 2004;350:1629e38.
[36] Hammond GL. Protein functions of plasma steroid-bindingproteins. Trends Endocrinol Metab 1995;6:298e304.
[37] Metz JR, Huising MO, Meek J, Taverne-Thiele AJ, WendelaarBonga SE, Flik G. Localization, expression and control ofadrenocorticotropic hormone in the nucleus preopticusand pituitary gland of common carp (Cyprinus carpio L.).J Endocrinol 2004;182:23e31.
[38] Nakane T, Szentendrei T, Stern L, Virmani M, Seely J, Kunos G.Effects of IL-1 and cortisol on beta-adrenergic receptors, cellproliferation, and differentiation in cultured human A549 lungtumour cells. J Immunol 1990;145:260e6.
[39] Holland JW, Pottinger TG, Secombes CJ. Recombinant inter-leukin-1b activates the hypothalamicepituitaryeinterrenalaxis in rainbow trout, Oncorhynchus mykiss. J Endocrinol2002;175:261e7.
[40] Zou J, Holland J, Pleguezuelos O, Cunningham C,Secombes CJ. Factors influencing the expression of interleu-kin-1b in cultured rainbow trout (Oncorhynchus mykiss) leuco-cytes. Dev Comp Immunol 2000;24:575e82.
[41] Redding JM, Patino R, Schreck CB. Clearance of corticoste-roids in yearling coho salmon, Oncorhynchus kisutch, in freshwater and seawater and after stress. Gen Comp Endocrinol1984;54:433e43.
[42] Barton BA, Ribas L, Acerete L, Tort L. Effects of chronic con-finement on physiological responses of juvenile gilthead seabream, Sparus aurata L., to acute handling. Aqua Res 2005;36:172e9.
[43] Schreck CB. Immunomodulation: endogenous factors. In:Iwama GK, Nakanishi T, editors. The fish immune system: or-ganism, pathogen, and environment. San Diego: AcademicPress; 1996. p. 311e37.
[44] Yada T, Nakanishi T. Interaction between endocrine and im-mune systems in fish. Int Rev Cytol 2002;220:35e92.
[45] Basu N, Nakano T, Grau EG, Iwama GK. The effects of cortisolon heat shock protein70 in two fish species. Gen Comp Endo-crinol 2001;124:97e105.
[46] McEwen BS. Protective and damaging effects of stress media-tors. N Engl J Med 1998;338:171e9.
[47] Schreck CB. Accumulation and long-term effects of stress infish. In: Moberg GP, Mench JA, editors. The biology of animalstress: basic principles and implications for animal welfare.New York: CABI Publishing; 2000. p. 147e58.
[48] Gayrard V, Alvinerie M, Toutain PL. Interspecies variations ofcorticosteroid-binding globulin parameter. Dom Anim Endocri-nol 1996;13:35e45.
204 M.D. Fast et al.




















